46
Biosynthesis of Nucleotides Biosynthesis of Nucleotides

Biosynthesis of Nucleotides
Biosynthesis of Nucleotides

Nucleotides actively participate in many biochemical reactions:
Nucleotides actively participate in many biochemical reactions:
• ATP and GTP as energy sources• ATP and GTP as energy sources
• Uridine derivatives of sugars participate in carbohydrate metabolism
• Uridine derivatives of sugars participate in carbohydrate metabolism
• Coenzymes (NAD, FAD, CoA) are nucleotide derivatives
• Coenzymes (NAD, FAD, CoA) are nucleotide derivatives
• [ATP], [ADP], [AMP] act as allosteric regulators of key enzymes
• [ATP], [ADP], [AMP] act as allosteric regulators of key enzymes
• Monomeric units of nucleic acids• Monomeric units of nucleic acids

Two Pathways for Nucleotide Biosynthesis:Two Pathways for Nucleotide Biosynthesis:
de novo pathway (anew; from scratch): nucleotides are constructed from simple precursors
de novo pathway (anew; from scratch): nucleotides are constructed from simple precursors
salvage pathways: recovery and recycling of nucleotides obtained in the dietsalvage pathways: recovery and recycling of nucleotides obtained in the diet

De Novo Biosynthesis of Purines:De Novo Biosynthesis of Purines:
Studied first in pigeons. Birds excrete nitrogen as the purine uric acid:Studied first in pigeons. Birds excrete nitrogen as the purine uric acid:
NNHH
OONN
NN
NNHH
HH
OO
OO
HH
Pigeons were fed isotopically labeled compounds and the distribution of labeled atoms examined in uric acid.
Pigeons were fed isotopically labeled compounds and the distribution of labeled atoms examined in uric acid.
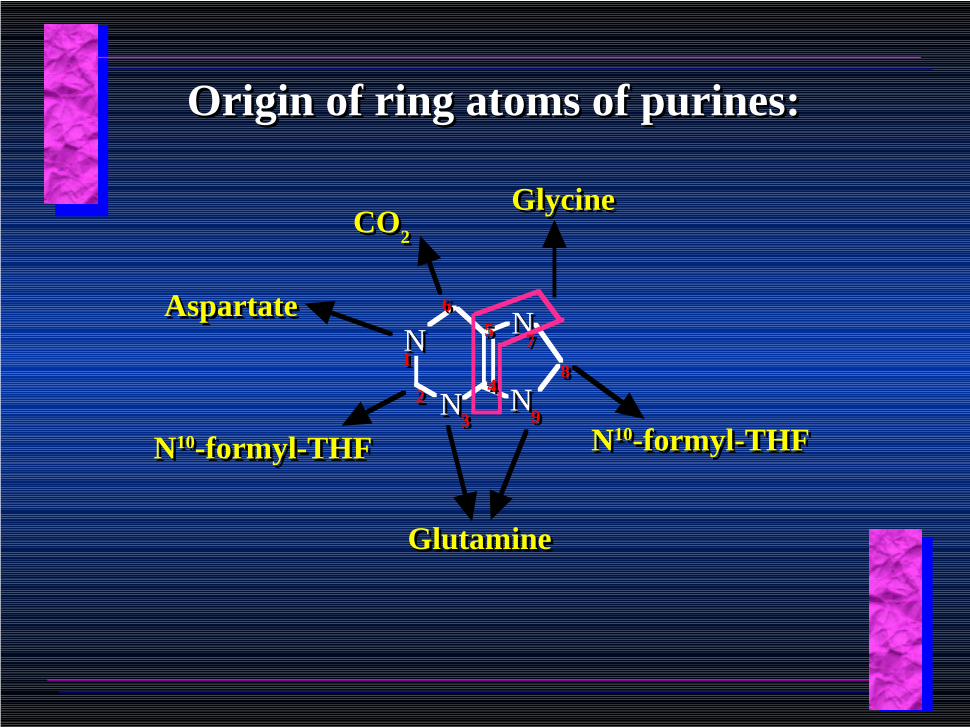
NN
NN
NN
NN
GlycineGlycine
GlutamineGlutamine
N10-formyl-THFN10-formyl-THF
CO2CO2
AspartateAspartate
N10-formyl-THFN10-formyl-THF
11
99
88
66
55
44
3322
77
Origin of ring atoms of purines:Origin of ring atoms of purines:

Atoms forming the purine ring are successively added to ribose-5-P.Atoms forming the purine ring are successively added to ribose-5-P.
Purines are thus directly synthesized as nucleotide derivatives by assembling the atoms comprising the purine ring directly on ribose.
Purines are thus directly synthesized as nucleotide derivatives by assembling the atoms comprising the purine ring directly on ribose.

