The substantia nigra of the human brainII. Patterns of loss of dopamine-containing neurons inParkinson’s disease
P. Damier,1,2 E. C. Hirsch,1 Y. Agid 1 and A. M. Graybiel2
1INSERM U289, Hoˆpital de la Salpeˆtriere, Paris, France Correspondence to: Dr P. Damier, INSERM U289, Hoˆpitaland 2Department of Brain and Cognitive Sciences, de la Salpeˆtriere, 47, boulevard de l’hoˆpital, 75013 Paris,Massachusetts Institute of Technology, Cambridge, FranceMassachusetts, USA E-mail: [email protected]
SummaryTo achieve accuracy in studying the patterns of loss ofmidbrain dopamine-containing neurons in Parkinson’sdisease, we used compartmental patterns of calbindinD28K immunostaining to subdivide the substantia nigrawith landmarks independent of the degenerative process.Within the substantia nigra pars compacta, we identifieddopamine-containing neurons in the calbindin-richregions (‘matrix’) and in five calbindin-poor pockets(‘nigrosomes’) defined by analysis of the three-dimensional networks formed by the calbindin-poorzones. These zones were recognizable in all of thebrains, despite severe loss of dopamine-containingneurons. The degree of loss of dopamine-containingneurons in the substantia nigra pars compacta wasrelated to the duration of the disease, and the cell lossfollowed a strict order. The degree of neuronal losswas significantly higher in the nigrosomes than in the
IntroductionIn Parkinson’s disease, the loss of dopamine-containingneurons in the midbrain is progressive and affects differentparts of the nigral complex to different degrees, the mostsevere loss occurring in the ventrolateral part of the substantianigra pars compacta (Hassler, 1938; Fearnley and Lees,1991). Other distributions of maximal neuronal loss in thenigral complex occur with the normal ageing process and inother degenerative disorders affecting the substantia nigra(Fearnley and Lees, 1991). These differences suggest thatthe regional selectivity of the lesions is specific to the diseaseprocess and is thus related to the mechanisms underlying theneurodegenerative changes. Precise knowledge of the patterns
matrix. Depletion was maximum (98%) in the mainpocket (nigrosome 1), located in the caudal andmediolateral part of the substantia nigra pars compacta.Progressively less cell loss was detectable in moremedial and more rostral nigrosomes, following thestereotyped order of nigrosome 1 > nigrosome 2 >nigrosome 4 > nigrosome 3 > nigrosome 5. A parallel,but lesser, caudorostral gradient of cell loss was observedfor dopamine-containing neurons included in the matrix.This pattern of neuronal loss was consistent from oneparkinsonian substantia nigra pars compacta to another.The spatiotemporal progression of neuronal loss relatedto disease duration can thus be drawn in the substantianigra pars compacta for each Parkinson’s diseasepatient: depletion begins in the main pocket (nigrosome1) and then spreads to other nigrosomes and the matrixalong rostral, medial and dorsal axes of progression.
of depletion of dopamine-containing neurons in Parkinson’sdisease may thus be crucial to understanding its pathogenesis.
We have identified such patterns by applying a newcalbindin D28K-based analysis that allowed us to plot thedistributions of nigral neurons in consistently identified nigralcompartments, the nigrosomes and the surrounding matrix(Damier et al., 1999). This method permitted the firstquantitative study of cell loss in individually distinctsubdivisions of the substantia nigra pars compacta inParkinson’s disease. Our results suggest that cell loss isconsistently greater in the nigrosomes than in the matrix, andthat, within the system of nigrosomes, there is an ordered
1438 P. Damieret al.
Fig. 1 Preservation of the compartmental organization of calbindin D28K immunostaining in the parkinsonian midbrain. Sections at rostral(R), intermediate (I) and caudal (C) levels through the midbrain of case P1060 (B). Sections from corresponding levels taken from acontrol midbrain (A). Symbols indicate different calbindin-poor zones (★, nigrosome 1;r, nigrosome 2;m, nigrosome 3;d, nigrosome4; j, nigrosome 5). CP5 cerebral peduncle; RN5 red nucleus; III5 exiting fibres of the third cranial nerve. Scalebar 5 3 mm.
pattern of loss consistent across Parkinson’s disease patients.This analytical method has great promise as a diagnostic andinvestigative tool.
Material and methodsBrain samplesFive brains from patients who died with a diagnosis ofidiopathic Parkinson’s disease [age6 SEM 5 72 6 4 years]were analysed and compared with brains from five controlpatients (846 4 years) who had no clinical or pathologicalhistory of neuropsychiatric disease. Disease duration wasdefined as the interval between first diagnosis and death. Theclinical diagnosis of Parkinson’s disease was confirmed byhistopathological examination performed on one hemisphereon the basis of severe loss of melanized neurons in thesubstantia nigra pars compacta and the presence of Lewybodies. The five control brains were analysed in the studyreported in the accompanying paper (Damieret al., 1999).
