Biotransformation of 1,1,1-Trichloroethane, Trichloromethane, andTetrachloromethane by a Clostridium sp.
RENE GALLI AND PERRY L. McCARTY*Environmental Engineering and Science, Department of Civil Engineering, StanJfrd University,
Stanford, California 943054020
Received 8 July 1988/Accepted 4 January 1989
A gram-positive, strictly anaerobic, motile, endospore-forming rod, tentatively identified as a proteolyticClostridium sp., was isolated from the effluent of an anaerobic suspended-growth bioreactor. The organism was
able to biotransform 1,1,1-trichloroethane, trichloromethane, and tetrachloromethane. 1,1,1-Trichloroethanewas completely transformed (.99.5%) by reductive dehalogenation to 1,1-dichloroethane (30 to 40%) and,presumably by other mechanisms, to acetic acid (7%) and unidentified products. The reductive dehalogenationof tetrachloromethane led to the intermediate trichloromethane, which was further transformed to dichloro-methane (8%) and unidentified products. The biotransformation occurred during the exponential growthphase, as well as during the stationary phase. Tetrachlorethene, trichloroethene, 1,1-dichloroethene, chloroet-hane, 1,1-dichloroethane, and dichloromethane were not biotransformed significantly by the organism.
Chlorinated aliphatic C1 and C, compounds are widelyused as solvents, degreasing agents, and intermediates inchemical synthesis and are commonly found as contami-nants of groundwater (27). Recent studies reporting thebiotransformation of chlorinated aliphatic compounds bymixed bacterial cultures under anaerobic conditions haveindicated the potential importance of anaerobic microorgan-isms in the transformation of halogenated compounds atcontaminated sites (2, 3, 11, 21, 22, 26). However, little isknown about the bacterial composition of mixed culturesenriched under methanogenic or denitrifying conditions, orabout the organism that may be responsible for the transfor-mations that have been observed. Often, a large variety ofanaerobic microorganisms are able to grow under givenconditions (13), and only certain ones might be capable ofcarrying out biotransformations of chlorinated aliphaticcompounds (7, 8, 16, 23).
In this report we describe the isolation and characteriza-tion of a Clostridiium sp. from a methanogenic, 1,1,1-trichlo-roethane (TCA)- and trichloroethene- transforming mixedculture which was able to dehalogenate chlorinated aliphaticcompounds.
MATERIALS AND METHODSChemicals and radiochemicals. The chlorinated aliphatic
compounds used in this study were TCA (99+%), 1,1-dichloroethane (DCA; 97%), 1,1-dichloroethene (99%), di-chloromethane (99.9%), tetrachloroethene (99.97%), andtrichloroethene (99+%) (all of which were obtained fromAldrich Chemical Co., Inc., Milwaukee, Wis.); tetrachlo-romethane (99.8%) and trichloromethane (99.5%) (both ofwhich were obtained from J. T. Baker Chemical Co., Phill-ipsburg, N.J.); and chloroethane (gas chromatographic [GC]standard, 0.2 mg/ml of methanol; Supelco Inc., Bellafonte,Pa.).["4C]TCA (Dupont, NEN Research Products, Boston,
Mass.) was diluted initially in methanol to 5.1 x 106 dpm/,ul(74.8 mg of TCA per ml of methanol).
Anaerobic facilities. Experiments were performed in ananaerobic glove box with an automatic airlock (Coy Labo-
* Corresponding author.
ratory Products Inc., Ann Arbor, Mich.) at 35°C. Theatmosphere inside the anaerobic chamber consisted of anN,-H2 gas mixture (95 and 5%, respectively), and traces ofoxygen were removed by two catalyst boxes containingpalladium-coated aluminum oxide. N2, N,-C02 (70 and 30%,respectively), and H2-C02 (70 and 30%, respectively) gasmixtures were scrubbed free of oxygen traces by passingthem through a heated copper filling at 300°C (Lindberg,Watertown, Wis.). All media were prepared inside theanaerobic chamber by using deionized water that was boiledfor 20 min under a stream of the N2-CO, gas mixture andcontained 1 mg of resazurine per liter as a redox indicator.Agar plates were incubated in an anaerobic system (Gas Pak;BBL Microbiology Systems, Cockeysville, Md.), and theGas Pak jar was stored inside the glove box.Organisms and culture conditions. Methylobacterium ther-
moautotrophicium Marburg (DSM 2133) was obtained fromL. Meile (Federal Institute of Technology, Zurich, Switzer-land) and was used as an indicator strain to test the cultiva-tion conditions for methanogenic bacteria. It was grown onmedium A at 60°C as described below and was kept in liquidculture at 4°C. Clostridiium sp. strain TCAIIB (which wasisolated in this study) was kept on agar slants containingmedium C at 35°C inside the anaerobic chamber.Medium A, which was used for the enrichment of anaer-
obic microorganisms, was essentially the same as that de-scribed by Kiener and Leisinger (17), which was a modifiedmedium of Balch et al. (1) for the cultivation of methano-genic bacteria. Sulfates were replaced with the correspond-ing chlorides with the same molarity. For solid media, 15 g ofagar per liter was added. After autoclaving, 16 ml of atitanium(III) citrate solution (28) per liter was added toagar-containing medium. Medium B (essentially the same asmedium A, but lacking tryptone and yeast extract) supple-mented with different substrates was used as describedbelow. Serum bottles with a volume of 70 ml were filled with20 ml of medium A or B, sealed with butyl rubber stoppersand aluminum crimp-top seals, and autoclaved. The inocu-lated bottles were pressurized aseptically with either H2-CO,or N, at 1 bar (0.987 atm).
