Bone Sialoprotein (BSP) is a Crucial Factor for the Expression ofOsteoblastic Phenotypes of Bone Marrow Cells Cultured on Type ICollagen Matrix
M. Mizuno,1 T. Imai,2 R. Fujisawa,1 H. Tani,2 Y. Kuboki 1
1Department of Biochemistry, School of Dentistry, Hokkaido University, Sapporo, 060 Japan2Department of Preventive Dentistry, School of Dentistry, Hokkaido University, Sapporo, Japan
Received: 17 February 1999 / Accepted: 14 December 1999
Abstract. In this study, we demonstrated that type I colla-gen matrix induced the expression of osteoblastic pheno-types of bone marrow cells, and that antibone sialoprotein(BSP) monoclonal antibody suppressed the expression ofthese phenotypes. On the other hand, BSP accelerated theexpression of osteoblastic phenotypes of bone marrow cells.The adherent bone marrow cells were harvested from ratfemur and cultured on type I collagen matrix gels in me-dium containing 15% fetal calf serum, neitherb-glycero-phosphate nor glucocorticoid. Cells showed osteoblasticphenotypes (high alkaline phosphatase activity, osteocalcinsynthesis, and responsiveness against parathyroid hormone)on collagen matrix gels at week 3 after the inoculation, andsimultaneously, BSP was detected in the conditioned me-dium by Western blotting using an anti-BSP monoclonalantibody. However, cells in the conventional culture dishesdid not show osteoblastic phenotypes during the experimen-tal period. To investigate the physiological function of BSPin osteoblastic differentiation, bone marrow cells were cul-tured on collagen matrix with an anti-BSP monoclonal an-tibody for 3 weeks. This treatment suppressed the expres-sion of the osteoblastic phenotypes, and the effect of theantibody was abolished by the addition of bovine bone BSP.Furthermore, bovine bone BSP stimulated the expression ofosteoblastic phenotypes of bone marrow cells. Our resultsindicate that BSP plays a crucial role in the expression ofosteoblastic phenotypes of bone marrow cells.
Key words: Bone sialoprotein — Osteoblasts — Bonemarrow cells.
Bone sialoprotein (BSP) is produced by osteoblasts [1–3],odontoblasts [4], and bone marrow stromal cells treatedwith dexamethasone [5]. Cells accumulate BSP in the ex-tracellular matrix, and the distribution of BSP is restricted tobone and mineralized tissuesin vivo [6–8]. In bone, BSP isone of the major noncollagenous proteins and is present atthe bone matrix [9]. BSP is characterized by the presence ofan Arg-Gly-Asp (RGD) sequence that binds to the integrinreceptors of cells [10, 11] and stimulates cell attachment[12]. This protein also contains a poly-Glu segment that issuggested to be a hydroxyapatite-binding region [13, 14].
From these features, BSP has been considered to play a rolein the maintenance of bone cells on mineralized matrix, andto be a nucleator of hydroxyapatite crystal precipitation[14]. Recently, we found that BSP stimulates the expressionof osteoblastic phenotypes of MC3T3-E1 osteoblastic cells[15]. This finding indicates that BSP regulates the expres-sion of osteoblastic phenotypes of bone cells.
The aim of this study was to investigate the function ofBSP in osteoblastic differentiation. We cultured bone mar-row cells with a specific antibody against BSP and foundthat the anti-BSP antibody suppressed the osteoblastic dif-ferentiation of bone marrow cells. We could thus concludethat BSP has a crucial role in the osteoblastic differentiationof bone marrow cells.
Materials and Methods
Biochemicals
Type I collagen was purchased from Nitta Gelatin Co. (Japan). Ratparathyroid hormone (PTH) and the cAMP enzyme immunoassaykit were purchased from Cosmo Bio Co. (Japan) and AmershamCo. (Japan), respectively.
Production of Anti-BSP Monoclonal Antibody
BALB/C mice were injected intraperitoneally with 50 mg of boneBSP that was purified from bovine bone, as described previously[15], and dissolved in phosphate-buffered saline (PBS) withoutadjuvant. The mice were boostered with 25 mg of antigen on days7 and 14 after the first injection. Three days after the third injec-tion, the mice were sacrificed and their spleens were removedunder sterile conditions. Hybridomas were prepared by the fusionof spleen cells with X63-AG8.653 myeloma cells in the presenceof 50% polyethylene glycol (Mr4 103). The media containinganti-BSP antibody from the hybridoma colonies were assayed byenzyme-linked immunoassay (ELISA) using purified bovine BSP.Media from wells showing a single colony were screened againstan extract of bovine bone, and the positive colonies were sub-cloned again. The hybridomas producing BSP antibody were suc-cessfully converted to ascites tumors by injecting hybridoma intothe peritoneal cavity of mice. Mice showed the evidence of tumorgrowth in approximately 7–10 days. Ascites fluid was obtainedunder sterile conditions on the 10th day. Immunoglobulin fromascites fluid was purified by affinity chromatography on a columnof BSP conjugated to Agarose (Biorad, Rockville Center, NY).Bound immunoglobins were eluted with 3.5 M sodium thiocya-Correspondence to:M. Mizuno
nate. The eluted antibody was dialyzed against two changes ofPBS and evaluated by direct ELISA for antibody activity.
Culture of Adherent Bone Marrow Cells
The adherent bone marrow cells were harvested from the femurs of6-week-old male Wister rats as described previously [16]. Thefemur was harvested aseptically and soft tissue was removed. Thenthe femur was passed through four washes of 10 minutes each witha culture medium containing kanamycin sulfate (600mg/ml) andpenicillin G (1000 U/ml). The marrow cells were flushed out usinga 5-ml syringe with an 18-gauge needle. The cells were collectedin a 25-cm2 plastic culture flask (Falcon Labware, Oxnard, CA)containing 5 ml of culture medium, and cultured until adherentbone marrow cells reached the subconfluent stage. Culture me-dium consisted of thea-modification of Eagle’s minimum essen-tial medium (aMEM), and 15% fetal calf serum and antibiotics(100 U/ml of penicillin G, and 60mg/ml of kanamycin sulfate).Adherent bone marrow cells (5 × 105) were plated on type I col-lagen matrix gel formed in a 35 mm dish and cultured. We used0.5% of type I collagen solution (Nitta Gelatin Co., Japan) and thecollagen concentration was adjusted to 0.35% before cell inocula-tion. Cells were maintained in a humidified atmosphere of 95% airand 5% CO2 at 37°C and the medium was changed every 2 days.For the experiment, one group of cells was cultured witha-MEMcontaining the anti-BSP monoclonal antibody, and the controlgroup of cells was cultured in medium containing mouse immu-noglobulin. The reactivity of the antibody was confirmed byELISA assay using bovine bone BSP.
Measurement of Alkaline Phosphatase (ALP) Activity andContent of DNA
The cell-matrix of bone marrow cells was washed in PBS threetimes to remove serum proteins, and was homogenized with 10mM Tris/HCl buffer (pH 7.4) containing 1 mM MgCl2, and uti-lized for the measurement of ALP activity [17]. One unit wasestimated to be 1.4 mg of p-nitrophenol liberated from p-nitrophenyl phosphate in 15 minutes. DNA content was deter-mined using bisbenzimidazole (Hoechst 33258) [18]. Briefly, cellpellets were homogenized with 50 mM Na2HPO4/2.0 M NaCl (pH7.4) and mixed with Hoechst 33258. The intensity of fluorescencewas measured at an excitation wavelength of 356 nm and emissionwavelength of 458 nm. All standard stock solution was stored at−20°C until use.
Western Blotting
Bone marrow cells were cultured on collagen gels for 1, 2, or 3weeks witha-MEM containing 15% fetal calf serum. Then cellswere cultured in serum-free medium for 24 hours and medium washarvested for Western blot analysis, followed by the concentrationby Centricon (Amicon, Japan). The medium was separated bySDS-polyacrylamide gel electrophoresis (SDS-PAGE) using12.5% gel, and transferred onto nitrocellulose membranes for 1hour at 30 mA. The membranes were blocked by incubation with20 mM Tris/500 mM NaCl (pH 7.5) containing 3% albumin for 30minutes at room temperature. The membrane was incubated withthe antibody against BSP overnight. Following washing and incu-bation of the membrane with horseradish peroxidase-conjugatedProtein A (1:1000), the membrane was developed using 3,3-diaminobenzidine.
Measurement of Osteocalcin Content in Conditioned Medium
Mouse osteocalcin was labeled with125I using Bolton-Hunter re-agent [19], and osteocalcin content in the medium was measuredby commercial radioimmunoassay (BTI, USA).
Northern Blotting
Total RNA was extracted from rat bone marrow cells using theacid guanidine-phenol-chloroform method [20]. Twenty micro-grams of RNA was separated by 1.4% denaturing agarose gelelectrophoresis in MOPS buffer. Then RNAs were transferred ontoa nylon membrane (Zeta Probe, Biorad) with a transblotter (Strata-gene, La Jolla, CA). The 300 bp cDNA probe of rat osteocalcinwas constructed by RT-PCR using oligonucleotide primers (for-ward: ATGAGGACCCTCTCTCTG, reverse: CTAAACGGTG-GTGCCATA), whose basic sequence was determined from the ratosteocalcin gene [21] and labeled with DIG-11-dUTP using a ran-dom primer labeling kit (Takara, Japan). Hybridization to RNAimmobilized on the nylon membrane was performed for 15 hoursat 50°C with a probe containing buffer (50% formamide, 5× SSC,50 mM NaPO4 (pH 7.0), 7% SDS, 2% blocking buffer, 0.1%lauroylsarcosine, and 100mg/ml of salmon sperm DNA) afterprehybridization at 50°C for 1 hour in the same buffer without theprobe. Membranes were washed in 2× SSC (1× SSC is 180 mMNaCl and 15 mM sodium citrate) containing 0.1% SDS for 10minutes at room temperature, followed by 0.1× SSC containing0.1% SDS for 10 minutes at 68°C. The detection of signals wasperformed by contact of the membrane with X-P film for 5 hours.
Responsiveness Against PTH of Bone Marrow Cells
Bone marrow cells were cultured with serum-free medium for 12hours and incubated with 1.0 mM isobutyl methylxanthine(IBMX) in serum-free medium for 15 minutes. Thereafter, cellswere incubated with 5 × 10−8 M 1-34 rat PTH and 1.0 mM IBMXfor 5 minutes. Cells were homogenized with 1 ml of phosphate-buffered saline and centrifuged at 4000 g for 20 minutes. Thesupernatant was harvested and washed with ethyl ether three times.The cAMP content in the supernatant was measured by immuno-assay, and referred to as the content of intracellular cAMP.
Results
Expression of Osteoblastic Phenotypes of Bone Marrow CellsCultured on Type I Collagen Matrix
We first investigated the morphological changes of bonemarrow cells in the plastic culture dishes or on the collagenmatrix gels by phase-contrast microscopy. Cells showed afibroblastic form in the plastic dishes from 1 to 3 weeksafter cells were inoculated (Fig. 1A). However, cells did notshow a definite fibroblastic form on the collagen matrixafter 1 week (Fig. 1B). At week 2, nodules were detected onthe collagen matrix (Fig. 1C) and precipitates were recog-nized in the nodules at 3 weeks (Fig. 1D). These noduleswere positive by von Kossa staining, indicating that pre-cipitates contained calcium. The morphological observationindicates that bone marrow cells formed mineralized tis-sues, a finding confirmed by measuring calcium content inthe matrix. Calcification proceeds by the precipitation ofhydroxyapatite, and a high content of calcium indicates ahigh magnitude of mineralization. At week 3, high calciumcontent was detected in the matrix, which meant that min-eralization in the cell layers occurred (Fig. 2). However, atweeks 1 and 2, calcium content was low (Fig. 2) and min-eralized tissues were not formed. Cells in the plastic dishesshowed low calcium content until 3 weeks (Fig. 2).
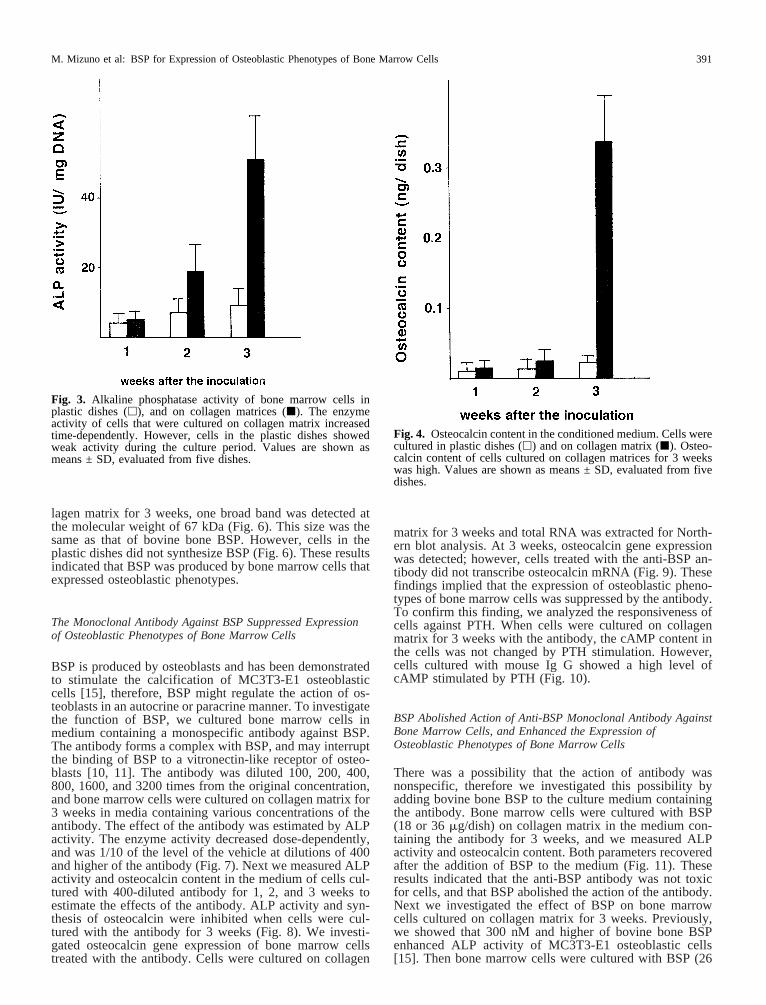
Mineralized tissue formation is a typical osteoblasticphenotype. Next we measured ALP activity and osteocalcincontent in the medium. High ALP activity is an osteoblasticphenotype. Its activity of cells cultured on collagen matrixwas low at 1 week and increased time-dependently. At week3, the enzyme activity was 12-fold higher than that of cells
M. Mizuno et al: BSP for Expression of Osteoblastic Phenotypes of Bone Marrow Cells 389
at week 1 (Fig. 3). However, cells in the plastic dishesshowed a low level of ALP activity until 3 weeks (Fig. 3).Osteocalcin synthesis is another osteoblastic phenotype andmore than 80% of total osteocalcin is secreted into the me-dium [22]. When bone marrow cells were cultured on col-lagen matrix for 3 weeks, osteocalcin was secreted into themedium (Fig. 4). We also investigated the responsiveness ofcells against parathyroid hormone (PTH) which elevates thelevel of cAMP in osteoblasts [23–25]. This is due to thepresence of PTH receptors on the cell membrane of osteo-blasts [26, 27], and this receptor is specific for osteoblasts.Bone marrow cells cultured for 1 and 2 weeks did not re-spond to PTH; however, cells cultured for 3 weeks on col-lagen matrix showed high cAMP content after the stimula-tion with PTH (Fig. 5). These findings demonstrated thatbone marrow cells cultured on collagen matrix for 3 weeksexpressed osteoblastic phenotypes.
Bone Marrow Cells Produced Bone Sialoprotein (BSP)
BSP is produced by osteoblasts and odontoblasts, and isconsidered to be a marker of osteoblastic phenotypes.Therefore, we attempted to detect BSP by Western blottingusing a monospecific antibody. The conditioned mediumwas concentrated and proteins in the medium were sepa-rated out by SDS-PAGE. After cells were cultured on col-
Fig. 1. Phase-contrast microscopic observation of bone marrowcells cultured for 3 weeks in conventional plastic dishes(A), andon type I collagen matrices for 1 week(B), 2 weeks(C), and 3
weeks(D). At week 2, cells on collagen matrix formed nodules(arrowhead), and at week 3, mineralized tissues were observed(arrow).
Fig. 2. Calcium contents in the cell matrix. Bone marrow cellswere cultured in conventional culture dishes (h) and on type Icollagen matrix (j). At week 3, matrices of cells cultured oncollagen gels showed high calcium content. Data are shown asmeans ± SD, evaluated from five dishes.
M. Mizuno et al: BSP for Expression of Osteoblastic Phenotypes of Bone Marrow Cells390
lagen matrix for 3 weeks, one broad band was detected atthe molecular weight of 67 kDa (Fig. 6). This size was thesame as that of bovine bone BSP. However, cells in theplastic dishes did not synthesize BSP (Fig. 6). These resultsindicated that BSP was produced by bone marrow cells thatexpressed osteoblastic phenotypes.
The Monoclonal Antibody Against BSP Suppressed Expressionof Osteoblastic Phenotypes of Bone Marrow Cells
BSP is produced by osteoblasts and has been demonstratedto stimulate the calcification of MC3T3-E1 osteoblasticcells [15], therefore, BSP might regulate the action of os-teoblasts in an autocrine or paracrine manner. To investigatethe function of BSP, we cultured bone marrow cells inmedium containing a monospecific antibody against BSP.The antibody forms a complex with BSP, and may interruptthe binding of BSP to a vitronectin-like receptor of osteo-blasts [10, 11]. The antibody was diluted 100, 200, 400,800, 1600, and 3200 times from the original concentration,and bone marrow cells were cultured on collagen matrix for3 weeks in media containing various concentrations of theantibody. The effect of the antibody was estimated by ALPactivity. The enzyme activity decreased dose-dependently,and was 1/10 of the level of the vehicle at dilutions of 400and higher of the antibody (Fig. 7). Next we measured ALPactivity and osteocalcin content in the medium of cells cul-tured with 400-diluted antibody for 1, 2, and 3 weeks toestimate the effects of the antibody. ALP activity and syn-thesis of osteocalcin were inhibited when cells were cul-tured with the antibody for 3 weeks (Fig. 8). We investi-gated osteocalcin gene expression of bone marrow cellstreated with the antibody. Cells were cultured on collagen
matrix for 3 weeks and total RNA was extracted for North-ern blot analysis. At 3 weeks, osteocalcin gene expressionwas detected; however, cells treated with the anti-BSP an-tibody did not transcribe osteocalcin mRNA (Fig. 9). Thesefindings implied that the expression of osteoblastic pheno-types of bone marrow cells was suppressed by the antibody.To confirm this finding, we analyzed the responsiveness ofcells against PTH. When cells were cultured on collagenmatrix for 3 weeks with the antibody, the cAMP content inthe cells was not changed by PTH stimulation. However,cells cultured with mouse Ig G showed a high level ofcAMP stimulated by PTH (Fig. 10).
BSP Abolished Action of Anti-BSP Monoclonal Antibody AgainstBone Marrow Cells, and Enhanced the Expression ofOsteoblastic Phenotypes of Bone Marrow Cells
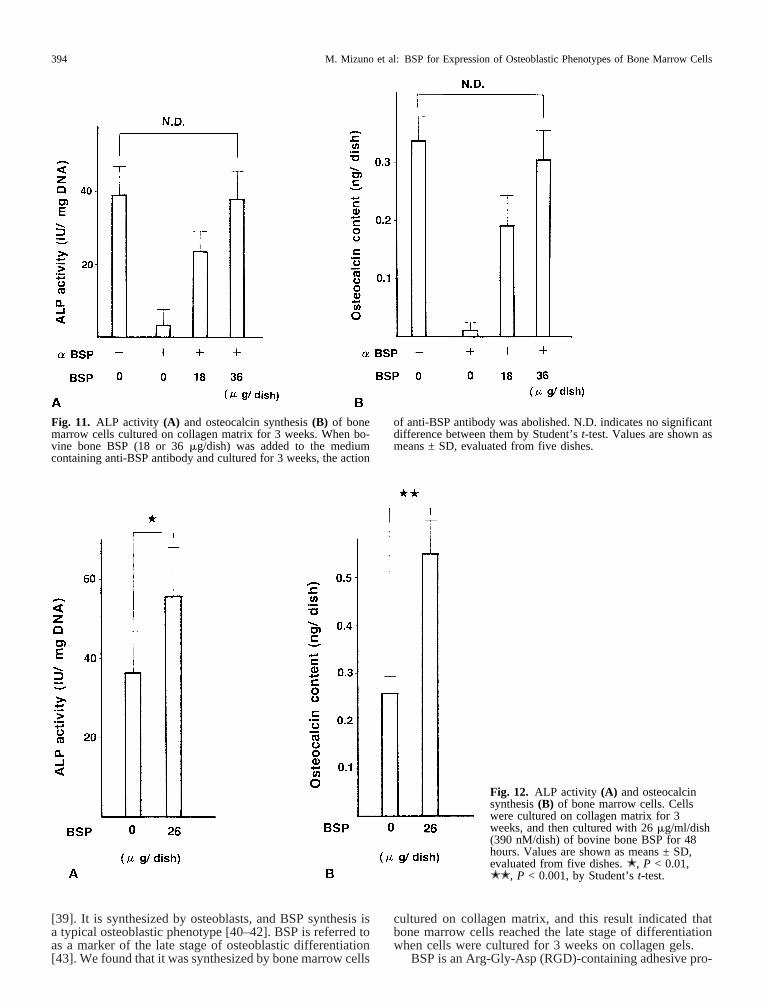
There was a possibility that the action of antibody wasnonspecific, therefore we investigated this possibility byadding bovine bone BSP to the culture medium containingthe antibody. Bone marrow cells were cultured with BSP(18 or 36mg/dish) on collagen matrix in the medium con-taining the antibody for 3 weeks, and we measured ALPactivity and osteocalcin content. Both parameters recoveredafter the addition of BSP to the medium (Fig. 11). Theseresults indicated that the anti-BSP antibody was not toxicfor cells, and that BSP abolished the action of the antibody.Next we investigated the effect of BSP on bone marrowcells cultured on collagen matrix for 3 weeks. Previously,we showed that 300 nM and higher of bovine bone BSPenhanced ALP activity of MC3T3-E1 osteoblastic cells[15]. Then bone marrow cells were cultured with BSP (26
Fig. 3. Alkaline phosphatase activity of bone marrow cells inplastic dishes (h), and on collagen matrices (j). The enzymeactivity of cells that were cultured on collagen matrix increasedtime-dependently. However, cells in the plastic dishes showedweak activity during the culture period. Values are shown asmeans ± SD, evaluated from five dishes.
Fig. 4. Osteocalcin content in the conditioned medium. Cells werecultured in plastic dishes (h) and on collagen matrix (j). Osteo-calcin content of cells cultured on collagen matrices for 3 weekswas high. Values are shown as means ± SD, evaluated from fivedishes.
M. Mizuno et al: BSP for Expression of Osteoblastic Phenotypes of Bone Marrow Cells 391
mg/ml/dish, 390 nM) for 48 hours and measured ALP ac-tivity and osteocalcin content in the medium. By this treat-ment, cells showed high ALP activity and osteocalcin con-tent in the medium (Fig. 12). These results indicated thatBSP accelerated the expression of osteoblastic phenotypes.
Discussion
The present study demonstrated that bone marrow cells cul-tured on type I collagen matrix gel expressed osteoblasticphenotypes, and that an anti-bone sialoprotein (BSP) mono-clonal antibody suppressed the expression of these pheno-types. Furthermore, bovine bone BSP enhanced the expres-sion of osteoblastic phenotypes of bone marrow cells.
Bone marrow cells on type I collagen matrix showed apolygonal form, and formed mineralized tissues at week 3after the inoculation. Bone marrow cells on collagen matrixshowed common morphological features of osteoblasts,since primary and established osteoblasts have a polygonalform [28, 29]. To confirm that bone marrow cells possessedthe osteoblastic features, we measured ALP activity andosteocalcin content in the medium. ALP activity and syn-thesis of osteocalcin are typical phenotypic markers of os-teoblasts. Osteocalcin is synthesized by osteoblasts at thestage of mineralized tissue formation [30]. Bone marrowcells cultured on type I collagen matrix showed high ALPactivity and high content of osteocalcin.
We investigated the responsiveness of cells against PTH.PTH responsiveness is known to correlate with expression
of osteoblastic phenotypes in a subpopulation of calvarialcells and in osteosarcoma cells [31, 32]. PTH receptors wererecognized on osteoblasts, and lining cells exist on bonematrix [26, 27], therefore, responsiveness against PTH is acommon feature of osteoblasts. In this study, we found high
Fig. 5. The intracellular cAMP content of cells stimulated byPTH. The cAMP level of cells that were cultured on collagenmatrices for 3 weeks, elevated by the stimulation with PTH (j).Values are shown as means ± SD, evaluated from three samples.
Fig. 6. Immunodetection of BSP in the conditioned medium ofbone marrow cells. Cells were cultured in plastic dishes for 1, 2,and 3 weeks (lanes 2, 4, and 6), or on collagen matrix for 1, 2, and3 weeks (lanes 3, 5, and 7). Bovine bone BSP was applied as thestandard (lane 1). Bone marrow cells cultured on collagen matrixfor 3 weeks produced BSP, and its molecular weight was the sameas that of bone BSP.
Fig. 7. Inhibition of ALP activity by various concentrations ofanti-BSP antibody. Bone marrow cells were cultured on collagenmatrices for 3 weeks with various concentrations of the antibody(diluted to 1/3200, 1/1600, 1/800, 1/400, 1/200, and 1/100 fromoriginal antibody solution). ALP activity decreased dose-dependently, and cells cultured with 1/400 diluted antibody solu-tion showed 1/10 of the activity of cells cultured without antibody.Values are shown as means ± SD, evaluated from three dishes.
M. Mizuno et al: BSP for Expression of Osteoblastic Phenotypes of Bone Marrow Cells392
intracellular cAMP content in bone marrow cells culturedon collagen matrix, and treated with PTH. These findingsindicated that bone marrow cells cultured on collagen ma-trix expressed the osteoblastic phenotypes.
Collagen matrices and other extracellular proteins havebeen used to support phenotypes and tissue-specific func-tions [33, 34], which indicates that extracellular matricesmaintain cell functions. The major organic component ofbone extracellular matrix is type I collagen, and it has beenshown that type I collagen supports the expression of bonecell phenotypes and enhances the mineralization [35]. Thesefindings are consistent with our results.
Bone marrow cells are the source of osteoblasts in boneand differentiate into osteoblasts after treatment with dexa-methasone [36] or vitamin C [37]. Furthermore, we foundthat bone marrow cells on collagen matrix form bonein vivo[38]. These findings indicate that bone marrow cells coulddifferentiate into osteoblasts as a result of the actions ofseveral soluble factors and extracellular matrix.
BSP is an extracellular matrix and a component of bone
Fig. 8. Inhibition of ALP activity (A) and osteocalcin production(B) by anti-BSP antibody. Bone marrow cells were cultured withanti-BSP antibody (h) and with mouse Ig G (j). The antibody
suppressed the expression of ALP and synthesis of osteocalcin atweek 3. Values are shown as means ± SD, evaluated from fivedishes.
Fig. 9. Expression of osteocalcin gene. Total RNA was extractedfrom bone marrow cells on collagen matrix for 3 weeks withanti-BSP antibody (lane 1) and with mouse Ig G (lane 2). Panel Ashows the RNA stained with ethidium bromide, and panel B dem-onstrates the expression of osteocalcin mRNA. Positions of a largesubunit and small subunit of ribosomal RNA, respectively, areindicated by 28S and 18S. Fig. 10. The intracellular cAMP content of cells stimulated by
PTH. Culture of cells with anti-BSP antibody caused responsive-ness against PTH to disappear. Values are shown as means ± SD,evaluated from three samples.
M. Mizuno et al: BSP for Expression of Osteoblastic Phenotypes of Bone Marrow Cells 393
[39]. It is synthesized by osteoblasts, and BSP synthesis isa typical osteoblastic phenotype [40–42]. BSP is referred toas a marker of the late stage of osteoblastic differentiation[43]. We found that it was synthesized by bone marrow cells
cultured on collagen matrix, and this result indicated thatbone marrow cells reached the late stage of differentiationwhen cells were cultured for 3 weeks on collagen gels.
BSP is an Arg-Gly-Asp (RGD)-containing adhesive pro-
Fig. 12. ALP activity (A) and osteocalcinsynthesis(B) of bone marrow cells. Cellswere cultured on collagen matrix for 3weeks, and then cultured with 26mg/ml/dish(390 nM/dish) of bovine bone BSP for 48hours. Values are shown as means ± SD,evaluated from five dishes.✭, P < 0.01,✭✭, P < 0.001, by Student’st-test.
Fig. 11. ALP activity (A) and osteocalcin synthesis(B) of bonemarrow cells cultured on collagen matrix for 3 weeks. When bo-vine bone BSP (18 or 36mg/dish) was added to the mediumcontaining anti-BSP antibody and cultured for 3 weeks, the action
of anti-BSP antibody was abolished. N.D. indicates no significantdifference between them by Student’st-test. Values are shown asmeans ± SD, evaluated from five dishes.
M. Mizuno et al: BSP for Expression of Osteoblastic Phenotypes of Bone Marrow Cells394
tein that mediates cell attachment of various cell typesthrough RGD-dependent as well as RGD-independent at-tachment [44, 45]. BSP also induces mineralization via theprecipitation of hydroxyapatite [13, 14]. These two func-tions indicate that BSP contributes to bone formation. Infact, BSP has a specific role during the initial phases of boneformation at the cartilage and bone interface [46]. Yao et al.[47] reported that addition of BSP to media induced theexpression of osteoblastic phenotypes, and we found thatBSP accelerated mineralization by MC3T3-E1 osteoblasticcells [15]. To investigate the function of BSP in the expres-sion of osteoblastic phenotypes, we attempted to abolish theinteraction of BSP with cells by the addition of a monospe-cific anti-BSP antibody to the medium. As a result of thistreatment, ALP activity, osteocalcin synthesis, and respon-siveness against PTH were inhibited. These findings areconsistent with the recent report that the anti-BSP antibodysuppressed the differentiation of osteoblasts [48]. Further-more, BSP enhanced ALP activity and osteocalcin synthesisof bone marrow cells, therefore, BSP is a crucial factor forthe osteoblastic differentiation.
BSP is an adhesive protein in extracellular matrix, andosteoblastic phenotypes are altered by adhesion protein-cellinteractions. For example, laminin-derived peptide pro-motes MC3T3-E1 osteoblastic cell differentiation [49], andfibronectin regulates osteoblastic differentiation [50, 51].These results support the findings that interactions of cellswith attachment proteins (BSP, laminin, fibronectin, etc.)could be a crucial signal for continued differentiation ofosteoblasts. These attachment proteins may substitute foreach otherin vivo because bone formation is not altered bya total lack of BSP expression in the BSP knockout mouse[52].
References
1. Ecarot-Charrier B, Bouchard F, Delloye C (1989) Bone sia-loprotein II synthesized by cultured osteoblasts contains tyro-sine sulfate. J Biol Chem 264:20049–20053
2. Nagata T, Bellows CG, Kasugai S, Butler WT, Sodek J (1991)Biosynthesis of bone proteins [SPP-1 (secreted phosphopro-tein-1. osteopontin), BSP (bone sialoprotein) and SPARC (os-teonectin)] in association with mineralized-tissue formationby fetal-rat calvarial cells in culture. Biochem J 274:513–520
3. Chen J, Shapiro HS, Wrana JL, Reimers S, Heersche JNM,Sodek J (1991) Localization of bone sialoprotein (BSP) ex-pression to sites of mineralized tissue formation in fetal rattissues by in situ hybridization. Matrix 11:133–143
4. Bianco P, Fisher LW, Young MF, Termine JD, Robey PG(1991) Expression of bone sialoprotein (BSP) in developinghuman tissues. Calcif Tissue Int 49:421–426
5. Kasugai S, Todescan R, Nagata T, Yao K-L, Butler WT,Sodek J (1991) Expression of bone matrix proteins associatedwith mineralized tissue formation by adult rat bone marrowcells in vitro: inductive effects of dexamethasone on the os-teoblastic phenotypes. J Cell Physiol 147:111–120
6. Franze´n A, Heinegård D (1985) Isolation and characterizationof two sialoproteins present only in bone calcified matrix.Biochem J 232:715–724
7. Fisher LW, Hawkins GR, Tuross N, Termine JD (1987) Pu-rification and partial characterization of small proteoglycans Iand II, bone sialoprotein I and II, and osteonectin from themineral compartment of developing human bone. J Biol Chem262:9702–9708
8. Fisher LW, McBride OW, Termine JD, Young MF (1990)Human bone sialoprotein. J Biol Chem 265:2347–2351
9. Chen J, Zhang Q, McCulloch CAG, Sodek J (1991) Immu-nohistochemical localization of bone sialoprotein in foetalporcine bone tissues: comparisons with secreted phosphopro-
tein-1 (SPP-1, osteopontin) and SPARC (osteonectin). Histo-chem J 23:281–289
10. Oldberg Å, Franzen A, Heinegård D (1988) The primarystructure of a cell-binding bone sialoprotein. J Biol Chem263:19430–19432
11. Oldberg A, Franzen A, Heinegård D, Pierschbacher M, Rous-lahti E (1988) Identification of a bone sialoprotein receptor inosteosarcoma cells. J Biol Chem 263:19433–19436
12. Somerman MJ, Fisher LW, Foster RA, Sauk JJ (1988) Humanbone sialoprotein I and II enhance fibroblast attachment invitro. Calcif Tissue Int 43:50–53
13. Hunter GK, Goldberg HA (1993) Nucleation of hydroxyapa-tite by bone sialoprotein. Proc Natl Acad Sci USA 90:8562–8565
14. Hunter GK, Goldberg HA (1994) Modulation of crystal for-mation by bone phosphoproteins: role of glutamic acid-richsequences in the nucleation of hydroxyapatite by bone sia-loprotein. Biochem J 302:175–179
15. Zhou H-Y, Takita H, Fujisawa R, Mizuno M, Kuboki Y(1995) Stimulation by bone sialoprotein of calcification inosteoblast-like MC3T3-E1 cells. Calcif Tissue Int 56:403–407
16. Hasegawa T, Oguchi H, Mizuno M, Kuboki Y (1994) Theeffect of the extracellular matrix on differentiation of bonemarrow stromal cells to osteoblasts. Jpn J Oral Biol 36:383–394
17. Kind PRN, King EJ (1954) Estimation of plasma phosphataseby determination of hydrolyzed phenol with amino-antipyrine.J Clin Pathol 7:322–326
18. Labrca C, Paigen K (1980) A simple, rapid, and sensitiveDNA assay procedure. Anal Biochem 102:344–352
19. Bolton AE, Hunter WM (1973) The labeling of proteins tohigh specific radioactivities by conjugation to a 125I-containing acylating agent. Biochem J 133:529–539
20. Chomczynski P, Sacchi N (1987) Single-step method of RNAisolation by acid guanidium thiocyanate-phenol-chloroformextraction. Anal Biochem 162:156–159
21. Yoon K, Rutledge SJC, Buenaga RF, Rodan GA (1988) Char-acterization of the rat osteocalcin gene: stimulation of pro-moter activity by 1,25-dihydroxyvitamin D3. Biochemistry27:8521–8526
22. Gerstenfeld LC, Chipman SD, Glowacki J, Lian JB (1987)Expression of differentiated function by mineralizing culturesof chicken osteoblasts. Dev Biol 122:49–60
23. Bellows CG, Ishida H, Aubin JE, Heersche JNM (1990) Para-thyroid hormone reversibly suppresses the differentiation ofosteoprogenitor cells into functional osteoblasts. Endocrinol-ogy 127:3111–3116
24. Murray TM, Rao LG, Muzaffar SA (1991) Dexamethasone-treated ROS 17/2.8 rat osteosarcoma cells are responsive tohuman carboxyterminal parathyroid hormone peptide hPTH(53–84): stimulation of alkaline phosphatase. Calcif Tissue Int49:120–123
25. Jongen JWJM, Bos MP, Van der Meer JM, Herrman-ErleeMPM (1993) Parathyroid hormone-induced changes in alka-line phosphatase expression in fetal calvarial osteoblasts: dif-ferences between rat and mouse. J Cell Physiol 155:36–43
26. Pliam NB, Nyiredy KO, Arnaud CD (1982) Parathyroid hor-mone receptors in avian bone cells. Proc Natl Acad Sci USA79:2061–2063
27. Silve CM, Hradek GT, Jones AL, Arnaud CD (1982) Para-thyroid hormone receptor in intact embryonic chicken bone:characterization and cellular localization. J Cell Biol 94:379–386
28. Bellows CG, Aubin JE, Heersche JNM, Antosz ME (1986)Mineralized bone nodules formed in vitro from enzymaticallyreleased rat calvaria cell populations. Calcif Tissue Int 38:143–154
29. Sudo H, Kodama H, Amagai Y, Yamamoto S, Kasai S (1983)In vitro differentiation and calcification in a new clonal os-teogenic cell line derived from newborn mouse calvaria. J CellBiol 96:191–198
30. Owen TA, Aronow M, Shalhoub V, Barone LM, Wilming L,Tassinari MS, Kennedy MB, Pockwinse S, Lian JB, Stein GS(1990) Progressive development of the rat osteoblast pheno-
M. Mizuno et al: BSP for Expression of Osteoblastic Phenotypes of Bone Marrow Cells 395
type in vitro: reciprocal relationships in expression of genesassociated with osteoblast proliferation and differentiationduring formation of the bone extracellular matrix. J Cell Phys-iol 143:420–430
31. Luben RA, Wong GL, Cohn DV (1976) Biochemical charac-terization with parathormone and calcitonin of isolated bonecells: provisional identification of osteoclasts and osteoblasts.Endocrinology 99:526–534
32. Majeska R, Rodan SB, Rodan GA (1980) Parathyroid hor-mone-responsive clonal cell lines from rat sarcoma. Endocri-nology 107:1494–1503
33. Rocha V, Ringo DL, Read DB (1985) Casein production dur-ing differentiation of mammary epitherial cells in collagen gelculture. Exp Cell Res 159:201–210
34. Reznikoff CA, Loretz LJ, Pesciotta DM, Oberley TD, Ignja-tovic MM (1987) Growth kinetics and differentiation in vitroof normal human uroepitherial cells on collagen gel substratesin defined medium. J Cell Physiol 131:285–301
35. Lynch MP, Stein JL, Stein GS, Lian JB (1995) The influenceof type I collagen on the development and maintenance of theosteoblast phenotype in primary and passaged rat calvarialosteoblasts: modification of expression of genes supportingcell growth, adhesion, and extracellular matrix mineralization.Exp Cell Res 216:35–45
36. Maniatopoulos C, Sodek J, Melcher AH (1988) Bone forma-tion in vitro by stromal cells obtained from bone marrow ofyoung adult rats. Cell Tissue Res 254:317–330
37. Malaval L, Modrowski D, Gupta AK, Aubin JE (1994) Cel-lular expression of bone-related proteins during in vitro os-teogenesis in rat bone marrow stromal cells cultures. J CellPhysiol 158:555–572
38. Mizuno M, Shindo M, Kobayashi D, Tsuruga E, Amemiya A,Kuboki Y (1997) Osteogenesis by bone marrow stromal cellsmaintained on type I collagen matrix gels in vivo. Bone 20:101–107
39. Fisher LW, Whitson SW, Avioli LV, Termine JD (1983) Ma-trix sialoprotein of developing bone. J Biol Chem 258:12723–12727
40. Midura RJ, McQeillan DJ, Benham KJ, Fisher LW, HascallVC (1990) A rat osteogenic cell line (UMR 106-01) synthe-sizes a highly sulfated form of bone sialoprotein. J Biol Chem265:5285–5291
41. Nagata T, Goldberg HA, Zhang Q, Domenicucci C, Sodek J(1991) Biosynthesis of bone proteins by fetal porcine cal-variae in vitro. Rapid association of sulfated sialoproteins (se-
creted phosphoprotein-1 and bone sialoprotein) and chondroi-tin sulfate proteoglycan (CS-PG III) with bone mineral. Ma-trix 11:86–100
42. Ibaraki K, Termine JD, Whitson SW, Young MF (1992) Bonematrix mRNA expression in differentiating fetal bovine os-teoblasts. J Bone Miner Res 7:743–754
43. Aubin JE, Bellows CG, Turksen K, Liu F, Heersche NM(1992) Analysis of the osteoblast lineage and regulation ofdifferentiation. In: Slabkin H, Price P (eds) Chemistry andbiology of mineralized tissues. Elsevier Science Publishers,Netherlands, pp 267–276
44. Grzesik WJ, Robey PG (1994) Bone matrix RGD glycopro-teins: immunolocalization and interaction with human primaryosteoblastic bone cells in vitro. J Bone Miner Res 9:487–496
45. Mintz KP, Grzesik WJ, Midura RJ, Robey PG, Termine JD,Fisher LW (1993) Purification and fragmentation of nonde-natured bone sialoprotein: evidence for a cryptic RGD-resistant cell attachment domain. J Bone Miner Res 8:985–995
46. Hultenby K, Reinholt FP, Norgård M, Wendel M, HeinegårdD (1994) Distribution and synthesis of bone sialoprotein inmetaphyseal bone of young rats show a distinctly differentpattern from that of osteopontin. Eur J Cell Biol 63:230–239
47. Yao K, Todescan R Jr, Sodek J (1994) Temporal changes inmatrix protein synthesis and mRNA expression during min-eralized tissue formation by adult rat bone marrow cells inculture. J Bone Miner Res 9:231–240
48. Cooper LF, Yiheikkila PK, Felton DA, Whitson SW (1998)Spatiotemporal assessment of fetal bovine osteoblast culturedifferentiation indicates a role for BSP in promoting differen-tiation. J Bone Miner Res 13:620–632
49. Vukicevic S, Luyten FP, Kleinman HK, Reddi AH (1990)Differentiation of canalicular cell processes in bone cells bybasement membrane matrix components: regulation by dis-crete domain of laminin. Cell 63:437–445
51. Moursi AM, Globus RK, Damsky CH (1997) Interactions be-tween integrin receptors and fibronectin are required for cal-varial osteoblast differentiation in vitro. J Cell Sci 110:2187–2196
52. Aubin JE, Gupta AK, Zirngbl R, Rossant J (1996) Knockoutmouse lacking bone sialoprotein expression have bone abnor-malities. J Bone Miner Res 11:S102
M. Mizuno et al: BSP for Expression of Osteoblastic Phenotypes of Bone Marrow Cells396