Midbrain blocks were fixed as described previously
(Damieret al., 1999). Transverse 40-µm-thick serial sectionswere cut from the tissue blocks on a sliding microtome,immersed free-floating in 0.25 M Tris buffer containing 0.1%sodium azide, and stored at 4°C.
Calbindin D28K and tyrosine hydroxylaseimmunohistochemistryFor three parkinsonian and three control midbrains, everythird section was processed for calbindin D28K
immunostaining and every ninth for tyrosine hydroxylase(TH) immunostaining. Each of the TH-stained sections wasimmediately adjacent to one of the calbindin-stained sections.In two other parkinsonian and two other control midbrainsadded for the quantitative analysis, every ninth section wasprepared by calbindin immunohistochemistry and every 36thfor TH. The protocol for the calbindin and THimmunohistochemistry was carried out as described byDamier and colleagues (Damieret al., 1999).
Dopaminergic lesions in parkinsonian midbrain 1439
Fig. 2 Regional and intranigral loss of dopamine-containing neurons in Parkinson’s disease. The colorimetric scale indicates theestimated amount of cell loss (least5 blue; most5 red). Cell losses in the different groups of the midbrain were calculated bycomparing mean numbers of TH-positive neurons in each group of five parkinsonian midbrains (disease duration5 7–32 years) withcorresponding means for five control midbrains. A85 dopaminergic cell group A8; CGS5 central grey substance; CP5 cerebralpeduncle; DBC5 decussation of brachium cunjunctivum; M5 medial group; Mv5 medioventral group; N5 nigrosome; RN5 rednucleus; SNpd5 substantia nigra pars dorsalis; SNpl5 substantia nigra pars lateralis; III5 exiting fibres of the third cranial nerve.
1440 P. Damieret al.
Table 1 Quantitative analysis of dopamine-containing neurons and their loss in parkinsonian midbrain
CGS A8 M Mv SN
Number 53036 438 143156 2693 91116 1582 151826 1916 287336 1202Cell loss (%) 9 31 7 46 79
P , 0.001
Values indicate number of cells (mean6 standard error of the mean) with percentage reduction compared with controls indicated below.P indicates statistical significance of cell loss (Tukey’spost hoccomparison). CGS5 central grey substance; A85 dopaminergic cellgroup A8; M 5 medial group; Mv5 medioventral group; SN5 substantia nigra.
Table 2 Rostrocaudal variation in percentage loss of dopamine-containing neurons in parkinsonian midbrain
P indicates statistical significance of the loss of neurons (Tukey’spost hoccomparison). CGS5 central grey substance; A85dopaminergic cell group A8; M5 medial group; Mv5 medioventral group; SN5 substantia nigra.
Mapping and quantitative analysis of dopamine-containing neuronsDopamine-containing neurons in the midbrain were identifiedas TH-positive neurons. Charts of the distributions ofdopamine-containing neurons were constructed from everyTH-stained section by plotting TH-positive neurons with acomputer-assisted image analysis device (HistoRag, Biocom,Les Ulis, France). Five dopaminergic cell groups wereidentified in the midbrain (Hirschet al., 1988), and thesubstantia nigra was subdivided into the substantia nigra parscompacta, formed by the matrix and five nigrosomes, thesubstantia nigra pars dorsalis and the substantia nigra parslateralis, according to the patterns of calbindinimmunostaining observed. This plan of subdivision isdescribed in detail in the accompanying paper (Damieret al., 1999).
The total numbers of dopamine-containing neurons in eachcell group were calculated for each patient by integratingvalues for individual sections over the entire length of themidbrain. The percentage of cell loss in Parkinson’s diseasewas calculated from these values by comparing means forparkinsonian and control midbrains. Split-cell counting errorswere corrected by applying the formula of Abercrombie(Abercrombie, 1946). The correction factor was 0.61; nosignificant differences were found between the sizes ofneuronal nuclei in control and Parkinson’s disease brains.
Statistical analysisStatistical analysis was done by analysis of variance (generallinear model from SAS software, SAS Institute Inc., Cary, NC,USA) based on three factors: (i) disease status (Parkinson’sdisease versus controls); (ii) midbrain subdivisions [centralgrey substance, dopaminergic group A8, medial group,medioventral group and the substantia nigra (Hirschet al.,
1988)] or subdivisions of the substantia nigra [substantianigra pars lateralis, substantia nigra pars dorsalis, and thematrix and nigrosomes 1–5 of the substantia nigra parscompacta (Damieret al., 1999)]; and (iii) rostrocaudallocation (rostral5 anterior to the level of exit of third cranialnerve fibres; intermediate5 at the level of their exit; caudal5posterior to the level of their exit). Interactions among thesefactors were added to the model. Forpost hoccomparisons,we used the Tukey method to correct for multiple test effects.
ResultsCompartmental pattern of calbindinimmunostaining in the parkinsonian midbrainThe conspicuous three-dimensional compartmentalorganization of calbindin immunostaining characteristic ofthe control midbrains was preserved in the parkinsonianmidbrain. Although the calbindin-poor zones were shrunken,all five nigrosomes found in the control midbrains wereidentifiable in the parkinsonian midbrain (Fig. 1). Thispreservation of the nigrosome/matrix organization of thesubstantia nigra pars compacta was the fundamental basisfor the quantitative study undertaken. Because we were ableto identify these nigral compartments and the associatedcalbindin-rich matrix, despite shrinkage and loss of landmarkpatterns based on TH immunostaining, we were able toidentify subgroups of dopamine-containing neurons and toquantify the cell loss characteristic of each.
Global analysis of the loss of dopamine-containing neurons in the parkinsonianmidbrainThere was a mean reduction of 64% (range 57–72%) in thetotal count of dopamine-containing neurons in the five cases
Dopaminergic lesions in parkinsonian midbrain 1441
Fig. 3 Distribution of dopamine-containing neurons in control andparkinsonian midbrains. Filled circles indicate mean counts fromthree control midbrains and filled diamonds indicate the meancounts from three parkinsonian midbrains. Bars indicate SEM.Values along the abscissa indicate distance (mm) to the level ofrostrally exiting fibres of the third cranial nerve. Values on theordinate indicate the number of TH-positive neurons. Areas underthe curves indicate the total numbers of neurons in these regions;indicator box below shows the area corresponding to 5500neurons.
of Parkinson’s disease compared with the total count for thecontrols (P , 0.0001). Across the midbrain as a whole, theloss of TH-positive neurons varied significantly (P , 0.01)along the rostrocaudal axis of the midbrain: there was 24%(range 0–65%) loss in the rostral third, 65% (36–83%) lossin the intermediate third and 72% (66–80%) loss in thecaudal third (Figs 2 and 3).
Cell losses were significantly different (P , 0.00001)among the five different midbrain groups of dopamine-
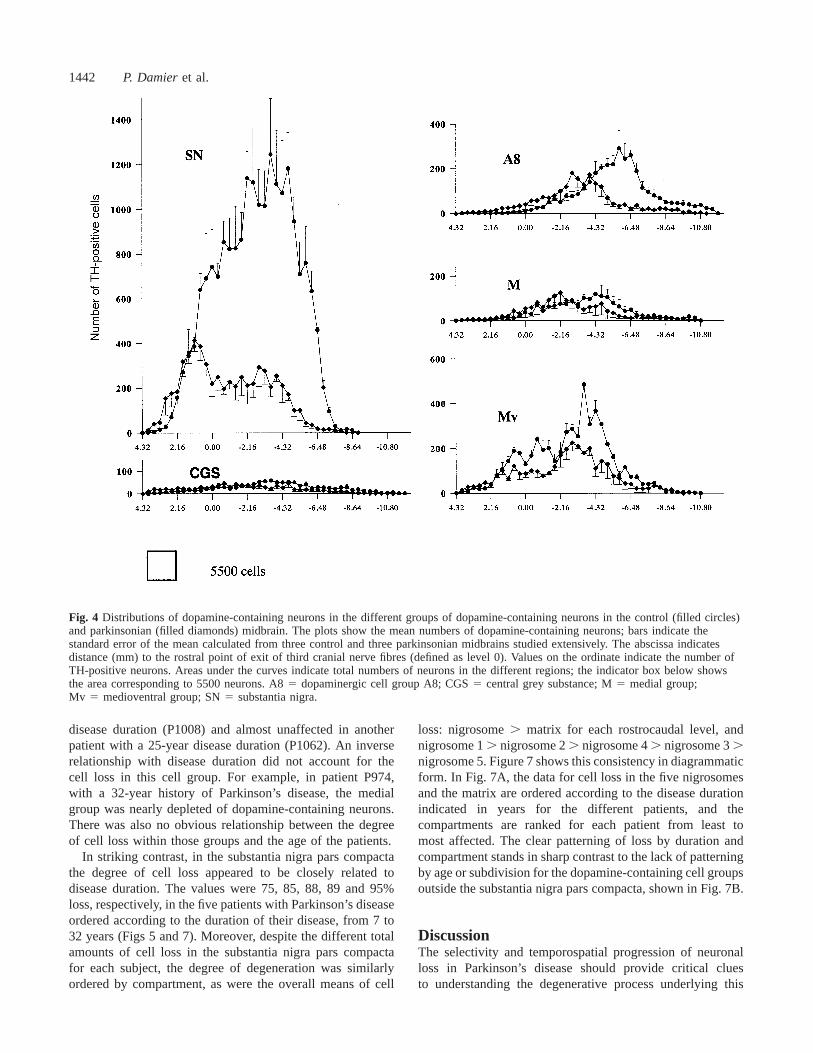
containing neurons that we identified. The average loss wasmaximal in the substantia nigra (79%, range 71–84%),intermediate in the medioventral group (46%, range 26–67%)and in dopaminergic group A8 (31%, range 0–66%), andlower in the central grey substance (9%, range 0–26%) andin the medial group (7%, range 0–41%) (Table 1). Withineach region there was a rostrocaudal gradient of increasingcell loss (Table 2; Figs 2 and 4).
Heterogeneous patterns of cell loss in thesubstantia nigra in Parkinson’s diseaseThere were highly significant differences (P , 0.00001) inthe extent of cell loss in the different subgroups of the nigralcomplex. Loss was higher in the substantia nigra parscompacta (86%, range 75–95%) than in the substantia nigrapars dorsalis (57%, range 44–67%), and there was a smallloss (21%, range 0–72), which did not reach statisticalsignificance, in the substantia nigra par lateralis (Table 3).
The calbindin-based definition of compartments withinthe substantia nigra pars compacta demonstrated furtherheterogeneity in neurodegenerative patterns. First, as shownin Table 3, the cell loss was significantly (P , 0.01) greaterin the nigrosomes (95%, range 87–99%) than in the matrix(80%, range 67–91%). This was true for each rostrocaudallevel for all nigrosomes, excepting only the caudal nigrallevels in patient P974, in which cell loss was nearly total inboth nigrosomes and matrix (Fig. 5). Secondly, there wereclear differences in the loss of dopamine-containing neuronsin the different nigrosomes (Table 3). The mean cell losswas maximal in nigrosome 1 (98%, range 93–100%). Thefew TH-positive neurons that did survive in nigrosome 1 didnot appear to have characteristic locations (e.g. in its dorsalor its ventral part). Nigrosome 2 and nigrosome 4 were thenext most affected, and nigrosome 3 and nigrosome 5 wereconsiderably less affected. The degree of loss of dopamine-containing neurons in the different nigrosomes was strictlyordered: nigrosome 1. nigrosome 2. nigrosome 4.nigrosome 3. nigrosome 5.
In the matrix of the substantia nigra pars compacta, thedecreases in dopamine-containing neurons seemed mainly tofollow a rostrocaudal gradient (Table 4; Figs 2, 5 and 6).Cell loss in the medial part of the matrix (77%, range57–91%) was slightly, but not significantly, lower than thatin the lateral part (86%, range 79–93%).
Individual differences in the loss of midbraindopamine-containing neuronsCell losses in the different dopamine-containing cell groupsof the midbrain were different from one patient to another(Figs 5 and 7). The extent of depletion showed no apparentrelation to disease duration in any of the cell groups exceptthe substantia nigra pars compacta. For example, the medialcell group was severely affected in a patient with a 7-year
1442 P. Damieret al.
Fig. 4 Distributions of dopamine-containing neurons in the different groups of dopamine-containing neurons in the control (filled circles)and parkinsonian (filled diamonds) midbrain. The plots show the mean numbers of dopamine-containing neurons; bars indicate thestandard error of the mean calculated from three control and three parkinsonian midbrains studied extensively. The abscissa indicatesdistance (mm) to the rostral point of exit of third cranial nerve fibres (defined as level 0). Values on the ordinate indicate the number ofTH-positive neurons. Areas under the curves indicate total numbers of neurons in the different regions; the indicator box below showsthe area corresponding to 5500 neurons. A85 dopaminergic cell group A8; CGS5 central grey substance; M5 medial group;Mv 5 medioventral group; SN5 substantia nigra.
disease duration (P1008) and almost unaffected in anotherpatient with a 25-year disease duration (P1062). An inverserelationship with disease duration did not account for thecell loss in this cell group. For example, in patient P974,with a 32-year history of Parkinson’s disease, the medialgroup was nearly depleted of dopamine-containing neurons.There was also no obvious relationship between the degreeof cell loss within those groups and the age of the patients.
