Calcium, Magnesium, and Phosphorus Balance in Human and Experimental Hypertension DAVID A. MCCARRON, M.D. SUMMARY The thesis that primary disturbances of divalent ion metabolism contribute to the development and maintenance of hypertension is addressed. Representative interactions of calcium, magnesium, and phosphorus with normal cardiovascular physiology are presented. Established and postulated abnormalities of divalent ion metabolism associated with human and experimental hyper- tension are reviewed. The influence of calcium balance on blood pressure development in the young spontaneously hypertensive rat is demonstrated by the results of a diet intervention study. Twelve male SHRs were randomized at 4 weeks of age to one of three diets that differed only in the calcium content (0.25%, 0.5%, and 4.0% by weight). The SHRs' blood pressures stratified inversely (p < 0.001) based upon the calcium content. The low calcium animals experienced a more rapid and greater rise in blood pressure between 4 and 20 weeks of age (p < 0.01). Blood pressures of the supplemented SHRs (4%) peaked at a lower value (174 vs 192 mm Hg, p < 0.01). After maturity, the 4% SHRs experienced an attenuation (p < 0.01) of their hypertension (154 ± 7 mm Hg, 4% SHR vs 176 ± 7 mm Hg, 0.5% SHR). It is proposed that membrane-associated bioavailable Ca 2+ is reduced in the SHR, and possibly in human, hypertension. Dietary calcium supplementation may reverse this defect, resulting in cell membrane stabilization and vascular smooth muscle relaxation. (Hypertension 4 (supp III): III-27-III-33, 1982) KEY WORDS • calcium and hypertension • magnesium and hypertension • phosphorus and hypertension • Ca 2+ and the SHR • spontaneously hypertensive rat urinary electrolytes and dietary Ca 2+ D IVALENT ions serve critical functions in car- I diovascular tissue. This report focuses on three of those ionic species: calcium, magne- sium, and phosphorus, and assesses the contributions of changes in their metabolic balance on the pathophysiology of hypertension. While each exerts direct actions on cardiovascular physiology, each also modifies one another's. In addition, the cardiovascular effects of sodium, potassium, and many vasoactive hormones may be influenced by the divalent ions. First, representative actions of these three ions on nor- mal cardiovascular physiology will be reviewed and established, and then postulated abnormalities of diva- lent ion metabolism associated with hypertension will be assessed. From the Division of Nephrology and Hypertension, Oregon Health Sciences University, Portland, Oregon. Supported, in part, by a grant-in-aid from the Oregon Heart Association, a grant-in-aid from the National Dairy Council, and Grant RR00334 from the General Clinical Research Branch of the Division of Research Resources, U.S. Public Health Service. Address for reprints: Dr. David A. McCarron, Division of Neph- rology, University of Oregon Health Science Center, 3181 South- west Sam Jackson Park Road, Portland, Oregon 97201. Calcium, Magnesium, and Potassium in Cardiovascular Physiology Calcium (Ca 2+ ) is essential to neurohurmoral con- trol,' volume regulation, 2 and vascular smooth muscle function. 3 Calcium's diverse effects on blood pressure control reflect the cation's central role in both mem- brane and cytosolic associated events. In conjunction with membrane receptors and intracellular calmodulin, Ca 2+ regulates cell-cell communication, neurotrans- mitter synthesis and release, and hormone receptor interactions that initiate cytosolic metabolism. 4 - 5 In the central nervous system, Ca 2+ is essential to the integra- tion of the various components of blood pressure con- trol. In the kidney, Ca 2+ modifies water and solute excretion through its effects on filtration rate and reab- sorptive processes. 6 Calcium is a critical element in normal vascular tissue physiology. The cation's contribution to blood pressure regulation is most prominent in its influence on vascular resistance. Membrane receptor binding is dependent on Ca 2+ . Ion fluxes, including those of so- dium and potassium, are altered by their interaction with Ca 2+ . 7 Calmodulin and Ca 2+ induce enzymatic pathways that range from phospholipase A to myosin 111-27 by guest on May 5, 2018 http://hyper.ahajournals.org/ Downloaded from
Transcript
Calcium, Magnesium, and Phosphorus Balance inHuman and Experimental Hypertension
DAVID A. MCCARRON, M.D.
SUMMARY The thesis that primary disturbances of divalent ion metabolism contribute to thedevelopment and maintenance of hypertension is addressed. Representative interactions of calcium,magnesium, and phosphorus with normal cardiovascular physiology are presented. Established andpostulated abnormalities of divalent ion metabolism associated with human and experimental hyper-tension are reviewed. The influence of calcium balance on blood pressure development in the youngspontaneously hypertensive rat is demonstrated by the results of a diet intervention study. Twelvemale SHRs were randomized at 4 weeks of age to one of three diets that differed only in the calciumcontent (0.25%, 0.5%, and 4.0% by weight). The SHRs' blood pressures stratified inversely (p <0.001) based upon the calcium content. The low calcium animals experienced a more rapid and greaterrise in blood pressure between 4 and 20 weeks of age (p < 0.01). Blood pressures of the supplementedSHRs (4%) peaked at a lower value (174 vs 192 mm Hg, p < 0.01). After maturity, the 4% SHRsexperienced an attenuation (p < 0.01) of their hypertension (154 ± 7 mm Hg, 4% SHR vs 176 ± 7mm Hg, 0.5% SHR). It is proposed that membrane-associated bioavailable Ca2+ is reduced in theSHR, and possibly in human, hypertension. Dietary calcium supplementation may reverse this defect,resulting in cell membrane stabilization and vascular smooth muscle relaxation.(Hypertension 4 (supp III): III-27-III-33, 1982)
KEY WORDS • calcium and hypertension • magnesium and hypertension •phosphorus and hypertension • Ca2+ and the SHR • spontaneously hypertensive raturinary electrolytes and dietary Ca2+
DIVALENT ions serve critical functions in car-I diovascular tissue. This report focuses on
three of those ionic species: calcium, magne-sium, and phosphorus, and assesses the contributionsof changes in their metabolic balance on thepathophysiology of hypertension. While each exertsdirect actions on cardiovascular physiology, each alsomodifies one another's. In addition, the cardiovasculareffects of sodium, potassium, and many vasoactivehormones may be influenced by the divalent ions.First, representative actions of these three ions on nor-mal cardiovascular physiology will be reviewed andestablished, and then postulated abnormalities of diva-lent ion metabolism associated with hypertension willbe assessed.
