Page 1
Advances in Environmental Biology, 10(9) September 2016, Pages: 20-29
AENSI Journals
Advances in Environmental Biology
ISSN-1995-0756 EISSN-1998-1066
Journal home page: http://www.aensiweb.com/AEB/
Copyright © 2016 by authors and Copyright, American-Eurasian Network for Scientific Information (AENSI Publication).
Bioecology of Culicidae (Diptera ; Nematocera) of Taksebt dam of Tizi-Ouzou (Algeria)
1Lounaci Z., 2Doumandji S., 2Doumandji- Mitiche B., 1Taguemout M.
1Department of Agronomy, University of Mouloud Mammeri, PB15000 Tizi Ouzou, Algeria. 2Department of Agricultural and Forest Zoology, Upper Agronomical National School, El Harrach., Algeria. Avenue Hassan Badi،El Harrach, Algeria.
Address For Correspondence: LOUNACI_Zohra, Department of Agronomy, University of Mouloud Mammeri,Tizi Ouzou, Algeria. Phone : +213 774015595 ; E-mail : [email protected] , Postal : Street Segna, 16112, Reghaia, Bt 8- Aprt. 23, Algiers, Algeria.
This work is licensed under the Creative Commons Attribution International License (CC BY). http://creativecommons.org/licenses/by/4.0/
Received 12 July 2016; Accepted 18 September 2016; Available online 22 September 2016
ABSTRACT Background: The Culicidae’s bioecological study is performed from April to August 2014 at Taksebt dam (Tizi-Ouzou). Whose it is
considered as a protected area of national importance. It is gathering in fact favourable conditions in settlement and multiplication of
terrestrial and aquatic insects and to maintaining wild mammals. As result to this remarkable diversity. Its biological functions confer on this potential wetland a capacity to be produced and nourish living matter and to become a tank of the biodiversity. Three natural sites
have been selected as stations and are the study object of their mosquitoes larvae population (upstream, near the dam’s dike and downstream
of the dam Taksebt). Capture of Culicidae larvae is made by use of a stringer net. Objective: It is underlined that not much works have been conducted on richness of Diptera Culicidae among Entomofauna settling humid ecosystems such as the dam Taksebt of Tizi Ouzou
in Algeria. So, the choice of the present work is made in view of Culicidae species importance within present arthropod fauna at level of
Taksebt dam. Their identification and study of their bio ecology are essential, to guide and to refine preventive fight method such as surveillance of larval habitats of pathogenic vectors agent’s species. Results: Eight species belonging to two sub-families are identified;
it is about Anopheles labranchiae, Culex hortensis, Culex impudicus, Culex theileri, Culex perexiguus, Culiseta longiareolata, Aedes
caspius and Aedes vexans. This study allows us to specify the distinctive morphological characters of species harvested at the larval stage. In terms of number by species and by station, it should be noted that Culex hortensis is strongly present in upstream, and in downstream
of the dam with respectively 419 individuals (65, 88%) and 356 individuals (97, 2%). These two environments represent breeding sites the
most favourable to larvae. By contrast, in the centre of the dam, Culex impudicus is the first place with a number of 39 individuals (65%). The populations dynamics of Culex hortensis revealed that species is developing in summer and seems to follow a classic thermophilic
trend. Conclusion: Based upon behavior and ecology of the identified mosquito species, studied various sites of the Taksebt Dam of Tizi
Ouzou seem to provide a suitable environment and breeding sites for the proliferation of the species of Culicidae in particular Anopheles labranchiae potential vectors of arbovirus and those being a problem of real nuisance for the close inhabitants.
KEYWORDS: Bioecology, Culicidae, Taksebt dam; population’s dynamics.
INTRODUCTION
In Algeria, Culicidae constitute the biting insects the most harmful to populations, some species may transmit
infectious diseases (malaria particularly).Regular anti-mosquito campaigns are being conducted against these
insects to reduce their nuisance at level of urban and rural centres. Efficiency of such struggle whether chemical
or biological, is dependent of ecology knowledge of these insects. It is a fact that undeniable success are obtained
against numerous outbreaks of malaria. However, it should be emphasised that malaria has not vanished from
Mediterranean Africa, which remains at hypo-endemic status and may reoccur at any time under its epidemic
form under influence of climatic circumstances favouring anopheline development [46, 25, 23]. Furthermore, it
would be appropriate to monitor not only the possible arrival of new vectors as Aedes albopictus which has been
recently established in Europe and in the major countries of Mediterranean Basin [31, 1, 43, 65, 49, 67]including
Page 2
21 Lounaci Z., et al, 2016
Advances in Environmental Biology, 10(9) September 2016, Pages: 20-29
Algeria in North-Africa [37]. It should also be pointed out the introduction of new pathogenic agents such as West
Nile virus, which is regularly circulating on the Mediterranean littoral which one of vectors is Culex pipiens [22,
56, 5, 19]. Europe is also at risk of introduction of mosquito-borne equine alphavirus from Americas [16]. and
Arboviruses particularly dengue, chikungunya, and Zika viruses. transmitted by the yellow fever mosquito, Aedes
aegypti [32, 2]. This rapid global spread was favored by international trade, especially of used tyres, and by its
physio-local and ecological plasticity, which allow the species to thrive in a wide range of climates and habitats
[67]. Other mosquitoes pathogenic agent’s vectors are a real scourge by their painful and annoying bites constitute
in fact a nuisance problem [10, 33]. It is within this context that initiation of mosquito’s study in the region is a
necessary prerequisite towards a more general study. In Algeria, ecological studies on Culididae remain
fragmentary [34, 33, 11]because it concerns above all systematic and morphometrics [6, 8, 14, 13, 44, 9, 35, 57,
59], biology and chemical struggle [51, 52, 7, 6, 15] with this in mind that involves our study on Culicidae of
Taksebt’s dam which constitutes an ecosystem whose objective is :
- to take an inventory of Culicidae species present in this humid area ;and determine biodiversity
- to study seasonal dynamics of preimaginal population (larvae and nymphs) of the main Culex hortensis.s’
species.
This bioecological study constitutes an essential contribution to all reasoned action in species preservation,
environments and preventing against Culicidae nuisance and vector-borne diseases.
MATERIAL AND METHODS
Study sites:
The present study took place in 2014 in Taksebt’s dam (36° 24’ 15”N; 4° 45’ 46”E). The dam is located on
Oued Aissi of approx. 10 km south-east of Tizi-Ouzou town (Fig 1). The whole site presents a capacity of 175
millions of m3 that is spreading on a surface of 550 hectares. It is fed by rain water and the melting snowpack of
Djurdjura. Oued Assi comes down from Djurdjura Massif, generally towards North-South direction until its
confluence into Oued Sebaou. This last one in turn flows into the Mediterranean. This region belongs to sub-
humid bioclimatic stage with temperate winter according to data of 1998 to 2013. The annual rainfall is of 808
mm. They are generally spreading from October to April with peaks in December, January and February.
Fig. 1: Taksebt’s dam of Tizi-Ouzou,A: [42], B, C: Google Earth, 2014
Page 3
22 Lounaci Z., et al, 2016
Advances in Environmental Biology, 10(9) September 2016, Pages: 20-29
Culicidae fauna monitoring was performed with help of 15 serial samplings made at intervals of 8 days on
duration of 4 months (May to August 2014). Three epigeous sites (in the open air) are retained as stations and are
subject of study of their mosquitoes larvae populations ; their choice is supported on representativeness of
Culicidae larvae in a productive sites batch, accessibility, sustainability and no treatment by chemical product.
(Fig. 1 and 2):
Station I:
is located on the upstream side of the dike (36°40'12.39"N ; 4° 6'51.44"E) at about 800 m to South of the
dike where small ponds are formed due to the water level decrease of the dam. This site is surrounded by trees
layer composed essentially of Populus nigra, populous alba and Nerium oleander (Fig.2a).
Station II:
is located near the dam’s dike36°40'31.24"N;4° 6'57.93"E). Three vegetative strata: herbaceous, shrubby and
tree are present (Fig. 2b).
Station III:
is located on the downstream of Taksebt’s dam (36°40'50.34"N, 4° 7'0.67"E). It is constituted of waters
coming from dam’s overflow that form small ponds. This station is rich in dense vegetation composed of
Asphodelus microcarpus, of Juncus acutus and of Typha angustifolia (Fig. 2c).
Fig. 2: Station of Taksebt’s dam. a: Upstream of the dam, b:near the dam’s dike, c: downstream of the dam
(Lounaci, 2014).
Sampling technique of Culicidae populations:
Sampling of preimaginal stages (larval) have been made according to the technique of dipping (dipper blow)
[54, 62, 48, 21]. The dipper of known capacity (c=1l) is put into water, then moved with care avoiding to make
turmoil. Ten blows of dipper are therefore made in different spots of the site with a space of 1 m. In laboratory;
larvae are selected by species, by stages and counted. For determination of those ones, we consulted works of
[53, 57, 58]. This one-time study has also been facilitated by use of identification software of Culicidae of
Mediterranean Africa performed by [18].
Ecological index:
Results obtained on bioecology of Culicidae in Taksebt’s dam are treated by ecological index. The specific
richness in species (S) the relative abundance expressed in percentage ((pi = ni x 100/ N is the ratio of individuals
number of Culicidae species ni to total number of individuals of all species inventoried N is frequency of
occurrence (constancy) of species in each of the study stations. Constancy (C) is calculated with index C = P x
100 / N, where P is the number collected containing species studied and N is the total number of collections made
[27, 50]. In each site studied, diversity index of Shannon-Weaver (H’) is the fairness (F) are calculated. The first,
H’=− ∑ 𝒑𝒊𝐋𝐨𝐠𝟐𝒑𝒊𝒊=𝑺𝒊=𝑰 where pi = ni /N is the relative frequency of individuals category according to i. is used to
measure and evaluate diversity of mosquito species for each of the three study stations. Equitability (E) = H’/
H’max (where H’max =log2S) was calculated to measure the distribution of individuals within species irrespective
of species richness. It varies from 0, if only one species dominates, to 1, if all species show similar abundance
[60].
Results:
In total 1062 larvae of Cuticidae are inventoried on 2014 in Taksebt’s dam (tab 1). Morphological
identifications show presence of 8 species belonging to two sub-families: Anophelinae and Culicinae. Among
captured species in all stations, Culex hortensis (72, 98%) and Culiseta longiareolata (19, 68 %) are the most
abundant
Page 4
23 Lounaci Z., et al, 2016
Advances in Environmental Biology, 10(9) September 2016, Pages: 20-29
Table 1: relative abundance, RA%; frequency occurrence (constancy), Co; of Culicidae larvae per sampling station in 2014in Taksebt’s dam
Station I : Upstream of the dam Station II : near the dam’s dike Station III : downstream of the dam -
Species Ni A.R.%
F.O.% Co
Ni A.R.%
F.O.%
Co
Ni
A.R.%
F.O.% Co
Anopheles labranchiae
_ _ _ _ 8 13,33 6,66 At _ _ _ _
Aedes caspius _ _ _ _ 7 11,66 13,33 At _ _ _ _
Aedes vexans _ _ _ _ 1 1,67 6,66 At _ _ _ _
Culex hortensis 419 65,88 93,33 O _ _ _ _ 356 97,2 86,66 O
Culex perexiguus 0 0 0 _ 4 6,67 6,66 At 0 0 0 _
Culex theileri 5 0,79 6,66 At 1 1,67 6,66 At 0 0 0 _
Culex impudicus 6 0,94 13,33 At 39 65 26,66 Ac 7 1,9 20 At
Culiseta
longiareolata
206 32,39 20 At _ _ _ _ 3 0.9 13,33 At
Totals 636 100 _ _ 60 100 _
_ 366 100 _ _
(A); Accidental; (Ac) : Accessory ; ( (O) : Omnipresent.
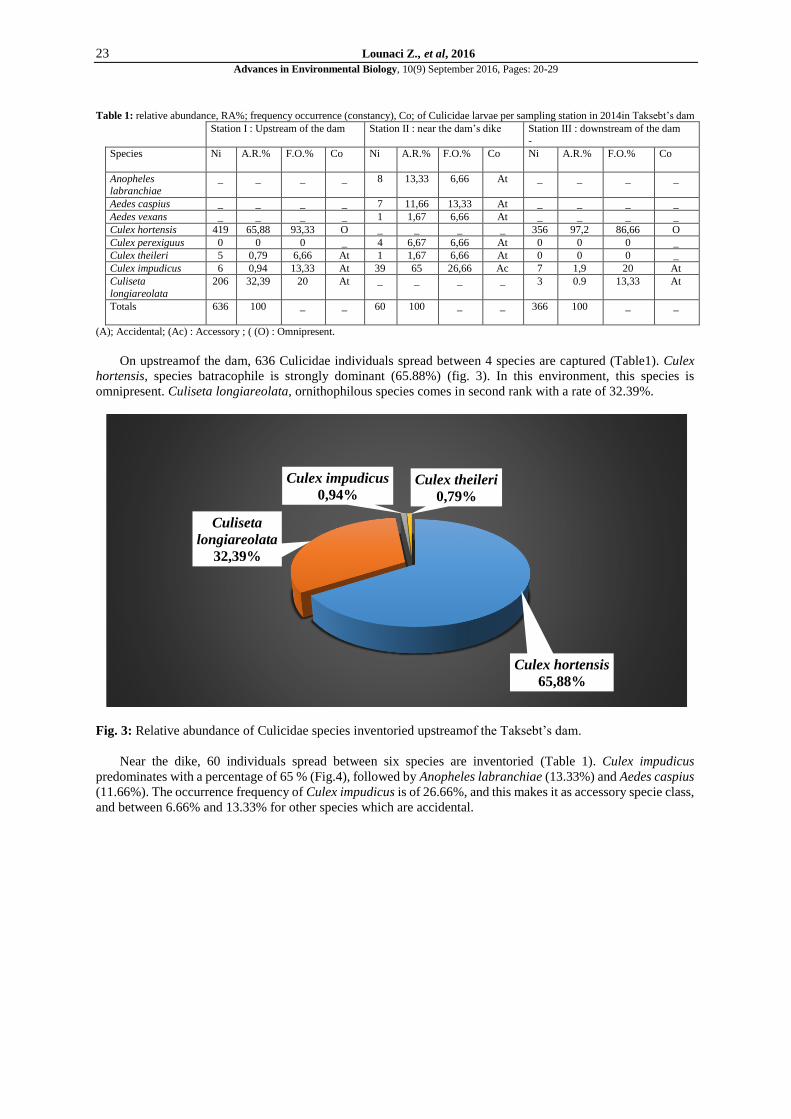
On upstreamof the dam, 636 Culicidae individuals spread between 4 species are captured (Table1). Culex
hortensis, species batracophile is strongly dominant (65.88%) (fig. 3). In this environment, this species is
omnipresent. Culiseta longiareolata, ornithophilous species comes in second rank with a rate of 32.39%.
Fig. 3: Relative abundance of Culicidae species inventoried upstreamof the Taksebt’s dam.
Near the dike, 60 individuals spread between six species are inventoried (Table 1). Culex impudicus
predominates with a percentage of 65 % (Fig.4), followed by Anopheles labranchiae (13.33%) and Aedes caspius
(11.66%). The occurrence frequency of Culex impudicus is of 26.66%, and this makes it as accessory specie class,
and between 6.66% and 13.33% for other species which are accidental.
Culex hortensis
65,88%
Culiseta
longiareolata
32,39%
Culex impudicus
0,94%Culex theileri
0,79%
Page 5
24 Lounaci Z., et al, 2016
Advances in Environmental Biology, 10(9) September 2016, Pages: 20-29
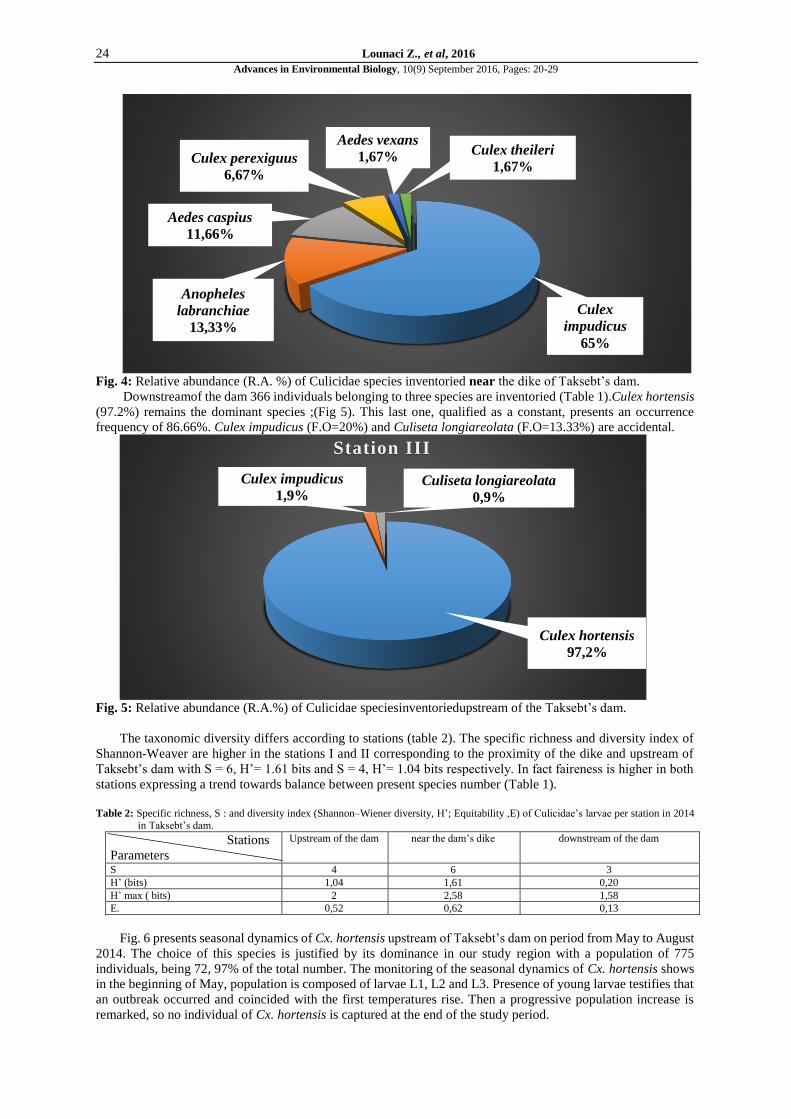
Fig. 4: Relative abundance (R.A. %) of Culicidae species inventoried near the dike of Taksebt’s dam.
Downstreamof the dam 366 individuals belonging to three species are inventoried (Table 1).Culex hortensis
(97.2%) remains the dominant species ;(Fig 5). This last one, qualified as a constant, presents an occurrence
frequency of 86.66%. Culex impudicus (F.O=20%) and Culiseta longiareolata (F.O=13.33%) are accidental.
Fig. 5: Relative abundance (R.A.%) of Culicidae speciesinventoriedupstream of the Taksebt’s dam.
The taxonomic diversity differs according to stations (table 2). The specific richness and diversity index of
Shannon-Weaver are higher in the stations I and II corresponding to the proximity of the dike and upstream of
Taksebt’s dam with S = 6, H’= 1.61 bits and S = 4, H’= 1.04 bits respectively. In fact faireness is higher in both
stations expressing a trend towards balance between present species number (Table 1).
Table 2: Specific richness, S : and diversity index (Shannon–Wiener diversity, H’; Equitability ,E) of Culicidae’s larvae per station in 2014
in Taksebt’s dam.
Stations
Parameters
Upstream of the dam near the dam’s dike downstream of the dam
S 4 6 3
H’ (bits) 1,04 1,61 0,20
H’ max ( bits) 2 2,58 1,58
E. 0,52 0,62 0,13
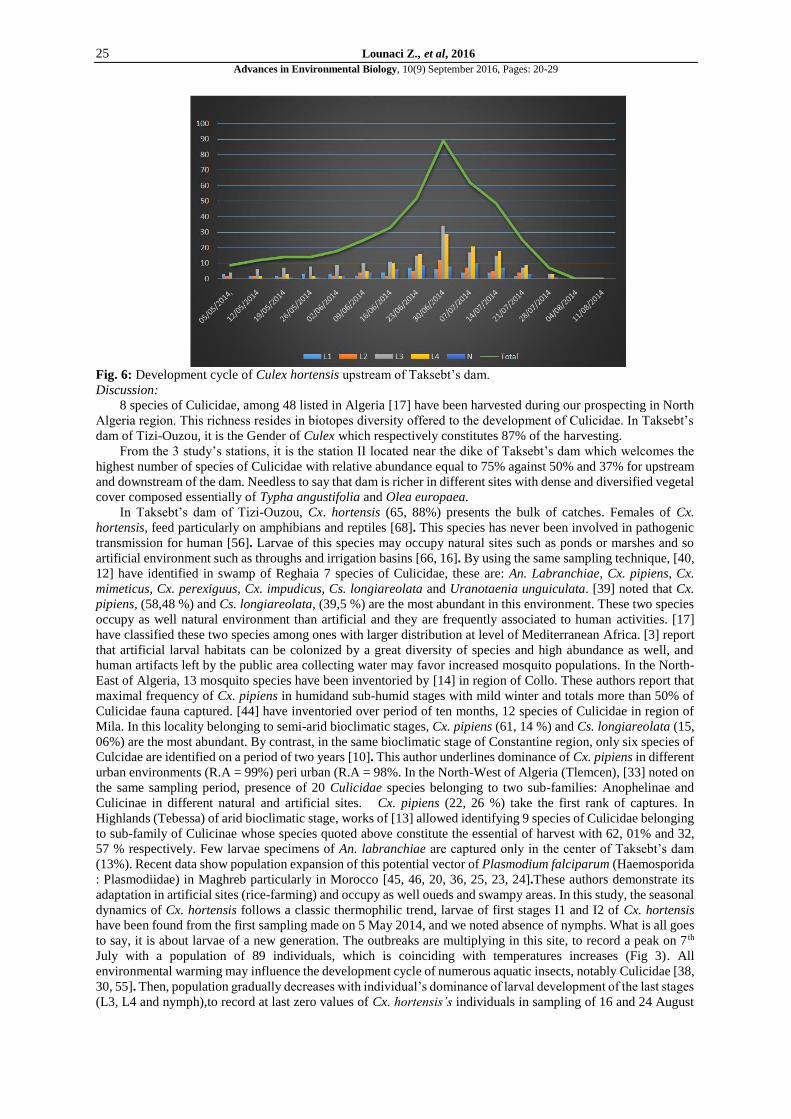
Fig. 6 presents seasonal dynamics of Cx. hortensis upstream of Taksebt’s dam on period from May to August
2014. The choice of this species is justified by its dominance in our study region with a population of 775
individuals, being 72, 97% of the total number. The monitoring of the seasonal dynamics of Cx. hortensis shows
in the beginning of May, population is composed of larvae L1, L2 and L3. Presence of young larvae testifies that
an outbreak occurred and coincided with the first temperatures rise. Then a progressive population increase is
remarked, so no individual of Cx. hortensis is captured at the end of the study period.
Culex
impudicus
65%
Anopheles
labranchiae
13,33%
Aedes caspius
11,66%
Culex perexiguus
6,67%
Aedes vexans
1,67%Culex theileri
1,67%
Culex hortensis
97,2%
Culex impudicus
1,9%Culiseta longiareolata
0,9%
Station III
Page 6
25 Lounaci Z., et al, 2016
Advances in Environmental Biology, 10(9) September 2016, Pages: 20-29
Fig. 6: Development cycle of Culex hortensis upstream of Taksebt’s dam.
Discussion:
8 species of Culicidae, among 48 listed in Algeria [17] have been harvested during our prospecting in North
Algeria region. This richness resides in biotopes diversity offered to the development of Culicidae. In Taksebt’s
dam of Tizi-Ouzou, it is the Gender of Culex which respectively constitutes 87% of the harvesting.
From the 3 study’s stations, it is the station II located near the dike of Taksebt’s dam which welcomes the
highest number of species of Culicidae with relative abundance equal to 75% against 50% and 37% for upstream
and downstream of the dam. Needless to say that dam is richer in different sites with dense and diversified vegetal
cover composed essentially of Typha angustifolia and Olea europaea.
In Taksebt’s dam of Tizi-Ouzou, Cx. hortensis (65, 88%) presents the bulk of catches. Females of Cx.
hortensis, feed particularly on amphibians and reptiles [68]. This species has never been involved in pathogenic
transmission for human [56]. Larvae of this species may occupy natural sites such as ponds or marshes and so
artificial environment such as throughs and irrigation basins [66, 16]. By using the same sampling technique, [40,
12] have identified in swamp of Reghaia 7 species of Culicidae, these are: An. Labranchiae, Cx. pipiens, Cx.
mimeticus, Cx. perexiguus, Cx. impudicus, Cs. longiareolata and Uranotaenia unguiculata. [39] noted that Cx.
pipiens, (58,48 %) and Cs. longiareolata, (39,5 %) are the most abundant in this environment. These two species
occupy as well natural environment than artificial and they are frequently associated to human activities. [17]
have classified these two species among ones with larger distribution at level of Mediterranean Africa. [3] report
that artificial larval habitats can be colonized by a great diversity of species and high abundance as well, and
human artifacts left by the public area collecting water may favor increased mosquito populations. In the North-
East of Algeria, 13 mosquito species have been inventoried by [14] in region of Collo. These authors report that
maximal frequency of Cx. pipiens in humidand sub-humid stages with mild winter and totals more than 50% of
Culicidae fauna captured. [44] have inventoried over period of ten months, 12 species of Culicidae in region of
Mila. In this locality belonging to semi-arid bioclimatic stages, Cx. pipiens (61, 14 %) and Cs. longiareolata (15,
06%) are the most abundant. By contrast, in the same bioclimatic stage of Constantine region, only six species of
Culcidae are identified on a period of two years [10]. This author underlines dominance of Cx. pipiens in different
urban environments (R.A = 99%) peri urban (R.A = 98%. In the North-West of Algeria (Tlemcen), [33] noted on
the same sampling period, presence of 20 Culicidae species belonging to two sub-families: Anophelinae and
Culicinae in different natural and artificial sites. Cx. pipiens (22, 26 %) take the first rank of captures. In
Highlands (Tebessa) of arid bioclimatic stage, works of [13] allowed identifying 9 species of Culicidae belonging
to sub-family of Culicinae whose species quoted above constitute the essential of harvest with 62, 01% and 32,
57 % respectively. Few larvae specimens of An. labranchiae are captured only in the center of Taksebt’s dam
(13%). Recent data show population expansion of this potential vector of Plasmodium falciparum (Haemosporida
: Plasmodiidae) in Maghreb particularly in Morocco [45, 46, 20, 36, 25, 23, 24].These authors demonstrate its
adaptation in artificial sites (rice-farming) and occupy as well oueds and swampy areas. In this study, the seasonal
dynamics of Cx. hortensis follows a classic thermophilic trend, larvae of first stages I1 and I2 of Cx. hortensis
have been found from the first sampling made on 5 May 2014, and we noted absence of nymphs. What is all goes
to say, it is about larvae of a new generation. The outbreaks are multiplying in this site, to record a peak on 7 th
July with a population of 89 individuals, which is coinciding with temperatures increases (Fig 3). All
environmental warming may influence the development cycle of numerous aquatic insects, notably Culicidae [38,
30, 55]. Then, population gradually decreases with individual’s dominance of larval development of the last stages
(L3, L4 and nymph),to record at last zero values of Cx. hortensis’s individuals in sampling of 16 and 24 August
Page 7
26 Lounaci Z., et al, 2016
Advances in Environmental Biology, 10(9) September 2016, Pages: 20-29
due to raising of temperature generating therefore, an intense water evaporation, and consequently a drop in water
level and to the drying of the sites. Temperature can favour preimaginal population development of Cx. hortensis
because in sites of Taksebt’s dam, the highest larvae populations are observed from May (Fig 3). ). [28, 64,] report
that temperature and food quantity intervene in regulation process of the larvae development speed of Ae caspius.
So the spring period is associated to a strong proliferation of preimaginal stages. Several studies demonstrate that
high temperatures seem to favour abundance of mosquito larvae and to reduce their development time, particularly
larvae of Culex species [29]. According to [11], larval development is faster when water temperature increases.
It determines Cx. pipiens larvae’s development speed acting principally on larval evolution duration and to a
lesser degree on nymphal evolution duration. [4] reports that larval development of Ae. aegypt is soaked not only
by low temperatures, but also by high temperatures. That is explaining shortage (decrease) of Cx. hortensis’s
larval number from end of July and also their absence in August. According to [38, 30, 26, 41], Any warming or
cooling of the environment can disturb development cycle of several aquatic insects notably in Culicidae.
Furthermore [63] underlines in addition of ecological factors influence on larval development, there are pregnant
females’ attractiveness according to a site that may explain presence of larval stages. According to [62]
oviposition is determined by the site’s structure which can be easy access for females.
Conclusion:
Bioecological study of Culicidae in Taksebt’s dam of Tizi-Ouzou is based on dipping technique use, during
a period of 4 months going from 5th May to 28 August 2014. Eight species, belonging to two sub-families are
identified. Those one of Anophelinae with a single species, in this case An. labranchiae; potential causal agent
of malaria and those one of Culicinae with seven species ; these are Cx. hortensis, Cx. impudicus, Cx. theileri,
Cx. perexiguus, Cs. longiareolata , Ae. vexans (virus vector of Tahyna), and at last Ae. caspius. Medical
entomologist and veterinarian think that these are the main vector of infectious diseases, quoting West Nil virus,
which is transmitted, to human by Ae. caspius. From the 3 study’s stations, it is the station II located near the dike
of Taksebt’s dam which welcomes the highest number of species of Culicidae. In terms of number by species and
by station, it should be noted that Cx. hortensis is strongly present in upstream, and in downstream of the dam.
One-time study carried out on population of Cx. hortensis is based on the monitoring of variation in time and
space of immature stages. So, larval number is relatively high in sites bordered by diversified vegetation.
Population explosion provoked by positive climate conditions in May and June allows to this species to settle
study sites. In summer period, from end of July, absence of larvae in the site would be due to a temporary nature
of these environments. In an additional study, other sampling techniques for mature stage capture are taken into
account. However, it is important to further studies on inventory, taxonomy and ecology of these organisms for
year-round in order to cover all seasons and being able to detect period of these mosquitoes’ intense proliferations.
Also it should be necessary to extend the study towards other regions, particularly on species with high
transmission risk or having origin of intense nuisance. That should allow us to put in place, permanently, the
appropriate means to fight and to control their populations. This issue is important because it concerns public
health. We need to bring attention on risk of vector agents’s arrival carrying exotic diseases from African Sahel
and this is due to the global warming. The whole information should obviously be reinforced by new studies on
the ground, with particularly a more detailed analysis about determinism of some processes; already would give
an image on spatio-temporal distribution of Culicidae species. This information is likely to be used in fighting
organisation against these vectors.
ACKNOWLEDGEMENTS
We heartily thank all the staff of Hydraulic Service Department of Wilaya Tizi-Ouzou for their collaboration
on the ground. The collection of Culicidae would not be possible without their assistance.
Contributions to knowledge Subject:
This study contributes to the knowledge of Culicidae fauna of the wetlands which present a context
particularly favorable to the developments of certain transmissible vectorial diseases. This study put the point on
the remarkable richness, abundance, the diversity and the monthly fluctuation of Culicidae in one of these zones
(Dam Taksebt Algeria) subjected to considerable ecological upheavals. This with a view to put at the disposal of
actors of fight against the diseases transmitted by the mosquitos adatabase reliable and brought up to date
necessary for planning, the follow-up and the evaluation of antivectorielle operations of fight.
REFERENCES
[1] Aguilar, V.P., J.G. Estrada-Franco, R. Navarro-Lopez, C. Ferro, A.D. Haddow and S.C. Weaver, 2011.
Endemic Venezuelan equine encephalitis in the Americas: hidden under the dengue umbrella. Future Virol.,
6: 721-740.
Page 8
27 Lounaci Z., et al, 2016
Advances in Environmental Biology, 10(9) September 2016, Pages: 20-29
[2] Ahmed, M.A.I., C.F.A. Vogel, 2016. Sulfoxaflor: A New Promising Insecticide in the Global Control of
Dengue and Zika Vector Aedes aegypti. (Diptera: Culicidae) Mosquito. Advances in Environmental Biology,
10(3): 171-177.
[3] Alencar, J., C.M. Ferreira, N.M. Serra-Freire, R.S. Mello, A.E. Guimarães, R. Hélcio, G. Santana, R.M.
Gleiser, 2016. Biodiversity and Temporal Distribution of Immature Culicidae in the Atlantic Forest, Riode
Janeiro State, Brazil. Plos One, 11(7):1-15.
[4] Bar-Zeev, M., 1958. The effect of temperature on the growth rate and survival of the immature stages of
Aedes aegypti (L.). Bull. Entomol. Res., 49: 157-163.
[5] Becker, N., D. Petric, M. Zgomba, C. Boase, C. Dahl, M. Madon and A. Kaiser, 2010. Mosquitoes and Their
Control, 2nd edition. Springer, Berlin Heidelberg, Berlin, p: 579.
[6] Bendali, F., 2006. Etude bioécologique, systématique et biochimique des Culicidae (Diptera - Nematocera)
de la région d’Annaba. Lutte biologique aniti-culicidienne. Thèse de Doctorat en Biologie animale.
Département de Biologie. Université d’Annaba, Algérie, p : 224.
[7] Bendali, F., F. Djebbar and N. Soltani, 2001. Efficacité comparée de quelques espèces de poissons à l'égard
de divers stades de Culex pipiens L. dans des conditions de laboratoire. Parasitica, 57(4) : 255-265.
[8] Bendali-Saoudi, F., W. Oudainia, L. Benmalek, A.Tahar and N. Soltani, 2013. Morphometry of Culex pipiens
pipiens Linneus, 1758 (Dipterae; Culicidae) principal vector of West Nile Virus, harvested from two zones,
humid, semiarid (East of Algeria). Annals of Biological Research, 4(10): 79-86.
[9] Berchi, S., 2000a. Bioécologie de Culex pipiens L. (Diptera, Culicidae) dans la région de Constantine et
perspectives de lutte. Thèse Doc. es-Sciences, univ. Conctantine, p : 133.
[10] Berchi, S., 2000b. Résistance de certaines populations de Culex pipiens pipiens L. au malathion à Canstantine
(Algerie) (Diptera, Culicidae). Bull. Soc. Ent. France, 105(2) : 125-129.
[11] Berchi, S., A. Aouati and K. Louadi, 2012. Typologie des gîtes propices au développement larvaire de Culex
pipiens L. 1758 (Diptera-Culicidae), source de nuisance à Constantine (Algérie). ecologia méditerranea,
38(2) : 5-16.
[12] Berrouane, F.Z., H. Idouhar-Saadi, Z. Lounaci, K. Souttou, K. Mahdi and S. Doumandji, 2016. Bio-
ecological relationship on Diptera’s order among invertebrates of Reghaia Lake (Algeria). Advances in
Environmental Biology, 10(5): 254-264.
[13] Bouabida H., F. Djebbar and N. Soltani, 2012. Etude systématique et écologique des Moustiques (Diptera:
Culicidae) dans la région de Tébessa (Algérie). Faunistic Entomology, 65: 99-103.
[14] Boudemagh, N., F. Bendali-Saoudi and N. Soltani, 2013. Inventory of Culicidae (Diptera: Nematocera) in
the region of Collo (North-East Algeria). Annals of Biological Research, 4(2): 94-99.
[15] Boudjelida, H., A. Bouaziz, T. Soin, G. Smagghe and N. Soltani, 2005. Effects of ecdysone agonist
halofenozide against Culex pipiens.Pestic. Biochem. Physiol., 83: 115-123.
[16] Boukraa, S., M.A. Grandiereb, T. Bawina, F.N. Raharimalalac, J.Y. Zimmer, E. Haubrugea, E. Thiryb and
F. Francis, 2016. Diversity and ecology survey of mosquitoes potential vectors inBelgian equestrian farms:
A threat prevention of mosquito-borneequine arboviruses. Preventive Veterinary Medicine, 124: 58-68.
[17] Brunhes, J., K. Hassaine, A. Rhaim and J.P. Hervy, 2000. Les Culicides de l’Afrique méditerranéenne:
espèces présentes et répartition (Diptera, Nematocera) . Bull. Soc. Ent. France, 105(2) : 195-204.
[18] Brunhes, J., A. Rhaim, B. Geoffroy, G. Angel and J.P. Hervy, 1999. Les Culicidae d’Afrique
méditerranéenne. Logiciel d’identification et d’enseignement, Montpellier, France, IRD & ITP, CD-Rom
collection didactique, IRD.
[19] Calzolari, M., P. Bonilauri, R. Bellini, A. Albieri, F. Defilippo, M. Tamba, M. Tassinari, A. Gelati, P.
Cordioli, P. Angelini and M. Dottori, 2013. Usutu Virus Persistenceand West Nile Virus Inactivity in the
Emilia-Romagna Region (Italy) in 2011.PLos ONE 8(5): e63978.
[20] Chlaida, M. and A. Bouzidi, 1995. Contribution à l’étude des Diptères Culicidae : Dynamique et cartographie
écologique de quelques espèces au sein de la retenue du barrage Al Massira (Sud de Setta) Bull. inst.
Sci.Rabat, 19: 83-92.
[21] Croset, H., B. Papierok, J. A. Rioux, A. Gabinaud, J. Cooserans and D. Arnaud, 1976. Absolute estimates of
larval population of Culicidae mosquitoes of capture – recapture, removal and dipping methods. Ecolog.,
Ent., 1: 251-256.
[22] Durand, B., G. Dauphin, J. Labie, H. Zeller and S. Zientara, 2005. Résultats d'une enquête sérologique sur
l'infection à virus West Nile chez les équidés dans le Var, en 2003 Environnement, Risques & Santé 4(2):
114-118.
[23] El Joubari, M., A. Louah and O. Himmi, 2014. Mosquitoes (Diptera, Culicidae) of Smir marshes (northwest
of Morocco): inventory and biotypology. Bull. Soc. Pathol. Exot., 107: 48-59.
[24] El Joubari, M., C. Faraj, A. Louah, O. Himmi, 2015. Sensibilité des moustiques Anopheles labranchiae,
Culex pipiens, Ochlerotatus detritus et Ochlerotatus caspius de la région de Smir (Nord-Ouest du Maroc)
aux organophosphorés utilisés en santé publique. Environnement, Risques & Santé, 14(1) : 72-9.
Page 9
28 Lounaci Z., et al, 2016
Advances in Environmental Biology, 10(9) September 2016, Pages: 20-29
[25] El Ouali Lalami, A., A. Hindi, A. Azzouzi, 2010. Inventaire et répartition saisonnière des Culicidae dans le
centre du Maroc. Entomology, 62(4) : 131-138.
[26] Elkaiem, B., 1972. Contribution à l’étude écologique et biologique des Culilcides Aedes detritus et Aedes
caspius. Bull. Soc. Scien. Nat. Phys. Maroc, 52, 3-4: 197-204.
[27] Frontier, S., 1983. Stratégie d’échantillonnage en écologie. Masson, Paris, p : 494.
[28] Gabinaud, A., 1975. Ecologie de deux Aedes halophiles du littoral français : Aedes (O) caspius, (Pallas, 1771)
et. Aedes (O) detritus (Haliday, 1833). Utilisation de la végétation comme indicateur biologique pour
l’établissement d’une carte écologique. Application en dynamique des populations. Ph D. Thèse. Université
de Montepellier, p : 474.
[29] Gardner, A.M., G.L. Hamer, A.M. Hines, C.M. Newman, E.D. Walker and M.O. Ruiz, 2012. Weather
variability affects abundance of larval Culex (Diptera: Culicidae) in storm water catch basins in suburban
Chicago. J. Med. Entomol., 49: 270-276.
[30] Gaud, J., 1953. Notes biogéographiques sur les Culicides du Maroc. Arch. Inst. Pasteur, Maroc, IV(7) : 443-
490.
[31] Gratz, N.G., 2004. Critical review of the vector status of Aedes albopictus. Medical and Veterinary
Entomology, 18: 215-227.
[32] Hahn, M.B., R.J. Eisen, L. Eisen, K.A. Boegler, C.G. Moore, J. Mc. Allister, H.M. Savage and J.P. Mutebi,
2016. Reported Distribution of Aedes (Stegomyia) aegypti and Aedes (Stegomyia) albopictus in the United
States, 1995-2016 (Diptera: Culicidae). Journal of Medical Entomology, pp: 1-7.
[33] Hassaine, K., 2002. Bioécologie et biotypologie des Culicidae (Diptera, Nematocera) de l’Afrique
méditerranéenne. Biologie des espèces les plus vulnérantes (Ae. caspius, Ae. detritus, Ae. mariae et Cx.
pipiens) dans la région occidentale algerienne. Thèse Doc. es-Sciences, Univ. Tlemcen, p: 191.
[34] Hassaine, K., S. Gourmala and G. Metge, 2001. Cinétique démographique des populations pré-imaginales
d’Aedes mariae (Diptera : Culicidae) des côtes occidentales algeriennes. Ann. Limnol., 37(1): 59-69.
[35] Himmi, O., M. Dakki, B. Trari and M.A. Elagbani, 1995. Les Culicidae du Maroc. Clés d'identification avec
données biologiques et écologiques. Travaux de l’Institut Scientifique, série Zoologie, Rabat pp: 44 - 50.
[36] Himmi, O., B. Trari, M.A. El Agbani and M. Dakki, 1998. Contribution à la connaissance de la cinétique et
des cycles biologiques des moustiques (Diptera, Culicidae) dans la région de Rabat-Kénitra (Maroc). Bulletin
de l'Institut Scientifique, Rabat, 21: 71-79.
[37] Izri, A., I. Bitam, and R.N. Charrel, 2011. First entomological documentation Clinical Microbiology and
Infection, 17: 1116-1118.
[38] Kirkpatrick, T.W., 1925. The mosquitoes of Egypt. Government Press Cairo, p: 244.
[39] Lounaci, Z. and B. Doumandji-Mitiche, 2004. Biosystématique des Culicidae (Diptera : Nematocera) dans
la partie sub-urbaine de l’Algérois, du marais de Réghaïa et de l’Oued Sébaou de Tizi Ouzou. 2ème Jour.
Protec. Végét., Alger : Inst. Nati. Agro., p: 45.
[40] Lounaci, Z., S. Doumandji, B. Doumandji-Mitiche and F.Z. Berrouane, 2014. Dipterans biodiversity of
agricultural and medico veterinary interest in the marsh of Reghaia (Algeria). International Journal of
Zoology and Research (IJZR), 4: 71-82.
[41] Macan, T., 1981. Modifications of pupulations of aquatic invertebrates and the quality of the water.r Ed.
Gauthier - Villars, Paris p: 275.
[42] MATEL, C., 2008. Etude relative à la délimitation et la caractérisation des zones de montagne et des massifs
montagneux. Massifs montagneux du Djurdjura. Phasen°2 .Analyse prospective de l’état des lieux du massif,
p: 173.
[43] Medlock, J.M., K.M. Hansford, F. Schaffner, V.Versteirt, G. Hendrickx, H. Zeller and W. Van Bortel, 2012.
A review of the invasive mosquitoes in Europe: Ecology, public health risks, and control options. Vector
Borne Zoonotic Diseases, 12: 435-447.
[44] Messai, N., S. Berchi, F. Boulknafed and K. Louadi, 2010. Inventaire systématique et diversité biologique
de Culicidae (Diptera: Nematocera) dans la région de Mila (Algérie). Faunistic Entomology, 63(3): 203- 206.
[45] Metge, G., 1986. Etude des écosystèmes hydromorphes (daya, merja) de la Meseta occidentale marocaine.
Typologie et système cartographique à objectif sanitaire appliquée aux populations d’Anopheles labranchiae
(Falleroni, 1926) Diptera, Culicidae, Anophelina. Thèse de doctorat 3e Cycle, Univ. d’Aix-Marseille III.
[46] Metge, G., 1991. Contribution à l’étude écologique d’Anopheles labranchiae au Maroc: Activité des imagos
et Dynamique des stades pré-imaginaux dans la région de Sidi- Bettache. Bull. Ecol., 22, 3- 4: 419- 426.
[47] Metge, G. and K. Hassaine, 1998. Study of environmental factors associated with oviposition by Aedes
caspius et Aedes detritus along a transect in Algeria. Journal of the American Mosquito Control Association,
14(3): 283-288.
[48] Papierok, B., H. Crosset et J.A. Rioux, 1975. Estimation de l‘effet des populations larvaires d‘Aedes (O)
cataphylla Dyar, 1916 (Diptera, Culicidae).II. Méthode utilisant le coup de louche ou dipping. Cahiers
O.R.S.T.O.M., série entomologie médicale et parasitologie, 13(1): 47-51.
Page 10
29 Lounaci Z., et al, 2016
Advances in Environmental Biology, 10(9) September 2016, Pages: 20-29
[49] Raharimalala, F.N., S. Boukraa, T. Bawin, S. Boyer and F. Francis, 2016. Molecular detection of six (endo-
) symbiotic bacteria in Belgian mosquitoes: first step towards the selection of appropriate paratransgenesis
candidates. Parasitol Res, 115: 1391-1399.
[50] Ramade, F., 2009. Eléments d’écologie -Ecologie fondamentale. Ed. Dunod, Paris, p: 689.
[51] Rehimi, N., 2004. Etude de la reproduction chez Culex pipiens pipiens. Aspects : morphologique, éthologique
et physiologique. Effets de quelques inhibiteurs du développement sur quelques paramètres biologiques.
Thèse de doctorat d’état en Sciences Naturelles, mention Biologie Animale. Université Badji Mokhtar
d’Annaba, Algérie, p: 195.
[52] Rehimi, N. and N. Soltani, 1999. Laboratory evaluation of Alsystin, a chitin synthesis inhibitor, against Culex
pipiens pipiens (Diptera: Culicidae): effects on development and cuticle secretion. J. Appl. Ent., 123: 437-
441.
[53] Rioux, J.A., 1958. Les Culicides du midi méditerranéen. Encyclopédie entomologique, Lechevalier, Paris, p:
303.
[54] Rioux, J.A., H. Croset, G. Gras, B. Juminer and G. Tesson, 1965. Les problèmes théoriques posés pour la
lutte contre Culex pipiens dans le sud de la France. Arch. Inst. Pasteur Tunisie, 42: 473-501.
[55] Ruybal, J.E., L.D. Kramer, A.M. Kilpatrick, 2016. Geographic variation in the response of Culex pipiens life
history traits to temperature, Parasites & Vectors, 9: 116.
[56] Schaffner, F., G. Angel, B. Geoffroy, J.P. Hervy, A. Rhaiem, J. Brunhes, 2001. The mosquitoes of Europe.
An identification and training programme. IRD Éditions & EID Méditerrannée, 1 CD ROM (Didactiques),
Montpellier, France.
[57] Senevet, G. and L. Andarelli, 1956. Les Anopheles de l’Afrique du Nord et du bassin méditerranéen.
Encyclopédie Entomologique, Lechevalier, Paris, p: 280.
[58] Senevet, G. and L. Andarelli, 1959. Les moustiques du Tassili des Ajjer. Arch. Inst. Pasteur Algérie, 37 (4):
598-602.
[59] Senevet, G. and L. Andarelli, 1960. Contribution à l’étude de la biologie des moustiques en Algérie. Arch.
Inst. Pasteur Algérie, 38(2): 306-326.
[60] Silver, J.B., 2008. Mosquito Ecology: Field Sampling Methods, 3rd edition. Springer, Dordrecht.
[61] Sinegre, G., 1974. Contribution à l’étude physiologique d’Aedes (O) caspius (Pallas 1771) (Nematocera :
Culicidae). Eclosion. Dormance. Développement. Fertilité. Thèse Doct. es- Science. Univ. Sci. Tech.
Languedoc, Montpellier.
[62] Subra, R., 1971. Etude écologique sur Culex pipiens fatigans Wiedmann, 1828 (Diptera, Culicidae) dans une
zone urbaine de Savane Soudanienne ouest- africaine. Dynamique des populations préimaginales. Cahier
O.R.T.O.M., Ser. Ent. Med. et Parasitol., 9: 73-102.
[63] Subra, R., 1973. Etudes écologiques sur Culex pipiens fatigansWiedmann, 1828 (Diptera, Culicidae) dansune
zone urbaine de savane soudanienne ouest-africaine. Dynamique des populations imaginales. Cah.
ORSTOM, ent. méd. et parasitol., 11: 79-100.
[64] Trari, B., M. Dakki, O. Himmi and M.A. El Agbani, 2002. Les moustiques (Diptera Culicidae) du Maroc.
Revue bibliographique (1916 – 2001) et inventaire des espèces. Bull. Soc. Pathol. Exot., 95: 329-334.
[65] Vaux, G.C. and J.M. Medlock, 2015. Current status of invasive mosquito surveillance in the UK. Vaux and
Medlock Parasites & Vectors, 8: 351.
[66] Versteirt, V., S. Boyer, D. Damiens, E.M. De Clercq, W. Dekoninck, E. Ducheyne, P. Grootaert, C. Garros,
T. Hance, G. Hendrickx, M. Coosemans, W. Van Bortel, 2013. Nationwide inventory of mosquito
biodiversity (Diptera: Culicidae) in Belgium, Europeonwide inventory of mosquito biodiversity (Diptera:
Culicidae) in Belgium, Europe. Bull. Entomol. Res., 103(2): 193-203.
[67] Wagner, S., A. Mathis, 2016. Laboratory colonisation of Aedes geniculatus. Journal of the European
Mosquito Control Association, 34: 1-4.
[68] White, G., C. Ottendorfer, S. Graham, T.R. Unnasch, 2011. Competency of reptilesand amphibians for
eastern equine encephalitis virus. Am. J. Trop. Med. Hyg., 85 : 421-425.






















![File PDF - BioEcology Srl · 2019-05-24 · 1rph frpphufldoh 62/)$& 75,2 &rglfh surgrwwr 893 8vl lghqwlilfdwl shuwlqhqwl ghood vrvwdq]d r ghood plvfhod h xvl vfrqvljoldwl 8vr ,qvhwwlflgd](https://static.documents.pub/doc/80x56/5e68ba28c6e565363b42ef18/file-pdf-bioecology-srl-2019-05-24-1rph-frpphufldoh-62-752-rglfh.jpg)