In striking contrast, in the substantia nigra pars compactathe degree of cell loss appeared to be closely related todisease duration. The values were 75, 85, 88, 89 and 95%loss, respectively, in the five patients with Parkinson’s diseaseordered according to the duration of their disease, from 7 to32 years (Figs 5 and 7). Moreover, despite the different totalamounts of cell loss in the substantia nigra pars compactafor each subject, the degree of degeneration was similarlyordered by compartment, as were the overall means of cell
loss: nigrosome. matrix for each rostrocaudal level, andnigrosome 1. nigrosome 2. nigrosome 4. nigrosome 3.nigrosome 5. Figure 7 shows this consistency in diagrammaticform. In Fig. 7A, the data for cell loss in the five nigrosomesand the matrix are ordered according to the disease durationindicated in years for the different patients, and thecompartments are ranked for each patient from least tomost affected. The clear patterning of loss by duration andcompartment stands in sharp contrast to the lack of patterningby age or subdivision for the dopamine-containing cell groupsoutside the substantia nigra pars compacta, shown in Fig. 7B.
DiscussionThe selectivity and temporospatial progression of neuronalloss in Parkinson’s disease should provide critical cluesto understanding the degenerative process underlying this
Dopaminergic lesions in parkinsonian midbrain 1443
Table 3 Quantitative analysis of dopamine-containing neurons and their loss in parkinsonian nigral complex
Substantia nigra
SNpl SNpd Snpc
Matrix Nigrosomes
Number 16926 479 129376 1116 118426 2344 22626 917
Values indicate number of cells (mean6 standard error of the mean) with percentage reduction compared with controls indicated below.P values indicate significance of the loss (Tukey’spost hoccomparison). SNpl5 substantia nigra pars lateralis; SNpd5 substantia nigrapars dorsalis; SNpc5 substantia nigra pars compacta; N5 nigrosome.
Table 4 Rostrocaudal variation in neuronal loss in thedifferent dopamine-containing cells of the midbraingroups
Values indicate percentage reduction in cell number relative tocontrol counts.P indicates significance level of loss (Tukey’spost hoccomparison). SNpl5 substantia nigra pars lateralis;SNpd5 substantia nigra pars dorsalis.
disorder. To investigate these factors reliably, it is essentialto have a way to assess the selectivity for differentsubpopulations of dopamine-containing neurons degeneratingduring the course of disease. Landmarks permitting suchanalysis have been defined at a regional level (Hirschet al.,1988), but they have been missing for the most vulnerablepart of the midbrain nigral complex, the large and complexpars compacta of the substantia nigra. In the first paper ofthis series, we demonstrated that the compartmental patternof calbindin D28K immunostaining can be used to delineatesubgroups of dopamine-containing neurons in the substantianigra pars compacta in a reliable way from one brain toanother (Damier et al., 1999). Here we confirm theobservations of Ito and colleagues (Itoet al., 1992) that thecalbindin-positive neuropil survives in the substantia nigrain Parkinson’s disease, and we further show that the distinctivecompartmental patterns of calbindin immunostaining definingthe nigrosomes and the matrix are preserved in the midbrainsof patients with Parkinson’s disease for as long as 30 yearsafter diagnosis. These patterns of calbindin staining, probablycorresponding to the distribution patterns of striatonigralafferent fibres, provided landmarks apparently independent
of the degenerative process in Parkinson’s disease withwhich we could recognize consistently different subgroupsof dopamine-containing neurons in the parkinsonian midbraindespite massive neuronal degeneration. The differential lossof dopamine-containing neurons in Parkinson’s disease couldtherefore be calculated with great accuracy for the differentparts of the substantia nigra pars compacta as well as for themidbrain as a whole.
Dopamine-containing cell groups in themidbrain show different patterns ofneurodegeneration in Parkinson’s diseaseNeuronal loss was uneven across the different dopamine-containing cell groups of the midbrain, both within individualcases of Parkinson’s disease and across cases. Theseobservations confirm the earlier reports (Hirschet al., 1988;Germanet al., 1989). In cell groups outside the substantianigra pars compacta, the degree of neuronal loss did notseem to be related to the duration of disease, and hence maynot reflect disease progression in Parkinson’s disease. Otherfactors related to ageing or to the associated degenerativeprocesses may be important in the aetiology of these lesions.For example, age-related loss of dopamine-containing neuronshas been found to be high in the dorsal part of the substantianigra (Fearnley and Lees, 1991), and loss in the medial andmedioventral midbrain has been shown in Alzheimer’s disease(Mann et al., 1987).