From the Division of Nephrology and Hypertension, OregonHealth Sciences University, Portland, Oregon.
Supported, in part, by a grant-in-aid from the Oregon HeartAssociation, a grant-in-aid from the National Dairy Council, andGrant RR00334 from the General Clinical Research Branch of theDivision of Research Resources, U.S. Public Health Service.
Address for reprints: Dr. David A. McCarron, Division of Neph-rology, University of Oregon Health Science Center, 3181 South-west Sam Jackson Park Road, Portland, Oregon 97201.
Calcium, Magnesium, and Potassiumin Cardiovascular Physiology
Calcium (Ca2+) is essential to neurohurmoral con-trol,' volume regulation,2 and vascular smooth musclefunction.3 Calcium's diverse effects on blood pressurecontrol reflect the cation's central role in both mem-brane and cytosolic associated events. In conjunctionwith membrane receptors and intracellular calmodulin,Ca2+ regulates cell-cell communication, neurotrans-mitter synthesis and release, and hormone receptorinteractions that initiate cytosolic metabolism.4-5 In thecentral nervous system, Ca2+ is essential to the integra-tion of the various components of blood pressure con-trol. In the kidney, Ca2+ modifies water and soluteexcretion through its effects on filtration rate and reab-sorptive processes.6
Calcium is a critical element in normal vasculartissue physiology. The cation's contribution to bloodpressure regulation is most prominent in its influenceon vascular resistance. Membrane receptor binding isdependent on Ca2+. Ion fluxes, including those of so-dium and potassium, are altered by their interactionwith Ca2+.7 Calmodulin and Ca2+ induce enzymaticpathways that range from phospholipase A to myosin
111-28 PERSPECTIVES IN HYPERTENSION SUPPL III, HYPERTENSION, VOL 4, No 5, SEPTEMBER-OCTOBER 1982
light chain kinase which catalyzes the final steps inthe activation of the contractile proteins, actin andmyosin.4 Ultimately, the intracellular function of thisprotein-cation complex is self-regulating,8 facili-tating vascular tissue relaxation under appropriatecircumstances.9
Like Ca2+, magnesium (Mg2+) is an essential ele-ment in normal cardivascular physiology. The synthe-sis and secretion of neurotransmitters is influenced bythe cation.l0 Via its direct effects on cardiac conductionand contractility as well as its influencing water andsolute excretion, Mg2+ can alter both cardiac outputand intravascular volume." Magnesium is required fornormal vascular smooth muscle physiology.10 Mem-brane-associated actions of Mg2+ include stabilizationof Ca2+ channels, activation of Ca2+ /Mg2+ ATPase,and modulation of Na+ /K+ ATPase activity.12 In thecytosol, the cation functions as a cofactor in the enzy-matic generation of cAMP, and catalyzes the actin-myosin-Ca2+ interaction, which ultimately determinesbasal and stimulated vascular tone.13 Within smoothmuscle cell vesicles, Mg2+ is important for ATP gen-eration in mitochondria and sequestration of both Ca2+
and K+.14 In essence, Mg2+ acts as a regulatory cofac-tor in the cascade of events that is initiated by cellstimulation and proceeds through ion fluxes, enzymat-ic induction, energy-dependent metabolic responses,and return of the cell to its basal state. Magnesium'scardiovascular effects parallel those of Ca2+.
The role of phosphorus (PO4=) in normal cardiovas-
cular physiology is equally as diverse and important asthatofCa2+ andMg2+. In all cells and organs, PO4
= isa prerequisite for normal plasma membrane synthesisand integrity.15 In addition, most energy requiringmetabolic function of a cell is dependent upon PO4
=
through the formation and degradation of high energybonds in ATP. As a consequence, membrane-associat-ed ion pumps, as well as receptor-ion channel interac-tions, involve PO4
=.16 Furthermore, the synthesis,storage, and release of local and systemic hormonesthat regulate cardiac output and vascular resistancerequire phosphorus. In vascular tissue, PO4
= is a vitalcofactor in the processes outlined above for Ca2+ andMg2+. Functionally, the primary roles of these threeionic species in vascular cell physiology is highlyintegrated.
Disorders of Calcium, Magnesium, and PhosphorusMetabolism in Human and Experimental
HypertensionAbnormalities of Ca2+ homeostasis have been identi-
fied in both human and experimental hypertension.17"19
The human data include clinical and epidemiologicalobservations. Low serum ionized calciums,20 elevatedparathyroid hormone levels,17 and an increase in renalcalcium excretion 19 21- 22 have been reported in hyper-tensive humans. Consistent with clinical associationsbetween abnormalities of calcium and PTH homeosta-sis and human hypertension is the observation thatovert hypercalcemia will develop in a subset of hyper-
tensives taking thiazide diuretics.23 This thiazide-relat-ed hypercalcemia is facilitated by PTH.24
The established relationship between hypertensionand hyperparathyroidism25 appears to reflect the requi-site stimulation of the parathyroid gland axis as a com-pensatory mechanism to protect calcium balance ofhypertensives. This interpretation of recent data is tobe contrasted with earlier reports26-27 that have con-cluded that hyperparathyroidism and its attendant hy-percalcemia were the cause of hypertension, and,therefore, the primary abnormality linking hypercalce-mia with the development of hypertension. Independ-ent of the issue of essential hypertension, it is apparentthat some hypercalcemia states may be associated withthe emergence of high blood pressure via effects onrenal28 and sympathetic nervous system function.29
Comparable disturbances of Ca2+ metabolism haverecently been characterized in the Aoki-Okamoto,spontaneously hypertensive rat (SHR).18-19-3o:" TheSHR's disorders have been extended to include abnor-mal intestinal transport of calcium32 as well as alteredmembrane binding of 45Ca2+ in RBC,33 cardiomyo-cytes,34 and adipocytes.35 The functional importance ofthe SHR's abnormalities in Ca2+ metabolism has beensuggested by the animal's blood pressure response todietary calcium supplementation. Placing the SHR ona supplemented Ca2+ diet will lower its blood pres-sure.19- 36 even though the diet is introduced at 10 to 12weeks of age after the hypertension is established. Thenormotensive, Wistar-Kyoto rat's blood pressure islikewise responsive to changes in dietary Ca2+, as ittracks inversely based upon the Ca2+ content in thediet.37-38
The first suggestion that the dietary intake of Ca2+
may be an important factor in the development of hy-pertension came from the epidemiologic observationsthat cardiovascular mortality and regional blood pres-sures were inversely related to water hardness.39-40
More recently, a review of the incidence of gestationalhypertension worldwide,41 as well as two preliminaryreports from the United States, have suggested thathypertensive individuals' Ca2+ intakes are significant-ly less than those of normotensives.42-43
The accumulated experience with calcium balanceand hypertension suggests that a modest reduction inCa2+ intake may exacerbate the abnormalities of Ca2+
metabolism associated with hypertension. The relativeCa2+ depletion that ensues may alter vascular smoothmuscle function and enhance vascular tone. Mainte-nance of an adequate exposure to Ca2+, conversely,may have protective effects on blood pressureregulation.
There is additional circumstantial evidence that rela-tive calcium depletion may increase blood pressure inhumans and animals. On a teleological basis, onewould reason that, were relative Ca2+ depletion toresult in an increase in vascular resistance, then thehormone responsible for improving Ca2+ balance, ifendowed with any vascular effects, would cause vaso-dilation. Specific vasodilating properties of parathy-roid hormone at both physiologic and pharmacologic
CALCIUM, MAGNESIUM, PHOSPHORUS, AND BP/McCarron 111-29
concentrations have been demonstrated in experimen-tal models4445 and humans.46
Magnesium homeostasis has not been characterizedin either human or experimental hypertension. This ispartly due to Mg2+ being primarily an intracellularcation and, therefore, difficult to assess. Though dif-fering in their total body content and compartmentaldistributions, Mg2+ and Ca2+ share many common de-terminants of metabolic balance. The abnormalities ofCa2+ homeostasis described above may have theircounterparts in Mg2+ metabolism and hypertension.
Magnesium restriction in experimental animals ac-celerates cardiovascular disease.47 In vitro depletion ofMg2+ enhances vascular reactivity to vasoconstric-tors.48 Magnesium excess stabilizes vascular mem-branes and reduces vascular tone.49 Epidemiologicalevidence also suggests that increasing Mg2+ exposurein the diet may afford humans protection against car-diovascular events.50
Serum PO4= is inversely correlated with blood pres-
sure in normotensive individuals.51 Compared to age-,sex-, and racially-matched controls, serum PO4
= lev-els of hypertensives are lower.20 In both humans andexperimental animals,52 severe phosphorus depletionis associated with hypotension. The levels of hypo-phosphatemia required, though, are extreme, produc-ing multiple organ failure.
As noted above, supplementation of the adolescentSHR's diet with Ca2+, beginning at 10 to 12 weeks ofage, lowers blood pressure after the animal reachesmaturity. The effects of earlier introduction of dietaryCa2+ supplementation on the development and mainte-nance of the SHR's hypertension is unknown. In addi-tion, the influence of dietary Ca2+ intake on the dailyurinary excretion of other electrolytes is also incom-pletely defined in experimental hypertension. The fol-lowing protocol sought to address these questions.
MethodsTwelve male Aoki-Okamoto SHRs were random-
ized to one of three diet regimens at 4 weeks of age.The synthetic diets differed only in their Ca2+ carbon-ate content; 0.25%, 0.5%, and 4% Ca2+ by weight ofthe food. The 0.25% represents a low-normal Ca2+
diet; 0.5%, a normal Ca2+ diet; and 4.0%, a supple-mented Ca2+ diet. Beginning at 7 weeks of age, and at4- to 8-week intervals thereafter up to 39 weeks of age,systolic blood pressures, weights, serum ionized cal-cium, and 24-hour urinary electrolyte and creatinineexcretion were measured. Serum creatinine was meas-ured at only 33 weeks of age.
Urine collections were carried out 1 to 2 days beforeblood pressure determinations. Tail-cuff systolic bloodpressures were recorded on a Narcobiosystem physio-graph. Four readings were averaged for each rat.Blood samples (2 ml) were withdrawn the followingday via subclavian venapuncture. Serum samples wereobtained between 12 and 33 weeks of age. Serum andurine chemistries were determined by previously de-
scribed techniques.19 Analysis of variance (repeatedmeasures) and t statistics were used in the dataassessment.
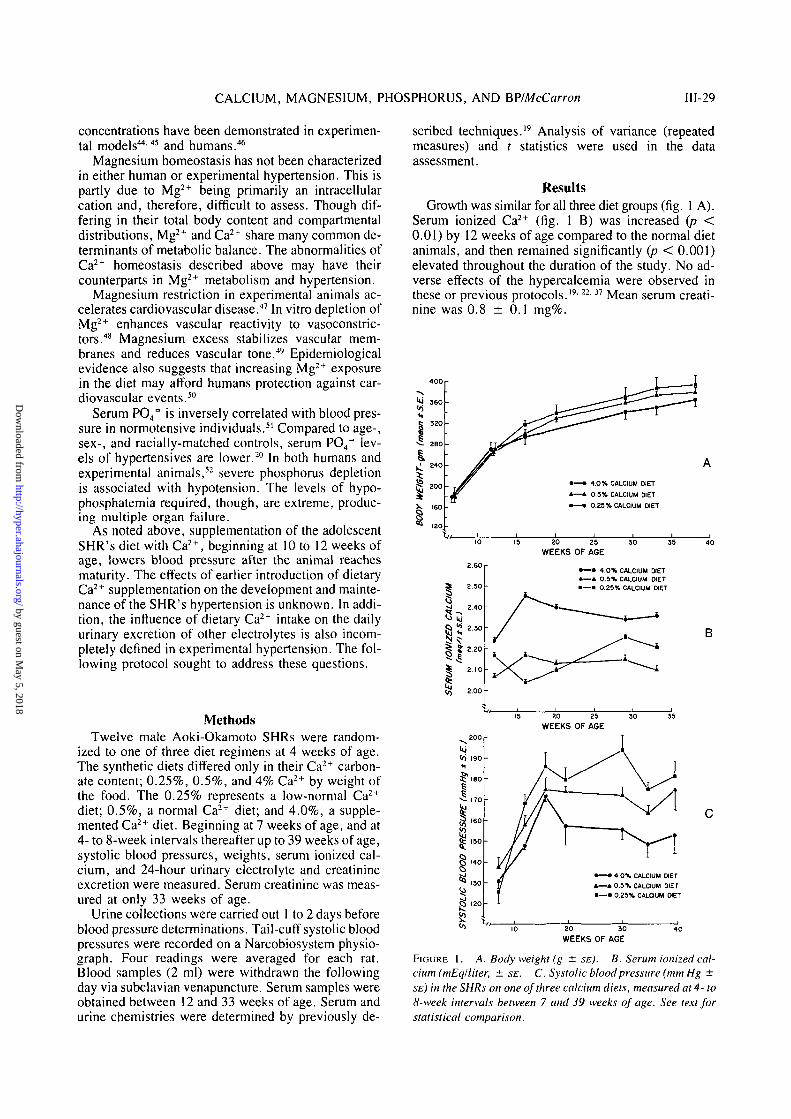
ResultsGrowth was similar for all three diet groups (fig. 1 A).
Serum ionized Ca2+ (fig. 1 B) was increased (p <0.01) by 12 weeks of age compared to the normal dietanimals, and then remained significantly (p < 0.001)elevated throughout the duration of the study. No ad-verse effects of the hypercalcemia were observed inthese or previous protocols.'9-22'37 Mean serum creati-nine was 0.8 ± 0.1 mg%.
FIGURE 1. A. Body weight (g ± SE). B. Serum ionized cal-cium (mEq/liter, ± SE. C. Systolic blood pressure (mm Hg ±SE) in the SHRs on one of three calcium diets, measured at 4- to8-week intervals between 7 and 39 weeks of age. See text forstatistical comparison.
111-30 PERSPECTIVES IN HYPERTENSION SUPPL III, HYPERTENSION, VOL 4, No 5, SEPTEMBER-OCTOBER 1982
Between 7 and 17 weeks of age, the rise in systolicblood pressure was similar on all three diets. From the17th week onward, the systolic pressure (fig. 1 C)tracked inversely (p < 0.001) with the Ca2+ content ofthe diet. The 4% animals' hypertension was the low-est, while that of 0.25% SHRs was the highest. At 39weeks of age, the 4% SHRs' systolic pressure was 154± 7 mm Hg, the 0.5% SHRs' was 176 ± 7 mm Hg,and the 0.25% SHRs' was 181 ± 5 mm Hg.
Urinary electrolyte excretions per 24 hours are re-corded in table 1. Urinary Ca2+ excretion increased (p< 0.001) in parallel with the Ca2+ content of the diet.For all three diet groups, UCaV increased over time (p< 0.001). Conversely, U ^ V stratified (p < 0.001)inversely based upon the percent dietary Ca2+. UMgVwas unchanged by the Ca2+ content of the diet. Potas-sium excretion was increased on the 4% Ca2+ diet (p< 0.01) when compared to both the 0.25% and 0.5%diets. Potassium excretion did not vary with the age ofthe animal. Urinary sodium excretion stratified ac-cording to the percentage of Ca2+ in the diet (p <0.02). By multiple range testing, UNaV differed be-tween all three diets, such that UNaV was less on the0.25% Ca2+ diet than on either the 0.5% (p < 0.05) or4% (p < 0.025) diets. As is apparent from table 1, thedifferences in UNaV on the three Ca2+ diets was evidentonly in the younger animals. Importantly, UNaV did notvary significantly over time on any of the three diets,as daily excretion was similar at all observation pointson the respective diets.
DiscussionModifying Ca2+ intake in experimental animals will
alter their blood pressures. Previous reports in normo-tensive3738 and hypertension rats,19-36 have demon-strated that supplementation of the diet with Ca2+ low-ers the blood pressure of adult SHRs and Wistar-Kyotorats. The protective effect occurs in conjunction withthe development of mild hypercalcemia in the rat. Thecurrent investigation extends these previous observa-tions. Introduction of Ca2+ supplementation at the timeof weaning reduces the peak systolic pressure that de-velops as well as attenuating the mature animal's hy-pertension. In addition, modest Ca2+ restriction accel-erates the early phase of developing hypertension inthe young SHR.
Urinary electrolyte excretion is modified by theCa2+ content of the diet. Predictably, UCaV rises andUp<>4 ^ decreases as dietary Ca2+ content increases. Theincreased UKV may reflect either altered renal handlingor, more simply, increased K+ consumption on the4% Ca2+ diet. Daily sodium excretion varied directlywith Ca2+ content of the diet. There was no variation inUNaV over time, however, thereby dissociating tempo-rally the effect on UNaV from that on the blood pressureattenuation induced by the high Ca2+ diet. Since theanimals were not pair-fed, a change in sodium balancecannot be excluded as a factor in the protective actionof the Ca2+ supplementation diet. Several factorsthough would suggest changes in sodium balance werenot responsible. Body weight was not different among
TABLE 1. Urinary Electrolyte Excretion (Mean ± so) per 24 Hours of Spontaneously Hypertensive Rats Grouped by Percentage of Calciumin the Diet
CALCIUM, MAGNESIUM, PHOSPHORUS, AND BP/McCarron 111-31
the diet groups. Urinary sodium excretion was alreadyincreased at 7 to 12 weeks of age at a time when the 4%animals systolic pressures were similar to the other dietgroups. Lastly, recent studies in the SHR fed 1% vs4% Ca2+ have found no change in total body sodiumcontent (Ritz E, University of Heidelberg, West Ger-many, personal communication, 1982) in the animalschronically fed the 4% diet. This latter observation,combined with our urinary excretion data, suggeststhat diet consumption increases as the Ca2+ content ofthe diet increases. The increased UNaV would thensimply reflect diet intake and not enhanced excretion.Such an interpretation would be most consistent withthe body weight, total body Na+, and blood pressuredata. Additional studies are needed to clarify preciselythe mechanism whereby UNaV stratifies with the per-centage of Ca2+ in the diet.
The specific cellular mechanism(s) whereby in-creasing dietary calcium lowers blood pressure in ex-perimental animals cannot be addressed by the currentstudy. Drawing upon our observations and recent in-vestigations of cellular Ca2+ metabolism in the SHR,however, a unifying theory can be advanced basedupon modifying bioavailable Ca2+ to vascular cellmembranes.
Numerous laboratories have reported cellular abnor-malities of Ca2+ handling in the SHRs. Defects havebeen identified in RBC, 3 3 " intestinal epithelial cells,32
adipocytes,35 aortic,5455 and vascular smooth musclecells.56"59 Mulvaney has recently demonstrated thatthese are intrinsic to the vascular tissue and not func-tional changes due to the increase in mean arterialpressure.60 The cellular abnormalities described to datehave included increased Ca2+ permeability of the plas-ma membrane,56 reduced 45Ca2+ binding to isolatedmembranes following incubation,3457 altered intracel-lular fluxes of Ca2+ between the cytosol and variousintracellular compartments,58 and increased total Ca2+
content of the cell.5961 The observtion that total cellCa2+ is increased, while membrane binding of 45Ca2+
is reduced, suggests that protein binding of Ca2+ isincreased, i.e., Ca2+ is bound more avidly to bindingsites in the plasma membranes of the cells and itsconstituent vesicles of cells. Such a proposed defectparallels the documented increases in extracellularbinding of Ca2+ noted in the SHR18'19 and humans.20
Critical to this interpretation of the literature, are thedata from the 45Ca2+ membrane binding stud-ies . 3 3 3 4 5 3 5 7 Those investigations have shown thatthere is a reduction in the number of available bindingsites for 45Ca2+ when the isotope is incubated withplasma membrane preparations. This observation hasbeen extrapolated so as to estimate total binding sites.33
Several authors have concluded that the total number,occupied and "free," are also reduced. However, anequally plausible interpretation is that total bindingsites are either normal or increased in number, but that"free" sites are reduced, secondary to an alteration inthe kinetics of the cation's binding to membrane pro-teins. This interpretation reconciles the otherwise dis-parate observations that total cell Ca2+ is increased
while binding of 45Ca2+ to plasma membranes isdecreased.
Basal and stimulated vascular smooth muscle tone isdependent upon Ca2+ fluxes through plasma mem-brane-associated channels. While a variety of endog-enous compounds and ions, as well as pharmacologicagents are known to modify Ca2+ channel fluxes, bioa-vailable calcium in the membrane may be the ultimateregulator of these fluxes.762-63 Recent studies havedemonstrated a specific inhibition of Ca2+ membranefluxes by membrane-available Ca2+.M Conditions thatresult in a reduction in bioavailable, membrane-associ-ated Ca2+ may stimulate Ca2+ fluxes, and result inenhanced permeability to Ca2+ and other ions with aconsequent increase in vascular tone.
Maintaining an adequate or supplemental Ca2+ in-take in the SHR may produce the in vivo counterpart ofthe in vitro incubation experiments that have demon-strated the specific inhibition by Ca2+ of calciumfluxes in smooth muscle cells.62'63 The net effect wouldbe a reduction in both basal and stimulated vasculartone. Restricting Ca2+ intake would theoretically pro-duce the opposite results, as membrane-available Ca2+
decreases, vascular ion fluxes increase, vasoconstric-tion occurs, resistance rises, and blood pressure goesup. One would predict that such effects would be ap-parent in normotensive36'37 as well as hypertensive ani-mals and humans. The influence of Ca2+ balance,though, may be magnified in the hypertensive animaldue to its inherent abnormality(ies) of cellular handlingof Ca2+.
These observations of Ca2+ balance, cellular Ca2+
defects, and blood pressure in the experimental modelmay have several implications for human hyperten-sion. First, as sodium intake, and thereby UNaV in-creases in humans, obligatory renal losses of Ca2+
ensue.64 The possibility that sodium exposure in thediet modifies blood pressure in those subjects who are"salt sensitive" via changes in Ca2+ balance needs tobe explored. Second, it is noteworthy that several ofthe societies that clearly differ in their prevalence ofhypertension in spite of similar sodium exposures inthe diet, also differ in their estimated Ca2+ intake.65""*7
Where average daily Ca2+ intake has been maintainednear or above 1000 mg per day, hypertension is infre-quent or nonexistent.66'67 Third, two established thera-peutic modalities that reduce blood pressure in humansalso induce positive Ca2+ balance. They are thiazidediuretics2468 and sodium restriction.69 The possiblecontribution of these changes in Ca2+ balance to thereduction in blood pressure associated with these treat-ment regimens should be explored.
Magnesium and PO4= balance may be equally im-
portant cofactors in this theoretical construct relatingCa2+ balance to hypertension. As noted above, thesetwo divalent ions would appear to facilitate, or be aprerequisite for, many cardiovascular actions of Ca2+.Factors, such as sodium excretion, which promoterenal Ca2+ losses also adversely affect the metabolicbalance of Mg2+ and PO4
=. As the Ca2+ balance issuesin human and experimental hypertension are pursued,
111-32 PERSPECTIVES IN HYPERTENSION SUPPL III, HYPERTENSION, VOL 4, No 5, SEPTEMBER-OCTOBER 1982
it will be essential to characterize equally as well theroles of Mg2+ and PO4
=balance.
ConclusionsDivalent ions are critical to normal cardiovascular
control. Limited observations in normal and hyperten-sive humans and experimental animals suggest thatalterations in the metabolic balance of Ca2+, Mg2+,and PO4
= may contribute to long-term variations inblood pressure. In the experimental model these ef-fects, primarily those of Ca2+, may be linked to identi-fied abnormalities in the cellular Ca2+ metabolism ofthe SHR's vascular tissue. The influence of positiveCa2+ balance, and the apparent generalized defect invascular cell's Ca2+ metabolism, need not be limited tothat tissue.70 They may be applicable to other recentlyhypothesized mechanisms that are believed to contrib-ute to the development of hypertension.70 This expand-ing area of high blood pressure research provides im-portant and obvious interfaces with many establishedand proposed neurohumoral, renal, and sodiummechanisms that have been associated with hyperten-sion.71 Rather than alterations of transcellular fluxesand/or intracellular concentrations of Na+ increasingintracellular Ca2+ and thereby enhancing vascularsmooth muscle tone, as proposed by Blaustein,71 pri-mary abnormalities of cellular Ca2+ metabolism72 maybe responsible for observed differences in cellular so-dium and potassium handling71"74 linked with hyperten-sion. Correction of the Ca2+ defects may then stabilizecell membrane9-a-w and modify vascular smooth mus-cle cell function including ion fluxes.8-9-75 In addition,this hypothesis poses provocative questions concern-ing possible nutritional measures that could be applica-ble to both the prevention and therapy of humanhypertension.
AcknowledgmentsThe author thanks Jennifer Paquet and Joni Utterback for prep-
aration of the manuscript. Cynthia D. Morris for statistical assis-tance, Janet Durow for technical assistance, and Patricia McCarronfor editorial assistance.
References1. Blaustein MP, Ratzlaff RW, Kendrick NK: The regulation of
intracellular calcium in presynaptic nerve terminals. Ann NYAcad Sci 307: 195, 1978
13. Ohhashi T, Azuma T: Contraction of arterial smooth muscleinduced by magnesium ions. Am J Physiol 242: C25, 1982
14. Racker F: Fluxes of Ca2 + and concepts. Federation Proc 39:2422, 1980
15. Lodish HF, Rothman JE: The assembly of cell membranes. SciAm 240: 43, 1979
16. DiPolo R: The influence of nucleotidcs on calcium fluxes. FedProc 35: 2579, 1976
17. McCarron DA, Pingree P, Rubin RJ, Gaucher SM, Molitch M,Kurtzik S: Enhanced parathyroid function in essential hyper-tension: A homeostatic response to a urinary calcium leak.Hypertension 2: 162, 1980
18. Wright G, Toraason MA, Barbe JS, Crouse W: The concentra-tion of ionic and total calcium in plasma of the spontaneouslyhypertensive rat. Can J Physiol Pharmacol 58: 1494, 1980
19. McCarron DA, Yung NN, Ugoretz BA, Krutzik S: Distur-bances of calcium metabolism in the spontaneously hyperten-sive rat. Hypertension 3: 1-162, 1981
20. McCarron DA: Alterations in serum ionized calcium and extra-cellular binding of calcium in essential hypertension (abstr).Kidney Int 21: 191. 1982
21. Bello J, Ruilope L. Alcaraz F, Garcia-Robles R, TresquerresJF, Alcazar JM, Barrientos, Sandho J, RodicioJL: Pathogene-sis of hypertension in renal lithiasis. Mineral Electrolyte Metab6: 302, 1981
23. Christensson T. Hellstrom K. Wenglc B: Hypcrcalccmia andprimary hypcrparathyroidism. Arch Intern Med 137: 1138.1977
24. Popvitzer MM, Subryan VL. Alfrey AC, Reeve EB, SchrierRW: The acute effects of chlorothiazidc on serum-ionized cal-cium: Evidence for a parathyroid hormone-dependent mecha-nism. J Clin Invest 55: 1295. 1975
25. Lafferty FW: Primary hyperparathyroidism, changing clinicalspectrum prevalence of hypertension and discriminate analysisof laboratory test. Arch Intern Med 141: 1761, 1981
26. Hellstrom J, Birk G. Edvall CA: Hypertension in hyperpara-thyroidism. Br J Urol 30: 13. 1958
27. Brinton GS, Jubiz W, Lagerquist LD: Hypertension in primaryhyperparathyroidism: the role of renin-angiotensin system. JClin Endocrinol Metab 41: 1025, 1975
28. Marone C, Beretta-Piccoli C, Weidmann P: Acute hypercalce-mic hypertension in man: role of hemodynamics, catechola-mines, and rcnin. Kidney Int 20: 92, 1981
29. Sowers JR. Barrett JD: Hormonal changes associated withhypertension in neoplasia-induced hypercalcemia. Am J Phys-iol 242: E330, 1982
30. Rodriguez-Sargent C. Cangiano JL, Opava-Spitzer S. Mar-tinez-Maldoneldo M: Renal Na + -K + -ATPase in Okamoto andDohl hypertensive rats. Hypertension 3: 11-86, 1981
31. Wexler BC, McMurtry JP: Kidney and bladder calculi in spon-taneously hypertensive rats. Br J Exp Path 62: 369, 1981
32. Toraason MA, Wright GL: Transport of calcium by duodenumof spontaneously hypertensive rat. Am J Physiol 241: G344,1981
37. McCarron DA: Blood pressure and calcium balance in theWistar-Kyoto rat. Life Sci 30: 683, 1982
38. Bclizan JM, Pineda O, Sainz E, Menendez LA, Villar J: Riseof blood pressure in calcium-deprived pregnant rats. Am JObstet Gynccol 141: 163, 1981
39. Neri LC, Johansen HL: Water hardness and cardiovascularmortality. Ann NY Acad Sci 304: 203, 1978
40. Stitt FW, Crawford MD, Clayton DG, Morris JN: Clinical andbiochemical indicators of cardiovascular disease among menliving in hard and soft water areas. Lancet 1: 122, 1973
41. Belizan JM, Villar J: The relationship between calcium intakeand edema-proteinuria and hypertension-gestosis: An hypoth-esis. Am J Clin Nutr 33: 2202, 1980
42. McCarron DA, Morris CD, Cole C: Dietary calcium and hu-man hypertension. Science (in press)
43. McCarron DA, Morris C: Calcium consumption in humanhypertension: report of a national survey. Clin Res 30: 338A,1982
44. Crass MF, Pang PKT: Parathyroid hormone: A coronary arteryvasodilator. Science 207: 1087, 1980
45. Ellison DE, McCarron DA: Infusion of bovine parathyroidhormone attenuates the pressure response to angiotensin II inspontaneously hypertensive rats. Clin Exp Hyperten (in press)
46. McCarron DA, Muther RS, Plant SB, Krutzike S: Parathyroidhormone: A determinant of post-transplant blood pressure reg-ulation. Am J Kidney Dis 1: 38, 1981
47. Burch GE, Giles TD: The importance of magnesium deficien-cy in cardiovascular disease. Am Heart J 94: 649, 1977
48. Altura BM, Altura BT: Magnesium and vascular tone andreactivity. Blood Vessels 15: 5, 1978
49. Altura BM, Altura BT: Ouabain, membrane Na + . K + -ATPaseand the intracellular action of magnesium ions in arterialsmooth muscle. Artery 3: 72, 1977
50. Manthey J, Stoeppler M, Morgenstern W, Nussel E, OpherkD, Weintraut A, Wesch H, Kiibler W: Magnesium and tracemetals: Risk factors for coronary heart disease? Circulation 64:722, 1981
52. Fuller TJ, Nichols WW, Brenner BJ, Peterson JC: Reversibledepression in myocardial performance in dogs with experimen-tal phosphorus deficiency. J Clin Invest 62: 1194, 1978
53. Postnov YV, Orlov SN, Pokudin NI: Decrease in calciumbinding by the red blood cell membranes in spontaneouslyhypertensive rats and in essential hypertension. Pfliigers Arch379: 191, 1979
54. Zsoter TT, Wolchinsky C, Henein NF, Ho LC: Calcium kinet-ic in the aorta of spontaneously hypertensive rats. CardiovascPrcs 11: 353, 1977
55. Bhalla RC, Webb RC, Singh D, Ashely T, Broch T: Calciumfluxes, calcium binding and adenosine cyclic 3; 5-monophos-phate-dependent protein kinase activity in the aorta of sponta-neously hypertensive and Kyoto Wistar normotensive rats.Mol Pharmacol 14: 468, 1978
56. Noon JP, Rice PJ, Baldessarini RJ: Calcium leakage as a causeof the high resting tension in vascular smooth muscle from thespontaneously hypertensive rat. Proc Nat Acad Sci 75: 1605,1978
57. Aoki K, Yamashita K, Tomita N, Tazumi K, Holta K: ATPaseactivity and Ca2 + binding activity of subcellular membrane ofarterial smooth muscle in spontaneously hypertensive rats. JpnHeart J 15: 180, 1974
58. Webb RC, Bhalla RC: Altered calcium sequestration by sub-cellular fractions of vascular smooth muscle from spontane-ously hypertensive rats. J Mol Cell Cardiol 8: 651, 1976
59. Wei JW, Janis RA, Daniel EE: Calcium accumulation andenzymatic activities of subcellular fractions from aortas andventricles of genetically hypertensive rats. Circ Res 39: 133,1976
60. Mulvany MJ, Korsgaard N, Nyborg N: Evidence that the in-creased calcium sensitivity of resistance vessels in spontane-ously hypertensive rats is an intrinsic defect of the vascularsmooth muscle. Clin Exp Hypertens 3: 749, 1981
61. Tobian L, Binion JT: Tissue cations and water in arterial hy-pertension. Circulation 5: 754, 1952
62. Hurwitz L, Weissimger J: Effects of variations in extracellularacetylcholine and calcium ion concentration on the operationallevel of calcium channels in intestinal smooth muscle. J Phar-macol Exp Ther 214: 581, 1980
63. Hurwitz L, McGuffee LJ, Smith PM, Little SA: Specific inhi-bition of calcium channels by calcium ions in smooth muscle. JPharmacol Exp Ther 220: 382, 1982
64. McCarron DA, Rankin LI, Bennett WM, Krutzik S, McClungMR, LuftFC: Urinary calcium excretion at extremes of sodiumintake in normal man. Am J Nephrol 1: 84, 1981
65. Sasaki N: High blood pressure and the salt intake of the Japa-nese. Jpn Heart J 3: 313, 1962
66: Mann GV, Schaffer RD, Anderson RS, Sandstead HH: Cardio-vascular disease in Masai. J Atheroscler Res 4: 289, 1964
67. Cerqueira MT. Fry M, Connor WE: The food and nutrientintakes of the Tarahumara Indians of Mexico. Am J Clin Nutr32: 905, 1979
68. Suki WN: Effects of diuretics on calcium metabolism. MineralElectrolyte Metab 2: 125, 1979
69. Porter RH, Cox BG, Heaney D, Hostter TH, Stinebaugh BJ,Suki WN: Treatment of hypoparathyroid patients with chlor-thalidone. N Engl J Med 298: 577, 1978
70. Bohr DF: What makes the pressure go up? A hypothesis.Hypertension 3: 11-160, 1981
71. Blaustein MP: Sodium ions, calcium ions, blood pressure reg-ulation and hypertension: a reassessment and a hypothesis. AmJ Physiol 232: C165, 1977
72. Webb RC, Bohr DF: Recent advances in the pathogenesis ofhypertension: consideration of structural, functional and meta-bolic vascular abnormalities resulting in elevated arterial resis-tance. Am Heart J 102: 251, 1981
73. Canessa M, Adragna N, Solomon HS, Connolly TM, TostesonDC: Increased sodium-lithium countertransport in red cells ofpatients with essential hypertension. N Engl J Med 302: 772.1980
74. Woods JW, Falk RJ, Pittman AW, Klemner PJ, Watson BS,Namboodiri K: Increased red-cell sodium-lithium counter-transport in normotensive sons of hypertensive parents. N EnglJ Med 306: 593, 1982
is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Hypertension doi: 10.1161/01.HYP.4.5_Pt_2.III27
1982;4:III27Hypertension.
http://hyper.ahajournals.org/content/4/5_Pt_2/III27World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://hyper.ahajournals.org//subscriptions/
is online at: Hypertension Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer process is available in the
Request Permissions in the middle column of the Web page under Services. Further information about thisOffice. Once the online version of the published article for which permission is being requested is located, click
can be obtained via RightsLink, a service of the Copyright Clearance Center, not the EditorialHypertension Requests for permissions to reproduce figures, tables, or portions of articles originally published inPermissions: