Revista Chilena de Historia Natural 79: 245-255, 2006 Bofedales: high altitude peatlands of the central Andes Bofedales: turberas de alta montaña de los Andes centrales FRANCISCO A. SQUEO 1, 3, 4 , BARRY G. WARNER 2, 3 , RAMÓN ARAVENA 2, 3 & DIANA ESPINOZA 1, 4 1 Departamento de Biología, Facultad de Ciencias, Universidad de La Serena, Casilla 599, La Serena, Chile 2 University of Waterloo, Waterloo, Ontario, Canada N2L 3G1; e-mail: [email protected]; [email protected]3 Center for Advanced Studies in Arid Zones (CEAZA), Casilla 599, La Serena, Chile 4 Institute of Ecology and Biodiversity (IEB); * e-mail for correspondence: [email protected]ABSTRACT There is an exceptional group of alpine peatlands in the world situated in the arid grasslands of the central Andes. The peatlands in northern Chile occur in the most arid part of their range. Members of the Juncaceae are the primary peat-forming plant species. Fresh and mildly saline groundwaters originate from glaciers, snowmelt and rain are the water sources for the northern Chile peatlands. Paleoecological investigations suggest that some peatlands are recent features of the landscape having developed within the last three thousand years or less. These peatlands are unique, extremely fragile water features sensitive to climate changes and human disturbances such as regional mining activity. Much more work is required to develop scientifically based sound management and conservation programs for the rare plants and animals that live in them and to ensure the future livelihoods of the indigenous peoples who depend on them. Key words: peatlands, arid grasslands, Altiplano, central Andes, South America. RESUMEN Existe un grupo excepcional de turberas (bofedales) de alta montaña en el mundo situados en la estepa árida de los Andes centrales. Los bofedales en el norte de Chile están presentes en la parte más árida de su rango. Las principales especies de plantas responsables de la formación de turba corresponden a miembros de Juncaceae. El agua fresca y medianamente salina de los bofedales proviene de agua subterránea asociada a riachuelos proveniente de glaciares, derretimiento de nieve y lluvia. Investigaciones paleoecológicas sugieren que algunos bofedales son integrantes recientes del paisaje, habiéndose desarrollado durante los últimos tres mil años o menos. Estos bofedales son entidades únicas, extremadamente frágiles por su dependencia del agua, sensibles a los cambios climáticos y vulnerables a la alteración humana tal como la actividad minera en la región. Se requiere mucho más trabajo para desarrollar programas de manejo y conservación, con sólidas bases científicas, de las plantas y animales que viven en ellos, y para asegurar la capacidad futura de pastoreo de la cual dependen los pueblos indígenas. Palabras clave: turberas, estepa árida, Altiplano, Andes centrales, América del Sur. INTRODUCTION The existence of wetlands, especially peat- accumulating wetlands, in arid environments seems counter intuitive. Indeed, wetlands exist in environments with low precipitation and soil moisture deficits. The wadis of southern Africa, the oases of the Middle East, and the billabongs of Australia are well known examples of arid land wetlands. Much less known, however, are the peatlands in the high Andean arid zone of the central Andes, which have been referred to as bofedales, vegas, cushion bogs, and wet grasslands. Despite hyper-aridity, intense solar radiation, high-velocity winds, hypoxia, daily frost, and a short growing season, bofedales are near the hydrological and altitudinal limits for plant life in the cold and arid grasslands of Perú, Bolivia, Chile and Argentina (Ruthsatz 1993, 2000, Squeo et al. 1994, 2006b, Villagrán & Castro 1997). These peatlands are like no other in the world. They have been referred to as “highland bogs” (Wilcox 1986, Ruthsatz 1993), but they
Transcript
245PEATLANDS OF THE CENTRAL ANDES Revista Chilena de Historia Natural79: 245-255, 2006
Bofedales: high altitude peatlands of the central Andes
Bofedales: turberas de alta montaña de los Andes centrales
FRANCISCO A. SQUEO1, 3, 4, BARRY G. WARNER2, 3, RAMÓN ARAVENA2, 3 & DIANA ESPINOZA1, 4
1 Departamento de Biología, Facultad de Ciencias, Universidad de La Serena, Casilla 599, La Serena, Chile2 University of Waterloo, Waterloo, Ontario, Canada N2L 3G1;
4 Institute of Ecology and Biodiversity (IEB);* e-mail for correspondence: [email protected]
ABSTRACT
There is an exceptional group of alpine peatlands in the world situated in the arid grasslands of the centralAndes. The peatlands in northern Chile occur in the most arid part of their range. Members of the Juncaceaeare the primary peat-forming plant species. Fresh and mildly saline groundwaters originate from glaciers,snowmelt and rain are the water sources for the northern Chile peatlands. Paleoecological investigationssuggest that some peatlands are recent features of the landscape having developed within the last threethousand years or less. These peatlands are unique, extremely fragile water features sensitive to climatechanges and human disturbances such as regional mining activity. Much more work is required to developscientifically based sound management and conservation programs for the rare plants and animals that live inthem and to ensure the future livelihoods of the indigenous peoples who depend on them.
Key words: peatlands, arid grasslands, Altiplano, central Andes, South America.
RESUMEN
Existe un grupo excepcional de turberas (bofedales) de alta montaña en el mundo situados en la estepa áridade los Andes centrales. Los bofedales en el norte de Chile están presentes en la parte más árida de su rango.Las principales especies de plantas responsables de la formación de turba corresponden a miembros deJuncaceae. El agua fresca y medianamente salina de los bofedales proviene de agua subterránea asociada ariachuelos proveniente de glaciares, derretimiento de nieve y lluvia. Investigaciones paleoecológicas sugierenque algunos bofedales son integrantes recientes del paisaje, habiéndose desarrollado durante los últimos tresmil años o menos. Estos bofedales son entidades únicas, extremadamente frágiles por su dependencia delagua, sensibles a los cambios climáticos y vulnerables a la alteración humana tal como la actividad minera enla región. Se requiere mucho más trabajo para desarrollar programas de manejo y conservación, con sólidasbases científicas, de las plantas y animales que viven en ellos, y para asegurar la capacidad futura de pastoreode la cual dependen los pueblos indígenas.
Palabras clave: turberas, estepa árida, Altiplano, Andes centrales, América del Sur.
INTRODUCTION
The existence of wetlands, especially peat-accumulating wetlands, in arid environmentsseems counter intuitive. Indeed, wetlands existin environments with low precipitation and soilmoisture deficits. The wadis of southern Africa,the oases of the Middle East, and the billabongsof Australia are well known examples of aridland wetlands. Much less known, however, arethe peatlands in the high Andean arid zone ofthe central Andes, which have been referred to
as bofedales, vegas, cushion bogs, and wetgrasslands. Despite hyper-aridity, intense solarradiation, high-velocity winds, hypoxia, dailyfrost, and a short growing season, bofedales arenear the hydrological and altitudinal limits forplant life in the cold and arid grasslands ofPerú, Bolivia, Chile and Argentina (Ruthsatz1993, 2000, Squeo et al . 1994, 2006b,Villagrán & Castro 1997).
These peatlands are like no other in theworld. They have been referred to as “highlandbogs” (Wilcox 1986, Ruthsatz 1993), but they
246 SQUEO ET AL.
are neither dominated by Sphagnum mosses norare they exclusively ombrogenous, as is typicalof true bogs in the Northern Hemisphere. Theironly similarity to northern bogs is themicrotopographic patterns of pools, lawns, andhummocks. Individual systems vary in extentfrom less than one hectare to in excess of 100sof hectares. Fresh and mildly salinegroundwater originate from glacier streams,snowmelt and rain are the water sources ofthese peatlands. Members of the Juncaceae,most common species being Oxychloe andinaand Patosia clandestina are the communitydominants and primary peat-formers (Ruthsatz1993, 2000, Squeo et al. 2006b).
The peatlands play a crit ical role insustaining a unique diversity of rare andendemic biota in the Cordillera de los Andes. Asmall number of mammals and bird species,about one-third of which is threatened, dependupon the peatlands for grazing, nesting andwater. Camelid species, wild vicuña (Vicugnavicugna) and guanaco (Lama guanicoe) are themost obvious mammalian inhabitants(Villagrán & Castro 1997).
Communities of native Aymara andAtacameños peoples are directly dependentupon the peatlands in this region whereconditions are so severe as to almost precludehuman habitation (Villagrán & Castro 1997,2003, Villagrán et al. 1999, 2003). Thepeatlands are used for grazing by theirdomestic herds of llamas (Lama glama) andalpacas (Vicugna pacos), which are the basis ofthe local indigenous economy. In other areas,the living surface layer of the peatlands isstripped away to expose underlying organic-rich mineral soils for cultivation. Drainage ofthe peatlands by hand-dug ditches to re-routewater to drier areas is undertaken to encourageexpansion of peatland and hence, the extent ofpastureland.
This paper focuses on the Oxychloe andPatosia-dominated peatlands in the most aridpart of their range in Chile. We assess the stateof current knowledge and focus on identifyingfactors contributing to their existence andcharacter. Such information is needed formanagement and conservation programsbecause there is growing pressure on water andassociated biological resources in this region.Potential conflict exists between industrialdevelopment and protection of these fragile
natural resources (Messerli et al. 1993). Stone(1992) asserted that fragmented andoversimplified views of fragile Andeanecosystems have led to mismanagement. Thedynamics of peatlands and their connectionwith water sources is not well understood. Noris it clear what the relationships are withclimate. However, legislation to protect thesefragile ecosystems is recognized by localgoverning bodies on water use in regions suchas Tarapacá and Antofagasta in Chile whereexploitation of water must have regard forpeatlands and for their groundwater rechargeareas (Dirección General de Aguas, Gobiernode Chile 1996).
There are examples of severely degraded andvanishing peatlands in northern Chile (Squeo etal. 1989, 1993, 1998, 2001, Arroyo et al. 1993,Villagrán & Castro 1997). Earle et al. (2003)suggested that degradation was associated withautoregulation processes, however, questionsremain as to whether there may be externallinkages with changes in regional precipitationor groundwater extraction for lowlandagriculture, urbanization, and mining? Are otherfactors, such as a decrease in regionalprecipitation in recent decades responsible forpeatland deterioration in this already water-stressed region? What is the connection betweenclimate and the regional hydrological and bioticresources? Is it possible that peatlandsdeterioration is part of the natural autogenicaging process of these sensitive ecosystems?
Where do these peatlands occur?
Bofedales are primarily restricted to the lowAlpine and sub-Alpine belts of the centralAndes at elevations between 3,200 to near5,000 m in the north and central part of theirrange and at elevations greater than 2800 m attheir southern limit (Fig. 1 and 2). Thegrassland and steppe straddle the volcanic andigneous rocks of the precordillera and westerncordillera ranges of the high Andes. The mostdistinctive geological feature in this region isthe Altiplano, a large flat plateau formed byMesozoic and Cenozoic sedimentary deposits,especially thick volcanic ashes laid down inlate Cenozoic times (Charrier 1997). TheAltiplano is among the highest plateaus in theworld. Glaciers descended onto the Altiplanofrom the surrounding mountain peaks and
247PEATLANDS OF THE CENTRAL ANDES
covered it with ice during Quaternary time.Large water bodies inundated the Altiplanoduring ice-free periods and eventually recededby the Late Quaternary (Clayton & Clapperton1997). Aeolian sand plains and dunes andwind-swept gravel and cobble plainscharacterize the modern landscape. Most of thebasins in the Altiplano are endorreic and arecharacterized by the occurrence of salt lakes
Fig. 1: Map showing the primary ecoregions of the central Andes (after Olson et al. 2001, WWF2001).Ubicación de las ecorregiones primarias de los Andes centrales (según Olson et al. 2001, WWF 2001).
and shallow open water wetlands (locallyreferred to as salares). Mechanical weatheringis intense, but the cold climate, aridity and lackof leaching, high relief and the continualdownward movement of mineral matter,detritus and water prevent the development ofmature soils and well-established plantcommunities (Wilcox 1986, Veit 1996,Abraham et al. 2000).
248 SQUEO ET AL.
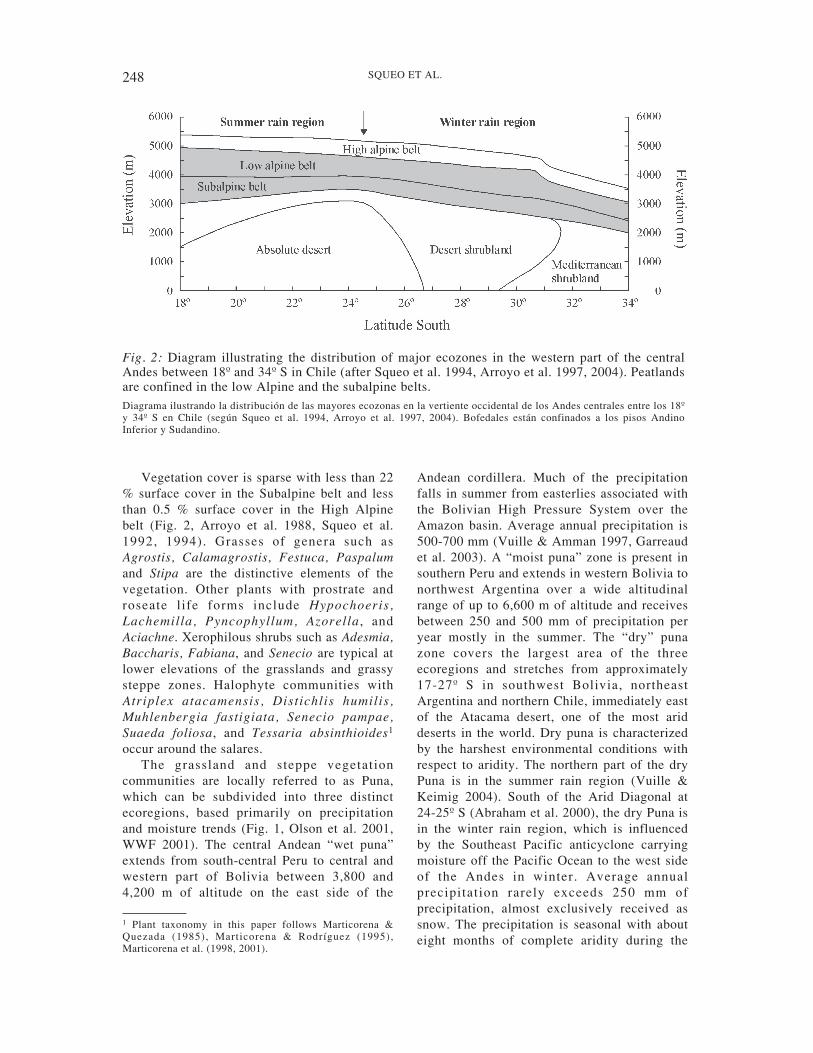
Vegetation cover is sparse with less than 22% surface cover in the Subalpine belt and lessthan 0.5 % surface cover in the High Alpinebelt (Fig. 2, Arroyo et al. 1988, Squeo et al.1992, 1994). Grasses of genera such asAgrostis, Calamagrostis, Festuca, Paspalumand Stipa are the distinctive elements of thevegetation. Other plants with prostrate androseate l ife forms include Hypochoeris,Lachemilla, Pyncophyllum, Azorella , andAciachne. Xerophilous shrubs such as Adesmia,Baccharis, Fabiana, and Senecio are typical atlower elevations of the grasslands and grassysteppe zones. Halophyte communities withAtriplex atacamensis, Distichlis humilis,Muhlenbergia fastigiata, Senecio pampae,Suaeda foliosa, and Tessaria absinthioides1
occur around the salares.The grassland and steppe vegetation
communities are locally referred to as Puna,which can be subdivided into three distinctecoregions, based primarily on precipitationand moisture trends (Fig. 1, Olson et al. 2001,WWF 2001). The central Andean “wet puna”extends from south-central Peru to central andwestern part of Bolivia between 3,800 and4,200 m of altitude on the east side of the
Andean cordillera. Much of the precipitationfalls in summer from easterlies associated withthe Bolivian High Pressure System over theAmazon basin. Average annual precipitation is500-700 mm (Vuille & Amman 1997, Garreaudet al. 2003). A “moist puna” zone is present insouthern Peru and extends in western Bolivia tonorthwest Argentina over a wide altitudinalrange of up to 6,600 m of altitude and receivesbetween 250 and 500 mm of precipitation peryear mostly in the summer. The “dry” punazone covers the largest area of the threeecoregions and stretches from approximately17-27º S in southwest Bolivia, northeastArgentina and northern Chile, immediately eastof the Atacama desert, one of the most ariddeserts in the world. Dry puna is characterizedby the harshest environmental conditions withrespect to aridity. The northern part of the dryPuna is in the summer rain region (Vuille &Keimig 2004). South of the Arid Diagonal at24-25º S (Abraham et al. 2000), the dry Puna isin the winter rain region, which is influencedby the Southeast Pacific anticyclone carryingmoisture off the Pacific Ocean to the west sideof the Andes in winter. Average annualprecipitation rarely exceeds 250 mm ofprecipitation, almost exclusively received assnow. The precipitation is seasonal with abouteight months of complete aridity during the
Fig. 2: Diagram illustrating the distribution of major ecozones in the western part of the centralAndes between 18º and 34º S in Chile (after Squeo et al. 1994, Arroyo et al. 1997, 2004). Peatlandsare confined in the low Alpine and the subalpine belts.Diagrama ilustrando la distribución de las mayores ecozonas en la vertiente occidental de los Andes centrales entre los 18ºy 34º S en Chile (según Squeo et al. 1994, Arroyo et al. 1997, 2004). Bofedales están confinados a los pisos AndinoInferior y Sudandino.
1 Plant taxonomy in this paper follows Marticorena &Quezada (1985), Marticorena & Rodríguez (1995),Marticorena et al. (1998, 2001).
249PEATLANDS OF THE CENTRAL ANDES
main growing season. Values as high as 400mm have been noted in the north and as low as50 mm at 24-25º S, increasing again to thesouthern limit (Arroyo et al. 1988). SouthernAndean steppe is the natural continuity of theAndean vegetation south of the dry Puna. It islocated in north-central Chile and west-centralArgentina. Precipitation is more abundant (over250 mm) and falls mostly as snow during thewinter (Squeo et al. 1994).
Peatlands in northern Chile
The peatlands are found from about 18o30’ S innorthern Chile at the southern limits of themoist Puna ecoregion, across the dry Punawhere most of the peatlands occur, to about 31ºS at their southern limit in the southern Andeansteppe ecoregion (Fig. 1, Squeo et al. 2001).They appear as green oases in valley bottoms,shallow basins and other low areas of relief inthis otherwise poorly vegetated and aridlandscape (Arroyo et al. 1988, Villagrán &Castro 2003).
The climate is characterized as subtropicalsemi-arid desert (Miller 1976). Conditions inChile are slightly wetter in the north of thisarea, and become progressively more arid tothe Arid Diagonal at 24-25º S. Lowatmospheric pressure, low air densities andminimal atmospheric humidity are typical ofthis high altitude region. In the north, rainfall isreceived mostly during the summer (December-March) when the regional easterlies bringmoisture over the Andes from the Amazonbasin. Local convective thunderstorms are mostcommon in late afternoon as a result of theintense surface heating by solar radiation(Aceituno 1997). There is virtually noprecipitation during the summer over the dryPuna between 24-25º S (Arroyo et al. 1988,Abraham et al . 2000). Potentialevapotranspiration is about 1,000 mm year-1,exceeding precipitation by a factor of five(Hastenrath & Kutzbach 1985, Risacher et al.1999). Most of the precipitation in the region isreceived as snowfall during the winter.However, a larger amount of snowfall isactually lost by sublimation (Vuille & Amman1997). Runoff from snowmelt and permanentglaciers is abundant and brief during the spring,except in areas where glacier meltwaters flowyear round.
Air temperatures are low with wide diurnalvariations resulting from intense surface heatingassociated with strong solar radiation and theintense radiative cooling during the night. Inhigh Andes at 30º S, the maximum andminimum extreme temperatures occur in January(18.7 ºC, -0.4 ºC) and July (8.3 ºC, -14.5 ºC),respectively. The minimum monthly temperatureof around -5 ºC during spring (end ofNovember) is a limiting factor that controls thebeginning of the growing season in this part ofChile (Squeo et al. 2006a, 2006b). The length oftime with and magnitude of freezingtemperatures increases with increasing altitude(Squeo et al. 1996).
Peatlands types in northern Chile
Three main groups of peatlands can berecognized based on their overall shape,hydrogeological setting and dominant source ofwaters (Fig. 3). The first group is slopingpeatlands that occur along steep valley bottomsand streams (Fig. 3A and 3B). These slopingpeatlands can be a few kilometers long andonly a few 10s of meters wide. Groundwaterdischarge from local flow systems seems to bethe dominant source of water with some inputof water from direct snowmelt and surfacefeeder streams (J.C. Aravena, unpublisheddata). Waters near the surface of these slopingpeatlands have high pH between 8-9 and lowconductivity 1-2 μS cm-1 (Iriarte et al. 1998)but low pH waters have also found in peatlandsin areas near ore deposits.
The second group, referred to as basinpeatlands, tends to be wider with a flat surfaceand includes those developed behind endmoraines and in cirque basins, shallowdepressions and other low areas of relief (Fig.3C and 3D). They can be up to a few hundredmeters wide and can have some slope relief butare not as steep as the sloping peatlands.Glacier streams originating from higherelevations and groundwater dischargeassociated with regional groundwater flowsystems, together create a complex hydrologyfor these peatlands. In the Andean steppeecoregion, there appears to be a strongrelationship between plant communities andsalinity. The water associated with Distichiamuschoides communities ranges between 19and 713 μS cm-1, which are linked to springs
250 SQUEO ET AL.
Fig. 3: Sketches of cross-sectional (A, C, E) and plan views (B, D, F) of the types of general peatlandform types in the high Andes of northern Chile based on geomorphological setting and hydrolo-gical conditions: (A, B) sloping peatland, (C, D) basin peatland, and (E, F) flat peatland. Arrowsindicate water flow directions.Esquemas de secciones verticales (A, C, E) y vistas superficiales (B, D, F) de los tipos de bofedales presentes en losAndes altos del norte de Chile basado en la geomorfología y condiciones hidrológicas: (A, B) bofedal de ladera, (C, D)bofedal de quebrada, y (E, F) bofedal plano. Las flechas indican las direcciones del flujo del agua.
(A) (B)
(C) (D)
(E) (F)
Cross-sectional view Aerial view
251PEATLANDS OF THE CENTRAL ANDES
and small streams that do not accumulate saltsduring the dry season. The Oxychloe andinacommunities are associated with more salineconditions with conductivity values between 22and 2,620 μS cm-1 and mixed Oxychloe andina-Deschampsia caespitosa communities grow inwater with conductivity values as high as 2300μS cm-1 (Espinoza, unpublished data).
The last group, are flat peatlands, which arelarge and expansive (Fig. 3E and 3F) coveringwide areas. These peatlands complexes includenatural systems and peatland areas created andrestored by human actions. The localinhabitants cut networks of shallow channels todivert water in and around pre-existing naturalpeatlands, which initialize peatland formationand overall peat landform expansion. Surfacewaters largely dominate the restored parts ofthese systems. The main characteristics of atypical bofedal in Chile are showed in Table 1.
Vegetation
The peatland vegetation contrasts sharply withsurrounding terrestrial communities by havingplant cover usually greater than 70 % and highplant productivity (biomass over 1,000 g m-2)(Squeo et al. 1993, 1994, 2006b). Three distinctvegetation communities are characteristic ofthese peatlands in Chile (Villagrán et al. 1983,Arroyo et al. 1988, Ruthsatz 2000, Villagrán &Castro 2003). Cushion plants of the Juncaceae,mainly Oxychloe andina and Patosiaclandestina and scattered Distichia muscoides
and D. filamentosa, dominate the lawn andhummock communities. Open water pools arethe second major community wherePotamogeton strictus, Myriophyllum quitenseand Ranunculus sp. grow in the dark dissolvedorganic carbon-rich waters. A thirdcharacteristic community is more typical ofpeatlands in the Southern Andean Steppeecoregion. Members of the Poaceae, primarilyDeschampsia caespitosa and Deyeuxia velutinaare dominants with species of the Cyperaceae,such as Carex spp. and Eleocharis and otherJuncaceae including Juncus spp. are moresecondary representatives. All three of thesecommunities may grade into one anotherforming mixed communities.
The peatland vegetation is controlled byfour main interacting ecological factors: (a)water quantity and seasonal availability,especially during dry periods, (b) favourableambient temperatures and occurrence of frostevents that control the duration of the growingseason, (c) water pH, availability of nutrients(mainly, N, P, K, Ca and Mg), and exposure totoxic elements such as As, B, Fe, and Al in thewater, and (d) biotic factors such as seeddispersion by animals, grazing and humanimpacts (Villagrán et al. 1983, Ruthsatz 1993,2000, Villagrán & Castro 2003, Squeo et al.2006b). Our ongoing work is characterizing thedifferent peatlands communities over theirgeographic range to understand more fully theecological linkages between a variety ofecological parameters and the plant
TABLE 1
Characteristics of typical bofedales in Chile. These characteristics may not apply to bofedales thathave been modified or disturbed by humans
Características de los bofedales típicos de Chile. Estas características podrían no ser aplicables a bofedales que han sidomodificados o alterados por el hombre
Characteristic Description
Geologic/physiographic setting Situated on flat terrain, in sloping valley bottoms and shallow broad basins
Peatland landform Flat or slightly raised centre
Hydrology Predominantly groundwater, and some stream/river influence, snow melt
Water table position At or slightly below the surface
Water pH typically pH 7-8
Dominant vegetation Dominated by members of Juncaceae (i.e., Oxychloe andina, Patosia clandestina),with some Gramineae and other herbaceous species
Soil composition Poorly decomposed Juncaceae peat
252 SQUEO ET AL.
communities. Our research approach includestracing the history of past plant communitiesand peatland development, and understandingthe rate and magnitude of changes in plantcommunities and their relationships with pasthydrological conditions, actual water sourcesused by plants, and plant productivity andwater availability (Squeo et al. 1993, 2006b,Earle et al. 2003).
Age and development
Even though these peatlands are unique in theworld because of their geographic andenvironmental settings and the unusualvegetation cover dominated by compactcolonies of Juncaceae, they are like other peatlandforms in their structure and ability to formpeat. They possess the typical diplotelmic soilstructure with a near surface oxygen-rich layerand a deeper oxygen-poor zone. The preservedrecord in the peat lends itself well to usingpaleoecological methods to trace the age, timeof origin, and developmental sequence leadingto present day conditions. Unfortunately, therehave been few such studies (Earle et al. 2003,Rech et al. 2003).
A paleoecological investigation of a slopingpeatland developed in a stream channel in theNevado Tres Cruces National Park near thesouthern limit of the dry Puna zone at 4,300 mof alti tude. showed the peatland wasunexpectedly young, dynamic and sensitive toenvironmental changes (Earle et al. 2003). Atransect of cores along the longitudinal axis ofthe peatland revealed up to 3.6 m of peat,organic muck and inorganic sediments belowthe surface. Organic matter began toaccumulate around 1,000 years ago. Animportant question revolves around the delay inonset of peat accumulation given that the areais known to have been ice-free during theHolocene. Earle et al. (2003) proposed thatgiven the mid-Holocene climate is thought tohave been dry (Grosjean et al. 1997, Grosjean2001), the peatland started to develop after achange to more wet conditions during the lateHolocene. The intercalation of limnic and sandand gravel sediments in the early phases at thestudy site reveals a complex history of low andhigh energy surface flow through the valley. Itseems the peat-forming Oxychloe andinastarted to develop only after modern climate
conditions (slightly drier) became establishedand water supplies and the energy of surfaceflow diminished in the narrow valley andhydrological conditions became morequiescent. It is also possible that intrinsicprocesses such as rapid peat growth in lowenergy stretches of the valley contributed to theestablishment of the present O. andinadominated wetland. Internal factors are also inlarge part responsible for peat degradationcurrently underway at the Nevado Tres Crucespeatland site (Earle et al. 2003).
We can confirm that onset of modern-daypeatland development has been within the lastcentury or two elsewhere in Chile, near thesouthern limit of bofedales (Warner et al.unpublished results). Cores from a basinpeatland in the Río Tres Quebradas (29º16’ S,70’04’ W) valley impounded behind an endmoraine produced ages of around 6,600 yearsold, however, the Oxychloe communities beganto spread out across the basin about 1,200 yearsago. It appears that peatlands in this part ofChile are relatively recent features too, and donot represent old ecosystems formed during theearly Holocene as is usually assumed forpeatlands with thick accumulations of peat suchas the Sphagnum-dominated systems in the southof Chile (Arroyo et al. 2005) or elsewhere in thenorthern hemisphere. Much more needs to belearned about how such water-dependantfeatures can form and are maintained in sucharid environments. In another of the fewexamples where peatlands have beenradiocarbon dated, we have found a complexstratigraphy of peat, sand and gravel, clay, andmarl deposits up to 15 m in thickness under abasin peatland in Collacaqua (20°00’ S, 68°45’W), near the centre of the dry Puna zone(Warner & Aravena, unpublished results). Aradiocarbon date of 8,600 yrs BP was obtainedon the bottom of the section, but it seems theOxychloe peat of the modern-day peatlandstarted to accumulate around 3,000 BP andbecame well established about 1,400 years ago,which is comparable to the record at the NevadoTres Cruces site about 800 km to the south.
The presence of old wetlands depositsformed at various times during the Holocenehas been documented by Grosjean et al. (1997)and Rech et al. (2002, 2003) in the Salar deAtacama basin. Grosjean et al . (1997)postulated that the middle Holocene wetlands,
253PEATLANDS OF THE CENTRAL ANDES
initiated during dry regional conditions, weredue to a rise in the local water table caused bydamming of the river canyon downstream ofthe wetlands. Rech et al. (2003) obtained awide range of dates in the same river canyonstudied previously by Grosjean and postulatedthat the wetlands were formed during highwater table conditions produced during a periodof wet regional conditions. These contrastinginterpretations (Grosjean 2001) highlight theneed for further studies to evaluate if thegroundwater regime that made possible theexistence of these Holocene wetlands wasrepresentative of a local groundwater flowsystem or the regional groundwater flowsystem. Recent evidence based on 18O and 2Hdata obtained in rivers and wetlands at differentaltitudes in the high part of the Huasco valleyseem to show that sloping wetlands found nearthe rivers are associated with a local flowsystems (Aravena et al. unpublished results).However, it is more likely that the largewetlands found at the foot of the Andesrepresentative of the types found around thehigh altitude salares are fed by regionalgroundwater flow systems.
Future of peatlands and management
The peatland in the Nevado Tres CrucesNational Park exhibits obvious degradation onits downslope end (Earle et al. 2003). Thestratigraphic analyses and dating suggest thatthe peatland played an increasing role in waterretention as the Oxychloe andina vegetationexpanded and accumulated peat under it. Theporous nature and extremely compact growth ofthe vegetation impeded drainage andevaporation probably contributed to retentionof large volumes of water, mostly during springrunoff. It is thought that further vegetationgrowth and peat accumulation would reduceflows and evaporative water losses over timeand likely reduced through flow to the lowerextremities of the peatland. Earle et al. (2003)have shown that autoregulation processesplayed a significant role in growth anddegradation of bofedales and it has to be takeninto account in studies dealing with theassessment of human impact or climate in thedevelopment of bofedales.
What is the future of peatlands elsewhere inthis water-stressed region, especially in light of
precipitation decreasing to nearly 50 % of whatit was 100 years ago in north-central Chile?Global climatic models (GCM) suggest thatprecipitation will continue to decrease at thesame rate over the next 50 years. We know thatplant species in the peatlands ecosystems arelong-lived species (a minimum of 20-50 years;Squeo et al. 1996a, 2006a). Radiocarbon datingof peat records reveal that instead of beingancient ecosystems that originated immediatelyafter deglaciation, peatlands are extremelyyoung features arising when local hydrologicaland geological factors come together to favourestablishment. Are the peatlands relicts ofancient ecosystems or are they temporaryecosystems that come and go as local wateravailability conditions change? In the presentscenario of reduced precipitation, the slopingpeatlands associated with discharge of localgroundwater flow system will be much moresensitive to water availability than the wetlandsassociated with regional groundwater flowsystems.
One important observation is emerging thatmay help to explain the climatic sensitivity ofthese peatlands in northern Chile. In thesouthern Andean steppe, measurements of plantbiomass production from year to year showchanges in response to water availability andlength of the growing season that are directlycontrolled by El Niño Southern Oscillation(ENSO) phenomena (Squeo et al. 2006a,2006b). In the winter rain region, El Niño(rainy) years increase water availability butreduce the growing season due to higheraccumulation of snow over the peatlands whichresults in lower plant productivity. Maximumplant productivity occurs in the summerimmediately following an El Niño summer,when water availability remains high and thelength of growing season remains the same. Wecan, therefore, expect that primary productivityin these peatlands, and hence capacity to formand accumulate peat will decrease after severalsuccessive La Niña (dry) years because of anexpected decrease in water availability. Aseries of successive La Niña years, wouldtherefore contribute to peatland degradation.However, winter rain has been declining duringthe last century in Chile, while summer rainsare globally increasing (Houghton et al. 2001).There may be an opposite trend in the quantityof precipitation and response of the bofedales
254 SQUEO ET AL.
vegetation depending on El Niño/La Niña northof the Arid Diagonal.
If autoregulation processes of the localhydrological cycle also are contributing todegradation of parts of the peatlands systemtoo, we can expect to see major decline inextent and accelerated deterioration of thedelicate peatlands ecosystems south of the AridDiagonal in the central Andes of Chile andArgentina. Much more studies are required tofurther understand the relationship betweenwater sources (regional versus localgroundwater flow systems) and plantproductivity in bofedales. This information iscrucial for the long term management of thesefragile ecosystems in a future scenario ofdecreasing precipitation and increasing waterdemand by the mining and agriculture sectors.
ACKNOWLEDGEMENTS
Financial support from the NationalCommission for Scientific and TechnicalResearch of Chile (CONICYT) and the NaturalSciences and Engineering Research Council ofCanada (NSERC) is gratefully acknowledged.
LITERATURE CITED
ABRAHAM EM, K GARLEFF, H LIEBRICHT, ACREGAIRAZ, F SCHÄBITZ, FA SQUEO, HSTINGL, H VEIT, & C VILLAGRÁN (2000).Geomorphology and paleoecology of the ariddiagonal in Southern South America. Zeitschrift fürAngewandte Geologie: Sonderheft SH 1: 55-61.
ACEITUNO P (1997) Aspectos generales del clima en elAltiplano sudamericano. In: González C (ed) ElAltiplano: Ciencia y Conciencia en los Andes: 63-69. Vicerrectoría Académica y Estudianti l ,Universidad de Chile, Santiago.
ARROYO MTK, FA SQUEO, J ARMESTO & CVILLAGRÁN (1988) Effects of aridity on plantdiversity in the northern chilean Andes: results of anatural experiment. Annals of the MissouriBotanical Garden 75: 55-78.
ARROYO MTK, JJ ARMESTO, FA SQUEO & JRGUTIÉRREZ (1993) Global change: flora andvegetation of Chile. In: Mooney H, E Fuentes & BKronberg (eds) Earth system responses to globalchange: contrast between North and South America:239-263. Academic Press, New York, New York,USA.
ARROYO MTK, FA SQUEO, H VEIT, L CAVIEDES, PLEÓN & E BELMONTE (1997) Flora andvegetat ion of Northern Chi lean Andes . In:González C (ed) El altiplano: ciencia y concienciaen los Andes: 167-178. Universidad de Chile,Santiago.
ARROYO MTK, FA SQUEO, L CAVIERES, & CMARTICORENA (2004) Chilenische Anden. In:Burga CA, F Kötzli & G Grabherr (eds) Gebirge derErde: Landschaft, Klima, Pflanzenwelt: 210-219.Eugen Ulmer Gmb and Co., Stuttgart, Germany.
ARROYO MTK, P PLISCOFF, M MIHOC & M KALIN-ARROYO (2005) The magellanic moorland. In:Fraser LH & PA Keddy (eds) The World’s largerwetlands: ecology and conservation: 424-445.Cambridge University Press, Cambridge, UnitedKingdom.
CHARRIER R (1997) Ciencias de la tierra y recursosmineros y energéticos en el Altiplano Chileno. In:González C (ed) El Altiplano: Ciencia y Concienciaen los Andes: 5-14. Universidad de Chile, Santiago.
CLAYTON JD & CM CLAPPERTON (1997) Broadsynchrony of a late-glacial advance and thehighstand of palaeolake Tauca in the BolivianAltiplano. Journal of Quaternary Science 12: 169-182.
DIRECCIÓN GENERAL DE AGUAS (1996) Delimitaciónde acuíferos de vegas y bofedales de las regiones deTarapacá y Antofagasta. Dirección General deAguas, Ministerio de Obras Públicas, Gobierno deChile, Santiago, Chile. 169pp.
EARLE L, BG WARNER & R ARAVENA (2003) Rapiddevelopment of an unusual peat-accumulatingecosystem in the Chilean Altiplano. QuaternaryResearch 59: 2-11.
GARREAUD R, M VUILLE & AC CLEMENT (2003) Theclimate of the Altiplano: observed current conditionsand mechanisms of past changes. Palaeogeography,Palaeoclimatology, Palaeoecology 194: 5-22.
GROSJEAN M (2001) Mid-Holocene climate in the south-central Andes: humid or dry? Science 292: 2391.
GROSJEAN M, BL VALERO-GARCÉS, MA GEYH, BMESSERLI, U SCHOTTERER, H. SCHRIER, & KKELTS (1997) Mid- and late-Holocenelimnogeology of Laguna del Negro Francisco,northern Chile, and its paleoclimatic implications.Holocene 7:151-159.
HASTENRATH S & J KUTZBACH (1985) LatePleistocene climate and water budget of the SouthAmerican Altiplano. Quaternary Research 24:249-256.
HOUGHTON JT, Y DING, DJ GRIGGS, M NOGUER, PJVAN DER LINDEN, X DAI, K MASKELL & CAJOHNSON (2001) Climate change 2001: thescientific basis, contribution of Working Group I tothe Third Assessment Report of theIntergovernmental Panel on Climate Change(IPCC). Cambridge University Press, Cambridge,United Kingdom. 881 pp.
IRIARTE S, I UGALDE & M VENEGAS (1998)Hidrogeología de la cuenca Laguna del NegroFrancisco (Región de Atacama). Servicio Nacionalde Geología y Minería, Ministerio de Economía,Gobierno de Chile, documento de trabajo 10,Santiago, Chile.
MARTICORENA C & M QUEZADA (1985) Catálogo dela flora vascular de Chile. Gayana Botánica 42: 1-155.
MARTICORENA C & R RODRÍGUEZ (1995) Flora deChile. Ediciones de la Universidad de Concepción,Concepción, Chile. 351pp.
MARTICORENA C, O MATTHEI, R RODRÍGUEZ, MTKARROYO, M MUÑOZ, FA SQUEO & GARANCIO (1998) Catálogo florístico de la Regiónde Antofagasta. Gayana Botánica (Chile) 55: 23-83.
MARTICORENA C, FA SQUEO, G ARANCIO & M
255PEATLANDS OF THE CENTRAL ANDES
MUÑOZ (2001) Catálogo de la flora de la IVRegión. In: Squeo FA, G Arancio & JR Gutiérrez(eds) Libro rojo de la flora nativa de la Región deCoquimbo y de los sitios prioritarios para suconservación: 105-142. Ediciones de la Universidadde La Serena, La Serena, Chile.
MESSERLI B, M GROSJEAN, G BONANI, A BURGI,MA GEYH, K GRAF, K RAMSEYER, HROMERO, U SCHOTTERER, H SCHREUER & MVUILLE (1993) Climate change and naturalresource dynamics of the Atacama Altiplano duringthe last 18,000 years: a preliminary synthesis.Mountain Research and Development 13: 117-127.
MILLER A (1976) The climate of Chile. In: SchwerdtfegerW (ed) Climate of Central and South America: 113-129. Elsevier Scientif ic Publishing Co.,Amsterdam, The Netherlands.
OLSON DM, E DINERSTEIN, ED WIKRAMANAYAKE,ND BURGESS, GVN POWELL, ECUNDERWOOD, JA D’AMICO, I ITOUA, HESTRAND, JC MORRISON, CJ LOUCKS, TFALLNUTT, TH RICKETTS, Y KURA, JFLAMOREUX, WW WETENGEL, P HEDAO & KRKASSEM (2001) Terrestrial ecoregions of the world:a new map of life on earth. Bioscience 51: 933-938.
RECH JA, J QUADE & JL BETANCOURT (2002) LateQuaternary paleohydrology of the central Atacamadesert (lat 22o -24o), Chile. Geological Society ofAmerica Bulletin 114: 334-348.
RISARCHER F, H ALONSO & C SALAZAR (1999)Geoquímica de aguas en cuencas cerradas: I, II y IIIregión, Chile. Dirección General de Aguas -Universidad Católica del Norte - Institut deRecherche Pour le Developpement. 89pp.
RUTHSATZ B (1993) Flora and ecological conditions ofhigh Andean peatlands of Chile between 18º00’(Arica) and 40º30’ (Osorno) south lat i tude.Phytocoenologia 25: 185-234.
RUTHSATZ B (2000) Die Hartpolstermoore derHochanden und ihre Artenviefalt. Ber. D. Reinh.-Tüxen-Ges. 12: 351-371.
SQUEO FA, J CEPEDA-PIZARRO & JE NOVOA (1989)Estudio del impacto minero sobre la flora y faunaandina: ejemplo de la relación empresa-universidad.Anales del Tercer Encuentro Científico sobre elMedio Ambiente (Chile) 1: 405-413.
SQUEO FA, H VEIT, G ARANCIO, JR GUTIÉRREZ,MTK ARROYO & N OLIVARES (1993) Spatialheterogeneity of high mountain vegetation in theandean desert zone of Chile (30º S). MountainResearch and Development 13: 203-209.
SQUEO FA, R OSORIO & G ARANCIO (1994) Flora delos Andes de Coquimbo: cordillera de Doña Ana.Ediciones de la Universidad de La Serena, LaSerena, Chile. 176 pp.
SQUEO FA, CE RADA, ME GARCÍA, AL PONCE, ALROJAS & A AZÓCAR (1996) Cold resistancemechanisms in high desert Andean plants.Oecologia 105: 552-555.
SQUEO FA, LA CAVIERES, G ARANCIO, JE NOVOA,O MATTHEI, C MARTICORENA, RRODRÍGUEZ, MTK ARROYO & M MUÑOZ(1998) Biodiversidad de la flora vascular en laRegión de Antofagasta, Chile. Revista Chilena deHistoria Natural 71: 571-591.
SQUEO FA, Y TRACOL, D LÓPEZ, JR GUTIÉRREZ,AM CÓRDOVA & JR EHLERINGER (2006a)ENSO effects on primary productivity in SouthernAtacama desert. Advances in Geosciences 6: 273-277.
SQUEO FA, E IBACACHE, BG WARNER, D ESPINOZA,R ARAVENA & JR GUTIÉRREZ (2006b)Productividad y diversidad florística de la Vega LosTambos, Cordillera de Doña Ana: variabilidad inter-anual, herbivoría y nivel freático. In: Cepeda J (ed)Geoecología de la Alta Montaña del Valle del Elqui:333-362. Ediciones Universidad de La Serena, LaSerena, Chile.
STONE PB (1992) The state of the world’s mountains: aglobal report. Zed Books Ltd., London, UnitedKinwdom. 391 pp.
VEIT H (1996) Southern westerlies during the Holocenededuced from geomorphological and pedologicalstudies in the Norte Chico, northern Chile (27-33°S). Palaeogeography, Palaeoclimatology,Palaeoecology 123: 107-119.
VILLAGRÁN C & V CASTRO (1997) Etnobotánica ymanejo ganadero de las vegas, bofedales yquebradas en el Loa superior, Andes deAntofagasta, Segunda Región, Chile. Chungará(Chile) 29: 275-304.
VILLAGRÁN C & V CASTRO (2003) Ciencia indígenade los Andes del norte de Chile. EditorialUniversitaria, Santiago, Chile. 362 pp.
VILLAGRÁN C, MTK ARROYO & C MARTICORENA(1983) Efectos de la desertización en la distribuciónde la flora andina de Chile. Revista Chilena deHistoria Natural 56: 137-157.
VILLAGRÁN C, V CASTRO, G SÁNCHEZ, FHINOJOSA & C LATORRE (1999) La tradiciónaltiplánica: estudio etnobotánico en los Andes deIquique, Primera Región, Chile. Chungará (Chile)13: 81-186.
VILLAGRÁN C, M ROMO & V CASTRO (2003)Etnobotánica del sur de los Andes de la PrimeraRegión de Chile: un enlace entre las culturasaltiplánicas y las de quebradas altas del Loasuperior. Chungará (Chile) 35: 73-124.
VUILLE M & F KEIMIG (2004) Interanual variability ofsummertime convective cloudiness andprecipitation in the central Andes derived fromISCCP-B3 data. Journal of Climate 17: 3334-3348.
VUILLE M & C AMMANN (1997) Regional snowfallpatterns in the high, arid Andes. Climatic Change36: 413-423.
WILCOX BP, MK WOOD, JT TROMBLE, & TJ WARD(1986) Grassland communities and soils on a highelevation grassland of central Peru. Phytologia 61:231-250.
WWF (2001) Terrestrial biorealms: Neotropical. WorldWildlife Fund, Glanz, Switzerland.(http:www.worldwildlife.org/wildworld/profiles)
Associate Editor: Javier FigueroaReceived January 8, 2005; accepted January 30, 2006















![Characterization and Comparison of Microbial Soil ...bofedales, vegas and bogs), which technically belong to a humid environment [7]. Peatlands are plant formations that are established](https://static.documents.pub/doc/80x56/5e99481d14e4f47d8f42020e/characterization-and-comparison-of-microbial-soil-bofedales-vegas-and-bogs.jpg)