In contrast to this apparently sporadic distribution ofneuronal loss in cell groups outside the substantia nigrapars compacta, loss of dopamine-containing neurons in thesubstantia nigra pars compacta, defined by its calbindin-positive neuropil, appeared to bear an orderly relationship todisease duration. This finding suggests that the loss of
1444 P. Damieret al.
Fig. 5 Individual patterns of depletion of dopamine-containing neurons in the five parkinsonian midbrains analysed. The data are presented in theorder of disease duration (shown at left), ranging from 7 to 32 years. The colorimetric scale indicates the degree of cell loss (blue5 least; red5most). DBC5 decussation of brachium conjunctivum; CP5 cerebral peduncle; RN5 red nucleus; III5 exiting fibres of third cranial nerve.
Dopaminergic lesions in parkinsonian midbrain 1445
Fig. 6 Distributions of dopamine-containing neurons in the different subdivisions of the substantia nigra defined on the basis of calbindinimmunostaining in the control (filled circles) and parkinsonian (filled diamonds) midbrain. Symbols indicate the mean numbers ofneurons; bars indicate the standard error of the mean calculated from the three control and three parkinsonian midbrains studied innear-serial sections. Values on the abscissa indicate distance (mm) to the level of the most rostrally exiting fibres of the third cranialnerve (defined as level 0). Values on the ordinate indicate the number of TH-positive neurons. Areas under the curves indicate totalnumbers of neurons in the different regions; the indicator box below shows an area corresponding to 5500 neurons. N5 nigrosome;SNpd5 substantia nigra pars dorsalis; SNpl5 substantia nigra pars lateralis.
dopamine-containing neurons in the substantia nigra parscompacta is more directly reflective of the degenerativeprocess in Parkinson’s disease than is degeneration in otherdopaminergic cell groups, and might even be solely aconsequence of the process underlying Parkinson’s disease.In the two cases with the shortest survival after diagnosis(7 years), degeneration within the substantia nigra parscompacta was already severe (75 and 82% cell loss). Thus,our sample missed three-quarters of the disease progression,as measured by estimated amounts of cell death. Thedifferences in amount of cell loss from the cases with adisease duration of 7 years to the next longer duration(21 years) also were small (75 and 82% versus 85% cellloss). Even so, there was a clear rank ordering in the amountof total cell loss across the cases, with a total range of15%. These findings, similar to those of Fearnley and Lees(Fearnley and Lees, 1991), give one estimate of the overall
lesion progression in Parkinson’s disease in terms of the celldeath endpoint. Further, they underline the great potential ofdeveloping neuroprotective measures in Parkinson’s disease.
Differential vulnerability of the nigralcompartments, nigrosomes and matrix inParkinson’s diseaseA major finding of this study is that, in every parkinsonianmidbrain, dopamine-containing neurons in nigrosomes weremore affected than dopamine-containing neurons of thematrix. Based on the subdivisions defined by calbindinpatterns in neurologically normal midbrains (Damieret al.,1999), dopamine-containing neurons of the substantia nigrapars compacta can be divided into two types: sparselydistributed neurons included in a calbindin-rich matrix
1446 P. Damieret al.
Fig. 7 Temporospatial pattern of the neuronal lesion inParkinson’s disease analysed for the substantia nigra parscompacta (A) and for the other midbrain dopaminergic groups(B). Individual cases of Parkinson’s disease are ordered from topto bottom in relation to increasing disease duration (7–32 years).In A, the subdivisions of the substantia nigra pars compacta areordered in terms of increasing cell loss based on mean values forall patients (matrix, nigrosome for each rostrocaudal level andnigrosome 5,nigrosome 3, nigrosome 4, nigrosome 2,nigrosome 1). InB, the remaining cell groups are ordered usingthe same rule (M, CGS, SNpl , A8 , Mv , SNpd). Notethat no particular pattern of vulnerability emerges inB but that inA there is a clear indication of temporospatial progression of theParkinson’s disease lesion from nigrosome 1 to nigrosome 2,nigrosome 4, nigrosome 3, nigrosome 5 and the matrix in all fivepatients. A85 dopaminergic cell group A8; CGS5 central greysubstance; M5 medial group; Ma5 matrix; N 5 nigrosome;Mv 5 medioventral group; SNpd5 substantia nigra pars dorsalis;SNpl 5 substantia nigra pars lateralis.
compartment, and densely packed neurons in five calbindin-poor nigrosomes. Our quantitative estimates in controlmidbrains suggest that the population of the matrix neuronsis normally ~1.4 times larger than the population of neuronsin the nigrosomes [mean5 59 000 in the matrix versusnearly 44 000 in nigrosomes (Damieret al., 1999)]. Theconsistent difference in cell loss in matrix and nigrosomesin all five parkinsonian patients, found at all levels in thesubstantia nigra pars compacta containing surviving neurons,suggests that differential nigrosome/matrix vulnerability maybe a basic attribute of the disease process in Parkinson’sdisease.