Phosphoribosylpyrophosphate (PRPP) is formed from ribose-5-P and ATP by PRPP synthetase. PRPP is the donor of the ribose ring of the nucleotides.
Phosphoribosylpyrophosphate (PRPP) is formed from ribose-5-P and ATP by PRPP synthetase. PRPP is the donor of the ribose ring of the nucleotides.
PRPPPRPP
PRPP also participates in pyrimidine biosynthesis and in the synthesis of histidine and tryptophan.
PRPP also participates in pyrimidine biosynthesis and in the synthesis of histidine and tryptophan.
NH2NH2
NHNHO=CO=C
CH2CH2
NH2NH2
CHCH
OO
NHNHHN=CHN=C
CH2CH2
NHNHCHCH
OO
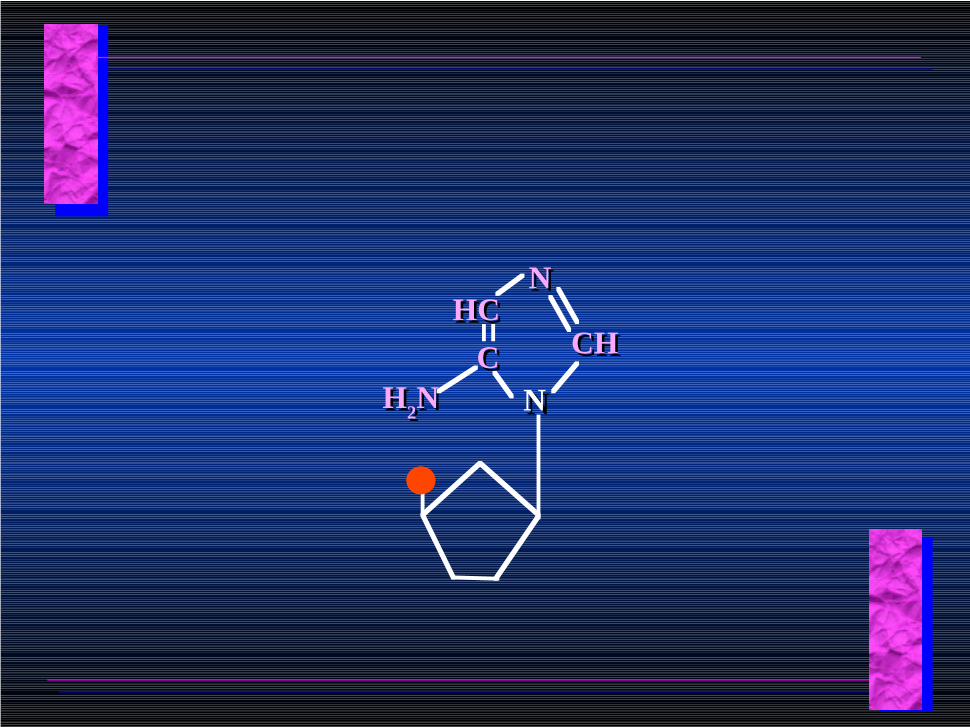
NNCC
HCHCNN
CHCH
H2NH2N
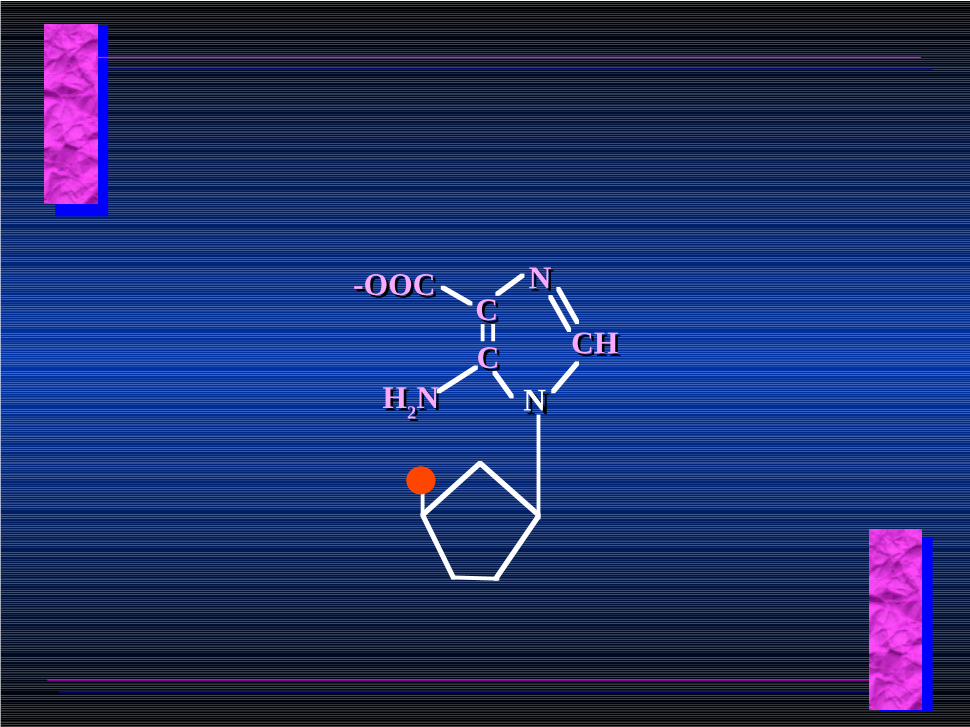
NNCCCC
NN
CHCH
H2NH2N
-OOC-OOC
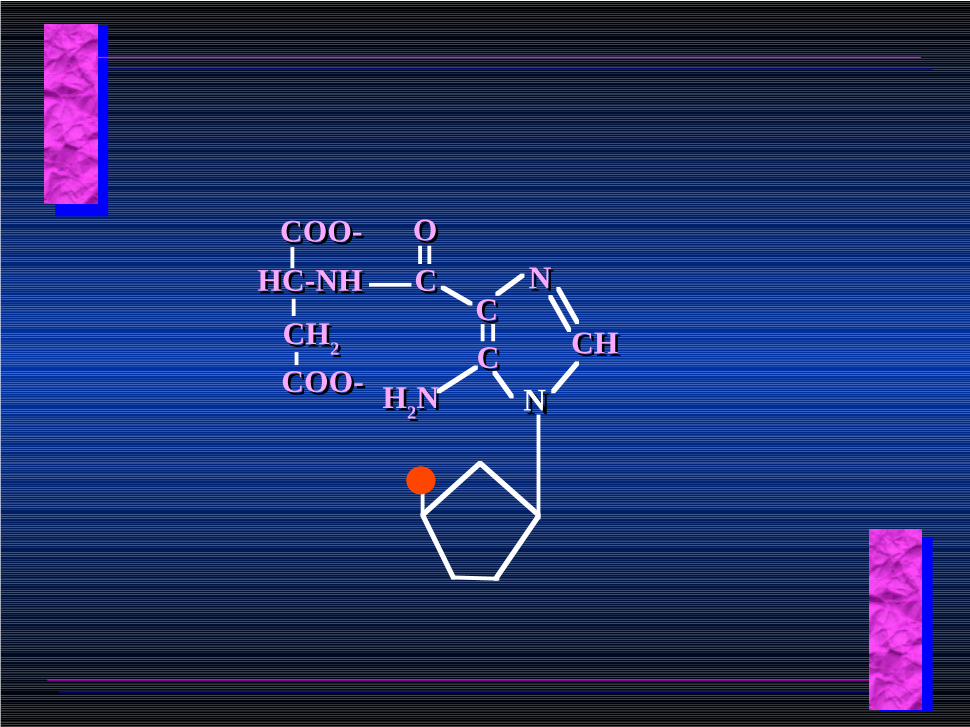
NNCCCC
NN
CHCH
H2NH2N
CC
OO
HC-NHHC-NH
CH2CH2
COO-COO-
COO-COO-
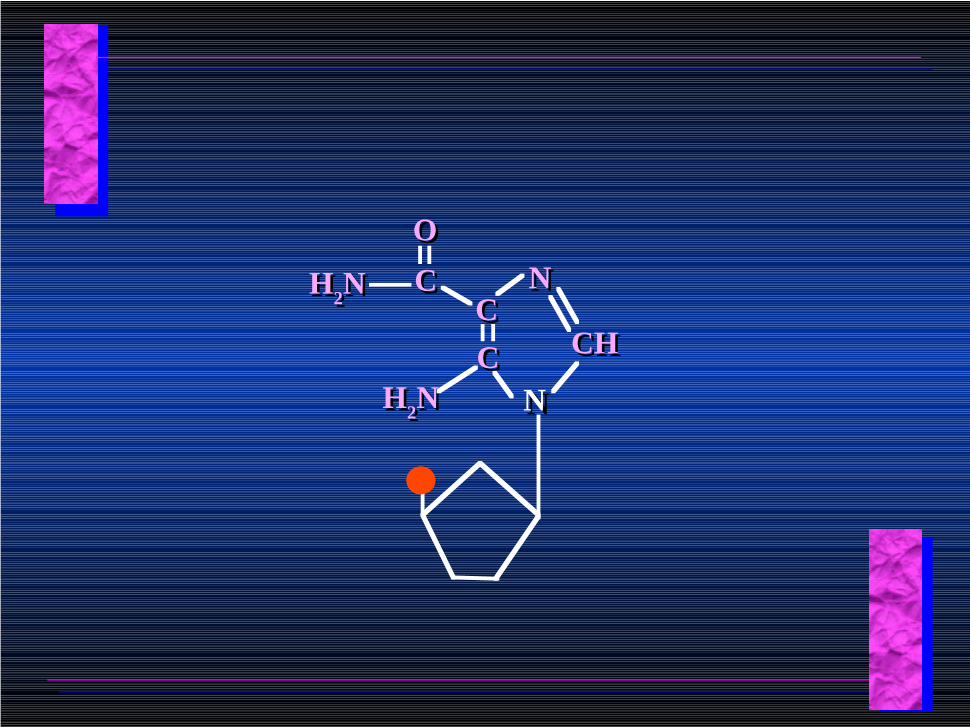
NNCCCC
NN
CHCH
H2NH2N
CC
OO
H2NH2N
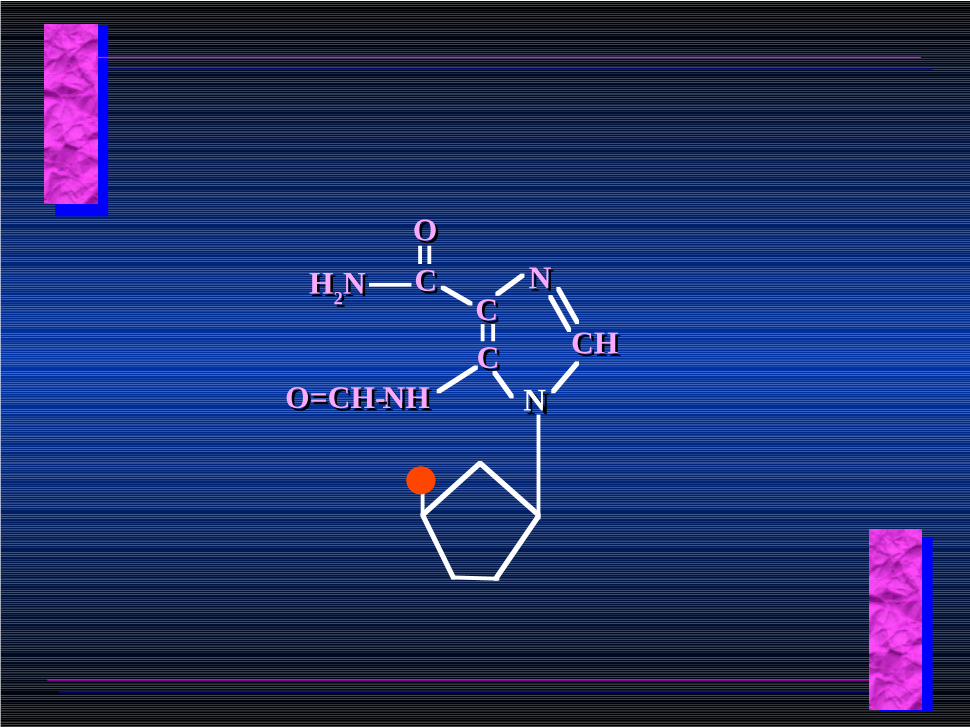
NNCCCC
NN
CHCH
NHNH
CC
OO
H2NH2N
O=CH-O=CH-
NNCCCC
NN
CHCH
NN
CC
OO
HNHN
HCHC
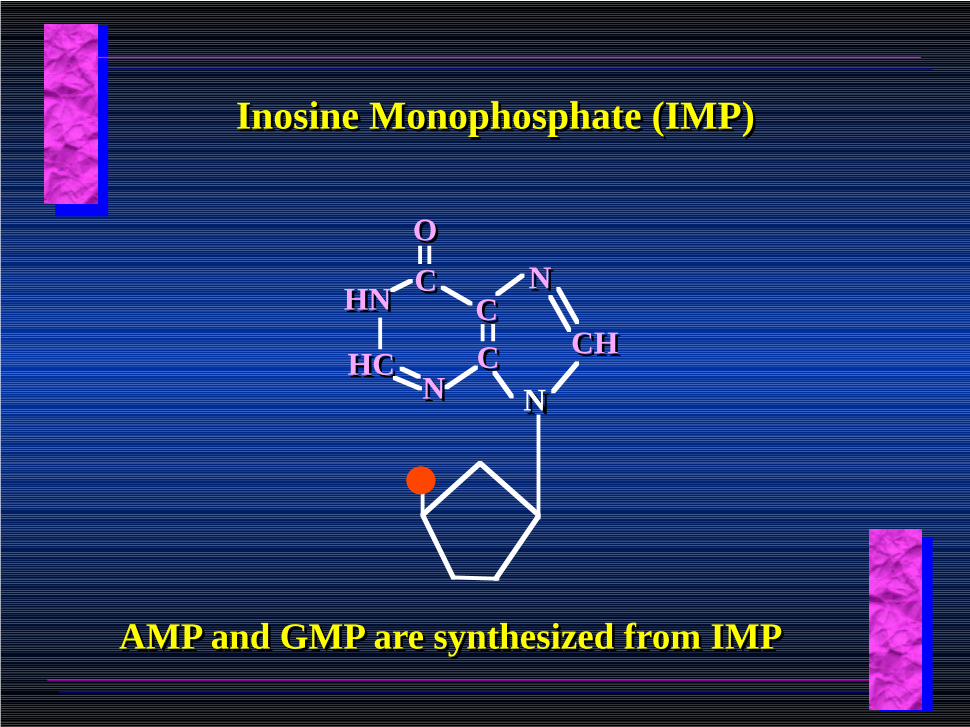
Inosine Monophosphate (IMP)Inosine Monophosphate (IMP)
AMP and GMP are synthesized from IMPAMP and GMP are synthesized from IMP
NNCCCC
NN
CHCH
NN
CC
OO
HNHN
HCHC
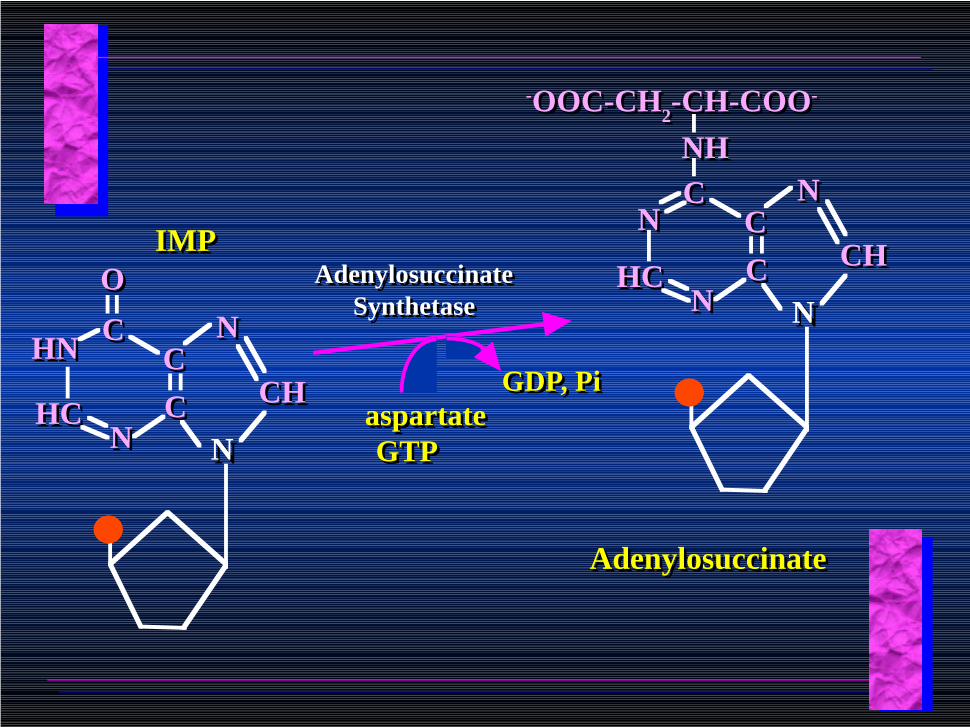
IMPIMP
aspartateGTP
aspartateGTP
Adenylosuccinate Synthetase
Adenylosuccinate Synthetase
GDP, PiGDP, Pi
NNCCCC
NN
CHCH
NN
CCNN
HCHC
NHNH
-OOC-CH2-CH-COO--OOC-CH2-CH-COO-
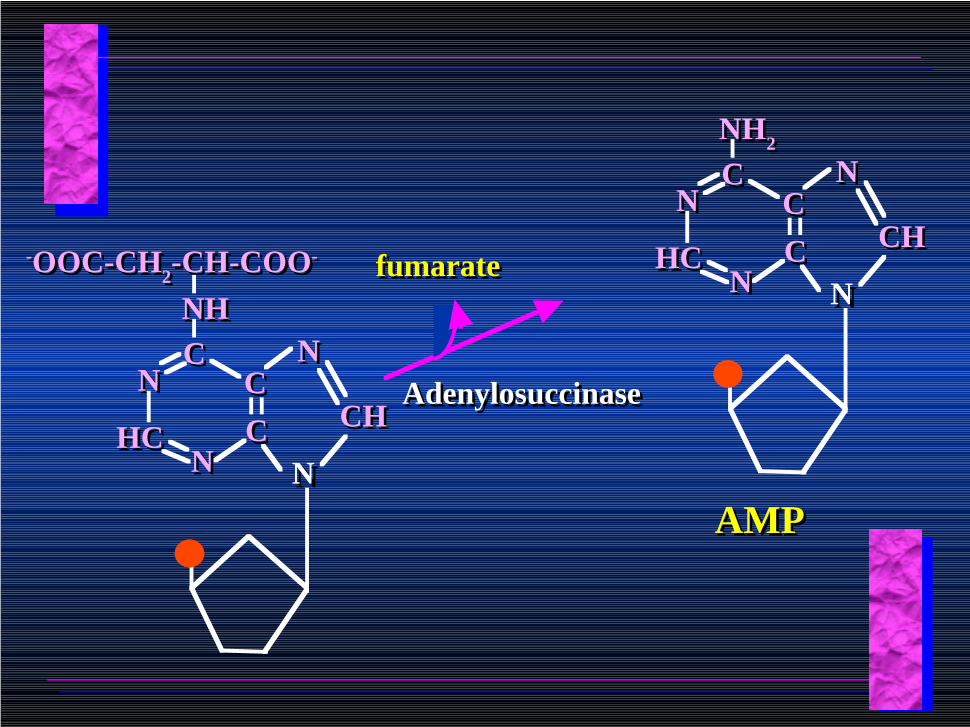
AdenylosuccinateAdenylosuccinate
NNCCCC
NN
CHCH
NN
CCNN
HCHC
NHNH
-OOC-CH2-CH-COO--OOC-CH2-CH-COO- fumaratefumarate
AdenylosuccinaseAdenylosuccinase
NNCCCC
NN
CHCH
NN
CCNN
HCHC
NH2NH2
AMPAMP
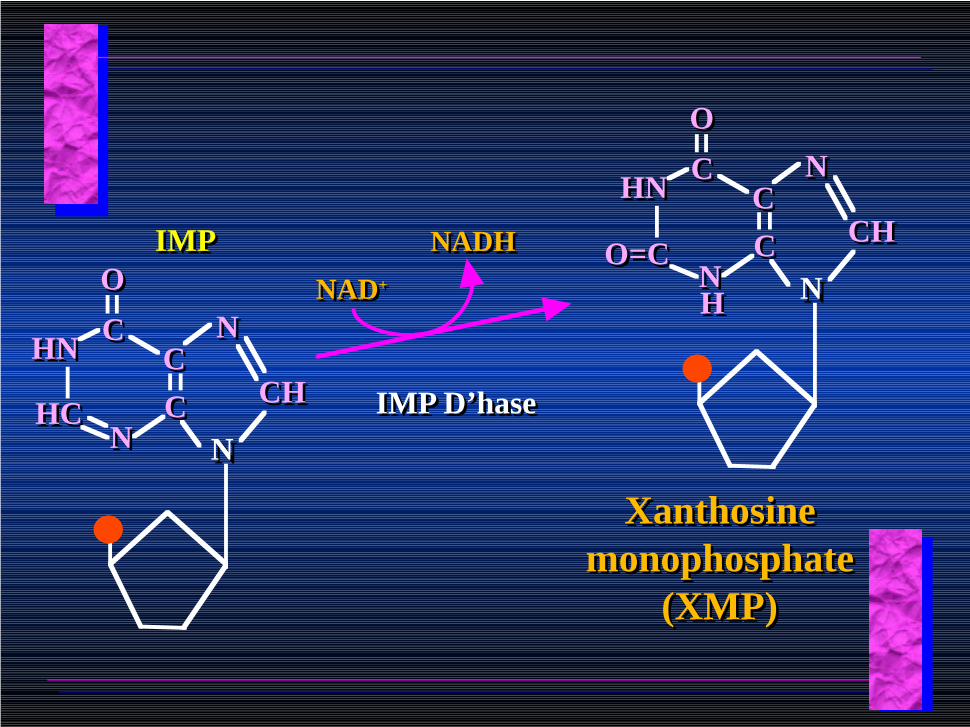
NNCCCC
NN
CHCH
NN
CC
OO
HNHN
HCHC
IMPIMP
NAD+NAD+
IMP D’haseIMP D’hase
NADHNADH
NNCCCC
NN
CHCH
NN
CC
OO
HNHN
O=CO=C
HH
Xanthosine monophosphate
(XMP)
Xanthosine monophosphate
(XMP)
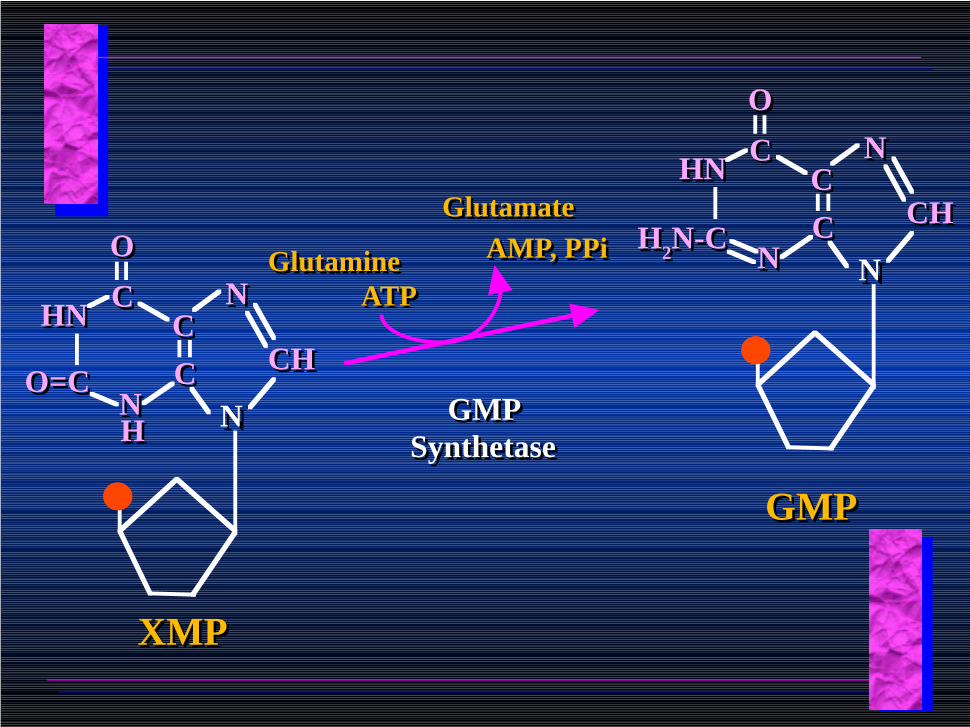
NNCCCC
NN
CHCH
NN
CC
OO
HNHN
O=CO=C
HH
XMPXMP
GMP Synthetase
GMP Synthetase
ATPATP
AMP, PPiAMP, PPiGlutamineGlutamine
GlutamateGlutamate
NNCCCC
NN
CHCH
NN
CC
OO
HNHN
H2N-CH2N-C
GMPGMP

Regulation of Purine Biosynthesis:Regulation of Purine Biosynthesis:
• PRPP synthetase is feedback inhibited by AMP, GMP and IMP.
• PRPP synthetase is feedback inhibited by AMP, GMP and IMP.
• Adenylosuccinate synthetase is inhibted by AMP.
• Adenylosuccinate synthetase is inhibted by AMP.
• IMP d’hase is inhibited by XMP and GMP.• IMP d’hase is inhibited by XMP and GMP.

Purine Salvage:Purine Salvage:
• During cellular metabolism and during digestion in animals, nucleic acids are degraded to mononucleotides, nucleosides, and free purine bases.
• During cellular metabolism and during digestion in animals, nucleic acids are degraded to mononucleotides, nucleosides, and free purine bases.
• Some purines are further degraded to uric acid, but a considerable fraction are directly converted back to purine ribonucleotides
• Some purines are further degraded to uric acid, but a considerable fraction are directly converted back to purine ribonucleotides

AMPAMP
AdenosineAdenosine
AdenineAdeninePRPPPRPP
PPiPPi
Adenine phosphoribosyl transferase
Adenine phosphoribosyl transferase
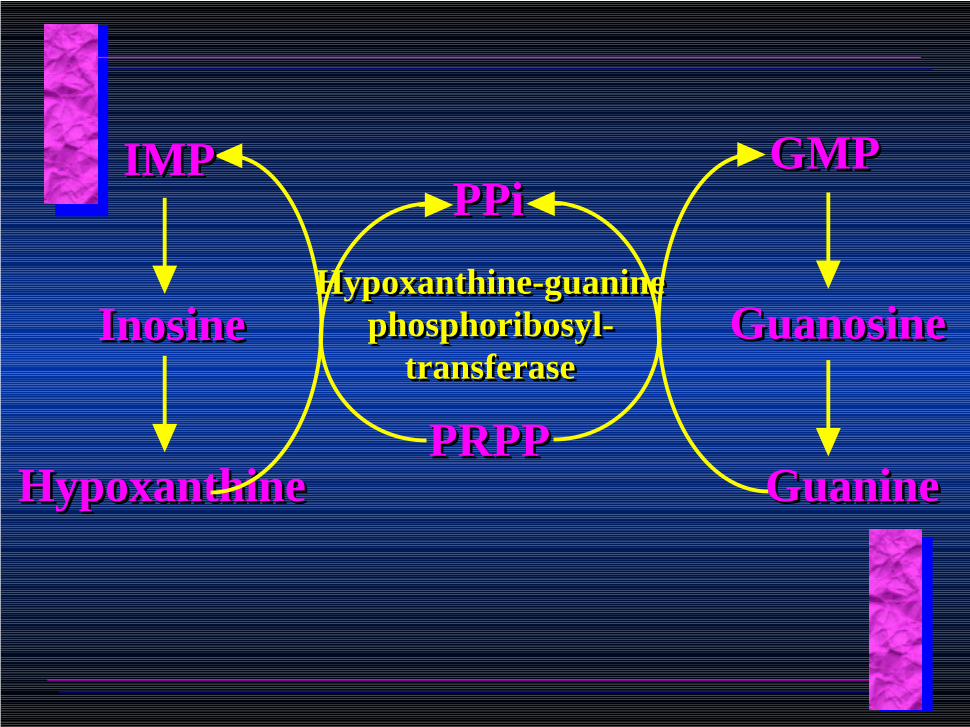
InosineInosine
HypoxanthineHypoxanthine
Hypoxanthine-guaninephosphoribosyl-
transferase
Hypoxanthine-guaninephosphoribosyl-
transferase
PRPPPRPP
PPiPPiGMPGMP
GuanosineGuanosine
GuanineGuanine
IMPIMP

Lesch-Nyhan Syndrome:Lesch-Nyhan Syndrome:
• Described by William Nyhan and Michael Lesch in 1964.
• Described by William Nyhan and Michael Lesch in 1964.
• Hereditary deficiency of hypoxanthine-guanine phosphoribosyltransferase. Disease affects mostly males.
• Hereditary deficiency of hypoxanthine-guanine phosphoribosyltransferase. Disease affects mostly males.
• Hypoxanthine and guanine are degraded to uric acid instead of being converted to IMP and GMP.
• Hypoxanthine and guanine are degraded to uric acid instead of being converted to IMP and GMP.
• Symptoms: mental retardation; spasticity; bizarre tendency to self-mutilate.
• Symptoms: mental retardation; spasticity; bizarre tendency to self-mutilate.

Pyrimidine BiosynthesisPyrimidine Biosynthesis

• The common pyrimidine ribonucleotides are cytidine-5’-monophosphate and uridine-5’-monophosphate
• The common pyrimidine ribonucleotides are cytidine-5’-monophosphate and uridine-5’-monophosphate
• The pyrimidine ring is synthesized first, then attached to ribose-5-phosphate
• The pyrimidine ring is synthesized first, then attached to ribose-5-phosphate
• Pyrimidine nucleotides are made from aspartate, PRPP and carbamoyl phosphate
• Pyrimidine nucleotides are made from aspartate, PRPP and carbamoyl phosphate
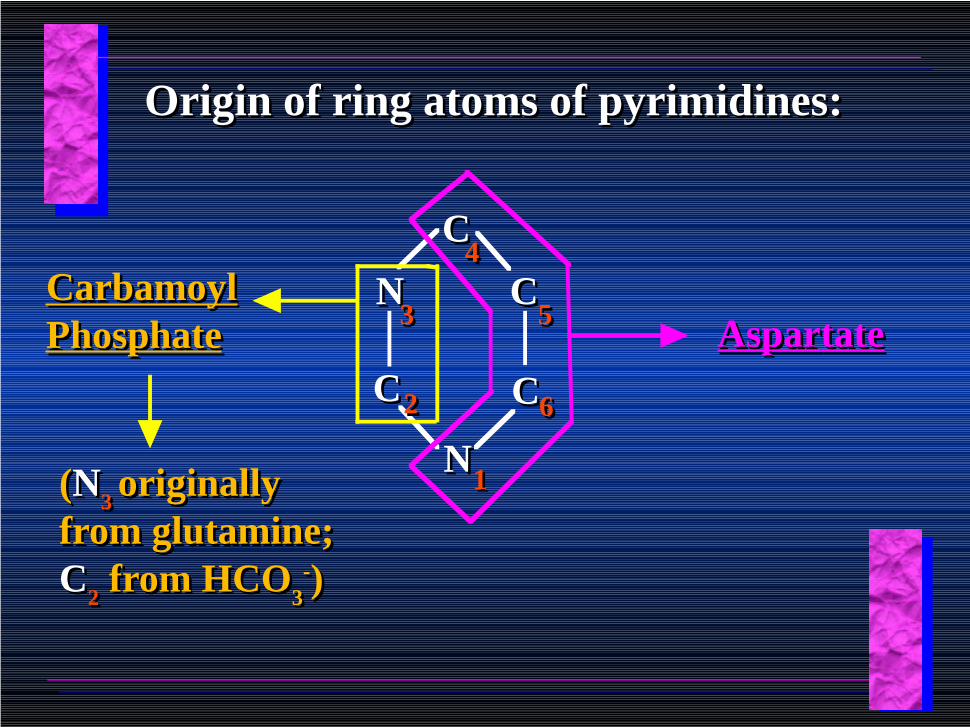
Origin of ring atoms of pyrimidines:Origin of ring atoms of pyrimidines:
CC
NN
CC
CCCC
NN
11
33
22
44
66
55 AspartateAspartateCarbamoyl PhosphateCarbamoyl Phosphate
(N3 originally
from glutamine; C
2 from HCO
3-)
(N3 originally
from glutamine; C
2 from HCO
3-)
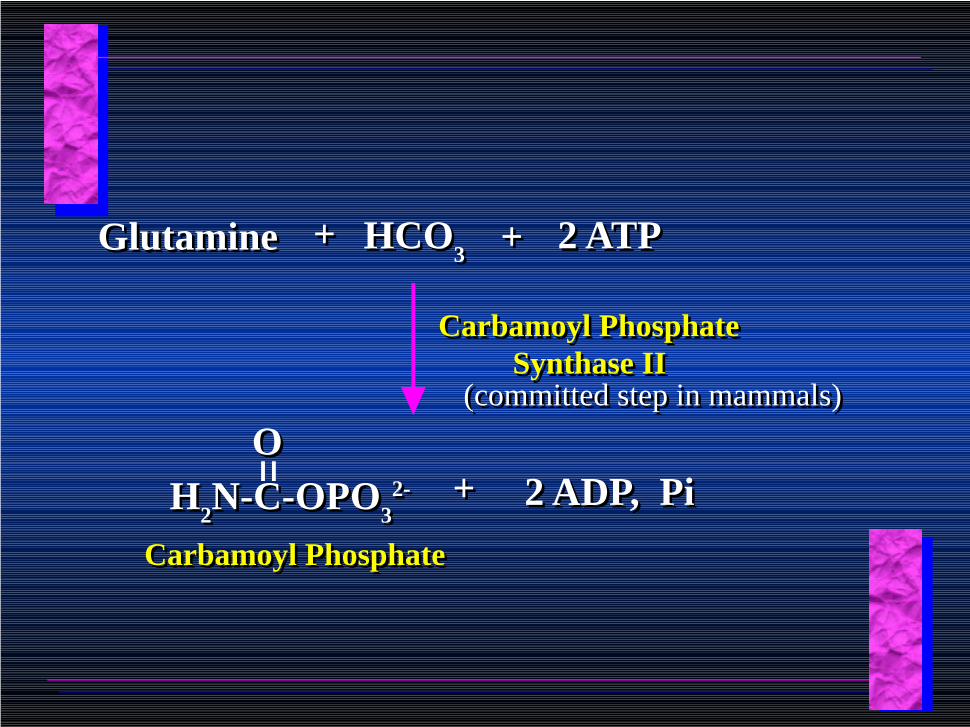
GlutamineGlutamine ++ HCO3
HCO3 ++ 2 ATP2 ATP
Carbamoyl Phosphate Synthase II
Carbamoyl Phosphate Synthase II
(committed step in mammals)(committed step in mammals)
H2N-C-OPO
32-H
2N-C-OPO
32-
OO++ 2 ADP, Pi2 ADP, Pi
Carbamoyl PhosphateCarbamoyl Phosphate
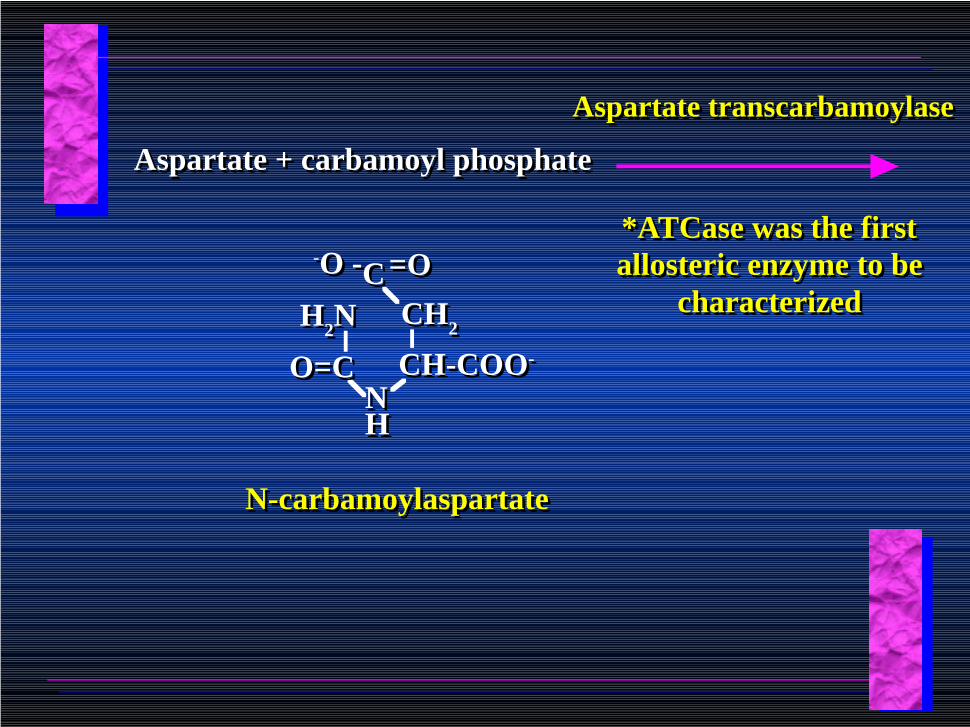
Aspartate + carbamoyl phosphateAspartate + carbamoyl phosphate
Aspartate transcarbamoylaseAspartate transcarbamoylase
CC =O=O-O --O -
H2NH2N
CH-COO-CH-COO-
CH2CH2
NNHH
O=CO=C
N-carbamoylaspartateN-carbamoylaspartate
*ATCase was the first allosteric enzyme to be
characterized
*ATCase was the first allosteric enzyme to be
characterized
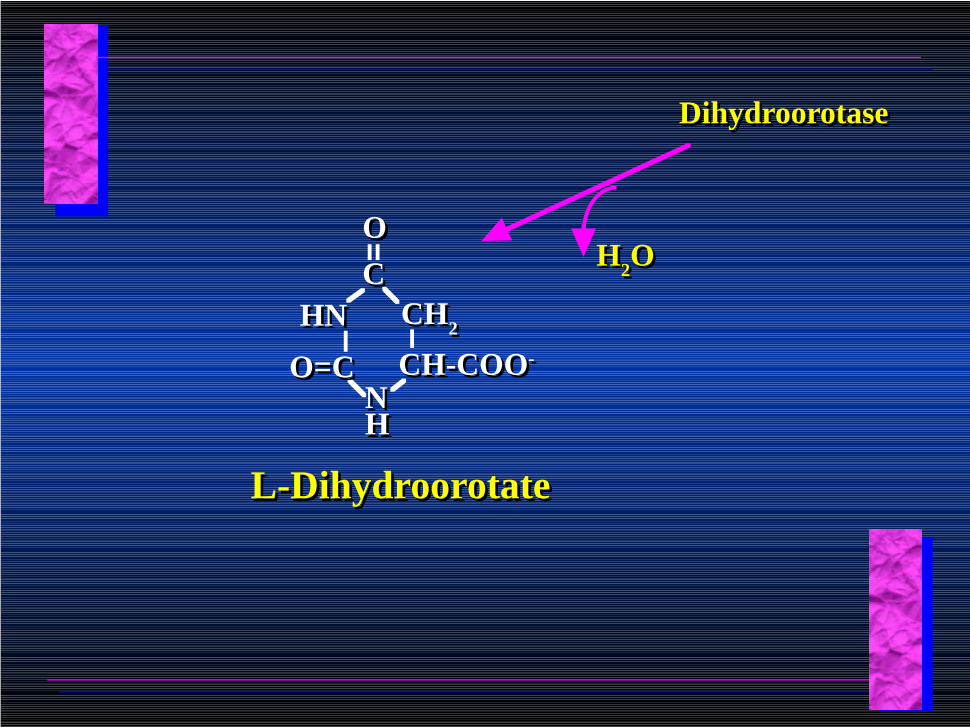
DihydroorotaseDihydroorotase
H2OH2O
CH-COO-CH-COO-
CCOO
HNHN CH2CH2
NNHH
O=CO=C
L-DihydroorotateL-Dihydroorotate
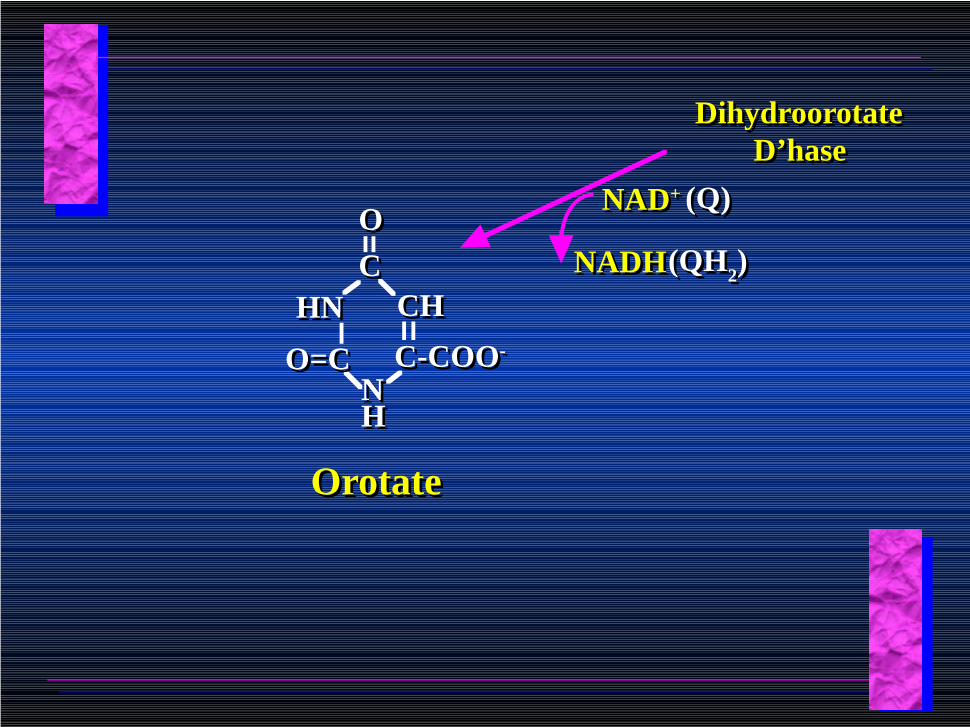
Dihydroorotate D’hase
Dihydroorotate D’hase
NADHNADH
NAD+NAD+ (Q)(Q)
(QH2)(QH2)
C-COO-C-COO-
CCOO
HNHN CHCH
NNHH
O=CO=C
OrotateOrotate
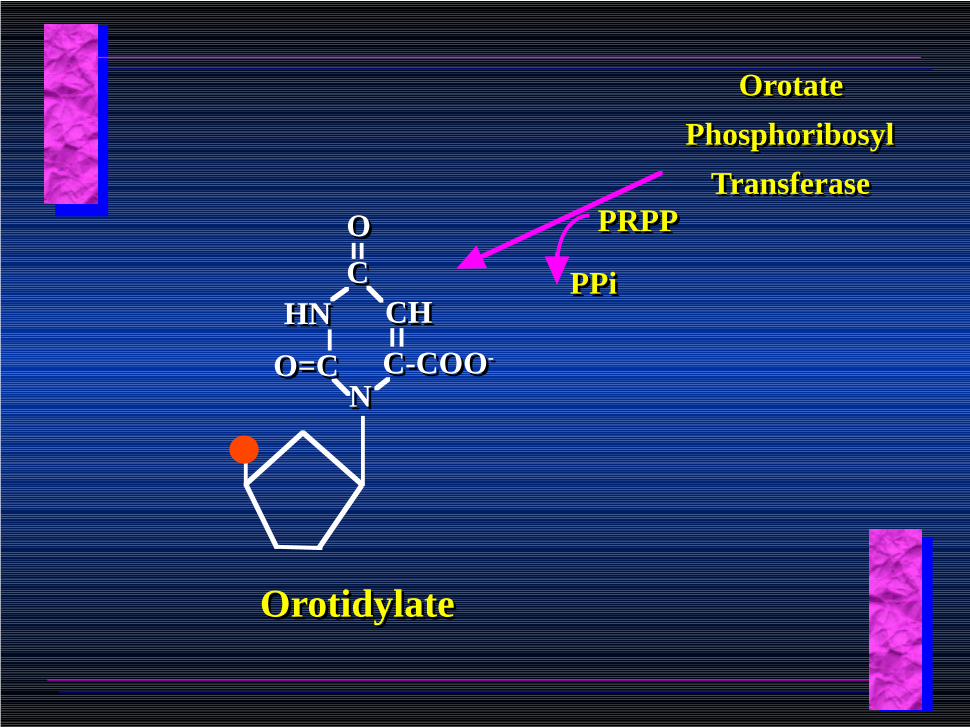
Orotate
Phosphoribosyl
Transferase
Orotate
Phosphoribosyl
Transferase
PPiPPi
PRPPPRPP
C-COO-C-COO-
CCOO
HNHN CHCH
NNO=CO=C
OrotidylateOrotidylate
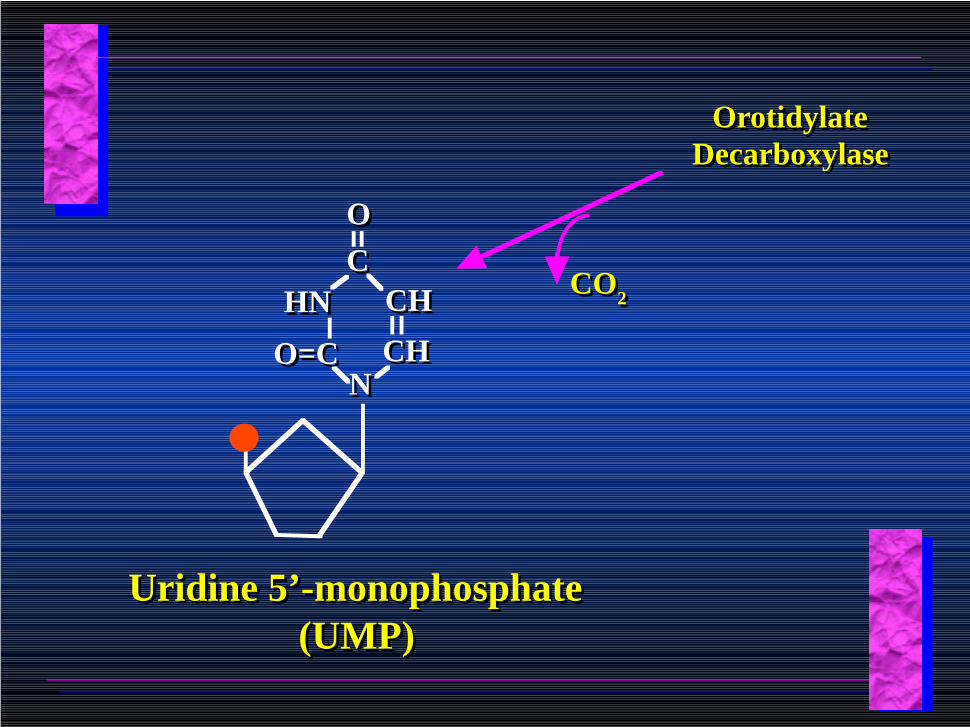
Orotidylate Decarboxylase
Orotidylate Decarboxylase
CO2CO2
CHCH
CCOO
HNHN CHCH
NNO=CO=C
Uridine 5’-monophosphate (UMP)
Uridine 5’-monophosphate (UMP)
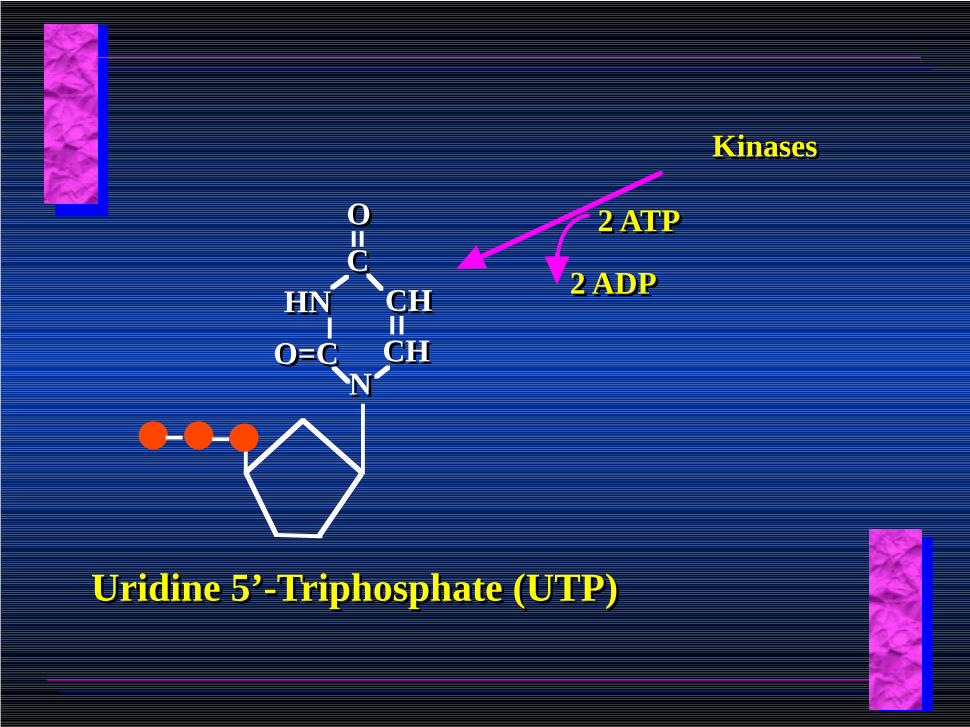
KinasesKinases
2 ADP2 ADP
2 ATP2 ATP
CHCH
CCOO
HNHN CHCH
NNO=CO=C
Uridine 5’-Triphosphate (UTP)Uridine 5’-Triphosphate (UTP)
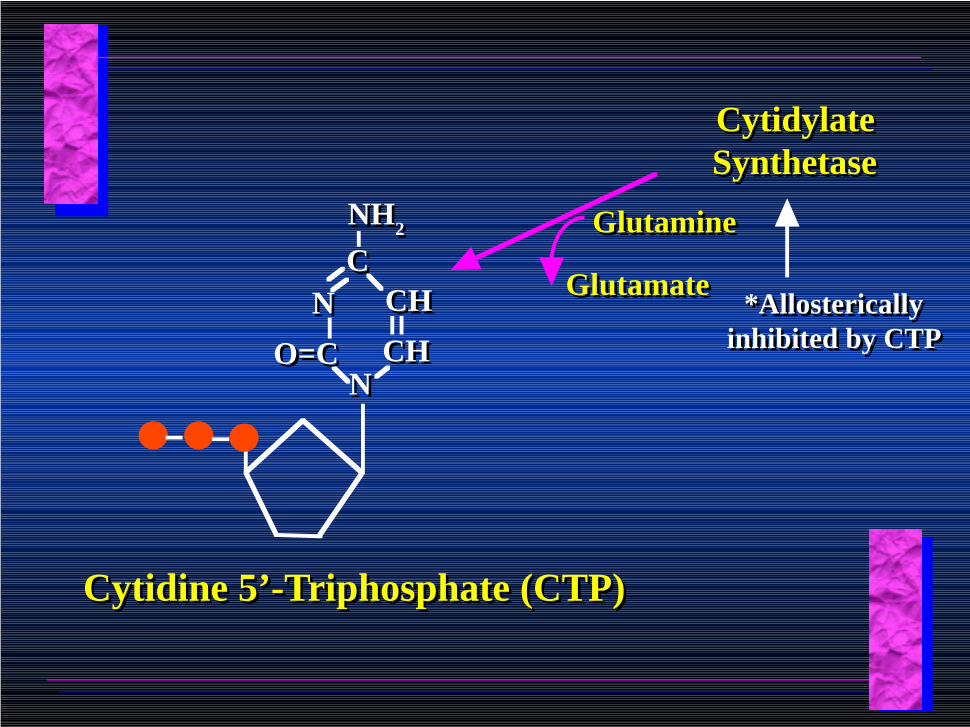
Cytidylate SynthetaseCytidylate Synthetase
*Allosterically inhibited by CTP
*Allosterically inhibited by CTP
GlutamateGlutamate
GlutamineGlutamine
CHCH
CCNH2NH2
NN CHCH
NNO=CO=C
Cytidine 5’-Triphosphate (CTP)Cytidine 5’-Triphosphate (CTP)

Mammalian pyrimidine synthesis is an example of metabolite channeling.Mammalian pyrimidine synthesis is an example of metabolite channeling.
In bacteria, the six enzymes of de novo pyrimidine synthesis are separate proteins.In bacteria, the six enzymes of de novo pyrimidine synthesis are separate proteins.
In mammals, the six activities are contained within three proteins. In mammals, the six activities are contained within three proteins.
CPS-II, asparate transcarbamoylase, dihydroorotaseare all contained within a single cytosolic protein. DHO d’hase is localized in the inner mito. membrane. Orotate phosphoribosyltransferase and OMP decarboxylase are contained with a single protein called OMP synthase.
CPS-II, asparate transcarbamoylase, dihydroorotaseare all contained within a single cytosolic protein. DHO d’hase is localized in the inner mito. membrane. Orotate phosphoribosyltransferase and OMP decarboxylase are contained with a single protein called OMP synthase.

Regulation of Pyrimidine Biosynthesis:Regulation of Pyrimidine Biosynthesis:
• Carbamoyl phosphate synthetase II is allosterically activated by PRPP and ATP. Pyrimidine nucleotides (UDP, UTP) inhibit.
• Carbamoyl phosphate synthetase II is allosterically activated by PRPP and ATP. Pyrimidine nucleotides (UDP, UTP) inhibit.
• Aspartate transcarbamoylase (ATCase) from E. coli is inhibited by pyrimidine nucleotides (CTP and UTP). ATP is an allosteric activator.
• Aspartate transcarbamoylase (ATCase) from E. coli is inhibited by pyrimidine nucleotides (CTP and UTP). ATP is an allosteric activator.

• Deoxyribonucleotides are synthesized by reduction of ribonucleosides
• Deoxyribonucleotides are synthesized by reduction of ribonucleosides
• All 4 ribonucleoside diphosphates (ADP, GDP, CDP, UDP) are substrates for Ribonucleotide Reductase
• All 4 ribonucleoside diphosphates (ADP, GDP, CDP, UDP) are substrates for Ribonucleotide Reductase
• Ribonucleotide Reductase has both a catalytic site and two allosteric sites. One allosteric site (Activity site) controls activity at the catalytic site. The second (Specificity site) determines which nucleoside diphosphate binds the active site.
• Ribonucleotide Reductase has both a catalytic site and two allosteric sites. One allosteric site (Activity site) controls activity at the catalytic site. The second (Specificity site) determines which nucleoside diphosphate binds the active site.

Ligand bound to Activity SiteLigand bound to Activity Site
Ligand bound to Specificity SiteLigand bound to Specificity Site
Activity of Catalytic SiteActivity of Catalytic Site
dATPdATP Enzyme InactiveEnzyme Inactive
ATPATP ATP or dATPATP or dATP CDP or UDPCDP or UDP
ATPATP dTTPdTTP GDPGDP
ATPATP dGTPdGTP ADPADP

dTMP is formed from dUMP:dTMP is formed from dUMP:
UMPUMP
Nucleoside Monophosphate Kinase
Nucleoside Monophosphate Kinase
dUMPdUMPUDPUDP
Nucleoside Diphosphate Kinase
Nucleoside Diphosphate Kinase
dUDPdUDP
Ribonucleotide ReductaseRibonucleotide Reductase
dUTPdUTP
dUTPasedUTPase
dTMPdTMPThymidylate SynthaseThymidylate Synthase

dTMP is also formed from dCDP:dTMP is also formed from dCDP:
Cytidine
deaminase
Cytidine
deaminase
dCDPdCDP dCMPdCMP dUMPdUMP
(activated by dCTP
inhibited by dTTP)
(activated by dCTP
inhibited by dTTP)
Of the 4 dNTPs, only dCTP does not interact with the regulatory sites on ribonucleotide reductase, instead it interacts with dCMP deaminase.
Of the 4 dNTPs, only dCTP does not interact with the regulatory sites on ribonucleotide reductase, instead it interacts with dCMP deaminase.
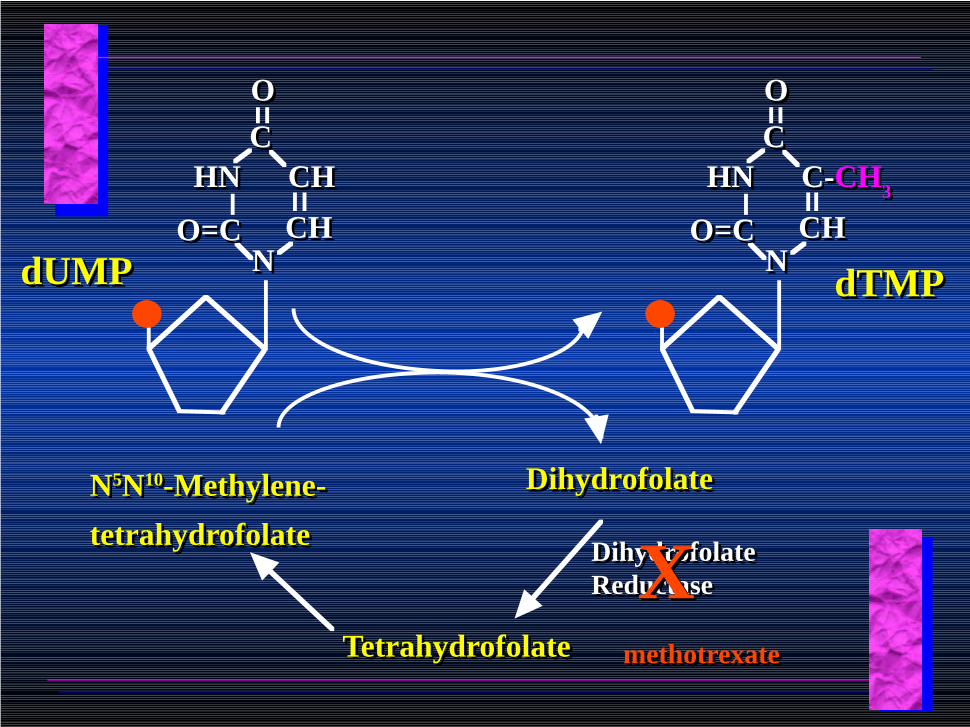
dUMPdUMPCHCH
CCOO
HNHN CHCH
NNO=CO=C
N5N10-Methylene-
tetrahydrofolate
N5N10-Methylene-
tetrahydrofolate
DihydrofolateDihydrofolate
TetrahydrofolateTetrahydrofolate
CHCH
CCOO
HNHN C-CH3C-CH3
NNO=CO=C
dTMPdTMP
Dihydrofolate ReductaseDihydrofolate Reductase
methotrexatemethotrexate
XX

dTMP can also be synthesized via salvage of thymidine:dTMP can also be synthesized via salvage of thymidine:
Thymidine Thymidine Thymidine
kinaseThymidine
kinase
ATPATP ADPADPdTMPdTMP
Radioactive thymidine is used for monitoring intracellular synthesis of DNA because it enters cells easily and its principle metabolic fate is salvage leading to incorporation into DNA.
Radioactive thymidine is used for monitoring intracellular synthesis of DNA because it enters cells easily and its principle metabolic fate is salvage leading to incorporation into DNA.

• Many anticancer drugs target DNA synthesis; particularly thymidylate synthesis
• Many anticancer drugs target DNA synthesis; particularly thymidylate synthesis
• Methotrexate and aminopterin inhibit dihydrofolate reductase; thymidylate cannot be formed thus DNA cannot be replicated
• Methotrexate and aminopterin inhibit dihydrofolate reductase; thymidylate cannot be formed thus DNA cannot be replicated
• 5-Fluorouracil is converted to 5-Fluorodeoxy- uridylate which binds tightly to thymidylase synthase and inhibits the enzyme.
• 5-Fluorouracil is converted to 5-Fluorodeoxy- uridylate which binds tightly to thymidylase synthase and inhibits the enzyme.

• Azaserine and acivicin are glutamine analogues also used as chemotheraputic agents
• Azaserine and acivicin are glutamine analogues also used as chemotheraputic agents

