For degradation studies, Clostridiium sp. strain TCAIIBwas grown at 35°C in medium C containing (per liter of
degassed water) 10.0 g of yeast extract, 5.0 g of tryptone,2.72 g of KH2PO4, 0.5 g of cysteine hydrochloride, 0.5 g ofNa2S 9H2O, and 10 ml of a solution with trace elements(17). The pH was adjusted to 7.5 prior to autoclaving. Forsolid media 15 g of agar was added per liter.
Biotransformation studies. Serum bottles with a total vol-ume of 118 ml were filled with 50 ml of medium C and sealedwith gas-tight Teflon (E. I. du Pont de Nemours & Co., Inc.,Wilmington, Del.) mininert valves (Precision SamplingCorp., Baton Rouge, La.). To prevent the mininert valvesfrom popping off during autoclaving, the bottles were fittedinto a metal rack. All mininert valves were pressurized totest them for leaks before they were used. Sterile stocksolutions of chlorinated aliphatic hydrocarbons were pre-pared in serum bottles that were completely filled withdegassed, deionized water which had been autoclaved. Thepure chlorinated compounds were injected with a microsy-ringe through the septa of the mininert valves. The volume ofstock solution injected into the serum bottles containingmedium C was calculated by using the Henry law constantsat 35°C (12) to obtain a concentration in liquid culture of 100,ug/liter. The bottles were inoculated and incubated in thevertical position in a 35°C chamber on a rotary shaker at 120rpm. It was verified that the medium did not alter the Henrylaw constants by comparing the concentrations of chlori-nated compounds in the headspace and those in the solutionsof bottles containing either medium C or water. Results ofpreliminary studies indicated that the incubation time toachieve equilibrium between headspace and liquid culture isless than 20 min, when the chlorinated compounds are
injected into prewarmed bottles.Periodically, sterile samples from the headspace were
taken inside the environmental chamber to be analyzed forvolatile halogenated compounds; samples were also takenfrom the liquid culture. These samples were assayed forprotein concentration. After each sampling, the bottles werefilled with the same volume of sterile gas mixture from theanaerobic chamber that was withdrawn by the samplingprocedure. The small loss of chlorinated compounds causedby the monitoring procedure was corrected for by calcula-tion.
Analytical methods. Protein concentration was measuredby the method of Bradford (4), with bovine serum albuminused as a standard. The bacteria were kept in a volume of 0.5to 1.0 ml of liquid culture, precipitated with 3 M trichloro-acetic acid (final concentration, 0.5 M), and collected bycentrifugation in a benchtop centrifuge (4,000 x g for 20min). The bacterial pellet was dissolved in 0.5 ml of 0.66 NNaOH for 48 h at 35°C and was then assayed for the proteinconcentration. All assays were performed in duplicate. Thestandard errors for the range of 10 to 60 Fg of protein werewithin 5%, and the detection limit was 2 ,ug of protein.Headspace concentrations of chlorinated aliphatic com-
pounds were determined by GC with a gas chromatograph(9610; Finnigan, Sunnyvale, Calif.) connected to a conduc-tivity detector (700A Hall; Tracor Instruments, Austin,Tex.) that was fitted to a fused-silica megabore column (30 mby 0.53 mm, film thickness, 1.8 ,um; DB-624; J+W ScientificInc., Rancho Cordova, Calif.) at 35°C. Sterile samples were
withdrawn from the serum bottles with a 1.0-ml gas-tightsyringe (Pressure Lok, series A-2) at 35°C and injecteddirectly into the gas chromatograph. Standard curves were
performed by analyzing serum bottles containing degassed,deionized water and the chlorinated compound. The stan-dard errors were within 10%, and the detection limits,corresponding to the concentrations in liquid culture, were
0.5 Vjg/liter for TCA, 1.0 kg/liter for DCA, 3.0 ,ug/liter forchloroethane, 0.2 Vg/liter for tetrachloromethane, 0.6 pug/liter for trichloromethane, and 1.0 pLg/liter for dichlorometh-ane.
Volatile acids in liquid culture were extracted with etherunder acidic conditions and analyzed with a gas chromato-graph (5730A; Hewlett-Packard Co., Palo Alto, Calif.) with aflame ionization detector and a 100/120 Chromosorb W AWglass column (190 cm by 4 mm, 10% SP-1000 and 1% H3PO4at 155°C; Supelco). A volatile acid standard mixture (Su-pelco) was used to identify and quantify the acids that wereproduced.[14C]TCA and [14C]DCA were separated by GC; and
fractions of the effluent, based on the retention times of thecompounds, were trapped in 10 ml ofACS liquid scintillationcounting solution (Amersham Corp., Arlington Heights, Ill.)(11, 26). Effluent bypassed the detector, which was done byusing a splitter part of the column, and was trapped inscintillation counting solution. 14C-labeled compounds wereused as standards. The recovery of [14C]TCA was 75%. 14Cwas assayed with a liquid scintillation spectrophotometer(Tricarb model 4530; Packard Instruments Co., DownersGrove, Ill.), as described by Vogel and McCarty (26).
Identification of chlorinated compounds was achieved bycomparing their retention times and the mass spectra withthose of pure compounds by using a gas chromatograph-mass spectrometer (model 5970; Hewlett-Packard) with a60-m thick-film capillary column (inner diameter, 0.32 mm;thickness, 1 mm; 30 to 180°C at a rate of 4°C/min; DB-5;J+W Scientific Inc.).Methane was determined by using a gas chromatograph
(model 5730A; Hewlett-Packard) with a flame ionizationdetector and a 60/80 Carbosieve glass column G (150 cm by3.2 mm at 120°C; Supelco).Hydrogen was measured by using a reduction gas detector
(Trace Analytical, Menlo Park, Calif.) at 280°C, with air usedas the carrier gas.Redox potentials were measured at 25°C with a platinum
electrode combined with a calomel reference electrode byusing supersaturated solutions of hydroquinone at pHs 4 and7 to calibrate the electrodes at 180 and 0 mV, respectively.
Separation and analysis of "4C-labeled compounds. Fromthe liquid cultures that were grown in the presence of[14C]TCA, 2 ml was removed and analyzed for the radioac-tivity that was associated with the biomass. Two 1-mlportions were centrifuged, and the bacterial pellets werewashed twice with medium C. To each pellet 1 ml of 1 NNaOH was added, and the tubes were sealed and incubatedfor 48 h at 35°C to dissolve the bacterial cells. Radioactivitywas then measured as described above.To the remaining culture 2.5 ml of 5 N KOH was added,
and the basic culture (pH 13) was extracted four times with10 ml of isooctane to remove volatile organic compounds.The isooctane fractions and the aqueous solution wereassayed for radioactivity. Volatile chlorinated compounds inthe isooctane fractions were separated by GC and trapped asdescribed above. The aqueous solution was analyzed for14CO2 (26).To make sure that all volatile organic compounds were
extracted, the aqueous solution was assayed before and afterit was purged with N2 for 1 h. The aqueous solution wasextracted once with ether, acidified with concentratedH2SO4 to a pH of 2, and then extracted three times with 10ml and three times with 20 ml of ether to remove the organicacids. The ether fractions were pooled and dried overanhydrous MgSO4. Acetic acid (20 ml) was added to the
acidic ether phase, and the volatile compounds were sepa-rated by fractional distillation. A total of four fractions wereobtained; they had boiling points of 35 to 1150C, 115 to1180C, 118 to 140°C, and >1400C (distillation residue). Theradioactivity was measured in each fraction.
RESULTSEnrichment and isolation of TCA-degrading microorgan-
isms. Medium A, a complex medium containing acetate,formate, yeast extract, tryptone, and vitamins that is widelyused in the cultivation of methanogenic bacteria, was inoc-ulated with liquid culture from two suspended-growth biore-actors that are known to biotransform trichloroethene tovinyl chloride and TCA to DCA and chloroethane (26; T. M.Vogel, Ph. D. thesis, Stanford University, Stanford, Calif.,1987). The bioreactors were fed acetone, isopropanol, andeither acetate or methanol semicontinuously under methano-genic conditions. The redox potentials and pH values of thecultures used as an inoculum were -100 mV and pH 7.5 forthe acetate-fed culture and -80 mV and pH 7.7 for themethanol-fed culture, whereas the values for medium Awere -390 mV and pH 7.3. Two series of serum bottles wereinoculated with each methanogenic culture and pressurizedwith N2 or H2-CO2. The bottles were incubated in thepresence of 100 p.g of TCA, trichloroethene, or tetrachlo-roethene per liter. Control bottles were incubated withoutthe addition of chlorinated aliphatic compounds. Methanol(10 mM) was added to bottles containing the methanol-fedinoculum. Bottles which were not inoculated, but that con-tained TCA, trichloroethene, or tetrachloroethene, served ascontrols. All cultures were incubated on a rotary shaker at20°C in the dark. The cultures were subcultured after 3 and6 weeks. Periodically, the bottles were assayed qualitativelyfor chlorinated aliphatic compounds, methane production,and a decrease in the H2-CO, pressure. After 5 to 8 days ofincubation, no TCA could be detected, but DCA was de-tected in all TCA-fed cultures. In cultures containing trichlo-roethene and tetrachloroethene, only minor concentrationsof transformation products (e.g., cis-1,2-dichloroethene)were detected. Methane was detected in all inoculatedbottles, indicating the presence of methanogenic bacteria.The TCA-transforming cultures were streaked onto agar
plates containing medium A. After 2 to 3 days, coloniesbecame visible; and single colonies were picked, dependingon the colony shape, to inoculate serum bottles containingmedium A and TCA. The plates were further incubated toallow the growth of slow-growing microorganisms. Theisolation of pure cultures was accomplished by seriallypicking single colonies from plates, growing them in liquidculture, and streaking the cultures onto the plates. Allisolates underwent at least three passages of the isolationprocedure. Based on their morphologies and ability to growon different substrates, 21 distinguishably different, TCA-biotransforming isolates were obtained.The isolates were tested for the presence of F420, a
coenzyme that is commonly found in methanogenic bacteria.They were compared with Methylobacteriium thermoau-totrophicum, which served as a reference, under the fluo-rescence microscope (6). No isolate contained coenzymeF420, and none of the isolates was able to produce significantamounts of methane or to grow on medium B. From theseresults it was concluded that the isolates belong to the groupof nonmethanogenic anaerobic heterotrophs. Seven strainsexhibited spore formation when they were examined micro-scopically and were able to grow after heat treatment (10 minat 70°C).
Characterization of strain TCAIIB. Strain TCAIIB wasderived from the acetate-fed methanogenic culture and wasisolated from an enrichment culture that was pressurizedwith the H,-CO, gas mixture. Since this strain biotrans-formed TCA about 2 to 10 times faster than the other isolatesdid, it was selected for further studies. Criteria for the purityof strain TCAIIB were microscopic examination and uni-form morphology of the colonies during growth on differentagar plates containing either medium A or medium B sup-plemented with 0.5% glucose, starch, Casamino Acids(Difco Laboratories, Detroit, Mich.), or casein and on eggyolk agar. The organism was a gram-positive, straight orslightly curved, motile rod (5 by 0.5 ,um after 1 day onmedium C at 35°C). During the early growth phase, up toseven cells were linked together, forming a chain. After 2days the cells became separated, and the length of a singlecell was about 3 p.m. The formation of terminal spores wasobserved microscopically before and after the spores werestained with malachite green and safranin. Spore formationoccurred after 5 days at 35°C only if the strain was grown onmedium A or medium B supplemented with yeast extract,tryptone, or amino acids (see below).
Strain TCAIIB was obligately anaerobic (did not grow onyeast extract or tryptone under aerobic conditions; El, >-50 mV) and was unable to carry out dissimilatory sulfate ornitrate reduction. On the basis of the characteristics de-scribed above, the organism was tentatively classified as aClostridiuim sp. (5, 14).Growth of Clostridium sp. strain TCAIIB. The growth of
strain TCAIIB was tested in liquid culture on medium Bwhich was supplemented with various substrates (0.5 g/liter). The strain grew on yeast extract, tryptone, casein,Casamino Acids, collagen, and albumin but not on sugars[cx-D-( + )-maltose, D-( + )-xylose, at-D-( + )-glucose, D-manni-tol, 1-lactose, or starch], metabolic intermediates (citricacid, DL-lactic acid, acetone, isopropanol, glutaric acid,DL-malic acid, or 2-ketoglutaric acid), or fatty acids (de-canoic acid, palmitic acid, or stearic acid). Strain TCAIIBgrew well on egg yolk agar, but was not able to liquefygelatin or to grow on pairs of amino acids (Stickland reac-tion). However, the organism was able to grow on a mixtureof 17 amino acids (amino acid standard H; Pierce ChemicalCo.). During growth on medium C (Fig. 1), as well as growthon medium B (lacking acetate and formate) supplementedwith the mixture of amino acids, acetic acid, butyric acid,isobutyric acid, and isovaleric acid were formed. Themethod used for the detection of volatile organic acids didnot allow the detection of formic acid or the quantification ofacetic acid. From the substrate spectrum of the organismand the formation of branched-chain fatty acids, it wasconcluded that strain TCAIIB belongs to the group ofproteolytic, nonsaccharolytic clostridia (14).The doubling time of strain TCAIIB growing on medium C
at 35°C was 3 to 4 h (Fig. 1), but no growth was observed at60°C. The hydrogen production rate was 0.45 pLmol per dayper liter on medium C, and hydrogen up to a concentration of5% in the headspace did not cause significant growth inhibi-tion.
Biotransformation of chlorinated aliphatic hydrocarbons.TCA (100 ,ug/liter) was degraded by Clostridium sp. strainTCAIIB within 13 days to below the detection limit of 0.5,ug/liter (Fig. 2). Between 30 and 40% of the TCA wasbiotransformed to DCA by reductive dehalogenation. DCAwas identified by GC-mass spectrometry (data not shown).TCA was not abiotically transformed in sterile controls, andDCA could not be further biotransformed by the organism.
FIG. 1. Growth and formation of volatile organic acids by Clostridiuim sp. strain TCAIIB onconcentration; A, isovaleric acid; A, isobutyric acid; O, butyric acid.
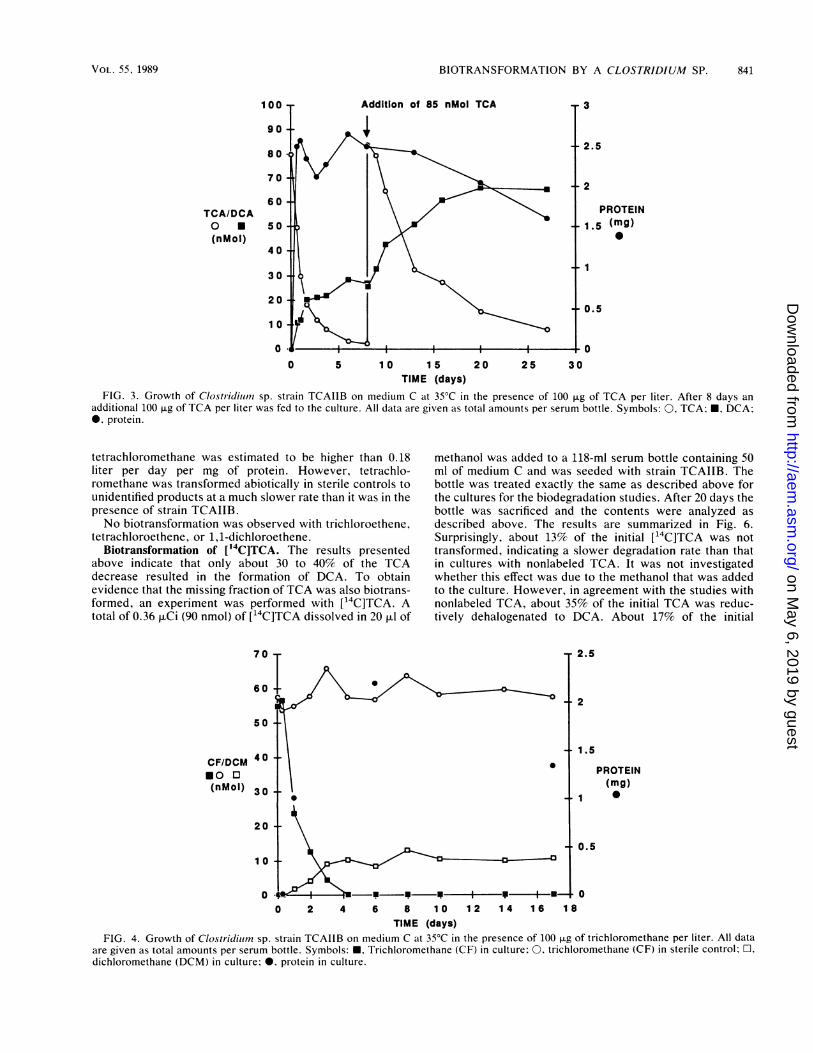
No significant chloroethane formation was observed in cul-tures grown in the presence of TCA or DCA, and chloroet-hane was not degraded by strain TCAIIB (data not shown).TCA (100 ,ug/liter) had no effect on the growth of theorganism, as could be seen by comparing it with a culturegrown without TCA. A duplicate bottle with TCA showingexactly the same results (Fig. 3) was fed an additional 100 1Lgof TCA per liter after 8 days. TCA was biotransformed toDCA at about the same ratio, but at a rate slower than thatobserved during the previous period. The klKl values basedon the Monod equation for substrate utilization were deter-mined by linear regression (3). For the period of 1 to 8 daysthe value was 0.013 liter per day per mg of protein (r = 0.98)and decreased to 0.0025 liter per day per mg of protein (r =0.99) after the second addition of TCA (8 to 27 days).
TCA/DCA00 U(nMol)
100
90
80
70
60
50
40
30
20
1 0
0
- 15
. 10
VOLATILEACIDS(mM)
El A A5
m o
medium C at 35°C. Symbols: *, Protein
Trichloromethane was completely (.99%) biotransformedby strain TCAIIB within 5 days, yielding about 20% of thetheoretical dichloromethane concentration (Fig. 4). Dichlo-romethane was identified by GC-mass spectrometry (datanot shown). Control experiments showed that dichlorometh-ane was not further degraded by the organism and trichlo-romethane was not abiotically transformed. The kIK, valuewas 0.020 liter per day per mg of protein (r = 0.99).The biotransformation of tetrachloromethane occurred
within the first day and led to the formation of trichlo-romethane, corresponding to about 55% of the initial tetra-chloromethane concentration (Fig. 5). As expected from theresults presented above, trichloromethane was further de-graded and dichloromethane was formed to about 8% of thetheoretical amount. The kIKs value for the disappearance of
3
2.5
2
PROTEIN1.5 (mg)
0
0.5
00 2 4 6 8 10 12 14 16 18 20
TIME (days)FIG. 2. Growth of Clostridium sp. strain TCAIIB on medium C at 35°C in the presence of 100 [±g of TCA per liter. All data are given as
total amounts per serum bottle. Symbols: 0, TCA in culture; O, TCA in sterile control; *, DCA in culture; 0, protein in culture.
FIG. 3. Growth of Clostridiin7? sp. strain TCAIIB on medium C at 35°C in the presence of 100 p.g of TCA per liter. After 8 days anadditional 100 p.g of TCA per liter was fed to the culture. All data are given as total amounts per serum bottle. Symbols: 0, TCA; *, DCA;*, protein.
tetrachloromethane was estimated to be higher than 0.18liter per day per mg of protein. However, tetrachlo-romethane was transformed abiotically in sterile controls tounidentified products at a much slower rate than it was in thepresence of strain TCAIIB.No biotransformation was observed with trichloroethene,
tetrachloroethene, or 1,1-dichloroethene.Biotransformation of ['4C]TCA. The results presented
above indicate that only about 30 to 40% of the TCAdecrease resulted in the formation of DCA. To obtainevidence that the missing fraction of TCA was also biotrans-formed, an experiment was performed with [14C]TCA. Atotal of 0.36 ,uCi (90 nmol) of [14C]TCA dissolved in 20 ,ul of
70
60
50
CF/DCM 4 0E0 0(nMol) 3 0
20
1 0
0
methanol was added to a 118-ml serum bottle containing 50ml of medium C and was seeded with strain TCAIIB. Thebottle was treated exactly the same as described above forthe cultures for the biodegradation studies. After 20 days thebottle was sacrificed and the contents were analyzed asdescribed above. The results are summarized in Fig. 6.Surprisingly, about 13% of the initial [14C]TCA was nottransformed, indicating a slower degradation rate than thatin cultures with nonlabeled TCA. It was not investigatedwhether this effect was due to the methanol that was addedto the culture. However, in agreement with the studies withnonlabeled TCA, about 35% of the initial TCA was reduc-tively dehalogenated to DCA. About 17% of the initial
2.5
2
1.5
PROTEIN(mg)
1 0
0.5
00 2 4 6 8 10 12 14 16 18
TIME (days)FIG. 4. Growth of Clostridilon sp. strain TCAIIB on medium C at 35°C in the presence of 100 p.g of trichloromethane per liter. All data
are given as total amounts per serum bottle. Symbols: *, Trichloromethane (CF) in culture: 0, trichloromethane (CF) in sterile control; O,dichloromethane (DCM) in culture; 0, protein in culture.
TIME (days)FIG. 5. Growth of Clostridiiin sp. strain TCAIIB on medium C at 35°C in the presence of 100 ,ug of tetrachloromethane per liter. All data
are given as total amounts per serum bottle. Symbols: U, Tetrachloromethane (CT) in culture; El, trichloromethane (CF) in culture: 0,tetrachloromethane (CT) in sterile control; A, dichloromethane (DCM) in culture; *, protein in culture.
radioactivity was found in the isooctane fraction, but did notpass the GC column. No chloroethane was detected in theisooctane fraction. No radioactivity was found to be associ-ated with biomass, and only a negligible amount was de-tected as "4CO2, indicating that strain TCAIIB is not able tomineralize TCA. The ether-extractable organic acid fractioncontained at least two organic acids. After microdistillationof the ether fraction containing nonlabeled acetic acid, about6% of the total radioactivity was found in the acetic acidfraction (boiling point, 115 to 118°C), whereas about 11%was found in the distillation residue (boiling point, >140°C).About 11% of the initial radioactivity was not extracted fromthe aqueous solution by the extraction procedure. The
results indicate that only about 35% of the TCA wasbiotransformed to DCA by reductive dehalogenation,whereas the main part was transformed, possibly by othermechanisms.
DISCUSSION
The isolation of microorganisms with biodegradation po-tential from anaerobic mixed cultures that are capable oftransforming halogenated C1 and C2 aliphatic compoundshas not been reported previously. Therefore, the role ofdifferent groups of anaerobic microorganisms in thebiotransformation of these compounds remains unclear. In
FIG. 6. Biotransformation of [14C]TCA by Clostridium sp. strain TCAIIB on medium C. The culture was sacrificed after an incubationtime of 20 days at 350C and analyzed for different radiolabeled compounds. Data are given as the percentage of total radioactivity that wasapplied.
this study, 21 TCA-transforming pure cultures were isolatedfrom methanogenic cultures that were fed acetone, isopro-panol, and either acetate or methanol semicontinuously.Since the medium used for the enrichment and isolation ofthe organisms did not contain nitrates or sulfates, the growthof methanogens and anaerobic heterotrophs was favored.No methanogenic organisms were isolated, but methaneformation was observed in enrichment cultures and metha-nogens were detected under the fluorescence microscope. Ofthe 21 isolated microorganisms, 7 produced endospores andpresumably belonged to the genus Clostridium. Because theclostridia are able to form spores, they are ubiquitouslyfound in the environment (14) and might play an importantrole in the dehalogenation of chlorinated compounds inanaerobic ecosystems. Most of the biotransformation ofTCA occurred during the stationary phase, indicating thestability of the dehalogenation activity even in resting cells.The biotransformation of TCA proceeded for longer than 1month at 35°C after growth in batch culture occurred.However, strain TCAIIB could not grow on the primarysubstrates that were used to feed the bioreactor from whichit was derived. Therefore, we assume that the organism isable to grow on decay products of microorganisms feedingon the primary substrates. The degradation of TCA by strainTCAIIB led to the accumulation of DCA, whereas DCA wasfurther degraded to chloroethane and CO2 in the methano-genic mixed culture (26). This indicates that in the mixedculture, other microorganisms are also involved in the over-all biotransformation of TCA. Trichloroethene and tetra-chloroethene were not significantly biotransformed by strainTCAIIB, although they could be transformed by the mixedculture (26). Tetrachloroethene was not biotransformed bytwo different strains of Clostridiuim (8). It thus appears thatin anaerobic mixed cultures, even when they are enrichedwith simple substrates such as acetate or methanol, manydifferent strains may participate in the dehalogenation reac-tions that are observed. Thus, caution is needed in general-izing about the role of individual organisms in dehalogena-tion reactions in such systems.
Reductive dehalogenations under anaerobic conditionshave been reported for many chlorinated compounds such aschlorinated fatty acids (15), various isomers of hexachloro-cyclohexane (16, 19), chlorinated aromatic compounds (10),dichlorodiphenyltrichloroethane (DDT) (19), and chlorinatedaliphatic C1 and C2 compounds (7, 11, 21, 22, 26). Chlori-nated aliphatic C1 and C2 compounds such as tetrachloroet-hene, TCA, tetrachloromethane, trichloromethane, and 1,2-dichloroethane can be reductively dehalogenated by purecultures of methanogens (7, 8), Desulfobacter auitotrophi-cum (7), and a 3-chlorobenzoate-degrading bacterium (8).The reductive dehalogenation of TCA and trichloromethane,leading to the accumulation of DCA and dichloromethane,respectively, has been shown by other investigators (7, 11,21). This observation might lead to the assumption that thereductive dehalogenation of these compounds shifts theproblem from compounds containing three chlorines tocompounds containing two chlorines in the molecule. Sincedichloromethane is readily mineralized under aerobic condi-tions by various methylotrophs (9) and DCA is slowlydehalogenated to chloroethane and CO2 under methanogenicconditions (26), however, it seems possible that TCA andtrichloromethane can be mineralized under appropriate con-ditions.
Strain TCAIIB transformed about 60 to 70% of the TCA toacetic acid and other unidentified products. We assume thatdegradation pathways other than reductive dehalogenation
might exist. A nucleophilic substitution for a chlorine atomby a hydroxy group would lead to the formation of aceticacid, which was apparently formed by the organism. Theoxidation of TCA would lead to 2-hydroxy-TCA, whichmight either be dehalogenated to 2-hydroxy-DCA, be oxi-dized to dichloroacetic acid or trichloroacetic acid, or both.However, no experiments were undertaken to prove thesehypotheses.
In sterile controls the tetrachloromethane concentrationdecreased significantly, but the decrease was much slowerthan that in seeded cultures. The abiotic transformation oftetrachloromethane has been shown to occur in reducedaqueous solutions containing Fe2+ ions or porphyrins (18),which were both present in medium C. No abiotic transfor-mation of TCA or trichloromethane was observed during thegiven period of time, as was expected from the reportedhalf-lives (18, 20, 25). However, no experiments were un-dertaken to investigate the possibility of transformation as aresult of a reaction of the chlorinated aliphatic compoundswith biomass decay products. For tetrachloromethane,which is a stronger electron acceptor than trichloromethaneor TCA (24), the kIKS value was estimated to be .0.18 literper day per mg of protein, whereas the values for trichlo-romethane and TCA were 0.020 and 0.013 liter per day permg of protein, respectively. It needs to be investigatedwhether strain TCAIIB is able to use chlorinated compoundsas electron acceptors for growth. Future studies should alsoelucidate biotransformation pathways and the possibility offavoring pathways that lead to nonhalogenated products.
ACKNOWLEDGMENTS
We thank B. I. Galli for excellent technical assistance and F.Haag for the performance of GC-mass spectrometry analysis.
This study was supported by grants from the Swiss NationalFoundation and grant ECE 8519243 from the National ScienceFoundation.
LITERATURE CITED1. Balch, W. E., G. E. Fox, C. R. Woese, and R. S. Wolfe. 1979.
Methanogens: reevaluation of a unique biological group. Micro-biol. Rev. 43:260-296.
2. Bouwer, E. J., and P. L. McCarty. 1983. Transformation of 1-and 2-carbon halogenated aliphatic organic compounds undermethanogenic conditions. Environ. Sci. Technol. 45:1286-1294.
3. Bouwer, E. J., and P. L. McCarty. 1985. Utilization rates oftrace halogenated organic compounds in acetate-grown bio-films. Biotechnol. Bioeng. 27:1564-1571.
4. Bradford, M. 1976. A rapid and sensitive method for thequantitation of microgram quantities of protein utilizing theprinciple of protein-dye binding. Anal. Biochem. 72:248-254.
5. Cato, E. P., W. L. George, and S. M. Finegold. 1986. GenusClostridium, p. 1141-1200. In P. H. A. Sneath, N. S. Mair,M. E. Sharpe, and J. G. Holt (ed.), Bergey's manual of system-atic bacteriology, vol. 2. The Williams & Wilkins Co., Balti-more.
6. Doddema, H. J., and G. D. Vogels. 1978. Improved identificationof methanogenic bacteria by fluorescence microscopy. Appl.Environ. Microbiol. 36:752-754.
7. Egli, C., R. Scholtz, A. M. Cook, and T. Leisinger. 1987.Anaerobic dechlorination of tetrachloromethane and 1,2-dichlo-roethane to degradable products by pure cultures of Desulfo-bacterium sp. and Methanobacteriuin sp. FEMS Microbiol.Lett. 43:257-261.
8. Fathepure, B. Z., J. P. Nengun, and S. A. Boyd. 1987. Anaerobicbacteria that dechlorinate perchloroethene. Appl. Environ. Mi-crobiol. 53:2671-2674.
9. Gailli, R., and T. Leisinger. 1986. Specialized bacterial strainsfor the removal of dichloromethane from industrial waste.Conserv. Recycl. 8:91-100.
10. Gibson, S. A., and J. M. Suflita. 1986. Extrapolation of biodeg-radation results to groundwater aquifers: reductive dehalogena-tion of aromatic compounds. Appl. Environ. Microbiol. 52:681-688.
11. Gossett, J. M. 1985. Anaerobic degradation of Cl and C,chlorinated hydrocarbons. Air Force Engineering and ServicesCenter. ESI-TR-85-38. Final Report. National Technical Infor-mation Service, Springfield, Va.
12. Gossett, J. M. 1987. Measurement of Henry's law constants forC1 and C. chlorinated hydrocarbons. 1987. Environ. Sci. Tech-nol. 21:202-208.
13. Gottschalk, G. 1981. The anaerobic way of life of procaryotes,p. 1415-1424. In M. P. Starr, H. Stolp, H. G. Trulmper, A.Balows, and H. G. Schlegel (ed.), The procaryotes: a handbookon habitat, isolation and identification of bacteria. Springer-Verlag, New York.
14. Gottschalk, G., J. R. Andreesen, and H. Hippe. 1981. The genusClostridinim (nonmedical aspects), p. 1767-1803. In M. P. Starr.H. Stolp, H. G. Trulmper, A. Balows, and H. G. Schlegel (ed.),The procaryotes: a handbook on habitat, isolation and identifi-cation of bacteria. Springer-Verlag. New York.
15. Hashimoto, H., and H. Simon. 1975. Reductive DehalogenierungP-halogenierter Fettsauren und stereoselective Hydrierung a-halogenierter, ot,3-ungesattigter Fettsauren durch Clostiidiininkhlvi'eri. Angew. Chem. 87:111-112.
16. Jagnow, G., K. Haider, and P.-C. Ellwardt. 1977. Anaerobicdechlorination and degradation of hexachlorocyclohexane iso-mers by anaerobic and facultative anaerobic bacteria. Arch.Microbiol. 115:285-292.
17. Kiener, A., and T. Leisinger. 1983. Oxygen sensitivity of meth-anogenic bacteria. Syst. Appl. Microbiol. 4:305-312.
18. Klecka, G. M., and S. J. Gonsior. 1984. Reductive dechlorina-tion of chlorinated methanes and ethanes by reduced iron (11)
porphyrins. Chemosphere 13:391-402.19. Lal, R., and D. M. Saxena. 1982. Accumulation, metabolism and
effects of organochlorine insecticides on microorganisms. Mi-crobiol. Rev. 46:95-127.
20. Mabey, W., and R. E. Robertson. 1978. Critical review ofhydrolysis of organic compounds in water under environmentalconditions. J. Phys. Chem. Ref. Data 7:383-415.
21. Parsons, F., and G. Barrio-Lage. 1985. Chlorinated organics insimulated groundwater environments. J. Am. Water WorksAssoc. 77:52-59.
22. Parsons, F., G. Barrio-Lage, and R. Rice. 1985. Biotransforma-tion of chlorinated organic solvents in static microcosms. Envi-ron. Toxicol. Chem. 4:739-742.
23. Shelton, D. R., and J. M. Tiedje. 1984. Isolation and partialcharacterization of bacteria in an anaerobic consortium thatmineralize 3-chlorobenzoic acid. Appl. Environ. Microbiol.48:840-848.
24. Vogel, T. M., C. S. Criddle, and P. L. McCarty. 1987. Trans-formations of halogenated aliphatic compounds. Environ. Sci.Technol. 21:722-736.
25. Vogel, T. M., and P. L. McCarty. 1987. Rate of abiotic forma-tion of 1,1-dichloroethylene from 1,1,1-trichloroethane ingroundwater. J. Contam. Hydrol. 1:299-308.
26. Vogel, T. M., and P. L. McCarty. 1988. Abiotic and biotictransformations of 1,1,1-trichloroethane under methanogenicconditions. Environ. Sci. Technol. 21:1208-1213.
27. Westerick, J. J., J. W. Mello, and R. F. Thomas. 1984. Thegroundwater supply survey. J. Am. Water Works Assoc. 76:52-59.
28. Zehnder, A. J. B., and K. Wuhrmann. 1976. Titanium (Ill)citrate as a nontoxic oxidation-reduction buffering system forthe culture of obligate anaerobes. Science 194:1165-1166.