Our data suggest that this differential loss of neurons innigrosomes may not solely reflect the high packing densityof nigrosomal neurons. For example, the loss of dopamine-containing neurons was moderate (56%) in the medioventralgroup, in which neurons are densely packed at the level ofexiting third cranial nerve fibres, in comparison with the84% loss of neurons at this level in the matrix, in whichneurons are sparsely distributed. Further, cell loss in differentnigrosomes ranged from 76 to 98% despite dense cell packingin each of them. The greater loss of dopamine-containingneurons in the nigrosomes than in the matrix also did notappear to be related solely to the preferential topographicallocations of these neurons in the midbrain. Whatever therostrocaudal level analysed, the degree of cell loss in thenigrosomes was higher than that in the immediatelysurrounding matrix. Similarly, a preferential ventral locationof many of the dopamine-containing neurons in thenigrosomes did not appear to account for the differentialvulnerability. Within a given nigrosome, the loss ofTH-positive neurons did not show a dorsoventral gradient,even in the large nigrosome 1, and neurons in more dorsalnigrosomes (e.g. nigrosome 4) were more affected thanneurons in the matrix ventral to them at the same rostrocaudallevel. The preferential lateral location of many dopamine-containing neurons belonging to nigrosomes was also nota sufficient correlate of the differential vulnerability ofnigrosomal neurons. Dopamine-containing neurons includedin the medially situated nigrosome 2, for example, were moreaffected (94%) than those of the medial matrix (77%) at thesame rostrocaudal level.
Among the different nigrosomes, there were also consistentdifferences in the amount of cell loss, the greatest loss alwaysbeing in nigrosome 1. These differential patterns of cell losssuggest that it is not only the environment common to allnigrosomes (e.g. low levels of calbindin-positive neuropil)that sets the threshold of vulnerability. The specialvulnerability of nigrosome 1 is in good accord with thepreferential loss found in the ventrolateral clusters by Hassler(Hassler, 1938) and by Fearnley and Lees (Fearnley andLees, 1991). This strong bias indicates that, in addition to agenerally greater vulnerability of neurons in nigrosomesthan in adjacent parts of the matrix, there is heightenedvulnerability of ventrocaudal nigral regions that are ofunknown origin but reflecting a well-known generaltopography of the disease. What our analysis adds to thistopographical analysis is the notion that factors contributingto neuronal loss interact in the substantia nigra pars compactawith dopamine-containing neurons of two types—those in thematrix and those in nigrosomes—with appreciably differentvulnerabilities in Parkinson’s disease.
Evidence for stereotyped temporospatialprogression of cell loss in Parkinson’s diseaseOne of the most interesting results of our nigrosome/matrixanalysis is the indication of orderly lesion progression in
Dopaminergic lesions in parkinsonian midbrain 1447
Parkinson’s disease. The lesions appeared to follow astereotyped temporospatial progression, beginning innigrosome 1, extending to other nigrosomes in the ordernigrosome 1, nigrosome 2, nigrosome 4, nigrosome 3,nigrosome 5, and then affecting the matrix, with a globalcaudal-to-rostral, lateral-to-medial and ventral-to-dorsaldirection of progression. Our case material was limited toonly five patients, but if this pattern appears in a largerpopulation of parkinsonian cases it may prove a signature ofthe neuropathology in Parkinson’s disease.
The general gradient loss observed in the substantia nigrapars compacta is probably related to the gradient depletionof dopaminergic terminals observed post-mortem in thestriatum of Parkinson’s disease patients (Kishet al., 1988)and in [18F]fluorodopa (Brookset al., 1990) or 11C-Win35 428 (Frostet al., 1993) PET scans, with a higher loss indorsal and caudal parts of the putamen than in the caudatenucleus. In relation to the somatotopic projection of corticalafferents to the putamen, one might expect an initialsymptomatology in the foot in Parkinson’s disease (Vidailhetet al., 1994), but other authors have found symptomsexpressed first in the arm (Schelosky and Poewe, 1990).We have no information about whether there are differentnigrostriatal projections from matrix neurons that might adddetail to this general mapping and that might therefore be ofgenuine significance in terms of clinical interpretation.
The particular temporospatial lesion progression that weobserved probably reflects the pathogenesis of Parkinson’sdisease. Because the nigrosome/matrix analysis refers tocompartmental subdivisions within the substantia nigra parscompacta, the most obvious conclusion would be thatcompartmental locality in the substantia nigra pars compactaitself is a key to differential vulnerability. If these differentlocalities have, as suggested above, different striatalprojection zones, then it is possible that the initial trigger forcell loss is at the level of the striatum, and that the effectmoves retrogradely back to the different nigral compartments.If the basis for the pathology is within the substantia nigrapars compacta, our results suggest three main types ofhypothesis: (i) that dopamine-containing neurons are exposedin Parkinson’s disease to variable amount of exogenous orendogenous toxins whose amounts increase from nigrosome1 to other nigrosomes and then to the matrix; (ii) that thedopamine-containing neurons are exposed to similar amountsof toxin, regardless of compartment, but that mechanisms ofdefence against such toxins vary in an orderly way from onecompartment of dopamine-containing neurons to the next;and (iii) that neurons in the different compartments havedifferent patterns of expression of genes implicated in thedisease process. Our results point to the possibility that thenigrosomes and matrix compartments of the substantia nigradiffer in term of their content of growth factors and receptors,compounds related to excitotoxicity, agents involved inoxidative metabolism, and potentially predisposing genessuch as those forα-synuclein (Polymeropouloset al., 1997)and parkin (Kitadaet al., 1998).
AcknowledgementsThis study was supported by NIH Javits Award NS25529,the National Parkinson Foundation, the Fondation pour laRecherche Me´dicale and the French Foreign Office(programme Lavoisier).
ReferencesAbercrombie M. Estimation of nuclear population from microtomesections. Anat Rec 1946; 94: 239–47.
Brooks DJ, Ibanez V, Sawle GV, Quinn N, Lees AJ, Mathias CJ,et al. Differing patterns of striatal 18F-dopa uptake in Parkinson’sdisease, multiple system atrophy, and progressive supranuclear palsy[see comments]. Ann Neurol 1990; 28: 547–55. Comment in: AnnNeurol 1991; 29: 689–90.
Damier P, Hirsch EC, Agid Y, Graybiel AM. The substantianigra of the human brain. I. Nigrosomes and nigral matrix, acompartmental organization based on calbindin D28Kimmunohistochemistry. Brain 1999; 122: 1421–36.
Fearnley JM, Lees AJ. Ageing and Parkinson’s disease: substantianigra regional selectivity. Brain 1991; 114: 2283–301.
Frost JJ, Rosier AJ, Reich SG, Smith JS, Ehlers MD, Snyder SH,et al. Positron emission tomographic imaging of the dopaminetransporter with11C-Win 35,428 reveals marked declines in mildParkinson’s disease [see comments]. Ann Neurol 1993; 34: 423–31. Comment in: Ann Neurol 1994; 35: 376–9.
German DC, Manaye K, Smith WK, Woodward DJ, Saper CB.Midbrain dopaminergic cell loss in Parkinson’s disease: computervisualization [see comments]. Ann Neurol 1989; 26: 507–14.Comment in: Ann Neurol 1990; 28: 111–2.
Hassler R. Zur Pathologie der paralysis agitans und despostenzephalitischen Parkinsonismus. J Psychol Neurol 1938; 48:387–476.
Hirsch EC, Graybiel AM, Agid YA. Melanized dopaminergicneurons are differentially susceptible to degeneration in Parkinson’sdisease. Nature 1988; 334: 345–8.
Ito H, Goto S, Sakamoto S, Hirano A. Calbindin-D28 in the basalganglia of patients with Parkinsonism. Ann Neurol 1992; 32: 543–50.
Kish SJ, Shannak K, Hornykiewicz O. Uneven pattern of dopamineloss in the striatum of patients with idiopathic Parkinson’s disease.Pathophysiologic and clinical implications. N Engl J Med 1988;318: 876–80.
Kitada T, Asakawa S, Hattori N, Matsumine H, Yamamura Y,Minoshima S, et al. Mutations in the parkin gene cause autosomalrecessive juvenile parkinsonism [see comments]. Nature 1998; 392:605–8. Comment in: Nature 1998; 392: 544–5.
Mann DM, Yates PO, Marcyniuk B. Dopaminergic neurotransmittersystems in Alzheimer’s disease and in Down’s syndrome at middleage. J Neurol Neurosurg Psychiatry 1987; 50: 341–4.
Polymeropoulos MH, Lavedan C, Leroy E, Ide SE, Dehejia A,Dutra A, et al. Mutation in the alpha-synuclein gene identified infamilies with Parkinson’s disease [see comments]. Science 1997;276: 2045–7. Comment in: Science 1997; 276; 1973, Comment in: