Organization for Flora Neotropica Cecropia Author(s): Cornelis C. Berg, Pilar Franco Rosselli, Diane W. Davidson Source: Flora Neotropica, Vol. 94, Cecropia (May 9, 2005), pp. 1-230 Published by: New York Botanical Garden Press on behalf of Organization for Flora Neotropica Stable URL: http://www.jstor.org/stable/4393938 . Accessed: 09/03/2011 09:12 Your use of the JSTOR archive indicates your acceptance of JSTOR's Terms and Conditions of Use, available at . http://www.jstor.org/page/info/about/policies/terms.jsp. JSTOR's Terms and Conditions of Use provides, in part, that unless you have obtained prior permission, you may not download an entire issue of a journal or multiple copies of articles, and you may use content in the JSTOR archive only for your personal, non-commercial use. Please contact the publisher regarding any further use of this work. Publisher contact information may be obtained at . http://www.jstor.org/action/showPublisher?publisherCode=nybg. . Each copy of any part of a JSTOR transmission must contain the same copyright notice that appears on the screen or printed page of such transmission. JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range of content in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new forms of scholarship. For more information about JSTOR, please contact [email protected]. New York Botanical Garden Press and Organization for Flora Neotropica are collaborating with JSTOR to digitize, preserve and extend access to Flora Neotropica. http://www.jstor.org
Transcript
Organization for Flora Neotropica
CecropiaAuthor(s): Cornelis C. Berg, Pilar Franco Rosselli, Diane W. DavidsonSource: Flora Neotropica, Vol. 94, Cecropia (May 9, 2005), pp. 1-230Published by: New York Botanical Garden Press on behalf of Organization for Flora NeotropicaStable URL: http://www.jstor.org/stable/4393938 .Accessed: 09/03/2011 09:12
Your use of the JSTOR archive indicates your acceptance of JSTOR's Terms and Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp. JSTOR's Terms and Conditions of Use provides, in part, that unlessyou have obtained prior permission, you may not download an entire issue of a journal or multiple copies of articles, and youmay use content in the JSTOR archive only for your personal, non-commercial use.
Please contact the publisher regarding any further use of this work. Publisher contact information may be obtained at .http://www.jstor.org/action/showPublisher?publisherCode=nybg. .
Each copy of any part of a JSTOR transmission must contain the same copyright notice that appears on the screen or printedpage of such transmission.
JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
New York Botanical Garden Press and Organization for Flora Neotropica are collaborating with JSTOR todigitize, preserve and extend access to Flora Neotropica.
CECROPIA CORNELIS C. BERG AND PILAR FRANCO ROSSELLI
WITH A CHAPTER WRITTEN BY
DIANE W. DAVIDSON
rRF)Pg( OF CANCER
FLORAL NEOTROPICA
C, E C R CE -
- -
- -- - -
---
Published for Organization for Flora Neotropica
by The New York Botanical Garden
Bronx, New York
Issued 9 May 2005
? 2005 by The New York Botanical Garden All rights reserved.
Published by The New York Botanical Garden Press
Bronx, NY 10458
International Standard Serial Number 0071-5794
The paper used in this publication meets the requirements of the American National Standard for Information Sciences - Permanence of Paper for Publications and Documents in Libraries and Archives, ANSI/NISO (Z39.48-1992).
Printed in the United States of America using soy-based ink on recycled paper.
MetLife Foundation is a Leadership Funder of The New York Botanical Garden Press.
Library of Congress Cataloging-in-Publication Data
Flora neotropica. - Monograph no. 1 - New York: Published for Organization for Flora Neotropica by The New York Botanical Garden, 1968-
v.: ill.; 26 cm.
Irregular. Each issue has distinctive title. Separately catalogued and classified in LC before monograph no. 40. ISSN 0071-5794 = Flora neotropica.
1. Botany - Latin America - Classification - Collected works. 2. Botany - Tropics - Classification - Collected works. 3. Botany Classification - Collected works. I. Organization for Flora Neotropica. II. New York Botanical Garden.
QK205.F58 581.98'012-dcl9 85-647083
Library of Congress [8508] ISBN 0-89327-461-5
03040506070809 10/98765432 1
CECROPIA CORNELIS C. BERG AND PILAR FRANCO ROSSELLI
Habit ......................................................5 Sap and Wax .....................................................7 Trichomes ......................................................7 Colors ......................................................8 Leaves ......................................................8 Development of Leaves ......................................................9 Trichilia ..................................................... 13 Stipules ..................................................... 14 Inflorescences ..................................................... 14 Spikes of Staminate Inflorescences ..................................................... 16 Staminate Flowers ..................................................... 16 Key to the Types of Stamen ..................................................... 19 Pollen ..................................................... 19 Spikes of Pistillate Inflorescences ................ ..................................... 20 Pistillate Flowers ..................................................... 20 Fruits and Seeds ..................................................... 20 Nature and Variation of Morphological Characters .................................................... 20
Anatomy ..................................................... 21 Protection of Flowers and Inflorescences ..................... ................................ 22 Phenology ..................................................... 22 Pollination ..................................................... 22 Dispersal ..................................................... 23 Survey of Myrmecophytism in the Genus ....................... .............................. 23 Organisms Other than Ants Associated with Cecropia ................................................... 23 Ecology ..................................................... 24 Distribution ..................................................... 25
Representation in the Major Phytogeographic Subdivisions ...................................... 26 Systematic Position ..................................................... 28 Taxonomic Differentiation ..................................................... 29 Uses ..................................................... 31 Vernacular Names ..................................................... 31 Conservation ..................................................... 32 Systematic Treatment ..................................................... 32
Keys to the Species ..................................................... 32 Key to the Species of Central America, Mexico, and the West Indies ............ .......... 32 Key to the Species of Colombia ............... ...................................... 33 Key to the Species of Venezuela ..................................................... 37
2 FLORA NEOTROPICA
Key to the Species of the Guianas ..................................................... 38 Key to the Species of Ecuador ....................................................... 38 Key to the Species of Peru ....................................................... 41 Key to the Species of Amazonian Brazil ....................................................... 44 Key to the Species of Bolivia ............. .......................................... 45 Key to the Species of Extra-Amazonian Brazil, Paraguay, and Argentina .......... ...... 46 Key to the Species of the Cecropia peltata-group ..................................................... 46
Excluded Name ....................................................... 188 Doubtful Names ....................................................... 188 Nomina Nuda ....................................................... 188 Acknowledgments ....................................................... 188 Literature Cited ....................................................... 189 Numerical List of Taxa ....................................................... 196 List of Exsiccatae ....................................................... 196 Cecropia and Its Biotic Defenses (by D. W. Davidson) ................................................ 214
Benefits to Cecropia-ants ....................................................... 214 Benefits to Cecropia ....................................................... 215 Interspecific Variation in Defense Investment ....................................................... 217 Ants Symbiotic with Cecropia ............... ........................................ 220 Coevolution of Ants and Cecropia .................... ................................... 223 Evolutionary Colonization, Host Shifts and Habitat Shifts ................... .................. 225
Index of Scientific Names ....................................................... 227 Index of Vernacular Names ....................................................... 229
ABSTRACT Berg, Cornelis C. (The Norwegian Arboretum/Botanical Institute, University of Ber-
gen, N-5259 Hjellestad, Norway; e-mail: [email protected]), Pilar Franco Ros- selli (t 3 Feb 2000; Universidad Nacional de Colombia, Instituto de Ciencias Naturales, Santafe de Bogota, Colombia), and Diane W. Davidson (University of Utah, 257 South, 1400 East, Salt Lake City, Utah 84112-0840, U.S.A.; e-mail: [email protected] .edu). Cecropia. Flora Neotropica Monograph 94: 1-230. 2005. Cecropia, with 61 species presently recognized, is the largest genus of the Cecropiaceae. It is a genus of pioneer trees in the more or less humid parts of the Neotropics. The majority of the species are myr- mecophytic. The genus is characterized by some unusual traits: spathes fully enclosing the flower-bearing parts of the inflorescences until anthesis, patches of dense indumentum (trichilia) producing Muillerian (food) bodies at the base of the petiole, and anthers be- coming detached at anthesis. The ecological role of the genus and its association with ants have been the subject of numerous studies, most of them cited in the present monograph. Although the taxonomy has been sorted out reasonably well, the coverage by collections and field notes are still insufficient in many cases to fully understand morphological and ecological variation and plasticity in relation to taxonomic delimitations, and in several cases, to prepare complete descriptions. The introductory chapter deals with taxonomic history, morphology, ecology, plant-animal relations, pollination, dispersal, distribution, classification, diversification, use, vernacular names, and conservation. Myrmecophily in Cecropia is treated in a separate chapter (by Davidson). In the present contribution, the new combination Cecropia schreberiana Miquel subsp. antillarum (Snethlage) C. C. Berg & P. Franco is made and numerous lectotypes or neotypes are designated as follows.
Neotypes for: Cecropia concolor Willdenow, C. herthae Diels, C. hololeuca Miquel, C. insignis Liebmann, C. litoralis Snethlage, C. maxima Snethlage, C. palmata Willdenow, C. pastasana Diels, and C. velutinella Diels.
Lectotypes for: Cecropia acutifolia Trecul, C. adenopus Miquel, C. amphichlora Stan- dley & L. 0. Williams, C. angulata I. W. Bailey, C. cyrtostachya Miquel, C. ferreyrae Cuatreccasas, C. ficifolia Snethlage, C. glaziovii Snethlage, C. inchuensis Cuatrecasas, C.
FOREWORD 3
megastachya Cuatrecasas, C. pachystachya Trecul, C. palmatisecta Cuatrecasas, C. por- venirensis Cuatrecasas, C. saxatilis Snethlage, C. scabra Martius, C. strigilosa Cuatreca- sas, C. strigosa Trecul, and C. ulei Snethlage.
RESUMEN Cecropia con 61 especies reconocidas en la actualidad, es el genero mas diversificado
de la familia Cecropiaceae. Sus especies son arboles pioneros en localidades huimedas y muy hdmedas del Neotr6pico. La mayorfa de las especies son mimerc6filas y se caracter- izan por rasgos extraordinarios: espatas que envuelven completamente las partes de la inflorescencia hasta la anthesis, parches de indumento denso (trichilia) que producen los cuerpos de Muller en la base del peciolo, los cuales sirven como fuente de alimento a las hormigas, y anteras que se separan en la antesis. El papel ecol6gico del genero y la aso- ciaci6n con las hormigas han sido fuente de nuemerosos estudios, la mayoria de los cuales se resefian en la presente monograffa. Aunque la taxonomfa esta razonablemente bien clarificada, el cubrimento geografico de las colecciones y las observaciones de campo son insuficientes en numerosos casos para comprender las variaciones morfologicas y ecol6- gicas y la plasticidad, y en otros casos para preparar decripciones completas. El capitulo inicial se relaciona con la historia taxonomica, la morfologia, la relaci6n planta-animal, la polinizaci6n, dispersion, distribuci6n, clasificaci6n, diversificaci6n, usos, nombres comu- nes y conservaci6n.
Cecropia y sus hormigas son tratadas en un capitulo aparte. En esta contribuci6n ig- ualmente se propone la nueva combinaci6n Cecropia schreberiana Miquel subsp. antil- larum (Snethlage) C. C. Berg & P. Franco y se designan numerosos lectotipos y neotipos.
RESUMO Cecropia, com 61 especies atualemente reconhecidas, e o maior genero das Cecropi-
aceae. Apresenta avores pioneiras nas partes umidas ou semi-hutmidas da regiao Neotrop- ical. A maioria das especies e mirmec6fita. 0 genero apresenta algumas caracteristicas impares: espatas cobrindo integralmente as partes florais das inflorescencias ate a antese, partes com denso indumento (triquilios) produtores dos corpu'sculos de Muller (alimentos) na base do peciolo, e anteras tomando-se destacadas na antese. 0 papel ecol6gico do genero e a associac,o com a formigas ja' foram assunto para numerosas pesquisas, muitas das quais referidas na presente monografia. Embora a taxonomia seja razoavelmente con- hecida, os dados existentes nas coleqc,es e notas de campo sao ainda insuficientes em muitos casos para serem esclarecidas a morfologia, variac6es ecol6gicas e plasticidade com respeito as delimita,6es taxonomicas, e em varios casos, na preparac,o de descric,oes completas. 0 capitulo introdut6rio aborda o hist6rico taxonomico, morfologia, ecologia, relac,o planta-animal, polinizac,o, dispersao, distribuicvo, classificaqao, diversificaqao, usos, nomes populares e conservaqao. A mirmecofilia em Cecropia e tratada num capitulo a parte. Na presente contribuicbo consta a nova combinaqao Cecropia schreberiana Miquel subsp. antillarum (Snethlage) C. C. Berg & P. Franco assim do designados numerosos lect6tipos e ne6tipos.
FOREWORD Pilar Franco Rosselli had agreed to participate in
the prepration of a monograph of Cecropia in con- junction with the preparation of a treatment of the genus for Colombia, which would have been a part of her doctoral thesis to be presented at the University of Utrecht (the Netherlands). Before the preparation
of the monograph came into a final stage, Pilar died (03.02.2000) in an accident while making a collection of a species of Cecropia. Our discussions on the rec- ognition and delimitation of some taxa had not fin- ished, in particular the inclusion of C. polyphlebia in C. angustifolia, the way to treat some taxa of the C. peltata-group, and the publication of a new species
4 FLORA NEOTROPICA
(referred to in the comment under C. membranacea). The first author is solely responsible for the final de- cisions on these matters and for the final version of the introduction.
INTRODUCTION "Jeder Naturforscher der im tropischen Amerika je gewesen, wird sich Cecropien ( ... ) wohl erinneren; dieselben gehoren zu den sonderbarsten Biumen je- ner Lander, . . ." (Schimper, 1888: 26).-"Every nat- uralist who has been in tropical America will remember Cecropia, belonging to the most conspic- uous trees of those countries".
The Neotropical genus Cecropia is the largest ge- nus of the Cecropiaceae, with 61 species presently recognized. It is the most important group of pioneer trees in more or less humid regions of the Neotropics and, therefore, very common in these regions. In spite of its common and conspicuous occurrence and its important ecological role, it has been studied little taxonomically and the studies available were not very useful.
There are several reasons for the slow progress of taxonomic knowledge. One reason was (and still is) the scarcity of collections and the poor quality of many of them. Most plant collectors avoid Cecropia because the plant parts are too large to easily make good herbarium specimens, most species are inhab- ited by aggressive ants, and most species are elements of secondary growth. Because the species are super- ficially so similar, even conscientious collectors nor- mally collected either only staminate or pistillate ma- terial of one species and left uncollected the other sex and the other five to seven or so species often occur- ring at a locality. The relative paucity of collections made (and still makes) it difficult to prepare descrip- tions of all details (such as those of stamens) for sev- eral species and to understand the extent of the morphological variation; to evaluate the nature of morphological differences; and to comprehend the ecological variation, the limits of the ranges of dis- tribution, and the reality of disjunctions. It is often still difficult to decide whether differences found in species described from different regions (or even from the same region) justify recognition at the species level. The number of collections of several recognized taxa is so small in relation to the variation that their circumscriptions are incomplete. Because important diagnostic characters were often not included in de- scriptions, even in the extensive ones provided by Cuatrecasas (1945, 1956), identification of material has often been impossible or at least difficult.
TAXONOMIC HISTORY The first accounts on Cecropia are from Marc-
grave (1648) and Piso (1658), the latter with an illus- tration (of "Ambalba") in which the characteristic fea- tures of the genus were depicted (Piso, 1658: 147). The genus name Cecropia was created by Loefling (1758) and is conserved against Coilotapalus Browne (1756). The name Ambaiba, (also) used by Barrere (1741) and Adanson (1763), was reintroduced by Kuntze (1891) who made several new combinations.
Linnaeus (1759) described Cecropia peltata, a name applied to many species for a long time. Two additional species were published by Willdenow (1806), of which C. palmata was also applied to var- ious species. Some additional species were described by Bertoloni (1840), Martius (1841), and Liebmann (1851). The first extensive collections of Cecropia, many specimens of several species, were made by Ruiz and Pavon and distributed to many herbaria, of- ten in mixed sets. This mixing of specimens of more than one species during distribution or mounting is a problem with many older (and even more recent) col- lections of Cecropia. The Ruiz and Pavon material was used for descriptions of several species by Trecul in 1847 and later in the same year by Klotzsch. The type material of the species described by Klotzsch in Berlin was destroyed. Due to the mixing of elements of the Ruiz and Pav6n collections, it is not easy to relate duplicates to the Klotzsch names. The species decribed by Trecul and Klotzsch were included in a survey of the genus by Miquel (1853) in Flora Bras- iliensis, who added several new ones and recognized 37 species for the Neotropics, some of them based on material in cultivation in Hortus Schonbrunn in Vi- enna. For a long time, Miquel's was the only com- prehensive taxonomic treatment for the neotropical region. A number of species were described by Hems- ley (1883), Richter (1897), Donnell Smith (1899), Rusby (1907, 1910), Huber (1910), Robinson (1912), Pittier (1917), and Bailey (1922) before Snethlage started to work on the genus and published a good number of new species (1923, 1924), many of them based on the collections of Ule, who consistently tried to collect sets of both staminate and pistillate mate- rial. In 1923, Snethlage also proposed a subdivision for the genus. Several additional species were pub- lished by Burret (1924), Mildbread (1925, 1933), Standley (1929, 1940a, 1940b), Macbride (1937), Diels (1941), Standley & Steyermark (1944), and Standley & Williams (1952). In the period 1932- 1970, Cuatrecasas made extensive collections of Ce- cropia in Colombia, providing the basis for the pub- lication of numerous Colombian species (1944,
MORPHOLOGY 5
1945). Collections made in later years, also from other parts of South America, provided the material for the establishment of a total of 68 additional spe- cies and several variety names (Cuatrecasas, 1949, 1951, 1956, 1959, 1967, 1971, 1976, 1982). Cuatre- casas planned to revise Cecropia and the related Coussapoa Aublet and Pourouma Aublet (Cuatreca- sas, 1945). Unfortunately, he could not realize his plans. Many of the names established by Cuatrecasas are synonymized in the present treatment. One reason is that he paid little attention to previously published species, possibly as he could not get hold of type ma- terial from European herbaria because of war and postwar circumstances when he started to publish. The other reason is that he did not or could not un- derstand the variation, plasticity, distribution, and ecological amplitudes of most Cecropia species, treat- ing them as "normal" species, clear-cut in their mor- phology, distribution, and ecology. More recently, poor understanding of the nature of variation and dis- tribution patterns in Cecropia caused premature es- tablishment of a number species (Berg & Franco Ros- selli, 1993, 1996).
In 1972 the senior author of this treatment started to work on the genus, initially on the taxa occurring in Amazonian Brazil, later extending the study to in- clude extra-Amazonian Brazil, the Guianas, Vene- zuela, and Peru (Berg, 1972, 1977a, 1978b, 1980, 1981, 1985, 1992, 1998, 2000, 2002; Berg & Carauta, 1986). More recently, these studies included collab- oration with Franco Rosselli in Ecuador and the entire Neotropics (Berg & Franco Rosselli, 1993, 1996).
Regional treatments of the genus include those for Peru (Macbride, 1937), Panama (Woodson & Schery, 1960), Costa Rica (Burger, 1977), Venezuela (Velas- quez, 1971), the species of Kcosniipata and Manu val- leys in Peru (Galiano-Sanchez, 1976), and the state of Rio de Janeiro in Brazil (Duarte, 1959).
The Cecropia-Azteca ant association received at- tention in several papers in the 19th century and the beginning of the 20th century (Muller, 1876; Schim- per, 1888, 1898; Buscalioni & Huber, 1900; Rettig, 1904; Ule, 1900; Ihering, 1907; Fiebrig, 1909; Bailey, 1922; Wheeler & Bequaert, 1929; Wheeler, 1942). Numerous more recent studies on the myrmecophytic traits have been published by Janzen (1969, 1973), Janzen & McKey (1977), Andrade (1984b), Andrade & Carauta (1979, 1982), Longino (1989a, 1989b, 1991a, 1991b), Davidson et al. (1988, 1991), and Davidson & McKey (1993). Other aspects of the bi- ology of Cecropia that have been studied include pop- ulation dynamics (Alvarez-Buyalla & Martinez- Ramos, 1992); the role as pioneer tree in treefall gaps,
forest edges, landslides, etc. (Brokaw, 1985, 1986, 1987, 1998; Guariguata, 1990; Didham & Lawton, 1999; Sposito & Santos, 2001a, 2001b); light regimes (Davidson & Fisher, 1991; Folgarait & Davidson, 1994); dispersal (Charles-Dominique, 1986); seed banks (Holthuijzen & Boerboom, 1982; Vazquez- Yanes & Smith, 1982; Alvarez-Buylla & Martinez, 1990; Garay-Arroyo & Alvarez-Buylla, 1997); con- trol of seed germination (Vazquez-Yanes & Smith, 1982; Vazquez-Yanes & Orozco-Segovia, 1986); seedling and sapling development and photosynthesis (Reekie & Bazzaz, 1989; Strauss-Debenedetti & Baz- zaz, 1991; Poorter & Oberbaurer, 1993); tannin con- tents (Coley, 1986); isozyme variation (Garay-Arroyo & Alvarez-Buylla, 1997); associations with various groups of beetles (Andrade, 1984a; Jolivet, 1987, 1989, 1990b); exploitation by leaf-cutting ants (Vas- concelos & Casimiro, 1987); and presence of lianas, hemiepiphytes, and hemiparasites (Putz, 1982; Clark & Clark, 1990; Jolivet, 1990a).
MORPHOLOGY
HABIT
Cecropia trees are few-branched, usually with a candelabrum-like branching system and with the ar- chitectural Model of "Rauh" (Halle & Oldeman, 1970; Halle et al., 1978). In some species of small trees the branching is reduced, so that the tree is usu- ally or mostly monocaul (as in C. megastachya and C. ulei). A tendency to monocauly is also found in some types of C. obtusifolia in western Colombia (Valle). In many species (e.g., C. concolor and C. fi- cifolia), flowering often starts in the monocaul state. Branch development is often initiated in seedlings, even in the axils of the first formed (opposite) leaves; prophylls are formed, and often the development of the first leaf begins but is arrested (if the seedling is not decapitated). In the axils of the leaves formed dur- ing later development, the axillary branch primordia do not produce more than one or two prophylls and a bud.
In Cecropia garciae and C. hispidissima, branch- ing often occurs at a height of 0.5-1 m; the branches depart at acute angles and the tree has a bushy habit. More commonly, the branches depart at more obtuse angles (at least the lower ones mostly at ca. 450), and the crown becomes more or less distinctly umbrella- shaped. However, in some species, as C. putumayonis and C. utcubambana, as well as forms of C. obtusi- folia, the crown is usually small and more or less glo- bose.
A remarkable variation in branching is found in
6 FLORA NEOTROPICA
Cecropia albicans. In this species, the branches tend to be short, departing at wide angles, or the tree some- times remains unbranched. In a population in Huan- uco, Peru, individuals rarely produce lateral branches departing at wide angles, but often produce branches departing at acute angles in one or two whorls.
Most Cecropia species form small to medium- sized trees, 5-15(-20) m tall; in others, like C. di- stachya, C. herthae, C. insignis, and C. sciadophylla, trees become taller, even to 40 m tall. In others (e.g., C. ulei) the height rarely surpasses 5 m. The size of the trees shows regional variation in some species, such as C. pachystachya, which is represented by a small type in restingas of eastern Brazil (Andrade & Carauta, 1981), and C. concolor near Manaus.
A conspicuous variation in habit is found in the co-occurring Cecropia concolor and C. polystachya in Santa Cruz, Bolivia, at the (climatic) limit of their distribution. Here, trees are robust with relatively short, thick stems and branches with short internodes and relatively dense and wide umbrella-shaped crowns. This tree shape is typical of montane species, such as C. telenitida or C. plicata. Individuals with this tree shape are found within short distances of others with the shape normally found in the two spe- cies. This variation in habit occurs occasionally in other species, e.g., a large tree of C. putumayonis with a broad umbrella-shaped crown (near Villavicencio, Meta, Colombia), apparently in the periphery of its range. Individuals of C. obtusifolia are atypically large-canopy trees in Veracruz, Mexico, and are therewith extending longevity (Alvarez-Buylla & Martinez-Ramos, 1992) and those of C. membrancea often are large-canopy trees outside their typical hab- itat (see p. 119).
The differences in tree shape and size appear to be related to longevity (see Sposito & Santos, 2001a, 2001b); the large or robust specimens (rare in some species, but common in others) are less ephemeral than normal in the genus (see Vester, 1997 for Ce- cropia distachya, C. ficifolia, and C. sciadophylla). According to Duarte (1959), trees of C. hololeuca can become more than 100 years old, whereas those of C. glaziovii have a lifespan of 30-40 years. The mono- caul treelets of C. ulei live for only a few years. This suggests that the tree shape as found in montane spe- cies could be (at least partly) ecologically defined (by climatic conditions, affecting growth rate) rather than genetically. From the variation in habit described above, it is clear that various factors play a role in shaping the trees of many Cecropia species.
All Cecropiaceae can produce adventitious roots. In Cecropia, they become stilt-roots, which are com-
mon and manifest, especially in large trees and in trees growing along rivers or in marshy places.
Although Cecropia trees often occur in sites rich in vines, they are rarely overgrown by them (cf. Clark & Clark, 1990), even individuals that are not inhab- ited by Azteca ants (cf. Putz, 1984; Putz & Holbrook, 1988). Vines can often be observed on the stem and the lower parts of the lower branches, but not higher up. This also applies to the non-myrmecophytic Af- rican sister-genus Musanga R. Brown with a habit similar to that of most Cecropia species. The angle of ca. 450 at which (at least) the lower branches depart from the trunk in these genera may affect the growth of vines on the tree. In (still) unbranched specimens, the rapid continuous growth and the continuous loss of the lower leaves (and handholds for vines) may reduce possibilities to become overgrown.
In most species, the internodes are hollow and contain sparse, usually whitish pith. Such internodes provide nesting space for ants. However, in some spe- cies the intemodes are usually or often filled with brown pith either completely (as Cecropia bullata and C. gabrielis) or partly (as C. schreberiana and C. te- lealba). In some other species, abundant brown pith is found occasionally (as in C. strigosa) or consis- tently in parts of the species range (in C. angustifolia). Abundant pith is found mostly in montane species, C. schreberiana being a distinct exception. The presence of abundant pith could be related to a low growth rate.
In young plants, the diameter of the stem often increases considerably upward, either gradually or more or less abruptly. The walls of the inflated inter- nodes are thin. In some species, Cecropia albicans and C. elongata, this phenomenon can also be en- countered in adult trees. Stems with inflated inter- nodes are also found in C. angustifolia in Ecuador.
Most species have distinct prostomata, thin spots in the upper part of the wall of the interuodes. They are located above the insertion of the petiole, as a small depression, often at the end of a longitudinal groove that starts just above the middle of the petiole. The development of the groove and prostoma (or "diaphragma") is described and discussed by Schim- per (1888).
In trees occupied by ants, the prostomata are mostly opened and thus present as small holes. By perforating prostomata, ant queens enter the stem and establish colonies. In non-myrmecophytic species, the prostomata can be either obvious, as in Cecropia scia- dophylla (see Bailey, 1922: 385) or are more or less indistinct to (virtually) absent, as in C. hololeuca (see Schimper, 1888: fig. 7; Schimper, 1898: fig. 82) and several Andean species in which the outer woody wall
MORPHOLOGY 7
of the internodes of leafy twigs (with short inter- nodes) is relatively thick; even if prostomata depres- sions can be detected, they cannot be penetrated by ants.
Whether occurrence of pith in the internodes and the absence of (distinct or functional) prostomata can be regarded as primitive states or as consequences of growth rate, which tends to be less in montane than in lowland species, is not clear, but these features af- fect inhabitation by ants.
The length of the internodes varies. In young plants, the internodes are long, but in many species the internodes become short (0.5-2 cm long) in adult trees, especially in the distal parts of the branches. In other species, the internodes remain long (>2 cm) in adult trees, especially those of the continuously fast growing, "weedy" species (see below). Shortening of the internodes is correlated with harder internode walls.
On the stem and main branches, the scars of the stipules are usually conspicuous and in some species (e.g., Cecropia annulata, C. engleriana, and C. lito- ralis) present as prominent annular ridges.
SAP AND WAX
Most parts, like young branches, petioles, and pe- duncles, exude a watery, often somewhat mucilagi- nous sap, when cut. Exposure to the air turns the sap black (and can cause persistent dark stains on cloth- ing). Ducts and cells containing the mucilaginous sap are common in Cecropiaceae (cf. Renner, 1907). In some montane non-myrmecophytic species (e.g., Cecropia telenitida), mucilage may fill the space in the terminal buds and in the upper internodes. This prevents inhabitation by ants but also prevents or re- duces occupation and damage by herbivorous insect larvae.
In several species, leafy twigs are covered by a waxy layer making them bluish. Such a layer is al- ways present in Cecropia andina; in some other spe- cies it is often, occasionally, or (in C. albicans) re- gionally present. This layer is found only on (sub)glabrous twigs. However, leafy twigs with waxy surfaces or dense (villous) indumentum can be found in individuals of the same species (e.g., C. telenitida).
Tannin is often abundant (Renner, 1907), as in the rachises of the spikes of the inflorescence, the apices of the perianths of pistillate flowers, and in the peri- carps.
TRICHOMES
The abundance and composition of the indumen- tum are important features in Cecropia, morphologi-
cally and partly also functionally. Six types of tri- chomes can be recognized.
1. Unicellular hairs which are relatively thick, straight, curved, uncinate, or crinkled. In some spe- cies these hairs can be setose, and, e.g., in Cecropia hispidissima and C. megastachya placed in sockets and resembling the urticating trichomes of Urtica- ceae. Such hairs easily break at the sockets, causing rough surfaces. They easily penetrate and irritate the skin. Uncinate hairs are very common. Their absence is a differentiating character.
2. Unicellular hairs which are very thin and in- terwoven, usually white but sometimes brownish. This arachnoid or cobwebby indumentum is found on several parts of the plant. Dense covers of arach- noid indumentum, causing white or whitish surfaces, are common on the spathe, the stipules, the petiole, and the upper and lower surfaces of the lamina. On the lower surface, this indumentum can be very short and confined to the areoles. Arachnoid indumentum of longe hairs is always present on the outer surface of the perianth of the pistillate flower, shorter hairs are often found inside. If arachnoid indumentum oc- curs on the outer surface of the perianth of staminate flowers, then it is usually short. Hairs as long as those on the perianth of the pistillate flower are found in some species, e.g., Cecropia glaziovii and C. palmata.
3. Pluricellular trichomes. They are either elon- gate and usually more or less clearly moniliform or shortened and more or less globular. They are mostly pale to dark brown or sometimes reddish or purplish. They are commonly found on young parts. In dry ma- terial, the indumentum of pluricellular trichomes is usually manifest as a powdery layer.
4. Cystolith hairs. These protruding cystoliths are found at the upper surface of the lamina and cause a rough surface.
5. Pearl glands (or "pearl bodies"). These pluri- cellular trichomes are ca. 1-2 mm long, (sub)clavate, and whitish opaque. According to Rickson (1976), they contain a few glycogen plastids and lipids. These trichomes are present in all (or most) species. Because they are apparently soon harvested by ants (cf. An- drade, 1 984a), they are not often observed in the field. They are abundantly present on young parts of plants grown in greenhouses. Pearl glands are also found in Coussapoa and Myrianthus P. Beauvois (other genera of the Cecropiaceae).
6. Mullerian bodies. These trichomes occur only in patches (or cushions) of dense indumentum (tri- chilia, see below). They are ellipsoid to oblongoid, mostly ca. 1-2 mm long, and whitish; in Cecropia chlorostachya they are pinkish and in C. hispidissima
8 FLORA NEOTROPICA
pink and ca. 3 mm long. Their inner cells are filled with glycogen plastids, and in the apex of the tri- chome the epidermis is provided with a stoma (Schimper, 1888; Renner, 1907; Rickson, 1976). They abscise when full-grown and are "pushed" upward to the surface of the trichilia by the surrounding dense indumentum, consisting largely of pluricellular tri- chomes. Moreover, this indumentum keeps the Mill- lerian bodies at the surface of the trichilia and delays dropping from the trichilium (Schimper, 1888). The bodies are rich in lipid and also contain protein and glycogen (Marshall & Rickson, 1973; Rickson, 1973). These trichomes can be regarded as modified pearl glands. A detailed account of the development of trichilia and Miillerian bodies was provided by Rickson (1976). The Mullerian bodies are the main food source for ants associated with Cecropia but are also eaten or collected by other animals: beetles (cf. Andrade, 1984a), bees, and small birds. In material grown in greenhouses (where Mullerian bodies are not removed by animals), the trichilia are often soon covered by fungi. Andrade (1982) suggested that Muillerian bodies could have a protective function equivalent to that of extrafloral nectaries. Food bodies are also found in the swollen and saccate base of the petiole of Pourouma formicarum and P myrmeco- phila (Benson, 1985; Berg et al., 1990). Whether these food bodies have the same structure and con- tents as Miillerian bodies is unknown.
The indumentum varies considerably within spe- cies. This is most conspicuous for the white arachnoid indumentum. Dense (villous or velutinous) indumen- tum on leafy twigs-as found in Cecropia angusti- folia, C. albicans, C. bullata, C. telenitida, and C. velutinella-may affect the inhabitation by ants, hindering locomotion. In some of these species (C. albicans and C. telenitida), both this type of dense indumentum and very smooth, waxy twig surfaces occur. Scabrous surfaces are caused by cystolith hairs, short and rigid unicellular hairs, or bases of bristle- like hairs.
COLORS
Red-coloring substances are quite common in var- ious plant parts, in particular the stipules, the spathes, and the main veins of the lamina. The concentration of these substances varies, even within species, col- oring the parts pale pink to dark red, or even dark purple to almost blackish. The coloring substances may be distributed equally or in patterns, e.g., lon- gitudinal stripes. Colors may fade with age. In some species (e.g., Cecropia membranacea) red-coloring substances are rare and all parts are usually green. In
several species, both red morphs (with red-colored stipules, etc.) and green morphs (with green-colored stipules, etc.) occur. Both morphs can be found side by side throughout the range of the species (as in C. heterochroma) or in separate parts of its distribution ranges (as in C. angustifolia). Proportions of red and green morphs vary within populations. Leaves of young plants of many species are often silvery mac- ulate.
Colors of plant parts of Cecropia can also be de- termined or influenced by dense indumentum. The white arachnoid indumentum can be so dense as to mask underlying reddish colors such that spathes, stipules, or leaf surfaces appear whitish. In some spe- cies-in particular several montane Andean species (C. albicans, C. maxima, C. telealba, and C. teleni- tida), but also in the Brazilian C. hololeuca, which occurs at low elevations-the layer of arachnoid in- dumentum on the upper surface of the lamina is usu- ally so dense that the trees can then be detected at great distances by their white leaves. More or less white and green surfaces may occur in the same spe- cies. In C. pastasana, the lamina is often distinctly white above in the southern part of its range but only slightly so in the northern part. In C. reticulata, the lamina is usually green but sometimes whitish above. In C. maxima, the lamina is normally white in pop- ulations occurring at 1500-2400 m, but in a popula- tion at ca. 1000 m (in Narifno, Colombia) the leaves are green above.
The brown to purplish (or blackish) pluricellular trichomes can be so dense that they impart these col- ors to the plant parts. In Cecropia angustifolia, brown trichomes on leafy twigs may turn black and make younger parts of the stems blackish.
In some species, young plant parts are covered by a bluish waxy cover; always in the stems of C. andina and sometimes in C. albicans and C. pastasana. Len- ticels, in particular large white and pink ones, may add to the color range, particularly on leafy twigs.
LEAVES
In all Cecropia species, the leaves of adult speci- mens are large and peltate, almost circular in circum- ference. The lamina is excentrically attached to the petiole. The venation is radiate and the lamina is usu- ally radially incised in between the radiating main veins. Incisions can run down to the petiole, and the leaf segments can then be petiolulate (in C. sciado- phylla). In some species (e.g., C. putumayonis and C. subintegra), the lower part of the lamina is hardly or not incised. The number of lobes or leaf segments vary from 5 to more than 20. Within the species, the
MORPHOLOGY 9
number of segments of the lamina mostly varies within certain limits.
In some species (e.g., Cecropia pachystachya and C. polystachya), the upper lamina segments are often or usually lobed; the lobes are usually more frequent and pronounced in subjuvenile stages than in adult specimens.
The main pattern of venation is rather uniform in the genus. However, the details of the venation may vary somewhat to considerably within many of the species. The variation is partly caused by shifting of features of the subjuvenile state to those of the adult state (or vice versa). In spite of the variation, the ve- nation provides important diagnostic characters, such as the number of secondaries of the main radiating veins, the branching of the secondaries, and the loop- connections of the secondaries in the margin or (sometimes closely) near the margin: in the descrip- tions and keys this is indicated as marginally and sub- marginally loop-connected. The venation between the secondaries is scalariform with parallel veinlets.
The lamina can be coriaceous (to subcoriaceous) or chartaceous (to subcoriaceous). Coriaceous lami- nas are often smooth, and chartaceous ones often more or less scabrous. Segments of coriaceous lami- nas are usually plicate in Cecropia plicata or some- times in C. insignis, C. strigosa, and C. telealba. In some species the upper surface is usually more or less bullate in C. bullata and often so in C. andina.
The length of the petiole (in adult trees) is often proportional to (and often approximately as long as) the length (and width) of the lamina. However, in some species, e.g., Cecropia marginalis and C. vir- gusa, the petiole is about half as long as the lamina, with a maximum length of ca. 40 cm in adult speci- mens.
In the majority of the species the lamina is folded in the bud and extended from the petiole; in Cecropia angustifolia and C. montana the lamina is often re- flexed around the relatively long petiole in the bud.
The circular area where the radiating veins meet at the upper surface of the lamina is in some species descriptions indicated as the "umbilicus."
DEVELOPMENT OF THE LEAVES
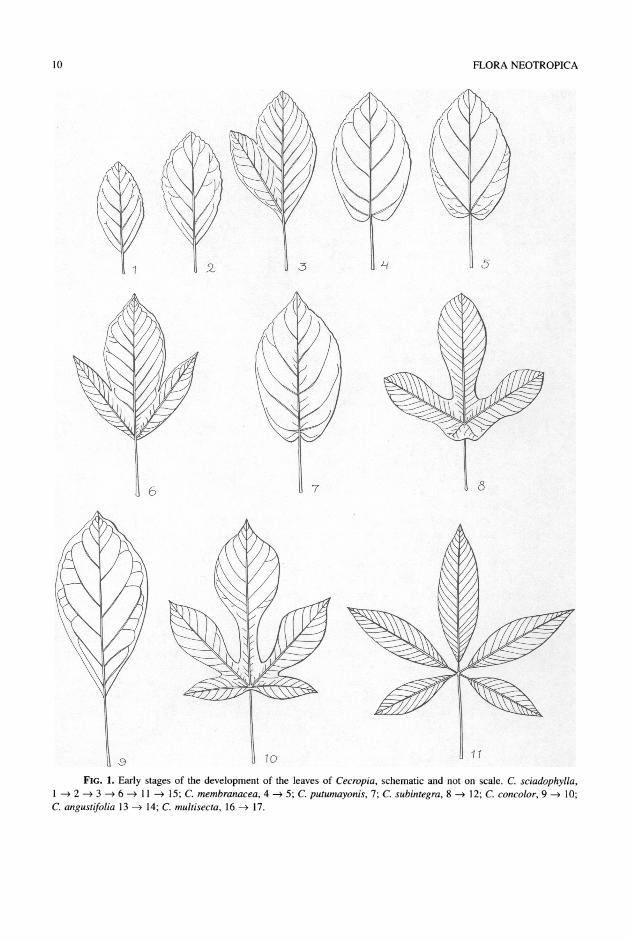
The cotyledons are small and somewhat cartilag- inous, and lack chlorophyll and venation. The first trophophylls are opposite. The two sets of opposite leaves are succeeded by leaves arranged in spirals. The first formed leaves are pinnately veined and entire and have a basally attached lamina and a short petiole (Figs. 1. 1, 1.2, 1.9). Later leaves have a longer petiole and the lamina lobed on one side or both sides
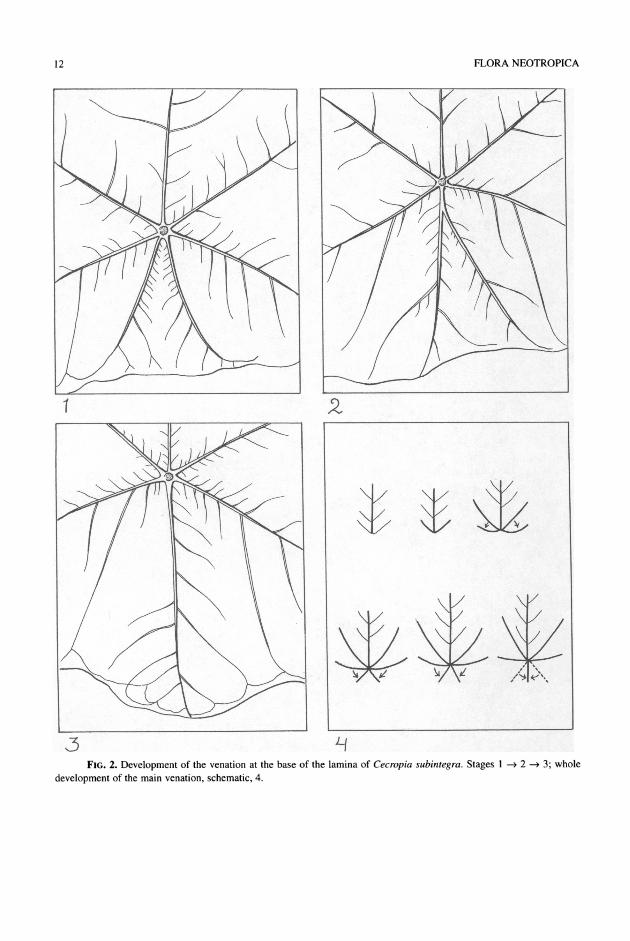
(Figs. 1.3, 1.6). Even later leaves have inequilateral laminas of which the corresponding lobes of both sides are not equally long. Lobation is correlated with a change in venation. Initially one or two lateral veins, usually not the basal ones, become stronger (Figs. 1.5, 1.13). However, the formation of lobes is soon asso- ciated with the basal pair of lateral veins. The tran- sition from the 3-lobate state to the 5-lobate one goes with the formation of strong veins departing from the base of the strong primary basal lateral veins (Fig. 1.8). The base of the lamina of the first formed leaves is acute to rounded (Figs. 1.1, 1.2, 1.9), de- pending on the species. Then the base becomes cor- date and successively peltate (Figs. 1.7, 1.9). Pelta- tion may precede lobation or may happen after the first pair or two of lobes have been formed. In species with laminas with the radial incisions down to or nearly down to the petiole, as Cecropia sciadophylla or C. concolor, the lamina becomes peltate when the sixth lobe is formed, opposite the midlobe (or mid- segment) (Figs. 1.14, 1.15). In all species the sixth lobe or segment is formed opposite the midsegment. The fact that there are often two veins running close together opposite the primary midrib, or that the mid- rib of the sixth lobe is often furcate, suggests that the midrib of the sixth lobe (and other lobes opposite the midsegment) is formed by fusion of two veins
(Fig. 2). The next step is the formation of a lamina with
seven lobes (Figs. 1.16, 1.17). The midribs of these lobes depart from the bases of the primary basal lat- eral veins. The increase of the number of lobes or segments is caused by the successive formation of branches from the base of the (original) pair of basal veins in basipetal direction: thus from 3 -X 5 -> 6 -*
7 -> 8 -> 9 -X 10 -lO 1 and so forth, up to 24 (in
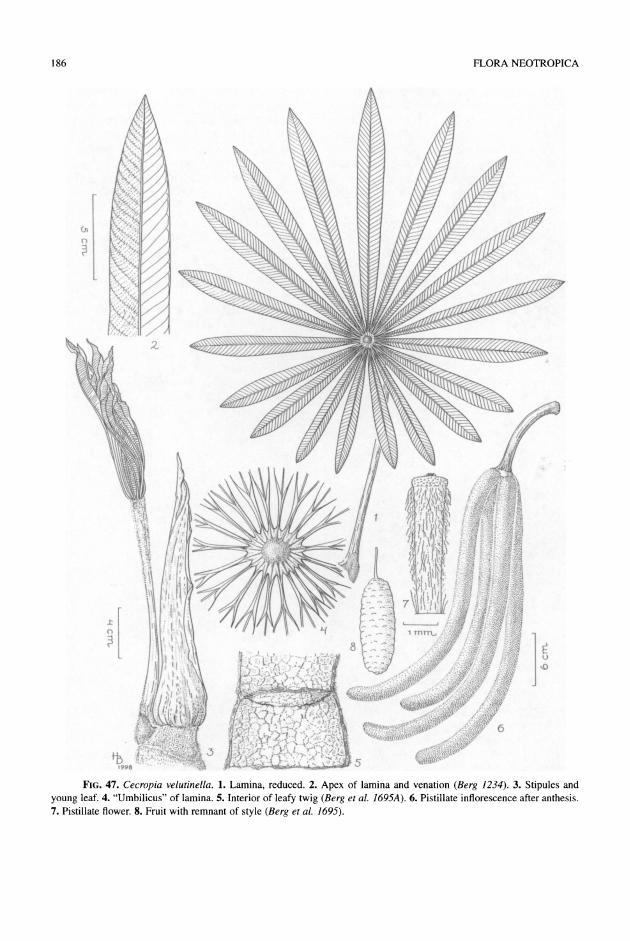
Cecropia velutinella). The even numbers of lobes are caused by the presence of a segment opposite the mid- segment. The attachment of the lamina to the petiole moves from the base toward the center of the lamina, but remaining more or less pronouncedly excentric.
These developments also affect other features of the venation, with an often considerable increase in the number of lateral veins (Figs. 1.2, 1.3, 1.13, 1.14).
The events of the development as described above are (at least in part) repeated on the first nodes of the branches.
Before branching, the leaves on the stem are often larger, are more deeply incised, and often have more segments and more lateral veins in the free part of the midsegment; in certain species the free parts of the segments are more pronouncedly lobate than those on the branches. Such subjuvenile traits can be retained
10 FLORA NEOTROPICA
~~~2 }
6 7 810
10~~~~~~~~~~~1 l 9 C 10
FIG. 1. Early stages of the development of the leaves of Cecropia, schematic and not on scale. C. sciadophylla, 1 2 -> 3 -* 6 -* 11 -> 15; C. membranacea, 4 -* 5; C. putumayonis, 7; C. subintegra, 8 -> 12; C. concolor, 9 -* 10; C. angustifolia 13 -* 14; C. multisecta, 16 -> 17.
MORPHOLOGY 11
1z 13
1 F6 171 FIG. 1. Continued
12 FLORA NEOTROPICA
1 2.
FIG. 2. Development of the venation at the base of the lamina of Cecropia subintegra. Stages 1 2 - 3; whole
development of the main venation, schematic, 4.
MORPHOLOGY 13
in leaves on the branches to some extent. A remark- able situation is found in Cecropia subintegra, in which the leaves of the main stem are deeply incised with numerous segments, whereas the leaves of the branches are subentire with relatively few radiating main veins.
Laminas with incisions down to the petiole or nearly so reach the basic shape of the lamina rather quickly in the process of leaf development. In these species, further development implies the increase of the number of segments, usually without showing in- termediate stages. This type of lamina is rather con- stant in its features. Another type of lamina mostly showing stability in its features is the one with rela- tively few (ca. 7) radiating main veins and a shallowly incised or (sub)entire base. Species with intermediate traits of the lamina often show considerable variation with regard to the number of segments and the extent of the incisions.
The various traits of the development of the lamina are presented in Figures 1 and 2.
TRICHILIA
In most species of Cecropia, one or two patches of dense indumentum are found abaxially at the broadened base of the petiole of the adult leaf. They consist of various types of trichomes: whitish (to brownish) unicellular and more or less stiff hairs of various lengths, dense and relatively long brown pluricellular trichomes, and Mullerian bodies (see above). These characteristic concentrations of tri- chomes are called trichilia. In literture the trichilium has been called "pulvinus," "giba," and, in the Spanish descriptions by Cuatrecasas (1945, 1956), "cojin" or "almohadilla basilar."
The Mtillerian bodies are usually initially hidden by relatively long and very dense brown pluricellular hairs, probably to prevent precocious harvesting (by animals other than Azteca ants inhabiting the tree). When "mature," the Muillerian bodies become de- tached from their bases and they are pushed upward to the surface of the trichilia by the surrounding dense indumentum and are "released" in the course of some weeks. A single trichilium may produce 2500-8000 Muillerian bodies (Janzen, 1973) and/or ca. 10 g Mill- lerian body tissue a week (Rickson, 1973) in C. pel- tata.
The unicellular hairs intermixed with the brown indumentum of pluricellular hairs vary in density and length: from about as long as to somewhat longer to conspicuously longer than the brown pluricellular trichomes. The trichilia may vary more or less in size
and shape. In some species (as Cecropia pastasana and C. reticulata), the trichilia cover the scrotiform swollen bases of the petioles. In some other species (as C. herthae), the trichilia cover shallow depres- sions at the base of the petiole.
The trichilia of Cecropia virgusa are hidden in lat- eral (depressions) pockets with a slit-shaped opening at the base of the petiole. The pockets develop as grooves departing from the base of the petiole and become longer and deeper before trichilia are formed.
In Cecropia hispidissima, unlike the other species, the pluricellular hairs are short and do not cover the relatively large (ca. 3 mm long) Miillerian bodies. The trichilia are often not clearly defined patches and dif- fer in size. They may even extend to the base of the stipules or may be almost absent. This species is in- habited by ants of the genus Pachycondyla, which harvest the Miillerian bodies when the trichilia are still enclosed by the stipules. These ants penetrate the node septum below the bud cover to enter the space containing the Mullerian bodies.
The majority of the species have a single trichil- ium, median and extending laterally. Cecropia mar- ginalis and C. strigosa usually or often have two lat- eral trichilia; they are fuly developed. Separate trichilia are often found in species in which trichilia are often or sometimes absent. In these species tri- chilia occur in a "primordial" state, i.e, as two small patches with only unicellular hairs, as small patches (of equal or unequal size) also with pluricellular hairs, or as larger patches also containing Mullerian bodies, and finally large fused patches. This appears to indi- cate that fused trichilia represent the advanced state. In species with consistently fully developed fused tri- chilia, the first formed trichilia on the sapling are al- ready fused. In these the first trichilia (initially usually without Mullerian bodies) appear on the leaves (5th- 10th formed) with long petioles and peltate laminas. However, in some species (C. latiloba and C. mem- branacea, both of periodically inundated habitats), trichilia may appear much later in the development of the trees, at about the 40th-5Oth leaf formed after the cotyledons (Berg, 1978b), as observed in material cul- tivated in a greenhouse from seeds obtained from an area near Manaus, Amazonas, Brazil. But, at least in aberrant types of C. membranacea, trichilia can al- ready be present in a very early stage of development. This variation could be related to differences in the degree of inundation.
Trichilia are always completely absent in some species: Cecropia hololeuca, C. pittieri, C. sciado- phylla, and C. tacuna. In several species, trichilia may
14 FLORA NEOTROPICA
be absent or present, and if present, in various states of development. In C. schreberiana, trichilia (often reduced and two per petiole base) are sometimes pres- ent but, according to Rickson (1977), without Mill- lerian bodies (or with perhaps abnormal ones; cf. Wheeler, 1942). In other species, absence, presence, and different degrees of development of the trichilia appear to be accidental, while in still others these states are characteristics of populations or certain parts of the distribution range. Clear correlation with age or habitat are wanting. Nor does absence, pres- ence, or state of the trichilia appear to be correlated with the amount of pith in the internodes and, therewith, the suitability for inhabitation by ants. In one of the morphological types of C. angustifolia, the internodes are filled with pith but the trichilia are mostly fully developed, yet in C. gabrielis, also in which the intemodes are filled with pith, trichilia may be absent, more or less reduced, or fully developed. These different stages of development also occur in C. telenitida, in which, however, the intemodes are clearly hollow. In C. albicans, trichilia are present only in some parts of the species' range and are as- sociated with differences in indumentum and tree shape.
Absence of trichilia or presence in various states is usually associated with species of montane or sub- montane habitats in the Andes. Trichilia are always absent in the lowland species Cecropia pittieri, C. hololeuca, and C. sciadophylla, and mostly in C. schreberiana.
In most other Cecropia species, the majority of which occur in lowland habitats, trichilia are always present and well-developed. Cecropia peltata is a re- markable exception, for the trichilia are well- developed throughout its wide range of distribution except in Jamaica, where trees with and without tri- chilia can be encountered. In Africa, Asia, and the Pacific, where this species is introduced, trichilia are absent or poorly developed.
The types of trichomes found in trichilia are com- mon in Cecropiaceae, but their concentration into more or less clearly defined patches and the transfor- mation of pearl glands into Mullerian bodies is char- acteristic only of Cecropia. The trichilium is one of the requisites for the association with ants. The pres- ence of ants can be related to prevention of damage by fungi and insects (cf. Bailey, 1922: 388). The hol- low leafy stems with thin internode walls and the ap- ical meristem are susceptible to damage. Their pro- tection, particularly in early stages of development, i.e., before branching takes place, promotes the rapid and continous growth of the pioneer tree.
STIPULES
The stipules in Cecropia are always fully amplex- icaul and fused. They vary in length from about 5 to 50 cm, often depending on the diameter of stems and leafy twigs in species. However, some species with relatively slender stems and branches have long stip- ules and then often terminal buds with a curved apex (e.g., C. marginalis and C. virgusa). The stipules are usually caducous, but in some species (e.g., C. chlo- rostachya and C. palmata) they remain temporarily attached to the stem, and in other species (e.g., C. angustifolia and C. engleriana) they are sometimes or often subpersistent, or they may be only subper- sistent in juvenile or subjuvenile stages (e.g., C. gar- ciae and C. velutinella). In some species (C. albicans and C. strigosa), the terminal buds are more or less inflated. In slow-growing species the stipules often enclose (and protect) the inflorescences until near an- thesis; and in C. hololeuca, in which the inflores- cences lack spathes, they are enclosed until anthesis. In general, the duration of the protection of young leaves and inflorescences is greater in species with longer stipules.
INFLORESCENCES
There are usually two inflorescences per leaf axil with the lateral bud between them (Fig. 3). One of the two inflorescences may be abortive.
In many species, each of the inflorescences is sub- tended by a basal bract, which is probably a prophyll. These bracts are sometimes inserted on the peduncle (e.g., in Cecropia albicans). They vary in length from ca. 0.5 to 8 cm and usually have the same texture and indumentum as the stipules. They are mostly cadu- cous and, therefore, not present in herbarium mate- rial. There are sometimes bracts at the bases of the spikes; they are small but occasionally grow to 2.5 cm long.
The inflorescence consists of a common peduncle bearing a number of spikes (or spadices) initially completely enclosed by a spathe (Fig. 3). At anthesis, the spathe opens and drops. Cecropia hololeuca is the only species without a spathe. Here, the spikes are subtended by one or two free bracts, to 8 cm long. They do not enclose the spikes. In this species, the spikes are enveloped by the terminal bud cover (formed by the stipules) until anthesis.
The spathe is usually somewhat longer than the longest spikes just before anthesis. Sometimes the spathe is shorter and then the spikes are curved, crin- kled, or coiled, but if straight then piercing the apex of the spathe. The spathe opens adaxially before drop- ping. In staminate inflorescences of Cecropia stri-
MORPHOLOGY 15
1 Z FIG. 3. Structure of the inflorescence of Cecropia,
schematic. 1. Pair of inflorescences with the axillary bud in between and subtended by basal bracts, the spikes fully en- veloped by a spathe, reduced/modified flowers in between the (stipes of the) spikes; 2. Spike with flowers and hairs, at the base of the spike a modified flower.
gosa, the spathe may remain hanging on the one or two longest spikes. The spathes are similar to the stip- ules in texture, color, and indumentum.
The spikes are sessile or stipitate; the stipe (or the pedicel or peduncle of some authors) is the basal part of the rachis. The flowers are densely set on the rachis and not intermixed with interfloral bracts. The rachis is often hairy, mostly with short stiff hairs, sometimes also with soft crinkled hairs. If perianths are basally fused, then stiff short hairs, similar to those on the rachis, are found on the perianths just above the fused parts.
The number of spikes of the pistillate inflores- cence is usually four but may be one (Cecropia ga- brielis) or as many as 20 (C. garciae and C. hispidis- sima).
The staminate inflorescences usually have more spikes, even up to 50 spikes in, e.g., Cecropia mar- ginalis and C. membranacea), or up to 100 in C. her- thae. In some species, e.g., C. palmata and C. putu- mayonis, four spikes are common. Inflorescences with less than four spikes are occasionally found.
The number of spikes of both staminate and pis- tillate inflorescences usually varies more or less within a species, more in staminate inflorescences than in pistillate ones. The first formed inflorescences on the plant and the branches tend to be relatively small and often have fewer and shorter spikes than the succeeding ones. Poorly developed trees also tend to
FIG. 4. Basic structure of the inflorescence of Cecropiaceae, as still largely present in Myrianthus, schematic. (Redrawn from Bull. Jard. Bot. Etat 46: 475. 1976.)
16 FLORA NEOTROPICA
have shorter and fewer spikes than vigorously grow- ing trees. Independent of the variation related to the stage or state of development of trees, the length of the peduncle, the length and the number of spikes, and/or the length of the stipes, may also vary from one part of the species range of distribution to another. In Cecropia obtusifolia, specimens with conspicu- ously different spikes are found side by side.
The diameter of the spikes may vary considerably, especially the staminate ones, partly in relation to the number of spikes per inflorescence. Usually the more spikes present, the thinner they are. Spikes, in partic- ular pistillate ones, are sometimes (dichotomously) branched. The spikes of the pistillate inflorescences of Cecropia goudotiana are two by two basally fused.
In both staminate and pistillate inflorescences, sol- itary flowers are present at or among the bases of the spikes (Fig. 3). They are more conspicuous in pistil- late inflorescences than in staminate ones. These flow- ers are often more or less reduced and modified, the pistillate ones often with more or less clearly cup- shaped perianth containing some fluid (nectar?, cf. Andrade, 1984c). The position of these flowers is of- ten irregular, however, with a tendency to occur in between two spikes. Moreover, minute swollen bracts may occur, often accompanying the solitary flowers.
The length of the peduncle, stipe, and spike varies considerably within the genus, and may vary in spe- cies as well. Depending on the length and the rigidity of the peduncle, the stipe, and the rachis, the stami- nate inflorescence may have an erect peduncle and erect spikes, an erect peduncle and pendulous spikes, or a pendulous peduncle and (thus) pendulous spikes. In the pistillate inflorescences, the peduncle is pen- dulous and with pendulous spikes, or it is erect with erect spikes. In the latter case the peduncle and spikes may still be erect when fruiting or they may curve downward when fruiting. The length of peduncle and spikes can, to some extent, be related to pollination (shedding of pollen) and dispersal (the group of ani- mals involved). Staminate inflorescences with long peduncles and/or long spikes can be more easily moved by the wind, promoting the release of pollen. Long and pendulous pistillate inflorescences in fruit are associated with dispersal by bats. Long to very long peduncles, in particular in pistillate inflores- cences, are more common in lowland than in montane species.
The structure of the inflorescence in Cecropia may be explained from a presumably basic type of inflo- rescence of Cecropiaceae as found in the African ge- nus Myrianthus (de Ruiter, 1976). This basic type is cymose and preserved in staminate inflorescences of
some Myrianthus species (Fig. 4). It has a peduncle with about four primary branches at the apex. Further ramification of these branches is usually repeatedly dichotomous. In a more reduced state, as can be found in some Myrianthus species and in many Coussapoa species (Akkermans & Berg, 1982; Berg et al., 1990), the ramification may be(come) dichotomous from the peduncle on. In the basic type of inflorescence, the flowers are concentrated on the distal branchlets (or only on the ultimate branchlets), thus forming branched spikes (or unbranched ones). Solitary flow- ers can be found at the top of the branches, i.e., at the bases of the dichotomies.
One could explain the structure of the inflores- cence of Cecropia by assuming a shortening of the proximal branches of the basic type of inflorescence and the elongation (and reduction of branching) of its ultimate flower-bearing parts. The most common number of spikes (four), more or less distinct pairs of (pistillate) spikes, occasional (dichotomous) ramifi- cation of spikes, and the most common position of solitary flowers can all be regarded as indications for a basically dichotomous ramification.
In many Cecropiaceae the inflorescences are enclosed in the terminal bud cover (formed by the stipules) until anthesis or just prior to anthesis. The development of a spathe fully enclosing the flower- bearing parts created a substitution for the original way of protection. This could be related to continuous rapid growth of the stems, too rapid to allow full de- velopment of inflorescences within the bud covers, and to the need of more or less continuous production of flowers and diaspores. Cecropia hololeuca is the only species in which the inflorescences are still pro- tected in the original way. The independence of the development of inflorescences inside the terminal bud cover allows long peduncles and/or spikes.
SPIKES OF STAMINATE INFLORESCENCES
The rachis of the spike is terete or, more often, more or less angular and mostly triangular in cross section. It is mostly hairy, usually only with short stiff hairs, sometimes also with arachnoid indumentum. The rachis contains mostly tannin, often concentrated in its outer cell layers.
The spikes are mostly pale yellow to whitish at anthesis. However, they are green in Cecropia chlo- rostachya and occasionally dark yellow to orange or even almost scarlet in C. polystachya.
STAMINATE FLOWERS
The flowers are sessile or (shortly) pedicellate. They are free in all species, except for Cecropia pur-
MORPHOLOGY 17
purascens, in which the upper parts of the perianths are connate. In C. heterochroma, the more or less fleshy apices of the perianths are interlocking and close off the slits among the flowers. In the other spe- cies the perianth apices are free from each other. The length of the flowers varies, partly in connection with their position on the rachis. They are relatively short at the apex and the base, and at sides of an angular rachis. The flowers on the angles tend to have longer perianths and have pedicels or longer pedicels than those on the sides. The flowers are 2-merous, very rarely 3-merous. The perianth is tubular, although in some species, as C. marginalis, the tubular part is short. In most species the tepals are almost entirely connate, leaving only a narrow slit-shaped aperture. But in C. marginalis the tepals are partly or alsmost entirely free. The apical part of the perianth is often more or less thickened. It is hollow in several species and often also contains (some) tannin. In contrast to the perianth of the pistillate flower, the apex of the perianth is mostly smooth and the part below the apex is often glabrous, or if hairy, then mostly with stiff short hairs or, if arachnoid indumentum is present, then it is mostly sparse and short. However, in a small number of species (as in those with flowers of the C. sciadophylla type, see below) the arachnoid indu- mentum below the apex is similar to that of the pis- tillate flower.
The staminate flowers of Cecropia show more var- iation than the pistillate flowers. This variation is largely related to the stamens. The anthers are in- trorse, and the filaments are flat or more or less thick- ened. In most species the filaments differ in length in the bud and then elongate one after the other, pushing the anthers through the slit-shaped aperture (Fig. 5. 1). The anthers may remain attached to the filament, but more often they become detached and subsequently reattached to the margins of the aperture of the pern- anth or remain connected with the filament in one way or another. The mechanical aspect of the detachment of the anthers is not quite clear. It can be explained by contraction of the filament after it has pushed the anther through the aperture, which is narrower than the anther. The filaments are usually distinctly shorter than the tube of the perianth after detachment of the anthers.
Several types of staminate flowers are found. But the state of the material makes it often difficult to determine for each species the type of flower, either because the material in a dried state often does not show clearly the features of the stamens and anthers or because the flowers are not in the right state of development. One needs material at the right phases
of development conserved in liquid. Moreover, two types of staminate flower may occur in the same spe- cies, sometimes in the same area, sometimes geo- graphically separated. The following types of stami- nate flowers can be recognized.
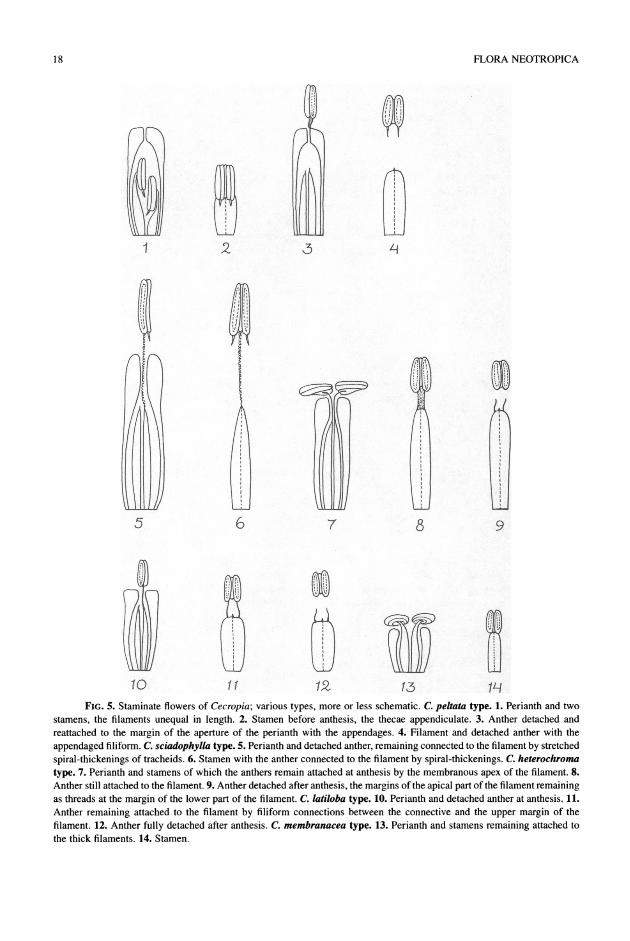
1. Cecropia peltata-type (Fig. 5.1-4). The anthers bear mostly ca. 0.1 mm subulate appendages at the bases on the thecae in between the locules. The an- thers are ca. 0.5-1 mm long. The perianths are tubular and mostly ca. 1-2 mm long, glabrous or sparsely hairy outside below the apex. The apex is plane to slightly convex. The filaments are relatively broad, flat or slightly thickened. The anthers become de- tached from the filament and reattached to the margins of the slit-shaped aperture by the (sticky) tips of the appendages. The appendages become thread-like, and thus the anthers loosely attached to the flower and movable by wind, so that pollen can be shed. The anther which is pushed first through the aperture is often removed by the second anther. This type is found in the majority of the species.
2. Cecropia latiloba-type (Fig. 5.10-12). The an- thers are appendiculate or not. At anthesis, they are detached from the middle of the filament but remain laterally loosely connected to the filament by a pair of filiform connections between the upper margin of the filament and the V-shaped base of the connective of the anther. The perianth is tubular with a plane apex and an aperture that opens somewhat more widely than in the C. peltata type. The perianth is relatively short, ca. 1-1.5 mm long, and bears short stiff hairs outside below the apex. The anthers are 0.6-1.2 mm long. The filaments are often somewhat narrower than those of the C. peltata-type. The anthers are so loosely attached that they can be moved by the wind. After the anther becomes fully detached, the filiform con- nections remain on the upper margin of the filaments. It is not clear how the anther becomes finally detached from the filament. This type of flower is found in C. albicans, C. latiloba, and C. utcubambana.
3. Cecropia sciadophylla-type (Fig. 5.5-6). The anthers are relatively long, 1-2 mm, and are appen- diculate or not. After detachment of the anther they remain (temporarily) loosely attached by stretched spiral thickenings of the tracheids of the (median) vas- cular bundle of the filament. The perianth is often relatively long, 1.5-4 mm, and it bears more or less dense arachnoid indumentum below the apex outside, as typical for the perianth of the pistillate flower. The apex is distinctly convex to plane with an narrow ap- erture. This type of staminate flower is found in C. glaziovii, C. palmata, C. saxicola, and C. sciado- phylla. However, in the former two species, the C.
18 FLORA NEOTROPICA
LI~~~~~~~LL
5 6 8 9
10 12 13 lL4
FIG. 5. Staminate flowers of Cecropia; various types, more or less schematic. C. peltata type. 1. Perianth and two stamens, the filaments unequal in length. 2. Stamen before anthesis, the thecae appendiculate. 3. Anther detached and reattached to the margin of the aperture of the perianth with the appendages. 4. Filament and detached anther with the appendaged filiform. C. sciadophylla type. 5. Perianth and detached anther, remaining connected to the filament by stretched spiral-thickenings of tracheids. 6. Stamen with the anther connected to the filament by spiral-thickenings. C. heterochroma type. 7. Perianth and stamens of which the anthers remain attached at anthesis by the membranous apex of the filament. 8. Anther still attached to the filament. 9. Anther detached after anthesis, the margins of the apical part of the filament remaining as threads at the margin of the lower part of the filament. C. latiloba type. 10. Perianth and detached anther at anthesis. 11. Anther remaining attached to the filament by filiform connections between the connective and the upper margin of the filament. 12. Anther fully detached after anthesis. C. membranacea type. 13. Perianth and stamens remaining attached to the thick filaments. 14. Stamen.
MORPHOLOGY 19
peltata-type of stamens is also found. The type de- scribed is found in thick spikes with relatively long flowers and anthers, the other type in more slender spikes with shorter flowers and anthers.
4. Cecropia heterochroma-type (Fig. 5.7-9). The anthers are pushed through the aperture (which is of- ten slightly wider than in the C. peltata-type) but they remain attached to the filament by its thin (membran- aceous) upper part. The anthers are appendiculate or not. The perianth is of the normal length, with a plane or slightly convex apex, and it is hairy or glabrous below the apex outside. The filaments are thick and before anthesis different in length. After anthesis the filaments fill the space inside the perianth and the apices of the filaments reach or even exceed the ap- erture. The anthers may subsequently remain attached to the filament only by two filiform connections run- ning from the upper margin of the filament to the connective of the anthers, like in flowers of the C. latiloba-type. This type of staminate flower is found in C. angustifolia, C. heterochroma, C. polystachya, and C. tacuna.
5. Cecropia membranacea-type (Fig. 5.13-14). The anthers remain attached to the filament at anthe- sis. The perianth is short, with a wide aperture, and it is glabrous. The anthers are short, 0.5-0.7 mm long, and are not appendiculate. The filaments are relatively narrow, (rather) thick, (almost) equally long before anthesis, and exceed the aperture after anthesis. The spikes are very slender and pendulous and thus are easily moved by wind to get pollen released from the anthers. This type of staminate flower is found in C. marginalis, C. membranacea, and C. strigosa.
One could regard the C. membranacea-type of flower as primitive. However, this is not correlated with primitive characters in vegetative parts. Not only for this type but also for the others, correlations with vegetative characters are not evident. Similarity in the type of staminate flower appears not always to be an indication of relationship.
The partial detachment of anthers, such that they remain loosely attached to the filament by stretched spiral thickenings, was discovered and de- scribed by Allemao (1860). The complete detach- ment of anthers which become secondarily attached to the flower by appendages of the thecae was dis- covered and described by Berg (1977b), who then also regarded the case of attachment of the anthers by spiral thickenings as new and undescribed. How- ever, he misinterpreted the loose attachment of the anthers by filiform connections between filaments and anthers as anthers remaining attached to the up- per margin of the filament by the stretched append- ages of the thecae.
Complete or partial detachment of the anthers, by which they become loosely attached to the flower, are clearly adaptations to wind pollination. In cases where the anthers remain firmly attached to the filaments, as in Cecropia membranacea, the spikes are very slender and pendulous and are easily moved by the wind. In other cases, too, slender and/or long pendulous spikes may promote release of pollen.
The staminate inflorescences (flowers) of many (or all?) species emit a (faint) sweetish smell; for some species, e.g., Cecropia strigosa, it is rather character- istic.
Key to the types of stamen 1. Anthers completely detached from the filaments and reattached to the margins of the aperture of the
perianth by sticky appendages of the anther ................................................................... 1. C. peltata-type 1. Anthers remaining connected to the filaments.
2. Anthers remaining loosely connected to the filaments. 3. Anthers remaining connected by stretched spiral thickenings of tracheids .......... ............ 3. C. sciadophylla-type 3. Anthers remaining connected by two filiform connections between the connective and upper
margin of the filament .................................................................. 2. C. latiloba-type 2. Anthers remaining fully connected to the filaments.
4. Anthers remaining attached by a membranaceous apex of the filament: stamens different in length before; perianth and anther of normal length; apperture of the perianth slit-shaped ..... 4. C. heterochroma-type
4. Anthers remaining attached without a membranaceous apex of the filament: stamens about equal in length before anthesis; perianth and anther shorter than normal; aperture of the perianth wide ........................................................... 5. C. membranacea-type
POLLEN
The pollen is dry. Clouds of pollen are released from the spikes when they are moved or touched. Pol- len of some species has been decribed by Barth
(1974). Cecropia pollen found in sediments play a role in palaeoecological studies revealing changes in vegetation and climate (Wijmstra, 1967; Liu & Col- invaux, 1985, 1988; Bush & Colinvaux, 1988; Col-
20 FLORA NEOTROPICA
invaux et al., 1988a, 1988b; Bush et al., 1990; Col- invaux, 1993; Wijninga, 1996) and in relation to agricultural history (Bush et al., 1989).
SPIKES OF PISTILLATE INFLORESCENCES
The spikes are mostly terete, less commonly more or less angular than in the staminate ones. The rachis is mostly hairy, with only stiff straight hairs, some- times also with (or rarely only with) arachnoid indu- mentum. Tannin is mostly present, often concentrated in the outer cell layers; but in several species, such as many of the Cecropia peltata-group, the outermost layers are free from tannin. The spikes are normally whitish until the spathe drops, but they soon turn green.
PISTILLATE FLOWERS
Most of the variation in the pistillate flowers is presented in Figure 6. The flowers are sessile, mostly free, but in several species of the Cecropia peltata- group, such as C. litoralis and C. peltata, they are or may be basally connate. If so, the stiff hairs com- monly arising from the rachis are situated on junc- tions of the adjacent perianths. The perianth is tubu- lar; the tepals are fused entirely. The length of the perianth is shorter at the base and the apex of the spike. The perianths are usually about 1 mm longer in the fruiting state than at anthesis. The upper part is more or less strongly thickened and the apical part is mostly more or less convex and is often muriculate to punctate or hispidulous. In all species arachnoid indumentum is present on the outside of the perianth, mostly only below the apical part, but in species of the C. peltata-group, usually also on the apex if the stigma is peltate (Fig. 6.4). The arachnoid indumen- tum may occur just below the apex or extend farther downward, sometimes even to near the base. In some species stiff short hairs may also occur below the ap- ical part outside. Arachnoid indumentum is often also found inside or just below the style channel. In many species this indumentum appears to be always pres- ent, and in others it may be sometimes absent or al- ways absent (as in several species of the C. peltata- group). The aperture, which is just large enough to allow the style (or stigma) through, is slit-shaped or (almost) circular. The style channel in the thickened apical part of the perianth varies in length in relation to the length of the apical part. The style also varies in length; long styles are often curved, and they may bear short hairs. The style is attached to the ovary apically or subapically. The stigma varies from pen- icillate to tongue-shaped to peltate. In species (or in- dividuals) of the C. peltata-group with distinctly pel- tate stigmas, the arachnoid indumentum extends to
near the aperture of the perianth. In these species (or individuals) the stigma is already outside the aperture long before anthesis, whereas otherwise the stigma becomes exposed just before anthesis. The presence of arachnoid indumentum on the apical part to near the aperture is apparently correlated with a peltate stigma and a difference in the ontogeny of the flower. In the C. peltata-group one can find transitions from the commonly peltate stigma through the subpeltate to tongue-shaped stigma to the penicillate one, and these are related to extension of arachnoid indumen- tum from below the apical part of the perianth toward the aperture of the perianth (Fig. 6). Tannin is com- mon in the perianth and is often concentrated in the upper thickened part. However, in some species it is scanty or in the species of the C. peltata-group usually absent. Tannin often accumulates in the pericarp, but it can be totally absent, as in C. longipes.
FRUITS AND SEEDS
The fruits are achenes enveloped by somewhat en- larged, at least in the upper part, more or less fleshy perianths, remaining greenish even at full maturity of the infructescence. The fruits are oblongoid, elliptic, (sub)obovoid or (sub)ovoid. The pericarp is more or less tuberculate in most species, but in several it is smooth. The color of the pericarp varies from dark to pale brown. The testa is very thin. The straight em- bryo with a rather long radicle and equal and flat cot- yledons is embedded in endosperm. Under certain conditions the viability of Cecropia seeds can be more than five years. Germination is triggered by full sun- light and changing temperatures (Holthuijzen & Boerboom, 1982; Vaizquez-Yanes & Smith, 1982).
NATURE AND VARIATION OF
MORPHOLOGICAL CHARACTERS
The general picture of morphological differentia- tion in the genus is a mixture of uniformity, as in the general habit and in the pistillate flowers, and of con- siderable variation, as in leaf characters and in the dimensions and numbers of spikes in the inflores- cence. Part of the variation in leaf characters, such as the number and depth of the incisions, the number of lateral veins in the midsegment of the lamina, and their loop-connections before or in the margin, are clearly related to the considerable change from juve- nile to adult features, often without reaching a state of stability in features. The variation in the number of spikes can be related to the basically open inflo- rescence in which the number of branches is not def- inite and may vary with vigor of growth. Whereas some of the variation appears to be related to ontog- eny, variation in other respects-such as the length of
ANATOMY 21
Fi(;. 6. Pistillate flowers of Cecropia, different types with regard to stigmas and the presence of arachnoid indu- mentum. schematic. 1. From C. latiloba. 2. From C. ulei. 3. From C. sciadophY/l1a. 4. From C. peltata.
stipules, the composition of the indumentum, and the absence, presence, and state of the trichilia-is not or is only partly so.
About 50% of the species are morphologically uniform; most of them have more or less small ranges of distribution and more or less narrow ecological am- plitudes. The more or less conspicuous variation in the other species is partly (geographically) clinal; it can be partly related to ecological conditions; or it cannot be linked to either distribution or ecology.
A remarkable type of variation is the co- occurrence of color variants in different proportions and of variants in the length of spikes (in Cecropia obtusifolia).
There are few exceptional characters such as the absence of a well-developed spathe in Cecropia holo- leuca, trichilia enclosed in "pockets" in C. virgusa, connate staminate flowers in C. purpurascens, and distinctly larger and pink Mullerian bodies in C. his- pidissima. There are some characters shared by few species which probably indicate taxonomic affinity, such as long irritating hairs that easily break off in C. garciae, C. hi.spidissima, and C. megastachya, a group of species showing affinities also in other fea- tures. However, the presence of anthers remaining at- tached to the filaments by spiral thickenings is found in the clearly related species C. glaziovii and C. pal- mata, but also in C. putumavonis, C. saxatilis, and C. sciadophvlla, which do not show affinities to each other or to the former set of species.
Hybridization may occur rather frequently (S. T. Trimble, pers. comm.). Hybrids with a clear mixture
of parental characters have not been encountered by the authors. However, individuals showing one (or at most two) features-e.g., of the indumentum, of a co- occurring species-are not uncommon (see, e.g., comments under Cecropia gabrielis and C. metensis). A proof of such results in hybridization was found at the campus of the Universidade Federal de Minas Gerais (Belo Horizonte, Brazil). A single fruiting spike from a specimen of C. polystachya in cultivation in Campinas (Sao Paulo, Brazil) was taken by J. Lom- bardi (BHCB) and from the seeds several plants were raised and four of them were planted. They were adult specimens in March 2001. Three of them show all the features of C. polystachva, but the fourth, a staminate one, almost fully the features of the parental species C. pachystachva. The latter differs from other mate- rial of C. pachystachya (indigenous in Campinas) in the pronounced lobation of the free part of the seg- ments of the lamina retained in the adult state. In this respect, that specimen resembles C. polvstachva. Other characters, such as numbers or dimensions ex- ceeding the "normal" ranges of the species, might also be linked to hybridization.
ANATOMY Comparative anatomical studies on the wood and/
or the leaves, including the genera of the Cecropi- aceae, have been carried out by Renner ( 1907) and by Bonsen & ter Welle (I1983). The wood anatomy of the neotropical genera was treated by ter Welle et al. (1992). The anatomy of the leaves of some Cecropia
22 FLORA NEOTROPICA
species has been described by Richter (1897). The idioblasts in the leaves of Cecropiaceae have been studied by Setoguchi et al. (1993). Members of the family accumulate silicon in contrast, but in other Urt- icales, abundant calcium and silicon.
PROTECTION OF FLOWERS AND INFLORESCENCES
Several features of the flowers and inflorescences probably protect the inflorescences and flowers against penetration and subsequent damage by the lar- vae of insects (cf. Berg, 1990).
Insect larvae are usually present in the spikes of staminate inflorescences after they have shed the spathe, but they are absent in the pistillate spikes of pistillate inflorescences. Andrade (1984a) presented an account of a species of Ophtalmoborus (Curcu- lionidae, the larvae of which are found in staminate inflorescences of C. pachystachya (= C. lyratiloba). These larvae destroy the rachises of the spikes.
Insects (adults and/or larvae) can always penetrate the spikes of staminate inflorescences through the ap- ertures of the perianths and in most species through the slits among the flowers as well. The slits among the flowers can be closed off by fusion of the upper parts of the perianths (in Cecropia purpurascens) or by dense, entangled arachnoid indumentum at the lat- eral surfaces of the perianths as in some other species (e.g., C. palmata and C. sciadophylla). A third way to close slits among the staminate flowers is found in C. heterochroma, in which the apices of the perianth interlock.
The perianths of pistillate flowers always bear long arachnoid indumentum outside, at least below the apex. This indumentum is interwoven with that of adjacent flowers and close slits among the flowers. Penetration through the aperture of the perianth is pre- vented by the small diameter of the aperture, just let- ting through the style. Moreover, short bristles on the style and the arachnoid indumentum at the inner sur- face of the perianth, in or just below the style channel, will contribute to barring access to the interior of the flower. In addition, tannin is usually more abundantly present in the pistillate flowers than in the staminate ones; it is concentrated in the thickened apical part of the perianth and in the wall of the ovary and devel- oping pericarp, and it provides additional chemical protection against herbivory (cf. Coley, 1986). These features probably explain the general absence of in- sect larvae in the spikes of the pistillate inflorescence.
Before anthesis the spikes are not only protected by the spathe, but initially also in the terminal bud.
The duration of this protection depends on the length of stipules and the length of the inflorescence. In Ce- cropia hololeuca, which lacks a well-developed spathe, the protection of the spikes until anthesis is carried out only by the stipules. Protection of inflo- rescences by the terminal bud is a common feature of Cecropiaceae.
PHENOLOGY Most species flower and bear fruit throughout the
year, but individuals often intermittently. In some spe- cies (e.g., Cecropia elongata), flowering and fruiting appears to be season-dependent either throughout or in part of their ranges of distribution, as is found for C. obtusa in populations in southeru Para (Brazil), which bear neither flowers nor fruits at the end of the dry season. According to field notes and data on col- lection labels, C. palmata and C. sciadophylla appear to flower throughout the year, but in French Guiana the fruiting is restricted to the period October-Janu- ary (the end of the dry season and the beginning of the rainy season) (Charles-Dominique, 1986). These data suggest that flowering and fruiting periods vary throughout their ranges. Milton (1991) reported sea- sonal fruiting (December-July) for C. insignis in Pan- ama. Seasonal flowering is clear in C. latiloba and C. membranacea, both inhabitants of seasonally flooded (riverside) vegetation. These species (at least in the middle part of the Amazon basin, e.g., near Manaus), flower when the water level is high and fruit when the water level is going down and river banks become available for seedling establishment.
POLLINATION Several traits of the staminate flowers and inflo-
rescences described and field observations indicate that Cecropia is in general adapted to wind pollina- tion, either by pendulous spikes which can be moved by wind to shed the pollen or by the special adaptation of detachment of anthers and their secondary attach- ment allowing the shedding of pollen by motion of the anthers. The pollen is dry and easily released by movement. It is likely that wind pollination is pre- dominant. Although observations of pollen transport by insects are wanting, small beetles and flies use sta- minate inflorescences as breeding sites (cf. Andrade, 1984a; Wheeler, 1942) and thus may play a small role in pollination. Insects (Epitragus sp. and Ophtalmo- borus sp.) eat pollen grains of C. pachystachya (= C. lyratiloba) but probably do not pollinate (Andrade, 1984a). In the same Cecropia species pistillate flow- ers produce small amounts of nectar (Andrade, 1980,
ORGANISMS OTHER THAN ANTS ASSOCIATED WITH CECROPIA 23
1984c), probably to catch pollen and not to attract insects.
DISPERSAL In many species of Cecropia, full-grown trees can
produce millions of seeds (Brokaw, 1998; Martinez- Ramos & Alvarez-Buylla, 1986). The abundance and the regular presence of fruits permits the genus to play a role as keystone food supply for frugivorous animals (Vepsalainen, 1999). The diaspores are dispersed mainly by animals: various birds, including insectiv- orous ones (Eiseman, 1961; Leck, 1972; Snow & Snow, 1971; Charles-Dominique, 1986; Vepsalainen, 1999) and toucans (Skutch, 1971); fruit bats (Charles- Dominique, 1986; Fleming, 1988; Fleming & Wil- liams, 1990; Lobova et al., 2003); monkeys (Eiseman, 1961; Terborgh, 1983; Stevenson et al., 2000); op- posums (Eiseman, 1961; Medellin, 1994); and fish (Gottsberger, 1978). They create or continuously re- plenish the seed banks.
Pendulous infructescences with long peduncles and spikes are associated with dispersal by bats, and those with short infructescences with dispersal by birds. Species occurring along rivers may be dis- persed by water, because parts of mature infructesc- ences drop from the rachis and float for a while, kept buoyant by the air in the arachnoid indumentum among the perianths (see Kubitzki & Ziburski, 1994).
SURVEY OF MYRMECOPHYTISM IN THE GENUS
For a thorough discussion of ants and Cecropia, see "Cecropia and Its Biotic Defenses" by Davidson (this volume, p. 214).
1. Species usually (or often) and throughout their geographic ranges inhabited by Azteca ants; trichilia with Mullerian bodies present and internodes (usu- ally) with scarce pith: Cecropia angulata*, C. annu- lata, C. concolor, C distachya, C. elongata, C. en- gleriana, C. ficifolia, C. garciae, C. glaziovii, C. goudotiana, C. granvilleana, C. herthae, C. hetero- chroma, C. idroboi*, C. insignis, C. kavanayensis*, C. latiloba, C. litoralis, C. longipes, C. marginalis, C. maxima, C. membranacea (p.p.; see below), C. me- tensis, C. montana, C. multisecta, C. mutisiana, C. obtusa, C. obtusifolia, C. pachystachya, C. palmata, C. pastasana, C. plicata (often), C. polystachya, C. purpurascens, C. putumayonis, C. reticulata, C. sa- rarensis, C. saxatilis*, C. silvae*, C. strigosa, C. sub- integra, C. ulei, C. utcubambana, and C. virgusa (44 species in total). Species with an asterisk (*) were assigned to this category based not on field observa-
tions by the first author but on other evidence, such as label data and perforated prostomata in herbarium material.
2. Species inhabited by Azteca ants, but not throughout their geographic ranges; either trichilia (with Mullerian bodies) absent or internodes with am- ple pith: Cecropia albicans, C. angustifolia, C. pel- tata, and C. telenitida.
3. Species inhabited by other genera of ants; tri- chilia with Mullerian bodies present, internodes with scarce pith: always Cecropia hispidissima and C. me- gastachya, sometimes C. membranacea.
4. Species never inhabited by Azteca ants; trichi- lia absent or poorly developed and/or internodes filled with ample pith: Cecropia andina, C. bullata, C. ga- brielis, C. hololeuca, C. pittieri, C. schreberiana, C. sciadophylla, C. tacuna, and C. telealba.
The absence of ants in Cecropia chlorostachya and the presence in C. velutinella is not certain, because of the few specimens examined. The dense indumen- tum on the leafy twigs and subpersistent stipules in these species may prevent or reduce occupation by ants (cf. Davidson et al., 1991).
In other species, internodes filled with mucilage or internodes with thick walls and smooth leafy twigs with a waxy layer (cf. Federle et al., 1997) may pre- vent (or reduce) occupation by ants (or select species) and attacks by leaf-cutter ants (Schimper, 1888). As a waxy surface of the leafy twig and dense villous indumentum can be found in the same species, also the type of indumentum may deter ants (and other insects).
Specimens of Cecropia species which are nor- mally occupied by Azteca ants may experience the absence of these ants or be inhabited by ants of var- ious other genera (see Wheeler, 1942).
ORGANISMS OTHER THAN ANTS ASSOCIATED WITH CECROPIA Cecropia leaves are favorite food for sloths, ani-
mals which can often be observed in Cecropia trees (as in C. polystachya, Kemper & Satore, 2000).
An extensive list of animals found on Cecropia was presented by Wheeler (1942). Attention has been drawn to association of groups of beetles and species of Cecropia by Andrade (1984a) and Jolivet (1990b). The latter author also drew attention to the only oc- casional presence of hemiparasitic plants (Lorantha- ceae) on Cecropia and the possible role of ants to explain this phenomenon. It is remarkable that the presence of Loranthaceae is quite rare, but if Loran- thaceae are present, then very locally and abundantly.
24 FLORA NEOTROPICA
Hollow internodes often contain scale insects, Coc- cidae (cf. Wheeler, 1942; Davidson, this volume), tended by the ants. The coccids are usually attached to the "ceilings" of the compartments of the stem, and young ones are moved to the next internode, mostly via the holes made in the nodal septa. The inhabiting ants obtain sugary exudates from these insects.
ECOLOGY Cecropia is one of the major pioneer tree genera
in regions of the Neotropics with more or less wet lowland and montane forest. There is a high degree of specialization to certain types of habitats, like mon- tane rather than lowland or non-inundated rather than inundated habitats. Within the major ecologic cate- gories species show more definite (fine-tuned) pref- erences to elevations, duration of inundation, and other conditions. Clear ecological variation is not un- common within species.
The ecological aspect in the differentiation and speciation appears to be more pronounced in Cecro- pia than in the other genera of Cecropiaceae, as well as in those of Moraceae.
Most of the Cecropia species are lowland species occurring at elevations up to 1000(-1300) m. Species that can be described as submontane are found at (700-)1000-1800(-2000) m, and the montane ones in cloud forest at 1600-2600 m.
The elevational amplitudes are narrow and/or well-defined for many species (as Cecropia albicans and C. megastachya) or wide and/or with unclear lim- its for others (as C. angustifolia). Some predomi- nantly lowland species, e.g., C. garciae and C. his- pidissima, extend to (or possibly occur disjunctly in) submontane habitats (see below) and the montane C. maxima is found in submontane habitats.
Most of the species are associated with non- inundated vegetation. Some species, Cecropia lati- loba and C. membranacea, are clearly elements of seasonally inundated riverside vegetation (cf. La- motte, 1990, 1992). These two species of medium- sized trees occur abundantly in riverside habitats with periodic inundation of long duration and are distinct in their requirements: C. membranacea is associated with nutrient-rich and well-aerated habitats and C. la- tiloba with poorer and/or less well-aerated habitats. The former can form extensive stands, the latter oc- curs often solitary or in small groups. Some other species may occur in temporary inundated vegetation, as C. metensis in Mauritia-swamps and C. litoralis (in its Amazonian area of distribution) along black- water rivers.
A few species are normally found on rocks, such
as Cecropia granvilleana on wet rocky slopes of a few inselbergs in French Guiana, C. goudotiana on rocky surfaces in the dry Upper Magdalena valley (Colombia), and C. saxicola in rocky areas in central Brazil and Bolivia. Cecropia idroboi is probably as- sociated with (sandstone) rocky areas in Caqueta and Meta (Colombia).
Several species, e.g., Cecropia distachya, C. her- thae, C. insignis, C. purpurascens, and C. sciado- phylla, are species commonly found in (large) treefall gaps in mature lowland forest (cf. Brokaw, 1986), as well as in other natural or man-made primary clear- ings. These species often become longlived tall trees (some to 30-40 m) and can form part of the forest canopy. These species, which can be described as for- est (gap) species, usually establish abundantly in pri- mary forest clearings (and can form almost pure stands, cf. Oldeman, 1974: tabs. 68-69), but hardly succeed in establishing or regenerating in subsequent (or secondary) clearing, or after use of the clearing for agriculture. In contrast to the next category, this group of species performs poorly, growing slowly, in greenhouse conditions (at Store Milde, Norway), pos- sibly due to absence of mycorrhizal fungi (see Bro- kaw, 1998: 95).
Others, which can be described as more or less pronouncedly "weedy," "capoeira," or "chagra" spe- cies are Cecropia concolor, C. ficifolia (p.p.), C. Ii- toralis (p.p.), C. obtusifolia, C. pachystachya, C. pel- tata, and C. ulei. These are small or medium-sized and more or less shortlived trees, rarely establishing in forest gaps but common and often abundant in sec- ondary growth, abandoned fields, etc.
Many species occur abundantly in the proper hab- itats throughout their ranges of distribution. However, some species, such as Cecropia annulata, C. putu- mayonis, and C. utcubambana, are (far) less common and occur more or less widely dispersed within their ranges of distribution, and, moreover, do not clearly behave as pioneer trees by forming dense stands in secondary growth. They appear to be associated with the forest habitat, establishing in small gaps or in the margins of larger treefall gaps and are apparently shade-tolerant. Cecropia ficifolia behaves similary in part of its range, including the foothills of the Andes. The indicated differences in ecological behavior also became evident in greenhouse experiments with some species of Cecropia, indicated as "gap" and "pioneer" species, under different light regimes and nutrient treatments (Folgarait & Davidson, 1994, 1995).
There appear to be two groups of montane (or sub- montane) species. One comprises species like Ce- cropia maxima, C. tacuna, C. telealba, and C. tele-
DISTRIBUTION 25
nitida, which occur rather evenly in the forest and do not evidently show the traits of pioneer species. They can become tall trees with broad crowns. The other group, as represented by C. andina and C. gabrielis, form usually smaller trees and tend to aggregate in more or less open sites, such as along streams. In general, montane species are not evidently colonizers of landslides, which appears to be in accordance with the results of a study by Guariguata (1990) on forest regeneration on landslides in Puerto Rico.
The categories that can be recognized with regard to ecological behavior can only be circumscribed roughly, as many species take (or can take) more or less intermediate positions. Moreover, they may be- have differently in different parts of their distribution ranges, as in the periphery of their ranges. Several species have (or may have) differentiated into eco- types, ecospecies, or ecological subspecies. This ap- pears to be clear in some members of the Cecropia peltata-group (see below). Cecropia litoralis is a common, "weedy" species of non-inundated lowlands of the coastal region of Ecuador and the adjacent coastal part of Colombia (Narifno). It also occurs in the Amazonian part of Ecuador and adjacent parts of Colombia and Peru. There it is apparently confined to the borders of the (more or less) black-water rivers. Cecropia engleriana is a small to medium-sized tree, not clearly associated with riverine habitats, in the southern part of its range of distribution, Bolivia and southern Peru. However, in part of the upper Amazon basin, along the Rio Napo and the upper Amazon River, this species is found mostly along rivers, in habitats with short periods of inundation, as a medium-sized to quite tall tree.
Differences of abundance of species in the same locality and habitat may be indicative of slight eco- logical preferences. For example, Cecropia poly- stachya and C. strigosa co-occur in the lowlands of eastern Peru, where the former apparently tolerates drier climatic conditions than the other.
In the upper Amazon basin, where up to 10 species of Cecropia can be found in the same area, the spe- cies, or in some cases pairs of species, are ecologi- cally complementary in occupation of the various habitats. This is less clear in the Pacific lowlands.
Most of the morphological differentiation is weakly or not at all related to ecological preferences. A few characters show relations to the ecological cat- egories as indicated above: coriaceous leaves in true lowland forest (canopy) species and in montane spe- cies; absence of the full set of myrmecophilous traits in montane species; dense arachnoid indumentum on the upper surface of the lamina in montane species;
and relatively short and or erect pistillate inflores- cences in lowland forest (canopy) species and mon- tane species. But shape of the lamina cannot be as- sociated with any of the categories. Deeply incised lamina with numerous segments as well as shallowly incised laminas with few segments can be found both in forest species and secondary growth species or in both montane and lowland species.
Ecological equivalents of Cecropia in Africa are the two species of Musanga in the Cecropiaceae (cf. de Ruiter, 1976; Janzen & McKey, 1977). In Asia, species of Macaranga Thouars (Euphorbiaceae) are such equivalents. The genus comprises myrmeco- phytes with remarkable similarities to Cecropia in ad- aptations to and variation in myrmecophytism (see Fiala & Maschwitz, 1992a, 1992b).
DISTRIBUTION Forty to fifty percent of the taxa of Cecropia can
be regarded as montane or submontane Andean, with a distinct concentration of species in the northern part of the Andes, in Colombia and Ecuador. Taking into account the lowland taxa that reach the eastern or western foothills of the Andes, the Andean region can be regarded as the center of species richness and also as the center of speciation. Only approximately 25% of the species occur more or less remotely from the Andean region. The representation of lowland and montane species in various parts of the Neotropics as well as the phytogeographic regions for the genus is presented in Figure 7.
The size of the ranges of distribution of Cecropia species vary considerably. Some lowland species have large ranges-e.g., C. membranacea, C. peltata, C. pachystachya, C. latiloba, and C. sciadophylla-oc- curring in at least two more or less distinct phytoge- ographic subdivisions (see below). Cecropia angus- tifolia is the only montane species with a large range: from the coastal mountain range in Venezuela through the Andes to Bolivia, and in Central America from Panama to Mexico, at elevations of (500-)800-2500 m. The majority of the species have medium-sized to small ranges and are usually confined to a single phy- togeographic subdivision. Some species have very small ranges, e.g., C. pittieri, an endemic of Cocos Island, and C. granvilleana, confined to some (two?) small populations on slopes of granite outcrops (in- selbergs) in French Guiana.
The distribution of a few species is disjunct: Ce- cropia membranacea, occurring in the Amazon basin, the lower Magdalena Valley, the Pacific coastal re- gion; and C. litoralis, common in the coastal region of Ecuador but also occurring along (black-water) riv-
26 FLORA NEOTROPICA
30 ~ ~ ~ ~ ~ ~ ~ ~ ~ ~~-0
2 i0~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~2
10~~~~~~~~~~~~~~.
10 4-~~~~~~~~~~~~~~~~~~~~~~~~~~~
20- 1;0 260 3~~~7 12 - -0
00 0 02000000000 500... .- '00 0 o/ s
FIG. 7. Distribution of Cecropia. Dashed lines show the major boundaries in the distribution. Numbers indicate the number of species per region (boldface for lowland species, italics for montane species).
ers in the Amazon basin. In some other cases, more or less segregated distribution is related to habitat spe- cialization, such as C. granvilleana, on some insel- bergs in French Guiana, C. kavanayensis on isolated table mountains in the Guayana region, C. saxicola in rocky areas in central Brazil and Bolivia, and C. latiloba on relatively nutrient-poor and not well- aerated periodically inundated soils. In a number of montane or submontane species, more or less pro- nounced discontinuities in the ranges of distribution appear to occur in spite of the presence of (presum- ably) suitable habitats in the gaps. For example, in C. telenitida, not recorded from the central part of the eastern slopes of the Andes in Ecuador, C. velutinella, recorded from Pastaza and Morona-Santiago in Ec- uador and from San Martfn in Peru, C. montana, known from Putumayo (Colombia) and adjacent Suc- umbios (Ecuador) and from Morona-Santiago (Ec- uador) southward to Madre de Dios (Peru). An anal- ysis of the species' distribution then recognized, and with the distribution data then known, was presented by Franco Rosselli & Berg (1997).
Some species of Cecropia have been introduced in neotropical regions outside their natural distribu- tions, e.g., C. peltata in the Bermudas and C. poly- stachya in Sao Paulo (Brazil), where they are natu- ralized.
Several species have also been introduced in the Palaeotropics, e.g., Cecropia peltata (and possibly C. pachystachya) in West Africa, Malaysia, and several Pacific islands, where it is (or they are) are natural- ized; in Indonesia, Taiwan, and China, where it is (or they are) probably still confined to botanical gardens.
Cecropia schreberiana was introduced in Mada- gascar, where it is naturalized, and C. obtusifolia in Hawai'i, also naturalized.
Cecropia polystachya is (or was) in cultivation in botanical gardens in South Africa and Singapore.
REPRESENTATION IN THE MAJOR PHYTOGEOGRAPHIC SUBDIVISIONS
1. West Indies. Two species are found: Cecropia peltata in Trinidad and in Jamaica and C. schreberi- ana in the Lesser and Greater Antilles, except for Ja-
DISTRIBUTION 27
maica. The former species has a type which often lacks trichilia in Jamaica and the widespread type with trichilia in Trinidad. In C. schreberiana two in- fraspecific entities can be distinguished: a subspecies in the Lesser Antilles and Puerto Rico, and a more pubescent subspecies in Cuba, Hispaniola, Puerto Rico, and the Virgin Islands.
2. Central America. Eleven species are recorded, of which only Cecropia pittieri is endemic, being con- fined to Cocos Island. Cecropia angustifolia is a mon- tane species with its main area in South America. The other species are lowland elements which also occur in Pacific Coastal region of South America. Cecropia longipes and C. heterochroma have rather small ranges of distribution, from eastern Panama to adja- cent Colombia. Cecropia hispidissima, C. garciae, and C. virgusa range from Ecuador to eastern Pan- ama, and C. obtusifolia and C. insignis from Ecuador to southern Mexico and Honduras, respectively. The most common species in Central America is C. pel- tata, which extends to Jamaica and to the northern part of South America, far into the Magdalena valley, in the Pacific lowlands down to Buenaventura, and through northern Venezuela to Trinidad, Suriname, and the northern fringe of the Amazon basin (Brazil). Cecropia membranacea, one of the most widespread species, occurs in the Amazon basin, the Pacific low- lands, the Cauca valley (up to Valle) and the Lower Magdalena valley, and extends just into eastern Pan- ama.
3. Pacific Coastal lowlands (Colombia-Ecua- dor). In addition to the nine species that also occur in Central America, this region contains Cecropia li- toralis, common in Ecuador and extending to the coastal part of Narifno (Colombia) and, remarkably enough, also known from the Amazonian region. Ce- cropia obtusifolia, already quite variable in Central America, exhibits confusing variation in Colombia. Some of the species, such as C. hispidissima and C. obtusifolia, extend into submontane conditions in the
western slopes of the Andes. On the other hand, some of the species that can be regarded as (sub)montane, like C. reticulata and C. angustifolia, extend into the lowlands, at elevations down to ca. 500 m. The upper elevational limits of the typically lowland species of this region vary from 100 to 1200 m, or sometimes up to 1500 m.
4. Andean region (Venezuela-Bolivia). This re- gion has the most diverse Cecropia flora, but it is still relatively poorly explored, due in part to limited ac- cess to many areas inhabited by these species. Many of these species are known only by a few collections, and ranges of distribution are often not clear. Several
regions, such as in Peru and Bolivia, have hardly been explored.
Most species have small to medium-sized ranges of distribution, mostly confined to either the western or the eastern slopes of the Andes. Cecropia angus- tifolia is an exception. In South America it ranges from the coastal mountain range of Venezuela to Bo- livia, occurring in Colombia and Ecuador on both sides of the Andes, but also in the Sierra de Macarena, and in western Ecuador and in Peru down to eleva- tions of about 500 m. This species is differentiated into a number of regional and ecological forms, dis- cussed under the species.
On the western slopes of the Andes, at elevations of 1700-2400 m, on the slopes at both sides of the upper drier part of the Cauca valley the white-leaved Cecropia telealba is found, and in Ecuador and ad- jacent Colombia (Narinio) the white-leaved C. max- ima. Species at lower elevations on these slopes are C. bullata and C. gabrielis, both from Pichincha (Ec- uador) to Antioquia (Colombia); C. megastachya, in Choc6 and Valle (Colombia); C. plicata, from Antio- quia to Valle; C. reticulata, from Antioquia to El Oro (Ecuador); and C. subintegra, from Valle (Colombia) to Carchi (Ecuador). The total number of montane and submontane species recorded for the western slopes of the Andes is nine.
In the upper Magdalena valley at middle eleva- tions two species are endemic: at low elevations the lithophytic Cecropia goudotiana, and at middle ele- vations C. mutisiana.
In the Planalto area of Antioquia, several montane species are represented, the widespread Cecropia an- gulata, C. gabrielis and C. plicata, both extending from the slopes of the Western Cordillera, the en- demic C. multisecta, and the white-leaved C. teleni- tida, a species from the Central and Eastern Cordil- leras.
On the eastern slopes of the Andes C. telenitida extends from Venezuela to Putumayo (Colombia) and apparently disjunctly from Morona-Santiago (Ecua- dor) to San Martin (Peru) at elevations of 1700-2400. The northern part of the range of C. telenitida largely coincide with that of the "palmatisecta form" of C. angustifolia, and at lower elevations that of C. sar- arensis (of which the southern limit is not known). In southern Colombia (Caqueta) the "palmatisecta form" of C. angustifolia is replaced by the "hachensis form," and two other species reach the northern limit of their distribution: the montane C. andina (extend- ing to northern Peru, at least to San Martin), and the submontane C. pastasana (extending to central Peru, at least to Junin). In the central part of Ecuador, two
28 FLORA NEOTROPICA
other submontane species C. utcubambana and C. ve- lutinella reach their northern limits; C. utcubambana extend to Madre de Dios and C. velutinella to San Martin, but is not found in southern Ecuador. Three montane species appear to be limited to Peru, C. al- bicans, ranging from San Martin at least to Pasco, C. chlorostachya, recently discovered in San Martin, and C. tacuna, ranging at least from Huainuco to Cuzco. In Bolivia, the probably endemic montane species C. elongata is known from Cochabamba, La Paz, and Santa Cruz. In this area C. strigosa is found at ele- vations of ca. 900-1800 and is a submontane element rather than a lowland one as it is in the northern part of its distribution range. The number of montane and submontane species on the eastern slopes of the An- des is 11 (12 if C. strigosa is included).
5. Amazonian region. In this region 24 species are found. Some of them are widespread and extend to other/adjacent regions: Cecropia distachya, C. la- tiloba and C. sciadophylla (extending to [parts of] the Guiana region), and C. membranacea (also occurring west of the Andes). Two species with the main area of distribution elsewhere occur in the peripheral parts of the Amazon basin: C. peltata in the northern fringe and C. pachystachya in the southern fringe. Cecropia litoralis, a common species of the southern part of the Pacific coastal region, is also in the northwestern part of the Amazon basin.
The other species are either (largely) confined to or distinctly concentrated in either the upper Amazon basin or the lower Amazon basin (mostly extending to the eastern part of the Guiana region). Elements of the upper Amazon basin are Cecropia annulata (southern), C. engleriana (southern to northern), C. ficifolia (widespread and down to the middle Amazon basin), C. herthae (northern), C. idroboi (northern), C. marginalis (northern), C. polystachya (southern to central), C. putumayonis (northern), C. strigosa (southern to central), and C. utcubambana (northern to central). The latter two species are submontane in the northern part of the range. Elements of the lower Amazon basin are C. concolor (extending to central Amazonia [Manaus region] and the southwestern part of the Amazon basin), C. obtusa (extending to the Guianas), C. palmata (extending to the eastern part of the Guianas, to northeastern Brazil, to central Ama- zonia [Manaus region], and to Amazonian Bolivia), Guianas, C. sylvae (with a small distribution range, extending to French Guiana). Cecropia purpurascens and C. ulei occupy more or less restricted areas in the central part of the Amazon basin.
6. Guiana region-Venezuela (Rio Orinoco) to Brazil (Amapa). Some predominantly Amazonian
species extend to this region: Cecropia distachya, C. latiloba, C. obtusa, C. palmata, C. sciadophylla, and C. silvae, and, moreover, C. peltata extending from northern Venezuela. The species confined to the Gui- ana region are C. granvilleana (confined to slopes of granite outcrops, inselbergs, in French Guiana), C. ka- vanayensis (montane in eastern Venezuela and the ad- jacent parts of Guyana and Brazil [Roraima]), and C. angulata (also in the eastern part of the region).
7. South America, South of the Amazon Basin. Five species occur in the region. The most widespread species is Cecropia pachystachya (eastern and central Brazil, northern Argentina and Paraguay, a member of the C. peltata-group). Cecropia saxicola occurs in central Brazil and extends to eastern Bolivia; it is usu- ally found in rocky areas. Two species are confined to more humid areas in eastern Brazil: C. glaziovii (closely related to C. palmata) and the white-leaved C. hololeuca (probably related to an Andean group of montane species). The Amazonian C. palmata ex- tends south of the Amazon basin to northeastern Bra- zil.
SYSTEMATIC POSITION Cecropia is one of the six genera which consti-
tuted the tribe Conocephaleae in the classification presented by Trecul (1847) and the subfamily Cono- cephaloideae in the Moraceae in that presented by En- gler (1889). The other genera included in these taxa were the Asian genus Conocephalus Blume, the Af- rican genera Musanga with 2 species and Myrianthus with 7 species (see de Ruiter, 1976), and the neotrop- ical genera Coussapoa with 48 hemiepiphytic species, Pourouma with 27 species of terrestrial trees (see Berg et al., 1990; Berg & Franco Rosselli, 1993, 1996). Chew Wee-Lek (1963) included Conocephalus in Poikilospermum Miquel, a genus until then treated in the Urticaceae. In his revision of Poikilospermum 20 species were recognized. The study by Chew Wee- Lek raised questions about the position of the six gen- era. Chew Wee-Lek suggested transferring the mi- crospermous genera to the Urticaceae and keep the macrospermous genera Myranthus and Pourouma in the Moraceae. Corner (1962) suggested transferring all six genera to the Urticaceae while Berg (1978a) placed them in a separate family, the Cecropiaceae. Berg's concept was accepted, e.g., by Cronquist (1981). Others (e.g., Judd et al., 1989) prefer to regard the group of six genera as a subdivision of a broadly construed family Urticaceae, also including the Mor- aceae; and Romaniuc-Neto (1999) reduced the group again to a subfamily of the Moraceae, but with the name Cecropioideae. The "technical" characters (to
TAXONOMIC DIFFERENTIATION 29
key the three families out) are few and not strong: milky sap (Moraceae), urticaceous stamens (Urtica- ceae and Moraceae p.p.), and basal ovules (Urticaceae and Cecropiaceae) versus apical ovules (Moraceae). However, the three families are clearly distinct in overall morphological (and ecological) diversifica- tion. The Cecropiaceae always have adventitious aer- ial roots, becoming stilt-roots in most genera or hold fasts for root-climbers (Poikilospermum), or adapta- tions to the hemiepiphytic life form (Coussapoa). The plants contain mucilaginous sap that turns black on exposure to the air. The stipules are fully amplexicaul (except in some species of Poikilospermum) and con- nate, often large and protecting developing inflores- cences. The plants are always dioecious (unlike Mor- aceae and Urticaceae), and they are microspermous or macrospermous (as in Moraceae). Pearl glands are common and the cystoliths accumulate silicon instead of calcium (Setoguchi et al., 1993).
Poikilospermum is morphologically and anatomi- cally clearly different from the other five genera (Berg, 1989; Renner, 1907). The two macrospermous genera, Pourouma and Myrianthus, constitute a pair of genera with clear morphological and ecological similarities. The same applies to Cecropia and Mu- sanga, but the differences between the inflorescences make clear that Musanga cannot be regarded as "a Cecropia without ants" (Janzen & McKey, 1977), but they are probably sister genera. Whereas Musanga is represented by only two species, a lowland and a montane one, speciation based on both morphological and ecological differentiation has been considerable in Cecropia.
TAXONOMIC DIFFERENTIATION The only attempt to establish an infrageneric clas-
sification was made by Snethlage (1923) to accom- modate the 42 species he recognized. It is based on the differences in indumentum on perianths of sta- minate flowers, the length of filaments, and the length of spikes of staminate and pistillate inflorescences.
I. Section Tomentosae. Perianths of the stami- nate flowers covered with white felted or arachnoid indumentum. A. Aequales. Filaments equally long. With
Cecropia concolor and C. palmata. B. Subequales. Filaments unequally long.
With Cecropia glaziovii, C. hololeuca, C. saxatilis, and C. sciadophylla.
C. Arachnoideae. Perianth with long arach- noid indumentum. With only Cecropia maxima.
II. Section Atomentosae. Perianths of the sta- minate flowers with short hairs or glabrous. Filaments unequally long, but in Polystachyae about equally long. D. Centrales. Staminate spikes medium-
sized, relatively thick. With five species: Cecropia litoralis, three species presently in the synonymy of C. peltata, and C. ulei.
E. Angulatae. Staminate spikes angular. With only Cecropia angulata.
F. Elongatae. Staminate and pistillate spikes very long, to 40 cm or more. With Cecropia elongata and C. mexicana (= C. obtusi- folia).
G. Abbreviatae. Spikes usually very short, or if medium-sized, then "elegant." With 16 species, including Cecropia angustifolia, C. ficifolia, C. obtusa, C. peltata, and C. pittieri.
H. Polystachyae. Staminate spikes very nu- merous. Pistillate spikes short and thick. Stamens short, almost equally long. With four species, including Cecropia multifiora (= C. strigosa).
Five species, including Cecropia montana, could not be included in one of these unranked groups.
This classification was strongly influenced by the small number of collections available at that time. The differentiating characters chosen cannot or can hardly be connected to differences in other parts. The pro- posed subdivision cannot be used effectively.
The morphological differentiation, as presently known, does not allow the establishment of a satis- factory subdivision of the genus. Therefore, the spe- cies are presented in alphabetical order in this mon- ograph. There are some morphologically isolated species, some groups of two or three species showing clear affinities, some larger groups showing more vague or rather clear affinities. The species with a more or less isolated position due to the absence of characteristic generic features are Cecropia hololeuca and C. sciadophylla. The latter is distinct by the ab- sence of trichilia, which might not be secondarily lost as in most other cases and can neither be related to occurrence on islands and in montane habitats. Ce- cropia hololeuca is distinct primarily by the absence of trichilia. It also lacks well-developed spathes. The absence of well-developed spathes might be second- ary. Cecropia hololeuca shows in its habit and its white laminas similarities to a group of montane An- dean species, which may also lack trichilia. The sets of two or three clearly related species comprise, e.g., C. glaziovii and C. palmata; C. obtusifolia and C.
30 FLORA NEOTROPICA
subintegra; C. angustifolia and C. montana; C. lati- loba and C. utcubambana; C. garciae, C. hispidis- sima, and C. megastachya; and C. membranacea, C. polystachya, and C. strigosa. The two larger groups of species with more or less clear affinities are the C. peltata-group and the C. telenitida-group.
Cecropia peltata-group. This group is character- ized by the presence of peltate stigmas. In contrast to the more common comose to penicillate stigmas, the peltate stigma is already outside the aperture of the perianth at an early stage of development of the flower, and as a consequence, the arachnoid indumen- tum of the perianth covers not only the perianth below the apex (as commonly), but also on the apex to near the aperture. However, this feature is not present in C. granvilleana, nor is it in all material ranked under C. sararensis and C. schreberiana. In the latter two species the stigma varies from peltate and subpeltate to penicillate-comose in combination with the pres- ence of arachnoid indumentum varying from present on the apex of the perianth to only below the apex. In the C. peltata-group the trees are usually small to medium-sized, rarely tall (as sometimes in C. engler- iana). The stems have prominent scars of the stipules. Tannin is absent or scarce in the perianths of the pis- tillate flower and arachnoid indumentum is mostly ab- sent in the style channel.
The morphologically most distinct taxa of this groups are Cecropia engleriana, C. granvilleana, and C. schreberiana. Cecropia engleriana is distinct by the large number of lateral veins in the free part of the midsegment of the lamina, normally at least 24 pairs, rarely about 21 pairs, whereas there are between 8 and 20, or rarely up to 24 pairs, in all other taxa. Cecropia granvilleana is distinct by the small number of lateral veins in the free part of the midsegment of the lamina (9-10 pairs, whereas in the other taxa nor- mally more than 10) and the consistent absence of peltate stigmata. Cecropia schreberiana is distinct in the remarkable absence of trichilia (or the occasional presence of poorly developed ones). Trichilia are present in all other species of the group. However, they are absent in part of the collections of C. peltata made in Jamaica (or also in part of the individuals introduced in the Old World). In C. schreberiana, the intemodes contain ample brown pith, this being white and scarce in the other taxa. The indumentum on the leafy twigs and the base of the petiole can be subvil- lous. In these three aberrant features, C. schreberiana resembles a number of Andean montane species. Within C. schreberiana two morphological entities can be distinguished (in the present treatment recog- nized as subspecies): subsp. schreberiana glabrous or
sparsely hairy on the leafy twigs and veins on the lamina beneath and penicillate-comose stigmata, and subsp. antillarum more hairy, at least at the venation of the lamina beneath and transitions from peltate to penicillate stigmas, therefore, becoming in general as- pect quite similar to the neighboring C. peltata. Sim- ilar variation with regard to stigma and indumentum appears to occur in the submontane C. sararensis. The northern glabrous form of C. sararensis can be readily distinguished from the other entities of the C. peltata- group (C. concolor, C. litoralis, C. metensis, C. pel- tata, and C. pachystachya), but the southern hairy form less easily.
The Amazonian Cecropia concolor can be distin- guished by the deeply incised lamina, but transitions to the leaf shape of C. pachystachya occur in the southern part of the Amazon basin. Cecropia litoralis and C. sararensis are distinct from C. metensis, C. pachystachya, and C. peltata by the relatively long spikes, in particular in the pistillate inflorescences. Cecropia metensis is very close to C. pachystachya, and essentially not distinguishable from types of the latter species with dense arachnoid indumentum on the petiole and other plant parts. The long whitish hairs in the trichilia characteristic of C. pachystachya and C. metensis are sometimes found in areas where the ranges of C. peltata and C. metensis meet (or over- lap?).
The two taxa with widest distribution, Cecropia peltata and C. pachystachya, cannot be distinguished satisfactorily from each other. Differences in the var- iation of the length of the stipules, the presence/ab- sence of lobation of the lamina segments, and the presence/absence of long white hairs in the trichilia can be used to determine the species morphologically.
Lobation of leaf segments, at least in juvenile stages, is found in Cecropia pachystachya, C. meten- sis, and C. concolor.
The taxa of the Cecropia peltata-group show over- lap in distribution only in some cases: C. concolor and C. pachystachya in the southern Amazon basin, C. concolor and C. engleriana in the southwestern part of the Amazon basin, and C. engleriana and C. litoralis in the northwestern part of the Amazon basin. Slight overlap of distribution ranges may occur for C. metensis and C. peltata, as well as for C. metensis and C. sararensis. But in all those cases intermediates are absent or rare.
In general, the taxa of the Cecropia peltata-group can be regarded as belonging to the "weedy" group of Cecropia species. Cecropia granvilleana is pro- nouncedly adapted to wet granite rock surfaces. Ce- cropia metensis and C. pachystachya are largely com-
VERNACULAR NAMES 31
ponents of savanna, cerrado, and restinga vegetation. Other species show variation with regard to ecological requirements. Cecropia engleriana and C. litoralis are in parts of their ranges of distribution upland spe- cies but in other parts more or less clearly associated with temporarily inundated habitats. Cecropia peltata is found both in regions with wet rain forest and in dry types of vegetation, as some of the inter-Andean valleys in Colombia.
The distribution patterns and ecological differen- tiation suggest segregation of a very widespread en- tity, resulting in various degrees of morphological dis- tinctiveness. The taxonomy seems to be expressed most adequately by treating the nine taxa as species, rather than to unite some of them fully, reduce others to subspecies, and maintain the rest as species. For the Cecropia peltata-group a separate key is con- structed below.
Cecropia telenitida-group. It comprises species of which the leafy twigs and the upper leaf surface are glabrous (or almost so) and the outer surface of the stipules and spathes are either glabrous or (partly) villous with long, more or less soft, whitish hairs. In many representatives of this group the upper leaf sur- face is often or usually covered with more or less dense arachnoid indumentum. In this group of species trichilia can be always absent, occur only in more or less reduced states (mostly without forming Miullerian bodies), or occur in states of being absent through being more or less reduced to occurring in two patches to well-developed and fused. In some of the species the different states can even be found on the same tree. Most of these species are not inhabited by Azteca ants. The internodes may contain ample brown pith in the internodes. The number of spikes of the pistillate inflorescences may be reduced to one or two. The group is distinctly concentrated in the northern Andean region and is associated with montane to sub- montane habitats. It comprises C. albicans, C. bul- lata, C. gabrielis, C. maxima, C. telealba, and C. te- lenitida. Cecropia hololeuca (from eastern Brazil), with its white laminas and pistillate inflorescences with one or two spikes, could be a member of this group. Cecropia schreberiana (included in the C. peltata-group) could be linked to this group, because of the presence of ample pith and absence of trichilia. The montane species C. multisecta and C. plicata, as well as the lowland species C. insignis, could be linked to the C. telenitida-group.
A more vaguely definable group, largely associ- ated with the Guayana region and with relatively few and often broad lamina segments as a common fea- ture, comprises Cecropia angulata, C. ficifolia, C.
distachya, C. kavanayensis, C. obtusa, and C. pur- purascens. It might also include C. saxatilis, which is, however, clearly distinct in the staminate flowers.
USES The use of the genus is limited to largely local uses
(cf. Duarte, 1959; Velasquez, 1971; Galiano-Sanchez, 1976; Garcia-Barriga, 1974). Decoctions of leaves are made to stimulate the cardiac system, to treat asthma and pneumonia, to treat diabetes, and as diuretic. Powder of leaves is used to control Parkinson's dis- ease. Extract of roots are used to heal wounds or ec- zema. Plasters of leaves are made to reduce swellings. Ash of leaves (of various Cecropia species, in partic- ular those C. sciadophylla) is mixed with leaves of coca (Erythroxylum spp.) to reduce acidity when these leaves are chewed (cf. Plowman, 1981, 1984). Branches are used to make percussion instruments (in Nariino, Colombia) or flutes. Cortex fibers are used for several purposes, such as ropes, bow strings, and hammocks. Leaves of some species are used as cattle fodder, to clean utensils, or to polish furniture. The spikes ("bananitas") are eaten or locally sold on mar- kets (e.g., in Santa Cruz, Bolivia). Wood is used to make cheap caskets or matches, or to make rafts. Sev- eral attempts to use the wood for the production of paper pulp have not been successful, partly because of problems with the regeneration of the raw material. Pith of the petioles is used to make ornamental head- gear (in Napo, Ecuador).
VERNACULAR NAMES Guarumo is the commonly used vernacular name
in Spanish-speaking countries in Central America, Mexico, the Caribbean, Colombia, and Ecuador. The variant guarumbo is encountered in several Central American countries, yagrumo in Venezuela, and yu- arumo in Colombia and Ecuador. In the Amazonian part of Peru, setico (or cetico) is the common vernac- ular name, but in the Andean part of this country ta- cuna (or tacona) is in common use. Imbaiiba (also embaliba or umbauiba) is the common name used in Brazil, in particular in the Amazonian part of the country, and ambaiba (or ambaiba) is more common in the southern part of the country, as well as in Par- aguay. Ambaibo is the common vernacular name in Bolivia. Bois canon is commonly used in the French- speaking countries of the West Indies and in French Guiana, whereas trlimpet tree is the most common vernacular name in the English-speaking countries. The name snake wood, used in Jamaica, is connected to the scientific name, which is derived from "Ce-
32 FLORA NEOTROPICA
crops" (or "Kekrops"), a Greek mythological creature, being half snake and half human. The less commonly used vernacular names, the common ones with adjec- tives, and unusual variants are listed under the spe- cies.
CONSERVATION The majority of the species are not endangered.
Many of them temporarily increase in number by the destruction of forest, but continuous clearing leads to reduction of numbers, first of the "forest" species, and finally also of the "weedy" species. However, a few species that are restricted to rocky places (as Cecropia goudotiana), have scattered occurrence, and/or pos- sess small ranges of distribution (as Cecropia multi- secta) can easily become extinct. Second in vulnera- bility are the "small gap" species such as C. putumayonis and C. utcubambana.
Tree, dioecious, terrestrial, usually with stilt-roots; intemodes (mostly) hollow; sap watery, turning black at exposure to the air. Leaves in spirals; lamina sim- ple, peltate, radially veined and incised; petiole usu- ally with one or two patches of dense indumentum of various trichomes (trichilia) at the base; stipules large, connate, fully amplexicaul, leaving circular scars. Inflorescences axillary, pedunculate digitate clusters of spikes, sometimes 1 spike, (usually) en- closed by a spathe until anthesis. Flowers unisexual, sessile (or short-pedicellate); tepals 2, rarely 3, mostly entirely connate, forming a tubular perianth with a narrow aperture; stamens 2, rarely 3, free; ovary free, stigma penicillate to peltate. Fruit a small achene, of- ten tuberculate, fully enclosed by a fleshy perianth; seed with endosperm; embryo small, with flat coty- ledons.
KEYS TO THE SPECIES
A general key to the species would contain too many weak entries because of dioecy and the varia- tion of characters that can be used to key out the spe- cies. Therefore, only keys for countries or larger regions have been constructed, as well as a key to the species of the C. peltata-group, but nonetheless, weak points remain in most of the keys.
Key to the species of Central America, Mexico, and the West Indies 1. Trichilia absent (or poorly developed, in two patches with or without Mullerian bodies).
2. Intemodes with ample brown pith; Cuba, Hispaniola, Puerto Rico, and Lesser Antilles ........... 50. C. schreberiana 2. Internodes with sparse whitish pith.
3. Incisions of the lamina down to 2/10-3/10 from the margin; Cocos Island . ........................ 42. C. pittieri 3. Incisions of the lamina down to 5/10-7/10 from the margin; Jamaica . . 41. C. peltata
1. Trichilia present. 4. Trichilia separate and enclosed in 2 pockets with slit- shaped apertures at the base of the petiole;
Panama ...................................................................... 61. C. virgusa 4. Trichilia fused, superficial.
5. Lamina without arachnoid indumentum in the areoles beneath (at most initially with arachnoid indumentum on the margin and/or main veins). 6. Lateral veins in the free part of the midsegment (10-)15-25(-50) pairs; Mexico to Panama
............................................................................................................................................... .37. C. obtusifolia 6. Lateral veins in the free part of the midsegment 10-13 pairs.
7. Lamina segments 6-8; spikes of the staminate inflorescence 4-5; spikes of the pistillate inflorescence (1-)2-4, 3-10 cm long at anthesis, to 15 cm long in fruit; Panama ................................................................ 19. C. heterochroma
7. Lamina segments (7-)8-10; spikes of the staminate inflorescence ca. 30-40; spikes of the pistillate inflorescence 4, 10-16 cm long at anthesis, to 30 cm long in fruit; Panama ................................................................... 31. C. membranacea
5. Lamina with arachnoid indumentum at least in the areoles beneath. 8. Lateral veins in the free part of the midsegment 25-50 pairs.
SYSTEMATIC TREATMENT 33
9. Leafy twigs and petioles villous to hirsute or hirtellous to strigose, but without irritating bristles; Mexico to Panama ............................................................... 4. C. angustifolia
9. Leafy twigs and petioles partly setose with irritating hairs. 10. Trichilia with bristles among the brown pluricellular trichomes, the Millerian
bodies pink, ca. 3 mm long; lamina with (9-)10-13(-15) segments, smooth above; Panama ......................................................... 20. C. hispidissima
10. Trichilia with short brown or long white soft hairs among the brown pluricellular trichomes, the Mullerian bodies white, ca. 1 mm long; lamina with 14-19 segments, usually scabridulous above; Panama .................................................................. 14. C. garciae
8. Lateral veins in the free part of the midsegment to ca. 25 pairs. 11. Lateral veins in the free part of the midsegment 5-8 pairs; spikes of the staminate
inflorescence 30-80; peduncle of the pistillate inflorescence 30-65 cm long; Panama ........................................................................................................................................... .27. C. longipes
11. Lateral veins in the free part of the midsegment (8-)10-25 pairs; spikes of the staminate inflorescence to ca. 25; peduncle of the pistillate inflorescence to 15 cm long. 12. Lamina usually with 7-10 segments and/or free part of the midsegment with up to 15
pairs of lateral veins. 13. Stipules usually to 10 cm long; Mexico to Panama, Jamaica, and Trinidad ..... 41. C. peltata 13. Stipules 10-50 cm long.
14. Stipules 10-15(-25) cm long; spikes of the inflorescences sessile; Panama ................................................................................................................. 47. C. reticulata
14. Stipules (15-)20-50 cm long; spikes of the inflorescences with stipes; Honduras to Panama ................................................... 23. C. insignis
12. Lamina usually with 10-13 segments and the free part of the midsegment usually with 15-25 pairs of lateral veins. 15. Indumentum of leafy twigs and petioles (partly) brownish; lamina (almost)
smooth above; spathes of the inflorescences densely hairy outside; Mexico to Panama ......................................................... 4. C. angustifolia
15. Indumentum of leafy twigs and petiole whitish; lamina ? scabrous above; spathes of the inflorescences ? sparsely hairy outside; Mexico to Panama
....................................................................................................................... 3 7 C . o b tu sifo lia
Key to the species of Colombia
1. Trichilia enclosed in 2 pockets with slit-shaped apertures at the base of the petiole; Pacific region ... 61. C. virgusa 1. Trichilia exposed (superficial) or absent.
2. Trichilia absent. 3. Incisions of the lamina down to the petiole, the segments often petiolulate; Amazonian region
.................................................................................................................................................. .51. C. sciadophylla 3. Incisions of the lamina not down to the petiole; Andean region or Pacific region.
4. Indumentum partly setose with irritating hairs; Pacific region ......................................... 20. C. hispidissima 4. Indumentum not partly setose, without irritating hairs; Andean region.
5. Lamina ? scabrous above; E Cordillera ............................................................... 2. C. andina 5. Lamina smooth above; W Cordillera ......................... ...................................... 13. C. gabrielis
2. Trichilia present. 6. Trichilia separate (in two lateral patches).
7. Lamina with ? dense arachnoid indumentum above. 8. Lateral veins in the free part of the midsegment 7-16 pairs; arachnoid indumentum in two
distinct layers on the lamina beneath; E and central Cordillera ....................................... 57. C. telenitida 8. Lateral veins in the free part of the midsegment ca. 15-25 pairs; arachnoid indumentum not
in distinct layers on the lamina beneath; W Cordillera ...................................................... 56. C. telealba 7. Lamina without arachnoid indumentum above.
9. Lamina chartaceous to subcoriaceous; spikes of the staminate inflorescence ca. (20-)30-50; spikes of the pistillate inflorescence 4(-7); Amazonian region ...................................... 28. C. marginalis
9. Lamina (sub)coriaceous; spikes of the staminate inflorescence 5-9; spikes of the pistillate inflorescence 1-3(-4); Andean region. 10. Leafy twigs hairy; arachnoid indumentum (almost) absent or in minute patches
surrounded with thick veinlets on lower surface of the lamina .................. ................. 43. C. plicata
34 FLORA NEOTROPICA
10. Leafy twigs glabrous; arachnoid indumentum in the areoles or also on veins on the lower surface of the lam ina ................................................................... 13. C. gabrielis
6. Trichilia fused. 11. Lamina (usually) with 10-20(-24) segments; lateral veins in the free part of the midsegment
mostly >25 pairs .............................................................. 48. C. sararensis 12. Setose (irritating) hairs present on young parts; Pacific region.
13. Trichilia not well-defined; Mullerian bodies pink, ca. 3 mm long .......... ....... 20. C. hispidissima 13. Trichilia well-defined; Mullerian bodies white, ca. 1 mm long.
14. Lateral veins in the free part of the midsegment ca. 35-50 pairs; lamina usually scabridulous above .................................................. 14. C. garciae
14. Lateral veins in the free part of the midsegment ca. 20-30 pairs; lamina smooth above ............................................................ 30. C. megastachya
16. Incisions of the lamina down to the petiole; Antioquia, 1300-1600 m ...... 34. C. multisecta 16. Incisions of the lamina not down to the petiole.
17. Lamina often white above, segments of young leaves patent; spikes of staminate inflorescence 4-6(-8); spikes of pistillate inflorescence (1-)2-3, the spathe 10-20 cm long ..................... ................................ 29. C. maxima
17. Lamina green above, segments of young leaves deflexed; spikes of staminate inflorescence 10-25(-30); spikes of pistillate inflorescence 3-8 (-12), the spathe 3-6(-8) cm long ...................................................... 4. C. angustifolia
15. Lamina ? scabrous above. 18. Incisions of the lamina down to the petiole or 1-2 cm from the petiole; stipules
20-30 cm long; Amazonian region ..................................................... 18. C. herthae 18. Incisions of the lamina not down the petiole, or if 1-2 cm from the petiole,
then the stipules <20 cm long. 19. Lateral veins in the free part of the midsegment usually 30-50 pairs;
spathes and stipules with dense arachnoid indumentum outside; E Cordillera to Amazonian region .................................................... 33. C. montana
19. Lateral veins in the free part usually 10-25 pairs, or if to 35(-43) pairs, then the spathes and stipules without dense arachnoid indumentum outside. 20. Spathes of the inflorescences sparsely hairy to subglabrous outside;
Pacific region ................................................. 37. C. obtusifolia 20. Spathes ? densely hairy outside; Andean region.
21. Lateral veins in the free part of the midsegment 10-21 pairs; pistillate inflorescences soon ? pendulous. 22. Upper margin of the rim of the trichilium lobate; Upper
Magdalena valley .................. ........................ 35. C. mutisiana 22. Upper rim of the trichilium entire; W Cordillera ........ 48. C. sararensis
21. Lateral veins in the free part of the midsegment often >21; pistillate inflorescences erect; widespread .......................... 4. C. angustifolia
1. Lamina with (5-)7-10(-12) segments; lateral veins in the free part of the midsegment mostly <25 pairs. 23. Lamina without arachnoid indumentum in the areoles beneath.
24. Lamina coriaceous, often ? plicate; Andean region, 1000-2300 m ....................... 43. C. plicata 24. Lamina chartaceous to subcoriaceous; plane; Amazonian and/or Pacific region.
25. Base of the lamima entire or shallowly lobed, the upper part 3-lobed. 26. Spikes of the staminate inflorescence 4-5; peduncle of the pistillate
inflorescence 25-45 cm long; Amazonian region ........................... 46. C. putumayonis 26. Spikes of the staminate inflorescence 7-12; peduncle of the pistillate
inflorescence 3-24 cm long; Pacific region ........................................ 54. C. subintegra 25. Base of the lamina ? distinctly incised.
27. Lateral veins in the free part of the midsegment usually 15-25(-40) pairs; Pacific region .............................................. 37. C. obtusifolia
27. Lateral veins in the free part of the midsegment 9-16 pairs. 28. Inflorescences (or at least the spikes) pendulous at anthesis; spikes of
the staminate inflorescence ca. 30-40; spikes of the pistillate
SYSTEMATIC TREATMENT 35
inflorescence 10-16 cm long at anthesis (in fruit to 30 cm long); Amazonian region and Pacific region, usually riverine ......... 31. C. membranacea
28. Inflorescences erect at anthesis; spikes of the staminate inflorescence 4-25(-30); spikes of the pistillate inflorescence 3-10 cm long at anthesis (in fruit to 15 cm long). 29. Lamina with 6-8 segments, the incisions down to 3/10-5/10
from the margin; Pacific region .................................. 19. C. heterochroma 29. Lamina with (8-)10-14 segments, the incisions down to 7/10-
8/10 from the margin; Andean region ............... ................. 4. C. angustifolia 23. Lamina with arachnoid indumentum in the areoles beneath.
30. Lateral veins of the free part of the midsegment marginally loop-connected. 31. Lateral veins in the free part of the midsegment 24-32 pairs; Amazonian region
..................................................................................................................... II. C. engleriana 31. Lateral veins in the free part of the midsegment 8-24 pairs.
32. Lamina (sub)coriaceous, often smooth above. 33. Lateral veins in the free part of the midsegment 8-10(-12) pairs;
Amazonian region ............................................. 22. C. idroboi 33. Lateral veins in the free part of the midsegment 12-24 (or more)
pairs. 34. Lateral veins in the free part of the midsegment usually 12-18
pairs; lamina smooth above; stigma comose; Amazonian region ................................................................................................. 9. C. distachya
34. Lateral veins in the free part of the midsegment > 18 (to 24 or more) pairs, or if < 18 pairs, then the lamina scabrous above and the stigma peltate; not Amazonian. 35. Spathe of the pistillate inflorescence 8-15 cm long;
peduncle of the staminate inflorescence usually >8 cm long and the spathe ca. 10-20 cm long; E Cordillera (E slopes, 300-1400 m) ...................................... 48. C. sararensis
35. Spathe of the pistillate inflorescence 3-6(-8) cm long; peduncle of the staminate inflorescence usually <8 cm long and the spathe usually 4-10 cm long; Andean region, widespread, mostly >1500 m ....................... .............. 4. C. angustifolia
32. Lamina chartaceous to subcoriaceous, mostly ? scabrous above. 36. Trichilia with white hairs distinctly longer than the brown
indumentum. 37. Petiole with dense arachnoid indumentum; spikes of the
staminate inflorescence 4-20; peduncle of the pistillate inflorescence 3-17 cm long; llanos region ............................. 32. C. metensis
37. Petiole without arachnoid indumentum; spikes of the staminate inflorescence ca. 30-80; peduncle of the pistillate inflorescence ca. 30-90 cm long; Pacific region .......................................... 27. C. longipes
36. Trichilia with white hairs about as long as the brown indumentum. 38. Stipules and spathes densely hairy outside; inflorescences erect;
lateral veins in the free part of the midsegment often >20 pairs; Andes, mostly >1500 m ......................... ................ 4. C. angustifolia
38. Stipules and spathes ? sparsely hairy outside; inflorescences with at least the spikes pendulous or the peduncle becoming deflexed; lateral veins in the free part of the midsegment 12-19 pairs; lowlands. 39. Lamina attached to the petiole at ca. 1/4 of the total length
from its base; spikes of the staminate inflorescence usually ca. 15-20, 14-20 cm long; stigma penicillate; Amazonian region, inundated places ..................................... 25. C. latiloba
39. Lamina attached to the petiole at clearly >1/4 of the total length from its base; spikes of the staminate inflorescence usually ca. 5-15 and 5-12 cm long; stigma peltate; Pacific region, in non-inundated and inundated places, and Amazonian region, in inundated places ........................... 26. C. litoralis
36 FLORA NEOTROPICA
30. Lateral veins of the midsegment loop-connected before the margin. 40. Trichilia on a subscrotiform (abaxially bulging) base of the petiole.
41. Trichilia laterally extended by lobes; spikes of the staminate inflorescence 4-12, 6-12 cm long; spathe of the pistillate inflorescence 5-9 cm long; E Cordillera .............................................. 47. C. reticulata
41. Trichilia laterally rounded; spikes of the staminate inflorescence 10-25, (6-)12-28 cm long; spathe of the pistillate inflorescence 10-17 cm long; E Cordillera ...................................................... 40. C. pastasana
40. Trichilia not on a subscrotiform (abaxially bulged) base of the petiole. 42. Lamina (sub)coriaceous and usually smooth above.
43. Upper surface of the lamina ? bullate, glabrous; leafy twigs and petioles villous to subsericeous to subhirsute; W Cordillera ............. 6. C. bullata
43. Upper surface of the lamina plane or plicate, or if slightly bullate and/ or with villous to subhirsute indumentum on the leafy twigs and petioles, then with arachnoid indumentum on the lamina above. 44. Upper surface of the lamina white or whitish with ? dense
(sometimes sparse) arachnoid indumentum. 45. Arachnoid indumentum on the lamina beneath in two
distinct layers (in the areoles and on the stiff hairs on the veins); spikes of the pistillate inflorescences mostly 4-5; E and central Cordilleras .............. 57. C. telenitida
45. Arachnoid indumentum on the lamina beneath not in two distinct layers; spikes of the pistillate inflorescence mostly 1- 3; W Cordillera .............. 56. C. telealba
44. Upper surface of the lamina without (or with very sparse) arachnoid indumentum. 46. Leafy twigs with ? dense arachnoid indumentum, spikes of
the staminate inflorescence 3-9, and those of the pistillate inflorescence l-3(-4); W Cordillera and Antioquia. 47. Lamina ? plicate and with 8-10(-I 1) segments; stipes
of the spikes of the staminate inflorescence 0.5-2.5 cm long; spikes of the pistillate inflorescence 2-3(-4) ................................................................................... 43. C. plicata
47. Lamina plane and with 7-8 segments; stipes of the spikes of the staminate inflorescence 0.4-0.5 cm long; spikes of the pistillate inflorescence 1 or 2 ............ 13. C. gabrielis
46. Leafy twigs without dense arachnoid indumentum, or if with such indumentum, then spikes of the staminate inflorescence 10-25(-40) and those of the pistillate inflorescence 3-8(-12). 48. Leafy twigs glabrous (often with a bluish waxy layer)
................................................................................ 13. C gabrielis 48. Leafy twigs hairy.
49. Stipules 22-50 cm long; leafy twigs sparsely hispidulous; spikes of the pistillate inflorescence 8- 18 cm long at anthesis, to 26 cm long in fruit; Pacific region ........... .................... 23. C. insignis
49. Stipules 6-25 cm long; leafy twigs ? densely hairy, hispidulous to subvillous; spikes of the pistillate inflorescence 1-7 cm long at anthesis, to 17 cm long in fruit; Andean region, widespread
. .................................................................... 4. C angustifolia 42. Lamina chartaceous to subcoriaceous and usually ? scabrous above.
50. Lateral veins in the midsegment mostly 15-35 pairs. 51. Spathes more or less densely hairy outside; Andean region,
widespread ............................. 4. C. angustifolia 51. Spathes sparsely hairy to subglabrous outside; Pacific region
............................................................................................. 37. C obtusifolia 50. Lateral veins in the midsegment 6-15(-17) pairs.
SYSTEMATIC TREATMENT 37
52. Upper surface of the lamina with ? dense (rarely sparse) arachnoid indumentum; W and central Cordilleras .............. 57. C. telenitida
52. Upper surface of the lamina without (or rarely with very sparse) arachnoid indumentum; lowland. 53. Stipules 6-8 cm long.
54. Lateral veins in the free part of the midsegment 7-10 pairs; spikes of the staminate inflorescence 4; spikes of the pistillate inflorescence often fused in pairs at the base; lithophytic, Tolima ............................... 16. C. goudotiana
54. Lateral veins in the free part of the midsegment (8-)10- 15(-17) pairs; spikes of the staminate inflorescence 10- 60; spikes of the pistillate inflorescence free at the base; not lithophytic, widespread ....................................... 41. C. peltata
53. Stipules 8-35 cm long. 55. Stipules sparsely hairy to subglabrous outside,
(usually) green; Amazonian region, inundated places ........................................................................ 3 1 . C . m em branacea
55. Stipules more or less densely hairy outside, often reddish or whitish. 56. Stipules 15-25(-35) cm long; Amazonian region
.......................................................................... 12 . C . ficifolia 56. Stipules 3-12 cm long; Pacific region, Cauca
valley (to Tolima) and N part of the country .... 41. C. peltata
Key to the species of Venezuela I . Trichilia absent.
2. Lamina with 10-15 segments; lowland, Amazonas, Bolivar, and Delta Amacuro ....................... 51. C. sciadophylla 2. Lamina with 7-10 segments; montane, Andes ....................... ........................................ 57. C. telenitida
1 . Trichilia present. 3. Arachnoid indumentum absent in the areoles of the lamina beneath.
4. Lamina chartaceous to subcoriaceous; stipules and spathes mostly greenish; spikes of staminate inflorescence ca. 0.2 cm thick; pistillate inflorescence pendulous at anthesis; Amazonas, inundated places ....................................................... 31. C. membranacea
4. Lamina (sub)coriaceous; stipules and spathes red; spikes of the staminate inflorescence 0.3-0.4(-l) cm thick; pistillate inflorescence erect at anthesis (to pendulous in fruit); Amazonas and Bolivar, non-inundated places ............................................................ 9. C. distachya
3. Arachnoid indumentum present in the areoles of the lamina beneath. 5. Stipules usually 15-35 cm long; indumentum (of the leafy twig) without uncinate hairs; Amazonas
and Bolfvar, (sub)montane ....................................................... 24. C. kavanayensis 5. Stipules usually to 15 cm long; indumentum (of the leafy twigs) with some of the hairs uncinate.
6. Lateral veins of the free part of the midsegment submarginally loop-connected. 7. Lamina with 6-7(-8) segments, the incisions down to ca. 5/10-9/10 from the margin; free
part of the midsegment spathulate to obovate and the lateral veins mostly unbranched; Amazonas .................................................. 12. C. fcifolia
7. Lamina with (7-)8-11 segments, the incisions down to ca. >5/10 from the margin, the free part of the midsegment oblong to elliptic to obovate and the lateral veins usually branched. 8. Lamina (sub)coriaceous and mostly smooth above; peduncle and spathe of the
inflorescences usually >10 cm long; stigma comose and apex of the perianth of the pistillate flower without arachnoid indumentum; Bolivar and Delta Amacuro, lowland ........................................................................................................................................ .3. C. angulata
8. Lamina chartaceous to subcoriaceous and ? scabrous above; peduncle and spathe of the inflorescences usually <10 cm long; stigma peltate and apex of the perianth of the pistillate flower with arachnoid indumentum; widespread, lowland (to montane) ........ 41. C. peltata
6. Lateral veins of the free part of the midsegment marginally loop-connected. 9. Lateral veins in the free part of the midsegment 9-17 pairs.
10. Petiole with dense arachnoid indumentum; the brown indumentum of the trichilium intermixed with distinctly longer white hairs; free parts of the lamina segments often ? lobate; llanos and savanna regions, widespread .............. .................... 32. C. metensis
38 FLORA NEOTROPICA
10. Petiole without or with sparse arachnoid indumentum; the brown indumentum of the trichilium intermixed with about equally long (or rarely longer) white hairs; free parts of the lamina segment entire; forested regions and riversides. 11. Lamina (sub)coriaceous and usually smooth above; lowland, Amazonas and
Bolivar ............................................................. 9. C. distachya 11. Lamina chartaceous to subcoriaceous.
12. Lamina attached to the petiole at ca. 1/4 of the total length from its base; stipules and spathes without arachnoid indumentum outside; stigma penicillate; Amazonas, Apure, Bolivar, and Delta Amacuro, inundated places
...................................................................................................................... 2 5 . C . la tilo b a 12. Lamina attached to the petiole at clearly >1/4 of the total length from its
base; stipules and spathes often with (dense) arachnoid indumentum outside; stigma peltate; widespread, non-inundated places ........................................ 41. C. peltata
9. Lateral veins in the free part of the midsegment usually 18-32 pairs. 13. Lateral veins in the free part of the midsegment 24-32 pairs; Amazonas, lowland
................................................................................................................................. .11. C. engleriana 13. Lateral veins in the free part of the midsegment 18-24(-25) pairs; Andes.
14. Spikes of the staminate inflorescence ca. 15-20; peduncle of the pistillate inflorescence 2-5 cm long and the spikes 3.5-7.5 cm long ........................... 4. C. angustifolia
14. Spikes of the staminate inflorescence 4-8; peduncle of the pistillate inflorescence 8-14 cm long and the spikes 11-33 cm long .................................................. 48. C. sararensis
Key to the species of the Guianas 1. Trichilia absent; incisions of the lamina down to the petiole and the segments ? clearly petiolulate
.............................................................................................................................................................. .51. C. sciadophylla l. Trichilia present; incisions of the lamina not down to the petiole and the segments not petiolulate.
2. Lamina with (12-)15-17 segments; lateral veins in the free part of the midsegment ca. 40-45 pairs; French Guiana and Suriname ..................................................................... 52. C. silvae
2. Lamina with 7-11 segments; lateral veins in the free part of the midsegment ca. 10-20 pairs. 3. Trichilia with whitish hairs 5-9 mm long; French Guiana (inselbergs) ........... ...................... 17. C. granvilleana 3. Trichilia with whitish hairs much shorter.
4. Lateral veins loop-connected in the margin of the lamina. 5. Stipules subpersistent; peduncle of the pistillate inflorescence 20-40 cm long; staminate
inflorescences with 4-6(-1 1) spikes, these 0.8-1.8 cm diam.; French Guiana and E Suriname ............... ............................................... 39. C. palmata
5. Stipules caducous; peduncle of the pistillate inflorescence to 16 cm long; staminate inflorescence usually with > 12 spikes, these 0.2-0.4 cm diam. 6. Lamina (usually) smooth above, (sub)coriaceous; French Guiana .............................. 9. C. distachya 6. Lamina scabrous above, chartaceous to subcoriaceous.
7. Stipules and spathes reddish, without arachnoid indumentum outside; stigma penicillate; widespread, in periodically inundated places ....................... ................ 25. C. latiloba
7. Stipules and spathes (usually) greenish or, if reddish, then with + dense arachnoid indumentum outside; stigma peltate; Guyana and Suriname, in non-inundated places ................................................................................................................................... .41. C. peltata
4. Lateral veins loop-connected (just) inside the margin of the lamina. 8. Stipules 12-20 cm long; Guyana ................................................................... 3. C. angulata 8. Stipules (usually) 3-12 cm long.
9. Upper surface of the lamina with ? dense arachnoid indumentum; stigma comose; widespread .............................................................. 36. C. obtusa
9. Upper surface of the lamina without arachnoid indumentum; stigma peltate; Guyana and Suriname .............................................................. 41. C. peltata
Key to the species of Ecuador 1. Trichilia enclosed in 2 pockets with slit-shaped apertures at the base of the petiole; coastal region ........ 61. C. virgusa 1. Trichilia exposed (superficial) or absent.
2. Trichilia absent.
SYSTEMATIC TREATMENT 39
3. Lamina usually with 10-15 segments; lateral veins in the free part of the midsegment ca. 25-40 (-45). 4. Leafy twigs hairy; coastal region .................................................................. 20. C. hispidissima 4. Leafy twigs (sub)glabrous; Amazonian region ................................................................. 51. C. sciadophylla
3. Lamina usually with 7-9 segments; lateral veins in the free part of the midsegment 10-18 pairs; montane. 5. Upper surface of the lamina smooth, subglabrous; spikes of the staminate inflorescence 3-6;
spikes of the pistillate inflorescence 1-2; Andes, W slopes .................................................... 13. C. gabrielis 5. Upper surface of the lamina ? scabrous; spikes of the staminate inflorescence 5-16; spikes of
the pistillate inflorescence (3-)4-5 spikes; Andes, E slopes. 6. Lamina (sub)coriaceous, the upper surface of the lamina with dense arachnoid indumentum;
main veins sparsely hairy beneath ............................................................. 57. C. telenitida 6. Lamina chartaceous to subcoriaceous, the upper surface without arachnoid indumentum;
smaller (and often also larger) veins beneath ? densely hairy ................. ............................ 2. C. andina 2. Trichilia present (sometimes poorly developed).
7. Lamina (usually) with 10-20(-24) segments; lateral veins in the free part of the midsegment mostly >28 pairs. 8. Setose (irritating) hairs present on young parts; coastal region.
9. Lamina (usually) ? scabrous above; trichilia well-defined; Mulllerian bodies white, ca. 1 mm long ............................................................. 14. C. garciae
9. Lamina smooth above; trichilia not well-defined; Mullerian bodies pink, ca. 3 mm long ...................................................................................................................................... .20. C. hispidissima
8. Setose hairs absent. 10. Lateral veins in the free part of the midsegment 50-80 pairs; Andes, E slopes .......... 60. C. velutinella 10. Lateral veins in the free part of the midsegment 15-50 pairs.
11. Lamina smooth above. 12. Lamina often white above, segments of young leaves patent; spikes of staminate
inflorescence 4-6(-8); spikes of pistillate inflorescence (1-)2-3, the spathe 10- 20 cm long ........................................ 29. C. maxima
12. Lamina green above, segments of young leaves deflexed; spikes of staminate inflorescence 10-25(-30); spikes of pistillate inflorescence 3-8(-12), the spathe 3-6(-8) cm long .............................................. 4. C. angustifolia
11. Lamina ? scabrous above. 13. Lateral veins in the free part of the midsegment usually 30-50 pairs; spathes and
stipules with dense arachnoid indumentum outside; Andes, E slopes to ca. 1000 m and Amazonian region ....................................... 33. C. montana
13. Lateral veins in the free part of the midsegment usually (I0-)15-25 pairs, or if to 35(-43) pairs, then the spathes and stipules without dense arachnoid indumentum outside. 14. Spathes of the inflorescences sparsely hairy to subglabrous outside; lamina
erect in the bud; coastal region .......... ......................... 37. C. obtusifolia 14. Spathes ? densely hairy outside; lamina often reflexed in the bud; Andean
region .................................... 4. C. angustifolia 7. Lamina with (6-)7-10(-l 1) segments; lateral veins in the free part of the midsegment mostly <25
pairs. 15. Lamina without arachnoid indumentum in the areoles beneath.
16. Base of the lamima entire or shallowly lobed, the upper part 3-lobed. 17. Spikes of the staminate inflorescence 4-5; peduncle of the pistillate inflorescence 25-
45 cm long; Amazonian region .......................................... 46. C. putumayonis 17. Spikes of the staminate inflorescence 7-12; peduncle of the pistillate inflorescence 3-
24 cm long; coastal region .................................................. 54. C. subintegra 16. Base of the lamina ? distinctly incised.
18. Lateral veins in the free part of the midsegment usually 15-25(-40) pairs; coastal
region .............................................. 37. C. obtusifolia 18. Lateral veins in the free part of the midsegment 9-16 pairs.
19. Spikes of the staminate inflorescence ca. 30-40; peduncle of the pistillate inflorescence 5-15(-20) cm long; stipules mostly greenish; Amazonian region (and coastal region), usually inundated places ...................................... 31. C. membranacea
19. Spikes of the staminate inflorescence 4-7(-15); peduncle of the pistillate
40 FLORA NEOTROPICA
inflorescence 15-50 cm long; stipules mostly reddish; Amazonian region (to Andean region), non-inundated places .............................................. 59. C. utcubambana
15. Lamina with arachnoid indumentum in the areoles beneath. 20. Trichilia separate.
21. Trichilia on a subscrotiform (abaxially bulging) base of the petiole; Andes, E slopes .............................................................................................................................. 40. C. pastasana
21. Trichilia not on a subscrotiform base of the petiole. 22. Leafy twigs glabrous and smooth, mostly with a bluish waxy layer; Andes.
23. Spikes of the staminate inflorescence 3-6; spikes of the pistillate inflorescence 1-2; Andes, W slopes ................................................. 13. C. gabrielis
23. Spikes of the staminate inflorescence (5-)6-16; spikes of the pistillate inflorescence 3-5; Andes, E slopes. 24. Lamina chartaceous to subcoriaceous, without arachnoid indumentum
above ............................................. 2. C. andina 24. Lamina (sub)coriaceous, with (often dense) arachnoid indumentum
above ............................................. 57. C. telenitida 22. Leafy twigs hairy and often ? scabrous, green or reddish-brown; lowlands.
25. Petiole as long as ca. half the diameter of the lamina; spikes of staminate inflorescence (20-)30-50; stipes of the spikes of the pistillate inflorescence to 0.5 cm long; Amazonian region ................................................. 28. C. marginalis
25. Petiole much longer than half the diameter of the lamina; spikes of the staminate inflorescence 5-9; stipes of the spikes of the pistillate inflorescence 0.5-1 cm long; coastal region ............................................ 23. C. insignis
20. Trichilia fused. 26. Trichilia on a subscrotiform (abaxially bulging) base of the petiole.
27. Trichilia laterally extended by lobes; spikes of the staminate inflorescence 4- 12, 6-12 cm long; spathe of the pistillate inflorescence 5-9 cm long; Andes, W slopes .................................................... 47. C. reticulata
27. Trichilia laterally rounded; spikes of the staminate inflorescence 10-25, (6-)12- 28 cm long; spathe of the pistillate inflorescence 10-17 cm long; Andes, E slopes .................................................... 40. C. pastasana
26. Trichilia not on a subscrotiform base of the petiole. 28. Lateral veins of the free part of the midsegment marginally loop-connected.
29. Lateral veins in the free part of the midsegment 21-32 pairs; Amazonian region .1................................................. l.C. engleriana
29. Lateral veins in the free part of the midsegment 12-19 pairs. 30. Lamina (sub)coriaceous, usually smooth above; Amazonian region,
non-inundated places ............................................. 9. C. distachya 30. Lamina chartaceous to subcoriaceous, ? scabrous above.
31. Lamina attached to the petiole at ca. 1/4 of the total length from its base; spikes of the staminate inflorescence usually ca. 15-20, usually 14-20 cm long; stigma penicillate; Amazonian region, inundated places .......... ............................... 25. C. latiloba
31. Lamina attached to the petiole at clearly >1/4 of the total length from its base; spikes of the staminate inflorescence usually ca. 5- 15, usually 5-12 cm long; stigma peltate; coastal region, usually in non-inundated places, and Amazonian region, in inundated places .......................................... 26. C. litoralis
28. Lateral veins of the free part of the midsegment submarginally loop-connected. 32. Lamina ? scabrous above; Amazonian region ......................... .............. 12. C. ficifolia 32. Lamina smooth above.
33. Lamina with (often dense) arachnoid indumentum above and leafy twigs either glabrous or villous; Andes, E slopes .............. .......... 57. C. telenitida
33. Lamina without arachnoid indumentum above or, if with that indumentum, then the leafy twigs hispidulous. 34. Incisions of the lamina down to 3/10-6/10 from the margin.
35. Upper surface of the lamina usually bullate; spikes of the staminate inflorescence 4-10; spikes of the pistillate inflorescence 1-2; Andes, W slopes ................................... 6. C. bullata
SYSTEMATIC TREATMENT 41
35. Upper surface of the lamina smooth; spikes of the staminate inflorescence 20-50; spikes of the pistillate inflorescence 4 (-7); Amazonian region ..................................... 28. C. marginalis
34. Incisions of the lamina (usually) down to 6/10-9/10 from the margin; upper surface smooth; spikes of the pistillate inflorescence (usually) 3-8. 36. Spikes of the staminate inflorescence 5-9; spikes of the
pistillate inflorescence 8-18 cm long; coastal region ...... 23. C. insignis 36. Spikes of the staminate inflorescence 10-50; spikes of the
pistillate inflorescence 3-8(-12) cm long; Andes and coastal region ..................................... 4. C. angustifolia
Key to the species of Peru 1. Trichilia absent.
2. Incisions of the lamina down to the petiole and the segments often petiolulate; Amazonian region ........................................................................................................................................................ ...51. C. sciadophylla
2. Incisions of the lamina not down to the petiole; Andean region. 3. Lamina with 7-10(-1 1) segments.
4. Lamina chartaceous to subcoriaceous, without arachnoid indumentum above ............ ............... 2. C. andina 4. Lamina (sub)coriaceous, with (often dense) arachnoid indumentum above .......................... 57. C. telenitida
3. Lamina with 10-1 8 segments. 5. Lateral veins in the midsegment of the lamina 15-35 pairs; upper surface of the lamina ?
scabrous; Andes ............................................................... 55. C. tacuna 5. Lateral veins in the midsegment of the lamina 10-16 pairs; the upper surface of the lamina
smooth; Andes .............................................................. 1. C. albicans 1. Trichilia present.
6. Trichilia separate (in 2 lateral patches). 7. Trichilia on a subscrotiform (abaxially bulging) base of the petiole; Andes .............................. 40. C. pastasana 7. Trichilia not on a subscrotiform base of the petiole.
8. Lamina (sub)coriaceous, with (often dense) arachnoid indumentum (and thus usually white) above; Andes ............................................................... 57. C. telenitida
8. Lamina chartaceous to subcoriaceous, without arachnoid indumentum above. 9. Leafy twigs (sub)glabrous, smooth, with a bluish waxy layer; Andes .............. .................... 2. C. andina 9. Leafy twigs hairy.
10. Petiole as long as ca. half the diameter of the lamina; terminal buds usually curved, not inflated; Amazonian region ........................................................ 28. C. marginalis
10. Petiole much longer than half the diameter of the lamina; terminal buds straight, ? inflated; Amazonian region to Andes ......................... ............................... 53. C. strigosa
6. Trichilia fused. 1 1. Lamina with 10-20(-24) segments.
12. Terminal buds + inflated; lateral veins in the free part of the midsegment 8-17 pairs. 13. Lamina coriaceous, with ? dense arachnoid indumentum and smooth above; Andes
(2200-2500 m) ............................................................................ 1. C. albicans 13. Lamina chartaceous to subcoriaceous and without arachnoid indumentum and scabrous
or smooth above; Amazonian region to Andes (to 1900 m) 53. C. strigosa 12. Terminal buds not inflated; lateral veins in the free part of the midsegment 15-80 pairs.
14. Lateral veins in the free part of the midsegment 50-80 pairs; Andes ........... ........... 60. C. velutinella 14. Lateral veins in the free part of the midsegment 14-50 pairs.
15. Incisions of the lamina down to the petiole or at most 2 cm from the petiole; Amazonian region ............................................................. 18. C. herthae
15. Incision of the lamina down to >2 cm from the petiole. 16. Lateral veins in the free part of the midsegment usually 30-50 pairs; leafy
twig whitish, green, or brownish; Amazonian region .................................. 33. C. montana 16. Lateral veins in the free part of the midsegment ca. 15-25 pairs, or if to 35
(-43) pairs, then the leafy twig usually turning blackish. 17. Upper segments of the lamina lobate; lamina erect in the bud; peduncle
of the inflorescences usually >8 cm long; Amazonian region (to Andes, to 1350 m)44 .................................................. C. polystachya
42 FLORA NEOTROPICA
17. Upper segments of the lamina entire; lamina often reflexed in the bud; peduncle of the inflorescences usually to 8 cm long; Andes (mostly 1200- 2400 m, sometimes down to 400 m) ....................... ......................... 4. C. angustifolia
11. Lamina with 7-10(-1 1) segments. 18. Lamina without arachnoid indumentum in the areoles beneath.
19. Base of the lamima entire or shallowly lobed, the upper part 3-lobed; Amazonian region ................................................................................................................................ .46. C. putumayonis 19. Base of the lamina ? distinctly incised. 20. Spikes of the staminate inflorescence ca. 4-7(-15); peduncle of the pistillate
inflorescence 15-50 cm long; stipules mostly reddish; Amazonian region, non- inundated places ........................................................ 59. C. utcubambana
20. Spikes of the staminate inflorescence ca. 25-50; peduncle of the pistillate inflorescence ca. 5-15(-20) cm long; stipules mostly greenish. 21. Terminal buds ? inflated; stipes of spikes of the staminate inflorescence (0.5-)
1-2.5 cm long; spikes of the pistillate inflorescence 3-7(-9); Amazonian region to Andes (to 1900 m), non-inundated places .................................... 53. C. strigosa
21. Terminal buds not inflated; stipes of spikes of the staminate inflorescence 0.5- 1 cm long; spikes of the pistillate inflorescence (2-)4; Amazonian region, usually inundated places ................................................................ 31. C. membranacea
18. Lamina with arachnoid indumentum in the areoles beneath. 22. Lateral veins in the free part of the midsegment marginally loop-connected.
23. Lateral veins in the free part of the midsegment 24-32 pairs. 24. Inflorescences pendulous, the peduncle usually 10-19 cm long; Amazonian
region ................................................. IIC. engleriana 24. Inflorescences erect, the peduncle usually 1-8 cm long; Andes (mostly 1200-
2200 m) ........................................................ 4. C. angustifolia 23. Lateral veins in the free part of the midsegment 12-24 pairs.
25. Incisions of the lamina down to 1.5 cm from the petiole. 26. Inflorescences pendulous, the peduncles usually 10-22 cm long;
Amazonian region .............................................. 8. C concolor 26. Inflorescences erect, the peduncles usually 1-8 cm long; Andes to
Amazonian region ([400-]1200-2400 m) . ........................... 4. C. angustifolia 25. Incisions of the lamina down to at most 4 cm from the petiole.
27. Lamina (sub)coriaceous, usually smooth above. 28. Free parts of the upper segments of the lamina obovate to
subobovate; Amazonian region ........................................... 9. C. distachya 28. Free parts of the upper segments of the lamina oblanceolate; Andes
(mostly 1200-2400 m) .................................................... 4. C. angustifolia 27. Lamina chartaceous to subcoriaceous, mostly ? scabrous above.
29. Spathes with dense arachnoid indumentum; peduncles usually 10-20 cm long, and lateral veins in the free part of the midsegment 21-32 pairs ............................................ 11. C. engleriana
29. Spathes without or with sparse arachnoid indumentum; peduncles to 10 cm long, and/or lateral veins in the free part of the midsegment to 20 pairs. 30. Lamina attached to the petiole at ca. 1/4 of the total length
from its base; spikes of the staminate inflorescence usually ca. 15-20, usually ca. 14-20 cm long; stigma penicillate; Amazonian region, inundated places ..................................... 25. C. latiloba
30. Lamina attached to the petiole at clearly > 1/4 of the total length from its base; spikes of the staminate inflorescence usually ca. 5-15 and/or usually 5-12 cm long; stigma peltate (lowland) or comose (montane). 31. Stipules and spathes chartaceous to membranaceous when
31. Stipules and spathes subcoriaceous when dry, densely hairy; stigma comose; Andes (mostly 1200-2400 m)
. .................................................................................. 4 . C angustifolia
SYSTEMATIC TREATMENT 43
22. Lateral veins in the free part of the midsegment submarginally loop-connected. 32. Trichilia on subscrotiform (abaxially bulging) base of the petiole.
33. Lamina with ? dense arachnoid indumentum above; spikes of the staminate inflorescence 10-25; Andes (900-2300 m) ............................................... 40. C. pastasana
33. Lamina without arachnoid indumentum above; spikes of the staminate inflorescence ca. 25-100; Amazonian region and Andes (to 1900 m) ........ 53. C. strigosa
32. Trichilia not on a subscrotiform base of the petiole. 34. Leafy twigs (sub)glabrous, smooth, bluish by a waxy layer; Andes ............... 2. C. andina 34. Leafy twigs ? hairy.
35. Upper lamina segments lobate; Amazonian region and Andes (to 1350 m) ....................................................................................... 44. C. polystachya
37. Stipules subpersistent; Mulllerian bodies pinkish when fresh; spikes of the staminate inflorescence 7-8; spikes of the pistillate inflorescence 4, often curved; Andes .............. 7. C. chlorostachya
37. Stipules caducous, or if subpersistent, then the Mullerian bodies white, the spikes of the staminate inflorescence (usually) 10- 25(-40), and the spikes of the pistillate inflorescence often >4, straight, and often thickened toward the base. 38. Lateral veins in the free part of the midsegment usually 12-
18 pairs; spikes of the pistillate inflorescence 2-4, usually 8-15 cm long at anthesis; Amazonian region ............... 9. C. distachya
38. Lateral veins in the free part of the midsegment usually 15- 35 pairs; spikes of the pistillate inflorescence often >4, 1- 7 cm long at anthesis; Andes .................................... 4. C. angustifolia
36. Lamina chartaceous to subcoriaceous. 39. Petiole as long as ca. half the diameter of the lamina; stipules
ca. 15-45 cm long, glabrous inside; terminal buds usually curved, glabrous inside; Amazonian region ...................... 28. C. marginalis
39. Petiole much longer than half the diameter of the lamina; stipules often < 15 cm long, or if longer, then often hairy inside; terminal buds straight. 40. Lateral veins in the free part of the midsegment
unbranched; incisions down to ca. 5/10, or if more deeply, then the midsegment broadly spathulate; spikes of the staminate inflorescence 8-13 and the peduncle of the staminate inflorescence usually 10-20 cm long; Amazonian region ........... ........................ 12. C. ficifolia
40. Lateral veins in the free part of the midsegment mostly branched; incisions down to 5/10-8/10, the midsegment obovate, elliptic, subobovate, oblong, or oblanceolate; spikes of the staminate inflorescence 4-7 or ca. 25-100. 41. Lateral veins in the free part of the midsegment
usually ca. 20-35 pairs, the midsegment usually oblanceolate; lamina often reflexed in the bud; Andes (mostly 1200-2400 m) ................................ 4. C. angustifolia
41. Lateral veins in the free part of the midsegment usually 8-17 pairs, the midsegment usually obovate to elliptic or subobovate; lamina erect in the bud; lowland to submontane. 42. Terminal buds + inflated; lamina often ?
bullate above; indumentum of the leafy twig puberulous to hirtellous or to subhirsute; with straight hairs; Amazonian region to Andes (to 1900 m) ........................... 53. C. strigosa
42. Terminal buds not inflated; lamina not bullate above; indumentum of the leafy twig partly
44 FLORA NEOTROPICA
(sub)setulose or puberulous to hispidulous with uncinate hairs; Amazonian region. 43. Spikes of the staminate inflorescence ca. 30-
40; peduncle of the pistillate inflorescence 5- 15(-20) cm long; leafy twig partly (sub)setulose; Amazonian region, usually inundated places .... ..... 31. C. membranacea
43. Spikes of the staminate inflorescence 4-7 (-15); peduncle of the pistillate inflorescence 15-50 cm long; leafy twig not (sub)setulose; Amazonian region, non- inundated places .... ..... 59. C. utcubambana
Key to the species of Amazonian Brazil 1. Trichilia absent; incisions of the lamina down to the petiole and the segments ? clearly petiolulate
.............................................................................................................................................................. .51. C. sciadophylla 1. Trichilia present; incisions of the lamina not down to the petiole and the segments not petiolulate.
2. Lamina with 12-15 segments. 3. Lateral veins in the free part of the midsegment ca. 40-45 pairs; NE Amazon basin .......... ............ 52. C. silvae 3. Lateral veins in the free part of the midsegment ca. 20-25 pairs; NE to central Amazon basin ......... 58. C. ulei
2. Lamina with up to 10(-12) segments. 4. Lamina incised down to 1.5(-2.5) cm from the petiole; lower, central and SW Amazon basin .... 8. C. concolor 4. Lamina incised down to at most 4 cm from the petiole.
5. Lateral veins of the free part of the midsegment marginally loop-connected. 6. Lateral veins in the free part of the midsegment of the lamina (21-)24-32 pairs; western
Amazon basin .1................................................................ L C. engleriana 6. Lateral veins in the free part of the midsegment 10-17 pairs.
7. Lamina attached to the petiole at ca. 1/4 of the total length from its base, chartaceous to subcoriaceous, scabrous above; stipules caducous; widespread, inundated places ...... 25. C. latiloba
7. Lamina attached to the petiole clearly > 1/4 of its total length from its base, (sub)coriaceous, smooth above, or if + scabrous, then the stipules often subpersistent; widespread, non-inundated places. 8. Inflorescences pendulous at anthesis; spathes white due to dense arachnoid
indumentum outside; stipules often subpersistent .................................................. 39. C. palmata 8. Inflorescences erect (at least at anthesis); spathes not white, without dense arachnoid
indumentum outside; stipules caducous .................................................................. 9. C. distachya 5. Lateral veins of the free part of the midsegment submarginally loop-connected.
9. Indumentum (of the leafy twig) without uncinate hairs; Roraima, submontane ....... 24. C. kavanayensis 9. Indumentum (of the leafy twig) with some of the hairs uncinate.
10. Lower surface of the lamina without arachnoid indumentum in the areoles. 11. Lamina (sub)coriaceous; stipules red; widespread, non-inundated places ......... 9. C. distachya 11. Lamina chartaceous (to subcoriaceous); stipules mostly green; widespread,
inundated places .............. ............................................ 31. C. membranacea 10. Lower surface of the lamina with arachnoid indumentum in the areoles.
12. Lamina (sub)coriaceous; petiole and usually also the upper surface of the lamina with dense arachnoid indumentum; lower to central Amazon basin ........... ......... 36. C. obtusa
12. Lamina chartaceous to subcoriaceous; petiole and usually also the upper surface of the lamina without or with sparse arachnoid indumentum. 13. Midsegment of the lamina oblong to elliptic; lateral veins in the midsegment
usually branched; spathes with ? dense arachnoid indumentum outside; N fringe of the Amazon basin ................................................ 41. C. peltata
13. Midsegment of the lamina broadly spathulate to obovate or ovate; lateral veins of the midsegment usually unbranched; spathes without or with sparse arachnoid indumentum outside. 14. Midsegment of the lamina ovate; stipules 8-16 cm long; central
Amazon basin ............................................ 45. C. purpurascens 14. Midsegment of the lamina broadly spathulate to obovate; stipules
usually ca. 15-25 cm long; W (to central) Amazon basin .................. 12. C. ficifolia
SYSTEMATIC TREATMENT 45
Key to the species of Bolivia
1. Trichilia absent; incisions of the lamina down to the petiole and the segments ? clearly petiolulate .............................................................................................................................................................. .51. C. sciadophylla
1. Trichilia present; incisions of the lamina not down to the petiole and the segments not petiolulate. 2. Trichilia separate (2 lateral patches); submontane ........................................................................ 53. C. strigosa 2. Trichilia fused.
3. Lamina with 10-16 segments; montane. 4. Spikes of the inflorescences 18-40 cm long at anthesis ..............................................0........... I. C. elongata 4. Spikes of the inflorescences 1.5-13 cm at anthesis, in fruit to 17 cm long ......................... 4. C. angustifolia
3. Lamina with (5-)7-10(-12) segments; lowland (or montane). 5. Lateral veins of the free part of the midsegment marginally loop-connected.
6. Lateral veins in the free part of the midsegment usually 24-32 pairs; lowland, widespread ......................................................................................................................................... .11. C. engleriana
6. Lateral veins in the free part of the midsegment 8-24 pairs. 7. Incisions of the lamina down to 1.5(-2.5) cm from the petiole; lowland (widespread)
........................................................................................................................................ .8. C. concolor 7. Incisions of the lamina down at most to ca. 4 cm from the petiole.
8. Lateral veins in the free part of the midsegment 8-10 pairs; lowland (Santa Cruz) ................................................................................................................................. 49. C saxatilis
8. Lateral veins in the free part of the midsegment 10-20 pairs. 9. Lamina attached to the petiole at ca. 1/4 of the total length from its base,
chartaceous; lowland, inundated places (Beni and Pando) ................................ 25. C. latiloba 9. Lamina attached to the petiole at clearly > 1/4 of its total length from its base,
subcoriaceous to coriaceous; lowland, non-inundated places. 10. Stipules with dense arachnoid indumentum outside; trunk with prominent
stipular scars; lowland (La Paz) ................................. ................... 5. C. annulata 10. Stipules without or with sparse arachnoid indumentum outside; trunk
without prominent stipular scars. 11. Inflorescences pendulous; spathes with dense arachnoid indumentum
outside; stipules often subpersistent; lowland (Santa Cruz) ............. 39. C. palmata 11. Inflorescences initially erect, pendulous in fruit; spathes without or
with sparse arachnoid indumentum outside; stipules always caducous; lowland (La Paz and Santa Cruz) ................................................ 9. C. distachya
5. Lateral veins in the free part of the midsegment submarginally loop-connected. 12. Lamina without arachnoid indumentum in the areoles beneath.
13. Spikes of the staminate inflorescence ca. 4-7(-15); peduncle of the pistillate inflorescence 15-50 cm long; stipules mostly reddish; lowland (Pando) ........ 59. C. utcubambana
13. Spikes of the staminate inflorescence ca. 25-50; peduncle of the pistillate inflorescence ca. 5-15(-20) cm long; stipules mostly greenish. 14. Terminal buds ? inflated; stipes of spikes of the staminate inflorescence (0.5-)1-
2.5 cm long; spikes of the pistillate inflorescence 3-9; submontane ............... 53. C. strigosa 14. Terminal buds not inflated; stipes of spikes of the staminate inflorescence 0.5-1
cm long; spikes of the pistillate inflorescence (2-)4; lowland, usually inundated places ................................................... 31. C. membranacea
12. Lamina with arachnoid indumentum in the areoles beneath; non-inundated places. 15. Upper segments of the lamina lobate; lowland (widespread) ............. ............... 44. C. polystachya 15. Upper segments of the lamina entire.
16. Lamina (sub)coriaceous; spikes of the staminate inflorescence 4-8, the peduncle 3-9 cm long; spathe of the pistillate inflorescence 6-8 cm long; lowland (Santa Cruz) .................................................... 49. C. saxatilis
16. Lamina chartaceous to subcoriaceous; spikes of the staminate inflorescence 8- 100, or if <8, then the peduncle usually 10-21 cm long; spathe of the pistillate inflorescence 8-18(-23) cm long. 17. Terminal buds ? inflated; lateral veins of the midsegment mostly branched;
spikes of the staminate inflorescence (15-)25-100; submontane ............ 53. C. strigosa 17. Terminal buds not inflated; lateral veins of the midsegment often
unbranched; spikes of the staminate inflorescence 8-15; lowland (Beni and/ or Pando).
46 FLORA NEOTROPICA
18. Stipules (12-)18-25(-35) cm long; midsegment of the lamina usually broadly spathulate ......... ................................... 12. C. ficifolia
18. Stipules 4-12 cm long; midsegment of the lamina oblong to obovate ................................................................................................... 59. C. utcubam bana
Key to the species of Extra-Amazonian Brazil, Paraguay, and Argentina 1. Trichilia absent; upper surface of the lamina white due to dense arachnoid indumentum; E Brazil ...... 21. C. hololeuca 1. Trichilia present; upper surface of the lamina not white, without dense arachnoid indumentum.
2. Lateral veins in the free part of the midsegment 8-10 pairs; central Brazil ............... ......................... 49. C. saxatilis 2. Lateral veins in the free part of the midsegment 10-20(-23) pairs.
3. Stipules white to greenish, with + dense arachnoid indumentum outside; stigma peltate, and arachnoid indumentum covering the apex of the perianth of the pistillate flower; anthers 0.5-0.7 mm long; spikes of the staminate inflorescence often >10 and often <1 cm diam.; Brazil (widespread), Argentina, and Paraguay ............................. .................................... 38. C. pachystachya
3. Stipules usually red(dish), without or with sparse arachnoid indumentum outside; stigma comose to penicillate, and arachnoid indumentum not covering the apex of the perianth of the pistillate flower; anthers 0.8-2 mm long; spikes of the staminate inflorescence often <10, often >1 cm diam. 4. Spathes with dense arachnoid indumentum outside (white); NE Brazil (Paraiba, Pernambuco,
and Sergipe) ............................................................... 39. C. palmata 4. Spathes without or with sparse arachnoid indumentum outside (reddish); Bahia to Rio Grande
do Sul .............................................................. 15. C. glaziovii
Key to the species of the Cecropia peltata-group 1. Trichilia absent or poorly developed (in 2 patches and/or without Mullerian bodies).
2. Internodes with ample brown pith; Cuba, Hispaniola, Puerto Rico, Lesser Antilles ......... ......... 50. C. schreberiana 2. Internodes with scarce whitish pith; Jamaica .......................................................................... 41. C. peltata
1. Trichilia present and well-developed (fused). 3. Incisions of the lamina (at least some) down to near (at most 1.5 cm or rarely 2.5 cm from) the petiole;
lower to SW Amazon basin ........................................................................ 8. C concolor 3. Incisions down to 9/10, at least 2 cm from the petiole.
4. Lateral veins in the free part of the midsegment (21-)24-32 pairs; upper Amazon basin, from Bolivia to Colombia .................................................................... 11. C. engleriana
4. Lateral veins in the free part of the midsegment 9-20(-24) pairs. 5. Trichilia with white (or brownish) unicellular hairs ? dense and distinctly longer than the
brown indumentum. 6. Stipules and mostly also the petiole with dense arachnoid indumentum.
7. Stipules to 13 cm long; lamina segments sometimes lobate; savannas from Colombia to Guyana ............................................................... 32. C. metensis
7. Stipules to 22 cm long; lamina segments often lobate; extra-Amazonian Brazil, Paraguay, Argentina, Paraguay, and S fringe of the Amazon basin ..................... 38. C. pachystachya
6. Stipules and petiole without or with sparse arachnoid indumentum. 8. Stipules 3-4 cm long; lateral veins in the free part of the midsegment 9-10 pairs;
French Guiana, inselbergs ......................................................... 17. C. granvilleana 8. Stipules 3-10(-12) cm long; lateral veins in the free part of the midsegment (8-)10-15
(-17) pairs; Jamaica, Central America, N South America, eastward to Suriname, Trinidad, Tobago, N fringe of Amazonian Brazil (Para, Roraima) ............. .................. 41. C. peltata
5. Trichilia with (inconspicuous) short white hairs. 9. Stipules usually to 10 cm long and spathes usually to 7 cm long; segments of the lamina
entire; Jamaica, Central America, N South America, eastward to Suriname, Trinidad, Tobago, N fringe of Amazonian Brazil (Para, Roraima) ..................................................... 41. C. peltata
9. Stipules usually 10-20 cm long and spathes mostly 8-19 cm long, if shorter, then the segments of the lamina often lobate to sinuate. 10. Spathes often ,0 cm long; segments of the lamina often lobate to sinuate; central to E
Brazil to Argentina, Paraguay, and S fringe of the Amazon basin .......... .......... 38. C. pachystachya 10. Spathes usually 10-19 cm long; segments of the lamina entire.
glabrous; E Cordillera, Colombia to Venezuela, mostly submontane to montane .......................................................................................................................... 48. C sararensis
11. Spathes sparsely hairy inside; leafy twigs densely hispidulous; Pacific Coastal lowlands of Ecuador and Colombia and NW Amazon basin (Colombia, Ecuador, and Peru) ....................................................... 26. C. litoralis
1. Cecropia albicans Trecul, Ann. Sci. Nat. Bot., Ser. 3, 8: 82. 1847 (Aug). Type. Peru. Without locality, without date (Y), Ruiz & Pavo'n s.n. (ho- lotype: FI-W, photograph seen; isotypes: F, G, LE).
Tree, to 20 m tall; branches (very) short and de- parting at angles of ca. 900 at the apex of the tree or longer and then departing in acute angles at the lower part of the tree. Leafy twigs 3-10 cm thick, blackish or reddish to green and then with a bluish waxy layer, densely (sub)hirsute or (sub)glabrous. Lamina coria- ceous, ca. 25 X 25 cm to 95 x 95 cm; the segments 11-13, the upper ones in deeply incised laminas ob- long (to subovate), the incisions down to 4/10 or to 8/10; apices acute to subacuminate; upper surface smooth, subglabrous or with ? dense arachnoid in- dumentum; lower surface rather sparsely to densely tomentose on the veins, arachnoid indumentum in the areoles and on the smaller veins or also on the main veins; lateral veins in the free part of the midsegment 10-16 pairs, submarginally loop-connected, in the lower part often not distinctly so, often ? branched; petiole ca. 30-75 cm long, puberulous and with sparse to rather dense arachnoid indumentum or subgla- brous; trichilia absent or present and fused, the brown indumentum intermixed with short to rather long white to pale brown (unicellular) hairs; stipules ca. (lO-)25-45 cm long, red to pinkish, sometimes with a bluish waxy layer, with only moniliform brown pluricellular hairs outside, glabrous inside; terminal buds inflated. Staminate inflorescences in pairs, erect, usually subtended by pinkish or greenish, caducous bracts to 4 cm long, attached at or ca. 1 cm above the base of the peduncle; peduncle 3-7 cm long, (sub)glabrous; spathe ca. 4-10 cm long, yellowish to pinkish, with only moniliform brown pluricellular hairs outside, glabrous inside; spikes (7-)15-24, 3.5- 9 x 0.3-0.7 cm, sessile or with stipes 0.2-0.3 cm long and sparsely hairy; rachis hairy. Staminate flowers: perianth tubular, 1-1.5 mm long, sparsely and min- utely puberulous on the lower part of the apex, the apex plane, ? clearly 2-lobed; filaments flat; anthers ca. 0.8 mm long, appendiculate, detached at anthesis, remaining attached to the filament by 2 filiform con- nections between the connective and the upper margin of the filament. Pistillate inflorescences in pairs, erect, usually subtended by pinkish, caducous bracts to 4
cm long, attached at or ca. 1 cm above the base of the peduncle; peduncle 1.5-2.5 (or to 14) cm long, sub- glabrous; spathe 5-11 cm long, the color and indu- mentum as in the staminate inflorescence; spikes 4- 7(-9), 4-10 X 1-1.2 cm, to 15 X 2 cm in fruit, sessile; rachis glabrous. Pistillateflowers: perianth ca. 2 mm long, with arachnoid indumentum below the apex out- side, also in the lower part of the style channel inside, the apex ? convex, punctate to submuriculate; style rather long, straight, unilaterally hairy; stigma co- mose. Fruit oblongoid, ca. 2.5 mm long, ? tubercu- late.
Distribution (Fig. 8.2). Peru (Pasco to San Mar- tin), in cloud forest, at 2200-2500 m.
Specimens examined. PERU. HuANUCO: Rd. Huan- uco-Tingo Maria, Carpish, ca. 2500 m, 23 Nov 1997 (6), Berg et al. 1730 (BG, COL, MOL); rd. Hudnuco-Tingo Maria, below Carpish, ca. 2400 m, 23 Nov 1997 (Y fl-fr), Berg et al. 1732 (BG, COL, MOL). PASCO: Prov. Oxa- pampa, rd. Oxapampa-Lluapi, km 3-4, ca. 2250 m, 29 Nov 1997 (Y fl-fr), Berg et al. 1757 (BG, COL, MOL), (6), Berg et al. 1758 (BG, COL, MOL); Prov. Oxapampa, rd. Oxa- pampa-Cerro de Pasco, 20-30 km W of Oxapampa, 2000- 2500 m, 3 Feb 1983 (Y fr), Gentry et al. 39956 (BG, MO, MOL, UMS). SAN MARTiN: Prov. Rioja, rd. Rioja-Poma- cocha, ca. km 379, ca. 2200 m, 8 Dec 1997 (d), Berg et al. 1800 (BG, COL, MOL); Prov. Mariscal Caceres, Abiseo Na- tional Park, Las Palmas, E of Gran Pajaten ruins, 2250 m, 17 Aug 1986 (? fl), Young 4071 (F, MOL). WITHOUT LO-
CALITY: (6), Ruiz & Pav6n s.n. (BM, K).
The material collected in Pasco, matching the type material, has been made from trees with the typical Cecropia tree shape, although the primary branches are short and do not branch, the stems are broadened toward the apex, the leafy twigs are densely hairy (and blackish), the lamina of adult trees is incised <5/10, and trichilia are present. Berg et al. 1800 from San Martin largely matches that from Pasco, but Young 4071 from San Martin matches the Huanuco material.
The material collected in Huanuco taken from trees more or less fastigiate in shape, with lateral branches departing (at one to three levels) in acute angles from the lower part of the stem, the stem is not broadened toward the apex, the leafy twigs are glabrous with a bluish waxy layer, the lamina is incised >5/10, and trichilia are absent. In other features the two types are largely similar, occur at similar elevations, and co- occur with C. tacuna both in Pasco and Huanuco.
48 FLORA NEOTROPICA
tAX~~~~~~~~
FIG. 8. Distribution maps. 1. Cecropia latiloba, circles; C. tacuna, hexagons; C. virgusa, triangles. 2. C. albicans,
hexagons; C. heterochroma, triangles; C marginalis, circles. 3. C. litoralis, circles; C. longipes, hexagons; C. mutisiana,
triangles. 4. C. obtusa. 5. C. chlorostachya, hexagon; C. obtusifolia, circles; C. pittieri, triangle. 6. C. granvilleana, open
circles; C. hololeuca, hexagons; C membranacea, circles; C. saxatilis, triangles. 7. C. garciae, triangles (question mark for uncertain locality); C. montana, circles.
SYSTEMATIC TREATMENT 49
Young 4071 also has glabrous leafy twigs and also lacks trichilia. Juvenile specimens of the Pasco type has laminas with incisions to 9/10, the free part of the midsegment has ca. 17-20 pairs of lateral veins, the dense subhirsute indumentum found in the leafy twigs occurs also on the petioles and on the main veins of the lamina beneath, and the stipules are pinkish. The peduncle of the pistillate inflorescence is usually very short (to 2.5 cm), but in Young 4071 the peduncles are much longer (to 9 cm). The trees in Pasco are usually inhabited by ants, those in Huainuco not.
2. Cecropia andina Cuatrecasas, Revista Acad. Col- omb. Ci. Exact. 6(22/23): 286. 1945. Type. Co- lombia. Putumayo: Nr. source of Rio Putumayo, Sibundoy Valley, 2200 m, 3 Jan 1941 (Y) Cuatre- casas 11662 (holotype: COL; isotype: F). Fig. 9
Tree, to 20 m tall. Leafy twigs 2-4 cm thick, bluish due to a waxy layer, glabrous; pith brown and copi- ous. Lamina subcoriaceous to chartaceous, ca. 40 X 40 cm to 80 X 80 cm (to 130 X 130 cm), the segments 7-9, the free parts of upper segments elliptic to su- bobovate, the incisions down to 4/10-7/10 or in the lower part of the lamina <4/10; apices acuminate to obtuse; upper surface ? scabrous, sometimes ? bul- late, hirtellous to puberulous to hispidulous subhir- sute; lower surface hirtellous to subtomentose on the veins or the primary veins subglabrous, the arachnoid indumentum in the areoles, often very short and sparse or only near the margin, sometimes absent; lat- eral veins ca. 10-15 pairs, submarginally loop- connected, most of them branched; petiole 30-85 cm long, (sub)glabrous, often ? bluish due to a waxy layer; trichilia absent, or if present, then separate and usually poorly developed, the brown indumentum in- termixed with dense short and rather long white hairs; stipules ca. 15-30 cm long, red to greenish, glabrous outside, hairy inside. Staminate inflorescences soli- tary or in pairs, the peduncle erect, the spikes pen- dulous; peduncle 6-10 cm long, glabrous or with sparse long white hairs, often bluish due to a waxy layer; spathe 18-30 cm long, green to yellowish or to pink, almost glabrous outside, densely (sub)sericeous inside; spikes 6-16, 9-27 X 0.3-0.6 cm, the stipes 0.6-1 cm long, glabrous; rachis with short hairs. Sta- minate flowers sessile or short-pedicellate; perianth tubular, 1.2-1.6 mm long, glabrous, the apex slightly convex to almost plane, slightly 2-lobed, smooth; fil- aments flat; anthers 0.6-0.8 mm long, appendiculate, detached at anthesis, reattached to the margins of the aperture by the appendages. Pistillate inforescences solitary or in pairs, pendulous or the peduncle initially
erect; peduncle 6-14 cm long, glabrous, often bluish due to a waxy layer; spathe ca. 15-22 cm long, the color and indumentum as in the staminate inflores- cences or glabrous inside; spikes (3 or) 4, 12-20 X 0.8-1 cm, to 35 X 1.2 cm in fruit, subsessile or with stipes to 1.5 cm long and glabrous; rachis hairy or glabrous. Pistillate flowers: perianth ca. 2 mm long, with arachnoid indumentum below the apex outside, also in the style channel inside, the apex convex to dome-shaped, smooth; style short, hairy. Fruit ellip- soid to ovoid, ca. 1.2-1.5 mm long, smooth.
Distribution (see Fig. 10.3). From Colombia (Ca- queta and Putumayo) to Peru (Amazonas and San Martin), eastern slopes of the Andes, in (cloud) forest, at 1600-2300 m.
Representative specimens examined. COLOMBIA. CAQUETA: Mun. Florencia, below Gabinete, km 47, 2000 m, 16 Oct 1993 (9 fl-fr), Franco et al. 4503 (BG). HUILA:
30 km NW of Palermo, ca. 2300 m, 10 Oct 1944 (9 fl-fr), Little 8788 (COL, F, U). PUTUMAYO: Rd. Mocoa-Sibundoy, El Mirador, 2000 m, 5 May 1994 (9 fl-fr), Franco et al. 5529 (BG).
ECUADOR. LOJA: Rd. Loja-Zamora, Quebrada Na- vidad, 30 Jan 1996 ( 9 fl), Merino et al. 4779 (LOJA). NAPO: Bermejo, valley of Rio Cosanga, 1800 m, 27 Dec 1939 (9 fl-fr), Asplund 10150 (S); Cosanga, ca. 20 km S of Baeza, ca. 1850 m, 20 Sep 1977 (d), Berg et al. 439 (BG, COL, QCA, U); rd. Baeza-Cotundo, 1990 m, 12 Aug 1975 (st), Little et al. 150 (Q, QAME, QCNE); Cant6n Quijos, be- tween Baeza & Cuyuja, 2400 m, 13 Jan 1992 (9 fl), Palacios et al. 9627 (BG, QCNE); nr. Baeza, ca. 2000 m, 23-24 May 1992 (9 fl), Valencia et al. 2306 (BG, QCA). TUNGURA- HUA: Rio Pastaza, Hda. Rio Verde Grande, ca. 1500 m, 26 Jul 1939 (9 fl-fr), Asplund 7849 (F, S), 4 Feb 1956, (6), Asplund 19205 (S); Hda. Michai, ca. 1500 m, 4 Feb 1956 (6), Asplund 19228 (S). ZAMORA-CHINCHIPE: Rd. Loja- Zamora, nr. Sabanilla, ca. 1700 m, 3 Jan 1991 (d), Berg et al. 1652 (BG, LOJA, QCA), (9 fl), Berg et al. 1653 (BG, LOJA, QCA); 18 km E of Loja, 1800 m, 3 Sep 1975 (9 fl- fr), Little et al. 213 (COL, LOJA, Q, QAME, QCNE); rd. Loja-Zamora, ca. km 13 E of the pass, before jct. with old rd., ca. 2030 m, 8 Mar 1989 (st), 0llgaard et al. 90836 (AAU, LOJA, QCA).
PERU. AMAZONAS: Prov. Bongara, Dtto. Sipabamba, Shilla, 1900 m, 5 May 1981 (9 fr), Young et al. 357 (BG, MO, NY. TEX). SAN MARTiN: Prov. Rioja, rd. Rioja-Po- macocha, nr. km 392, 1600-1700 m, 9 Dec 1997 (9 fl-fr), Berg et al. 1796 (BG, COL, MOL), (d), Berg et al. 1797 (BG, COL, MOL); rd. Pedro Ruiz-Moyobamba, km 390, Venceremos, 1770-2150 m, 5-7 Aug 1983 (st), D. N. Smith et al. 4657 (MO).
This species shows close affinities to Cecropia pastasana, as discussed under the latter. Cecropia an- dina can be distinguished from C. pastasana by the absence of trichilia (or by very poorly developed
50 FLORA NEOTROPICA
f~~~~~~~~~~~~~~~~~~~~~~~
4 ~ ~ ~ ~ ~ ~ ~ ~ ~~~- \
FIG. 9. Cecropia andina. 1. Lamina, reduced. 2. Apex of lamina and venation (Berg et al. 1652). 3. Stipules and young leaf (Berg et al. 1653). 4. Lower surface of lamina. 5. Staminate inflorescence with spathe. 6. Staminate inflorescence at anthesis. 7. Staminate flower and stamen (Berg et al. 1652). 8. Filament after detachement of anther (Berg et al. 1797). 9. Pistillate inflorescence with spathe. 10. Pistillate inflorescence at anthesis (Berg et al. 1653). 11. Pistillate flower and pistil. 12. Fruit (Franco et al. 4503).
SYSTEMATIC TREATMENT 51
X~~~~~~~~~~~~~~
4.0-~~~~~~~~~~~~~~~~~~~~
FI(G. 10. Distribution maps. 1. Cecropia angustifolia, circles (open circles for material of not entirely certain identity); C. kavanxensis, triangles; C. purpurascens, hexagons. 2. C. annulata, open circle; C. ficifolia, circles; C. telealba, triangles. 3. C. ancdina, triangles; C. distachsva, circles. 4. C. engleriana, circles; C. maxima, triangles. 5. C. hispidissima. 6. C. herthae, triangles; C. idroboi, hexagons; C. insignis, circles.
52 FLORA NEOTROPICA
ones) and usually also by the smooth leafy twigs with a bluish waxy layer. In the southern part of the species range (Zamora-Chinchipe and Peru), in adult speci- mens the incisions of the lamina run down to at most 5/10 of the distance between margin and umbilicus (or less in the lower part of the lamina) and the lower surface of the lamina is conspicuously hairy. In the (possibly disjunct) northern part of the species range, the lamina is more deeply incised and less conspicu- ously hairy beneath. In the Sibundoy valley (Putu- mayo, Colombia) and near Loja (Ecuador), C. andina extends somewhat onto the western side of the wa- tershed.
Vernacular name. Peru: chanchan (Amazonas).
3. Cecropia angulata I. W. Bailey, Bot. Gaz. (Craw- fordsville) 74: 378. t. 1-7. 1922; Veldsquez, Acta Bot. Venez. 6: 39, t. 4. 1971. Type. Guyana. Kar- tabo region, Barakara, Jul-Aug 1920 (d), Bailey 9 (lectotype, here designated: GH).
Tree, to 20 m tall. Leafy twigs 3-6 cm thick, green, puberulous to hispidulous, some of the hairs uncinate. Lamina (sub)coriaceous, ca. 55 X 55 cm to 100 X 100 cm (to 120 X 120 cm), the segments (8-)9-1 1, the free parts of upper segments obovate, the incisions down to ca. 7/10-9/10; apices obtuse to rounded; up- per surface usually smooth, sometimes scabridulous, usually subglabrous or sometimes scabrous, initially often also with sparse arachnoid indumentum; lower surface puberulous on the veins, and with sparse longer uncinate hairs, with arachnoid indumentum in the areoles and on the smaller veins, or sometimes extending to the main veins; lateral veins in the free part of the midsegment 15-20 pairs, submarginally loop-connected, unbranched or the lower ones branched; petiole 50-110 cm long, sparsely puberu- lous to hirtellous or toward the apex often densely hirtellous, or also with ? dense ( ? persisting) arach- noid indumentum; trichilia fused, the brown indu- mentum intermixed with short white hairs; stipules 12-20 cm long, color unknown, outside ? densely puberulous to hirtellous or sometimes also with arach- noid indumentum, inside (rather) sparsely hairy. Sta- minate inflorescences in pairs, the peduncle curved downward and the spikes pendulous at anthesis; pe- duncle (6-)10-15 cm long, puberulous to hispidulous or also with arachnoid indumentum; spathe 12-16 cm long, color unknown, outside sparsely to rather densely puberulous to hirtellous or also with arach- noid indumentum, inside glabrous; spikes ca. 8-20, 4-14 x 0.3-0.6 cm long, ? angular; stipes 0.8-2 cm
long, glabrous; rachis glabrous. Staminate flowers: perianth tubular, 1-2 mm long, sparsely minutely pu- berulous below the apex, the apex almost plane; an- thers 0.6-0.8 mm long, appendiculate, detached at an- thesis, reattached to the margins of the aperture by the appendages (?). Pistillate inflorescences in pairs or solitary, initially erect, in fruit pendulous; peduncle 3-14(-18) cm long, puberulous to hispidulous; spathe 6-13 cm long, the color unknown, puberulous to hir- tellous, or also with ? dense arachnoid indumentum outside, glabrous inside; spikes (2-)4(-6), (3-)7-9 X
0.4-0.6 cm, to 16 X 1.2 cm in fruit, subsessile or with stipes to 0.6 cm long and subglabrous; rachis hairy. Pistillate flowers: perianth ca. 1.5 mm long, with arachnoid indumentum below the apex, also below the style channel inside, the apex convex, glabrous and smooth or muriculate; style short; stigma comose. Fruit oblongoid, ca. 2.5 mm long, tuberculate.
Distribution (see Fig. 11.2). Guyana and north- eastern Venezuela, in forest and in secondary growth, at low elevations.
Representative specimens examined. VENEZUELA. BOLivAR: Dtto. Sifontes, Concesi6n Minera Cristina Cua- tro, 11 Aug 1985 (Y fl-fr), Aymard et al. 4146 (MO, US); Dtto. Sifontes, 3-12 km SE of Campamento CVG Las Flo- res, 16-17 Sep 1989 (Y fl-fr), Colella et al. 1444 (BG, VEN); Dtto. Roscio, San Martin de Turumban, Aug 1979 (Y fl-fr), Delascio et al. 8010 (VEN); Mun. Sucre, 2 km S of Santa Maria de Erebato, Jan 1989 (Y fl-fr), Fernandez et al. 5007 (BG, MO); Mun. Cedenio, Serrania de Maigualida, 20 km E of San Jose de Kayama, Apr 1989 (Y fl), Ferndndez 5505 (MO, NY); rd. El Dorado-Santa Elena de Uairen, be- tween km 88 & Piedra La Virgen, 30 Jan 1981 (Y fl-fr), Marcano Berti et al. 144-981 (U, VEN); Rio Caura, Cafno
La Ceiba (Cafio Cumaca), 9-26 May 1988 (d), Stergios et al. 12888 (BG, NY, VEN); Reserva Forestal del Imataca, Sep 1967 (? fl-fr), Velasquez 252 (US, VEN), (Y fl), Velds- quez 253 (VEN). DELTA AMACURO: Depto. Antonio Diaz,
nr. Curiapo, Feb 1987 (Y fl-fr), Ferndndez 3964 (BG, MO, NY, VEN); Cafno Araguao, Aug 1954 (Y fr + d), Gines 5156 (US); Rio Winikina, 16 Jan 1983 (st), Wilbert 68325
Aug 1920 (st or juv), Bailey 3 & 17 (GH); Northwest Distr., Port Kaituma, Aug 1967 (st), Davis 239 & 240 (K, NY); Northwest Distr., Waini R., 3-18 Apr 1923, De La Cruz 3752 (F, GH, MO, NY, US); Mazaruni Station, 19 Jun 1942 (9 fl-fr), Fanshawe 732 (FD 3468) (K, NY); Essequibo R., Bartica, 12 Aug 1950 (d), Fanshawe 2990 (FD 6320) (K, NY, S, U); Demerara-Mahaica Region, 1-2 km S of Loo Cr., on Linden Hwy., 2 Mar 1989 (9 fl-fr), Gillespie et al. 803 (BG); Kangaruma, 23-27 Jun 1921 (9 fl-fr), Gleason 198 (GH, NY, US); Kamarang, 21 Aug 1977 (d), Maas et al. 2589 (NY, U); Essequibo R., Kurupukari, 3 Oct 1937 (9 fl), A. C. Smith 2164 (A, G, K, MO, NY, P, S, U, US); nr. Mabura
anulta exgos;Csiva, ranle;C le, icls.3.Cbulaa 0rage;Cguoin,oe ice utsca
heagn; ucbabaa crce.0
Hill sawmill, 31 Oct 1982 ( fl-fr), Stoffers et al. 159 (MO, U); Upper Mazaruni basin, Kukui R., Suruagupuh, 12 Sep 1960 (9 fl-fr), Tillett et al. 45398 (BG, K, NY); Kamarang R., above Utschi mouth, 25 Oct 1960 (9 fl-fr), Tillett et al. 45804 (BG, NY).
This species appears to be closely related to Ce- cropia obtusa. It matches C. obtusa in many features, but can be told apart by the differences presented be- low:
C. angulata C. obtusa Stipules 12-20cm 7-12(-15) cm Segments (8-)9-11 6-8(-9) Incisions 7/10-9/10 5/10-7/10
(-8/10) Pairs lat. veins 15-19 9-15 Lamina (Sub)coriaceous Chartaceous to
subcoriaceous d peduncle (6-)10-15 cm 4-8.5 cm
54 FLORA NEOTROPICA
With the listed differences the two taxa can be dis- tinguished, although a few specimens (such as Maas et al. 7501, included in C. obtusa) tend to be more or less intermediate. The flowers of C. angulata resem- ble those of both C. obtusa and C. ficifolia. In the shape, the texture, and the smooth upper surface of the lamina, C. angulata also resembles C. distachya, from which it can be usually distinguished by the sub- marginally loop-connected lateral veins and the more or less dense arachnoid indumentum on the petiole, features only occasionally found in C. distachya. Al- though C. angulata shows strong morphological sim- ilarities to both C. distachya and C. obtusa, it should be maintained as a distinct taxon on the species level, at least for the time being, awaiting comparative field studies.
4. Cecropia angustifolia Trecul, Ann. Sci. Nat. Bot., ser. 3, 8: 83. 1847 (Aug). Type. Peru. Without lo- cality, (Y), Ruiz & Pav6n s.n. (holotype: FI-W, photograph ex FI-W in G; isotypes: B, G, OXF).
Figs. 12, 13, 14
Cecropia acutifolia Trecul, Ann. Sci. Nat. Bot., ser. 3, 8: 81. 1847 (Aug). Type. Peru. Without locality, (d + Y), Ruiz & Pav6n s.n. (FI-W, photographs seen, mixed collection consisting of leaves of C. angusti- folia and inflorescences of C. strigosa, the leaf on the sheet with the staminate inflorescence here des- ignated as the lectotype).
Cecropia digitata Klotzsch, Linnaea 20: 534. 1847 (Oct). Type. Peru. Without locality, (6), Ruiz & Pa- von s.n. (holotype: B, destroyed, photograph ex B in F, G, MO; isotypes: B, G, OXF).
Cecropia tubulosa Klotzsch, Linnaea 20: 534. 1847 (Oct). Type. Peru. Huanuco: Chinchoa or Cuchero, Ruiz & Pavon s.n. (holotype: B-n.v., destroyed; is- otypes: probably the holotype and isotypes of C. an- gustifolia).
Cecropia polyphlebia Donnell Smith, Bot. Gaz. (Craw- fordsville) 27: 442. 1899; Burger, Fieldiana, Bot. 40: 127, t. 23. 1977. Type. Costa Rica. San Jose: La Palma, 1460 m, Sep 1898 (9), Tonduz 12642 = in herb. Donnell-Smith 7411 (holotype: US; isotypes: F, G, GH, K).
Cecropia sylvicola Standley & Steyermark, Publ. Field Mus. Nat. Hist., Bot. Ser. 23: 153. 1944. Type. Gua- temala. Alta Verapaz: Rd. Tactic to the divide on rd. to Tamahu, Apr 1941 (d), Standley 90762 (holotype: F).
Cecropia coriacea Cuatrecasas, Revista Acad. Colomb. Ci. Exact. 6(21): 66. 1944; Not. Fl. Colomb. VI.
(Trab. Com. Bot. etc. Colomb.) 41. 1944. Type. Co- lombia. Huila: Between Gabinete & Andalucia, 2200-2300 m, 25 Mar 1940 (Y), Cuatrecasas 8697 (holotype: COL; isotype: F).
Cecropia strigilosa Cuatrecasas, Revista Acad. Colomb. Ci. Exact. 6(22/23): 282. 1945. Type. Colombia. Valle: Rio Calima, El Cairo, between Darien & Me- diacanoa, 1700 m, 6-7 Jan 1943 (5), Cuatrecasas 13872 (COL), (9) Cuatrecasas 13939 (not traced), Cuatrecasas 13872 here designated as lectotype
(COL). Cecropia caucana Cuatrecasas, Revista Acad. Colomb.
Cecropia danielis Cuatrecasas, Revista Acad. Colomb. Ci. Exact. 6(22/23): 290. 1945. Type. Colombia. An- tioquia: San Antonio del Prado, 1870 m, 7 Aug 1943 (5), Hno. Daniel 2843 (holotype: COL; isotypes: F, GH, MEDEL).
Cecropia palmatisecta Cuatrecasas, Revista Acad. Col- omb. Ci. Exact. 6(22/23): 297. 1945; Velasquez, Acta Bot. Venez. 6: 49, t. 10. 1971. Syntypes. Colombia. Caqueta: Sucre, La Portada, 1200-1350 m, 5 Apr 1940 (5), Cuatrecasas 9157 (COL), (9), Cuatre- casas 9158 (not traced), Cuatrecasas 9157, here des- ignated as lectotype (COL).
Cecropia hachensis Cuatrecasas, Revista Acad. Colomb. Ci. Excact. 6(22/23): 298. 1945; Berg & Franco Ros- selli, Fl. Ecuador 48: 23. 1993. Type. Colombia. Ca- quetd: Rio Hacha, Caj6n de Pulido, 1700-1750 m, 26 Mar 1940 (9), Cuatrecasas 8782 (holotype: COL; isotype: F).
Cecropia moniquirana Cuatrecasas, Revista Acad. Col- omb. Ci. Exact. 6(22/23): 298. 1945. Type. Colom- bia. Boyaca: Nr. Moniquira, 1580 m, 25 Feb 1940 (9), Perez Arbelaez & Cuatrecasas 8135 (holotype: COL; isotypes: F, US).
Cecropia philipsonii Cuatrecasas, Revista Acad. Col- omb. Ci. Exact. 9(36/37): 382. 1956. Type. Colom- bia. Meta: Sierra de la Macarena, North Ridge, 29 Dec 1949 (5), Philipson & Idrobo 1983 (holotype: US; isotypes: BM, COL).
Cecropia villosa C. C. Berg & P. Franco, Fl. Ecuador 48: 53. 1993. Type. Ecuador. Zamora-Chinchipe: Rd. Loja-Zamora, nr. Sabanilla, 1600-1700 m, 3 Jan 1991 (9 fl-fr), Berg et al. 1651 (holotype: QCA; isotypes: BG, GB, LOJA).
Tree, to 20(-25) m tall. Leafy twigs (1-)2-5 cm
thick, green to brownish to often (turning) blackish, hispidulous to hirtellous to strigose to subvelutinous or to villous, usually also with dense brown to black- ish pluricellular hairs, sometimes also with arachnoid indumentum; internodes with sparse (whitish) pith or sometimes partly to entirely filled with brown pith. Lamina chartaceous to coriaceous, ca. (20 X 20 cm
to) 45 X 45 cm to 75 X 75 cm (to 90 X 90 cm), the
SYSTEMATIC TREATMENT 55
FIG. 12. Cecropia angustifolia. 1. Lamina, reduced (Berg 1279). 2. Lamina (Berg 1248). 3. Apex of lamina and venation. 4. Stipules and base of petiole with trichilium (Berg et al. 44J). 5. Stipules and young leaf (Berg 1281). 6. Pair of staminate inflorescences with spathes and base of petiole with trichilum (Hurtado et al. 2581). 7. Staminate inflorescence at anthesis and base of petiole with trichilium (Gentry et al. 63245). 8. Staminate flower and stamen (Berg et al. 1633). 9. Pistillate inflorescence with spathe (Dodson et a!. 10541). 10. Pistillate inflorescence at anthesis and base of petiole with trichilium (Berg et a!. 1204). 11. Pistillate flower and pistil (Berg et a!. 1632). 12. Fruit (Franco et a!. 4475).
56 FLORA NEOTROPICA
/4~~~~~~~~~~~~~~~~~~~~~~~~~~~4
I /~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~I
FIG. 13. Cecropia angustifolia. 1. Lamina, reduced. 2. Apex of lamina and venation. 3. Stipules, young leaf, staminate inflorescences with spathes, and base of petiole with trichilium. 4. Pair of staminate inflorescences with spathes and just at
anthesis. 5. Staminate flower. 6. Stamen and filament (Berg et al. 1753). 7. Pair of pistillate inflorescences with spathes and
base of petiole with trichilum. 8. Pairs of pistillate inflorescences at anthesis and base of petiole with trichilium. 9. Pistillate
flower and style. 10. Fruit (Berg et al. 1750). From Caldasia 24: 232. 2002.
SYSTEMATIC TREATMENT 57
FIG. 14. Cecropia angustifolia. 1. Lamina, reduced. 2. Apex of lamina and venation. 3. Stipules. 4. Staminate inflorescence with spathe and young leaf. 5. Staminate flower and stamen (Berg et al. 1650). 6. Pair of pistillate inflorescences at anthesis. 7. Pistillate flower. 8. Fruit (Berg et al. 1651). (From Fl. Ecuador 48: 54. 1993, modified.)
58 FLORA NEOTROPICA
segments (8-)10-14, in young leaves often reflexed, the free parts of upper segments (ob)lanceolate to ob- long to subobovate, the incisions down to (5/10-) 6/10-9/10 (or to near the petiole); apices obtuse to rounded or subacute to short-acuminate; upper sur- face scabrous or smooth, densely to sparsely hispi- dulous or puberulous or partly to hirtellous or to (sub)strigillose, also with ? dense brown pluricel- lular hairs and sometimes sparse arachnoid indumen- tum, the "umbilicus" sometimes villous; lower sur- face sparsely to densely (minutely) puberulous (with straight to uncinate hairs) to hirtellous to subtomen- tose or to subhirsute on the (main) veins, with arach- noid indumentum confined to the areoles, extending to the smaller veins, or (almost) absent; lateral veins in the free part of the midsegment (12-)15-35(-43) pairs, submarginally or marginally loop-connected, branched or unbranched; petiole 20-70 cm long, sparsely to densely puberulous to hirtellous to sub- hirsute or to villous, or sometimes also with sparse to dense arachnoid indumentum; trichilia fused, the brown indumentum intermixed with short (or some- times rather long) white to brownish (unicellular) hairs; stipules 6-25(-30) cm long, caducous or sub- persistent, green to yellowish, reddish to purplish, brownish or black, sparsely to densely white to brownish puberulous to hirtellous or partly to largely subhirsute to sericeous to subvelutinous or to villous, also with ? dense brown pluricellular hairs outside, sparsely to densely white- to pale brown-sericeous (or glabrous) inside. Staminate inflorescences in pairs, erect, often subtended by subpersistent bracts, to 5 cm long; peduncle 1-8(-12) cm long, puberulous to sub- hispid to hirtellous to subhirsute or to subvelutinous and often (initially) with dense brown pluricellular hairs and often also sparse arachnoid indumentum, sometimes subglabrous; spathe 4-14 cm long, green- ish to yellowish to brownish or reddish to purplish, densely white- to brownish-puberulous to -hirtellous or partly to -subhirsute to -strigose or to villous and with dense brown pluricellular hairs or also with sparse to rather dense arachnoid indumentum outside, sparsely to densely (sub)sericeous or glabrous inside; spikes 10-25(-50), (l.5-)3-11(-13) X 0.3-0.8 cm, sessile or with stipes to 0.7(-1.2) cm long, (sub)glabrous to puberulous to hirtellous; rachis hairy (or glabrous). Staminateflowers: perianth tubular, 1- 2 mm long, with short and stiff hairs on the ribs, arachnoid indumentum below the apex, or glabrous, the apex plane to convex, smooth or muriculate, the aperture circular or slit-shaped; filaments flat or ?
swollen; anthers 0.5-0.9 mm long, appendiculate or not, detached and reattached to the margins of the
aperture by the appendages or remaining attached to the filament at anthesis. Pistillate inflorescences in pairs, erect, often subtended by subpersistent bracts, to 3 cm long; peduncle 1-8(-12) cm long, the indu- mentum as in the staminate inflorescence; spathe 3- 6(-8) cm long, color and indumentum as in the sta- minate inflorescence; spikes 3-8(-12), 1-7 X 0.4-0.8 cm, to 17 X 0.8-2 cm in fruit, often broadened toward the base, sessile; rachis hairy (or glabrous). Pistillate
flowers: perianth 1.5-2.5 mm long, with arachnoid indumentum below the apex outside, also in the lower part of and/or below the style channel inside, the apex convex to almost plane, muriculate to minutely his- pidulous or glabrous, the aperture circular or slit- shaped; style rather short; stigma comose. Fruit ellip- soid to oblongoid to ovoid 1.5-2.5 mm long, tuberculate or smooth.
Distribution (see Fig. 10.1). From Guatemala (or probably Mexico: Puebla) to the coastal mountain range of Venezuela and through the Andes (and some adjacent low mountain ranges) to Bolivia, in wet montane or wet to relatively dry submontane forest, at ca. (400-)800-2400 m.
Representative specimens examined. MEXICO. PUEBLA: Nr. Cuerrero, 15 mi E of Tezuitlan, 16 Jul 1947 (6 + 9 fl-fr), Converse s.n. (US); rd. Tlapacoyan (Veracruz) to San Jose Acateno, 1000 m, 22 Oct 1968 (6), Pennington et al. 9273 (A, K, NY).
GUATEMALA. ALTA VERAPAZ: E of Tactic, rd. to Ta- mahu, 1500-1550 m, 9 Apr 1939 (st), Standley 71248 (F); rd. between Tactic & the divide on rd. to Tamahu, 1500- 1600 m, 1-7 Apr 1941 (6), Standley 90576 (F). HUEHUE-
TENAGO: Nr. Maxbal, 17 mi N of Barillas, Sierra de los Cuchumatanes, 1500 m, 15-16 Jul 1942 (juv), Steyermark 48888 (US).
HONDURAS. CORTES: Parque Nacional Cusuco, nr. Centro de Visitantes, 18 km W of San Pedro Sula, 1620 m, 21 Mar 1993 (6), Dario 355 (BG, EAP). OLANCHO: La Muralla, ca. 10 km N of La Uni6n, ca. 1350 m, 26 Oct 1996 (6), Berg 1806 (EAP).
NICARAGUA. JINOTEGA: Macizos de Penias Blancas, nr. Finca El Cielo, 1200-1400 m, 13-18 Jan 1979 (3 fl-fr), Stevens 11688 (BG, MO); rd. Matagalpa-Jinotega, ca. 4 km NW of Aranjuez, ca. 1400 m, 8 Dec 1983 (9 fl), Stevens 22520 (MO, NY, TEX). MATAGALPA: Between Disparate de Potter & Arajuez, 1300 m, 12 Jan 1963 (d), L. 0. Wil- liams et al. 23663 (F, G).
COSTA RICA. ALAJUELA: Rio Petias Blancas, 1540- 1575 m, 18 Jun 1985 (9 fr), Haber et al. 1658 (MO); 2 km N of La Balsa de San Ram6n, 800 m, 13 Feb 1977 (d + 9 fl-fr), Lent 4101 (AAU, MO, NY). CARTAGO: 10 km S of Tapantf, above Rio Grande de Orosi, 1600 m, 1 Sep 1971 (9 fl-fr), Burger et al. 8428 (BM, EAP, G, MO, NY, PMA); Cerro Dean, 3 km E of Cachi, 1400 m, 23 Apr 1969 (9 fl), Lent 1614 (S); 0.5 km S of Petias Blancas de Cachi, 1400
SYSTEMATIC TREATMENT 59
m, 5 Sep 1971 (Y fl-fr), Lent 2120 (BM, EAP, G); S limit of Monumento Nacional Guayabo, sector Las Ventanas, 1100 m, 7 Jul 1992 (5), Rivera 1934 (K); 15 km NW of Turrialba, ca. 1000 m, 13 Aug 1967 (Y fr), Taylor 4393 (NY). HEREDIA: 3 km W of Vara Blanca, 1750 m, 8 Aug 1971 (? fl-fr), Lent 2052 (GH, U). PUNTARENAS: Cerro Pando, above Rio Cot6n & Rio Negro, 1000-1800 m, 19- 21 Feb 1982 (st), Barringer et al. 1607 (LL); Monteverde, 1500 m, 15 Apr 1981 (5), Haber 455 (MO). SAN JOSE: Cord. de Talamanca, between Eusa Mato & El Empalme, 2200-2400 m, 3 Mar 1966 (9 fl), Blum et al. 2225 (MO); Cerro El Espino (Alto Mata de Cania-Azahar), Patarra, 1600-1800 m, 13 Nov 1983 (5), Chacon et al. 1563 (BG, MO); Las Tablas, Rfo Cotoncito, 10 Dec 1983 (5), Chacon et al. 1816 (BG, COL, MO, NY); Rfo Para Blanco, 1500 m, 4 Feb 1973 (5), Lent 3154 (COL, U).
PANAMA. CHIRIQUI: Cerro Horqueta, ca. 2300 m, 24 Jul 1966 (2 fl-fr), Blum et al. 2600 (MO); Cerro Colorado, above 1000 m, 9 Aug 1984 ( 9 fl-fr), Schmalzel et al. 2026 (BG).
COLOMBIA. ANTIOQUIA: Mun. La Ceja, 3 km N of La Ceja, 2400 m, 21 Jan 1994 (5), Callejas et al. 11111 (BG, COL, HUA); Mun. Medellin, Santa Elena, 1700 m, 24 Mar 1994 (5), Franco et al. 4577 (BG, HUA), (9 fl-fr), Franco et al. 4578 (BG, HUA), (5), Franco et al. 4580 (BG, HUA), (5), Franco et al. 4582 (BG, HUA); Mun. Cocormn, Vrda. Viao, 1950 m, 26 Mar 1994 (5), Franco et al. 4591 (BG, HUA); Mun. Frontino, Cgto. Nutibara, rd. Nutibara- La Blanquita, 1700 m, 4 Mar 1995 (9 fl-fr), Franco et al. 5547 (BG, COL, HUA, LP, MO, US); Mun. Frontino, rd. Nutibara-Frontino, km 14, 1750 m, 5 Mar 1995 (d), Franco et al. 5564 (COL, HUA, LP, MO, US); Mun. Anorf, Vrda. El Roble, 1400 m, 8 Mar 1995 (5), Franco et al. 5568 (BG, COL, HUA, LP, MO, US); Mun. Anorf, rd. Anorf-El Roble, 1600 m, 8 Mar 1995 (9 fl-fr), Franco et al. 5570 (BG, COL, HA, LP, MO, NY, QCA). BOYACA: Mun. Pajarito, below Corinto, 1200 m, 29 Jan 1995 (9 fl-fr), Franco et al. 4641 (BG, COL, HUA, MO, US); between Moniquira & Gach- antivd, 1200-1600 m, 14 May 1996 (st), Rangel et al. 13234 (COL). CALDAS: Manizales, 2153 m, Jul 1938 (5), Duque- Jaramillo 4048 (COL, NY). CAQUETA: Mun. Florencia, be- low Gabinete, km 17, 2200 m, 15 Oct 1993 (5), Franco et al. 4493 (BG), km 45, 2150 m, 15 Oct 1993 (9 fl-fr), Franco et al. 4495 (BG, COL), 15 Oct 1993 (5), Franco et al. 4496 (BG); Mun. Florencia, rd. Florencia-Gabinete, km 49, 1900 m, 16 Oct 1993 (5), Franco et al. 4502 (COL), ca. 1050 m, 18 Oct 1993 (9 fl-fr), Franco et al. 4513 (BG, COL), ca. 1750 m, 18 Oct 1993 (9 fl-fr), Franco et al. 4515 (BG, COL); trail Las Guacamayas-Ramos, km 59, ca. 1600 m, 29 Apr 1944 (9 fl-fr), Little 7763 (COL, US). CASANARE: Nr. Sacama, 1100 m, 27 Jan 1995 (5), Franco et al. 4637 (COL); Sacama, Vrda. Guavarin, 1750 m, 27 Jan 1995 (9 fl-fr), Franco et al. 4638 (BG, COL, HUA, MO, US). CAUCA: Between Popayan & Piendam6, 1800 m, 31 Dec 1942 (9 fl), Cuatrecasas 13855 (F, US, VALLE); valley of Rio Palac6, rd. to Totor6, Quebrada de Palac6, 1820-1960 m, 15 Oct 1961 (5), Cuatrecasas et al. 26487 (COL, P, US), (9 fl-fr), Cuatrecasas et al. 26491 (COL, P, US); rd. La Plata-Purace, 12 km from Santa Cecilia, Reserva Meren-
berg, 15 Jul 1977 (6), Idrobo 8753 (COL); Rio San Joaquin, 1400-1500 m, 29-30 Jun 1922 (6), Killip 7874 (GH, K, NY, US); Popayan, 21 Jul 1960 (9 fl), Schultes 22508 (COL, GH); El Tambo, 1700 m, 8 Oct 1939 (9 fl), Sneidern 2218 (A, LL, MICH, NY, S, TEX, US). CESAR: Sierra de Perijd, E of Manaure, El Podrido, 1550-1600 m, 16 Nov 1959 (9 fl-fr), Cuatrecasas et al. 25393 (COL, US), 17 Nov 1959 (6), Cuatrecasas et al. 25420 (COL, US); Mun. Manaure, rd. Manaure-El Cinco, 1850 m, 10 Nov 1993 (d), Franco et al. 4522 (BG), (9 fl), Franco et al. 4523 (BG). CHOC6: Mun. San Jose del Palmar, Vrda. El Sinai, 1650 m, 19 Mar 1994 (9 fl-fr), Franco et al. 4562 (BG, HUA); Emisora La Sirena, 3 km W of La Mansa, 2300-2400 m, 16 Jan 1979 (9), Gentry et al. 24195 (BG, COL, MO). CUNDINA- MARCA: Mun. Tena, Vrda. El Rosario, rd. to Laguna de Pe- dro Palo, 2100 m, 7 Aug 1984 (9 fl), Barrera 228 (COL); Granada, Subia, 2225 m, 12 Mar 1993 (6), Franco et al. 4343 (BG, COL); Guayabetal, rd. Guayabetal-Villavicen- cio, 1030-1150 m, 29 May 1993 (9 fl-fr), Franco etal. 4456 (BG) & 4457 (BG); rd. Caqueza-Puente Quetame, 13 Mar 1985 (9 fl-fr), Galeano et al. 642 & 645 (COL); rd. Bogota- Laguna de Pedro Palo, ca. 2100 m, 18 Mar 1985 (9 fl-fr), Galeano et al. 675 (COL). HUILA: Finca Merenberg, E of Volcan Purac6, nr. Cauca border, 2290 m, 3 Apr 1986 (st), Gentry et al. 53945 (BG). MAGDALENA: Sierra Nevada de Santa Marta, above Minca, 1680-1690 m, 16 Jan 1989 (9 fl), Gentry et al. 64746 (MO); Sierra Nevada de Santa Marta, Finca Los Arroyitos, 1600-1700 m, 4 Oct 1972 (d), Kirk- bride 2381 (BG, COL, NY, US); Mun. Santa Marta, San Lorenzo, 2240 m, 14-15 Sep 1969 (st), Lozano C. 1078 (COL). META: Mun. Villavicencio, rd. to Guayabetal, 900- 1000 m, 13 Feb 1995 (9 fl-fr), Franco et al. 4677 (BG, COL, HUA, NY, US); Sierra de La Macarena, Macizo Ren- jifo, 1300-1900 m, 6-20 Feb 1951 (9 fr), Idrobo et al. 1092 (COL, LE, U, US), (6), Idrobo et al. 1093 (COL, GH, US); Sierra de La Macarena, Central Mtns., North Ridge, 29 Dec 1949 (9 fl-fr), Philipson et al. 1982 (BM, COL, US). NA- RINO: Mun. Ricaurte, Reserva Natural La Planada, 1800 m, May 1989 (9 fr), Beltran 13 (MO, PSO); Mun. Ricaurte, nr. Ricaurte, 1400 m, 8 Oct 1993 (6), Franco et al. 4465 (BG); Mun. Ricaurte, Reserva La Planada, 1800 m, 8 Oct 1993 (st), Franco et al. 4468 (BG); Mun. Ricaurte, nr. Ri- caurte, 1400 m, 8 Oct 1993 (9 fr), Franco et al. 4475 (BG); Samaniego, 1535 m, Aug 1964 (6), Mora 0. 3185 (PSO); Mun. San Francisco, rd. Pasto-Mocoa, between El Mirador & San Francisco, 1500-2000 m, 27 Nov 1967 (9 fl-fr), Mora 0. 4474 (COL); Mun. Leiva, nr. Leiva, 1500 m, 1 Sep 1991 (6), Ramfrez P 4167 (COL). NORTE DE SANTANDER:
Ocafna region, nr. Apasica, Quebrada del Guarumal, 1900- 2000 m, 26 Sep 1969 (6), Cuatrecasas et al. 27950 (COL, NY, P, US); Ocafia region, Cercanfas de Apasica, La Playa, 1700 m, 26 Sep 1969 (9 fl-fr), Cuatrecasas et al. 27953 (COL, NY, P, US); Mun. Toledo, Vrda. El Encanto, 1800 m, 25 Jan 1995 (6), Franco et al. 4625 (BG, COL, HUA, US). PUTUMAYO: Mocoa, Vrda. La Campucana, Finca La Mar- iposa, 1350 m, 20 Apr 1994 (d), Franco et al. 5186 (BG, COL), (9 fl-fr), Franco et al. 5206 (BG, COL), 28 Apr 1994 (st), Franco et al. 5458 (BG), (6), Franco et al. 5497 (BG), (9 fl-fr), Franco et al. 5498 (BG); Mun. Mocoa, between
60 FLORA NEOTROPICA
La Campucana & San Antonio, 1000 m, 2 May 1994 (5), Franco et al. 5515 (BG); Mun. Mocoa, Cgto. El Pepino, rd. El Pepino-Sibundoy, Vrda. Las Mesas, 1250 m, 3-4 May 1994 (5), Franco et al. 5528 (COL); rd. Mocoa-Pitalito, Sajonia, 1600 m, 5 May 1994 (9 fl-fr), Franco et al. 5536 (BG); rd. San Juan-Pitalito, between Suacira & Santo Do- mingo, 2000 m, 5 May 1994 (9 fl-fr), Franco et al. 5541 (BG). QUINDIO: Mun. Montenegro, Vrda. Morelia Baja, 1240 m, 21 Apr 1991 (9 fl), Agudelo et al. 1429 (COL); Mun. Pijao, rd. Pijao-Chili, 2800 m, 13 May 1990 (5), W- lez et al. 1693 (COL); Mun. Pijad, Vrda. Carniceres, 1840 m, 24 Aug 1993 (9 fr), Wlez et al. 3370 (COL). RISAR-
ALDA: Mun. Santa Rosa, Vrda. El Crucero, Hda. La Colina, 1725 m, 24 Jul 1980 (5), Idrobo et al. 9975 (COL, U). SANTANDER: Mun. Oiba, Vrda. El Guayabito, ca. 1400 m, 22 Jan 1995 (9 fl-fr), Franco et al. 4605 (BG, COL, HUA, US); nr. Aratoca, ca. 1800 m, 22 Jan 1995 (9 fl-fr), Franco et al. 4606 (BG, COL, HUA, US). TOLIMA: Mun. Santa Isabel, Finca La Cima, 1860 m, 6 Aug 1980 (9 fl), Idrobo et al. 10680 (COL). VALLE: Rio Sanquinini, La Laguna, 1250-1400 m, 10-20 Dec 1943 (5), Cuatrecasas 15540 (F, VALLE), (9 fr), Cuatrecasas 15550 (COL, F, US); Rio Cali, confluence of Rio Pichindecito & Rio Pichinde, 1580-1650 m, 7 Nov 1944 (9 fl-fr), Cuatrecasas 18785 (COL, F, VALLE), (9), Cuatrecasas 18786 (F, US, VALLE), (9 fl- fr), Cuatrecasas 18787 (F, US, VALLE); Rio Pichinde, Que- brada de Juntas, 2100 m, 23 Sep 1946 (st), Cuatrecasas 22015 (MO, VALLE); Monte La Guarida, above La Car- bonera, between Las Brisas & Alban, 1950-2000 m, 16-24 Oct 1946 (5), Cuatrecasas 22511 (F, US); Quebrada del Tigre, between Las Brisas & La Marina, 1670-1730 m, 28 Oct 1946 (9 fr), Cuatrecasas 22682 (F, US, VALLE); Alto del Dinde, between Cartago & Alcala, 1200-1260 m, 16 Nov 1946 (5), Cuatrecasas 22891 (F, US, VALLE), (9 fr), Cuatrecasas 22902 (F, US); Mun. Argelia, rd. El Cedro-La Argelia-La Aurora, 1900 m, 11 Oct 1983 (5), Devia 313 (COL, MO, TULV); Mun. Calima, Vrda. Campo Alegre, Finca Los Bambues, 900 m, 5 Oct 1983 (9 fl), Devia 406 (BG, MO, TULV); Reserva Forestal de Yotoco, 1200-1900 m, Jul 1993 (5), Escobar M. s.n. (VALLE); Mun. Trujillo, rd. Trujillo-Rio Frio, km 1, 1260 m, 11 Sep 1993 (9 fl-fr), Franco et a!. 4177 (BG), (5), Franco et al. 44811 (BG); Mun. Trujillo, rd. Trujillo, Naranjal, km 20, Rfo Sanquinini, 1 I Sep 1993 (d), Franco et al. 4488 (BG); Mun. Yotoco, rd. Media Canoa-Dari6n, El Caney, 1500 m, 15 Mar 1994 (5), Franco et al. 4525 (BG), (9 fl-fr), Franco et al. 4526 (BG), (d), Franco et al. 4527 (BG); 1 km E of Bitaco, ca. 1500 m, 16 Nov 1963 (9 fr), Hutchinson et al. 3009 (F, NY, US), (d), Hutchinson et al. 3028 (COL, K, MO, NY, P, US), (d), Hutchinson et al. 3035 (COL, MICH, NY, US); Monte Frio, Yanaconas, 1700-1850 m, 27 Feb-i Mar 1939 (5), Killip et al. 33712 (US); Mun. Darien, Vrda. El Vergel, 1550, 20 Jun 1970 (9 fl-fr), Mahecha 281 (UDBC); Mun. Darien, Alto Calima, 1970 (9 fl-fr), Mahecha s.n. (UDBC 5780); rd. Cali-Pichinde, 1700 m, 15 Nov-6 Dec 1979 (d), Rooden et al. 251 (BG, MO, NY, U).
VENEZUELA. ARAGUA: Parque Nacional Henri Pit- tier, Rancho Grande, 1400 m, 10 Aug 1953 (9 fr), Little 15437 (NY, VEN); rd. El Pauji-El Soccoro, S of El Consejo,
1350-1400 m, 16 Jul 1979 (9 fl-fr), Steyermark et al. 118157 (U, VEN); nr. Estaci6n Rancho Grande, 800-1300 m, Aug 1966 (6), Veldsquez 92 (US, VEN, (9 fl-fr), Velds- quez 93 (US, VEN), 26 Dec 1975 (6), Wood 429 (VEN). BARINAS: Mun. Pedreza, Alto del Aguada, 1200-1300 m, 18 Feb 1955 (9 fl), Bernardi 1966 (NY); Altamira-Calderas, 850-1600 m, 1 Jun 1957 (9 fr), Bernardi 6878 (NY). DIs- TRITO FEDERAL: Nr. Tovar, 1860 (6), Fendler s.n. (G); Parque Nacional El Avila, Quebrada Quintero, Altamira, 6 Sep 1975 (9 fl-fr), Manara s.n. (VEN); Cerro Naiguata, Quebrada Besenilla, 900-1000 m, 3 Nov 1963 (6), Stey- ermark 91899 (U); between rd. Tovar-Junquito & Hda. El Lim6n, 1300-1500 m, 12 Feb 1966 (9 fl), Steyermark 94778 (NY, US, VEN); Parque Nacional Avila, nr. Estaci6n Los Venados, 900-1000 m, Mar 1967 (d), Veldsquez 244 (US, VEN). FALCON: Dtto. Bolivar, Sierra de San Luis, Cerro Galicia, 1500 m, 29 Mar 1984 (9 fr), Plowman et al. 13448 (BG, K, F, MO, NY), (6), Plowman et al. 13449 (BG, F, K, MO, NY). MERIDA: Mun. El Carrizal, El Rinc6n, 1400-1600 m, 23 Jun 1954 (9 fl-fr), Bernardi 1295 (F, NY); Mesa Bolivar, 1450 m, I Nov 1959 (9 fl-fr), Bernardi 7629 (VEN); Mun. Zerpa, Dtto. Andres Bello, La Azulita, 1500 m, 31 Oct 1979 (9 fl-fr), Marcano Berti et al. 280-979 (U); above La Azulita, 1375-1920 m, 25 Apr 1944 (st), Steyer- mark 56101 (VEN); nr. Merida, Feb 1970 (9 fr), Veldsquez 2093 (US). MIRANDA: Los Guayabitos, SE of Baruta, 11 Oct 1969 (9 fl-fr), Steyermark 91644 (NY, US); Los Guay- abitos, 1200-1400 m, Nov 1967 (6), Veldsquez 249 (VEN), (9 fl-fr), Veldsquez 250 (VEN). SucRE: Peninsula de Paria, NW of Irapa, 1060-1270 m, 3 Mar 1966 (9 fl), Steyermark 94971 (F, US); Peninsula de Paria, Dtto. Cagigal, between El Paujil & El Brasil, 850-890 m, 21 Feb 1980 (9 fr), Stey- ermark et al. 121434 (MO, NY, U, VEN). YARACUY: Sierra Aroa, Cerro Tigre, 10 km E of Aroa, 1000 m, 3 Apr 1980 (9 fl-fr), Liesner et al. 9987 (MO, U, VEN). ZULIA: Dtto. Mara, nr. Puesto El Bosque, 1450-1600 m, 10-15 Dec 1982 (9 fl-fr), Bunting et al. 12087 (MO, U).
ECUADOR. CARCHI: Nr. Guadalupe, RR Ibarra-San Lorenzo, 900 m, 8 Feb 1981 (9 fl-fr), Berg 1248 (AAU, BG, COL, MO, QCA, U); ca. 6 km above Maldonado, 2275 m, 23 May 1993 (st), Boyle et al. 1916 (BG). COTOPAXI: Rd. Quevedo-Latacunga, nr. Esperanza (El Tigre), 1500 m, 16 Feb 1981 (6), Berg 1263 (BG, COL, NY, QCA, U); rd. Quevedo-Latacunga, nr. El Palmar, ca. 800 m, 17 Feb 1981 (9 fl-fr), Berg 1279 (AAU, BG, COL, MO, QCA, U), (6), Berg 1281 (AAU, BG, COL, GB, MO, NY, QCA, U). EL
ORO: Rd. Pifias-Santa Rosa, km 12.5, ca. 600 m, 7 Oct 1979 (st), Dodson et al. 8941 (MO), 7 Oct 1979 (6), Dodson et al. 8953 (MO, QCNE); Viscaya, nr. Zaruma, 1250-1500 m, 24 Sep 1980 (9 fl-fr), Dodson et al. 10541 (MO, QCNE). IMBABURA: Between Parambas & Collapi, 1000 m, 4 Jul 1949 (6), Acosta S. 12777 (F). LOJA: Nr. Alamor, ca. 1250 m, 29 Dec 1997 (9 fl-fr), Lozano et al. 901 (BG, LOJA). Los Rios/PIcHINCHA: El Centinela, at crest of Montafias de Ila, rd. Patricia Pilar-24 de Mayo, km 12, 600 m, 27 Nov 1978 (st), Dodson et al. 7305 (BG, QCNE), (6), Dodson et al. 7335 (BG, QCNE). MANABi: Machalilla National Park, San Sebastian, 550 m, 21 Jan 1991 (6), Gentry et al. 72522 (MO, QCNE). MORONA-SANTIAGO: Rd. Lim6n-Quenca,
SYSTEMATIC TREATMENT 61
ca. 15 km, Plan del Milagro, 1700 m, 30 Jan 1981 (9 fl-fr), Berg 1233 (BG, COL, QCA, U); rd. Gualaquiza-San Juan Bosco, ca. km 13, ca, 1300 m, 16 Feb 1994 (9 fl-fr), Berg et al. 1692 (BG, QCA). NAPO: Cant6n Baeza, Bermeja, 2200-2650 m, 14 Dec 1993 (9 fl), Alvarez 881 (QCA, QCNE); Cant6n Archidona rd. Hollin-Loreto, km 32, Chal- lua Yacu, 1200 m, 14-16 Sep 1989 (9 fl-fr), Hurtado et al. 2457 (MO, QCNE), 14-16 Sep 1989 (d), Hurtado et al. 2581 (BG, MO, QCNE); El Codo Alto, 1300, 12 Sep 1990 (i), Jaramillo et al. 12580 (QCA); Cant6n El Chaco, Rio Quijos, Finca El Ave Brava, 1800-1900 m, 7-10 Sep 1990 (i), Palacios 5363 (BG, QCNE, MO). PASTAZA: Nr. Mera, Rio Mangayuca, ca. 1100 m, 9 Dec 1955 (9 fl-fr), Asplund 18745 (S); Upper Rio Pastaza, San Francisco, ca. 1100 m, 4 Feb 1956 (d), Asplund 19215 (S); Tnte. Hugo Ortiz, 20 km NNE of Puyo, 950 m, 30 Jan 1981 (9 fl), Berg et al. 1117 (AAU, BG, COL, GB, MO, QCA, U); 2 km NE of Mera, 1100 m, 27 Feb-19 Mar 1985 (9 fl-fr), Neill et al. 5994 (BG, MO, QAME, QCNE). PICHINCHA: Nr. Tandapi, 1350-1800 m, 11 Sep 1977 (6), Berg et al. 417 (BG, COL, MO, NY, QCA, U), (9 fl-fr), Berg et al. 418 (AAU, BG, COL, MO, QCA, U); old rd. Quito-Santo Domingo de los Colorados, km 59, nr. Palmeras, ca. 1800 m, 14 Dec 1990 (9 fl-fr), Berg et al. 1646 (BG, QCA); old rd. Quito-Santo Domingo de los Colorados, km 59, Reserva Floristica- Ecol6gica Rio Guajalito, 1800-2000 m, 24 Aug 1985 (9 fl- fr), Jaramillo et al. 8145 (BG, G, GB, GH, K, MO, NY, QCA, US); 5-6 km above Nanegalito, 1700 m, 21 Jul 1980 (9 fl-fr), Holm-Nielsen et al. 24409 (AAU). SUCUMBiOS: Rd. Baeza-Lago Agrio, nr. Santa Rosa, 1500-1600 m, 21 Sep 1977 (d), Berg et al. 441 (BG, COL, QCA); Cant6n Gonzalo Pizarro, Campo Bermejo 6 Norte, 30 km NW of Lago Agrio, 1050 m, 31 Mar 1990 (st), Cerc'n et al. 9418 (BG); 3.5 km N of Borja, 1850 m, 20 Sep 1980 (d), Holm- Nielsen et al. 26350 (AAU). TUNGURAHUA: Rd. Bafios- Puyo, between Rio Verde & Rio Negro, ca. 1400 m, 10 Jan 1981 (9 fl-fr), Berg 1204 (BG, COL, QCA); Banios, Los Llanganates, 18 km from Topo, 1800-2000, 9 Mar 1995 (9 fl-fr), H. Vargas et al. 355 (BG, QCNE). ZAMORA-
CHINCHIPE: Rd. Loja-Zamora, nr. San Ram6n (hydro- electric station), ca. 1900 m, 3 Jan 1991 (d), Berg et al. 1650 (BG, LOJA, QCA); rd. Loja-Zamora, km 28, 1900 m, 15 Feb 1994 (9 fl), Berg et al. 1688 (BG); rd. Loja-Zamora, nr. Sabanilla, 1600-1700 m, 4 Sep 1975 (9 fl-fr), Little et al. 227 (COL, LOJA, Q).
PERU. CAJAMARCA: La Palma, 10 km NW of Chirinos, 1780 m, 5 Feb 1988 (st), Gentry et al. 61217 (BG, MO); Jaena, E.P.S. El Champe, 1500-1800 m, 3 May 1980 (9 fr), Rios T 100 (MOL); Prov. Cutervo, Quebrada de Santa Rosa, W of rd. San Andres-Santo Tomas, 2400 m, 4 Nov 1991 (J), Sanchez V et al. 6060 (F). Cuzco: Prov. Paucartambo, rd. Pilcopata-Paucartambo, km 17, 900 m, 2 Aug 1988 (9
fl-fr), Berg et al. 1630 (BG, COL, Centro de Medicina An- dina, Cuzco); Prov. Paucartambo, rd. Pilcopata-Paucar- tambo, ca. 1600 m, 2 Aug 1988 (9 fl-fr), Berg et al. 1632 (BG, COL, Centro de Medicina Andina, Cuzco); Prov. Pau- cartambo, rd. Pilcopata-Paucartambo, ca. 1600 m, 2 Aug 1988 (d), Berg et al. 1633 (BG, COL, Centro de Medicina Andina, Cuzco); Prov. Quispicanchis, Camanti, Mariri, 720
m, 9 Sep 1990 (Y fl-fr), Timand 913 (BG, MO), (d), Timand 918 (MO); Prov. Quispicanchis, between Inambari & Quince Mil, 500-600 m, s.d. (Y fl-fr), Vargas C. 16517 (US). HUANUCO: Rd. Huanuco-Tingo Maria, below Chin- chao, ca. 1800 m, 24 Nov 1997 (6), Berg et al. 1737 (BG, COL, MOL); nr. Tingo Maria, 9 Aug 1940 (Y fl-fr), Asplund 12928 (S). JUNiN: Above San Ram6n, 1400-1450 m, 8-12 Jun 1929 (6), Killip et al. 24737 (F, NY, US); La Merced, 1350 m, 27 Aug-I Sep 1923 (9), Macbride 5741 (F). MA- DRE DE Dios: Prov. Manu, nr. Shintuya, ca. 500 m, 30 Jul 1988 (juv), Berg et al. 1610A (BG). PASCO: Rd. Oxapampa- Llaupi-Cerro de Pasco, ca. 13 km, ca. 2100 m, 29 Nov 1997 (6), Berg et al. 1759 (BG, COL, MOL); rd. Oxapampa- Llaupi-Cerro de Pasco, ca. 13 km, ca. 1950 m, 29 Nov 1997 (9 fl-fr), Berg et al. 1762 (BG, COL, MOL); 20 km NE of Villa Rica, rd. to Cacazu, 1600 m, 4 Jul 1988 (6), Gentry et al. 63245 (BG, MO, MOL); Prov. Oxapampa, Gran Pa- jonal, W of Chequitavo, 1200 m, 10 Apr 1984 (9 fl-fr), D. N. Smith 6905 (BG, MO, MOL). PUNO: Prov. Sandia between Quinsacu & Munu, 5 Aug 1965 (6), Vargas C. 16381 (US). SAN MARTiN: Prov. Rioja, rd. Rioja-Pomacocha, nr. km 391-392, ca. 1750 m, 8 Dec 1997 (d), Berg et al. 1798 (BG, COL, MOL); Prov. Rioja, rd. Rioja-Pomacocha, nr. km 378, ca. 2200 m, 8 Dec 1997 (6), Berg et al. 1801 (BG, COL, MOL), km 390-394, 1910-2040 m, s.d. (9 fl-fr), D. N. Smith 4525 (BG, MO). UCAYALI: Rd. Pucallpa-Tingo Maria, ca. km 145, near Huipoca, ca. 400 m, 26 Nov 1997 (9 fl-fr), Berg & Franco 1750 (BG, COL, MOL); rd. Pu- callpa-Tingo Maria, nr. Boquer6n, ca. 400 m, 26 Nov 1997 (d), Berg & Franco 1753 (BG, COL, MOL). WITHOUT LOCALITY: 1784 (st), Ruiz & Pavon s.n. (G).
BOLIVIA. BENI: Prov. Ballivian, rd. Yucumo-Caran- avi, ca. km 13, ca. 900 m, 4 Mar 1994 (st), Berg 1713 (BG, COL, LPB); Prov. Ballivian, Serrania Pil6n Lajas, 24 km SW of Yucumo, rd. Yucumo-Quiquibey, km 24, 850-900 m, 3-7 May 1991 (st), Killeen et al. 3072 (BOLV, USZ); rd. Yucumo-Rurrenabaque, km 28, Rio Hondo, 12 Mar 1989 (9 fl-fr), D. N. Smith et al. 12856 (LPB). COCHABAMBA: Rd. Cochabamba-Villa Tunari, km 96, 1800 m, 6, Dec 1995 (6), Berg et al. 1724 (BG, BOLV, COL, LPB, USZ); Prov. Chapare, Locotal, 1600 m, 5 Feb 1929 (9 fl-fr), Steinbach 9030 (BM, E, F, G, GH, K, MO, NY, S, U), 20 Feb 1929 (st), Steinbach s.n. (US). LA PAZ: Prov. Sud Yungas, Chu- lumani, rd. to Ocobaya, 1800 m, 28 Jun 1987 (9 fl-fr), Beck 12082 (BG, LPB); rd. Yucumo-Caranavi, nr. La Cascada, ca. 5 km SSW of Quiquibey, ca. 1100 m, 4 Mar 1994 (9 fl- fr), Berg 1715 (BG, COL, LPB); San Carlos, Rio Mapiri, 750 m, 7 Sep 1907 (st), Buchtien 2118 (NY, US); Prov. Sud Yungas, Rio Boopi, San Bartolom6, 1-22 Jul 1939 (6), Kru- koff 10304 (A, F, G, K, MICH, MO, NY, S, U, US); Prov. Larejaca, Tuiri, 12-30 Sep 1939 (6), Krukoff 10811 (A, G, K, MICH, MO, NY, S, U, US).
This species is highly variable not only morpho- logically but also ecologically, considering the wide elevational range inhabited and the occurrence in rel-
atively dry habitats in some regions and in very wet habitats in others. Attempts to distinguish (at least at the subspecific level) formal entities related to distri-
62 FLORA NEOTROPICA
bution and ecology have failed, partly because of the rather confusing patterns in the morphological differ- entiation and partly because of lack of sufficient col- lections from many parts of its range of distribution. But it is possible to recognize some more or less dis- tinct informal entities, representing extremes of the variation.
In material from Venezuela and the eastern Cor- dillera in Colombia, southward to Caqueta, and from the Sierra de la Macarena, the indumentum is (rather) sparse and the upper surface of the lamina is usually smooth. The lamina is usually coriaceous to subcor- iaceous, and the reticulum veinlets are often thick, and, therefore, the areoles small beneath. This mate- rial includes the types of Cecropia coriacea, C. mon- iquirana, C. palmatisecta, and C. philipsonii.
Another type is found in the western part of Co- lombia from Antioquia to Narinlo and adjacent Ec- uador (Carchi). It is largely associated with relatively dry areas in the western Cordillera, the Cauca Valley, and parts of the central Cordillera. It is distinctly hairy (hirtellous to subtomentose) on the veins of the lam- ina beneath, scabrous above, and at least in the south- ern and northern part of its range of distribution often without arachnoid indumentum in the areoles of the lamina beneath. This type is most clearly represented by the material described as Cecropia caucana and C. danielis, but it also includes the type material of C. strigilosa. In the Cauca Valley the stipules and spathes are usually reddish to purplish, and the pis- tillate inflorescences can be ? pendulous at fruit, due to relatively long peduncles. South of the Cauca Valley (Narinlo and adjacent Ecuador) the stipules and spathes are greenish to yellowish, the peduncle of the pistillate inflorescences is short, the arachnoid indu- mentum in the areoles is often lacking, the internodes can be completely filled by brown pith, and the num- ber of lamina segments is relatively small, mostly 8- 9, as in the type of C. caucana. In (northern) Antio- quia (as in the type of C. danielis), the smaller veins of the lamina are often prominent beneath, whereas normally (almost) plane, the arachnoid indumentum is often lacking in the areoles of the lamina beneath, but in contrast to the material occurring south of the Cauca Valley, the number of lamina segments is 9- 1 1. In adjacent wetter parts of Colombia and Ecuador, the lamina has usually 11-13 segments and the in- dumentum on the veins beneath is short and incon- spicuous.
On the eastern slopes of the Andes, from Caqueta' to Morona-Santiago, three morphological types can be distinguished. One of them resembling the type of the wetter parts in western Colombia and Ecuador, a
second one with relatively few (ca. 9) lamina seg- ments and densely hairy, sericeous to subhirsute on the stipules and often also on the spathes, and a third one (only found in Caqueta') with rather dense arach- noid indumentum on the leafy twigs and the petioles. The second type has been described as Cecropia hachensis and occurs in two color morphs, at least in parts of its range of distribution: a green morph and a reddish to purplish one.
Material in the coastal region of Ecuador, in El Oro and Manabf, at 550-1500 m, shows similarities to Cecropia montana in the whitish stipules and spathes and in the presence of rather long white uni- cellular hairs in the trichilia, but the number of lateral veins in the free part of the midsegment is up to ca. 20 pairs, thus distinctly less than the minimum num- ber in C. montana.
In the southern part of the species range at least three types can be recognized. The most prominent one is found in Bolivia and adjacent Peru. It has blackish and subpersistent stipules and incisions down to ca. 6/10-7/10 or incisions down to near the petiole. More northward in Peru, the stipules vary in color from brown to greenish and are either caducous or subpersistent. The latter type comprises the type of the species. In some specimens from Bolivia and Peru (as in the type of the species) the lamina has narrow segments, to 4 cm broad in medium-sized leaves. In Bolivia (Cochabamba) a purplish morph occurs be- sides the predominant green morph. In the majority of the material from Peru, the number of lateral veins in the free part of the midsegment varies from ca. 23 to 35 pairs, but in the third type the number varies between 15 and 20 pairs. The third type is represented by Berg et al. 1632, 1750, and 1753, the latter two found at ca. 400 m.
A remarkable type is represented by material from southern Ecuador and northen Peru on the eastern slopes of the Andes and from Costa Rica and western Panama. Material from Ecuador was recently de- scribed as Cecropia villosa (Berg & Franco Rosselli, 1993). Although in general features quite similar to C. angustifolia, it is distinct in villous indumentum and internodes entirely filled with brown pith. Further studies on the genus revealed that this material did not differ clearly from collections from Costa Rica which were named C. polyphlebia and were made from trees not inhabited by ants (Longino, 1989a); the Ecuadorian trees are also not occupied by ants. Ants are absent in spite of the common presence of trichilia, producing abundant Miillerian bodies (cf. Janzen, 1973: 17, fig. 1). The absence of ants is likely caused by the ample pith in the internodes. In C. an-
SYSTEMATIC TREATMENT 63
gustifolia the pith in the intemodes is usually very sparse, rarely ample (as in Berg et al. 1759). The un- usual disjunction between two sets of specimens and the fact that they fill gaps in the distribution of C. angustifolia led finally to the decision to include the material initially recognized as C. polyphlebia and C. villosa in C. angustifolia. This villous type tends to have more lateral veins [ca. 25-35(-40) pairs] in the free part of the midsegment of the lamina than is usual in C. angustifolia. It is remarkable not only that this villous type with ample pith arose disjunctly, but also that both in Ecuador and Central America it is mor- phologically linked to a hairy type without ample pith. In Ecuador, this type includes the type of C. hachensis, and in Central America it is represented by collections with mostly brownish indumentum from Nicaragua and Honduras.
This species is closely related to Cecropia mon- tana. The differences between the two species are dis- cussed under the latter.
5. Cecropia annulata C. C. Berg & P. Franco, Novon 6: 245. 1996. Type. Bolivia. La Paz: Nr. Sapecho, 1 Mar 1994 (Y fl-fr), Berg 1704 (holotype: LPB; isotypes: BG, COL, MO, NY).
Tree, to 25 m tall; trunk with prominent (annular) scars of the stipules. Leafy twigs 2-4.5 cm thick, (dark) green, hispidulous with curved to uncinate hairs. Lamina subcoriaceous to coriaceous, ca. 30 X 30 cm to 75 X 75 cm, the segments 8-10, the free parts of the upper segments obovate to elliptic, the upper ones sometimes slightly lobate, the incisions down to 6/10-8/10(-9/10); apices obtuse; upper sur- face smooth to scabridulous, sparsely to rather densely minutely puberulous to strigillose on the (main) veins, initially also with sparse arachnoid in- dumentum; lower surface sparsely puberulous to stri- gillose with straight, curved, or uncinate hairs on the (main) veins, with arachnoid indumentum (almost) confined to the areoles or almost absent; lateral veins 11-16(-20) pairs, marginally loop-connected, the lower ones branched; petiole ca. 25-70 cm long sparsely (minutely) puberulous and also with sparse arachnoid indumentum; trichilia fused, the brown in- dumentum intermixed with (rather) short white hairs;
stipules 12-20(-28) cm long, orange-red to pinkish or partly whitish, caducous, (appressed-)puberulous to hirtellous, or on the ribs to subhirsute, also with dense arachnoid indumentum and rather dense brown pluricellular hairs outside, ? densely hairy inside. Staminate inflorescences solitary or in pairs, the pe- duncle erect, the spikes ? spreading to pendulous; peduncle 9-13 cm long, with sparse arachnoid indu- mentum and brown pluricellular hairs; spathe 12-15 cm long, white, with dense arachnoid indumentum and sparse brown pluricellular hairs outside, glabrous or sparsely hairy inside; spikes ca. 10-25, 6-17 X ca. 0.3 cm (yellow to pale orange), with stipes 0.8-1.3 cm long and sparsely puberulous in the upper part; rachis hairy. Staminate flowers: perianth tubular, 1- 1.5 mm long, glabrous, the apex slightly convex, smooth; filaments flat; anthers ca. 0.8 mm long, ap- pendiculate, detached at anthesis (?). Pistillate info- rescences solitary or in pairs, pendulous; peduncle 9.5-19 cm long, sparsely puberulous to hirtellous (to subhispid); spathe not seen; spikes 4-5(-6), 9-17 X
(0.5-)0.7-0.8 cm, sessile or with stipes to 0.5 cm long and minutely puberulous; rachis hairy. Pistillate;flow- ers: perianth ca. 1.5-2 mm long, with arachnoid in- dumentum below the apex or also on the margin of the apex outside, also below the style channel inside, the apex ? convex, punctate to muriculate; style rather short; stigma tongue-shaped to subpeltate. Fruit ellipsoid, ca. 1.8 mm long, smooth.
Distribution (see Fig. 10.2). Bolivia (La Paz and Beni), in non-inundated forest and secondary growth, at low elevations.
Specimens examined. BOLIVIA. BENI: Prov. Balli- vidn, Serranfa del Pil6n, 8-10 km from, Yucumo, 19 May 1989 (9 fl-fr), D. N. Smith et al. 13264 (BG, BOLV, LPB, USZ). LA PAZ: Prov. Sud Yungas, nr. Sapecho, 26 Feb-i Mar 1994 (st), Berg 1698A (BG), (d), Berg 1699 (AAU, BG, COL, LPB, MO), (6) Berg 1701 (BG, COL, LPB); nr. Tucupi (= Tullupi), ca. 30 km SE of Palos Blancos, nr. Rio Beni, 5 Mar 1994 (9), Berg 1717 (AAU, BG, COL, LPB, MO, NY).
This species is probably closely related to Cecro- pia polystachya. It differs from the latter in the mar- ginally loop-connected lateral veins in the free part of the midsegment, the (very) sparse arachnoid indu- mentum on the petiole, the (rather) short white (uni- cellular) hairs in the trichilia, and the entire (or rarely slightly) lobate midsegment. However, C. annulata also resembles C. concolor subsp. engleriana, from which it differs in the less deeply incised lamina, the smaller number of lateral veins in the free part of the midsegment, and the non-peltate stigmas. The char- acters of this species look like a mixture of those of
64 FLORA NEOTROPICA
the two related species named above and might in- dicate an origin by hybridization. The species is rel- atively rare in secondary growth. In the field, it can be easily recognized by the orange to pinkish young leaves. The filaments of the stamens can be distinctly shorter than the perianth or they are longer, with the apices beyond the aperture; this might imply that the anthers are not detached before anthesis.
Vernacular name. Bolivia: ambaibo negro (Beni).
6. Cecropia bullata C. C. Berg & P. Franco, Fl. Ec- uador 48: 14, t. 3. 1993. Type. Ecuador. Pichincha: Rio Guajalito, ca. 2 km N of Palmeras, rd. Quito- San Juan-Chiriboga-Enpalme, km 59, ca. 1850 m, 15 Dec 1990 (Y), Berg et al. 1648 (holotype: QCA; isotype: BG). Fig. 15
Tree, to 20 m tall. Leafy twigs 2-4 cm thick, densely white to brownish villous to subsericeous or to subhirsute; internodes entirely or partly filled with brown pith. Lamina subcoriaceous (when dry often brittle), ca. 30 X 30 cm to 70 X 70 cm, the segments 7-9, the free parts of the upper segments elliptic to obovate, the incisions in the upper part of the lamina down to 5/10-6/10, the lower part of the lamina lobed; apices (of the upper segments) acuminate; upper sur- face smooth, bullate, (sub)glabrous; lower surface with concave areoles, ? densely (sub)hirsute to hir- tellous to subtomentose or sparsely hirtellous to pu- berulous (to subglabrous) on the veins, on the main veins often also with elongate brown pluricellular hairs, with (sparse) arachnoid indumentum in the ar- eoles, mostly extending to the reticulum or also dense arachnoid indumentum largely covering the area be- tween the secondary veins, often extending to the main veins; lateral veins in the free part of the mid- segment 10-20 pairs, submarginally loop-connected, unbranched (or the lower ones branched); petiole ca. 40-55 cm long, puberulous to hirtellous or also whit- ish to brownish hirsute to (sub)villous, often also with rather dense arachnoid indumentum (or subglabrous); trichilia fused, the brown indumentum intermixed with sparse short whitish to brownish (unicellular) hairs; stipules (8-)20-36 cm long, pink to dull red, densely whitish to brownish villous to subhirsute to subsericeous or also with short arachnoid indumen- tum outside, sparsely hairy to glabrous inside. Sta- minate inflorescences in pairs or solitary, the peduncle erect, the spikes erect to ? spreading; peduncle ca. 6.5 cm long, white-hirsute; spathe pink to dull red, ? densely villous to subhirsute to hirtellous outside, gla- brous inside; spikes 4-10, 6-15 X 0.7-0.8 cm, with
stipes 0.3-1.5 cm long and subsericeous or with arachnoid indumentum; rachis hairy. Staminateflow- ers: perianth tubular, ca. 2 mm long, glabrous or pu- berulous below the apex, the apex plane to slightly convex, hispidulous to muriculate; filaments swollen; anthers 0.5-0.7 mm long, appendiculate, remaining attached to the filament at anthesis (?) Pistillate inflo- rescences (usually) solitary, erect at anthesis to pen- dulous in fruit; peduncle ca. 5-12 cm long, at least in the upper part or the lower part hirsute to subvillous, also with short hairs and/or sparse arachnoid indu- mentum; spathe 10-14 cm long, the color and indu- mentum as in the staminate inflorescence; spikes 2 or 1, 8-12 X ca. 1 cm, to 21 X ca. 2-3.5 cm in fruit, subsessile or with stipes ca. 0.5 cm long and hirtellous to hirsute; rachis hairy. Pistillateflowers: perianth ca. 4-5 mm long, with arachnoid indumentum below the apex, sometimes also sparsely in the style channel inside, sometimes also short stiff hairs just below the apex outside, the apex slightly convex, muriculate to smooth or hispidulous around the aperture; style rather long, straight, hairy; stigma penicillate. Fruit oblongoid, ca. 3 mm long, smooth.
Distribution (see Fig. 11.3). On the western slopes of the western Cordillera in Colombia, from Antioquia to Narinlo, and in northwestern Ecuador, in (sub)montane (cloud) forest, at ca. 1700-2000 m.
Specimens examined. COLOMBIA. ANTIOQUIA: Mun. Frontino, Cgto. Nutibara, rd. Nutibara-La Blanquita, Murri region, 1600 m, 4 Mar 1955 (9 fl-fr), Franco et al. 5550 (BG, COL, HUA), 1650 m, (5), Franco et al. 5553 (COL, HUA). CHOCO: Mun. San Jose del Palmar, Vrda. El Sinai, ca. 1800 m, 19 Mar 1994 (d), Franco et al. 4559 (BG, HUA), (9 fl-fr), Franco et al. 4561 (BG, HUA). NA- RINO: Mun. Ricaurte, Reserva Natural La Planada, ca. 1700 m, 19 Jun 1990 (5), Benavides 11138 (PSO); Mun. Ri- caurte, rd. to Reserva Natural La Planada, 1800 m, 8 Oct 1993 (9 fr), Franco et al. 4467 (BG), 22 Feb 1995 (9 fl- fr), Franco et al. 4698 (COL); La Planada Reserva, 7 km from Chucunes, 1800 m, 7 Jan 1988 (9 fl-fr), Gentry et al. 60519 (BG, MO, PSO); Mun. Ricaurte, Reserva Natural La Planada, 20 Jul 1987 (9 fl-fr), Restrepo et al. 378 (MO), 14 Jan 1989 (5), Restrepo 441 (MO). VALLE: Mun. El Silen- cio, Hda. Himalaya, W of Yumbo, 1840 m, 4 Feb 1989 (5), Gentry et al. 65484 (BG, MO); Mun. Cali, rd. Cali-Bue- naventura km 18, rd. to Dapa, km 4, Finca Zingara, 1900 m, 10 Jul 1944 (9 fl-fr), Giraldo-Gensini et al. 411 (NY, TULV).
ECUADOR. CARCHI: Cant6n Mira, N of El Carmen, rd. to Chical, 2000-2200 m, 10 Feb 1992 (9 fl-fr), Gudinio et al. 9720 (QCNE); Cant6n Mira, rd. to Chical, N of El Carmen, 2000-2200 m, 10 Feb 1992 (9 fl-fr), Palacios et al. 9720 (BG, QCNE). PICHINCHA: Nr. Saloya, 1800 m, 9 Sep 1943 (st), Acosta S. 5848 (F); rd. Atenas-Tandapi, 1700 m, 3 Nov 1991 (d), Jaramillo et al. 14521 (NY, QCA); rd.
FIG. 15. Cecropia bullata. 1. Lamina, reduced. 2. Apex of lamina and venation (Berg et al. 1648). 3. Stipules. 4. Staminate inflorescence with spathe. 5. Staminate inflorescence at anthesis. 6. Staminate flower and stamen (Franco et al. 4559). 7. Pistillate inflorescence with spathe (Franco et al. 4461). 8. Pistillate inflorescence at anthesis and base of petiole with trichilium (Zak 1312). 9. Pistillate inflorescence in fruit and trichilium (Franco et al. 4467). 10. Pistillate flower. 11. Fruit (Franco et al. 4561). (From Fl. Ecuador 48: 15. 1993, modified.)
66 FLORA NEOTROPICA
Quito-San Juan-Chiriboga-Empalme, Rio Guajalito, ca. 2 km N of Palmeras, km 59, 1800-1900 m, 29 Feb 1992 (d), Jaramillo et al. 14651 (QCA); km 59, 15 km NE of rd., ca. 1700 m, 23 Sep 1986 (Y fl-fr), Zak 1312 (BG, MO, NY,
QCA).
This species is closely related to both Cecropia telealba and C. gabrielis, from which it can be easily distinguished by the surfaces of the lamina: distinctly bullate above and with concave areoles beneath. From C. telealba it can also be distinguished by the absence of dense arachnoid indumentum on the upper surface of the lamina and from C. gabrielis by the hairy (vil- lous to subhirsute) leafy twigs. The leaves are more distinctly bullate and with less dense arachnoid in- dumentum on the lamina beneath in the southern part of the range of distribution (Ecuador and Narifio) than in the northern part of the range (Valle to Antioquia). Since the villous indumentum is sometimes less pro- nouncedly present in the northern part and the stipules and spathes of C. gabrielis can be (partly) sparsely (sub)villous in Antioquia, C. bullata can (only) be distinguished by the presence of villous indumentum on the leafy tiwgs (and mostly also on the petioles). The trees are not inhabited by ants.
7. Cecropia chlorostachya C. C. Berg & P. Franco, Caldasia 24: 233. 2002. Type. Peru. San Martin: Prov. Rioja, rd. Rioja-Pomacocha, nr. km 391- 392, ca. 2000 m, 8 Dec 1997 (Y fl-fr), Berg & Franco 1802 (holotype: MOL; isotypes: BG, COL). Fig. 16
Tree, to 22 m tall. Leafy twigs 2-3.5 cm thick, whitish (due to the indumentum), green, white- to brownish-subvillous to -subhirsute to -hirtellous, sometimes only near the scars of the stipules, also with dense arachnoid indumentum and brown pluri- cellular hairs. Lamina (sub)coriaceous, ca. 50 X 50 cm to 60 X 60 cm, the segments 7-10, the free parts of upper segments elliptic to oblong, the incisions in the upper part of the lamina down to ca. 6/10, apices acuminate; upper surface (almost) smooth, rather sparsely puberulous to hirtellous, also sparse arach- noid indumentum; lower surface rather sparsely sub- villous to subhirsute to hirtellous on the veins, with arachnoid indumentum in the areoles, extending to the main veins, on the main veins also brown pluri- cellular hairs, most densely near the petiole; lateral veins in the free part of the midsegment ca. 15-20 pairs, submarginally loop-connected, mostly unbran- ched; petiole ca. 25-55 cm long, white subvillous to subhirsute and with dense arachnoid indumentum; tri- chilia fused, the brown indumentum intermixed with
short and (partly with) long white hairs, the Miillerian bodies pinkish; stipules 15-32 cm long, subpersistent, pinkish, densely (sub)villous, also with dense arach- noid indumentum and brown pluricellular hairs out- side, sparsely to (partly) densely hairy inside. Stami- nate inflorescences in pairs, erect, or the spikes ? spreading; peduncle ca. 3-6 cm, white, subhirsute to hirtellous and with dense arachnoid indumentum; spathe ca. 12-15 cm long, white, (sub)villous, with dense arachnoid indumentum and with brown pluri- cellular hairs outside, inside hairy; spikes 7-8, 15-17 X 0.6-0.8 cm, before anthesis yellow, at anthesis green, with stipes 0.5-1 cm long and hirtellous; rachis hairy with stiff and crinkled (arachnoid) hairs. Sta- minateflowers: perianth tubular, 1.8-2 mm long, with arachnoid indumentum below the apex, the apex plane, sparsely muriculate; anthers ca. 0.8 mm long, with short appendages, detached at anthesis, remain- ing attached to the filament by 2 filiform connections between the connective and the upper margin of the filament. Pistillate inflorescences in pairs or solitary, erect to ? spreading with the spikes straight to curved upward, subtended by whitish, caducous bracts, to 3 cm long; peduncle ca. 7-10 cm long, the indumentum as in the staminate inflorescence; spathe 10-12 cm long, pinkish, the indumentum as in the staminate in- florescence; spikes 4, 4-12 X ca. 0.-0.5, to 15 X 1 cm in fruit, subsessile or with stipes to 0.4 cm long with dense arachnoid indumentum; rachis hairy with stiff and crinkled hairs. Pistillate flowers: flowers of- ten basally connate; perianth ca. 2.5 mm long, with arachnoid hairs below the apex outside, and below the style channel inside, the apex plane to slightly convex and muriculate; style short; stigma comose. Fruit el- lipsoid to oblongoid, ca. 1.5-2 cm long, (almost) smooth.
Distribution (see Fig. 8.5). Northern Andean Peru (San Martin and Amazonas), in cloud forest, at ca. 1750-2000 m.
Specimens examined. PERU. AMAZONAS: Prov. Chachapoyas, Rodrigues Mendoza, Cochamal, Montania de Yanamonte, 4 Jul 1991 (? fl-fr), C. Diaz et al. 4567 (MO). SAN MARTiN: Prov. Rioja, rd. Rioja-Pomacocha, ca. 1750 m, 8 Dec 1997 (d), Berg et al. 1799 (BG); Prov. Rioja, rd. Pedro Ruiz-Moyobamba, km 390, Venceremos, 2100 m, 7- 9 Aug 1983 (i), D. N. Smith et al. 4686 (BG).
This species is distinguished by the dense and white indumentum on various parts, the subpersistent stipules, and the green spikes of the staminate inflo- rescence. Green spikes sometimes occur in Cecropia distachya, but only before anthesis (as in Berg et al. 1597). The species is probably not myrmecophytic.
SYSTEMATIC TREATMENT 67
FIG. 16. Cecropia chlorostachya. 1. Lamina, reduced (D. N. Smith et al. 4686). 2. Apex of lamina and venation. 3.
Stipuesadbaeofetioewihtrihilim(CDia et al. 4567).4.Stminatinflrescecew
ils c a sa Smith et al. 4686).6.Staminateflower(lngi
stn7Smafanarecnone(gt.J . .Pt i
'I" ? \ a
'0
FIG. 16. Cecropia chlorostachya. 1. Lamina, reduced (D. N. Smith et al. 4686). 2. Apex of lamina and venation. 3. Stipules and base of petiole with trichilium (C. Diaz et al. 4567). 4. Staminate inflorescence with spathe. 5. Pair of staminate inflorescences at anthesis and base of petiole with trichilium (D. N. Smith et al. 4686). 6. Staminate flower (longitudinal section). 7. Stamen and filament after detachment of anther (Berg et al. 1799). 8. Pistillate inflorescence with spathe (C. Diaz et al4567). 9. Pair of pistillate inflorescences at anthesis and base of petiole with trichilium. 10. Pistillate flower and style. 11. Fruit (Berg et al. 1802). (From Caldasia 24: 234. 2002.)
68 FLORA NEOTROPICA
8. Cecropia concolor Willdenow, Sp. P1. 4(2): 652. 1806; Berg, Acta Amazonica 8(2): 164. 1978. Am- baiba concolor (Willdenow) Kuntze, Revis Gen. P1. 2: 624. 1891. Type. Brazil. Para: Without lo- cality, (Y), Sieber in herb. Hoffinannsegg s.n. (ho- lotype: B in herb. Willdenow, destroyed, photo- graphs in F, G, MO, US), herewith replaced by: Brazil. Para: Mun. Belem, Mocambo, 8 Oct 1995 (6), Berg 1721 (neotype: MG; isoneotypes: B, BG, COL, INPA, NY, RB, U, UB). Fig. 17
Cecropia leucocoma Miquel, in Martius, Fl. Bras. 4(1): 142. 1853. Ambaiba leucocoma (Miquel) Kuntze, Revis. Gen. P1. 2: 624. 1891. Type. Brazil. Ama- zonas: Nr. Manaus (= Barra), Dec 1850-Mar 1851 ( i ), Spruce 1322 or s.n. (holotype: M; isotypes: BM, E, F, G, GH, K, LE, NY, P).
Cecropia obovata Rusby, Bull. New York Bot. Gard. 6: 498. 1910. Type. Bolivia. La Paz: San Buenaventura, 15 Nov 1901 (Y), R. S. Williams 645 (holotype: NY; isotypes: BM, K).
Cecropia maranhensis Snethlage, Notizbl. Bot. Gart. Berlin-Dahlem 9: 172. 1924. Syntypes. Brazil. Mar- anhao: Turiaqu, 2-3 Jan 1924 (d) (Y), Snethlage 384 (B, destroyed, photographs ex B in F, G, MICH, MO), (cd), Snethlage 388 (B, destroyed, photographs ex B in F, G, GH, MICH, MO; isotype: F).
Tree, to 15 m tall; stem with prominent stipular scars. Leafy twigs 1.5-4 cm thick, green with yellow spots to purplish-brown, hispidulous, with curved or uncinate hairs, or also with sparse to rather dense arachnoid indumentum. Lamina subcoriaceous to chartaceous, ca. 30 X 30 cm to 65 X 65 cm, the segments (7-)8-10(-11), the free parts of the upper segments oblong to subobovate to oblanceolate, entire or ? sinuate (or if juvenile, then to lobate), the in- cisions down to 1.5(-2.5) cm from the petiole; apices short-acuminate to (sub)acute to obtuse; upper surface smooth to scabridulous, sparsely puberulous to sub- hispidulous, also with sparse to rather dense arach- noid indumentum, along the main veins often with brown pluricellular hairs; lower surface minutely puberulous on the main veins, with arachnoid indu- mentum in the areoles, on the smaller veins, or, at least initially, also on the main veins; lateral veins in the free part of the midsegment 16-24 (or if juvenile to 30) pairs, marginally loop-connected, mostly branched; petiole 25-50 cm long, sparsely puberulous and/or with sparse to (rather) dense arachnoid indu- mentum; trichilia fused, the brown indumentum in- termixed with (sparse) short white hairs; stipules 7- 15 cm long, white to pale yellow or pinkish, caducous or subpersistent, puberulous or sparsely sericeous, of- ten with ? dense arachnoid indumentum, sometimes (also) with ? dense brown pluricellular hairs outside,
? densely sericeous inside. Staminate inflorescences in pairs or solitary, pendulous; peduncle (3.5-)10-19 cm long, sparsely puberulous to hirtellous and with sparse to rather dense arachnoid indumentum; spathe 10-20 cm long, white to yellowish or pinkish, puber- ulous to hirtellous and often with dense arachnoid in- dumentum outside, glabrous or (densely) sericeous inside; spikes (8-)15-20, (1-)6-15 X 0.2-0.4 cm, with stipes 0.2-2 cm long and hairy or subglabrous; rachis hairy. Staminateflowers: perianth tubular, 0.8- 1.5 mm long, glabrous, the apex almost plane; fila- ments slightly swollen; anthers 0.5-0.6 mm long, ap- pendiculate, detached at anthesis, reattached at the margins of the aperture by the appendages (?). Pistil- late inflorescences in pairs or solitary, pendulous; pe- duncle (3.5-)10-22 cm long, puberulous to hirtellous and with sparse to rather dense arachnoid indumen- tum; spathe 10-17 cm long, the color and indumen- tum as in the staminate inflorescence; spikes 4, (2-) 5-18 X 0.4-1 cm, to 29 X 1-1.5 cm in fruit, sessile or with stipes to 1(-1.5) cm long and hairy: rachis hairy. Pistillate flowers: perianth 1.5-2.5 mm long, with arachnoid indumentum on and below the apex, absent inside, the apex plane; style short; stigma (sub)peltate. Fruit ellipsoid to ovoid to oblongoid, 1.5-2.5 mm long, ? tuberculate.
Distribution (Fig. 18.1). Lower and central Am- azon basin, extending to Bolivia and southern Ama- zonian Peru (Cuzco and Madre de Dios), common in secondary growth, in non-inundated places or (in the southwestern part of its range) sometimes in tempo- rary inundated places, at low elevations.
Representative specimens examined. PERU. Cuzco: Prov. Paucartambo, between Pilcopata & Atalaya, 29 Jul 1988 (d), Berg et al. 1605 (BG, COL, Centro de Medicina Andina, Cuzco); Prov. Paucartambo, rd. Pilcopata-Paucar- tambo, km 12, 2 Aug 1988 (9 fl-fr), Berg et al. 1628 (BG, COL, Centro de Medicina Andina, Cuzco); Prov. Paucar- tambo, rd. Pilcopata-Atalaya, halfway, 24 Oct 1984 (9 fr), Maas et al. 6160 (BG, U); Prov. Paucartambo, between Tono & Pifiipifii (= Kosnipata), 23 Nov 1965 (6), Vargas C. 16900 (US). MADRE DE DIos: Prov. Manu, nr. Shintuya, 30 Jul 1988 (9 fl-fr), Berg et al. 1609 (BG, COL, Centro de Medicina Andina, Cuzco); Parque Nacional Manu, Rfo Manu, Cocha Cashu Station, 19 Sep 1986 (6), Foster 11435 (BG, MOL, US), (9 fl), Foster 11444 (BG); Parque Na- cional Manu, Rio Manu, Tayakome, 29 Sep 1986 (9), Foster et al. 11528 BG, US).
BRAZIL. AMAPA: Estrada do Marucca, 6 Nov 1979 (6), Rabelo 62 (MG). AMAZONAS: Manaus, INPA, 29 Aug 1973 (d), Berg 245 (U), (9 fl-fr), Berg 246 (A, MO, U, US); rd. Manaus-Caracarai, km 65, 21 Sep 1973 (st), Berg 279 (U), (9 fl-fr), Berg 280 (K, U); Rio Solimoes, Espfrito Santo, between Boca de Januaca & Boca de Manaquiri, 3
SYSTEMATIC TREATMENT 69
I9,'
XIE ~~ ~
6 H '10 8 9
FIG. 17. Cecropia concolor. 1. Lamina, reduced. 2. Apex of lamina and venation (Berg et al. 1628). Cecropia engleriana. 3. Stipules and bases of petioles with trichilia (Berg et al. 1629). 4. Staminate inflorescence with spathe and bases of petioles with trichilia (Alarcon 74). 5. Staminate inflorescence at anthesis (Berg 1605) 6. Staminate flower and stamen (Franco et al. 4688). 7. Pistillate inflorescence after anthesis and base of petiole with trichilium (Gentry 12442). 8. Pistillate flower (Berg et al. 1629). 9. Pistil (Berg et al. 1104). 10. Fruit (Berg 1700).
FIG. 18. Distribution maps. 1. Cecropia concolor, triangles; C. elongata, hexagons; C. pachystachya, circles; both C. concolor and C. pachystachya, crosses. 2. C. megastachya, hexagons; C. metensis, triangles; C. palmata, circles; C. strigosa, open circles. 3. C. peltata, circles; C. schreberiana subsp. schreberiana, triangles; C. schreberiana subsp. antil- larum, hexagons; both subsp. schreberiana and subsp. antillarum, crosses. 4. C. polystachya, circles; C. velutinella, triangles. 5. C. putumayonis, circles; C. reticulata, triangles. 6. C. gabrielis, triangles; C. sararensis, circles. 7. C. telenitida. 8. C. pastasana, circles; C. subintegra, triangles.
SYSTEMATIC TREATMENT 71
Oct 1973 (Y fr), Berg et al. P17590 (C, COL, F, INPA, K, MG, MO, NY, P, S, U, US); Manaus, Reserva Ducke, 28 Sep 1962 (i), Duarte 7190 (INPA, K, RB); Manaus, Feb 1904 (cd + Y fl-fr), Huber (MG) 4175 (G, MG); nr. Manaus, rd. Taruma-Ponta Negra, km 13, 6 Sep 1973 (Y fl), Lleras et al. P17553 (C, F, INPA, K, MG, MO, NY, S, U); Mun. Manaus, ca. 90 km N of Manaus, Distrito Agropecuaria da SUFRAMA, 16 Jul 1992 (Y fl), Nee 43005 (INPA, K, NY); nr. Manaus, Ponta Negra, 16 Mar 1967 (3), Prance et al. 4665 (F, INPA, NY, S, U); rd. Manaus-Itacoatiara, km 26, Reserva Florestal Ducke, 26 Apr 1994 (Y fl-fr), Ribeiro et al. 1293 (BG, INPA); rd. Torquato-Tapaj6s, km 38, 19 Feb 1968 (Y fl), Rodrigues et al. 8421 (F, INPA, US). MAR- ANHAO: Mun. Monqho, Rio Turiaqu, nr. Urutuway, 12 Feb 1985 (c), Balee et al. 809 (BG, MG, NY); Mun. Moncho, Urutawy, Ka'apor Indian Reserve, Rio Turiacu, 28 Sep 1986 (Y fl-fr), Balee 2670 (NY); Sao Bento, 13 Nov 1923 (Y), Snethlage 233 (F); Alto Alegria, 17 Nov 1923 (Y), Sneth- lage 338 (F); Turiacu, 2-3 Jan 1924 (6), Snethlage 383 (F), 385 (F), 386 (F). MATO GROSSO: Nucleo Pioneiro de Hum- boldt, Rio Aripuana, 21 Oct 1973 (Y fl), Berg et al. P.1981 7 (F, INPA, K, MO, NY, P, S, U), (6), Berg et al. P 19818 (F, INPA, K, NY, U, US, S), 11 Oct 1973 (st), Berg s.n. (RB). PARA: Sao Miguel do Guama, Oct 1906 (Y fl), Goeldi (MG) 7777 (BM, MG, G, P, S, U); Belem, 23 Aug 1906 (6), Huber (MG) 7285 (BM, G); Tucurui, BR.230, km 50,5 Nov 1979 (? fl-fr), M. E F da Silva et al. 15 (INPA, MG) (6), M. F E da Silva et al. 16 (INPA, MG).
BOLIVIA. BENI: Prov. Ballivian, Espiritu, RioYacuma, 9 Sep 1986 (6), Beck 5933 (LPB, MO), (Y fl-fr), Beck 5934 (BG, LPB, MO); Prov. Yacuma, Rio Curiraba, 4 Apr 1986 (9 fl-fr), M. Moraes R. 800 (BG, LPB, MO); Prov. Ballivian, rd. Yucumo-Rurrenabeque, km 28, nr. Rio Hondo, 12 Mar 1989 (9 fl-fr), D. N. Smith et al. 12856 (BG, BOLV, LPB, MO, USZ); Prov. Moxos, rd. San Borja-Trinidad, km 27, 6 Sep 1990 (6), D. N. Smith et al. 14336 (BG, MO, USZ); Prov. Ballivian, nr. San Borja, 30 Sep 1976 (9 fl-fr), Ter- ceros et al. 368 (BOLV, LPB, MO); Prov. Cercado, Ibiato, 7 Dec 1991 (st), Townsend 6 (BG, LPB); Trinidad-Misiones Guarayos, Sep 1926 (9 fl-fr), Werdermann 2566 (K, LPB, MO, S). COCHABAMBA: Nr. Chipiri, ca. 5 km N of Villa Tunari, 6 Dec 1955 (d), Berg et al. 1728 (BG, BOLV, COL, LPB, USZ); Prov. Chapare, between Villa Tunari & Puerto Villaroel, nr. Chimore, 30 Dec 1982 (6), Ferndndez C. 7913 (G, NY); Prov. Carrasco, Estaci6n Valle de Sajta UMSS, 28 Oct 1991 (9 fl), Galarza et al. 2 (BOLV). LA PAZ: Prov. Sud Yungas, nr. Sapecho, 26 Feb-2 Mar 1994 (6), Berg 1697 (BG, COL, LPB), (st, juv), Berg 1 703A (BG), (st, juv), Berg 1707A (BG), (6), Berg et al. 1708 (BG, LPB). PANDO:
Prov. Manupiri, rd. Altagracia-San Miquel-San Pedro, 23 Oct 1989 (9 fl), Beck et al. 19665 (BG, LPB); Prov. Nicolas Suarez, nr. Porvenir, 10 Jan 1983 (6), Fernandez C. et al. 8163 (G, NY); Prov. Nicolas Suarez, Campoana, 14 Jan 1983 (9 fl), Ferndndez C. et al. 8238 (G, NY). SANTA CRUZ: Rio Pirai, Jan 1911 (9 fl-fr), Herzog 1508 (G, S); Prov. Andres Ibafiz, 5.5 km NE of Cotoca, 1 km N of El Campanero, 28 Nov 1990 (9 fl-fr), Nee 40049 (BG, LPB, MO, NY, USZ); Jardin Botanico de Santa Cruz, rd. Santa Cruz-Cotoca, km 12, 19 Dec 1991 (6), Nee 42151 (BG,
LPB, MO, NY); Prov. Ichilo, Rfo Ibabo, Reserva Forestal Chore, 16-18 Aug 1990 (? fr), Neill et al. 9345 (MO, NY, QCNE, USZ); Prov. 0. Santiestevan, 15 km NNE of Mon- tero, 9 Oct 1990 ( fl-fr), Saldias et al. 1236 (LPB, USZ); Prov. Velasco, Parque Nacional Noel Kempff M., campa- mento Los Fierros, 30 Oct 1993 (Y fl-fr), Saldias 3356 (USZ); Prov. Sara, Buena Vista, 3 Oct 1925 (d), Steinbach 7268 (A, BM, F, G, GH, K, MO, S, U), (Y fl-fr), Steinbach 7268bis (A, BM, E, F, G, K, MO, NY, S, U).
In subjuvenile material, the incisions of the lamina are often not down to (near) the petiole, but up to 4 cm from it. This feature is occasionally retained even at flowering state. This could be problematic in areas where the ranges of Cecropia concolor and C. pachy- stachya (could) overlap in the southern part of the Amazon basin or where the taxa may intergrade. To be on the safe side, collections from Maranhao and Amazonas with less deeply incised laminas have been included in C. pachystachya in the present treatment. The lamina segments are often rather pronouncedly lobate in (sub)juvenile plants. In this respect the spe- cies resembles C. metensis and C. pachystachya, but it clearly differs from C. engleriana, in which lobation of lamina segments in (sub)juvenile states is less ap- parent or common. The number of lateral veins in juvenile material can be occasionally up to 30 pairs, exceeding the maximum of 24 pairs of adult material and thus extending into the normal range of adult ma- terial of C. engleriana. The trees are small to medium-sized (to ca. 15 m tall). In the middle and lower Amazon basin, they may already start to pro- duce flowers on trees of 1-1.5 m tall while still un- branched. At the southern limits of species range in Bolivia (Santa Cruz), the habit may deviate. Here, the trees are mostly quite robust with relatively thick trunks and the leaves more crowded due to short in- temodes. In Amazonian Brazil and adjacent Amazo- nian Bolivia (Beni), the petiole and the upper surface of the lamina, the petiole, and the stipules are (ini- tially) covered with ? dense arachnoid indumentum. In other parts of Amazonian Bolivia (Cochabamba and Santa Cruz) and southern Amazonian Peru (Cuzco and Madre de Dios) the arachnoid indumen- tum is sparse or lacking on these parts. The type with sparse arachnoid indumentum on the petiole (etc.) has often, or in some areas always, subpersistent stipules. In the conspicuous variation of the tree habit and the regional presence of subpersistent stipules C. conco- lor resembles C. engleriana. A collection made in Colombia [Vichada: Mun. Puerto Carrenlo, Cerro El Bita, airea del Jardin Botainico, 30 Jan 1997 (2 fr), H. Garcia et al. 60 (COL)], far outside the known range of distribution, may have been introduced.
Tree, to 30 m tall. Leafy twigs 1.5-6 cm thick, dark green to red-brown to blackish, hispidulous to puber- ulous to subhirtellous, partly with uncinate hairs. Lamina (sub)coriaceous, ca. 20 X 20 cm to 85 X 85 cm, the segments (5-)7-10(-12), the free parts of up- per segments obovate to subobovate, the incisions down to 6/10-9/10; apices acute to acuminate or to rounded; upper surface smooth (to scabridulous), pu- berulous (to hispidulous), initially often also sparse with arachnoid indumentum; lower surface (minutely) puberulous (with uncinate hairs) on the (main) veins, sometimes also with sparse longer uncinate hairs, with arachnoid indumentum in the areoles, sometimes sparse (to nearly absent), sometimes (initially) also on the main veins; lateral veins in the free part of the midsegment 12-18(-25) pairs, usually marginally or sometimes submarginally loop-connected, most (or some) of them branched; petiole 15-60 cm long, sparsely puberulous, sometimes (initially) also with arachnoid indumentum, or subglabrous; trichilia fused, the brown indumentum intermixed with short white (to brownish unicellular) hairs; stipules (6-) 10- 25 cm long, dark red-brown to brown or to bright red, densely to sparsely puberulous or also with arachnoid indumentum outside, sparsely hairy to subglabrous inside. Staminate inflorescences in pairs, the peduncle erect (or slightly deflexed), the spikes erect to + spreading (or to pendulous); peduncle 2-12(-15) cm long, often at the apex ? strongly broadened, sparsely hispidulous to hirtellous; spathe (7-)10-18 cm long, dark red-brown to red, densely to sparsely puberulous to hirtellous or also with sparse to rather dense arachnoid indumentum outside, glabrous in-
side; spikes (8-)15-25(-50), (0.5-)6-14 X 0.3-0.4 or 0.5-1 cm, ? angular, (sometimes green just before anthesis), the stipes 0.3-1.5 cm long, puberulous, sometimes only with brown pluricellular hairs, or gla- brous; rachis hairy or glabrous. Staminate flowers: perianth tubular, ca. 1-1.5 mm long, with sparse straight hairs and/or sparse to rather dense arachnoid indumentum below the apex, the apex slightly convex to plane, muriculate or (also) with sparse brown pluri- cellular hairs; filaments swollen; anthers 0.6-0.7 mm long, detached at anthesis (?). Pistillate inflorescences in pairs or solitary, erect or deflexed to pendulous in fruit; peduncle (2-)5-13 cm long, hispidulous to hir- tellous; spathe dark red-brown to red, 6-15 cm long, densely to sparsely puberulous to hirtellous or also with arachnoid indumentum outside, glabrous inside; spikes 2-4, (3-)8-15 X 0.6-0.8 or ca. 2.5 cm, to 22 X 1.5(-2) cm or to 38 X 4 cm in fruit, sessile or with stipes to 0.5 cm long and subglabrous; rachis glabrous or sparsely hairy. Pistillate flowers: perianth ca. 1.5- 3.5 mm long, with arachnoid indumentum below the apex, also in the lower part of style channel inside, the apex plane with (a rim around) a circular aperture or strongly convex with a slit-shaped aperture, muri- culate to smooth; style long; stigma comose. Fruit ellipsoid to oblongoid, 2.5-4.5 mm long, tuberculate or smooth.
Distribution (Fig. 10.3). Throughout the Amazon basin, extending to easterm Venezuela, and to French Guiana; common in primary forest, also in secondary growth, in non-inundated (or inundated) places, at low elevations.
Representative specimens examined. COLOMBIA. AMAZONAS: Rio Igara-Parana, 17 km from La Chorrera, 15 Jul 1974 (st), Gasche et al. 222 (COL); Mun. Tarapaca, Parque Nacional Amacayacu, Rio Catuh6, Lorena, 13 Jul 1992 (st), Rudas et al. 5268 (COL). CAQUETA: Nr. Arara- cuara, 12 Jul 1986 (Y fl), Berg et al. 1553 (BG), 29 Feb 1992 (Y fl-fr), Correa et al. 384 (COL). GUAVIARE: Sierra de Chiribiquete, Campamento Norte, 8 Dec 1990 (Y fl-fr), Galeano et al. 2204 (BG, COL, MO); Mun. San Jose de Guaviare, La Libertad, 11 Aug 1989 (cd), Marulanda et al. 1204 (HUA); Sierra de Chiribiquete, 26 Aug 1992 (st), Pa- lacios et al. 2655 (COL). META: Nr. La Macarena, Rio Mor- rocoy, 11 Feb 1995 (Y fl-fr), Franco et al. 4667 (BG, COL); Alto de Bengala, Dec 1993 (? fl), Stevenson 779 (COL). VAUPES: Type collection of C. richardii.
VENEZUELA. AMAZONAS: Depto. Rio Negro, Rio Mawarinuma, Neblina Base Camp, 2 Dec 1984 (Y fl), Boom et al. 5252 (BG, VEN); Depto. Atabapo, Cucurital de Can- ame, Middle Caflo Caname, 30 Apr-I May 1979 (9 fl-fr), Davidse et al. 16969 (MO, U, VEN); Depto. Atabapo, Rio Ocamo, Santa Maria de Los Guaicas, 25 Feb 1981 (9 fr), Gua'nchez 786 (VEN); Hauchica, 11 km NE of San Carlos
SYSTEMATIC TREATMENT 73
o 1 6] 1 9
199'3'~~~~~~~~~~~~~
FIG. 19. Cecropia distachya. 1. Lamina, reduced. 2. Apex of lamina and venation (Berg et al. 1626). 3. Stipules and trichilium (Berg et al. 1627). 4. Staminate inflorescence with spathe and base of petiole with trichilium. 5. Staminate inflorescence at anthesis and base of petiole with trichilium (Berg et al. 1626). 6. Staminate flower and stamen (Berg et al.
1597). 7. Pistillate inflorescence with spathe and trichilium. 8. Pistillate inflorescence at fruit and trichilium. 9. Pistillate flower. 10. Fruit with remnant of style (Berg et al. 1627).
74 FLORA NEOTROPICA
de Rio Negro, 14 Nov 1977 (9 fl-fr), Liesner 3481 (MO, NY, U, VEN); Rfo Caura, Canlo La Ceiba (Canlo Cumaca), 9-26 May 1988 (5), Stergios et al. 12888 (BG, NY); Rio Paragua, Raudales de Maihia, 1 Jan 1962 (st), Steyermark 90526 (NY, US); San Carlos de Rfo Negro, 20 km S of confluence of Rio Negro & Brazo Casiquiare, 10 Nov 1979 (5), Uhl 51 (MO). BOLiVAR: Rio Tirica, 29 Aug 1954 (9 fl-fr), Bernardi 1612 (NY); Rio Parguaza, just below Raudal Maraca, ca. 1 10 km above river mouth, 29 Dec 1955 (9 fl- fr) Wurdack et al. 40997 (BG, MO, NY).
FRENCH GUIANA. Regina region, Mt. Tortue, 11 km WNW of Approuague R., 18 Jun 1988 (5), Feuillet 10312 (BG); Basin de l'Approuague, rd. Regina-St.-Georges, 24 Nov 1995 (9 fl-fr), Granville et al. 13104 (BG, G); Bassin de l'Oyapock, Cr. Gabaret, 16 Sep 1996 (9 fl-fr), Tostain 263, 270 (BG).
ECUADOR. NAPO: Parque Nacional Yasunf, nr. Heli- puerto de Amo Sur, 16-19 Jan 1988 (9 fl), Ceron 3408 (BG, QAME, QCNE). SUCUMBfOS: Cant6n Shushufindi, San Roque, 14 Dec 1997 (st), E. Freire et al. 2784 (QCNE); Reserva Faunistica Cuyabeno, nr. Laguna Grande, 19 Nov 1991 (5), Palacios et al. 9312 (BG, QCNE), 15 Mar 1990 (9 fr), Valencia et al. 375 (BG, QCA) & 67967 (AAU). ZAMORA-CHINCHIPE: Cant6n Nangaritza, Rio Nangaritza, Miazi, 29 Jul 1993 (st), Gentry 80664 (QCNE).
PERU. AMAZONAS: Rio Cenepa, SE of Quebrada Kay- amas, 28 Dec 1972 (9 fr), Berlin 736 (GH); Rio Santiago, E of Caterpiza, ca. 65 km N of Pinglo, 7 Nov 1979 (9 fl- fr), Huashikat 1189 (BG, MO), 17 Jan 1980 (9 fr), Hu- ashikat 1828 (MO); Prov. Bagua, Dtto. Imaza, Rio Marafi6n, Yamayakat, Feb 1995 (st), R. Vdsquez et al. 19938 & 19973 (MO); Prov. Bagua, Cascadas de Mayasito, 5 Sep 1962 (d), Wurdack 1849 (F, NY, US, USM). Cuzco: Prov. Paucar- tambo, rd. Pilcopata-Paucartambo, km 12, 2 Aug 1988 (5), Berg et al. 1626 (BG, COL, Centro de Medicina Andina, Cuzco), (9 fl-fr), Berg et al. 1627 (BG, COL, Centro de Medicina Andina, Cuzco); Prov. Paucartambo, nr. Fortaleza- Patria, 18 Jan 1975 (9 fl-fr), Galeano S. 9 (US, USM). HUANUCO: Prov. Pachitea, 20-24 km SE of Puerto Inca, 1 1 Sep 1988 (9 fr), Morawetz et al. 11-11988 (BG). LORETO:
Nr. Iquitos, 8 Nov 1940 (9 fl-fr), Asplund 14413 (S); Prov. Requena, rd. Jenaro Herrera-Angamos, km 11, 18 Jul 1988 (5), Berg et al. 1597 (AAU, BG, COL), (9 fl), Berg et al. 1598 (AAU, BG, COL); Prov. Loreto, Campamento Petro- lero San Jacinto, Rio Tigre, 6 Sep 1979 (5), Diaz etal. 1470 (BG, MO); Prov. Maynas, nr. Sucusari, 20 Feb 1991 (9 fl- fr), Pipoly et al. 13115 (BG, MO), (st), Pipoly et al. 13141 (BG, MO, NY); Prov. Requena, Jenaro Herrera, 6 Sep 1983 (5), Spichiger et al. 3050 (G, NY), 7 Sep 1983 (9 fl), Spi- chiger 3053 (G); Prov. Maynas, Allpahuayo, Nov 1990 (9
fl), R. Vasquez et al. 14725 (BG, MO, NY). MADRE DE
Dios: Tambopata, 23 Feb 1984 (5), Gentry et al. 46069 (MO); Tambopata Nature Reserve, jct. of Rio La Torre & Rio Tambopata, 31 May 1987 (d), Gentry et al. 58001 (MO). SAN MARTiN: Prov. San Martin, rd. Tarapoto-Yuri- maguas, ca. km 35, 5 Dec 1997 (9 fl-fr), Berg et al. 1781 (BG, COL, MOL).
BRAZIL. ACRE: Reserva INCRA Santa Luzia, BR.364, km 40, 5-19 Oct 1984 (9 fr), Campbell et al. 6806 (BG);
Cruzeiro do Sul, Estrada Alemanha, 7 May 1971 (6), Maas et al. P12771 (INPA, K, MG, MO, NY, U, S, US). AMA- ZONAS: Rd. Manaus-Caracari, km 65, 21 Sep 1973 (6), Berg 282 (U); Rio Jurua, Carauari, 32 km from P6rto Gav- iao, 15-26 Oct 1980 (st), Lisboa et al. 1897 (MG); Mun. Humaita, between Rio Livramento & Rio Ipixuna, 7-18 Nov 1934 (9 fl-fr), Krukoff 7214 (A, BM, F, G, LE, K, MICH, MO, NY, RB, S, U); Mun. Sao Paulo de Olivenqa, Belem Cr., 26 Oct-i l Dec 1936 (9 fl-fr), Krukoff 8921 (A, BM, F, G, LE, K, MO, NY, S, U); Rio Negro, Communidade Apa- recida, opposite mouth of Igarape Tuari, 6 Nov 1987 (6), Maas et al. 6935 (BG, INPA); Rio Cuieras, 2 km below mouth of Rio Brancinho, 12 Sep 1973 (9 fl), Prance et al. 17827 (C, COL, F, INPA, K, MG, MICH, MO, NY, S, U, US), (9 fl-fr), Prance et al. 17840 (COL, F, INPA, K, MO, NY, P, S, U, US); Rio Cuieras, Jarada, 16 Sep 1973 (d), Prance et al. 18008 (C, COL, F, INPA, K, MICH, MG, MO, NY, P, S, TEX, U); Mun. Manaus, rd. Manaus-Itacoatiara, km 26, Reserva Florestal Ducke, 15 Sep 1994 (d), Ribeiro et al. 1424A (BG, INPA). MATO GROSSO: Nucleo Pioneiro de Humboldt, Rio Aripuana, 22 Oct 1973 (9 fr), Berg et al. P 19839 (COL, F, INPA, K, NY, U, US); rd. Vilhena-Jufna, km 134, Estaqao Ecol6gica Ique-Juruena, 24 Aug 1982 (d fr), F A. M. dos Santos et al. 14228 (RB, UEC). PARA: Serra do Cachimbo, nr. Cachoeira Curua, rd. Cuiaba-Santarem, ca. km 875, 30 Oct 1977 (9 fl-fr), Berg et al. 773 (COL, F, K, MG, MO, NY, U, US).
BOLIVIA. LA PAZ: Prov. Abel Iturralde, Puerto Mos- coso, Rio Heath, 25 Jul 1995 (9 fl-fr), Helme et al. 898 (LPB). SANTA CRUZ: Prov. Velasco, Parque Nacional Noel Kempff M., 13 Nov 1993 (9 fl-fr), Arroyo et al. 326 (USZ); Prov. Velasco, Parque Nacional Noel Kempff M., Meseta de Huanchaca, 18 Aug 1996 (9 fl-fr), Carri6n et al. 133 (BG); Prov. Velasco, Parque Nacional Noel Kempff M., campa- mento Huanchaca I, 26 May 1994 (d), H. Gonzdles et al. 42 (BG); Prov. Guayaros, Rio Negro, 13 Jul 1993 (9 fl-fr), I. G. Vargas et al. 2695 (USZ).
In Peru (Cuzco, Huainuco, Junin, and San Martin), the spikes of the pistillate inflorescences reach to 38 X 4 cm in fruit and the spikes of the staminate inflo- rescences are also thicker than usually found else- where. In other parts of the species' range, the spikes are and remain much shorter. Two of these collections with large spikes match the material of Cecropia di- stachya in most features, but are clearly distinct in the shape of the lamina. Moreover, they have unusual arachnoid indumentum on the lamina above. They may represent a distinct type of C. distachya (de- scribed below) in which some (sub)juvenile traits are retained or they may represent a hybrid (with many maternal and few paternal features; see "Nature and Variation of Morphological Characters").
Lamina coriaceous, incisions in the upper part down to ca. 5/10, the lower part slightly lobed; upper surface with sparse to rather dense arachnoid indu- mentum; lateral veins in the free part of the midseg-
SYSTEMATIC TREATMENT 75
ment 9-12 pairs, marginally to submarginally loop- connected. Pistillate inflorescences with the peduncle 2-5.5 cm long, the upper part broadened in fruit; spathe ca. 8 cm long; spikes 2, 5-6 (or more?) X 1- 1.3 cm, to 32 X 2 cm or to 35 X 3.5 cm in fruit. Pistillate flowers: perianth ca. 2.5 mm long. Fruit ob- longoid, 4-5 mm long.
Description based on the following collections: PERU. HUANUCO: Prov. Leoncio Prado, Dtto. Rupa Rupa, E of Tingo Maria, nr. Cerro Quemada, 7 Sep 1978 (? fr), Schunke V 10579 (BG, US). JUNIN: Pi- chanaki, between Satipo and bridge over Rio Chan- chamayo, 900-1000 m, 28 Jun 1982 (9 fl-fr), Gentry et al. 37251 (MO).
Collections with features transitional to the more typical type occur, as with regard to the inflorescence (Berg et al. 1627) or the leaf shape (Morawetz et al. 11-11988).
The spikes of the staminate inflorescences can be green before anthesis (as in Berg et al. 1597). In most specimens the apex of the perianth of the pistillate flower is plane with a rim around the round aperture and ? angular in circumference. In some collections (e.g., Bernardi 1612 and Liesner 3481) the apex of the perianth of the pistillate flower is (strongly) con- vex, circular in outline, and with a slit-shaped aper- ture. Some collections have up to 12 lamina segments and/or submarginally loop-connected lateral veins; these features can be related to the (sub)juvenile state.
10. Cecropia elongata Rusby, Bull. New York Bot. Gard. 4: 446. 1907. Type. Bolivia. Without locality (probably Prov. Nor Yungas, Coripata), (6d), Bang 2260 (holotype: NY; isotypes: BM, G, GH, K, LE, MO, US, WIS).
Tree, to 15 m tall. Leafy twigs 2-9(-15) cm thick, purplish brown, puberulous to hispidulous, with straight hairs and also dense brown pluricellular tri- chomes. Lamina chartaceous to subcoriaceous, ca. 30 x 30 cm to 90 x 90 cm (to 105 X 105 cm), the segments (8-)10-16, oblanceolate, the incisions down to 7/10-9/10; apices subacute to acuminate; up- per surface ? scabrous, hispidulous; lower surface puberulous to subtomentellous to subtomentose on the smaller veins to almost glabrous on the main
veins, with arachnoid indumentum in the areoles and on the smaller veins; lateral veins in the free part of the midsegment (18-)20-25(-40) pairs, submargin- ally (in the lower part of the segment faintly) loop- connected, unbranched; petiole 20-80(-105) cm long, sparsely puberulous to hispidulous to subglabrous; trichilia fused, the brown indumentum intermixed with short white hairs; stipules ca. (10-)20-40 cm long, red-brown to greenish, sparsely strigillose and with dense brown pluricellular trichomes or also with arachnoid indumentum outside, (sparsely) sericeous inside. Staminate inflorescences in pairs, the peduncle erect, the spikes ? spreading to pendulous; peduncle ca. 7-25 cm long, hirtellous to subhirsute; spathe at least 15 cm long, color unknown, densely brownish to whitish subhirtellous to subsericeous outside, ? sparsely hairy inside; spikes 13-25, 18-27 X ca. 0.4- 0.6 cm, the stipes 0.4-1 cm and glabrous or sparsely hairy; rachis sparsely hairy. Staminate flowers: peri- anth tubular, 1.5-2 mm long, with sparse to dense short arachnoid indumentum below the apex, the apex slightly convex to plane, punctate to smooth, or ini- tially also with brown pluricellular trichomes; fila- ments flat; anthers ca. 0.6-0.8 mm long, without ap- pendages, remaining attached to the filaments at anthesis. Pistillate inflorescences in pairs or solitary, erect to spreading to subpendulous in fruit; peduncle (10-)20-25 cm long, sparsely to rather densely hir- tellous to subhirsute; spathe at least 20 cm long, the color unknown, the indumentum as in the staminate inflorescence or more densely hairy inside; spikes 4- 9, ca. 25-40 X 1 cm, to 47 X 1.5 cm in fruit, the stipes 0.3-1.5 cm long and glabrous; rachis glabrous. Pistillate flowers: perianth ca. 1.5-2 mm long, with arachnoid indumentum below the apex outside, ab- sent inside, the apex convex, punctate to muriculate, the aperture slit-shaped; style long, straight; stigma comose. Fruit ellipsoid to oblongoid, 2-2.5 mm long, tuberculate, brown.
Distribution (Fig. 18.1). Bolivia (Cochabamba, La Paz, and Santa Cruz), in montane forest, at 1600- 2400 m.
Specimens examined. BOLIVIA. COCHABAMBA: Rd. Coachabamba-Villa Tunari, ca. km 84, 2400 m, 6 Dec 1995 ( fl-fr), Berg et al. 1722 (BG, BOLV, COL, LPB), ca. km 102, 2000 m, 6 Dec 1995 (d), Berg et al. 1726 (BG, BOLV, COL, LPB, USZ). LA PAZ: Prov. Nor Yungas, rd. Yolosa- Chuspipata, 2000, 11 Mar 1987 (6), Beck 13549 (BG, LPB, MO); Prov. Sud Yungas, rd. Huancane-San Isidro, km 7, 2250, 11 Feb 1991 (st), Beck 19723 (BG); Chulumani, 1800 m, Aug 1933 (6), M. Cdrdenas 1165 (NY); Prov. Inquisivi, Khora-Mikilpirhua, 24 Km N of Choquetanga, 1900 m, 14 Jul 1994 (6), Salinas 3145 (BG); Prov. SudYungas, 2.5 km
76 FLORA NEOTROPICA
NW of Yanacachi, 2120 m, 30 May 1987 (6), Seidel 994 (LPB); Prov. Nor Yungas, 13.5 km NW of Chuspipata, rd. to Yolosa, 2100 m, 5 Oct 1984 ( Y fr), Solomon et al. 12459 (BG, LPB, MO); valley of Rfo Zongo, 1650 m, 29 Apr 1981 (9 fl-fr), Vdzquez Avila 354 (LPB, RB), 18 Nov 1981 (9 fl-fr), Zuloaga et al. 1833 (LPB). SANTA CRUZ: Prov. Flor- ida, 5 km E of Santa Rosa de Lima, 1470 m, 3 Jul 1996 (6), Saldias et al. 4363 (NY); Prov. Caballero, K'arawasi, 1900 m, 20 Jul 1996 (st), Saldfas et al. 4442 (USZ).
This species certainly does not belong to the group of montane species (including the white-leaved ones) found more northward in the Andes. It might be re- lated to Cecropia angustifolia. The stems and
branches are often conspicuously broadened toward the apices. The apex of the terminal bud cover (the stipules) is often torn off by elongation of the corre- sponding leaf. Flowering (and fruiting) might be largely seasonal. Populations examined in Bolivia (Cochabamba) in December (1995) had only very few trees with staminate inflorescences and the pistillate inflorescences were largely in the same state of de- velopment. The trees are inhabited by ants.
11. Cecropia engleriana Snethlage, Notizbl. Bot.
Gart. Berlin-Dahlem 8: 365. 1923; Berg, Acta Amazonica 8(2): 166. 1978. Cecropia concolor Willdenow subsp. engleriana (Snethlage) C. C. Berg & P. Franco, Fl. Ecuador 48: 17. 1993. Type. Brazil. Acre: Rio Acre, Seringal Sao Francisco, Jun 191 1, Ule 9313 (holotype: B (with staminate inflorescences), destroyed; isotypes: G, K; under the same number in MG a specimen with pistillate inflorescences). Fig. 17.3-10
Cecropia pacis Cuatrecasas, Revista Acad. Colomb. Ci. Exact. 6(22/23): 288. 1945. Type. Colombia. Putu- mayo: Rio Putumayo, between mouth of Rio Gua- mues & Puerto Asis, Cuatrecasas 11244 (holotype: COL; isotype: F).
Cecropia yarinensis Cuatrecasas, Revista Acad. Col- omb. Ci. Exact. 9(36/37): 339. 1956. Type. Peru. Ucayali: Prov. Coronel Portillo, Pucallpa, 4 km from Yarina, rd. to Huanuco, 8 Apr 1953 (9), Ferreyra 9044 (holotype: US; isotype: UMS).
Tree, to 20(-30) m tall; trunk with prominent stip- ular scars. Leafy twigs 1.5-4 cm thick, green with yellow spots to reddish to purplish brown, hispidu- lous, with curved or uncinate hairs, or also with sparse to rather dense arachnoid indumentum. Lamina sub- coriaceous to chartaceous, ca. 30 X 30 cm to 65 X 65 cm, the segments (7-)8-10(-1 1), the free parts of the upper segments oblong to subobovate to oblan- ceolate, entire or ? sinuate (to lobate), the incisions
down to (6/1-)8/10-9/10 (2-4 cm, or sometimes 8 cm from the petiole); apices short-acuminate to (sub)acute to obtuse; upper surface smooth to scabri- dulous, sparsely puberulous to subhispidulous, also with sparse arachnoid indumentum, along the main veins often with brown pluricellular hairs; lower sur- face minutely puberulous on the main veins, with arachnoid indumentum in the areoles, on the smaller veins and/or at least initially also on the main veins; lateral veins in the free part of the midsegment (21-) 24-32 pairs, marginally loop-connected, mostly branched; petiole 25-50 cm long, sparsely puberulous and/or also with sparse to (rather) dense arachnoid indumentum; trichilia fused, the brown indumentum intermixed with (sparse) short white hairs; stipules 8- 25 cm long, white to pale yellow or pinkish, caducous or occasionally subpersistent, puberulous or sparsely sericeous, often with ? dense arachnoid indumen- tum, sometimes (also) with ? dense brown pluricel- lular hairs outside, ? densely sericeous inside. Sta- minate inflorescences in pairs or solitary, pendulous; peduncle (3.5-)10-19 cm long, sparsely puberulous to hirtellous and with sparse to rather dense arachnoid indumentum; spathe 10-20 cm long, white to yellow- ish or pinkisk, puberulous to hirtellous and often with dense arachnoid indumentum outside, glabrous or (densely) sericeous inside; spikes (8-)15-20, (1-) 10- 20 X 0.2-0.4 cm, the stipes 0.2-2 cm long, hairy or subglabrous; rachis hairy. Staminateflowers: perianth tubular, 0.8-1.5 mm long, glabrous, the apex slightly convex; filaments flat, anthers 0.5-0.6 mm long, ap- pendiculate, detached at anthesis, reattached to the margins of the aperture by the appendages. Pistillate inflorescences in pairs or solitary, pendulous; pedun- cle (3.5-)6-22 cm long, puberulous to hirtellous and with sparse to rather dense arachnoid indumentum; spathe 10-17 cm long, the color and indumentum as in the staminate inflorescence; spikes 4, (2-)5-18 X
0.4-1 cm, to 37 X 1-1.5 cm in fruit, sessile or with stipes to 1(-1.5) cm long and hairy. Pistillateflowers: perianth ca. 2 mm long, with arachnoid indumentum on and below the apex outside, absent inside, the apex plane to slightly concave or slightly convex; style short; stigma peltate (to subpelate or tongue-shaped). Fruit ellipsoid to oblongoid to ovoid, 1.5-2.5(-3) mm long, ? tuberculate or smooth.
Distribution (Fig. 10.4). Upper Amazon basin, from Colombia to Bolivia, often, at least in the north- ern part of its range, associated with riverine habitats, at low elevations.
cayacu, Quebrada de Agua Pudre, 19 Nov 1991 (st), Pipoly et al. 16410 (COL, MO), 22 Mar 1992 (9 fr), Rudas et al. 3489 (COL, MO); Rio Caraparana, nr. El Encanto, 22-28 May 1942 (9 fl-fr), Schultes 3821 (COL, ECON, F); Dtto. Puerto Narifno, Parque Nacional Amacayacu, 9 Oct 1989 (6), R. Vdsquez et al. 12685 (BG, COL, MO). CAQUETA:
Mun. Florencia, rd. to Gabinete, km 1, 18 Oct 1993 (9 fl- fr), Franco et al. 4516 (BG), (6), Franco et al. 4517 (BG). CAUCA: Rd. Mocoa-Pitalito, nr. Rio Caqueta, 18 Feb 1995 (9 fl-fr), Franco et al. 4690 (COL). PUTUMAYO: Between Mocoa & Urcusique, 9 Jan 1945 (9 fl-fr), Ewan 16732 (F, US); Mun. Mocoa, rd. Mocoa-Villa Garz6n, El Pepino, 18 Feb 1995 (d), Franco et al. 4687 (COL, HUA, LP, MO, NY, US), (6), Franco et al. 4688 (COL, HUA); rd. Mocoa- Puerto Asis, 3 May 1994 (9 fl-fr), Franco et al. 5517 (COL); rd. Garz6n-Puerto Asis, 3 May 1994 (6), Franco et al. 5520
Batata, 2 km NE of Salto Colorado, 55 km SE of Puerto Ayacucho, Oct 1989 (9), Sanoja et al. 3329 (BG); Rio Ca- siquiare, nr. Rio Orinoco, Jan-Feb 1969 (9 fl-fr), Velasquez et al. 501 (US) = Farinias et al. 501 (VEN), (d), Velasquez et al. 506 (US) = Farifnas et al. 506 (VEN); Upper Rio Orinoco, La Esmeralda, 19 May 1942 (9 fr), LI. Williams 15516 (US).
ECUADOR. MORONA-SANTIAGO: Centro Shuar Yu- kutis, 22 Apr 1989 (st), Bennett et al. 3801 (QCNE); rd. Zamora-Gualaquiza, Estaci6n Exp. El Padmi, ca. 5 km N of Los Encuentros, 4 Jan 1991 (6), Berg et al. 1657 (BG, LOJA, QCA). NAPO: Nr. Tena, 7 Jan 1940 (9 fl-fr), Asplund 10290 (S); Rio Napo, Descanso, 25 km from Coca, 1 Mar 1980 (9 fl-fr), Berg et al. 1104 (AAU, BG, COL, GB, MO, QCA, U); Parque Nacional Yasuni, Pozo petrolero Daimi 2, 26 May-8 Jun 1988 (9 fr), Cer6n et al. 3847 (BG, MO, QCNE); Afiangu, Parque Nacional Yasuni, 1-15 Feb 1986 (st), Korning et al. 47692 (AAU, BG, QCA); Reserva Biol- 6gica Jatun Sacha, 8 km E of Misahualli, 25 Dec 1990 (6), Neill et al. 9742 (BG, QCA, QCNE); nr. Reserva Biol6gica Jatun Sacha, rd. Puerto Napo-Ahuano, 25 Dec 1990 (d), Neill et al. 9744 (BG, QCA, QCNE). SUCUMBiOS: Rio Aguarico, ca. 0?0', 76'40-45'W, 22 Feb 1980 (d), Berg et al. 1075 (AAU, BG, COL, GB, K, MO, NY, QCA, TUR, U, WIS); 17 km W of Lumbaqui, 4 Nov 1974 (9 fl-fr), Gentry 12442 (MO, QCA, U).
PERU. AMAZONAS: Quebrada Cikan-inci, W of Huam- pami, 18 Dec 1972 (9 fl-fr), Berlin 592 (AAU, MO, NY, USM); Prov. Bagua, Dtto. Imaza, Rio Marafi6n, Yamayakat, 6 Mar 1966 (9 fl-fr), N. Jaramillo et al. 579 (MO); Rio Cenepa, Quebrada Huampami, 29 May 1973 (9 fr), Kayap 832 (BG, F, NY); Quebrada Satik-entsa, 16 Jul 1974 (d), Kayap 1099 (MO, U, USM); Rio Cenepa, nr. Huampami, ca. 5 km E of Chavez Valdivia, 3 Aug 1978 (9 fl-fr), Kujikat 132 (BG, MO). Cuzco: Prov. Paucartambo, rd. Pilcopata- Paucartambo, km 16, 2 Aug 1988 (9 fl-fr), Berg et al. 1629 (BG, COL, Centro de Medicina Andina, Cuzco). HUANUCO: Prov. Pachitea, ca. 26 km S of Puerto Inca, 27-28 Nov 1988 (st), Johann 2/26 (BG), 2/35 (BG); Prov. Puerto Inca, Dtto. Yupapichis, DANTAS, 16-31 Mar 1991 (d), Tello 1583 (G, NY), 1-15 Apr 1991 (9 fl-fr), Tello 1731 (G, NY); Shapa-
jilla, 28 May 1939 (6), Woytkowski 23 (F). LORETO: Prov. Maynas, Dtto. Indiana, Explorama Inn, 21 Apr 1992 (Y fl- fr), Grdndez et al. 4091 (MO); Jenero Herrera, 25 Nov 1987 (Y fl), Lamotte 325 (BG), (6), Lamotte 326 (BG); Rfo Am- piyacu, nr. Pebas, 24 Apr 1977 (Y fl), Plowman 7020 (ECON, F, US); mouth of Rio Santiago, s.d. (Y fl-fr), Tess- mann 4058 (G, NY, US); Iquitos, s.d. (Y fr), Tessmann 5044 (NY, US); Prov. Alta Amazonas, Andoas, 20 Nov 1980 (6), R. Va'squez et al. 776 (NY); Prov. Maynas, Pto. Alianza, (Qda. Tamshiyacu), 29 May 1981 (Y fl-fr), R. Vdsquez et al. 1856 (BG, MO). MADRE DE DIos: Prov. Tambopata, Comunidade Nativa Infierno, Hermosa Chica, 14 Feb 1989 (6), Alexiades et al. 290 (BG); Prov. Tambopata, 30 km SSW of Puerto Maldonado, confluence of Rio La Torre & Rio Tambopata, 26 Jun 1980 (Y fr), Barbour 5820 (BG, MO); Prov. Manu, nr. Shintuya, 30 Jul 1988 (Y fl-fr), Berg et al. 1610 (BG, COL, Centro de Medicina Andina, Cuzco); Prov. Manu, Manu Park, Cocha Cashu, 8 Sep 1986 (st), Niu- niez 6044 (BG), Aug-Sep 1989 (st), Nuliiez et al. 11276 (MO). SAN MARTiN: Prov. Lamas, rd. Lamas-Pamashto, 4 Dec 1997 (Y fl-fr), Berg et al. 1765 (BG, COL, MOL), (d), Berg et al. 1766 (BG, COL, MOL); Prov. Moyobamba, rd. Moy- obamba-Rioja, ca. km 10, 8 Dec 1997 (Y fl-fr), Berg et al. 1792 (BG, COL, MOL); rd. Rioja-Nueva Cajamarca, ca. km 5, 8 Dec 1997 (6), Berg et al. 1793 (BG, COL, MOL); rd. Tarapoto-Juanjui, 7 Oct 1984 (Y fl-fr), Maas et al. 5983 (U). UCAYALI: Nueva Requena, 11 km from Campo Verde (km 30 on rd. Pucallpa-Lima), 4 May 1988 (Y fl-fr), Arana 7 (K), 11 Aug 1988 (6), Berg et al. 1636 (BG, COL, K); Prov. Coronel Portillo, nr. Pucallpa, 4 km from Yarina, rd. to Huanuco, 8 Apr 1953 (6), Ferreyra 9045 (USM); Prov. Coronel Portillo, nr. Pucallpa, Yarinacocha, 19 Dec 1968 (d), Ferreyra 17629 (US, USM).
BRAZIL. ACRE: Mun. Marechal Taumaturgo, Rio Ju- rua, Fazenda Paraguay, 3 Apr 1993 (6), Daly et al. 7726 (NY); Mun. Madureira, Rio Macaua, Col6nia Bom Futuro, 30 Mar 1994 (Y fl-fr), Lima et al. 529 (NY). AMAZONAS:
Mun. Sao Paulo de Oliven,a, nr. Palmares, 11 Sep-26 Oct 1936 (6), Krukoff 8061 (A, BM, F, G, K, LE, MICH, MO, NY, P, S, U, US), (Y fl), Krukoff 8336 (A, BM, F, G, K, LE, MO, NY, S, U). ROND6NIA: Parque Indigena Aripuana, Jan 1982 (Y fl-fr), Coimbra 71B (MG); Estrada de Ferro Ma- deira, Mamore, 10 Sep 1923 (6), Kuhlmann 407 (RB).
BOLIVIA. BENI: Prov. Ballividn, Misi6n Fatima, San Borja, Rio Maniqui, 28 May 1988 (6), Beck et al. 16662 (BG, LPB, MO); Prov. Moxos, 130 km S of San Ignacio, 10 Sep 1991 (st), Del Aguila et al. 90 (LPB, MO); Prov. Bal- livian, Triunfo, 27 Jul 1976 (Y fl), Meneces et al. 45 (BOLV, MO); Prov. Ballividn, rd. Yucumo-Rurrenabaque, 20-25 Jun 1989 (9 fr), D. N. Smith et al. 13564 (BG, BOLV, LPB, MO, USZ); Prov. Ballivian, rd. Yucumo-Rurrenabaque, km 34, 7-14 Jul 1990 (6), D. N. Smith et al. 14126 (BG). Co- CHABAMBA: Rd. Santa Cruz-Villa Tunari, km 240, 12-14 Jul 1989 (9 fr), D. N. Smith et al. 13713 (BG, LPB, MO, USZ). LA PAZ: Prov. Iturralde, Luisita, 22 Feb 1984 (9 fl- fr), Beck et al. 9931 (BG, LPB, MO, USZ); Prov. Sud Yun- gas, rd. Sapecho-Yucumo, km 1.5, 28 Feb 1994 (9 fl-fr), Berg 1700 (AAU, BG, COL, LPB, NY); Prov. Sud Yungas, nr. Sapecho, 3 Mar 1994 (6), Berg 1710 (BG, COL, LPB,
78 FLORA NEOTROPICA
NY); Prov. Sud Yungas, Serrania de Marimonos, Colonia Tarapaca', 23 Jul 1993 (Y fl-fr), Seidel et al. 7316 (BG, LPB). PANDO: 54 km SW of Cobija, Triunfo, 23 Jul 1980 (Y fl- fr), Pennington et al. 31 (BG, F, K, LPB). SANTA CRUZ:
Prov. Ichilo, Reserva Forestal Chore, Rio Itabo, 16-18 Aug 1990 (Y fl-fr), Neill et al. 9345 (LPB, MO, QCNE).
The trees of the species are normally small to medium-sized but may sometimes, as in flood-plain forest (in Ecuador), become up to 30 m tall. In areas where Cecropia engleriana and C. concolor co-occur, Bolivia and southem Peru (Cuzco and Madre de Dios), more or less clearly transitional features can be found in C. engleriana, such as subpersistent stipules (Cuzco), some of the incisions of the lamina down to near the petiole, and the number of lateral veins in the free part of the midsegment in the range normal in C. concolor. In some specimens collected in San Martin, the lamina have ca. 21 pairs of lateral veins in the free part of the relatively short midsegment.
12. Cecropia ficifolia Snethlage, Notizbl. Bot. Gart. Berlin-Dahlem 8: 365. 1923; Berg, Acta Amazon- ica 8(2): 171. 1978. Type. Brazil. Acre: Rio Jurua'- Mirim, Jun 1901, Ule 5588 (holotype: B (with sta- minate inflorescences), destroyed, in G, K, L, and MG specimens with pistillate inflorescences, the one in MG here designated as the lectotype).
Cecropia mocoana Cuatrecasas, Bot. Mus. Leafl. Har- vard Univ. 14: 24. 1949. Type. Colombia. Putumayo: Nr. Mocoa, 3-7 Dec 1942 (ci), Schultes & C. E. Smith 2003 (holotype: F).
Cecropia porvenirensis Cuatrecasas, Bot. Mus. Leafl. Harvard Univ. 14: 25. 1949. Type. Colombia. Putu- mayo: Puerto Porvenir, nr. Puerto Ospina, 20 May 1942, Schultes 3697 (holotype: F, consisting of leaves of C. ficifolia, here designated as the lecto- type; isolectotype: GH) and pistillate inflorescences of C. membranacea; in COL only pistillate inflores- cences of C. membranacea.
Cecropia ferreyrae Cuatrecasas, Revista Acad. Colomb. Ci. Exact. 9(36/37): 338. 1956. Type. Peru. Ucayali: Nr. Pucallpa, Ferreyra 9032 (holotype: US, mixed collection, a leaf belonging to C. ficifolia, herewith designated as the lectotype, and a staminate inflores- cence belonging to C. membranacea).
Tree, to 15(-20) m tall. Leafy twigs 2-6 cm thick, green, subhispid to strigose (with uncinate hairs) to subhirsute, occasionally glabrous. Lamina charta- ceous, ca. 45 X 45 cm to 90 X 90 cm, the segments 6-8, the incisions down to ca. (3/10-)5/10-7/10(-8/ 10), if the incisions beyond 5/10, then the free part of the midsegment broadly spathulate to obovate; apices obtuse to rounded, sometimes shortly acuminate; up- per surface scabrous (or scabridulous), hispidulous to substrig(ill)ose (or to hirtellous); lower surface sparsely to densely (sub)hirsute (to subvillous) on the veins, with arachnoid indumentum in the areoles (sometimes very sparse), often also on the smaller veins, sometimes extending to the main veins; lateral veins in the free part of the midsegment 6-12, sub- marginally loop-connected, mostly unbranched; pet- iole 35-80 cm long, subhispid to subhirsute (to sub- villous) and with short arachnoid indumentum, occasionally glabrous; trichilia fused, the brown in- dumentum intermixed with dense rather long white hairs; stipules (12-)18-25(-35) cm long, reddish or greenish to yellowish (to orange) or whitish, subser- iceous to subhirsute (to subvillous) and with sparse or sometimes with very dense arachnoid indumentum outside, densely (sub)sericeous inside. Staminate in- florescences in pairs, the peduncle erect, the spikes pendulous; peduncle 6-12(-18) cm long, hispidulous to (sub)hirsute, occasionally glabrous; spathe 10-18 cm long, green to pale yellow or whitish, substrigose to subsericeous to subhirsute (to subvillous) and with sparse arachnoid indumentum outside, pilose to sub- sericeous inside; spikes 8-13, 8-10 X 0.3-0.5 cm, with stipes to 1.5 cm long and hispidulous; rachis hairy, the hairs relatively long. Staminateflowers ses- sile or short-pedicellate; perianth tubular, ca. 1.5-2 mm long, sometimes with sparse and short arachnoid
SYSTEMATIC TREATMENT 79
FI.2.Cecropia ficifolia. 1. Lamina, reduced (Franco et al. 4519). 2. Lamina, reduced. 3. Apex of lamina and venation (Gentry et al. 39501). 4. Stipules (Croat 18177). 5. Base of petiole with trichilium (Palacios 5673). 6. Staminate inflorescence with spathe. 7. Staminate inflorescence at anthesis and base of petiole with trichilium. 8. Staminate flower. 9. Stamen (Berg et al. 1066). 10. Pistillate inflorescences with spathes and base of petiole with trichilium. 11. Pistillate inflorescence at anthesis. 12. Pistillate inflorescence after anthesis (Berg 1224). 13. Pistillate flowers. 14. Fruit (Berg et al. 1637).
80 FLORA NEOTROPICA
indumentum below the apex, the apex plane and gla- brous; filaments flat; anthers ca. 0.6-0.8 mm long, appendiculate, detached at anthesis, reattached to the margins of the aperture by the appandages. Pistillate inflorescences in pairs, erect, becoming pendulous in fruit, occasionally subtended by caducous bracts, to 6 cm long; peduncle (5-)9-21 cm long, hispidulous to (sub)hirsute, occasionally glabrous; spathe 9-14 cm long, the color and indumentum as in the stami- nate inflorescence; spikes 4-5(-8), (2-)6-12 X 0.6- 0.8 cm, to 25 X 1.7 cm in fruit, sessile or sometimes with stipes to 0.8 cm long and puberulous; rachis hairy, the hairs relatively long. Pistillate flowers: per- ianth 2.5 mm long, with arachnoid indumentum be- low the apex outside, also in the lower part of and below the style channel inside, the apex slightly con- vex to plane or in fruit ? concave, muriculate, the aperture circular, surrounded by a low rim; style rather long, S-shaped or straight; stigma penicillate. Fruit oblongoid, ca. 1.5-2.5 cm long, ? tuberculate.
Distribution (Fig. 10.2). Upper Amazon basin and the llanos region of Colombia, in forest, in par- ticular in the southem part of the range of distribution common in secondary growth, in non-inundated places, at elevations to 1200 m.
Representative specimens examined. COLOMBIA. AMAZONAS: Leticia, Quebrada de Arara, 27 Jan 1969 (Y fl- fr), Cuatrecasas et al. 27244 (COL, K, NY, US); Villa Bit- tencourt, nr. Colombia-Brazil border, 5-6 Oct 1952 (st), Garcia-Barriga 14722 (COL); between Leticia & Rio Amaca-yacu, nr. Isla Santa Soffa, 15 Aug 1972 (Y fl), Id- robo 6520 (AAU, COL); Mun. Leticia, Parque Nacional Natural Amacayacu, nr. Quebrada de Agua Pudre, 21 Mar 1992 (Y fl), Rudas et al. 3353 (COL, MO); Rfo Igara- parana, nr. Cgto. La Chorrera, 7 Oct 1973 (d), Sastre 2404 (COL, G, U), 4 Jun 1974 (6), Sastre 3110 (COL, G, U). AMAZONAS/VAUPES: Rfo Apaporis, Jino-Goje, between Rfo Piraparana & Rfo Popeyaka, 3-11 Sep 1952 (Y fl-fr), Garcia-Barriga 14442 (US); Rfo Apaporis, Soratama, be- tween Rfo Pacoa & Rio Kananari, 11 Jul 1951 (6d), Schultes et al. 13028 (US); Rio Apaporis, Raudal de Jinjirimo, 25 Nov 1951 (6), Schultes et al. 14555 (COL, GH, U, US). CAQUETA: Nr. Araracuara, 11 Jul 1986 ( fl-fr), Berg et al. 1550 (BG, COL); Florencia, 29 Mar 1940 (Y fl-fr), Cuatre- casas 8816 (COL, F); Rio Orteguaza, San Luis, Quebrada de Miramar, 15 Jan 1969 (Y fl-fr), Cuatrecasas et al. 27125 (COL, F, G, K, NY, US), (6), Cuatrecasas 27126 (COL, F, K, NY, US); Florencia, between Rfo Hacha & Rfo La Yuca, 25 Jan 1968 (Y fl-fr), Cuatrecasas et al. 27236 (COL, P, US); Mun. Florencia, rd. to Gabinete, km 1.5, 18 Oct 1993 (9 fl-fr), Franco et al. 4519 (BG, TULV); Solano, 8 km SE of Tres Esquinas, Rfo Caqueta, below mouth of Rfo Orte- guaza, 2 Mar 1945 (9 fl-fr), Little et al. 9498 (COL, F, P, US); Rfo Orteguaza, 9 km S of Florencia, 25 Jan 1969 (9 fl-fr), Plowman et al. 2272 (GH, K, S, US); Morelia, 22 Nov
1941 (9 fl), Sneidern A.1352 (A, K, LL, MICH, NY, US). GUAINIA: Serrania de Naquen, Maimachi, 11 Apr 1993 (9 fl), Madrinidn et al. 982 (COL), (c), Madrinidn et al. 1000 (COL). META: Mun. Puerto L6pez, Alto Menagua, 10 Feb 1995 (9 fl-fr), Franco et al. 4658 (BG, COL, HUA, MO, US); nr. La Macarena, Rio Moccoroy, 11 Feb 1995 (6), Franco et al. 4664 (BG, COL, HUA, MO, US); Sabanas de San Juan de Arama, Rio Guejar, Los Micos, 22 Jan 1951 (9 fl-fr), Idrobo et al. 1216 (COL, GH, U, US); Sierra La Ma- carena, mouth of Rio Duda, 7 Oct 1975 (9 fl-fr), Idrobo 8394 (AAU, COL); Sierra de La Macarena, Rfo Guapaya, 29 Nov 1949 (9 fr), Philipson et al. 1623 (BM, COL, US). PUTUMAYO: Rio Putumayo, Pifiunia Negra, 20 Nov 1940 (d fl), Cuatrecasas 10702 (COL); Mun. Puerto Asis, rd. to Kan- akas, 16 Feb 1995 (9 fl-fr), Franco et al. 4682 (COL); Mo- coa, between La Campucana & San Antonio, 1000 m, 2 May 1994 (9 fl-fr), Franco et al. 5511 (BG, COL). VAUPES: Rio Unilla, Calamar, 30 Oct 1939 (9 fl-fr), Cuatrecasas 7328 (COL, US); Rio Guaviare, San Jos6 del Guaviare, 6 Nov 1939 (9 fl-fr), Cuatrecasas 7479 (COL, US); nr. Miraflores, 6-29 Jan 1944 (9 fl-fr), Gutierrez V et al. 542 (COL, F, GH, MEDEL); nr. Miti, 17 Sep 1951 (i), Schultes et al. 13904 (COL, US); Rio Apaporis, Jinogoj6, 8 Mar 1952 (9 fl-fr), Schultes et al. 15876 (US).
VENEZUELA. AMAZONAS: Depto. Rio Negro, Rio Mawarinuma, 7 km ESE of Puerto Chimo, 9-14 Jul 1984 (9 fl-fr), Davidse et al. 27161 (BG, NY); Depto. Rio Negro, Neblina Base Camp, on Rio Mawarinuma, 17 Jul 1984 (G), Davidse et al. 27450 (BG, MO, NY); 8 km NE of San Carlos de Rio Negro, 1 Dec 1977 (9 fl-fr), Liesner 4076 (MO, U, VEN); 12 km NE of San Carlos de Rio Negro, 14 Apr 1979 (d), Liesner 6617 (MO, U, VEN); 20-25 km SE of Puerto Ayacucho, 4 Nov 1980 (i), Maas et al. 5105 (F, K, NY, U, VEN); Rio Casiquiare, just above Capibara, 2 Apr 1953 (9 fl), Maguire et al. 34787 (BG); Depto. Atures, rd. Gavilan- Rfo Cataniapo (damsite), 11 May 1980 (i), Steyermark et al. 122299 (MO, U, VEN); Depto. Atabapo, nr. Culebra, Rio Cunucunuma, 22-29 Mar 1983 (6), Steyermark et al. 129060 (COL, U, VEN); Yavita, 26 Jan 1942 (9 fl), LI. Wil- liams 13983 (G, US, VEN); Solano, Rfo Casiquiare, 12 Mar 1942 (9 fl-fr), Ll. Williams 14760 (F, G, US); Rio Guainia, Yavita-Pimichin trail, nr. Pimichin, 2 Jul 1959 (6), Wurdack et al. 43287 (BG, NY, US).
ECUADOR. MORONA-SANTIAGO: Nr. Lim6n (Gral. Plaza), 1200 m, 27 Jan 1981 (9 fr), Berg 1224 (BG, COL, QCA, U); rd. Lim6n (Gral. Plaza)-La Uni6n, km 7-10, 18 Feb 1987 (9 fl-fr), Bohlin et al. 1468 (BG, GB, QCA, S); rd. Sucua-Macas, km 10, 24 Sep 1979 (6), Holm-Nielsen et al. 20426 (AAU); 6 km SE of Gualaquiza, 16 Sep 1975 (9 fl-fr), Little et al. 386 (COL, LOJA, Q, QAME, QCNE). NAPO: Tena, 10 Oct 1939 (9 fl-fr), Asplund 9198 (S); rd. Coca-Auca, 26 Feb 1980 (9 fl-fr), Berg et al. 1089 (AAU, BG, COL, QCA, U); Rio Napo, 15 km W of Coca, 18-20 Apr 1985 (9 fl), Neill et al. 6336 (BG, MO, QCNE); Ani- angu, Parque Nacional Yasunf, nr. Tiputini trail, 14-22 Jan 1985 (9 fl-fr), 0llgaard et al. 57100 (AAU, BG, QCA); Reserva Biol6gica Jatun Sacha, 8 km E of Puerto Misahualli, 1-15 Sep 1987 (9 fl-fr), Palacios 1891 (BG, GB, K, MO, NY, QAME, QCNE); Cant6n El Chaco, Codo Sinclair, 16-
SYSTEMATIC TREATMENT 81
20 Sep 1990 (9 fl-fr), Palacios 5673 (BG, QCNE). PAS- TAZA: Mera, ca. 1100 m, 13 Dec 1955 (9 fl-fr), Asplund 18829 (S); nr. Puyo, 19 Dec 1955 (9 fl-fr), Asplund 18892 (S); nr. Mera, 1160 m, Harling 3809 (S). SucuMBios: Rio Cuyabeno, ca. 0?10'S, 75?55'W, 20 Feb 1980 (d), Berg et al. 1066 (AAU, BG, COL, GB, MO, NY, QCA, TUR, U, WIS), 20 Feb 1980 (9 fl-fr), Berg et al. 1071 (AAU, BG, COL, NY, QCA, U); Lumbaqui, ca. 1000 m, 13 Aug 1975 (9 fl-fr), Little et al. 167 (Q, QAME, QCNE); Reserva Faun- istica Cuyabeno, between Tarapoa & Tipishca, Bellavista, 14 Nov 1991 (9 fl-fr), Palacios et al. 8884 (BG, QCNE). ZAMORA-CHINCHIPE: Rfo Nangaritza, nr. Nangaritza, 4 Dec 1990 (9 fl-fr), Neill et al. 9504 (BG, QCA, QCNE).
PERU. AMAZONAS: Rio Cenepa, 10 km E of Huam- pami, 10 Oct 1972 (9 fl), Berlin 229 (USM); Rio Santiago, nr. Caterpiza, 29 Aug 1979 (6), Huashikat 253 (MO), 6 Oct 1979 (9 fl-fr), Huashikat 793 (MO); Quebrada Kayamas, 3 Mar 1973 (9 fl-fr), Kayap 442 (BG, MO). HUANUCO: Puerto Bermddez, 14-17 Jul 1929 (st), Killip et al. 26614 (NY); Prov. Pachitea, Dtto. Honoria, Rio Pachitea, rd. to Ayamiria, 27 Dec 1966 (9 fl-fr), Schunke V 1417 (F, MOL); Prov. Pachitea, Dtto. Honoria, Isla del Pacanasi, 14 Nov 1967 (d), Schunke V 2313 (F, US). LORETO: Rd. Iquitos- Nauta, km 2, 4 Jul 1988 (9 fl-fr), Berg et al. 1573 (BG, USM); 4 km SW of Iquitos, 15-16 Jul 1972 (6), Croat 18128 (BG, MO); Prov. Requena, Jenaro Herrera, 9 Dec 1977 (9 fl-fr), Gentry et al. 21300 (F, MO, USM), (d), Gentry et al. 21333 (MO, USM); Prov. Maynas, Rio Am- piyacu, Pebas, 30 Mar 1977 (6), Plowman et al. 6495 (F, US, USM); Prov. Maynas, Rio Yaguayacu, Brillo Nuevo, 12 Apr 1977 ( 9 fl-fr), Plowman etal. 6805 (F, US, USM); Prov. Maynas, Rio Nanay, Mishana, 27 Oct 1980 (9 fl-fr), R. Vds- quez et al. 676 (MO, NY, TEX, USM); Prov. Alto Ama- zonas, Yurimaguas, 18 Sep 1981 (d), R. Vdsquez et al. 2568 (F, MO. MADRE DE Dios: Parque Nacional Manu, Rio
Manu, above Rio Sotileja, 11 Oct 1986 (9 fl), Foster et al. 11777 (BG, LPB, US, USM); Prov. Tambopata, Reserva Tambopata, 15 Aug 1990 (9 fl-fr), Reynel et al. 5144 (BG, MO); Prov. Tambopata, Pampas de Heath, 25 Feb 1990 (9 fl-fr), Gentry et al. 69558 (BG, MO); Prov. Tambopata, nr. Sandoval, 15 Jan 1967, Vargas C. 18572 (US). PASCO: Prov. Oxapampa, Panjil, 12 km from Puerto Inca, 27 Sep 1982 (6), D. N. Smith et al. 2427 (MO). SAN MARTiN: Prov. San Martin, rd. Yurimaguas, ca. km 24, 5 Dec 1977 (6), Berg et al. 1776 (BG, COL, MOL), ca. km 27, 5 Dec 1997 (9 fl- fr), Berg et al. 1778 (BG, COL, MOL); Prov. Lamas, rd. Tarapoto-Moyobamba, ca. km 70, 8 Dec 1997 (9 fl-fr), Berg et al. 1789 (BG, COL, MOL); rd. Yurimaguas-Tara- poto, km 50, 12 Oct 1985 ( 9 fl-fr), Gentry et al. 52282 (BG, F, MO); rd. Tarapoto-Yurimaguas, km 68, trail Pintoyacillo- Nuevo Lamas-San Miguel de Shanusi, 2 Oct 1986 (9 fl-fr), Knapp et al. 8482 (BG, QCNE, USM); San Roque, 13 Jan 1930 (9), Ll. Williams 7372 (F). UCAYALI: Nueva Requena, 11 km from Campo Verde (at km 33 rd. Pucallpa-Lima), 11 Aug 1988 (9 fl-fr), Berg et al. 1637 (BG, COL, K, MOL); nr. Pucallpa, (9 fl-fr), Ferreyra 9043 (US, USM); between Aguaytia & San Alejandro, 9 Nov 1964 (9 fr), Ferreyra 16102 (US, USM); rd. Pucallpa-Tingo Maria, Bosque Al-
BRAZIL. ACRE: Nr. Cruzeiro do Sul, 21-28 Aug 1978 (61), Benson 8304 (INPA, UEC, US); Mun. Cruzeiro do Sul, Rio Jurua, Seringal Sao Joao, 14 Mar 1992 (9 fl-fr), Daly et al. 7418 (BG, NY), 19 Mar 1992 (6), Daly et al. 7529 (BG, NY); Rio Acre, Antimary, 30 Mar 1904 (9 fl-fr), Huber MG 4245 (BM, G, MG, U); Cruzeiro do Sul, Estrada Alemanha, 27 May 1971 (9 fl-fr), Maas et al. P13319 (INPA, K, MO, NY, S, U, US). AMAZONAS: Rio Negro, Cucui, 4 May 1975 (6), Cavalcante 3097 (MG), (9 fl-fr), Cavalcante 3098 (MG); Codajas, Ig. do Engenho, 19 Apr 1952, (9 fr), E. Ferreira 58/239 (INPA, U); Mun. Tefe, Jauata, 21 May 1933 (9 fr), Krukoff 4517 (A, G, K, NY, S, U, US); Mun. Humaita, nr. Tres Casas, 14 Sep-Il Oct 1934 (9 fl), Krukoff 6420 (NY); Mun. Sao Paulo de Olivenga, nr. Palmares, 11 Sep-26 Oct 1936 (d), Krukoff 8333 (A, BM, F, K, LE, MICH, MO, NY, P, S, U, US); nr. Uaupes, 10 Feb 1963 (9 fl-fr), Lanna Sobrinho 451 = Castellanos 23756 (GUA, RB, US); Rio Javari, Palmeiras Army Post, 2 Aug 1973 (9 fl-fr), Lleras P 17033 (COL, F, INPA, K, MG, MO, NY, U); Rio Javari, Angamo Garrison, between Angamo & Natua, 5 Aug 1973 (d), Lleras P17161 (INPA, K, NY, U); Rio Negro, Aperecida, Igarape Tuari, 6 Nov 1987 (6), Maas et al. 6934 (INPA, NY); Tapuruquara, 17 Oct 1971 (9 fl- fr), Prance et al. 15375 (INPA); Manacapuru, 4 Apr 1957 (6'), Rodrigues 403 = INPA 5282 (INPA, U). MATO GROSSO: Rio Aripuana, nr. Igarapezinho, 11 Oct 1973 (9 fr), Berg et al. P 18419 (AAU, COL, EAP, F, GH, INPA, K, MG, MICH, MO, NY, P, S, U, US); Nucleo Pioneiro de Humboldt, Rio Aripuana, 14 Oct 1973 (9 fl), Berg et al. P 18526 (F, INPA, K, MG, NY, P, S, U, US); Presidente Marques, Rio Mamore, 11 Oct 1923 (9 fl), Kuhlmann 591 = (RB) 19890 (RB); Nucleo Pioneiro de Humboldt, Rio Aripuana, 9 Oct 1973 (6), Prance et al. 18294 (F, K, NY, S, U). ROND6NIA: Nr. Sao Loren9o mines, 26 Nov 1968 (61), Prance et al. 8910 (INPA, K, MG, MO, NY, S, U, US).
BOLIVIA. BENI: Prov. Vaca Diez, nr. Alto Ivon, Dec 1992 (9 fl-fr), Bergeron et al. 753 (BG), 25 Jan 1984 (9 fl- fr), Boom 4264 (NY); Prov. Vaca Diez, rd. Riberalta-Guay- aramerin, km 20, 20 Apr 1979 (d fl-fr), Krapovickas et al. 35179 (RB); Prov. Vaca Diez, rd. Riberalta-Guayaramerfn, km 3.3, 7 Sep 1981 (9 fl-fr), Solomon 6165 (LPB, MO, U). PANDO: Prov. Manuripi, 12 km W of Conquista-Trampolin, 6 Oct 1991 (st), Beck et al. 20168 (BG, LPB); prov Nicolas Suarez, nr. Porvenir, 10 Jan 1983 (9 fl), Ferndndez C. et al. 8152 (G, NY); Cobija, Jan 1912 (9 fl-fr), Ule 9311 (F, G, MG).
This is a very variable species in the leaf apex, the
leaf shape, and the indumentum of the lamina, the
stipules, and the inflorescences. Arachnoid indumen-
tum may be sparse or very dense. In Ecuador and
Colombia, the lower leaf surface is often white by dense arachnoid indumentum. Moreover, the stipules and spathes are usually green to yellowish or whitish
because of the cover by dense arachnoid indumentum, and other types of indumentum may be denser and
82 FLORA NEOTROPICA
more conspicuous as well. In Colombia, the main veins of the lamina beneath and the petiole are often densely hirsute to subvillous. In the northern part of the distribution range, as in Meta and Caqueta (Co- lombia) and San Martin (Peru), the incisions of the lamina are <5/10 down to the petiole and the number of lateral veins in the free part of the midsegment is often down to only 6 pairs. Moreover, the lamina can be very large (to 95 cm long). These specimens are quite distinct from the more typical type, normally with the characteristic broadly spathulate free part of the midsegment. The type with dense indumentum and shallowly incised laminas tends to be ecologically different as well, as it does not occur in dense stands in secondary growth, but rather is dispersed, often in margins of tree fall gaps. In areas (mainly in or close to the foot hills of the Andes) where the species oc- curs as treelets in small gaps or in the periphery of larger tree fall gaps, it is rare in man-made secondary growth, whereas elsewhere (in the Amazon basin) it is common. Therefore, the species may comprise two subspecies. The leafy twigs and petioles are glabrous in the two collections from Guainia (Colombia). The species shows clear morphological similarities to Cecropia obtusa. Prance et al. 15375, from Amazo- nian Brazil, shows features transitional to C. obtusa. Subjuvenile leaves of C. kavanayensis are quite sim- ilar to the relatively deeply incised type of leaf of C. ficifolia.
Vernacular names and use. Colombia: cetico blanco, yarumo de raton (Amazonas); jaibena, kYraY- kai iikai (Witoto, Amazonas); udagomo (Mui.). Ven- ezuela: toc'kori, yagrumo, yagrumo blanco (Ama- zonas). Ecuador: anduchi dundo, arduchina durdu, dundu (Quichua, Napo); huarumo (Napo). Peru: tsake, suu (Amazonas); setico blanco (Loreto); sutiik (Jivaro, Loreto). Brazil: imbauba branca (Amazonas; Tukuna Indians use leaf petioles to make flutes for puberty rites).
Cecropia diguensis Cuatrecasas forma albicans Cuatre- casas, Revista Acad. Colomb. Ci. Exact. 9(36/37): 333. 1956. Type. Colombia. Valle: Rfo San Juan, be- low Queremal, 1350-1400 m, 19 Mar 1947 (d), Cuatrecasas 23884 (holotype: US; isotypes: F, US, VALLE).
Cecropia monostachya C. C. Berg, Nord. J. Bot. 1: 485. 1981; Berg & Franco Rosselli, Fl. Ecuador 48: 38, t. 6. 1993. Type. Ecuador. Pichincha: Old rd. Quito- Santo Domingo de los Colorados, km 45, ca. 1600 m, 10 Sep 1977 (Y), Berg et al. 416 (holotype: U; isotypes: AAU, NY, QCA).
Tree, to 15 m tall. Leafy twigs 1.5-3 cm thick, reddish to purple or greenish, glabrous and then often with a bluish (waxy) layer, or occasionally with ? dense arachnoid indumentum; intemodes completely or largely filled with brown pith. Lamina coriaceous, ca. (20 X 20 cm to) 35 X 35 cm to 70 X 70 cm, the segments 7-8, the free parts the upper segments ob- ovate to elliptic, the incisions down to 5/10-8/10, in the upper part, less deeply in the lower part; apices acute, acuminate to rounded; upper surface smooth, subglabrous or puberulous with sparse white hairs on the main veins and "umbilicus," occasionally with rather dense arachnoid indumentum; lower surface glabrous or sparsely puberulous to hirtellous to sub- hirsute on the veins, with arachnoid indumentum in the areoles, also on the smaller veins, sometimes ex- tending to the main veins; lateral veins in the free part of the midsegment 12-18 pairs, submarginally loop- connected, sometimes branched; petiole (10-)20-50 cm long, glabrous or sometimes sparsely hirtellous to (sub)hirsute toward the apex and the base or with arachnoid indumentum; trichilia absent or separate and poorly developed or fused and well-developed, the brown indumentum intermixed with very short white or sometimes long hairs; stipules ca. 15-30 cm long, pinkish to red or greenish, glabrous or some- times (partly) sparsely subvillous and with brown pluricellular hairs or also arachnoid indumentum out- side, glabrous inside. Staminate inflorescences soli- tary or in pairs, erect; peduncle 5-7.5 cm long, almost glabrous with very few long and appressed hairs; spathe 9-19 cm long, pale brown to reddish or green- ish, (sub)glabrous (or subvillous) outside, glabrous (or subvillous) inside; spikes 3-6, 6.5-15 X 0.5-1.5 cm, with stipes 0.4-0.5 cm long and glabrous; rachis hairy. Staminate flowers with pedicels to 1 mm long; perianth tubular, 2-2.4 mm long, with short straight hairs or sometimes also with arachnoid indumentum below the apex, the apex slightly convex, glabrous or sparsely muriculate; filaments flat; anthers 0.6-0.8 mm long, appendiculate, detached at anthesis (?). Pis- tillate inflorescences solitary or in pairs, erect; pedun- cle 3.5-13 cm long, glabrous or sometimes with arachnoid indumentum; spathe 13-26 cm long, the color and indumentum as in the staminate inflores- cence; spikes 1 or 2, 8-15 X ca. 1 cm, to 25 X ca. 2.5 cm in fruit, subsessile; rachis hairy. Pistillateflow-
SYSTEMATIC TREATMENT 83
FIG. 21. Cecropia gabrielis. 1. Leafy twig (Berg et al. 416). 2. Base of petiole with trichilium (Webster et al. 27296). 3. Staminate inflorescence with spathe and at anthesis (Berg 1291). 4. Pistillate inflorescence with at anthesis. 5. Apices of pistillate flowers (Berg et al. 416). 6. Staminate flower. 7. Stamen and filament after detachment of anther (Franco et al. 5563). 8. Pistillate flower. 9. Fruit with persistent style (Franco et al. 4469). (By T. Schipper and Hendrieke Berg. From Nord. J. Bot. 1: 486. 1981, modified.)
84 FLORA NEOTROPICA
ers: perianth ca. 3-4(-6) mm long, with arachnoid hairs below the apex outside, sometimes also in the style channel inside, the apex slightly convex to plane, muriculate or smooth; aperture slit-shaped; style long, S-shaped to straight, hairy; stigma penicillate. Fruit ovoid, ca. 3 mm long, finely tuberculate to smooth.
Distribution (Fig. 18.6). Western Colombia (An- tioquia to Nariiio) and northwestern Ecuador, in mon- tane forest, at 1200-2000 m.
Representative specimens examined. COLOMBIA. ANTIOQUIA: Mun. Cocorna, Vrda. Viao, 1950 m, 26 Mar 1994 (Y fl-fr), Franco et al. 4589 (BG, HUA), (d), Franco et al. 4590 (BG, HUA); nr. Yarumal, 1900 m, 27 Mar 1994 (6), Franco et al. 4594 (BG, HUA). NARINO: Mun. Ri- caurte, rd. to Reserva Natural La Planada, 1800 m, 8 Oct 1993 (6), Franco et al. 4466 (BG, COL), (Y fl-fr), Franco et al. 4469 (BG, COL), (6), Franco et al. 4471 (BG, COL), 22 Feb 1995 (Y fl-fr), Franco et al. 5542 (BG, COL), (6), Franco et al. 5543 (BG, COL, HUA, LP, MO, US); Mun. Barbacoas, Cgto. Altaquer, Vrda. El Barro, 1325 m, 6 Dec 1993 (st), Franco et al. 4959 (COL), 10 Dec 1993 (6), Franco et al. 5107 (COL), 11 Dec 1993 (9 fl), Franco et al. 5142 (COL); Reserva Natural La Planada, above Chu- cunes, 1700 m, 3 Jan 1989 (9 fl-fr), Gentry et al. 64399 (PSO); Reserva Natural La Planada, 1800 m, 14 Jan 1989 (9 fr), Restrepo 440 (MO, PSO); Ricaurte, 1300 m, 12 Apr 1941 (9 fl-fr), Sneidern A. 563 (A, COL, K, LL, MICH, NY, S). VALLE: Mun. Dagua, rd. to Queremal, ca. 76 km from Buenaventura, 1300 m, 17 Mar 1994 (9 fl-fr), Franco et al.
4556 (BG, TULV); Rio Bitaco, 1 km E of Bitaco, ca. 1500 m, 16 Nov 1963 (9 fl-fr), Hutchinson et al. 3039 (COL, F).
ECUADOR. CARCHI: Nr. Maldonado, ca. 1500 m, 5 Sep 1981 (9 fl-fr), Balslev 1984 (QCA); Rio Blanco, above Chical, 1300-1500 m, 25 Sep 1979 (6), Gentry et al. 26527 (MO). PICHINCHA: Rd. Empalme-Chiriboga, ca. 1350 m, 21 Feb 1981 (6), Berg 1291 (AAU, QCA, U, US), 9 Feb 1994 (9 fl-fr), Berg 1687 (AAU, BG, COL, QCNE); Cant6n Quito, Chirobiga, Reserva Forestal La Favorita, 1600-1800 m, 8 Feb 1990 (st), Ceron et al. 8593 (MO); Maquipucuna, 5 km E of Nanegal, 1550 m, 9 May 1990 (6), Gentry et al. 69901 (QCNE); Nanegal, Reserva Ecol6gica Maquipucuna, 1250 m, 9 Jan 1992 (9 fl-fr), Quelal 65 (BG, NY, QCNE); rd. Quito-Manabf, Sep 1874 (st), Sodiro s.n. (Q); Nanegal, Sep 1902 (st), Sodiro s.n. (Q); between Hda. El Carmen (Maquipucuna) & Marianitas, Rio Umacacha, 1200-1500 m, 2 Sep 1989 (9 fl-fr), Webster et al. 27296 (BG, MO, QCA); Reserva Maquipucuna, trail Rio Umacacha-Hda. El Carmen, 4.5 km SE of Nanegal, ca. 1250 m, 16 Sep 1989 (d), Webster et al. 27635 (BG, QCA), 17 Sep 1989 (9 fl- fr), Webster et al. 27689 (BG, QCA).
In Colombia, this species has sometimes two spikes in the pistillate inflorescence. All states of de- velopment of the trichilia, from absent to well- developed, can be found, even on the same plant. Tri- chilia appear to be consistently present in the northern
part of the species range, whereas very often com- pletely absent in the southern part of its range. The leafy twigs and the upper surface of the lamina are occasionally covered with white arachnoid indumen- tum (Webster et al. 27635). This species is very closely related to C. telealba, and probably (more dis- tantly) also related to C. bullata. It can be distin- guished from the former by usual absence of arach- noid indumentum on the upper surface of the lamina and from the latter by the absence of villous indu- mentum on the leafy twigs and petioles. The trees are not inhabited by ants.
Tree, to 12 m tall, often with a short trunk. Leafy twigs 3-5 cm thick, dark green to dark purple; his- pidulous to hispid to setose with irritating bristles. Lamina (sub)coriaceous, ca. (55 X 55 cm to) 70 X 70 cm to 11O X 110 cm, the segments 14-19, lan- ceolate, the incisions down to the petiole; apices acu- minate; margin in dry material ? revolute; upper sur- face scabridulous (to scabrous or to smooth), very sparsely hispidulous, the "umbilicus" (sub)setose; lower surface with arachnoid indumentum in the ar- eoles and on the smaller veins, extending to the pri- mary and lateral veins, these also with brown pluri- cellular hairs; lateral veins ca. 35-50 pairs, submarginally loop-connected, unbranched; petiole ca. 50-100 cm long, with ? dense arachnoid indu- mentum and setose, often only adaxially at the base and in the uppermost part; trichilia fused, the (pale) brown indumentum intermixed with short brown (uni- cellular) hairs or sometimes also sparsely setose; stip- ules 18-30 cm long, dark red-brown or dark purple- red, sparsely to rather densely setose with appressed or patent irritating bristles outside, glabrous inside. Staminate inflorescences in pairs, erect; peduncle 4- 12.5 cm long, hispidulous or partly (short-)setose; spathe 10-15 cm long, dark red, sparsely setose with appressed to patent bristles outside, glabrous inside; spikes ca. 15-35, 4-13 X ca. 0.3 cm, (sub)sessile; rachis hairy. Staminate flowers: perianth tubular, ca. 1 mm long, with sparse short arachnoid indumentum below the apex, the apex plane, muriculate to hispi- dulous or smooth; filaments flat; anthers ca. 0.7-1 mm long, appendiculate, detached at anthesis (?). Pistil- late inflorescences in pairs, erect; peduncle (3-)6-13 cm long, sparsely to rather densely hispidulous to pu-
SYSTEMATIC TREATMENT 85
.~~~~~~~~~~~~~~~ ~ ~~~~~~~~~~~~~A
floer 11 FriIFac ta.49)
/ I nT
FIG. 22. Cecropia garciae. 1. Lamina, reduced. 2. Apex of lamina and venation (Jaramillo et al. 13740). 3. Stipules and young leaf (Franco et al. 4535). 4. Twig with opened prostoma and base of petiole with trichilium (Tipaz et al. 1258). 5. Pair of staminate inflorescences with spathes and base of petiole with trichilium. 6. Staminate inflorescence at anthesis (Franco et-al. 5176). 7. Staminate flower and stamen (Franco et al. 4535). 8. Pair of pistillate inflorescences with spathes and base of petiole with trichilium (Franco et al. 51 77). 9. Pistillate inflorescence at anthesis (Tipaz et al. 1258). 10. Pistillate flower. 11. Fruit (Franco et al. 4693).
86 FLORA NEOTROPICA
berulous and/or partly (short-)setose, often also with sparse to dense arachnoid indumentum; spathe 8-12 cm long, the color and indumentum as in the stami- nate inflorescence; spikes 5-21, 5-18 X 0.3-0.5 cm, to 21 X ca. 1.5 cm in fruit, (sub)sessile; rachis (sub)glabrous. Pistillate flowers: perianth ca. 1.5-2.5 mm long, with arachnoid indumentum below the apex, also below the style channel inside, the apex convex, muriculate, the aperture slightly slit-shaped; style short; stigma comose. Fruit ellipsoid to obovoid or to ovoid, 1.2-1.7 mm long, tuberculate.
Distribution (Fig. 8.7). From western Ecuador to eastern Panama, in forest and secondary growth, at elevations to 1050(-1800) m.
COLOMBIA. ANTIOQUIA: Mun. Frontino, Parque Na- cional Natural Las Orquideas, 20 Jun 1982 (6), Bernal et al. 359 (COL); Mun. Frontino, Cgto. Nutibara, rd. Nutibara- La Blanquita, Murri region, 4 Mar 1995 (6), Franco et al. 5559 (BG, COL, HUA). CAUCA: Mun. GuapI, Parque Na- cional Natural de Gorgona, 1 May 1986 (6), Lozano C. et al. 5082 (COL); Mun. Guapi, Parque Nacional Natural de Gorgona, rd. to Playa Blanca, 5 Sep 1987 (9 fl-fr), Lozano C. et al. 5572 (COL, MO). NARINO: Mun. Barbacoas, nr. El Diviso, 7 Oct 1993 (st), Franco et al. 4464 (BG, COL, HUA, LP, MO, NY, P), 21 Feb 1995 (9 fl-fr), Franco et al. 4693 (COL, HUA, LP, MO, NY, P); Mun. Barbacoas, rd. Tumaco-Junin, Cgto. Junin, 12 Dec 1993 (6), Franco et al. 5176 (BG), (9 fl), Franco et al. 5177 (BG); rd. Tumaco- Tuquerres, W of Junin, 25 Nov 1981 (d), Gentry et al. 34941 (BG, MO); 2-3 km S of Barbacoas, on rd. to Junfn, 7 Jan 1989 ( 9 fl), Gentry et al. 64550 (PSO); rd. Barbacoas- Junin, 1050 m, 7 Aug 1962 (6), Mora 0. 2311 (COL, US); RR Tumaco-El Divisio, km 86, 27 Jul 1952 (9 fl-fr), Romero-Castanieda 3324 (COL). RISARALDA: Mun. Mistrat6, Cgto. Puerto de Oro, Chirrincha, 28 Sep 1991 (9 fl-fr), Franco et al. 3607 (BG, COL). VALLE: Rio Calima, La Trojita, 19 Feb 1944 (9 fl-fr), Cuatrecasas 16467 (COL, F, VALLE); Rio Cajambre, Quebrada de Ord6tiez, 11 May 1944 (9 fl-fr), Cuatrecasas 17272 (F, US, VALLE); Rio Anchicaya, Sabaletas, 13 Nov 1945 (6), Cuatrecasas 19804 (F, US); nr. Buenaventura, 15 Mar 1994 (d), Franco et al. 4535 (BG, TULV); Mun. Buenaventura, rd. to Sabaletas, km 12, 17 Mar 1994 (9 fl-fr), Franco et al. 4551 (BG); Color- ada, Buenaventura Bay, 3 Jun 1944 (d), Killip et al. 38713 (COL, F, US, VALLE).
ECUADOR. CARCHI: Reserva Indigena Awa, Gualpi Alto, 1800 m, 15-28 Jun 1991 (9 fr), Rubio et al. 1535 (BG, QCNE); Tobar Donoso, 19-28 Jun 1992 (d fl-fr), Ti- paz et al. 1258 (MO, QCNE). ESMERALDAS: Between Ma- taje & Molina, 5 Sep 1991 (9 fr), Jaramillo et al. 13740 (GB, NY, QCA); nr. Tobar Donoso, jct. of Rfo Juan & Rio Camumbi, 28 Jul 1966 (9 fr), Jdtiva et al. 1151 (MO, NY, S, US); nr. Borb6n, 2 Aug 1967 (6 + 9 fl-fr), Jdtiva 2105 (MO, NY, S, U); Borb6n, 3 km S of Rio Cayapas, 19 Sep
1965 (9 fl-fr), Little et al. 21097 (MO, NY, Q, QAME, QCNE, US); Cant6n San Lorenzo, Reserva Indfgena Awi, Rfo Mira, 10 km W of Alto Tambo, 16-26 Mar 1991 (d), Rubio et al. 1263 (MO, QCNE); Reserva Ecol6gica Cota- cachi Cayapas, Rio Santiago, Angostura, 17-26 Jul 1994 (st), Tirado et al. 1104 (QCNE).
This species is in most characters more or less similar to Cecropia hispidissima, but they are clearly distinct in the trichilia. In C. garciae, the trichilia are well-defined, only sometimes with sparse irritating bristles, and the Miullerian bodies are small and whit- ish, whereas in C. hispidissima the Muillerian bodies are large and pinkish being produced in ? loosely defined trichilia with numerous irritating bristles. In C. garciae, the number of segments of the lamina is usually 15-19, whereas it is usually 12-15 in C. his- pidissima. Moreover, the lamina is usually scabridu- lous and plane in C. garciae, but smooth and often ? plicate in C. hispidissima. Cecropia garciae is nor- mally found at elevations below 1000 m. The single collection from a much higher elevation in Carchi (Ecuador) was made in an area where other lowland species (of moraceous genera) have also been found at elevations above the normal upper limit of lowland species. The trees of this species are normally inhab- ited by ants of the genus Azteca. The identity of Bris- tan 121, the only collection from Panama, is not quite certain.
15. Cecropia glaziovii Snethlage, Notizbl. Bot. Gart. Berlin-Dahlem 8: 358. 1923; Berg & Carauta, Al- bertoa 1(1): 9. 1986. Syntypes. Brazil. Rio de Ja- neiro: Between Petropolis & Sao Antonio, 1879 (6), Glaziou 11559 (B, destroyed; isotypes: C, GH, LE, P); and Mun. Rio de Janeiro, Floresta da Tijuca, nr. Pies, 1891 (9), Glaziou 18499 (B, de- stroyed; isotypes: A, C, G, H, K, LE, P); Glaziou 18499 (9) at P here designated here as the lecto- type).
Cecropia macranthera Warburg ex Snethlage, Notizbl. Bot. Gart. Berlin-Dahlem 8: 357. 1923; Warburg in Glaziou, Bull. Soc. Bot. France 59(Mem. 3): 645. 1913, nomen. Type. Brazil. Rio de Janeiro, Serra dos Orgaos, 1879 (6), Glaziou 12173 (holotype: B, de- stroyed; isotype: P, comprising vegetative parts of Pourouma guianensis Aublet subsp. guianensis and a staminate inflorescence of C. glaziovii).
Tree, to 20 m tall. Leafy twigs 2-5 cm thick, green, grayish green, or sometimes partly bluish, hispidulous to hirtellous, with curved to uncinate hairs. Lamina
SYSTEMATIC TREATMENT 87
subcoriaceous to chartaceous, ca. 25 X 25 cm to 70 X 70 cm, the segments 8-12, the free parts of the upper segments subobovate (to subpandurate), the in- cisions down to 5/10-8/10(-9/10); apices rounded to acuminate; upper surface scabrous, hispidulous to strigose; lower surface minutely puberulous (to sub- tomentellous) on the main veins, subtomentellous (to subtomentose) on the smaller veins, with arachnoid indumentum in the areoles or also on the smaller veins and concentrated along the margin; lateral veins in the free part of the midsegment 12-16(-23) pairs, sub- marginally loop-connected, often branched; petiole 20-55 cm long, (minutely) puberulous (mainly with curved to uncinate hairs) to subhispidulous or subgl- abrous; trichilia fused, the brown indumentum inter- mixed with ? dense rather short white to brownish (unicellular) hairs; stipules (7-)15-27 cm long, bright to dark red to purplish, sometimes greenish, sparsely puberulous, sparsely to rather densely hirtellous or sometimes ? densely pale yellow- to white-hirsute, or also with sparse arachnoid indumentum outside, glabrous or sparsely to densely hairy inside; terminal buds slightly inflated. Staminate infiorescences in pairs, the peduncle erect to pendulous, the spikes + spreading to pendulous; peduncle 2-13 cm long, his- pidulous to subglabrous; spathe 9-22 cm long, dark to pale red or greenish, hirtellous and with sparse arachnoid indumentum outside, glabrous inside; spikes (4-)8-12(-25), (5-)8-22 X (0.3-)0.5-l.6(-2) cm, yellow, sometimes pink, with stipes 0.5-2 cm long, glabrous and often with a bluish waxy layer or hispidulous, rarely sessile; rachis hairy or glabrous. Staminate flowers: perianth tubular, (1.5-)2-2.5 mm
long, with dense arachnoid indumentum below the apex, the apex ? convex to almost plane, muriculate or smooth; anthers 0.8-2 mm long, at anthesis de- tached, remaining attached to the filament by stretched spiral thickenings (or not detached, or de- tached and reattached to the margins of the aperture by the appendages?). Pistillate inflorescences in pairs, erect, becoming pendulous in fruit; peduncle 1.5- 20(-28) cm long, hispidulous to puberulous to sub- glabrous; spathe ca. 10-15 cm long, the color and indumentum as in the staminate inflorescence; spikes 4-9, 7-12 X 0.4-0.5 cm, to 30 x 1.2 cm in fruit, sessile or with stipes to 0.5(-1.5) cm long and sparsely puberulous to subglabrous; rachis hairy (or subglabrous). Pistillate flowers: perianth ca. 1.5 mm long, with arachnoid indumentum below the apex, also in the lower part of the style channel inside, the apex plane to slightly convex, muriculate; style rather short; stigma comose. Fruit ellipsoid to oblongoid, ca. 2-2.5 mm long, smooth, pale or dark brown.
Distribution (Fig. 11.1). Eastern Brazil, from Ba- hia to Rio Grande do Sul, in moist forest, common in secondary growth, at elevations to 1300 m.
Representative specimens examined. BRAZIL. BA- HIA: Rd. Mi9anga-Ciu, 15 Feb 1980 (? fl-fr), Arauijo 224 (RB); between rd. Sao Jose-Buerarema & Una, km 29, 1 Apr 1980 ( fr), Berg et al. 1146 (BG, K, NY, U); nr. Bahia, 1839 (d), Blanchet s.n. (BM, F, G); Ilheus, 12 May 1981 (Y), Hage et al. 662 (CEPEC, GUA); Mun. Marau, rd. BR.030, between Ubaitaba & Marad, km 33, 5 Feb 1979 (Y fl), Mori et al. 11357 (K, NY, U); Mun. Jussari, Serra do Teimoso, 7.5 km N of Jussari, 9 Feb 1998 (Y fl-fr), Thomas et al. 11774 (NY). ESPiRITO SANTO: Vila Velha, 30-31 Aug 1980 (Y), Andrade 118 (GUA); Santa Teresa, 16 Nov 1988 (d), Boudet Fernandes et al. 2623 (GUA), (Y fl-fr), Boudet Fernandes et al. 2624 (GUA); Mun. Conceicao do Castelo, rd. BR.262, 5 Dec 1984 (Y fl-fr), Hatschbach et al. 48643 (BG, US); between Vila Verde & Domingo Martins, 29 May 1993 (d), Simoes et al. 73 (GUA); Reserva Florestal Lin- hares, 21 May 1993 (Y fl-fr), Sposito 82 (CRVD). MINAS
GERAIS: Mun. Vicosa, between the centre of Vicosa & the University, 10 Sep 1967 (d), Carauta 414A (GUA, RB, U, US), (Y fl-fr), Carauta 414B (GUA, RB, U, US); Mun. Ar- axa, between Araxd & Barreira, 23 Mar 2001 (5), Ernani Diaz 446 (GUA); Juiz de Fora, 3 Sep 1970 (Y fr), Krieger 9054 (GUA); Vi,osa, Agricultural College, 3 Dec 1958 (st), Irwin 2199 (MICH, NY, US); 15 Feb 1939 (Y fl-fr), Mexia 4349 (A, BM, F, GH, K, MICH, MO, NY, S, U, US); Car- angola, Oct 1991 (d), Leoni 1699 (GUA). PARANA: Volta Grande, 1 Feb 1904 (d), Dusen 3542 (GH, S); Therezina, 26 Jan 1911 (st), Dusen 11217 (GH, S); Volta Grande, 31 Mar 1912 (d), Duse'n 14043 (GH, K, MICH, NY, P, S); Mun. Morretes, Est. Marumbi, 8 Feb 1985 (9 fl-fr), Hatsch- bach 48892 (BG, US); Mun. Morretes, Passa Sete, 5 Jul 1988 (9 fr), Hatschbach et al. 52158 (BG); Mun. Paran- angud, PR.53, 4 km N of Rio Guaraguacu, 5 Jul 1988 (d), Hatschbach et al. 52159 (BG); Paranagud, Ilha das Cabras, 14 Jun 1986 (9 fl-fr), W S. Souza et al. s.n. (UEC). RiO DE
JANEIRO: Nr. Cabo Frio, 16 Mar 1980 (d), Berg etal. 1134 (BG, K, RB, U), (9 fl-fr), Berg et al. 1136 (U); Mun. Rio de Janeiro, Serra Carioca, Estrada da Vista Chinesa, 19 Dec 1963 (9 fl-fr), Carauta 208 (GUA, K, NY, US); Mun. Ter- es6polis, Granja Comari, 10 Feb 1964 (9 fr), Carauta 211B (GUA, U, US); Mun. Resende, Vila de Itatiaia, 1 Jan 1968 (d), Carauta 553 (GUA); Mun. Rio de Janeiro, Estrada da Vista Chinesa, km 2, 21 Oct 1968 (9 fl-fr), Carauta 645 (E, F, GUA, MICH, SP, U); Mun. Petr6polis, Represa do Rio Caxambu Pequeno, 19 Jul 1978 (), Carauta 2976 (F, GUA, RB); rd. Barra do Pirai-Valen,a, 4 Mar 1980 (d), Carauta et al. 3425 (GUA); Mun. Rio de Janeiro, Gruta de Imprensa, Sao Gabriel, 3 Nov 1962 (9 fl-fr), Cuatrecasas et al. 26626 (US), (d), Cuatrecasas et al. 26627 (P, US); nr. Rio de Ja- neiro, GCvea, 19 Aug 1890 (5), Glaziou 18500 (C, G, K, P, US), (5), Glaziou 18501 (BG, C, G, K); Mun. Mage, Par- aiso, 9 Nov 1985 (d), Gomes 4 (RB); Angra dos Reis, 22 Mar 1951 (d), M. Kuhlmann 2673 (SP), (9 fl), M. Kuhl- mann 2674 (SP). Rio GRANDE DO SUL: Picada Verao, 20 Jan 1991 (9 fl-fr), Quadros etal. 1679 (GUA), (5), Quadros
88 FLORA NEOTROPICA
et al. 1680 (GUA); Os6rio, Maquine, 22 Dec 1979 (d), Wae- chter et al. 1516 (GUA). SANTA CATARINA: Mun. Brusque, Mata do Hoffmann, 8 May 1951 (9 fr), Klein 113 (US); Mun. Itapoa, Reserva Volta Velha, 17 Feb 1993 (9 fr), Ne- grelle et al. A. 743 (GUA); Mun. Palhoqa, Morro do Cavalo, 11 Mar 1953 (9 fl-fr), Reitz et al. 319 (GUA, US); Mun. Itajaf, Morro da Fazenda, 28 Apr 1954 (9 fl-fr), Reitz et al. 1797 (NY, US); Itajaf, Morro da Ressacada, 29 Mar 1956 (6), Reitz et al. 2902 (NY, US); Mun. Sao Francisco do Sul, Garuva, Tres Barras, 19 Dec 1957 (6), Reitz et al. 5736 (GUA); nr. Tubarao, May 1889 (9 fl-fr), Ule 1196 (P); Rio Novo, Orleans, 14 Jan 1993 (9 fl-fr), Zanette et al. 1438 (GUA). SAO PAULO: Mun. Ubatuba, Rio Escuro, Rio Es- curo, Praia Dura, 18 Dec 1979 (6), Benson 10837 (K, NY, UEC); Campinas, 10 Jul 1968 (6), Carauta et al. 623 (GUA), (9 fl-fr), Carauta et al. 624 (C, GUA, U, US); Mun. Iguap6, Pinheiro, 11 Jun 1982 (9 fl-fr), Hatschbach 44969 (U, US); Sao Paulo, Botanical Garden, 2 Sep 1949 (6), W
Hoehne et al. 3325 (GUA, US, SP, SPF) = M. Kuhlmann 3326 (SP); Jacarei, 30 Sep 1949 (6), W Hoehne et al. (SPF) 12787 (SPF); Mun. Ubatuba, Praia Dura, 26 Mar 1978 (6), Joly 7349 (GUA, RB, UEC); Reserva Florestal das Fontes do Ipiranga, 7 Jun 1988 (9 fl-fr), Romaniuc 760, 15 Jan 1989 (6), Romaniuc 787 (SP); Cubatao, BR.207, km 68, 14 Dec 1990 (d), Romaniuc et al. 1015 (SP); Tapiraf, Serra de Paranapicaba, 10 Feb 1992 (9 fl-fr), Romaniuc et al. 1273 (SP), (d), Romaniuc et at. 1275 (SP); Santos, Mata da La- goa, 2 May 1994 (9 fl-fr), Romaniuc et al. 1396 (SP); Tim- buri, Fazenda Dominiciana, 14 Jun 1995 (9 fl-fr), Tama- shiro et al. 1255 (SP).
Cecropia glaziovii is a rather variable species in
terms of the length of the peduncle and (even on the same tree) the number and diameter of the spikes of the staminate inflorescence. The peduncle of the pis- tillate inflorescence is mostly to ca. 10 cm long, some- times to ca. 20 cm long, or occasionally to 30 cm long in fruit. The outer surface of the stipule is normally rather sparsely puberulous to hirtellous, sometimes densely hirtellous, or occasionally densely hirsute (e.g., Berg et al. 1146 and Tamashiro et al. 1255). The uncommon features tend to occur in the periph- ery of the known distribution range of the species. The leaves on the stem are often large (to ca. 100 X 100 cm) before branching and the lamina is often more deeply incised (down to ca. 9/10, even down to 1.5-3 cm from the petiole); the free part of the mid- segment has ca. 20-23 pairs of lateral veins, and the arachnoid indument is often sparse and (almost) con- fined to the margin beneath. Cecropia glaziovii is very closely related to C. palmata, showing distinct simi- larities in the shape and indumentum of the leaves, in both the pistillate and staminate inflorescences, and in the staminate flowers. The two species appear to be allopatric. More or less clear differences are as follows: (1) The white arachnoid indumentum is
sparse on the spathes in C. glaziovii, but dense on those of C. palmata; (2) the stipules are mostly 15- 25 cm long and caducous in C. glaziovii, but 7-15 cm long and mostly subpersistent in C. palmata; and (3) the staminate inflorescences of C. glaziovii often have >4 spreading to pendulous spikes, but those of C. palmata normally have only 4 pendulous ones. In ma- terial with thick spikes, long flowers, and stamens > 1 mm long, the detached anthers remain attached to the filament by stretched spiral thickenings. However, it is not clear what happens to the smaller anthers (>1 mm long) of short flowers (in more slender spikes); they may remain attached to the (for the greater part ? swollen) filament or become detached and re- attached by the appendages; see under C. palmata. The collections Glaziou 18500-18507 are partly mixed, comprising material of C. glaziovii and C. pachystachya.
Cecropia arbelaezii Cuatrecasas, Revista Acad. Colomb. Ci. Exact. 6(21): 65. 1944; Not. Fl. Colomb. VI. (Trab. Col. Bot. etc. Colomb.) 44. 1944. Type. Co- lombia. Tolima: Curvas de Gualanday, between Iba- gue & Girardot, 22 Jul 1939 (d), Cuatrecasas & Perez Arbeldez 6503 (holotype: COL; isotypes: F, US).
Tree, to 5(-10) m tall. Leafy twigs 1.5-2.5 cm thick, dark green to brownish, hirtellous to subhispi- dulous with curved hairs. Lamina chartaceous, ca. 25 X 25 cm to 35 X 35 cm (to 60 X 60 cm), the segments 6-7, the upper segments elliptic, the incisions in the upper part of the lamina down to 5/10-6/10; apices rounded; upper surface scabridulous (or scabrous), somewhat bullate, densely puberulous to subhispi- dulous; lower surface densely hirtellous to subtomen- tose on the veins, the arachnoid indumentum not in the areoles, sparsely to rather densely covering the hairs on the veins, largely disappearing, remain- ing along the margin; lateral veins in the free part of the midsegment 7-10 pairs, submarginally loop- connected, branched; petiole 15-30(-50) cm long, puberulous to hirtellous; trichilia fused, the brown in- dumentum intermixed with rather long grayish to white hairs; stipules 4-6 cm long, green, densely grayish hirtellous to subtomentose outside, sericeous inside. Staminate inflorescences in pairs or solitary, pendulous; peduncle 6-11 cm long, hirtellous to sub- setulose; spathe 10-16 cm long, green, densely hir- tellous outside, sparsely hairy inside; spikes 4, 10-17
SYSTEMATIC TREATMENT 89
-%N~~~~~~~~~~~~~~~~~~~~~N
3~~~~~~~~~~~~
I mn ~ ~ 1mn
FIG. 23. Cecropia goudotiana. 1. Leafy twig with stipules, lamina, and trichilium. 2. Apex of lamina and venation. 3. Pair of staminate inflorescences with spathes and base of petiole with trichilium. 4. Staminate inflorescences at anthesis and bases of petioles with trichilia. 5. Staminate flower and stamen (Franco et al. 4652). 6. Pair of pistillate inflorescences with spathes and base of petiole with trichilum. 7. Pair of pistillate inflorescences after anthesis and base of petiole with trichilium (Franco et al. 4650). 8. Pistillate flower. 9. Fruit (Franco et al. 4651).
90 FLORA NEOTROPICA
X 0.4-0.5 cm, with stipes 0.3-0.5 cm long and densely puberulous; rachis sparsely hairy. Staminate flowers: perianth tubular, 1-1.5 mm long, glabrous, the apex plane; filaments ? swollen; anthers 0.5-0.6 mm long, short-appendiculate, remaining attached to the filament at anthesis (?). Pistillate inflorescences in pairs or solitary, erect to deflexed, soon pendulous; peduncle 4-12 cm long, hirtellous to subsetulose; spathe 7-11 cm long, the color and indumentum as in the staminate inflorescence; spikes (2-)4(-6), often two by two fused at the base, 7-10 X 0.4-0.5 cm, to 15 X 0.7-0.8 cm in fruit, sessile; rachis (sparsely) hairy, sometimes partly with arachnoid hairs, or (sub)glabrous. Pistillate flowers: perianth 1-1.5 mm long, with arachnoid hairs below the apex, absent in- side, the apex ? convex to plane, glabrous and smooth; style rather long; stigma comose. Fruit ob- longoid, ca. 1.2-1.5 mm long, tuberculate.
Distribution (Fig. 11.3). Colombia, probably con- fined to Tolima, on rock escarpments in areas with dry forest, at low elevations.
Specimens examined. COLOMBIA. TOLIMA: Mun. Piedras, rd. Piedras-Doima, km 5, 7 Feb 1995 (d fl-fr), Franco et al. 4650 (BG, COL, HUA, LP, US); Mun. Vena- dillo, rd. to Merida, 7 Feb 1995 (Y fl-fr), Franco et al. 4651 (BG, COL, HUA, LP, US), (d), Franco et al. 4652 (BG, COL, HUA, LP, US), (Y fl-fr), Franco et al. 4653 (BG, COL, HUA, LP, US); Mun. Venadillo, Vrda. La Sierrita, Finca El Cedro, 12 Aug 1980 ( fl-fr), Idrobo et al. 10988 (COL); Mariquita, 7 Jan 1918 (Y fl), Pennell 3635 (A).
The species is relatively rare in its range of distri- bution, the northern part of the dry upper Magdalena valley; it is confined to rocks and rocky escarpments.
17. Cecropia granvilleana C. C. Berg, Bull. Mus. Hist. Nat. (Paris), ser. 4, sect. B, Adansonia 7: 255, t. 1. 1985; Berg in Gorts-van Rijn, Fl. Guianas, ser. A., fasc. 11: 97, t. 20. 1992. Type. French Gui- ana. Montagne de Trinit6, 18 Jan 1984 (6), Gran- ville et al. 6000 (holotype: P; isotypes: BG, BR, CAY, G, NY, U). Fig. 24
Tree, to 10 m tall, the trunk with prominent stip- ular scars. Leafy twigs 1.5-2 cm thick, green, puber- ulous to hispidulous, partly with uncinate hairs. Lam- ina chartaceous to subcoriaceous, ca. 25 X 25 cm to 40 X 40 cm, the segments 9-10, oblong to lanceolate, the incisions down to ca. 6/10-8/10; apices acute to subacuminate; upper surface scabrous to scabridu- lous, hispidulous; lower surface hirtellous to puber- ulous on the veins, with arachnoid indumentum in the areoles and on the smaller veins; lateral veins
in the free part of the midsegment 9-10 pairs, mar- ginally to submarginally loop-connected, unbranched or branched; petiole ca. 15-35 cm long, minutely pu- berulous; trichilia fused, the brown indumentum in- termixed with 5-9 mm long white hairs; stipules 3-4 cm long, pink to dark red, puberulous to hirtellous and with sparse arachnoid indumentum outside, (sub)glabrous inside. Staminate inflorescences in pairs, erect; peduncle 8-9 cm long, sparsely puberu- lous; spathe 4-6 cm long, pale pink to yellow, sparsely puberulous to hirtellous and with sparse to rather dense arachnoid indumentum outside, glabrous inside; spikes 6-9, 2-4.5 X 0.3-0.4 cm, with stipes 0.3-0.4 cm long and sparsely hairy; rachis hairy. Sta- minateflowers: perianth tubular, 1-1.5 mm long, with arachnoid indumentum below the apex, the apex slightly convex to plane, glabrous; filaments flat; an- thers ca. 0.5-0.8 mm long, detached at anthesis, reat- tached to the margins of the aperture by the append- ages. Pistillate inflorescences in pairs, erect; peduncle 6-10 cm long, hispidulous; spathe ca. 5-8 cm long, the color and indumentum as in the staminate inflo- rescence; spikes 4(-6), (at anthesis) 4-7 X 0.5-0.8 cm, sessile; rachis glabrous. Pistillate flowers: peri- anth ca. 1.5 mm long, with arachnoid indumentum below the apex, the apex muriculate to sparsely min- utely puberulous; stigma subpeltate. Fruit unknown.
Distribution (Fig. 8.6). French Guiana, on steep, wet slopes of granite rocks (inselbergs).
Specimens examined. FRENCH GUIANA: Montagne de Trinite, 12 Feb 1984 (9 fl-fr), Granville et al. 6611 (BG, CAY, G, P, U); Haute Marouini, Rocher Koutou, 16 Aug 1987 (st), Granville et al. 9319 (US), 17 Aug 1987 (6), Granville et al. 9392 (BG, INPA, P, U, US).
This species is known from only two localities and is found in small patches at bases of inselbergs. It shows affinities to Cecropia peltata.
18. Cecropia herthae Diels, Notizbl. Bot. Gart. Berlin-Dahlem 15: 367. 1941. Type. Ecuador. Pas- taza: Mera, ca. 1100 m, 3 Dec 1938 (9) Schultze- Rhonhof 3072 (holotype: B, destroyed, duplicates not traced); here replaced by: Ecuador. Napo: Re- serva Biologica Jatun Sacha, 8 km E of Mishualli, 25 Dec 1990 (9), Neill & Berg 9743 (neotype: QCNE; isoneotypes: BG, GB, MO, NY). Fig. 25
Cecropia congesta Cuatrecasas, Revista Acad. Colomb. Ci. Exact. 6(22/23): 299. 1945. Type. Colombia. Pu- tumayo: Rio San Miguel, between Rio Conejo & Rio Hormiga, 15 Dec 1940 (D fr), Cuatrecasas 11069 (holotype: COL; isotype: F).
SYSTEMATIC TREATMENT 91
0.5mm I 9 3 f GS0mm
FIG. 24. Cecropia granvilleana. 1. Leafy twig with stipules, lamina, trichilia, and staminate inflorescences. 2. Sta- minate flower and stamen (Granville et al. 6000). 3. Pistillate flower and stigma. 4. Fruit with persistent style (Granville et al. 6611). (By H. Rypkema. From Bull. Mus. Natl. Hist. Nat., Paris, s6r. 4, 7[Sect. B, Adansonia 3]: 256. 1985.)
92 FLORA NEOTROPICA
I~~~~~~~4
SS; X
2;!~~~~~~~~~~I
FIG. 25. Cecropia herthae. 1. Lamina, reduced. 2. Apex of lamina and venation. 3. Stipules and bases of petioles with trichilia (Neill et al. 9743). 4. Staminate inflorescence with spathe. 5. Staminate inflorescence at anthesis (Zak et al. 4404). 6. Staminate flower and stamen (Neill et al. 9743A). 7. Pistillate inflorescence with spathe (Zak et al. 4471). 8. Pistillate inflorescence at anthesis and bases of petioles with trichilia. 9. Pistillate flower (Neill et al. 9743).
SYSTEMATIC TREATMENT 93
Tree, to 30 m tall. Leafy twigs 2-8 cm thick, ini- tially dark green with large pinkish lenticels, the older parts often blackish, hispidulous. Lamina coriaceous to subcoriaceous, ca. 60 X 60 cm to 80 X 80 cm (to 100 X 100 cm), the segments (10-)14-16, oblance- olate, the incisions down to 1-2 cm from the petiole or down to the petiole (and subpetiolulate); apices (sub)acuminate to acute; upper surface scabrous to scabridulous and hispidulous or smooth and subgla- brous to sparsely puberulous (to hirtellous); lower surface puberulous (with uncinate and straight hairs) or partly subhirtellous, with arachnoid indumentum confined to the areoles; lateral veins in the free part of the midsegment ca. 30-40 pairs, marginally loop- connected, unbranched (or faintly branched); petiole ca. 50-75(-l 10) cm long, puberulous (or adaxially partly hirtellous) or also with sparse to rather dense arachnoid indumentum; trichilia fused (slightly sunken into the base of the petiole), only with brown pluricellular hairs; stipules 20-30 cm long, pink to pale dull red, caducous or subpersistent, puberulous and with sparse short arachnoid indumentum outside, glabrous inside. Staminate inflorescences in pairs, the peduncle erect, the spikes erect to ? deflexed; pe- duncle 3.5-9 cm long, hispidulous and with sparse arachnoid indumentum; spathe (not mature) 6-7 cm long, sparsely minutely puberulous outside, glabrous inside; spikes ca. 50-100, (4-)8-14 X 0.2-0.4 cm, with stipes 0.3-1 cm long and glabrous or puberulous; rachis hairy. Staminate flowers: perianth tubular, ca. 1-1.2 mm long, glabrous, the apex slightly convex, sparsely muriculate; filaments flat; anthers ca. 0.4 mm long, appendiculate, detached at anthesis (?). Pistil- late inflorescences in pairs, erect (to ? spreading); peduncle 3-4 cm long, hispidulous to hirtellous and with sparse arachnoid indumentum; spathe ca. 7-8 cm long, the indumentum as in the staminate inflores- cence; spikes (7-)9-12, (in fruit) 5-12 X 1.5-2 cm, sessile or with stipes to 0.5 cm long and puberulous; rachis glabrous. Pistillate flowers: perianth ca. 2 mm long, with arachnoid indumentum below the apex, also below the style channel inside, the apex convex, dome-shaped, smooth or muriculate, the aperture slit- shaped; style short; stigma comose. Fruit oblongoid, ca. 4 mm long, almost smooth, pink.
Distribution (Fig. 10.6). Northern Amazonian Ec- uador and adjacent Colombia (Putumayo) and Peru (Loreto), in forest, at low elevations.
Specimens examined. COLOMBIA. AMAZONAS:
Mun. Leticia, Parque Nacional Amacayacu, Quebrada de Agua Pudre, 15 Nov 1991 (Y fl), Pipoly et al. 16272 (COL). PUTUMAYO: Mun. Orito, Vrda. El Parafso, 17 Feb 1995 (Y fr), Franco et al. 5574 (BG, COL).
ECUADOR. MORONA-SANTIAGO: Pozo petrolero Garza, ca. 35 km NE of Montalvo, 2-12 Jul 1985 (6), Zak et al. 4404 (MO, QCNE), (Y fl), Zak et al. 4471 (BG, MO, QCNE). NAPO: Rio Napo, nr. Santa Rosa, Rio Huambuno, 4 Mar 1990 (st), Berg et al. 1644 (BG, COL, QCA); Parque Nacional Yasuni, Pozo petrolero Daimi 2, 26 May-8 Jun 1988 (Y fl-fr), Ceron et al. 4021 (BG, QCNE); Cant6n Fran- cisco Orellana, Huashito, 3-21 Nov 1989 (Y fr), Gudino 179 (BG, MO, QCNE); Parque Nacional Yasuni, rd. Pom- peya Sur-Iro, km 38.7, 20 Oct 1994 (st), Jaramillo et al. 17110 (QCA); Cascada de San Rafael, 1400 m, 1 Aug 1975 (st), Little et al. 234A (Q); Reserva Biol6gica Jatun Sacha, 8 km E of Misahualli, 26 Dec 1990 (d), Neill et al. 9743A (BG), 8 Aug 1987 (st), Palacios 1774 (QAME, QCNE), 14 Aug 1987 (Y fl-fr), Palacios 1888 (BG, MO, QAME, QCNE), 22 Sep 1989 (st), Palacios 4494 (BG, MO, QAME, QCNE), 21 Oct 1989 (st), Palacios et al. 4684 (BG, QCNE). PASTAZA: Cant6n Pastaza, Rio Lliquino, 1 km N of Pozo Villano 2, 29 Aug 1997 (d), A. Alvarez et al. 2269 (BG). SUCUMBiOS: Shushufindi, 3 Aug 1975 (st), Little et al. 35 (Q, QCNE), 28 Dec 1974 (d), Vickers 92 (F).
The present description of the vegetative parts largely matches the description of this species by Diels (1941) and is in accordance with his comment that Cecropia herthae resembles C. sciadophylla. However, there is a discrepancy between the present description of the pistillate inflorescence and Diels's description of the peduncle as 12-15 cm long, his- pidulous to hirtellous; spikes 5, 10-13 cm long; per- ianth with arachnoid indumentum below the apex; stigma penicillate. At least the difference in the length of the peduncle cannot be easily explained, if one does not assume that the type collection was a mixed collection. This species is, at first sight, very similar to C. sciadophylla, in particular to the type with non- petiolate leaf segments. However, C. herthae is clearly distinct in the presence of trichilia. Both spe- cies are also ecologically similar; they can become tall trees and can be found side by side.
19. C. heterochroma C. C. Berg & P. Franco, Novon 6: 246, t. 11. 1996. Type. Panama. Veraguas: Rd. Escuela Agricola Alto Piedra-Rio Dos Bocas, km 10, 26 Jul 1974 (9), Croat 25880 (holotype: MO; isotype: BG). Fig. 26
Tree, to 8 m tall. Leafy twigs 2-5 cm thick, green or purplish, densely hirtellous with uncinate hairs. Lamina chartaceous, ca. 35 X 35 cm to 75 x 75 cm, green or purplish beneath, the segments 6-8, the in-
94 FLORA NEOTROPICA
/ ~~~~~~~1W
I /0
FIG. 26. Cecropia heterochroma. 1. Lamina, reduced. 2. Apex of lamina and venation (Nevers 4953). 3. Stipules and base of petiole with trichilium (Berg 299). 4. Pair of staminate inflorescence at anthesis (Nevers 3752). 5. Staminate flower. 6. Stamen (Berg et al. 393). 7. Pistillate inflorescence at anthesis and base of petiole with trichilium. 8. Pistillate inflorescence with spathe (Nevers 4953). 9. Pistillate flower (Herrera et al. 1763). 10. Fruit with persistent style (Nee et al. 11004). (From Novon 6: 247. 1996, modified.)
SYSTEMATIC TREATMENT 95
cisions down to 3/10-5/10; apices subacuminate to rounded; upper surface minutely hispidulous, scabri- dulous; lower surface minutely puberulous with curved hairs on the veins, the arachnoid indumentum very sparse, mainly on the main veins and the margin, not in the areoles, soon disappearing; lateral veins in the free part of the midsegment 10-12 pairs, submar- ginally loop-connected, some of them branched; pet- iole 30-55 cm long, green or purplish, puberulous, partly hirtellous to subhispid with uncinate hairs; tri- chilia fused, the brown indumentum intermixed with short white hairs; stipules 5-10 cm long, green or reddish, subhirtellous to subhispid with uncinate hairs outside, glabrous inside. Staminate inflorescences in pairs, erect; peduncle 3-6.5 cm long, reddish or pur- plish, hirtellous to subhirsute to subhispid; spathe 8- 14 cm long, reddish, purplish, or greenish, sparsely hirtellous and often with dense brown pluricellular hairs outside, glabrous inside; spikes 4-5, 3-6.5 X 0.3-0.4 cm, with stipes to 0.5 cm long and sparsely puberulous to glabrous; rachis glabrous. Staminate flowers: perianth tubular, 2-3 mm long, glabrous, the apex 5-angular in circumference, adhering to apices of adjacent perianths, slightly concave to plane, but the margins of the aperture bent upward, glabrous; filaments flat; anthers 0.6-0.8 mm long, oblong to lanceolate in outline, appendiculate or not, remaining attached at anthesis. Pistillate inflorescences solitary, erect; peduncle 8-12 cm, red to purplish, puberulous to hirtellous; spathe ca. 10-15 cm, the color and in- dumentum as in the staminate ones; spikes (l-)2-4, 3-10 X 0.4-0.6 cm, to 15 X ca. 1 cm in fruit, (sub)sessile; rachis glabrous. Pistillate flowers: perianth 1.5-2 mm long, with arachnoid hairs below the apex, also below the style channel inside, the apex slightly convex, glabrous, the aperture circular, sur- rounded with a low rim; style rather long; stigma co- mose. Fruit ellipsoid, ca. 2-3 mm long, smooth, brown.
Distribution (Fig. 8.2). From eastern Panama to Colombia, in forest margins and in secondary growth, at elevations to 1000 m.
Representative specimens examined. PANAMA. Bo- CAS DEL TORO: Isla Bastimentos, 22 Mar 1993 (9 fl-fr), Foster et al. 14728 (SCZ). COLON: Santa Rita Ridge rd., between Transisthmian Hwy. & Agua Clara, 11 Dec 1973 (9 fl-fr), Berg 299 (BG); Santa Rita Ridge, 1 Mar 1971 (9
fl), Croat 13887 (BG, MO); Santa Rita Ridge rd., 4-6 km from Transisthmian Hwy., 13 Apr 1976 (6), Croat 34288 (MO); Santa Rita Ridge rd., 21-26 km from Transisthmian Hwy., 4 Jul 1982 (9 fr), Knapp 5844 (BG, MO). DARIEN:
Parque Nacional Darien, nr. Cruce de Mono, 5 Nov 1989 (juv), Fisher 52 (BG). PANAMA: Cerro Jefe, 30 Aug 1977
(Y fl-fr), Berg et al. 393 (BG), (juv), Berg et al. 393A (BG), (d), Berg et al. 394 (BG); El Llano-Carti rd., km 7, 3 Sep 1977 (d), Berg et al. 401 (BG); Campo Tres, 5 km NE of Altos de Pacora, 9 Mar 1973 (6), Busey 822 (BG, MO); Cerro Jefe, 25 Aug 1972 (Y fr), Correa et al. 1806 (MO, PMA); 3 mi N of Cerro Azul, 26 Jul 1970 (Y fl-fr), Croat 11587 (MO); El Llano-Cartf rd., km 8.7, 31 Oct 1977 (Y fl), Folsom et al. 6161 (BG, MO); Cerro Jefe, 13 Sep 1970 ( fl-fr), Foster et al. 1896 (PMA, US), 1 Jan 1972 (9 fl- fr), Gentry et al. 3447 (MO, NY); Cerro Jefe, 29 Jul 1967 (9 fl-fr + 6), Kirkbride et al. 21 (MO, NY); El Llano-Cartf rd., km 18, 2 Mar 1975 (9 fl-fr), Mori et al. 5122 (BG, MO); El Llano-Carti rd., km 14, 28 Mar 1974 (9 fl), Nee et al. 11004 (MO, NY, PMA). SAN BLAS: Nusagandi, 16 Aug 1989 (9 fl), Fisher 19 & 20 (BG); Rio Play6n, Chico, 27 Aug 1994 (9 fl), H. Herrera et al. 1763 (BG); Nusagandi, NW of Punta Mama, 13 Aug 1984 (9 fl-fr), Nevers et al. 3731 (MO, PMA); El Llano-Carti rd., continental divide, 25 Aug 1984 (6), Nevers 3752 (MO); El Llano-Cartf rd., km 19.1, 4 Mar 1985 (9 fl), Nevers et al. 4953 (MO, PMA); El Llano-Cartf rd., 13 Mar 1986 (6), Nevers et al. 7378 (BG, MO, PMA). VERAGUAS: Rio Dos Bocas, 15.6 km NW of Santa Fe, 31 Aug 1974 (6), Croat 27627 (MO).
COLOMBIA. VALLE: Mun. Buenaventura, Bajo Cal- ima region, rd. Buenaventura-Malaga, km 51.3, 9 Feb 1990 (6), Croat et al. 70406 (BG, MO).
This species has a morph with the lamina purplish underneath and a morph with the lamina pale green underneath; they are found side by side. The species is probably related to Cecropia obtusifolia.
20. Cecropia hispidissima Cuatrecasas, Revista Acad. Colomb. Ci. Exact. 9(36/37): 325. 1956. Type. Colombia. Valle: Rio San Juan, below Quer- emal, 27 Mar 1947 (9), Cuatrecasas 23986 (ho- lotype: US; isotype: F). Fig. 27
Tree, to 10(-25) m tall, often with a short trunk. Leafy twigs 3-9 cm thick, green, setose with irritating hairs, sometimes subsetose to subvillous, or also with arachnoid indumentum. Lamina coriaceous, ca. 40 X
40 cm to 85 X 85 cm (to 1 5 X 1 5 cm), the segments (9-)10-13(-15), lanceolate, the incisions down to 2-5 cm from the petiole or to the petiole and then often almost petiolulate; apices (sub)acuminate to acute; upper surface smooth or nearly so, (sub)glabrous, the "umbilicus" usually (sub)setose; lower surface with brown pluricellular hairs or often also with arachnoid indumentum and sometimes ? sparsely setose on the main veins, with arachnoid indumentum in the areoles and on the smaller veins, often (initially) extended to the main veins; lateral veins in the free part of the midsegment 25-40(-44) pairs, submarginally loop- connected, unbranched; petiole 30-85(-100) cm long, with sparse to dense arachnoid indumentum and fili-
96 FLORA NEOTROPICA
\ 0.I,44.
4.1 C
;EgAt
FIG. 27. Cecropia"hispidissima. 1. Lamina, reduced. 2. Apex of lamina and venation1(Croat 11588). 3. Stipules,
form brown pluricellular hairs, usually also (partly) setose (with irritating hairs) to hirsute, glabrescent; trichilia fused, their margins often not well-defined, sometimes extending to the base of the stipules or almost lacking, the brown indumentum intermixed with irritating bristles, the Muillerian bodies relatively large (ca. 3 mm) and pink; stipules 12-40 cm long, dark red-brown, dark purple-red (to blackish), green, or sometimes whitish, densely subhispid to hirsute to substrigose (to subvillous) and with sparse to dense arachnoid indumentum outside, glabrous inside. Sta- minate inflorescences in pairs, erect; peduncle 8-12 cm long, very densely hispid; spathe 5-14 cm long, yellowish green or reddish brown to dark red to dark purple (to blackish), densely hispid to setose or to hirsute, sometimes subvillous and with dense arach- noid indumentum outside, glabrous inside; spikes ca. 15-65, (3-)7-13 X (0.3-)0.7-1 cm, with stipes 0.3- 0.4 cm long and hispid; rachis hairy. Staminate flow- ers sessile or short-pedicellate; perianth tubular, ca. 2-2.5 mm long, glabrous or with arachnoid hairs be- low the apex, the apex slightly convex to plane; filaments slightly swollen; anthers ca. 0.7-0.8 mm long, appendiculate, detached at anthesis, reattached to the margins of the aperture by the appendages (?). Pistillate inflorescences in pairs, erect; peduncle 6- 18 cm long, densely setose to hirsute (to subvillous) and with dense arachnoid indumentum, glabrescent; spathe ca. 4-13 cm long, the color and indumentum as in the staminate inflorescence; spikes 6-20, 3-12 x ca. 0.4 cm, to 19 X 1-1.5 cm in fruit, (sub)sessile; rachis (sparsely) hairy. Pistillateflowers: perianth, ca. 2.5 mm long, with arachnoid indumentum below the apex outside, absent inside, the apex slightly convex, muriculate or glabrous; style long, straight; stigma penicillate. Fruit ellipsoid to ovoid, ca. 1.2-1.5 mm long, ? tuberculate.
Distribution (Fig. 10.5). From eastern Panama to western Ecuador, in forest or secondary growth, at elevations to 1500 m.
Representative specimens examined. PANAMA. COLON: Rio Escandaloso, nr. Mina Boquer6n Numero 2, 14 Jul 1979 (d), Antonio 1309 (MO); Santa Rita Ridge rd., between Transisthmian Hwy. & Agua Clara, 11 Dec 1973 ( Y fl-fr), Berg 298 (BG); nr. Rio Piedras, rd. to Puerto Bello, 16 Jul 1966 (d), Blum 2530 (MO); Salud, 2 Aug 1971 (Y fr), Lao 181 (MO, PMA); Pipeline rd., 8 km NW of Gamboa, 26 Aug 1973 (D fl-fr), Nee 6643 (AAU, MO, NY, PMA). PANAMA: El Llano-Carti rd., 3 Sep 1977 (Y fl-fr), Berg et al. 402 (BG); Cerro Jefe, 27 Jan 1966 (Y fl), Blum et al. 2099 (MO); Cerro Azul, 3 mi N of Cerro Azul (vill.), 26 Jul 1970 (5), Croat 11588 (MO, NY); Cerro Jefe, ca. 1000 m, 22 Sep 1972 (D fl-fr), Gentry 6149 (GH, MO); El Llano-
Carti rd., km 5, 11 Nov 1973 (Y fl-fr), Nee 7949 (AAU, GH, MO, PMA); above El Valle, rd. to La Mesa, 12 Feb 1974 (st), Tyson 6951 (PMA). SAN BLAS: Rd. Nusagandi-Cartf, 17 Jul 1986 (5), McDonagh et al. 79 (BM); El Llano-Carti rd., continental divide, 25 Aug 1984 (Y fl), Nevers 3754 (MO).
COLOMBIA. ANTIOQUIA: Mun. Anorf, 12-16 km NW of Anorf, rd. to Dos Bocas, ca. 1000 m, 18 Nov 1989 (5), Callejas et al. 8753 (HUA, NY); Mun. San Rafael, nr. Cam- pamento Versalles, 1350 m, 23 Mar 1994 (5), Franco et al. 4576 (BG, HUA); Mun. San Luis, Rio Claro, 25 Mar 1994 (Y fl-fr), Franco et al. 4586 (BG, HUA); Valdivia, 1250 m, 27 Mar 1994 (Y fr), Franco et al. 4598 (BG, HUA); Mun. Anorf, Vrda. El Roble, 1400 m, 8 Mar 1995 (5), Franco et al. 5565 (BG, COL, HUA, LP); Mun. San Rafael, Vrda. La Luz, Rio Nare, 1300 m, 10 Jun 1993 (5), D. Sdnchez S. 1972 (MEDEL); valley of Rio Anorf, between Dos Bocas & Anorf, 23 km W of Zaragoza, 27 Aug 1976 (9 fl-fr), Shep- herd 596 (COL, MO); Mun. San Carlos, rd. Juanes-Alto Samana, km 12.6, 20 May 1988 (9 fl-fr), Zarucchi et al. 6756 (BG, MO). CHOc6: Mun. San Jose del Palmar, ca. 1 km from San Jos6 del Palmar, 1200 m, 19 Mar 1994 (9 fl- fr), Franco et al. 4569 (BG, HUA); 10 km W of Istmo de San Pablo, W of Las Animas, 12 Jan 1979 (9 fl-fr), Gentry et al. 24088 (BG, COL, HUA, MO); rd. Medellin-Quibd6, ca. 20 km W from top of divide of Cord. Occidental, 900- 1000 m, 3 Jan 1981 (9 fl-fr), Gentry et al. 30051 (COL, MO); nr. San Jos6 del Palmar, 1380 m, 11 Nov 1985 (9 fl- fr), Lozano C. et al. 4907 (COL, MO). NARINO: Nr. Alta- quer, 1150 m, 21 Feb 1995 (9 fl-fr), Franco et al. 4697 (COL, HUA). RISARALDA: Mun. Mistrato, rd. to Puerto de Oro, Rio Currumai, 29 May 1990 (9 fr), Orozco et al. 2181 (COL, MO). VALLE: Rio San Juan, below Queremal, 1300- 1500 m, 27 Mar 1947 (5), Cuatrecasas 23987 (K, US); nr. Buenaventura, 15 Mar 1994 (5), Franco et al. 4535 (BG); Mun. Buenaventura, rd. to Sabaletas, km 22, 17 Mar 1994 (9 fl-fr), Franco et al. 4552 (BG, HUA), km 24, (9 fl-fr), Franco et al. 4554 (BG, HUA); Rio Anchiyaca, nr. CVC hydroelectric plant, 15 Dec 1981 (9 fl-fr), Gentry 35671 (BG, COL, MO).
ECUADOR. AZUAY: Rd. La Troncal-Zhud, Manta Real, Rio Patul, 1200 m, 11-12 Jul 1991 (9 fl-fr), Yanez et al. 300 (QCA). CANAR: Rd. La Troncal-Suscal, ca. 20 km, 3 Feb 1981 (5), Berg 1239 (AAU, BG, COL, GB, MO, QCA, U). COTOPAXI: Rd. Quevedo-Latacunga, Tenefuerte, Rio Pilal6, 750-1300 m, 21 Feb 1982 (5), Dodson et al. 12769 (MO, QCNE). EL ORO: Rd. Pifias-Santa Rosa, km 11, 8 Nov 1979 (st), Dodson et al. 9005 (MO). ESMERAL- DAS: Cant6n Quininde, Bilsa Biological Station, 40 km NW of Quininde, 12 Mar 1997 (st), Clark et al. 4089 (BG). Los Rios/PICHINCHA: El Centinela, at crest of Montafias de Ila, rd. Patricia Pilar-24 de Mayo, km 12, 27 Nov 1978 (juv), Dodson 7303 (BG, MO, QCNE, SELBY). PICHINCHA: Rd. Quito-Puerto Quito, ca. 93 km from Nanegalito, 7 Feb 1982 (5), Berg et al. 1682 (AAU, BG, COL, QCNE); rd. Santo Domingo de los Colorados-Quevedo, km 21, 30 Jan 1979 (9 fl-fr), Dodson et al. 7413 (MO, QCNE, U); Reserva Flo- restal ENDESA, rd. Quito-Puerto Quito, km 113, 10 km from rd., 22-23 Nov 1984 (juv), Jaramillo 7436 (AAU, GB,
98 FLORA NEOTROPICA
MO, NY, QCA); Cant6n Pedro Vicente Maldonado, Reserva Rio Silanche, Nov 1996 (st), Pennington et al. 15230 (BG).
The species is very closely related to Cecropia garciae, as discussed under the latter. This species shows a variation in the trichilia from almost absent to even extended to the base of the stipules. In Valle (Colombia), a lowland and a submontane type appear to be present, probably morphologically distinct by different color of the stipules (green vs. dark red to [dark] purplish). The stipules are occasionally whitish due to dense arachnoid indumentum. The character- istic stiff (setose) hairs are absent in some collections made in Choco and Valle, being replaced by weaker hairs providing a subvillous indumentum. Ants are often absent, or if present, then a species of Pachy- condyla. These ants perforate the node bearing the stipules forming the apical bud cover and they harvest the (large and pink) Muillerian bodies when still en- closed in the bud.
21. Cecropia hololeuca Miquel, in Martius, Fl. Bras. 4(1): 148. 1853; Berg & Carauta, Albertoa 1(1): 6. 1986. Ambaiba hololeuca (Miquel) Kuntze, Re- vis. Gen. P1. 2: 624. 1891. Type. Brazil. Without locality, Pohl s.n. (holotype: W, destroyed), neo- type here designated: Brazil. Rio de Janeiro: Mun. Rio de Janeiro, Corcovado, Aug 1890 (Y fl-fr), Glaziou 18497 (IAN; isoneotypes: A, BM, C, E, G, K, NY, P).
Cecropia candida Snethlage, Notizbl. Bot. Gart. Berlin- Dahlem 8: 367. 1923. Type. Brazil. Rio de Janeiro, Serra de Macae, Ule 4863 (holotype: B, destroyed, photograph ex B in MO).
Tree, to 25 m tall. Leafy twigs 2-8 cm thick, green or slightly bluish to brown, glabrous, hirsute to vil- lous, and/or with arachnoid indumentum or some- times with only brown pluricellular hairs. Lamina coriaceous, ca. 40 X 40 cm to 120 X 120 cm, the segments 8-10, the free parts of the upper segments oblong to subobovate to oblanceolate, the incisions down to 7/10-9/10; apices rounded to obtuse to short- acuminate; upper surface glabrous, with ? dense arachnoid indumentum; lower surface with arachnoid indumentum in the areoles and on the smaller veins, or also on the main veins; lateral veins in the free part of the midsegment 12-15 pairs, submarginally loop- connected, branched or unbranched; petiole 30-95 cm long, glabrous, with arachnoid indumentum or also hirsute to subvillous at the base, or only with brown pluricellular hairs; trichilia absent; stipules 10-40 cm long, yellowish white to brownish, (partly) pale yel-
low to white-sericeous or -villous or also (or only) with dense brown pluricellular hairs and/or with (sometimes dense) arachnoid indumentum outside, densely sericeous inside. Staminate inflorescences in pairs, erect; peduncle 5-12 cm long, red to purplish, glabrous or (sub)villous at the base or also at the apex, with dense brown pluricellular hairs and/or sparse arachnoid indumentum; spathe lacking or with only bracts on the upper part of the peduncle, these to 0.5 cm long, caducous; spikes 9-17, 5-15 X 0.5-1 cm, often ? moniliform, wine-red to dark purple to al- most black, with stipes 0.8-2.5 cm long and glabrous; rachis hairy, with straight hairs or also arachnoid ones. Staminate flowers: perianth tubular, ca. 2 mm long, with short and stiff hairs in the ribes below the apex or glabrous, the apex almost plane; fil- aments flat; anthers ca. 0.6-0.8 mm long, short- appendiculate, detached and reattached to margins of the aperture at anthesis (?). Pistillate inforescences in pairs, erect, pendulous in fruit; peduncle 5-17 cm long, glabrous or (sub)villous, red(dish); spathe ab- sent, but with 1 or 2 bracts on the upper part or the apex of the peduncle, these to 1.5(-8) cm long, ca- ducous; spikes 1 or 2 (rarely 3), 5-12 X 0.5-1 cm, to 35 x 3 cm in fruit, initially red to purplish, turning blackish, sessile or with stipes to 1.5 cm long and glabrous or hairy; rachis hairy. Pistillateflowers: per- ianth ca. 2-2.5 mm long, with arachnoid indumentum below the apex outside, also in the lower part of the style channel inside, the apex ? convex, muriculate; style rather long, straight; stigma penicillate. Fruit el- lipsoid to oblongoid, ca. 3-4 mm long, ? tuberculate, dark brown.
Distribution (Fig. 8.6). Eastern Brazil, in moist forest, in mountainous and hilly regions, at elevations to 1300 m.
Representative specimens examined. BRAZIL. BA-
HIA: Rd. from Rd. Sao Jose-Buerarema to Una, km 6, 1 Apr 1980 (Y fl-fr), Berg et al. 1142 (BG, K, NY, U); rd. Itabela- Guaratinga, km 20-25, 31 Mar 1989 (st), Mattos Silva et al. 2674 (G); Santa Cruz de Cabralia, Reserva Biol6gica do Pau-brasil, 16 Sep 1971 (Y fl-fr), T S. Santos 1950 (U). ESPiRITO SANTO: Santa Teresa, 21 Nov 1988 (6), Bausen s.n. (GUA); Linhares, Reserva Florestal de CVRD, Est. Fla- mengo, 10 Feb 1981 (6), Folli 310 (BG, CRVD, GUA, MG). MINAs GERAIS: Juiz de Fora, Oct 1934 (6), Brade 14130 (RB); Viqosa, Fazenda Criciuma, 9 Sep 1967 (6 + Y fl-fr), Carauta 404 (GUA, K, RB, US); rd. BR.262, Mun. Man- huacu, 4 Dec 1984 (9 fl-fr), Hatschbach et al. 48598 (BG, C, MO); 5-10 km N of Teofilo Otoni, 15 Jul 1988 (9 fr), Hatschbach et al. 52174 (BG); Esta,co Experimental Co- ronel Pacheco, 11 Dec 1944 (st), Heringer 1685 (US); Mun. Marliera, Parque Estadual do Rio Doce, 18 Sep 1975 (6), Heringer et al. 15054 (US), 19 Sep 1975 (9 fr), Heringer
SYSTEMATIC TREATMENT 99
et al. 15057 (BG, MO, US); Carangola, 1 Sep 1990 (5), Leoni 1218 (GUA), (9 fl-fr), Leoni 1219 (GUA); Viqosa, 24 Feb 1930 (9 fl-fr), Mexia 4389 (BM, F, G, GB, K, MO, NY, S, U); Caldas, Sep 1867 (9 fl-fr), Regnell III.1100 (S); Ri- berao, Rio Novo, Sep 1894 (d), in herb. Schwacke 11128 (RB). RiO DE JANEIRO: Mun. Rio de Janeiro, Estrada da Vista Chinesa, 13 Mar 1963 (5), Angeli 350 (GUA, MG, RB, US); Cachoeiras do Macacu, Morro do Reservat6rio, 28 Feb 1968 (5), Carauta 581 (GUA, U, US); Rio de Ja- neiro, Estrada do Sumare, 15 May 1968 (9 fl-fr), Carauta 601 (GUA); Rio de Janeiro, Pedra de Gavea, 26 Feb 1977 (5), Carauta et al. 2328 (RB); Campos, Morro do Coco, Cruz da Serra, Morro do Bai, 13 Dec 1982 (d), Carauta et al. 4423 (GUA); Duas Barras, 13 Dec 1088 (d), Carauta et al. 5683 (GUA); Barra Mansa, Ribeirao de Sao Joaquim, Fazenda Campo Alegre, 18 Apr 1989 (5), Carauta et al. 5816 (GUA); Resende, Rio Palmital, 22 Feb 1966 (st), Cas- tellanos 25695 (GUA); Corcovado, Tijuca, 21 Jan 1891 (5), Glaziou 18498 (A, C, G, IAN, K, P); Larangeiras, 7 Sep 1890 (9 fr), Glaziou s.n. (P); Rio de Janeiro, Corcovado, Sumare, 20 Apr 1922 (5), Kuhlmann (RB) 3664 (RB, U); Rio de Janeiro, Horto Florestal, 20 Feb 1929 (9 fl), Kuhl- mann (RB) 111679 (GUA, RB, U); Rio de Janeiro, Estrada do Redentor, Tijuca, 1939 (9 fl-fr), Kuhlmann (RB) 136990 (RB, U); Rio de Janeiro, Corcovado, Sumare, Jun 1928 (5), Kuhlmann (RB) 141261 (RB), (9 fl-fr), Kuhlmann (RB) 141262 (RB); Mun. Mage, Paraiso, 2 Oct 1984 (9 fl-fr), Martinelli et al. 9908 (GUA, K, NY, RB). SAO PAULO:
Mun. Serra Negra, Alto da Serra, ca. 1000 m, 22 Nov 1991 (9 fl-fr), Barros et al. 2385 (SP); Represa do Jaguari, 31 Dec 1979 (9 fl-fr), Benson 10878 (UEC); Aruja, 12 Jul 1981 (9 fl-fr), Custodio Filho 654 (SP); Sao Jose dos Campos, 30 Aug 1949 (d), W Hoehne et al. 2004 (SP, SPF, US), (9
fl-fr), W Hoehne et al. 2005 (SP, SPF); Ibiti, 27 Jul 1943 (9
fl-fr), M. Kuhlmann 1326 (SP); Mun. Santa Izabel, Igarata, 27 Sep 1950 (9 fl-fr), M. Kuhlmann 2567 (NY, SP); Cam- pinas, 8 Jul 1988 (st), Matthes 2 (GUA); Braganqa Paulista, Fazenda San Antonio, 21 Sep 1991 (9 fl-fr), Mello-Silva et al. 547 (SP, SPF); Mogi das Cruzes, Rodovia dos Trabal- hadores, 22 Apr 1992 (5), Romaniuc et al. 1300 (SP); Mun. Amparo, rd. Amparo-Pedreira, 5 Apr 1993 (st), Romaniuc et al. 1366 (SP).
Cecropia hololeuca is the only species of the ge- nus lacking the characteristic spathe, fully enclosing the spikes before anthesis. Instead of the spathe, one or sometimes two bracts are found at the apex or just below the apex of the peduncle. These bracts are
mostly to 1.5 cm long, sometimes to 8 cm long. The
inflorescence remains enclosed in the apical bud until anthesis. This species resembles several Andean mon- tane species in the dense white arachnoid indumen- tum on the upper surface of the lamina and some of
them in the absence of trichilia and/or the presence of either smooth or densely hairy leafy twigs.
22. Cecropia idroboi Cuatrecasas, Revista Acad. Colomb. Ci. Exact. 9(36/37): 335. 1956. Type. Co- lombia. Vaupes: Mesa La Lindosa, 15-20 km S of San Jose del Guaviare, 13-15 Dec 1950 (Y), Id- robo & Schultes 662 (holotype: US; isotypes: BM, COL, GH, MO, NY, U, US).
Tree, to 10 m tall. Leafy twigs 1.5-2.5 cm thick, hispidulous to puberulous. Lamina (sub)coriaceous, ca. 15 X 15 cm to 40 X 40 cm, the segments 7-8, the free parts of upper segments (broadly) obovate to elliptic, the incisions down to 3/10-5/10; apices rounded or apiculate; upper surface smooth to scabri- dulous, sparsely hispidulous to strigillose; lower sur- face puberulous on the main veins or also on the smaller veins, with arachnoid indumentum in the ar- eoles or also sparsely to densely on the lateral veins or also on the main veins, sometimes the whole sur- face with dense arachnoid indumentum; lateral veins in the free part of the midsegment 8-10(-12) pairs, marginally loop-connected, most of them branched; petiole ca. 10-40 cm long, sparsely puberulous or also with dense arachnoid indumentum; trichilia fused, the brown indumentum intermixed with sparse to rather dense short brownish or whitish unicellular hairs, sometimes with dense arachnoid indumentum; stip- ules 3-7 cm long, dark red (?), sparsely puberulous to hirtellous, sometimes with dense arachnoid indu- mentum outside, hairy inside. Staminate inflores- cences in pairs or solitary, erect (?); peduncle 6-8 cm long, hispidulous; spathe 5-6 cm long, dark red, sparsely puberulous (to subglabrous) outside, gla- brous inside; spikes ca. 15, 1.5-5 X 0.2-0.3 cm, with stipes to 0.3 cm long and densely puberulous; rachis hairy. Staminate flowers: perianth tubular, 0.8-1 mm long, with sparse arachnoid indumentum below the apex, the apex almost plane, glabrous; anthers 0.5- 0.6 cm long, detached at anthesis (?). Pistillate info- rescences in pairs, erect (or deflexed in fruit?); pe- duncle 3-6.5 cm long, hirtellous to puberulous or also with ? dense arachnoid indumentum; spathe 4-5 cm long, the color and indumentum as in the staminate inflorescence; spikes 4-5, 3-6 X ca. 0.8 cm, to 9 x ca. 1.3 cm in fruit, (sub)sessile. Pistillateflowers: per- ianth 1.5-2 mm long, with arachnoid indumentum be- low the apex, also in the lower part of the style chan- nel inside, the apex convex, muriculate to granulate, the aperture slit-shaped; style long, S-shaped; stigma comose. Fruit oblongoid, ca. 2 mm long, smooth, dark brown.
100 FLORA NEOTROPICA
Distribution (Fig. 10.6). Colombia (Caqueta, Meta, and Vaupes), on sandstone rocks, at low ele- vations.
Specimens examined. COLOMBIA. CAQUETA: Sierra de Chiribiquete, 7 Dec 1990 (Y), Castroviejo et al. 11978 (COL). META: La Macarena, Rio Guayabero, Jan-Mar 1959 (? fl-fr), Garcia-Barriga et al. 17053 (COL, NY, US). VAUPES: Rio Kuduyari, Cerro Yapoboda, 5 Oct 1951 (Y fl- fr), Schultes et al. 14240 (COL, US), 5-6 Oct 1951 (Y fl- fr), Schultes 14370 (COL, GH, US), 17 Nov 1952 (Y fl-fr), Schultes et al. 18500 (BG, K, MO, NY), 9 Oct 1966 (5), Schultes 24272 (COL).
This species shows strong morphological affinities to Cecropia distachya; these two taxa might prove to be conspecific. Cecropia idroboi differs from C. dis- tachya in the less deeply incised lamina, the smaller number of lateral veins in the free part of the midseg- ment, and the shorter spikes of the pistillate inflores- cence. The trees remain smaller than on average in C. distachya. Cecropia idroboi is probably ecologically distinct, as a species growing on rocks or in rocky places. However, specimens which could be referred to C. distachya without doubt, have been collected in the same area and in (almost) similar habitats. Garcfa- Barriga 17053 matches C. idroboi in most features, but it is quite distinct in the dense arachnoid indu- mentum on the petiole, lower surface of the lamina, and the peduncle, and in this respect similar to C. metensis, found in the same locality.
23. Cecropia insignis Liebmann, Kongel. Danske Vi- dens. Selsk. Naturvidensk. Math Afh., ser. 5, 2: 281. 1851; Burger, Fieldiana, Bot. 40: 125, t. 23. 1977; Croat, Fl. Barro Colorado Island 346, t. 194. 1978; Berg & Franco Rosselli, Fl. Ecuador 48: 29. 1993. Type. Nicaragua. Rio San Juan, Oersted s.n. (type material not yet traced. Oersted 5806 (C), also from Rio San Juan does not match Lieb- mann's description and belongs to C. peltata), herewith replaced by: Nicaragua. Rio San Juan: Nr. Caiio Chontaleno, 20 km NE of El Castillo, 7- 9 Mar 1978 (9), Neill 3392 (neotype: MO; iso- neotype: BG). Fig. 28
Cecropia eximia Cuatrecasas, Revista Acad. Colomb. Ci. Exact. 6(22/23): 287. 1945. Type. Colombia. Valle: Rio Calima, La Trojita, 19 Feb-10 Mar 1944 (Y), Cuatrecasas 16830 (holotype: COL; isotypes: F, US).
Cecropia sandersoniana P. H. Allen, The rain forest of Golfo Dulce 162 & 409 (as C. standleyana). 1956. Type. Costa Rica. Puntarenas: Nr. Golfito, 4 Jan 1953 (9), Allen 6688 (holotype: EAP; isotype: US).
Cecropia polyandrophora Cuatrecasas, Phytologia 20: 465. 1971. Type. Colombia. Antioquia: between Villa Artega & Chigorod6, La Pradera, 3 Oct 1961 (i), Cuatrecasas & Willard 26187 (holotype: US; isotypes: COL, US).
Tree, to 25(-40) m tall. Leafy twigs 2-5 cm thick, reddish-brown to blackish, (rather) sparsely hispidu- lous. Lamina coriaceous (and sometimes ? plicate) to subcoriaceous, ca. 50 X 35 cm to 100 X 85 cm (to 120 X 100 cm), the segments 7-8(-9), the free parts of upper segments elliptic to (sub)obovate to oblong, the incisions down to ca. 7/10; apices obtuse; upper surface smooth, initially with sparse to rather dense arachnoid indumentum; lower surface with sparse to dense brown pluricellular hairs on the main veins, with arachnoid indumentum in the areoles, on the smaller veins, and (at least initially) also on the main veins; lateral veins in the free part of the midseg- ment, 12-18(-24) pairs, submarginally loop- connected, unbranched; petiole 35-80 cm long, red- brown to grayish, with dense brown pluricellular hairs and dense arachnoid indumentum; trichilia fused or separate, the brown indumentum intermixed with short stiff white to brownish (unicellular) hairs; stipules 22-50 cm long, pinkish to red to red-brown, caducous (or subpersistent), subsericeous to hirtel- lous to hispidulous and with dense brown pluricel- lular hairs outside, + densely sericeous inside. Sta- minate inflorescences in pairs, erect; peduncle 3-7 cm long, hispidulous to puberulous; spathe 12-16(- 22) cm long, pinkish to red to red-brown, puberulous to hispidulous or to strigillose or also brown pluri- cellular hairs and/or (rather) sparse arachnoid indu- mentum outside, sparsely to densely hairy or gla- brous inside; spikes 5-9 and 8-10 X 0.6-1.2 cm or ca. 15-35 and 6.5-14.5 X 0.3-0.5 cm, with stipes 0.5-2 cm long and glabrous; rachis hairy. Staminate flowers: perianth tubular, ca. 1.5-2 mm long, with short and stiff hairs below the apex, the apex almost plane; filaments flat; anthers ca. 0.7 mm long, appen- diculate, detached at anthesis (?). Pistillate inflores- cences in pairs or solitary, initially erect, pendulous in fruit; peduncle 9-15 cm long, green, hispid or subglabrous; spathe 10-22 cm long, the color and in- dumentum as in the staminate inflorescence; spikes (2-)4-5(-7), 8-18 X ca. 0.8 cm, to 26 X ca. 1 cm in fruit, with stipes 0.5-1 cm long and glabrous; rachis hairy. Pistillate flowers: perianth ca. 2 mm long, with arachnoid indumentum below the apex, also in the upper part of the style channel inside, the apex convex or almost plane, (sub)muriculate, ? angular in circumference, the apical part long, the aperture slit-shaped to circular, bordered by a rim; style long,
SYSTEMATIC TREATMENT 101
gi~~~~~~~~~ Tn .+
5crn.
FIG. 28. Cecropia insignis. 1. Lamina, reduced. 2. Apex of lamina and venation (Berg 1240). 3. Stipules, young leaf, and base of petiole with trichilium (Berg 1260). 4. Staminate inflorescence with spathe. 5. Staminate inflorescence at anthesis. 6. Staminate flower and stamen (Berg 1240). 7. Pistillate inflorescences, one with spathe, the other at anthesis, and base of petiole with trichilium (Berg 1260). 8. Pistillate flower. 9. Fruit (Franco et al. 5560).
102 FLORA NEOTROPICA
curved; stigma truncate, comose. Fruit oblongoid to ellipsoid, 2-2.5 mm long, ? tuberculate.
Distribution (Fig. 10.6). From Honduras to the Pacific coastal region of Ecuador, and the Magdalena valley (up to Tolima) and the valley of Rio Porce and Rio Medellin to near Medellin, in (treefall gaps in) primary upland forest, riverine forest, and in second- ary growth, at elevations to 1000 m, in Costa Rica to 1400 m.
Representative specimens examined. HONDURAS. OLANCHO: Between Campamento & border Olancho/Mor- azan, rd. Tegucigalpaca-Juticalpa, ca. km 100-101, 27 Oct 1996 (Y), Berg 1809 (EAP).
NICARAGUA. BLUEFIELDS: Base of Cerro San Isidro, Rfo Kama-Rio Escondido, 25 Mar 1966 (d), Proctor et al. 27245 (NY). JINOTEGA: Las Brisas, Comarca Kilambe, 930 m, 13 Jul 1980 (Y fr), Sandino 168 (BG, MO). ZELAYA: S of Rio Wawa, 60 km NW of Puerto Cabezas, 17 Mar 1972 (? fl), Little 25201 (MO, US); nr. Rfo Wawa, 60 km NW of Puerto Cabezas, 27 Mar 1971 (5), Little 25340 (MO, US); 20 km W of Awas Tingni, S of Rio Wawa, 1 Apr 1971 (Y fl), Little et al. 25356 (MO); 20 km NW of Alamicamba, 16 Apr 1971 (st), Little 25382 (MO, US); new rd. Siuna-Ma- tagalpa, ca. 10.1 km beyond Rio Uli, nr. Wani, 6 May 1978 (Y fl-fr), Stevens 8813 (MO).
COSTA RICA. CARTAGO: Turrialba, ( Y fl), Blum 2218 (MO); 0.5 km NE of Tapanti, 1360 m, 11 Jun 1967 (Y fl), Lent 1031 (MO, NY); 0.5 km S of Penias Blancas de Cachi, 1400 m, 5 Sep 1971 (Y fr), Lent 2119 (MO, NY); Turrialba, 1100 m, 7 Jul 1992 (Y fl-fr), Rivera 1933 (K). PUNTAR-
ENAS: Finca Loma Linda, 1 mi SW of Cafnas Gordas, 1150 m, 26-27 Feb 1973 (Y fl), Croat 22273 (MO); Parque Na- cional Corcovado, Sirena Woods, 30 Apr 1989 (? fl-fr), Ker- nan 1064 (BG); San Vito, Las Cruces Botanical Garden, 1250-1275 m, 4 Jul 1994 ( fl-fr), Kress et al. 94-422 (US); Osa Peninsula, Aguabuena, 3.5 km W of Rinc6n, 19 Jan 1993 (9 fl), Thomsen 541 (BG), 17 Apr 1993 (st), Thomsen 873 (BG).
PANAMA. COCLE: Cerro Pil6n, 1968 (st), Lallathin 5065 (MO). COLON: 10 mi N of Gamboa, 14 May 1973 (st), Gentry 7451 (BG); Salud, 4 Aug 1971 (9 fl), Lao et al. 211 (H, PMA). Los SANTOS: 12 mi S of Macaracas, 22 Feb 1966 (5), Blum et al. 2176 (MO); 16 mi S of Macaracas, Quebrada Bejuco, 22 Jan 1966 (d), Tyson et al. 2927 (MO). PANAMA: Nr. El Llano, 3 Sep 1977 (st), Berg et al. 410A (BG); Barro Colorado Is., 9 Jan 1969 (5), Croat 7023 (MO), 2 Feb 1971 (5), Croat 13211 (MO), 29 Dec 1970 (5), Fos- ter 2052 (PMA); J.W.T.C. Headquarters, 1 Apr 1956 (9), Johnston 1758 (A, MO). SAN BLAS: Nusagandi, 18 Aug 1989 (st), Fisher 33 (BG).
COLOMBIA. ANTIOQUIA: Rio Le6n (or Bacubd), be- tween Villa Arteaga & Chigorod6, La Pedrara, 3 Oct 1961 (9 fl-fr), Cuatrecasas et al. 26186 (COL, US); Mun. San Luis, Rio Claro, 25 Mar 1994 (d), Franco et al. 4585 (BG, COL, HUA); Mun. Taraza, Cgto. El Doce, Finca Las Mer- cedes, 28 Mar 1994 (d), Franco et al. 4600 (BG, COL, HUA); Mun. Frontino, Cgto. Nutibara, rd. Nutibara-La
Blanquita, Murri region, 4 Mar 1995 (9 fl-fr), Franco et al. 5560 (BG, COL, HUA, LP, MO); Mun. San Luis, Quebrada La Cristalina, 26 Feb 1987 (9 fl-fr), Ramirez et al. 691 (COL); valley of Rio Anorf, 23 km W of Zaragoza, between Dos Bocas & Anorn, 31 Aug 1976 (st), Shepherd 606 (COL, HUA); Mun. Zaragoza, Cgto. Providencia, 9 Feb 1971 (9 fl-fr), Soejarto et al. 2661 (GH, HUA); Mun. San Carlos, Cgto. El Jordan, Trocha Los Planes, 9 Apr 1991 (9 fr), Ma. Veldsquez et al. 293 (HUA). BOYACA: Mun. Puerto Boyaca, Vrda. Puerto Pinz6n, 17 Jan 1998 (st), Cortes et al. 1546 (UDBC). CHOCO: Colombia-Panama border, Alto del Lim6, 23 Sep 1979 (9 fl-fr), Barbosa 1191 (COL), (d), Barbosa 1194 (COL); Mun. Riosucio, Uraba region, Cerros del Cuchillo, 18 Nov 1987 (9 fl), Cdrdenas 897 (JAUM, MO); nr. Bahia Solano, 4 Apr 1990 (9 fl-fr), Franco et al. 3006 (COL); Mun. San Jose del Palmar, 1-2 km W of San Jos6 del Palmar, 19 Mar 1994 (d), Franco et al. 4567 (BG, COL, HUA), (9 fl), Franco et al. 4568 (BG, COL, HUA); Nuquf, Alto de Buey, 1000 m, 23 Jun 1940 (9 fl-fr), Snei- dern A.61 (A, COL, K, LL, MEDEL, MICH, NY, S, US); Rio Jurad6, 4 Oct 1940 (9 fl-fr), Sneidern A.245 (A, COL, NY); Parque Nacional Natural Los Katios, Alto del Lim6n, 9 May 1983 (9 fl-fr), Zuluaga R. 682 (COL). TOLIMA:
Mariquita, 17 May 1990 (9 fl-fr), Franco et al. 2965 (COL), 7 Feb 1995 (9 fl-fr), Franco et al. 4654 (BG, COL, HUA). VALLE: Between El Aguacate & Quebrada de la Yuca, 8 Feb 1944 (9 fl), Cuatrecasas 16096 (F, US, VALLE); Rio Calima, Aguaclara, 23 May 1946 (9 fr), Cuatrecasas 21262 (F, US, VALLE); Rio Calima, La Brea, 25 May 1946 (st), Cuatrecasas 21315 (US); Rio Anchicaya, Rio Blanco, 29 Mar 1949 (9 fl-fr), Cuatrecasas 24009 (US), (d), Cuatre- casas 24010 (F); Mun. Cisneros, 15 Mar 1994 (d), Franco et al. 4532 (BG, COL, HUA).
ECUADOR. ESMERALDAS: Rd. Santo Domingo de los Colorados-Quininde, km 171-188, Rio Cocola, 17 Feb 1950 (st), Acosta S. 16221 (F); Fila Bilsa, 7 km E of San Jose de Bilsa, ca. 80 km S of Esmeraldas, 30 Jan 1991 (st), Gentry et al. 72939A (MO); between Rio Blanco & Rio Chipo, 18 Aug 1967 (9 fl-fr), Ja'tiva et al. 2233 (NY); 3 km E of Quinind6, 10 Apr 1943 (st), Little 6220 (Q, US); Rio Guayl- labamba, 10 km E of Quinind6, 4 Oct 1965 (d), Little et al. 21224 (NY, Q, QAME, QCNE, US); Reserva Ecol6gica Cotacachi-Cayapas, Charco Vicente, Rio San Miguel, 6-9 Sep 1993 (st), Palacios et al. 11164 (QCNE); Anchayacu, Eloy Alfaro, Mayronga, 21 Oct 1993 (9 fr), Pennington et al. 14176 (BG); Cant6n Eloy Alfaro, Reserva Ecol6gica Co- tacahi Cayapas, Rio Santiago, Angostura, 19-24 Sep 1994 (st), Tirado et al. 1443 (QCNE). IMBABURA: Nr. Lita, 7 Feb 1981 (5), Berg 1240 (BG, COL, QCA, U), 9 Feb 1981 (9 fl-fr), Berg 1260 (AAU, BG, COL, MO, NY, QCA, U). Los Rios: Rfo Palenque Biological Station, rd. Quevedo-Santo Domingo de los Colorados, km 56, 8 Jun 1974 (9 fl-fr), Dodson 5579 (US), 11 Aug 1977 (9 fl-fr), Dodson et al. 6834 (AAU, QCA, US). PICHINCHA: Cant6n Pedro Vicente Maldonado, Reserva Rio Silanche, Nov 1996 (9 fl), Pen- nington et al. 15246 (BG); Cant6n Pedro Vicente Maldon- ado, Reserva Forestal Rio Pitzara, Nov 1997 (st), Pennington et al. 16112 (BG).
SYSTEMATIC TREATMENT 103
The staminate inflorescences have only 5-9 spikes, 0.6-1.2 cm diam., in the northern and southern part of the distribution range. However, in Colombia (Antioquia, Choc6, and Valle) the staminate inflores- cences have ca. 15-35 spikes, 0.2-0.5 cm diam. This very different type of staminate inflorescence is rep- resented by the collections Cuatrecasas et al. 26187 and Franco et al. 4532, 4585, and 4600, which are in their vegetative parts similar to the other collections. Material with pistillate inflorescences from the same region are not different. This species shows morpho- logical and ecological similarities to the Amazonian Cecropia distachya, both becoming tall forest trees with coriaceous and mostly smooth laminas. Cecro- pia insignis also shows close morphological affinities to the montane C. plicata; the latter is distinct by the presence of arachnoid indumentum on the leafy twigs and more numerous segments of the lamina.
24. Cecropia kavanayensis Cuatrecasas, Fieldiana, Bot. 28: 210. 1951; Velasquez, Acta Bot. Venez. 6: 41, t. 6. 1971; Berg in Gorts-van Rijn, Fl. Guianas, ser. A, fasc. 11: 97. 1992. Type. Vene- zuela, Bolivar: Between Santa Teresita de Kavan- ayen & Rio Pacairao, 20-21 Nov 1944 (6), Stey- ermark 62754 (holotype: US; isotype: F).
Cecropia auyantepuiana Cuatrecasas, Acta Bot. Venez. 2: 203. 1967; Veldsquez, Acta Bot. Venez. 6: 41, t. 5. 1971. Type. Venezuela. Bolfvar: Cerro Auyan- tepui, Rio Lomita camp, 1800 m, 10 May 1964 (6), Steyermark 93643 (holotype: US).
Cecropia steyennarkii Cuatrecasas, Bol. Soc. Venez. Ci. Nat. 32: 321. 1976. Type. Venezuela. Bolivar: Mes- eta del Jaua, Cerro Jaua, 1810-1880 m, 28 Feb-5 Mar 1974 (6), Steyermark 109842 (holotype: US; isotypes: K, U).
Tree, to 15(-30) m tall. Leafy twigs 4-15 cm thick, puberulous, sometimes also with sparse arachnoid in- dumentum, uncinate hairs lacking. Lamina coria- ceous, ca. 40 X 40 cm to 90 X 90 cm, the segments 7-8, the free parts of the upper segments elliptic to oblong to (sub)obovate or to ovate, the incisions down to 5/10-6/10(-8/10?); apices rounded to obtuse; up- per surface smooth to scabrous, sparsely to rather densely hispidulous (or if juvenile, then densely hir- tellous); lower surface hirtellous to tomentellous to puberulous (without uncinate hairs) on the veins, with arachnoid indumentum in the areoles, also on the smaller veins and along the margin, sometimes ex- tending to the main veins; lateral veins in the free part of the midsegment 12-16 pairs, submarginally loop- connected, (the lower ones) branched; petiole 40-85
cm long, dark purple, sparsely puberulous to hirtel- lous or to hirsute and with (rather) dense arachnoid indumentum; trichilia fused, the brown indumentum intermixed with short brownish to whitish (unicellu- lar) hairs; stipules 15-35 cm long, red-brown, densely to rather sparsely hirtellous to subhirsute and often also rather sparse arachnoid indumentum outside, sparsely to densely subsericeous inside. Staminate in- florescences in pairs, the peduncle erect (?); peduncle 4-10 cm long, sparsely puberulous; spathe 9-12 cm long, red-brown, puberulous to hirtellous to subhir- sute, also with ? dense brown pluricellular hairs, and often with sparse to dense arachnoid indumentum out- side, glabrous inside; spikes 4-15, 3-7 X 0.3-0.7 cm, with stipes to 1.2 cm long and subglabrous; rachis hairy. Staminate flowers: perianth tubular, 1.5-2.5 mm long, with sparse straight hairs or arachnoid in- dumentum below the apex, the apex almost plane, smooth; filaments flat; anthers 0.5-0.7 mm long, ap- pendiculate, detached at anthesis, reattached to the margins of the aperture by the appendages. Pistillate inflorescences in pairs, pendulous; peduncle 9-12 cm long, glabrous to sparsely puberulous to hirtellous; spathe 9-11 cm long, the color and indumentum as in the staminate inflorescence; spikes 4, (4-)8-10 X ca. 1.2 cm, to 22 X 1.8 cm in fruit, (sub)sessile or with spikes to 0.4 cm long and puberulous; rachis hairy. Pistillate flowers: perianth 4-4.5 mm long, with arachnoid indumentum below the apex outside, also below the style channel inside, the apex convex, smooth or muriculate; style long, ? S-shaped; stigma penicillate. Fruit oblongoid, ca. 3 mm long, tubercu- late.
Distribution (Fig. 10.1). Eastern Venezuela and adjacent Brazil (Roraima), in forest, at 1000-1900 m.
Specimens examined. VENEZUELA. AMAZONAS:
Depto. Rio Negro, Cerro de La Neblina Camp V, 1250 m, 21-24 Mar 1984 (d), Liesner et al. 16945 (BG), (Y fl-fr), Liesner et al. 16951 (BG). BOLiVAR: Apacara, Rio Caroni, 1500 m, 12 Nov 1946 (st), Cardona 1951 (US, VEN); Mun. Cedetio, Serranfa de Maigualida, 20 km E of San Jose de Kayama, 1250 m, Apr 1989 (Y fl-fr), Ferndndez 5505 (BG, MO); Mun. Roscio, N of San Juan de Kamoirin, ca. 10 km S of La Ciudadela, 1300 m, 6 Mar (Y fr), Huber 9235 (BG, NY, VEN); rd. Kavanayen-Santa Elena, km 13, 1200 m, Maas et al. 5384 (K, MO, U); Depto. Sifontes, Parupa, To- ron Meru, 1300 m, 25 Apr 1986 (d), Pic6n 1108 (BG); Cerro Uananapan, S of Uei-tepui, 1330-1450 m, 25 Apr 1960 (st), Steyernark et al. 758 (NY, US); Mt. Roraima, between Quebrada Ka-hua-parn & Glycon swamp, 1220- 1980 m, 1 Oct 1944 (st), Steyermark 59016 (F, NY); Cerro Venamo, Rfo Venamo, nr. Guyana border, 900-1000 m, Stey- ermark et al. 92863 (US, VEN); drainage of Rfo Cuyuni, 130-131 km S of El Dorado, ca. 1300 m, 19-26 Dec 1970
104 FLORA NEOTROPICA
(Y fl-fr), Steyermark et al. 104065 (K, NY, S, U, VEN); Meseta del Jaua, Cerro Jaua, 1810-1880 m, 28 Feb-5 Mar 1974 ($ fl-fr), Steyennark et al. 109843 (K, U, US); Mt. Roraima, Arabapu, ca. 1400 m, 1 Jan 1928 (st, juv), Tate 250 (NY).
BRAZIL. RORAIMA: Maita Mtns., 3?15'N, 63?38'W, 12 Feb 1971 (d), Prance et al. 10473 (INPA, NY, U, US).
This species shows morphological affinities to both Cecropia obtusa and C. ficifolia. It is myrme- cophytic.
Cecropia orinocensis Standley, Publ. Field Mus. Nat. Hist., Bot. Ser. 22: 14. 1940; Velasquez, Acta Bot. Venez. 6: 47, t. 9. 1971. Type. Venezuela. Amazonas: Rio Orinoco, mouth of Cafio Carida, 12-24 Jan 1930 (Y), Holt & Gehriger 248 (holotype: US; isotype: NY).
Tree, to 15 (-20) m tall, the trunk with rather prom- inent stipular scars. Leafy twigs 1.5-3 cm thick, green to red-brown to purplish black, hispidulous. Lamina chartaceous, ca. 20 X 20 cm to 55 X 55 cm (to 70 X 70 cm), the segments 8-9, the free parts of upper seg- ments elliptic to oblong, the incisions (in the upper part of the lamina) down to 5/10-7/10, the lamina attached to the petiole rather close to its base; apices obtuse; upper surface ? scabrous, hirtellous to his- pidulous; lower surface puberulous on the veins and with dense or sparse arachnoid indumentum in the areoles; lateral veins in the free part of the midseg- ment 12-14 pairs, marginally loop-connected, often branched, running (almost) straight into the margin; petiole 25-60 cm long, puberulous; trichilia fused, with only brown indumentum; stipules 8-13(-22), bright to dark red to brownish (or green), caducous or subpersistent, puberulous or sparsely subsericeous outside, sparsely villous inside. Staminate inflores- cences in pairs, the peduncle erect and the spikes pen- dulous; peduncle 4-10 cm, hispidulous to hirtellous to subhirsute; spathe (5-)9-12(-18) cm long, pink to red, puberulous or also with (rather sparse) arachnoid
indumentum outside, glabrous inside; spikes (6-)15- 20, (3-)14-20 X 0.3 cm, with stipes (0.2-)0.4-0.6 cm long and hairy; rachis hairy. Staminate flowers: perianth tubular, 0.6-0.8 mm long, sparsely puberu- lous just below the apex, the apex slightly convex to plane, sparsely puberulous on the margin, the aperture surrounded by a rim; filaments flat; anthers 0.3-0.5 mm long, remaining attached to the filament by 2 fili- form connections between the connective and the up- per margin of the filament at anthesis. Pistillate inflo- rescences in pairs, deflexed to pendulous; peduncle 5-17 cm long, hispidulous to hirtellous to subhirsute; spathe 7-15 cm long, the color and indumentum as in the staminate inflorescence; spikes 4-6, 3-12 X 0.5-0.8 cm, to 25 X 1.5-2 cm in fruit, sessile (or with stipes to 0.5 cm long and glabrous; rachis glabrous. Pistillate flowers: perianth tubular, 4 mm long, with arachnoid indumentum below the apex outside, also below the style channel (or absent) inside, the apex slightly convex to slightly concave, smooth or punc- tate, the aperture slit-shaped, bordered by a rim; style long, curved; stigma comose. Fruit oblongoid, ca. 3 mm long, finely tuberculate to (almost) smooth.
Distribution (Fig. 8.1). Amazon basin, the basin of Rio Orinoco, and in the Guianas, in periodically inundated places, especially along black-water rivers (on nutrient-poor and not well-aerated soil) and on sand savannas, at low elevations.
Representative specimens examined. COLOMBIA. AMAZONAS: Mun. Leticia, Quebrada de Arara, 27 Jan 1969 (d), Cuatrecasas et al. 27250 (COL, F, G, K, NY, US); Rio Loretoyacu, Nov 1945 (d), Duque-Jaramillo 2030 (COL); Rio Yari, nr. mouth of Quebrada El Mochilero, 23 Apr 1986 (st), Galeano et al. 1093 (COL); Rio Caquetd, Isla El Tigre, 23 Sep 1988 (st), M. Sdnchez S. et al. 1473 (BG, U); Rio Putumayo, between Rio Igara Parand & Rio Yaguas, 20 Jun 1942 (st), Schultes 3996 (ECON, F); Trapecio Amaz6nico, Rfo Loretoyacu, Oct 1945 (d), Schultes 6720 (COL, F); Amacayacu National Park, Matamata, 21 Jan 1989 (Y fr), Tye C.92 (BG, K); Puerto Narifno, Parque Nacional Ama- cayacu, 2 Aug 1989 (st), R. Vdsquez et al. 12449 (COL, MO). AMAZONAS/VAUPES: Rio Apaporis, Jinogoj6, 20 Jun 1952 (Y fl-fr), Schultes et al. 16811 (ECON, US). CA- QUETA: Rio Yari, nr. Araracuara, 15 Jul 1986 (Y fr), Berg et al. 1562 (BG); Rio Yari, nr. mouth of Quebrada El Mo- chilero, 25 Apr 1986 (? fl-fr), Galeano et al. 1093 (COL); Rio Orteguaza, between Peneya & Getucha, 28 Jul 1926 (st), Woronow et al. 6302 (LE). PUTUMAYO: Rio Caucaya, be- tween Vivianacocha & La Peinilla, 10-14 May 1993 (Y fr), Bernal et al. 2039 (COL).
VENEZUELA. AMAZONAS: Depto. Atabapo, Rio Parn, Sep 1989 (Y fr), Delgado 720 (BG, NY); Rio Casiquiare, Jan-Feb 1969 (Y fl-fr), Farinas et al. 502 (US, VEN); Depto. Atabapo, Cafno Jayuwapuey, affluent of Rio Ocamo, Jan 1990 (2 fl), Ferndndez 6775 (BG, NY); Brazo Casi-
FIG. 29. Cecropia latiloba. 1. Leaf, stipules, and trichilia. 2. Apex of lamina and venation. 3. Pair of staminate inflorescences and base of petiole with trichilium. 4. Staminate inflorescence at anthesis and base of petiole with trichilium
(Berg et al. 1057). 5. Staminate flower and stamen after detachment of anther (Prance et al. 20925). 6. Pistillate inflorescence at anthesis and base of petiole with trichilium (Berg et al. 1032). 7. Pistillate inflorescence after anthesis and base of petiole with trichilium (Berg et al. 1575). 8. Pistillate flower. 9. Fruit (Berg et al. 1562).
106 FLORA NEOTROPICA
quiare, 1-20 Feb 1931, Holt et al. 657 (NY, US); Rio Siapa, between mouth & campamento de los Yanomani- Emonitheri, 17-20 Sep 1986 (Y fl-fr), Stergios et al. 9401 (BG, MO, NY, VEN); Lower Rio Casiquiare, Solano, 10 Mar 1942 (? fr), LI. Williams 14733 (F, G); Upper Rio Casi- quiare, Capihuara, 3 Jun 1942 (? fr), Ll. Williams 15765 (G, US); Rio Casiquiare, between mouth of Rio Siapa & mouth of Rio Paciba, 26 Jul 1959 (Y fr), Wurdack et al. 43634 (BG, NY). APURE: Dtto. San Fernando, between Rfo Ori- noco, opposite Isla Peladura & lagunas ca. 1 km inland, 25- 26 Apr 1977 (6), Davidse et al. 12211A (MO, U, VEN); Dtto. San Fernando, mouth of Rio Arauca, 14-15 May 1977 (Y fl), Davidse et al. 13258 (MO, VEN); Dtto. Pedro Ca- mejo, Rio Orinoco, opposite Isla Pantallo, 24-25 Feb 1978 (d), Davidse et al. 14500 (MO, U, VEN); Dtto. Pedro Ca- mejo, Rio Orinoco, Isla El Fraile, 3 Nov 1989 (Y fr), Duno et al. 174 (BG). BOLiVAR: Mun. Raul Leoni, 25 km N of Macizo Ichun, Alto Rio Paragua, Apr 1988 (Y fl-fr), Fer- ndndez 4399 (MO); Mun. Raul Leoni, 50 km SW of Macizo Guaiquinima, 72 km W of Karnn, Nov 1988 (Y fr), Fer- ndndez et al. 4819 (MO); Rio Nichare, between mouth & Canio Sarrapio, 3-10 Aug 1985 (Y fr), Homer et al. 73 (MO); Rfo Oris, nr. Campamento Turumban, 13 May-13 Jun 1987 (6), Stergios 10800 (BG, NY); La Paragua, 25 Mar 1940 (6), Ll. Williams 12689 (F, K, S, US, VEN). DELTA AMACURO: Dtto. Antonio Diaz, Rio Acure, between Las Margaritas & Curiapo, 31 Jan 1980 (6), A. Femnandez et al. 3527 (U); Sacupana, Apr 1986 (Y fl), Rusby et al. 273 (A, BM, E, F, G, GH, K, MICH, MO, NY, VEN); Rio Cu- yubini, between la Paloma & Moron, 20 Nov 1960 (6), Steyermark 87693 (BG, NY); between La Margarita & Puerto Miranda, Rio Acure, 26 Nov 1960 (d), Steyermark 87808 (BG, NY, VEN); Depto. Antonio Diaz, Rio Cuyubini, between mouths of Rio El Toro & Rio Mecoro-uabanoco, 3 Nov 1980 (Y fr), Trujillo et al. 17307 (NY, U, VEN).
GUYANA. Mazaruni R., Puruni R., 21 Apr 1953 (Y fl- fr), "J.B." 85 (= FD 7769) (K, NY, U); U. Takutu-U. Es- sequibo region, Rewa R., 0-5 km S of confluence with Kwi- taro R., 25 Feb 1997 (6), Clarke 3937 (BG, US); Cuyuni- Mazaruni region, Cuyuni R., between Aurora & 4 km upstream, 8 Oct 1989 (Y fl-fr), Gillespie et al. 2195 (BG, COL); Essequibo R., Rockstone, 31 Jul 1921 (Y fr), Gleason 864 (GH, NY); Demerara-Mahaica region, 23 km E & W of Linden Hwy., within 3 km of second tollgate, 16 Aug 1993 (6), Henkel et al. 2584 (BG); Upper Takutu-Upper Esse- quibo region, S of Rupununi Savanna, Kidekperdana Cr., 8 km SE of Aishalton, 26 Nov 1993 (Y fl-fr), Henkel et al. 3519 (BG); Upper Takutu-Upper Essequibo region, Kassa- kaityu, 24 Mar 1994 (6), Henkel et al. 5318 (BG); Kakunu Mtns., Rupununi R., Crabwood Cr., 27 Jun 1995 (6), Jansen-Jacobs et al. 4231 (BG, U); Rupununi R., SW of Mt. Makarapan, 10 Sep 1988 (Y fl-fr), Maas et al. 7339 (COL, K, MO, NY, U); Berbice-Corantyne region, Potoko Cr., 24 Apr 1990 (Y fr), McDowell 2455 (NY).
SURINAME. Kwattaweg, W of Paramaribo, 21 Dec 1948 (6), BBS 223 (NY, U); Coppename R., nr. Kaaiman- stone, 7 Sep 1933 (9 fl-fr), Lanjouw 720 (U); Distr. Nickeri, Kabalebo Dam project area, 8 Sep 1980 (9 fl-fr), Lindeman & Girts-van Rijn et al. 288 (NY, RB, U); Wilhelmina Mtns.,
confluence of Lucie R. & Oost R., 12 Jul 1963 (2 fr), Ma- guire et al. 54150 (BG, NY, US); Corantijn R., 3 Aug 1935 (6), Rombouts 64 (U); Coppename R., nr. Heidoti, 24 Apr 1956 (6), Schulz (LBB) 7616 (F, U); 1 km NW of Voltzberg, 10 Feb 1965 (6), Schulz et al. (LBB) 10621 (U, US); Litanie R., Dec 1903 (6), Versteeg 418 (U); Oelemari R., 19 Mar 1963 (6), Wessels Boer 971 (U), (2 fl-fr), Wessels Boer 972 (U); confluence of Paloemeu R. & Tapanahoni R., 13 Apr 1963 (2 fl-fr), Wessels Boer 1256 (U).
FRENCH GUIANA. Comte R., Apr 1961 (2 fl-fr), Au- bre'ville 354 (P, U); Inini Basin, Saut Sonnelle, 23 Jan 1994 (6), Cremers 13269 (BG), (st), Cremers 13270 (BG); Ap- prouage R., Tortue Cr., 25 Jan 1967 (2), Oldeman 2321 (P, U, US); Maripasoula, 25 Jan 1984 (2 fl), Prevost 1468 (U); Belingui, S of Maroni, Beiman Cr., 21 Nov 1984 (2 fl-fr), Prevost 1693 (INPA, P, S, U), (6), Prevost 1700 (INPA, P, S, U).
ECUADOR. NAPO: Parque Nacional Yasuni, rd. Pom- peya Sur-Iro, km 38.4, 17 May 1994 (st), Jaramillo et al. 16518 (QCA), 16525 (QCA). SUCUMBiOS: Reserva Faun- istica Cuyabeno, Rio Cuyabeno, upstream from Laguna Grande, 11 Mar 1990 (2 fr), Balslev et al. 97162 (AAU, BG, QCA, QCNE); Rfo Cuyabeno, ca. 0'20'S, 75?55'W, 15 Feb 1980 (2 fl-fr), Berg et al. 1032 (BG, COL, QCA, U), 18 Feb 1980 (2 fl-fr), Berg et al. 1056 (AAU, BG, COL, GB, MO, NY, QCA, TUR, U), (6), Berg et al. 1057 (AAU, BG, COL, GB, MO, QCA, TUR, U).
PERU. LORETO: Nr. Iquitos, 6 Nov 1940 (2 fl-fr), As- plund 14350 (S), 14 Nov 1940 (6), Asplund 14499 (S); Prov. Maynas, Caballo Cocha, 24 Apr 1982 (2 fr), Ayala et al. 3310 (BG, MO, NY); Prov. Requena, Rio Ucayali, Huarmi Isla Cocha, 14 Dec 1987 (2 fr), Ayala et al. 5930 (F, MO); rd. Iquitos-Maynas, km 2, 4 Jul 1988 (6), Berg et al. 1570 (BG), (2 fr), Berg et al. 1575 (BG, COL, USM); Prov. Re- quena, Reserva Nacional Pacaya Samiria, 5 May 1983 (2 fr), Del Carpio 2087 (USM); Prov. Maynas, Rfo Yaguasy- acu, 9 Nov 1977 (2 fl), Gentry et al. 20514 (F, MO, NY, USM); Prov. Maynas, Rio Curaray, nr. Buena Vista, 16 Nov 1986 (2 fl), Kalliola et al. P5-4 (BG, USM) Prov. Maynas, Rio Amazonas, Isla Rondifia, 25 Mar 1977 (2 fr), Plowman et al. 6433 (F, GH, US, USM); Prov. Alta Amazonas, An- doas, 20 Nov 1980 (d), R. Va'squez et al. 776 (USM). MA- DRE DE DIos: Prov. Tambopata, 15 km ENE of Puerto Mal- donado, 1991 (2 fr), Fisher 179 (BG); Parque Nacional Manu, Rio Manu, Rio Cumerjali, 21 Oct 1986 (6), Foster et al. 11960 (BG, LPB, MOL, US, USM); Manu, Pakitza, 19 Jan 1987 (2 fl-fr), Nuiniez 6896 (BG); Prov. Tambopata, Las Piedras, Quebrada Gamitana, 26 Jan 1991 (2 fr), Ti- mand et al. 1406 (MO). UCAYALI: Prov. Coronel Portillo, Yarinacocha, Dec 1968 (2 fr), Ferreyra 17625 (US, USM); Prov. Coronel Portillo, Boquer6n del Padre Abad, 50 km E of Aguaytia, 9 Aug 1988 (2 fl), Gentry et al. 29531 (BG); Prov. Coronel Portillo, Lago Yarinacocha, 12 Sep 1980 (6), Maas et al. 4603 (BG, USM); Rio Ucayali, Masea, Nov 1923 (2 fl-fr), Tessmann 3307 (NY).
BRAZIL. AcRE: Rio Jurua, nr. Colonia Rodriguez Al- vez, 29 Sep-10 Oct 1986 (6), Campbell et al. 11161 (BG); Mun. Rio Branco, Lago Amapa, 3 Dec 1982 (2 fl-fr), L. Coelho et al. 1908 (INPA, NY); Mun. Bujari, Rio Branco,
SYSTEMATIC TREATMENT 107
Riozinho do Andira, 24 Mar 1995 (Y fl-fr), Pardo et al. 75 (BG, NY); Mun. Mancio Lima, Rio Moa, Parque Nacional da Serra do Divisor, 8 May 196 (Y fr), M. Silveira et al. 1310 (NY); Rio Jurua, Fortaleza, Nov 1901 (Y fl-fr), Ule 5938 (G, L, MG). AMAPA: Rio Jarn, Igarape Grande, ca. 1 km N of Arumanduba, 24 Dec 1961 (Y fr), Egler 45948 (BG, MG, NY, U, US). AMAZONAS: Rio Solimoes, 1-2 km E of Boca de Januaca, 2 Oct 1973 ( ? fr), Berg et al. P17586 (F, INPA, K, MG, MO, S, U, US); Parand do Autaz-Mirim, Lago do Caioe, 26 Aug 1973 (Y fr), Berg et al. P 19769 (C, EAP, F, INPA, K, MG, MO, NY, S, U, US); Mun. Novo Ayroa, Rio Negro, Arquipelago de Anavilhanas, 24 Jan 1992 (d), L. V Ferreira 109 (BG, K, NY); Manacapurui, 6 Mar 1904 (? fl-fr), Huber (MG) 4179 (BM, G, MG, U); Lower Rio Purus, 18 Mar 1904 (Y fl-fr), Huber (MG) 4195 (BM, G, MG, U); Mun. Humaita, nr. Livramento, on Rio Livra- mento, 12 Oct-6 Nov 1934 (d), Krukoff 6718 (A, BM, F, G, K, MICH, MO, NY, RB, S, U, US); Mun. Sao Paulo de Oliven,a, nr. Palmares, 11 Sep-26 Oct 1936 (d), Krukoff 8407 (A, BM, F, G, K, MICH, MO, NY, P, S, U, US); Rio Negro, Anavilhanas Is., 2?34'S, 60?19'W, 7 Aug 1991 (Y fr), Mori et al. 21959 (BG, NY, S); Mun. Maraa, Rio Japua, nr. liha Nova, 3 Dec 1982 (d), Plowman et al. 12160 (INPA, MG, NY, RB); nr. Tototobi, 27 Feb 1969 (Y fl-fr), Prance et al. 10289 (INPA, K, MG, NY, S, U, US); nr. Manaus, Rio Solimoes, Furo de Xiborena, 4 Apr 1974 (6), Prance et al. 20925 (GH, INPA, K, MG, MO, NY, P, S, U, US), (Y fl-fr), Prance et al. 20927 (INPA, K, MG, MO, NY, S, U, US), (d), Prance et al. 20929 (C, F, INPA, K, MG, MO, NY, P, U, US). PARA: Rio Trombetas, 1 km S of Cachoeira Porteira, 2 Jun 1974 (Y fl-fr), Campbell et al. P22441 (INPA, NY, U); Mun. Oriximina, Rio Trombetas, Minera,co Santa Pa- tricia, 7 Jul 1980 (Y fl), L. V Ferreira et al. 1341 (INPA, MG, NY, U, US); Mun. Tucuruf, Rio Tocantins, 1-5 km upriver from Represa Tucuruf, (Y fl-fr), Plowman et al. 9883 (F, GH, INPA, MG, MICH, MO, NY, U, US), 22 Mar 1980 (d), Plowman et al. 9886 (F, GH, MG, MO, NY, U, US); Rio Maicuru, Lageira, 19 Jul 1981 (Y fr), Strudwick et al. 3187 (INPA, MG, NY). ROND6NIA: Porto Velho, Rio Machado, 27 Nov 1977 (Y fl), Goulding 7 (INPA); Rio Ma- deira, Calama, Apr 1980 (st), Goulding Ila (MG); Rio Ma- chado, Jan 1981 (Y fr), Goulding 1252 (MG); Upper Rio Machado, nr. Tabajara, 24 Nov 1931 (d), Krukoff 1438 (A, BM, G, K, MICH, NY, P, S, U). RORAIMA: Ilha de Maraca, Mun. Alto Alegre, Rio Uraricuera, Furo Santa Rosa, 8 Jun 1986 (d), Hopkins et al. 872 (BG, INPA, NY); Ilha de Mar- aca, Furu Parand do Firmino, 3 Jul 1987 (Y fl-fr), Milliken et al. 388 (BG, E, K); Rio Uraricuera, between 63010' & 63?30'W, 13 Mar 1979 (d), Pires et al. 16963a (U), 16963b (NY); Rio Mucajai, 24 Mar 1971 (' fr), Prance etal. 11207 (INPA, K, MG, NY, S, U, US).
BOLIVIA. BENI: Prov. Yacuma, Rio Yacuma, Espiritu, 9 Apr 1981 (Y fr), Beck et al. 5292 (LPB), 28 Feb 1990 (Y fl-fr), Moraes R. et al. 1239 (BG, LPB, MO, USZ); Prov. Marban, Parque Isiboro-Secure, 5 Jun 1992 (st), Seidel et al. 6674 (LPB). PANDO: Prov. Manuripi, Rio Manuripi, Puerto Rico, 23 Jan 1983 (9 fr), Ferndndez C. et al. 8410 (G, NY). SANTA CRUZ: Prov. Velasco, Reserva Ecol6gica El Refugio,
Rfo Paragua, between Campamento El Refugio & Campa- mento Toledo, 7 Feb 1995 (9 fr), Guillen et al. 3170 (USZ).
In the upper Amazon basin, particularly in the northwestern part, the spikes of the fruiting pistillate inflorescence are short, usually to 10 cm long. In the Guianas, Venezuelan Guayana, and parts of Brazil, the fruiting spikes can be to 25 cm long, and the pedun- cles are usually longer as well. The species is com- monly found in periodically inundated areas along black-water rivers or in depressions with sandy and poorly drained soil away from white-water rivers (cf. Lamotte, 1992). It is also found in periodically in- undated white sand savannas in the Guianas. It is eco- logically clearly distinct from Cecropia membrana- cea, the other species of the genus that tolerates extended inundation.
Cecropia latiloba and C. utcubambana are mor- phologically very close and almost certainly closely related.
Vernacular names and use. Venezuela: simbra- potro (Apure); tokorodek (Pen6m, Bolivar); uaru (Warao, Delta Amacuro). Guyana: wanasoro. Suri- name: bospapaja. Peru: kabeari (Machiguenga, Madre de Dios); setico colorado (Ucayali). Brazil: imbauiba branca, imbauba da varzea (Amazonas); imbauba branca (Mato Grosso); imbaubarana (Rondonia); to- kori (Uaica'-Mucajai, Roraima).
In Madre de Dios, bark fibers are used to make arrow strings (D. W. Yu, pers. comm.).
26. Cecropia litoralis Snethlage, Notizbl. Bot. Gart. Berlin-Dahlem 8: 392. 1923. Type. Ecuador. "Coastal Plain," without locality, Rimbach 15 (ho- lotype: B, destroyed, duplicates not found; pho- tographs in F, G, GH, MICH, MO), herewith re- placed by: Ecuador. El Oro: Between Pasaje & Uzhcurrumi, 2 Feb 1981 (9), Berg 1238 (neotype: QCA; isoneotypes: BG, COL, U). Fig. 30
Tree, to 10(-25) m tall, the trunk with prominent stipular scars. Leafy twigs 1-3 cm thick, green with conspicuous lenticels, densely hispidulous (with curved to straight hairs). Lamina chartaceous to sub- coriaceous, ca. 40 X 40 cm to 75 X 75 cm, the seg- ments 7-9, the free parts of upper segments elliptic to oblong to (sub)obovate, the incisions down to 6/ 10-8/10; apices obtuse to short-acuminate; upper sur- face scabridulous, sparsely hispidulous and (initially) with sparse arachnoid indumentum; lower surface (minutely) puberulous to subtomentellous (or partly to hirtellous), with arachnoid indumentum in the areoles and on the smaller veins, extending to the
108 FLORA NEOTROPICA
1014~
FIG. 30. Cecropia litoralis. 1. Leaf. 2. Apex of lamina and venation. 3. Stipules and base of petiole with trichilium (Berg et al. 1043). 4. Leafy twig with staminate inflorescences in various stages of development (with spathes and at anthesis) and bases of petioles with trichilia (Balslev et al. 84740). 5. Staminate flower and stamen (Balslev et al. 97161). 6. Pistillate inflorescence with spathe (Dodson 7334). 7. Pistillate inflorescence at anthesis. 8. Pistillate inflorescence after anthesis and base of petiole with trichilium. 9. Pistillate flower. 10. Fruit (Berg et al. 1046).
SYSTEMATIC TREATMENT 109
main veins; lateral veins in the free part of the mid- segment 13-19 pairs, marginally loop-connected, mostly branched; petiole 25-75 cm long, puberulous to hirtellous or also with arachnoid indumentum; tri- chilia fused, the brown indumentum intermixed with short white hairs; stipules 8-17 cm long, yellowish green to reddish, when dry usually chartaceous to membranaceous, puberulous to strig(ill)ose and with sparse arachnoid indumentum outside, (rather) sparsely sericeous inside. Staminate inflorescences solitary or in pairs, the peduncle erect to deflexed, the spikes pendulous; peduncle 6-16 cm long, puberu- lous to hispidulous and/or (initially) with brown plur- icellular hairs and sparse arachnoid indumentum; spathe 7-14 cm long, white to/or pinkish or yellow- ish, when dry usually chartaceous to membranaceous, sparsely puberulous or partly to strig(ill)ose to sub- hirtellous and with very sparse to rather dense arach- noid indumentum outside, glabrous inside; spikes 6- 15, 5-12 X 0.2-0.3 cm, with stipes 0.2-1 cm long and hirtellous to puberulous; rachis glabrous. Stami- nate flowers: perianth tubular, ca. 1 mm long, gla- brous, the apex plane; filaments slightly swollen; an- thers ca. 0.5 mm long, (short-)appendiculate, at anthesis remaining attached to the narrowed apex of the filament (but after anthesis detached). Pistillate inflorescences solitary or in pairs, erect to deflexed, becoming pendulous in fruit; peduncle 9-16 cm long, sparsely puberulous to hirtellous below the spathe and with sparse to dense arachnoid indumentum, some- times subglabrous; spathe 8-19 cm long, the color, texture, and indumentum as in the staminate inflores- cence; spikes 4(-5), 7-16 X 0.4-0.6, to 28(-37) X 1-1.5 cm in fruit, with stipes 0.3-1 cm long and pu- berulous to glabrous; rachis hairy. Pistillate;flowers: perianth ca. 1.5 mm long, with arachnoid indumen- tum on and below the apex, also in the style channel inside, the apex plane; stigma peltate. Fruit oblon- goid, 1.8-2 mm long, tuberculate.
Distribution (Fig. 8.3). Coastal regions of Ecua- dor and of Narinlo (Colombia); very common in wet to relatively dry forest, coastal swamps, and second- ary growth; at elevations to 1500 m. Also occurring in the upper Amazon basin, probably discontinuously, known from Colombia (Caqueta' and Putumayo), Ec- uador (Sucumbios), and Peru (Loreto), along (black- water) rivers in places that are not inundated or inundated only during short periods, often in associ- ation with Cecropia latiloba.
Representative specimens examined. COLOMBIA. CAQUETA: Rio Caqueta, below mouth of Rio Orteguaza, Solano, 8 km SE of Tres Esquinas, 8 Mar 1945 (Y fl), Little et al. 9684 (COL). NARINO: Mun. Tumaco, rd. to Pasto,
Vrda. El Tigre, 23 Feb 1995 (9 fl-fr), Franco et al. 5544 (BG, COL, HUA, US), (6), Franco et al. 5545 (BG, COL, HUA, US), (9 fl-fr), Franco et al. 5546 (BG, COL, HUA, US). PUTUMAYO: Mun. Puerto Asfs, rd. to Kanakas, 16 Feb 1995 (9 fl-fr), Franco et al. 4681 (BG, COL, HUA, LP, MO, NY, US).
ECUADOR. EL ORO: Nr. Piedras, 23 Nov-16 Dec 1978 (9 fl-fl), Albert de Escobar 984 (QCA, TEX, U); be- tween Pasaje & Uzhcurrumi, 2 Feb 1981 (6), Berg 1237 (BG, COL, QCA); 30 km SW of Santa Rosa, rd. to Pinias, 6 Oct 1979 (9 fl), Dodson et al. 8869 (F, MO); Puyango, 29 Dec 1997 (9 fl-fr), P. Lozano et al. 902 (BG, LOJA), (d), P Lozano et al. 903 (BG, LOJA); nr. Portovelo, 6-15 Oct 1918 (6), Rose 23411 (GH, NY, US). ESMERALDAS: Can- t6n San Lorenzo, La Tola, 21 Nov 1993 (6), Alvarez 740 (QCNE); nr. Borb6n, 6 Aug 1967 (9), Jditiva et al. 2160 (NY, S); Rio Santiago, between Borb6n & Palma, 10 Aug 1967 (9 fl-fr), Jdtiva et al. 2223 (AAU, NY), (6) Jdtiva et al. 2224 (AAU, MO, NY); nr. Esmeraldas, 29 Sep 1965 (9 fl-fr), Little 21176 (NY, QAME, QCNE, US). GUAYAS:
Cerro Azul, W of Guayaquil, 9 Feb 1955 (9 fr), Asplund 15377 (S); nr. Naranjito, 6-7 Jun 1945 (9 fl-fr), Camp E.3634 (BG, NY, S); Cant6n Naranjal, Taura, Reserva Ecol- 6gica Manglares-Churute, 22 Feb 1991 (6), Ceron et al. 13470 (QCNE); rd. Guayaquil-Salinas, km 15, 2 Nov 1995 (9 fl-fr), Clark et al. 1530 (BG, QCNE). LOJA: Rd. Ala- mor-Arenillas, nr. bridge over Rio Puyango, 30 Dec 1997 (d6), P. Lozano et al. 912 (BG, LOJA). Guayachuma, rd. Loja-Machala, ca. 1400 m, 23 Sep 1967 (9 fl), Sparre 18866 (S). Los Rios: Rio Palenque Biological Station, rd. Quevedo-Santo Domingo de los Colorados, km 56, 23 Jul 1975 (6), Dodson 5880 (QCA, MO, US), 1 Dec 1978 (9 fr), Dodson 7334 (BG, QCNE); Cant6n Vinces, rd. Quev- edo-Palenque, km 70, Jauneche forest, 26 Mar 1980 (9 fr), Dodson et al. 9922 (BG, F, MO); 18 km S of Empalme, nr. Guayas border, 5 Feb 1974 (9 fl-fr), Gentry 9673 (MO); between Babahoyo & Montalvo, 18 Feb 1967 (6), Sparre 14534 (S). NAPO: Cant6n Pompeya, La Joya de los Sachas, Isla de Pompeya, 6 Ec 1992 (9 fl-fr), Gudifno et al. 2021 (BG, QCNE); Laguna de Yuturi, 28 Feb 1990 (9 fl-fr), Jar- amillo et al. 11475 (GB, NY, QCA). PICHINCHA: Santo Do- mingo de los Colorados, 22 Aug 1986 (d), Vivar et al. 2844 (LOJA). SUCUMBiOS: Reserva Faunistica Cuyabeno, Rio Cuyabeno, between Laguna Grande & 5 km upstream, 11 Mar 1990 (6), Balslev et al. 97160 (AAU, BG, QCA, QCNE); Rio Cuyabeno, ca. 0?10'S, 75?55'W, 17 Feb 1980 (6), Berg et al. 1043 (BG, COL, GB, MO, QCA, TUR, U), (9 fl-fr), Berg et al. 1046 (AAU, BG, COL, GB, MO, QCA, TUR, U).
PERU. LORETO: Nr. Iquitos, 30 Nov 1940 (9 fl-fr), Asplund 14810 (S); Prov. Requena, nr. Jenaro Herrera, Co- cha Supay, Arboretum de Braga, 18 Jul 1988 (9 fl-fr), Berg et al. 1586 (BG, USM), 16 Dec 1987 (9 fr), Lamotte 375 (BG); Prov. Alto Amazonas, Andoas, Rio Pastaza, 10 Nov 1980 (6), R. Vdsquez et al. 776 (BG).
Cecropia litoralis is closely related to C. engleri- ana, which is sympatric with C. litoralis in the north- western part of the Amazon basin. Cecropia litoralis
110 FLORA NEOTROPICA
can be easily distinguished from C. engleriana by the smaller number of lateral veins, to ca. 15 pairs, but in the other features the two taxa cannot be distin- guished. One could argue that these two taxa are sub- species rather than species (see "Ecology," discussion of the C. peltata-group). It is on phytogeographical and ecological considerations that they are kept dis- tinct at the species level for the time being. In the coastal region of Ecuador and the adjacent part of Colombia, C. litoralis is a ? weedy species in vari- ous habitats but not clearly associated with riverine habitats. However, in the Amazon basin it is common along Rio Cuyabeno, a black-water tributary of Rio Aguarico, and is there associated with C. latiloba. The species has also been collected in Peru (Loreto) in a C. latiloba habitat. The association with C. lati- loba indicates ecological preference different from that of C. engleriana, which is an upland species, or if it is found in riverine habitats, then rather in asso- ciation with C. membranacea.
27. Cecropia longipes Pittier, Contr. U.S. Natl. Herb. 18: 227. 1917; Croat, Fl. Barro Colorado Island 347, t. 195. 1978. Type. Panama. Panama': Nr. Ta- bernilla, 6 Jul 1911 (d), Pittier 3823 (holotype: US).
Tree, to 18 m tall. Leafy twigs 1.5-4 cm thick, green, hispid(ulous) with uncinate hairs to hirtellous. Lamina chartaceous, ca. 20 X 20 cm to 70 X 70 cm, the segments 7-10, the free parts of the upper seg- ments ovate to elliptic, the incisions in the upper part of the lamina down to 4/10-6/10; apices rounded to short-acuminate; upper surface ? scabrous, puberu- lous to hispidulous; lower surface puberulous to sub- tomentose on the veins, with arachnoid indumentum in the areoles or also on the reticulum; lateral veins in the free part of the midsegment 5-8 pairs, margin- ally loop-connected, often branched; petiole ca. 20- 60 cm long, puberulous to hirtellous; trichilia fused, the brown indumentum intermixed with dense long whitish hairs; stipules 4-9 cm long, color unknown, subhirsute to hirtellous or to subsericeous outside, sparsely hairy inside. Staminate inflorescences in pairs, at least the spikes pendulous (?); peduncle 5- 15 cm long, hirtellous to subhirsute; spathe 12-17 cm long, color unknown, puberulous to hirtellous outside, glabrous inside; spikes ca. 30-80, + crinkled, 4-15 x 0.1-0.2 cm, with stipes ca. 0.4-1 cm long and hairy; rachis hairy. Staminate flowers: perianth tu- bular, ca. 0.5-0.7 mm long, with arachnoid indumen- tum below the apex, the apex almost plane; stamens not seen. Pistillate inflorescences in pairs, pendulous;
peduncle ca. 30-90 cm long, hirtellous; spathe 7-12 cm long, the color unknown, the indumentum as in the staminate inflorescence; spikes 6-12, often ? crinkled, 4-10 X 0.6-0.8 cm, to 8 X 1.1 cm in fruit, with stipes 0.3-0.8 cm long and puberulous to hirtel- lous; rachis hairy. Pistillateflowers: perianth ca. 1.5- 2 mm long, with arachnoid indumentum below the apex outside, also in the style channel inside, the apex + convex, muriculate; style short, muriculate; stigma comose. Fruit subobovoid to oblongoid, ca. 2 mm long, smooth, pale brown.
Distribution (Fig. 8.3). Eastern Panama and northwestern Colombia, mostly in secondary growth, at low elevations.
Representative specimens examined. PANAMA. COLON: S of Fort Sherman, 27 Jul 1955 (9 fl-fr), L M. Johnston 1503 (A, MO); nr. Madden Dam, 17 Jul 1938 (9 fr), Woodson et al. 1312 (A, MO, NY). DARIEN: Cruce de Mono, 5 Nov 1989 (st), Fisher 53 (BG); Rio Tuquesa, lower Tuquesa Mining camp, 5 Jul 1975 (9 fr), Mori 6998 (BG, MO). PANAMA: El Llano-Carti rd., nr. El Llano, 1 Sep 1977 (9 fl-fr), Berg 410 (BG); 1.5 mi beyond intersection of USARSO chemical training camp area, 15 Jun 1967 (9 fl- fr), Correa et al. 27 (COL, MO, PMA, SCZ); Barro Colo- rado Is., 30 Apr 1970 (9 fl), Croat 10115 (COL, MO, SCZ), 23 Jun 1971 (6), Croat 15083 (MO, NY, SCZ); Capitana, Chepo, 9 Jun 1971 (6), Lao 120 (MO, PMA, SCZ), (9 fl- fr), Lao 121 (H, MO, PMA); nr. El Real, 22 May 1969 (9 fl), Lazor et al. 3440 (MO); Curundu, 20 Jul 1985 (9 fl-fr), Nevers et al. 5994 (BG, MO); 2 mi E of Santa Fe, 18 Jul 1966 (9 fr), Tyson et al. 4854 (MO, SCZ).
COLOMBIA. ANTIOQUIA: Uraba-Chigorod6- Malag6n, Cafno Malag6n, 22 Mar 1986 (9 fl-fr), Renteria et al. 4735 (JAUM). BOLiVAR: Mun. Turbaco, 8 Jun 1982 (9 fl), Cuadros V 1374 (COL, MO, US); Rfo San Jorge, 4 Jul 1948 (9 fl), Romero-Castaneda 1137 (COL, F, US). CHocO: Mun. Acandi: Vrda. Coquita, El Paramo, Quebrada Zardi, 26 May 1989 (9 fl-fr), Betancur et al. 1255 (HUA, MO); Mun. Turbo, rd. Tap6n del Dari6n, between Rio Le6n & Lomas Aisladas, km 37, 28 May 1984 (9 fl-fr), Brand 1201 (JAUM, MO); Mun. Riosucio, Cgto. La Honda, Vrda. La Balsa, Canal La Balsa, 15 Apr 1987 (6), Echavarria et al. 149 (JAUM); Mun. Riosucio, rd. Peye-Sautata, 5 Jun 1976 (9 fl), Forero et al. 1890 (COL, MO); Mun. Acandi, Cgto. Ungufa, Rio Cuti, 27 Jul 1957 (9 fl), Romero- Castaneda 6453 (COL); Parque Nacional Natural Los Ka- tios, Sautana, 3 May 1982 (6), Zuluaga R. 194 (COL).
This species is quite distinct because of long pe- duncles of the pistillate inflorescences. It does not show clear affinities to other species from the same region.
lombia. Putumayo: Rio Putumayo, between Rio Giiamues & Puerto Asis, 21 Dec 1940 (Y), Cua- trecasas 11243 (holotype: COL; isotype: F).
Fig. 31
Tree, to 25(-40) m tall. Leafy twigs 2-5 cm thick, green, hispidulous or hirtellous to hirsute. Lamina subcoriaceous to chartaceous, ca. 50 X 50 cm to 85 X 85 cm (to 120 X 120 cm), the segments 7-8, the free parts of the upper segments ovate, the incisions down to 3/10-5/10; apices rounded; upper surface (almost) smooth, almost glabrous, with sparse brown pluricellular hairs and sparse long, unicellular hairs; lower surface densely to rather sparsely hirtellous or subhirsute on the main veins, with arachnoid indumentum in the areoles and on the smaller veins; lateral veins in the free part of the midsegment 6-15 pairs, submarginally loop-connected, many of them branched; petiole relatively short, in adult specimens ca. half the diameter of the lamina, 25-45(-60) cm long, densely hirsute, denser toward the apex, often also with sparse to dense arachnoid indumentum; tri- chilia separate or occasionally fused, the brown in- dumentum intermixed with dense short or long white hairs; stipules ca. 15-45 cm long, caducous, yellow- ish to dark red to red-brown or pale green, with sparse unicellular hairs at the base or confined to the margin or largely ? densely hirsute to subvillous outside, glabrous inside; terminal buds usually ? curved. Sta- minate inflorescences in pairs, the peduncle erect, the spikes erect to pendulous; peduncle 6-9 cm long, sparsely to densely hirtellous to subhirsute; spathe 8- 13(-16) cm long, dark red to red-brown to orange or to pink, or pale green, puberulous or hirtellous to sub- hirsute outside, glabrous inside; spikes ca. (20-)30- 50, 6-11 X 0.2-0.3 cm, with stipes 0.5-1.5 cm long and tomentose; rachis hairy. Staminate flowers: per- ianth deeply 2-parted, 1.2-1.3 mm long, glabrous, the apex convex; filaments swollen; anthers 0.6-0.7 mm long, without appendages, remaining attached at an- thesis. Pistillate inflorescences in pairs, sometimes solitary, initially erect, pendulous in fruit; peduncle 7-8(-17) cm long, subhispidulous to hirtellous, more densely so at the apex; spathe 6-11(-14) cm long, the color and indumentum as in the staminate inflores- cence; spikes 4(-7), ca. 8-12 x ca. 0.5 cm, to 22 X
1.5 cm in fruit, sessile or with stipes to 0.5 cm long and tomentose; rachis glabrous. Pistillate flowers: perianth 1.3-1.5 mm long, with arachnoid indumen- tum below the apex outside, absent inside, the apex convex to subconical, sometimes 2-lobed, granulate or muriculate and with a few short bristles near the aperture, the aperture slit-shaped; style rather long,
almost straight; stigma penicillate. Fruit oblongoid, ca. 2 mm long, smooth.
Distribution (Fig. 8.2). From Colombia (Meta) through Amazonian Ecuador to Peru (Amazonas), in non-inundated or occasionally or briefly inundated places, mostly in secondary growth, at elevations to ca. 1200(-1500?) m. It is the most common species in Amazonian Ecuador.
Representative specimens examined. COLOMBIA. CAQUETA: Mun. Florencia, rd. to Gabinete, 18 Oct 1993 (Y fl), Franco et al. 4508 (BG, COL). META: Mun. Villavicen- cio, rd. to Guayabetal, 900-1000 m, 13 Feb 1995 (Y fl-fr), Franco et al. 4676 (BG, COL). PUTUMAYO: Mun. Puerto Asis, rd. to Kanakas, 16 Feb 1995 (Y fl-fr), Franco et al. 4680 (BG, COL); rd. Mocoa-Pasto, nr. Mocoa, 18 Feb 1995 (Y fl-fr), Franco et al. 4686 (BG, COL, HUA, US); rd. Mo- coa-Puerto Asis, 3 May 1994 (? fl-fr), Franco et al. 5518 (BG, COL), (6), Franco et al. 5519 (BG, COL).
ECUADOR. MORONA-SANTIAGO: Rio Blanco, be- tween Macas & Sucua, 26 Jan 1981 (6), Berg 1217 (BG, COL, QCA, U); rd. Zamora-Gualaquiza, Estaci6n Exp. El Padmi, ca. 5 km N of Los Encuentros, 4 Jan 1991 (Y fl-fr), Berg et al. 1659 (BG, LOJA, QCA); Cord. de Cutucu, 5-10 km E of Logronlo, 1200-1500 m, 7-9 Oct 1975 (? fl-fr), Little et al. 669 (COL, LOJA, Q, QAME, QCNE); 11.7 km SW of Taisha, 11 Sep 1976 (Y fl-fr), Ortega U. 148 (Q). NAPO: Rio Payamino, ca. 10 km NW Coca, 27 Feb 1980 (Y fl), Berg et al. 1091 (AAU, BG, COL, MO, QCA, U); Reserva Biol6gica Jatun Sacha, 8 km E of Misahualli, 4 Sep 1987 (6), Cer6n etal. 2149 (BG, QAME, QCNE); rd. Tena- Rfo Pano, km 6-12, 5 Aug 1984 (Y fl-fr), Dodson et al. 15011 (QCNE); San Jos6 de Payamino, 40 km W of Coca, Rio Tutapishu, 5 Nov 1982 (Y fl-fr), Irvine 210 (BG, F, QCA, QCNE); Archidona, 1865 (6), Isern SPB (GB, MA); rd. Puyo-Puerto Napo, km 18, 8 Apr 1969 (Y fl-fr), Lugo 1065 (BG, GB); Armenia Vieja, Rio Napo, 12 km SW of Coca, 12 Jan 1973 (6), Lugo 2650 (BG, GB); Cafi6n de los Monos, 15 km N of Coca, Rio Coca, 18-20 Apr 1985 (Y fl-fr), Neill 6337 (BG, MO, QAME, QCNE); Afiangu, Parque Nacional Yasuni, 30 Jun-9 Jul 1982 (st), SEF 10108 (AAU, BG, QCA). PASTAZA: Between Mera & Moravia, ca. 1000 m, 16 Dec 1955 (Y fl-fr), Asplund 18867 (S); Mera, ca. 1100 m, 21 Dec 1955 (6), Asplund 18944 (S); Rio Bo- bonaza, Cuansha-Nalpi, between Cabeceras & Canelos, 9 Feb 1971 (Y fl-fr), Lugo 1450 (BG, GB); Rio Bobonaza, at Rio Tinguiza, SE of Pacayacu, 15 Mar 1971 (6), Lugo 1685 (BG, GB); Rio Bobonaza, Puerto Ubilla, nr. Pacayacu, 17 Mar 1971 (d), Lugo 1705 (BG, GB). SUCUMBfOS: Rio Cuy- abeno, ca. 0?20'S, 75?55'W, 20 Feb 1980 (Y fl-fr), Berg et al. 1072 (AAU, BG, COL, GB, K, MO, NY, QCA, TUR, U), 1073 (BG, COL, QCA); Rio Aguarico, ca. 0?20'S, 75?05'W, 21 Feb 1980 (6), Berg et al. 1074 (AAU, BG, COL, GB, K, MO, NY, QCA, TUR, U, WIS); Rio Aguarico, nr. confluence with Rio Pavayacu, Sep 1981 (Y fl-fr), Bravo et al. 291 (QCA); Rio Aguarico, Dureno, 22 Feb 1980 (Y fl-fr), Holm-Nielsen et al. 21754 (AAU, MO). ZAMORA-
CHINCHIPE: Rio Nangaritza, nr. Nangaritza, 12 Dec 1990
112 FLORA NEOTROPICA
fX;~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~Vv
5z~ ~~~~~~~~~~~~~~~~~/
FIG. 31. Cecropia marginalis. 1. Leaf, reduced. 2. Apex of lamina and venation (Berg et al. 1072). 3. Stipules (Dodson et al. 15011). 4. Pair of staminate inflorescences with spathes and base of petiole with trichilium. 5. Pair of staminate
inflorescences, one with spathe, the other at anthesis, and base of petiole with trichilium (Berg et al. 1074). 6. Staminate flower and stamen (Franco et al. 4508). 7. Pistillate inflorescence with spathe and base of petiole with trichilium. 8. Pistillate inflorescence at anthesis and base of petiole with trichilium. 9. Pistillate inflorescence after anthesis and base of petiole with trichilium. 10. Pistillate flower. 11. Fruit with remnant of style (Berg et al. 1072). 12. Trichilia (Berg et al. 1072).
SYSTEMATIC TREATMENT 113
(d), Neill et al. 9703 (AAU, BG, QCNE); Cant6n Nanga- titza, Pachicutza, 18 Oct 1991 ( Y fl-fr), Palacios et al. 8293 (BG, QCNE, USZ).
PERU. AMAZONAS: Quebrada Aintami, 1 May 1973 (? fl-fr), Kayap 680 (GH); Quebrada Huampami, 5 Jul 1974 (Y fl), Kayap 1052 (BG); Rfo Santiago, nr. La Poza, 12 Nov 1979 (st), Tunqui 16 (MO); Rfo Santiago, nr. Caterpiza, 14 Dec 1979 (Y fl), Tunqui 350 (MO); Prov. Bagua, Dtto. Im- aza, Rio Comaina, Kusu-kubaim, 17 Aug 1994 (d), R. Vds- quez et al. 18822 (MO).
The species is quite uniform except for the length of the hairs. It has longer hairs and often also denser indumentum on the leafy twigs, stipules, spathes, pet- iole, lower surface of the lamina, and in the trichilia in Colombia than in collections made southward. The trichilia are occasionally fused (as in Franco et al. 5519). Cecropia marginalis shows affinities to C. stri- gosa.
Vernacular names. Ecuador: suu or tsu (Shuar, Morona-Santiago); tselan dundu (Quichua, Napo). Peru: sui, suu, tsake kumpari (Amazonas).
29. Cecropia maxima Snethlage, Notizbl. Bot. Gart. Berlin-Dahlem 8: 360. 1923. Type. Ecuador. Pi- chincha: Poluahua (= Pululahua?), ( Y + d ?), So- diro 2a and 3 (syntypes: B, destroyed, duplicates not traced; photographs in F, G, GH, MO), herewith replaced by: Ecuador. Pichincha: Rd. Quito-Chiriboga-Empalme, km 50, 2000-2100 m, 21 Jul 1987 (Y), Zak et al. 2198 (neotype: QCA; isoneotypes: BG, G, GB, GH, K, L, MO, NY, TEX, US). Fig. 32
Tree, to 30 m tall. Leafy twigs 4-9 cm thick, dark brown with pale brown to orange lenticels, sparsely puberulous to hirtellous and with rather sparse arach- noid indumentum and brown pluricellular hairs. Lam- ina coriaceous, ca. 40 X 40 cm to 90 X 90 cm, the segments 10-15, the free parts of the upper segments subobovate to (ob)lanceolate, the incisions down to 5/10-8/10; apices rounded; upper surface (almost) smooth, sparsely hirtellous on the main veins, on the whole surface dense or sometimes (very) sparse arachnoid indumentum; lower surface (initially) with brown pluricellular hairs, with arachnoid indumen- tum (almost) confined to the areoles, but (initially) also present on the main veins; lateral veins in the free part of the midsegment 15-27 pairs, submargin- ally (to marginally) loop-connected, most of them un- branched; reticulum with rather thick veinlets; petiole ca. 50-90 cm long, red to red-brown, sparsely puber- ulous and with brown pluricellular hairs, moreover, with (rather) dense arachnoid indumentum; trichilia
fused, the brown indumentum intermixed with dense short whitish hairs; stipules ca. 20-40 cm long, dull dark red, sparsely to densely hirtellous puberulous and with dark brown pluricellular hairs or also ? sparse arachnoid indumentum outside, densely (sub)sericeous (or glabrous) inside. Staminate inflo- rescences often solitary, the erect or the spikes ? spreading; peduncle 6.5-12 cm long, (sub)glabrous; spathe 10-15 cm long, dull red to purplish, hirtellous and with brown pluricellular hairs or also arachnoid indumentum outside, sparsely hairy or glabrous in- side; spikes 4-6(-8), 8-12.5 X 0.6-1 cm, (sub)- sessile; rachis with stiff hairs and crinkled (arachnoid) hairs. Staminate flowers sessile or shortly pedicellate; perianth tubular, ca. 2-3 mm long, puberulous below the apex, the apex plane to slightly convex with short hairs near the aperture; filaments flat; anthers ca. 1 mm long, appendiculate, detached at anthesis (?). Pis- tillate inflorescences solitary, erect, or the peduncle deflexed in fruit, subtended by caducous (or subper- sistent) bracts with dense arachnoid indumentum and to 8 cm long; peduncle 3-15 cm long, sparsely hir- tellous and with brown pluricellular hairs and also arachnoid indumentum; spathe ca. 10-20 cm long, the color and indumentum as in the staminate inflores- cence; spikes (l-)2-3, 12-18 X ca. 1.5 cm, to 48 X 2.8 cm in fruit, (sub)sessile; rachis glabrous. Pistillate flowers: perianth ca. 3-4 mm long, with arachnoid hairs below the apex, absent inside, the apex slightly convex, muriculate to puberulous around the aperture; style long, ? S-shaped, minutely puberulous; stigma comose. Fruit oblongoid, 2-2.5 mm long, smooth, pale brown.
Distribution (Fig. 10.4). From western Ecuador (Cotopaxi) to Colombia (Narifno), in montane forest; at 1500-2400 m, or in Narifio also at 900-1200 m (in vegetation with the physiognomy of montane forest).
Specimens examined. COLOMBIA: NARINO: Mun. Barbacoas, Cgto. Altaquer, rd. El Barro-Junfn, El Mirador, 1150 m, 12 Dec 1993 (Y fl-fr), Franco et al. 5175 (BG); Mun. Barbacoas, rd. Junfn-El Diviso, 900 m (!), 21 Feb 1995 (Y fr), Franco et al. 4694 (BG, COL, HA, US); rd Mun. Mallama, Quebrada La Chorrera, 2350 m, 27 Dec 1993 (d), Ramfrez P et al. 6061 (COL).
ECUADOR. COTOPAXI: Rd. Pilal6-Quevedo, between Pilal6 & Macuchi, 1500 m, 29 Jul 1980 (st), Holm-Nielsen et al. 24706 (AAU, BG). IMBABURA: Cotacachi, above Apuela, 2000 m, 24 Apr 1992 (d), Palacios et al. 10036 (MO, QCNE). PICHINCHA: Between San Juan & Chiriboga, ca. 2250 m, 3 May 1955 (d), Asplund 16167 (S); Guarumal, ca. 2100 m, 20 Jan 1956 (d), Asplund 19039 (S); rd. Quito- San Juan-Chiriboga-Empalme, ca. 7 km before Chiriboga, ca. 2300 m, 14 Dec 1990 (Y fl-fr), Berg et al. 1645 (BG, QCA); Calacali, Reserva Geobotanica del Pululahua, 1600
114 FLORA NEOTROPICA
Q,~~~~~~~,, Ip I
FIG. 32. Cecropia maxima. 1. Leaf, reduced. 2. Apex of lamina and venation (Ervik 36971). 3. Stipules and young leaf (Franco et al. 4694). 4. Staminate inflorescence with spathe. 5. Staminate inflorescence and base of petiole with trichilium (Palacios et al. 10036). 6. Staminate flower. 7. Stamen (Ervik 36871). 8. Pistillate inflorescence with spathe and base of petiole with trichilium (Cero'n et al. 4776). 9. Pistillate inflorescence after anthesis. 10. Pistillate flower (Berg et al. 1645). 11. Fruit (Franco et al. 4694).
SYSTEMATIC TREATMENT 115
m, 7 Sep 1988 (? fl-fr), Cer6n et al. 4776 (BG, MO, QCNE); rd. Nano-Mindo, Los Cedros, 2000 m, 27 Aug 1991 (d), Ervik 36871 (BG, QCNE); rd. Quito-Tandapi- Santo Domingo de los Colorados, 9.5 km E of Tandapi, 1900 m, 28 Sep 1979 (? fl-fr), Gentry et al. 26662 (AAU, BG, MO).
This white-leaved species is conspicuous in mon- tane forest on the western slopes of the Andes in Ec- uador and adjacent Colombia. However, in Nariiio (Colombia), the species also occurs at distinctly lower elevations (900-1200 m) in forest showing physiognomic similarities to montane forest. At these low elevations the trees lack the typically white upper surface of the lamina as found at higher ele- vations. The trees are usually inhabited by ants. There is a clear discrepancy between the present de- scription of the staminate flowers and that by Sneth- lage (1923), who stated that the perianth is 3-4 mm long and anther 2-3 mm long. Because of these fea- tures, Cecropia maxima was included in sect. Tomen- tosae. The differences in the descriptions cannot be explained.
30. Cecropia megastachya Cuatrecasas, Revista Acad. Colomb. Ci. Exact. 6(22/23): 284. 1945. Type. Colombia. Valle: Piedra de Moler, 1000 m, 19 Jul 1943 (d + Y), Cuatrecasas 14910 (sheet with pistillate inflorescence, stipules, and leaf here designated as lectotype: COL; isolectotype: COL).
Tree, to 17 m tall, often unbranched. Leafy twigs 5-8 cm thick, pale green with conspicuous white to pinkish lenticels, hirtellous to hirsute to subsetose. Lamina coriaceous, ca. 50 X 50 cm to 140 X 140 cm (to 170 x 170 cm), the segments 10-12, the free parts of the upper segments oblanceolate, the incisions down to 8/10-9/10 or to 1-2 cm from the petiole; apices acute to subacuminate; upper surface smooth with rather sparse arachnoid indumentum or (sub)- glabrous; lower surface sparsely minutely puberulous on the main veins largely with brown pluricellular hairs), with (very) sparse to dense arachnoid indu- mentum in the areoles and on the smaller veins, ex- tending to the main veins or not; lateral veins in the free part of the midsegment 20-30 pairs, submargin- ally loop-connected, unbranched; reticulum veinlets thick; petiole 70-120 cm long, with ? sparse arach- noid indumentum and (especially at the basal and upper part) hirsute to subsetose or setose (with irri- tating hairs); trichilia fused, the brown indumentum intermixed with sparse bristles; stipules ca. 20-70 cm long, partly pinkish to red, largely whitish, (sub)- setose and with dense arachnoid indumentum outside,
(sub)glabrous or subsericeous on the lower part in- side. Staminate inflorescences solitary or in pairs, sometimes subtended by caducous bracts, to 1 cm long, the peduncle deflexed and the spikes pendulous; peduncle 10-18 cm long, green, (sub)setose; spathe 28-45 cm long, white to pinkish, hirsute to (sub)setose and with dense arachnoid indumentum outside, glabrous inside; spikes 12-32, 25-36 X ca. 0.6-1 cm, with stipes 0.2-1 cm long and subsetose or with arachnoid indumentum; rachis with short stiff hairs and arachnoid indumentum. Staminate flowers with pedicels to 1.2 mm long, these hispidulous to puberulous or glabrous; perianth tubular, 1.5-2 mm long, puberulous to hispidulous below the apex, the apex plane, hispidulous, mainly along the margins of the aperture; filaments flat; anthers 0.6-0.7 mm long, appendiculate, detached at anthesis, reattached to the margins of the aperture by the appendages. Pistillate inflorescences solitary, deflexed to pendulous; pedun- cle green, 8-18 cm long; spathe ca. 30-40 cm long, the color and indumentum as in the staminate inflo- rescence; spikes 2 or 3, ca. 25-40 X ca. 2 cm, to 60 X 3 cm in fruit, (sub)sessile; rachis hairy only with stiff short hairs. Pistillate flowers: perianth 3.5-4.5 mm long, with arachnoid indumentum below the apex outside, also in the style channel inside, the apex ? convex, punctate to muriculate; style long, straight, minutely hairy; stigma comose. Fruit oblongoid to subovoid, ca. 3 mm long, finely tuberculate, pale- brown.
Distribution (Fig. 18.2). Colombia, on the west- ern slopes of the western Cordillera, from Antioquia to Valle, in forest, at ca. 400-1300 m.
Specimens examined. COLOMBIA. ANTIOQUIA: Mun. Frontino, Cgto. Nutibara, rd. Nutibara-La Blanquita, Murri region, 1250 m, 4 Mar 1995 (Y fl-fr), Franco et al. 5555 (BG, COL, HUA, US), (d), Franco et al. 5556 (BG, COL, HUA, US); Mun. Frontino, Cgto. Nutibara, valley of Rio Cuevas, 1000 m, 13 Apr 1987 (Y fl-fr), D. Sdnchez S. et al. 1125 (MEDEL). CHOc6: Mun. Trujillo, rd. Trujillo- Naranjal, km 32, Rio Sanquininf, ca. 900 m, 11 Sep 1993 (Y fl-fr), Franco et al. 4485 (BG), (d), Franco et al. 4486 (BG); Mun. San Jose del Palmar, ca. 1 km W of San Jos6 del Palmar, ca. 900 m, 19 Mar 1994 (cd), Franco et al. 4564 (BG, HUA), (Y fl-fr), Franco et al. 4565 (BG, HUA). VALLE: Rio Anchicaya, nr. CVC hydroelectric plant, 400- 500 m, 16 Dec 1981 (Y fl-fr), Gentry 35680 (BG, COL, MO, TULV).
The habit and the presence of irritating bristles are indications of close relationship to Cecropia hispidis- sima. Both species are not inhabited by ants of the genus Azteca, but by representatives of the genus Pa- chycondyla.
Cecropia setico Snethlage ex Macbride, Publ. Field Mus. Nat. Hist., Bot. Ser. 13(2.2): 289. 1937. Type. Peru. Loreto: Rio Marafi6n, nr. Uanana, 1 Aug 1924 (Y), Tessmann 3713 (holotype: B, destroyed, pho- tographs in F, MO; isotypes: NY, S).
Cecropia occidentalis Cuatrecasas, Revista Acad. Col- omb. Ci. Exact. 6(22/23): 277. 1945; 7(27): t. 5.6, t. 6.2. 1947. Type. Colombia. Valle: Rio Calima, La Trojita, 22 Feb 1944 (6), Cuatrecasas 16347 (lec- totype, Berg & Franco Rosselli, Fl. Ecuador 48: 36. 1993: COL; isolectotype: F).
Tree, to 25(-35) m tall. Leafy twigs 2-4 cm thick, usually pale to dark green (to reddish), hispid to (sub)setulose. Lamina chartaceous to subcoriaceous, ca. 40 X 40 cm to 80 X 80 cm, the segments (7-)8- 10, the free parts of upper segments obovate, the in- cisions down to ca. 5/10-7/10; apices acuminate; up- per surface smooth, puberulous on the veins and with very sparse arachnoid indumentum or glabrous; lower surface sparsely to densely minutely puberulous, also with longer uncinate hairs or almost glabrous on the veins, the arachnoid indumentum confined to the mar- gin, occasionally present in the areoles; lateral veins in the free part of the midsegment 10-13 pairs, sub- marginally loop-connected, several of them branched; petiole 40-75 cm long, glabrous; trichilia fused, the
brown indumentum intermixed with sparse short white hairs; stipules 8-20(-26) cm long, usually green, sometimes dull red, almost glabrous or (very) sparsely hirtellous to hirsute outside, sparsely to rather densely sericeous inside. Staminate inflores- cences in pairs, pendulous; peduncle 4-6 cm long pu- berulous to hirtellous; spathe 8-10 cm long, yellow- ish, hirtellous outside, glabrous inside; spikes ca. 30- 40, 7-11(-16) X 0.2 cm, with stipes 0.5-1 cm long and hirtellous; rachis hairy. Staminate flowers: peri- anth cup-shaped, 0.8 mm long, at the upper part with stiff hairs; filaments swollen; anthers ca. 0.4-0.5 mm long, not appendiculate, remaining attached to the fil- ament at anthesis. Pistillate inflorescences in pairs, pendulous; peduncle 5-15(-20) cm long, hispid, sometimes only just below the spathe; spathe 12-18 cm long, the color and indumentum as in the stami- nate inflorescence; spikes (2-)4, (7-)10-16 X 0.4-0.6 cm, to 30 X 1.4 cm in fruit, with stipes 0.5-1 cm long and hirtellous; rachis glabrous. Pistillateflowers: per- ianth ca. 1.5-2 mm long, with arachnoid indumentum below the apex, also below the aperture inside, the apex slightly convex, minutely and often ? sparsely muriculate; style rather long, straight; stigma penicil- late. Fruit oblongoid, ca. 2.5 mm long, smooth.
Distribution (Fig. 8.6). Upper Amazon basin (from Bolivia to Venezuela), the Pacific coastal region of Ecuador and Colombia, extending to eastern Pan- ama, in the lower Rio Magdalena valley (Colombia), and the lower Cauca valley (Colombia), common in periodically inundated places along (white-water) riv- ers, also in non-inundated places, but then often near rivers, at elevations to ca. 1000(-1250) m.
Representative specimens examined. PANAMA. DARIEN: Cerro Pirre, Rio Pirre, S of El Real, 750-1030 m, 11 Aug 1962 (6), Duke 5355 (MO); Cruce de Mono, 5 Nov 1989 (Y fr), Fisher 55 (BG); nr. El Real, Rio Tuira, 7 Nov 1989 (9 fl-fr), Fisher 56 (BG).
COLOMBIA. AMAZONAS: Mun. Leticia, Cgto. Puerto Santander, Rio Caqueta, Santa Isabel, 7 Apr 1994 (9 fl), Ca'rdenas et al. 4599 (COL); between Leticia & Rio Ama- cayacu, Isla Santa Soffa, 15 Aug 1972 (D fr), Idrobo 6518 (AAU, COL); Rio Igara-Parana, 15 km below La Chorrera, 18 May 1975 (d), Idrobo 8315 (AAU, COL); Rio Caqueta, nr. mouth of Quebrada Quinch6, 7 May 1988 (st), M. Sdn- chez S. et al. 280 (BG, U); Rio Caqueta, Isla El Tigre, 19 Sep 1988 (st), M. Sanchez S. et al. 1321 (BG, U); Rio Ama- zonas, above Leticia, Isla de Monos, 25 Apr 1987 (9 fl-fr), Gentry et al. 57097 (BG, MO); Dtto. Puerto Narinio, Parque Nacional Amacayacu, 2 Aug 1989 (d), R. Vdsquez et al. 12447 (COL, MO). ANTIOQUIA: Parque Nacional Natural Los Katios, Rio Atrato, Isla de Tanggara, 24 Oct 1979 (9 fl), Barbosa 1334 (COL); Mun. Necoclf, Reserva Indigena Caiman Nuevo, Rio Caiman, Aug 1992 (6), Castano et al.
SYSTEMATIC TREATMENT 117
FIG. 33. Cecropia membranacea. 1. Leaf, reduced. 2. Apex of lamina and venation (Berg et al. 1092). 3. Twig, longitudinal section. 4. Stipules (Oldeman et al. 90). 5. Pair of staminate inflorescences with spathes and base of petiole with trichilium (Balslev 2280). 6. Staminate inflorescence at anthesis (Berg et al. 1090). 7. Staminate flower and stamen (Ferreira s.n.). 8. Pair of pistillate inflorescences with spathes and base of petiole with trichilium. 9. Pair of pistillate inflorescences at anthesis. 10. Pistillate inflorescence after anthesis (Berg et al. 1092). 11. Pistillate flower. 12. Fruit (Berg et al. P.17584). (From Fl. Ecuador 48: 37. 1993, modified.)
118 FLORA NEOTROPICA
200 (HUA); between Villa Arteaga & Chigorod6, 1 Oct 1961 (6), Cuatrecasas et al. 26107 (COL, P, US), (9 fl-fr), Cua- trecasas et al. 26113 (COL, P, US); Taraza', Cgto. El Doce, 28 Mar 1994 (9 fl), Franco et al. 4601 (BG, HUA); Rfo Atrato, rd. to Codelsa, km 9-10 from entrance, 11 Mar 1970 (9 fl-fr), Idrobo 6318 (COL); Casabe, 30 Oct 1979 (9 fl- fr), Renterfa et al. 1929 (COL, HUA, JAUM, MO). BOL- iVAR: Rio Magdalena, 18 Dec 1969 (d), Idrobo 6300 (COL), (9 fl), Idrobo 6301 (COL). CAQUETA: Mouth of Rfo Yari, nr. Araracuara, 15 Jul 1986 (9 fl-fr), Berg et al. 1563 (BG, COL); rd. Puerto Rico-San Vincente del Caguan, 4 Oct 1975 (9 fl), Cabrera 3753 (F); Rio Orteguaza, Ve- necia, 31 May 1940 (9 fl), Cuatrecasas 8953 (COL, F); Florencia, El Venado, 25 Jan 1969 (9 fl-fr), Cuatrecasas et al. 27240 (COL, NY, US); Rfo Caqueta, Araracuara, Centro Experimental las Penias, 14 Mar 1991 (9 fl), Franco et al. 3349 (COL); ca. 25 km N of Solano, on Rio Caqueta, 8 km SE of Tres Esquinas, below mouth of Rio Orteguaza, 14 Mar 1945 (st), Little et al. 9787 (COL, US); Morelia, 2 Oct 1941 (6), Sneidern A.1053 (A, MICH, NY, S, US). CAUCA: Nr. San Francisco, 23 Sep 1968 (9 fl), Espinal 2746 (MEDEL). CESAR: Rfo Magdalena, 15 Oct 1969 (6), Irusta & Fortoul Ltd. s.n. (COL). CHOC6: Rfo San Juan, Quebrada del Ta- paral, 30 May 1946 (st), Cuatrecasas 21511 (F, VALLE); Rfo San Juan, between Palestina & Agua Negra, 1 Jun 1946 (9 fl), Cuatrecasas 21577 (F, VALLE); Rfo Atrato, below Quibd6, 3 Apr 1958 (st), Cuatrecasas et al. 24203 (US); Rio Salaquf, 17-29 Oct 1989 (d), Mahecha etal. 6244 (UDBC); Rio Taparal, tributary of Rfo San Juan, 15 Nov-6 Dec 1979 (6 fl-fr), Rooden et al. 675 (BG, NY, U); Nuquf, Alto de Buey, 1000 m, 26 Jun 1940 (6), Sneidern A.72 (A, LL, MICH, NY, US). META: Rio Meta, Bocas del Tua, 6 Nov 1938 (st), Cuatrecasas 4490 (COL, F, S); nr. La Macarena, 11 Feb 1995 (6), Franco et al. 4669 (BG, COL, HUA, MO, US); Reserva Nacional La Macarena, Rfo Guayabero, 13 Mar 1959 (6), Jaramillo M. 2153 (COL); nr. Puerto Lopez, 30 Jul 1944 (9 fl-fr), Little 8337 (COL, US); Parque Na- cional Natural Tinigua, Serranfa Chamusa, Apr 1991 (6), Stevenson 294 (COL). NORTE DE SANTANDER: Rio Mag- dalena, mouth of Rfo Lebrija, 17 Dec 1969 (9 fl-fr), Idrobo 6298 (AAU, COL), (6), Idrobo 6299 (COL). PUTUMAYO:
Rfo Putumayo, Puerto Ospina, 29 Nov 1940 (9 fl-fr), Cua- trecasas 10867 (COL, F); Rfo San Miquel, between Bermeja & Rio Conejo, 13 Dec 1940 (9 fl-fr), Cuatrecasas 11063 (COL, F); Mun. Orito, Vrda. El Parafso, 17 Feb 1995 (d), Franco et al. 4683 (BG, COL, HUA, LP, MO, NY, US); Mocoa, Jul 1989 (9 fr), Mahecha et al. s.n. (UDBC 7853); Rfo Caucaya, nr. Puerto Jaramillo, 18 May 1942 (6), Schul- tes 3778 (COL, GH). SANTANDER: Rio Magdalena, nr. Bar- ranca Bermeja, 20 Apr 1926 (st), Bosse' 7633 (LE); Mun. Barrancabermeja, Cgto. Chucurf, 23 Apr 1983 (d), Moreno I (COL). VALLE: Mun. Buenaventura, Cgto. El Tigre, Que- brada Tatama, 26 Oct 1979 (9 fl-fr), Cuadros V 891 (MO, TULV), (d), Cuadros V 892 (MO, TULV); Rio Calima, La Trojita, 22 Feb 1944 (9 fl-fr), Cuatrecasas 16349 (VALLE); Rfo Calima, Pail6n, 23 May 1946 (st), Cuatrecasas 21242 (F, VALLE); Mun. Versalles, Vrda. El Cedro, 1000 m, 1 Sep 1983 (9 fl-fr), Devia 301 (COL, TULV); Mun. Trujillo, rd. Trujillo-Naranjal, Rfo Sanquininf, 1250 m, 11 Sep 1993 (9
fl-fr), Franco et al. 4487 (BG); Buenaventura, 7-9 May 1922 (st), Killip 5312 (NY, US); Mun. Buenaventura, San Isidro, 15 Nov-6 Dec 1979 (Y fl-fr), Rooden et al. 341 (BG, NY, U), (d), Rooden et al. 344 (BG, MO, NY, U). VAUPES: Upper Rio Guaviare, 9 Nov 1939 (9 fl-fr), Cuatrecasas 7617 (COL, US).
VENEZUELA. AMAZONAS: Nr. San Carlos de Rfo Ne- gro, 14 Dec 1978 (9 fl-fr), Clark 6880 (MO, QCNE); San Carlos de Rio Negro, 21 Mar-17 Apr 1988 (9 fl), Delascio et al. 9304 (VEN); base of Cerro Duida, Jan-Feb 1969 (st), Farifias 349 (MO, NY); 5-6 km NE of San Carlos de Rio Negro, 12 Apr 1979 (9 fl-fr), Liesner 6573 (MO, U); San Carlos de Rio Negro, ca. 20 km S of confluence of Rio Negro & Brazo Casiquiare, 1986-1988 (6), Uhl 520 (MO); So- lano, Lower Rio Casiquiare, 10 Mar 1942 (9 fr), LI. Wil- liams 14733 (US).
ECUADOR. Los Rios: Rio Palenque Biological Sta- tion, rd. Quevedo-Santo Domingo de los Colorados, km 56, 2 Oct 1976 (st), Dodson et al. 6235 (QCA). NAPO: Between Tena & Archidona, 12 Oct 1939 (9 fl-fr), Asplund 9261 (S); Rio Payamino, nr. Coca, 27 Feb 1980 (c), Berg et al. 1090 (AAU, BG, COL, MO, NY, QCA, TUR, U), (9 fl-fr), Berg et al. 1092 (AAU, BG, COL, MO, QCA, TUR, U); Reserva Biol6gica Jatun Sacha, 8 km E of Mishualli, 29 Jan 1989 (9 fl), Ceron 6081A (BG); Rio Napo, Santa Rosa, 28 Apr 1972 (d ), Lugo 2003 (BG, GB); Rio Sumino, tributary of the Rio Napo, ca. 5 km NE of Santa Rosa, 2 May 1972 (4), Lugo 2104 (BG, GB); Mun. Orellana, Parque Nacional Yasuni, Rio Indillama, Comuna Pompeya, 26 Aug 1992 (9 fl-fr), Neill et al. 10107 (BG, QCNE), (6), Neill et al. 10130 (BG, QCNE); Cononaco, 27 May 1976 (9 fl), Oldeman et al. 90 (QCA); Parque Nacional Yasuni, Afiangu, nr. mouth of Rio Aniangu, 30 Jun-9 Jul 1982 (st), SEF 10099 (AAU, QCA, QCNE, U). PASTAZA: Lorocachi, Rio Curaray, 29 May 1980 (9 fl-fr), Jaramillo 31352 (AAU). SUCUMBiOS: Nr. mouth of Rio San Miguel, Aug 1980 (9 fl-fr), Andrade 33149 (AAU); nr. Limoncocha, 9 Apr 1982 (6), Balslev 2280 (QCA, NY); Reserva Faunistica Cuyabeno, nr. Laguna Grande, 1988-1990 (st), Christensen 82750 (AAU); Rfo Aguarico, 75?22'W, 0?37'S, island midway between Zan- cudo & Lagarto Cocha, 30 Aug 1979 (6 + 9 fl-fr), Holm- Nielsen et al. 20205 (AAU); Santa Cecilia, rd. Lago Agrio- Baeza, ca. 16 km W of Lago Agrio, 27 Feb 1973 (6), Lugo 3542 (BG, GB).
PERU. AMAZONAS: Rio Numpatkin, Kusu, 10 Mar 1975 (9 fl-fr), Ancuash 50 (GH); Prov. Bagua, Yamayakat, 29 Jan 1996 (9 fl-fr), Jaramillo et al. 994 (MO); Rio Ce- nepa, Quabrada Huampami, 16 May 1973 (9 fl-fr), Kayap 805 (F, GH, USM). Cuzco: Prov. Paucartambo, Pilcopata, 9 Feb 1975 (9 fl-fr), Plowman et al. 5100 (GH, US); Prov. Quispicanchis, between Suamsari & Fortaleza, 9 Dec 1966 (9 fl-fr), Vargas C. 18486 (US). HUANUCO: Nr. Tingo Maria, Cueva de las Lachusas, 16 Jul 1957 (9 fl-fr), Ellen- berg 2385 (U); Prov. Puerto Inca, Dtto. Yuyapichis, DAN- TAS, 1-15 Dec 1988 (9 fl), Kroell S. 190 (G). LORETO: Rd. Jenaro Herrera-Angamos, km 10, 18 Jul 1988 (9 fl-fr), Berg et al. 1600 (BG, UMS); nr. mouth of Rio Nanay, 29 Sep 1972 (9 fl-fr), Croat 20818 (F, MO, NY); Prov. Maynas, Rio Gueppi, above Puerto Peru, 14 May 1978 (9 fl), Gentry
SYSTEMATIC TREATMENT 119
et al. 21817 (F, G, U, USM); Prov. Maynas, Rio Amazonas, between Yanomono & mouth of Rfo Manati, 29 Jun 1983 (Y fl-fr), Gentry 42327 (BG, MO); Allapahuayo, 39 km SW of Iquitos, 24 Feb 1988 (d), Gentry et al. 61909 (MO); Rio Sucusari, nr. Rio Napo, 15 Jul 1981 (Y fr), Hahn 6 (MO); Iquitos, 23 Feb 1924 (Y fl-fr), Kuhlmann 1486 = RB 19903 (RB); Prov. Alto Amazonas, Rio Huasaga, nr. Wishintsa, 16- 26 Jun 1986 (Y fl), Lewis 11206 (MO); Prov. Maynas, Yan- amono, 21 May 1983 (Y fl), R. Vdsquez et al. 4136 (NY). SAN MARTiN: Prov. San Martin, rd. Tarapoto-Lamas, ca. km 7, 4 Dec 1997 (6), Berg et al. 1771 (BG, COL, MOL); Prov. San Martin, rd. Tarapoto-Chazuta, km 13, nr. bridge over Rio Mayo, Juan Guerra, 6 Dec 1997 (Y fl), Berg et al. 1782 (BG, COL, MOL). MADRE DE DIos: Prov. Tambo- pata, Comunidad Nativo de Infierno, Hermosa Chica, 24 Feb 1989 (st), Alexiades et al. 353 (BG, MG, MO); Rfo Tam- bopata, 40 km upriver from Puerto Maldonado, 15 Jan 1991 (Y fl-fr), Alexiades et al. 1097 (BG, K); Prov. Tambopata, 15 km ENE of Puerto Maldonado, 1991 (6), Fisher 177 (BG); Parque Nacional Manu, Rio Manu, Cocha Cashu Sta- tion, 3 Nov 1986 (d), Foster 12084 (BG), 7 Nov 1986 (st), Foster et al. 12202 (INPA, MOL); Parque Nacional Manu, Rio Manu, Pakitsa Station, 26 Dec 1988 (Y fl), Foster et al. 12825 (BG, LPB), (d), Foster etal. 12827(BG, LPB); Prov. Manu, Cocha Juarez, 28 Sep 1989 (Y fl-fr), Foster et al. 13284 (BG, F); Prov. Tambopata, Rio Heath, Peru-Bolivia border, 2 Mar 1990 (? fl-fr), Gentry et al. 69785 (MO); Prov. Manu, Rio Salvaci6n, 5 Dec 1991 (Y fl), Ntiuiez et al. 14627 (BG); Prov. Tambopata, Cuzco Amaz6nico, 1 Jun 1989 (Y), Phillips et al. 172 (MO). PAsco: Prov. Oxa- pampa, Rfo Pichis valley, nr. Paujil, 10 km downriver from Puerto Bermudez, 28 Sep 1982 (d), Foster et al. 8948 (MO, NY, U, USM). SAN MARTfN: Rd. Tocache-Tingo Maria, N of El Progreso, 15 Mar 1979 (Y fl), Gentry et al. 25762 (BG, MO). UCAYALI: Prov. Coronel Portillo, Pucallpa, Pu- callpillo, 8 Apr 1953 (Y fl-fr), Ferreyra 9030 (USM); Prov. Coronel Portillo, nr. Pucallpa, Isla Progresso, (6), Ferreyra 9044 (USM).
BRAZIL. AcRE: Upper Rio Moa, Fazenda Arizona, 24-30 1984 (st), Campbell et al. 8197 (BG); Serra de Moa, 30 Apr 1971 ( fl-fr), Maas et al. P 12674 (INPA, K, MO, NY, U, US); Rio Jurua-Mirim, nr. Porangaba, 24 May 1971 (Y fl-fr), Maas et al. P.13281 (INPA, MO, K, NY, P, S, U, US); Mun. Mancio Lima, Rio Moa, Parque Nacional da Serra do Divisor, 13 May 1996 (Y fr), M. Silveira et al. 1327 & 1329 (NY). AMAZONAS: Rio Solimoes, S of Ma- naus, 2 Oct 1973 ( fr), Berg et al. P 17584 (C, COL, INPA, MG, NY, P, U, US); Rio Solimoes, 10 km E of Boca de Januaca, 2 Oct 1973 (Y fl-fr), Berg et al. P 17585 (F, INPA, K, NY, S, U); Mun. Humaita, Rio Madeira, Igarape do Ban- heiro, 2 km N of Humaita, 2 May 1975 (Y fl-fr), Gottsberger 35-2575 (INPA); Tef6, Rio Solimoes, 23 Jul 1972 (Y fl-fr), Krieger et al. 12284 (GUA, INPA); Mun. Sao Paulo de Oli- ven,a, nr. Palmares, 11 Sep-26 Oct 1936 (9 fl), Krukoff 8571 (A, BM, F, G, K, MICH, MO, NY, P, S, U, US); Rio Solim6es, nr. Manaquiri, 27 Jul 1979 (9 fl-fr), Rodrigues 8892 (INPA); Rio Solimoes, Lago do Capitari, 25 Jun 1988 (9 fr), Ziburski 88/50 (INPA). PARA: Mun. Afua, Rio Ca- juuna, 4 Dec 1992 (st), Maciel et al. 2113 (MG).
BOLIVIA. BENI: Prov. Moxos, Rio Chirizi, 130 km S of San Ignacio, 12 Oct 1991 (st), Del Aguila et al. 168 (BOLV); Prov. Yacuma, Estaci6n Biol6gica Beni, Rio Cu- riraba, 25 Dec 1987 (9 fl), Moraes R. 982 (BG, LPB, MO, USZ); Prov. Ballivian, rd. Yucumo-Rurrenabaque, km 38, 7-14 Apr 1989 (9 fl-fr), D. N. Smith et al. 12897 (LPB, MO); Prov. Ballivian, rd. Yucumo-Rurrenabaque, km 35, Mar-Jul 1990 (st), D. N. Smith et al. 14349 (BG, MO). COCHABAMBA: Prov. Carrasco, Estaci6n Valle del Sajta UMSS, 29 Oct 1991 (st), Atahuachi et al. 23 (BOLV, USZ); nr. Chipiri, ca. 5 km N of Villa Tunari, 6 Dec 1995 (9 fl), Berg et al. 1729 (BG, BOLV, COL, LPB, USZ). LA PAZ:
Nr. Sapecho, 26 Feb 1994 (9 fl-fr), Berg 1698 (BG, COL, LPB). PANDO: Prov. Nicolas Suarez, "en la zona del Na- reuda, junto a la barraca San Jose," 12 Jan 1983 (d), Fer- ndndez C. et al. 8203 (G); Prov. Manupiri, nr. Independen- cia, 23 Aug 1985 (st), Moraes R. 263 (BG, LPB). SANTA
CRUZ: Prov. Ichilo, Rio Ichilo, Reserva Forestal El Chor6, 15 Mar 1990 (9 fl-fr), Saldias 1038 (USZ).
Cecropia membranacea and C. litoralis are the only lowland species represented both in the Amazon basin and in the Pacific Coastal region. The species is abundant along white-water rivers, as a component of early successional stages on well-aerated soil (cf. Lamotte, 1992). However, it can also be found in flood plain forest, although often in wet places and/ or inundated for short periods, or (less often) far away from rivers. In such places the species is often less ephemeral and can become large trees. The white arachnoid indumentum on the lower surface of the lamina is normally confined to the margin; sometimes some sparse arachnoid is present in the areoles, but it soon disappears. However, in Cuatrecasas et al. 26107 from Antioquia (Colombia) the arachnoid in- dumentum in the areoles appears to be more persist- ent. Some collections from Ecuador (Cer6n 712 and Hurtado et al. 2403), erroneously included in C. id- roboi by Berg & Franco Rosselli (1993: 27), and one from Colombia, Putumayo (Franco et al. 4679), are distinct by the presence of minute arachnoid indu- mentum in the areoles of the lamina, which is thicker (coriaceous to subcoriaceous) than normal in C. mem- branacea. They largely match C. membranacea in other morphological vegetative parts. Collections with staminate inflorescences, which could make af- finities clearer, are wanting. The collections examined have not been made from the typical C. membranacea habitat, and they may represent a type with aberrant phenology. They are presently included in C. mem- branacea with some doubt.
Specimens examined. COLOMBIA. PUTUMAYO:
Puerto Asis, on rd. to Kanakas, 6 Feb 1995 (9 fl-fr), Franco et al. 4679 (BG, COL).
ECUADOR. NAPO: Reserva Biol6gica Jatun Sacha, 8
120 FLORA NEOTROPICA
km E of Misahualli, 17 Jan-6 Feb 1987 (Y fl-fr), Cer6n 712 (BG, QCNE), 22 Feb 1990 (st), Ceron et al. 8865 (BG); Cant6n Archidona, rd. Hollin-Loreto, km 65, Huaticocha, 14-19 Jul 1989 (9 fl-fr), Hurtado et al. 2403 (BG, QCNE).
The incomplete collection from Peru. Loreto, Santa Rosa, nr. Yurimaguas, 11 Nov 1929 (9 fl), Ll. Williams 4949 (F), for which the vernacular name "pungara" is noted, might also belong to this type.
In addition to this type of Cecropia membranacea with arachnoid indumentum on the lower surface of the lamina, two other types have to be mentioned: one indicated as "pungara" in southern Amazonian Peru (Madre de Dios) and the other, which can be indicated as "herrerensis," in northern Amazonian Peru (Lor- eto); see page 220. The latter is probably represented by Berg et al. 1600, which was not made in a typical C. membranacea habitat and distinct by reddish stip- ules. With the very scarce and incomplete material available, these two types can hardly or not be distin- guished morphologically from typical C. membra- nacea. They may produce trichilia earlier than normal in the species and they are not inhabited by Azteca ants. The ecology of these types (may) deviates more or less, also with regard to light regimes, and conse- quently habitat preferences (Davidson & Fisher, 1991). Cecropia membranacea is in cultivation in the Botanical Garden of Universidad Nacional Agraria, La Molina, Lima (Peru).
Vernacular names. Colombia: guanibarae, uda- gomo (Mui., Amazonas); yarumo corante (Ama- zonas); guanibarae, yarumo (Caqueta'); yarumo ma- cho (Choco). Venezuela: yagrumo rojo (Amazonas). Ecuador: bokarawa (Auca, Napo); dondo, tsitica dundu (Quichua, Napo). Peru: setico blanco (Huain- uco); suui, sutl, sutlk, or sutiik (Jivaro, Loreto); ka'a- bona or taa-bona (Ese-ejhal, Madre de Dios). Bolivia: ambaibo blanco (Beni).
32. Cecropia metensis Cuatrecasas, Revista Acad. Colomb. Ci. Exact. 9(36/37): 338. 1956. Type. Co- lombia. Meta: Puerto L6pez, 29-30 Jul 1946 (9 fl-fr), Jaramillo M. et al. 378 (holotype: US; iso-
type: COL).
Cecropia peltata Linnaeus var. candida Velasquez, Acta Bot. Venez. 6: 54, t. 12. 1971. Syntypes. Venezuela. Guarico: Nr. Calabozo, Morichal Villasmil, nr. Es- taci6n Biol6gica de Los Llanos, 8 Aug 1966 (9), Veldsquez 82 (VEN; isosyntype: US), (d), Veldsquez 83 (VEN; isosyntype: US).
Tree, to 12(-20) m tall, the trunk with prominent stipular scars. Leafy twigs 1-3 cm thick, dark green
to purplish brown with conspicuous lenticels (or white), densely hispidulous to puberulous with curved to straight hairs, sometimes also with dense arachnoid indumentum. Lamina chartaceous to subcoriaceous, ca. 15 X 15 cm to 60 X 60 cm, the segments (7-)8- 10(-1 1), the free parts of upper segments elliptic to oblong to (sub)obovate (when juvenile often ? lo- bate), the incisions down to 6/10-8/10; apices obtuse to short-acuminate; upper surface ? scabrous, sparsely to rather densely hispidulous and (initially) with sparse arachnoid indumentum; lower surface (minutely) puberulous with longer uncinate hairs, arachnoid indumentum in the areoles and on the smaller veins, extending to the main veins; lateral veins in the free part of the midsegment 9-16 pairs, marginally loop-connected, mostly branched; petiole 10-50 cm long, densely to sparsely puberulous to hir- tellous and with ? dense (or sparse, occasionally very sparse) arachnoid indumentum; trichilia fused, the brown indumentum intermixed with short or often long white hairs; stipules 6-13 cm long, white, hir- tellous to subhirsute and with dense arachnoid indu- mentum outside, sericeous inside. Staminate inflores- cences in pairs or solitary, the peduncle erect to + deflexed, the spikes erect to pendulous; peduncle 6- 15 cm long, ? densely puberulous to hirtellous and with ? dense (or sparse) arachnoid indumentum; spathe 7-11 cm long, white, hirtellous to subhirsute or to puberulous and with (very) dense arachnoid in- dumentum outside, sericeous inside; spikes (4-)6-20, 3-12 X 0.2-0.3 cm, with stipes 0.1-0.8 cm long and densely puberulous or with arachnoid indumentum; rachis hairy. Staminate flowers: perianth tubular, ca. 1 mm long, glabrous, the apex plane to slightly con- vex; filaments flat; anthers ca. 0.5 mm long, appen- diculate, detached at anthesis, reattached to the mar- gins of the aperture by the appendages. Pistillate inflorescences in pairs or solitary, erect to deflexed, becoming pendulous in fruit; peduncle 3-12(-17) cm long, ? densely puberulous to hirtellous to hispidu- lous and with ? dense (or sparse) arachnoid indu- mentum, sometimes subglabrous; spathe 8-19 cm long, the color and indumentum as in the staminate inflorescence; spikes 2-4, 5-9 X 0.4-0.6 cm, to 16 X 1.3 cm in fruit, sessile (or with stipes 0.5-0.8 cm long and sparsely puberulous and with arachnoid in- dumentum); rachis glabrous (but hairs at the base of the free part of the perianth). Pistillateflowers basally connate; perianth ca. 1.5 mm long, with arachnoid indumentum below the apex outside, absent (or pres- ent in the style channel) inside, the apex slightly con- vex; stigma peltate. Fruit oblongoid, 1.8-2 mm long, tuberculate.
SYSTEMATIC TREATMENT 121
Distribution (Fig. 18.2). The Ilanos region of Co- lombia and Venezuela and savannas of Guyana, in (white sand) savannas, gallery forests and swamps (morichales), at low elevations.
Representative specimens examined. COLOMBIA. CASANARE: Orocue, Canlo San Miguel, Jul 1989 (9 fl-fr), Mahecha 5588 (UDBC), (5), Mahecha 5589 (UDBC). META: Mun. Remolinos, Centro Recreacional Cafamllanos, Cafno Matamata, 30 Sep 1995 (5), Bottia I (COL); rd. Puerto Gaitan-San Pedro de Arimena, Alto Neblinas, 15 Mar 1986 (5), Forero et al. 10239 (COL); Mun. Villavicen- cio, rd. Villavicencio-Puerto L6pez, 10 Feb 1995 (9 fl-fr), Franco et al. 4655 (COL), (5), Franco et al. 4656 (COL); Mun. Puerto L6pez, Alto Menagua, 10 Feb 1995 (5), Franco et al. 4660 (COL); Mun. Puerto L6pez, Puenta La Balsa, 10 Feb 1995 (9 fl), Franco et al. 4661 (BG, COL, HUA, MO, US), (juv), Franco et al. 4662 (COL, HUA, MO, US); nr. La Macarena, 11 Feb 1995 (5), Franco et al. 4465 (BG, COL, HUA, MO, US); nr. La Macarena, nr. rapids of Rio Guayabero, 1 1 Feb 1995 (5), Franco et al. 4670 (BG, COL, HUA, MO, US), (9 fl-fr), Franco et al. 4671 (BG, COL, HUA, NY, US); San Juan de Arama, Rio Giiejar, 5- 20 Dec 1950 (5), Idrobo et al. 604 (US); San Martfn, 22 Jan 1972 (5), Idrobo et al. 6489 (AAU, COL); Puerto L6- pez, 27 Jul 1944 (9 fr), Little et al. 8242 (COL, F, P, US); 10 km S of Puerto Gaitan, 12 Mar 1971 (9 fl), Pinto et al. 1165 (COL, U); Parque Nacional Natural Tinigua, Serrania Chamusa, Apr 1992 (5), Stevenson 361 (COL); Rio Melda, Finca La Florida, Nov 1993 (5), Stevenson 545 (COL).
VENEZUELA. AMAZONAS: Depto. Atures, Rio Catan- iapo, 25 Jul 1981 (5), Castillo 1337 (VEN); 8 km S of Puerto Ayacucho, 13-15 Apr 1978 (9 fl-fr), Davidse et al. 14998 (MO, U, VEN); Depto. Atabapo, 15 km SW of Santa Barbara del Orinoco, 26 Jun 1979 (9 fl-fr), Huber 3869 (VEN). ANZOATEGUI: Bella Vista, Rfo Orinoco, 19 Nov 1970 (st), Morillo 113 (VEN); Rio Guaraguara, nr. Santome, 17 Jun 1941 (9 fl-fr), Pittier 14842 (US), (5), Pittier 14845 (VEN), 23 Jun 1941 (5), Pittier 14878 (US); nr. La Leona, Rio Chives, 25 Jun 1941 (5), Pittier 14907 (VEN). APURE:
Dtto. Pedro Camejo, 22 km WNW of Buena Vista, 2-3 km N of Isla Algarrobe, 17 Feb 1978 (9 fl), Davidse et al. 14302 (MO, U, VEN); Dtto. Pedro Camejo, ca. 62 km NNE of Puerto Pdez, 28 Feb 1978 (5), Davidse etal. 14637(MO, NY, U, VEN); Dtto. Pedro Camejo, ca. 53 km NE of Puerto Paez, 21 Feb 1979 (9 fl), Davidse et al. 15555 (MO, U, VEN); Dtto. Pedro Camejo, 16 km NW of Mata de Guan- abana, between Rio Meta & Rio Cinaruco, 27 Feb 1979 (9
fr), Davidse et al. 15831 (MO, NY, U, VEN); Dtto. Pedro Camejo/Anchagua, rd. San Fernando-Puerto Paez, between Rio Capanaparo & Cafio La Guardia, 7 Jul 1988 (d), G6mez et al. 216 (VEN). BARINAS: Corozo, La Vizcaina, 5 May 1980 (5), Marcano Berti et al. 94-980 (U). BOLiVAR: Mun. Piar, Sector Las Patillas, Feb 1987 (9 fl), Fernandez 2993 (BG, VEN). GUARICO: Morichal Largo, 17 km SE of Cal- abozo, 15 Dec 1982 (5), Brito 83 (MO, NY, VEN); nr. Par- mana, 3-10 Apr 1950 (d), Croizat 196 (F); Dtto. Miranda, Morichal El Recreo, Jul 1982 ( 9 fl-fr), Delascio et al. 11720 (VEN); nr. Calabozo, Morichal Villasmil, nr. Estaci6n Biol-
6gica de Los Llanos, (Y fl-fr), Veldsquez 84 (US); rd. El Sombrero-Calabozo, 8 Aug 1966 (9 fl-fr), Veldsquez 89 (US, VEN); nr. Calabozo, nr. Laguna de los Patos, Estaci6n Biol6gica de Los Llanos, 8 Aug 1966 (9 fl-fr), Veldsquez 90 (US), (d ), Veldsquez 91 (US). SucRE: Montafia de Mo- chima, 20-21 km SW of Cumana, 16 Sep 1973 (9 fl-fr), Steyermark et al. 108727 (MO, NY, VEN).
GUYANA. Nr. Aishalton Hospital, 19 Nov 1982 (st), Stoffers et al. 457 (BG, NY, U); Lake Awakawau, 5 km SE of Aishalton, 20 Nov 1982 (d), Stoffers et al. 516 (U).
BRAZIL. RORAIMA: 12 km S of Boa Vista, 13 Apr 1989 (d), Guillaumet et al. 5850 (INPA), (9 fl-fr), Guillau- met et al. 5851 (INPA).
Cecropia metensis is characterized by dense white arachnoid indumentum on stipules, spathes, and usu- ally also petioles and peduncles, and by its occurrence in savanna regions. The presence of arachnoid indu- mentum distinguishes C. metensis from C. sararensis in most cases, and if not by this arachnoid indumen- tum, then by the dense indumentum of straight to un- cinate hairs on the petiole. Although the areas of C. metensis and C. sararensis touch (or slightly over- lap?) each other, intermediates between them are rare, suggesting the presence of rather solid genetic barri- ers. The collections Franco et al. 4655, 4656, and 4663 lack more or less dense arachnoid indumentum on the petioles, but both the stipules and spathes are covered by dense arachnoid indumentum. Moreover, the relatively long hairs are not present in the trichilia of Franco et al. 4663. Because of these features, these collections could be regarded as transitional to C. sar- arensis. Material of C. sararensis from Arauca also shows features more or less transitional to C. meten- Sis.
33. Cecropia montana Snethlage, Notizbl. Bot. Gart. Berlin-Dahlem 8: 368. 1923. Type. Peru. San Mar- tin: Tarapoto, Cerro de Escaler, 1300 m, Mar 1903 (9), Ule 6845 (holotype: B, destroyed, fragment and photograph in F; isotype: MG, photographs in F, GH). Fig. 34
Tree, to 25 m tall. Leafy twigs 1.5-4 cm thick, green to red-brown or brownish to whitish, hispidu- lous or hispid to subhirtellous. Lamina chartaceous to subcoriaceous, ca. 35 x 35 cm to 85 x 85 cm, often
122 FLORA NEOTROPICA
~~~~~~~~~~~4\* ~ ~~~~~~~~~~~~~~~~~~~~~~~~~~~~4
10 ~9 3
FIG. 34. Cecropia montana. 1. Lamina, reduced. 2. Apex of lamina and venation (Berg et al. 1661). 3. Young leaf (Palacios 8178). 4. Base of petiole with trichilium (Neill et al. 9704). 5. Stipules (Berg et al. 1656). 6. Staminate inflorescence with spathe (Palacios 8178). 7. Staminate inflorescence at anthesis and base of petiole with trichilium (Berg et al. 1661). 8. Staminate flower and stamen (Berg et al. 1656). 9. Pistillate inflorescence with spathe. 10. Pistillate inflorescence at anthesis, opened prostoma, and base of petiole with trichilium (Neill et al. 9704). 11. Pistillate inflorescence after anthesis (Berg et al. 1662). 12. Pistillate flower. 13. Fruit (Franco et al. 4684).
SYSTEMATIC TREATMENT 123
reflexed in young leaves, the segments (1I-)12-15, the free parts of upper segments oblanceolate (to su- bobovate), the incisions down to ca. 7/10-9/10; apices obtuse to subacute; margin with long, weak, white hairs; upper surface scabrous, densely hispidulous or partly hirtellous to strigose; lower surface (rather) densely puberulous to hirtellous to subtomentose and with (rather) dense brown pluricellular hairs on the main veins, rather sparsely subtomentose on the smaller veins, with arachnoid indumentum very short in the areoles and much longer and loosely covering the areas between the lateral veins, often extending to the lateral veins or the main veins; lateral veins in the free part of the midsegment (25-)30-50 pairs, of- ten >35 pairs, submarginally to marginally loop- connected, most of them unbranched; petiole ca. 30- 70 cm long, puberulous to hirtellous, toward the apex subhirsute; trichilia fused, the brown indumentum in- termixed with dense short and rather long white hairs; stipules 5-12 cm long, white to pinkish, subhirsute to subsericeous to villous and with (very) dense arach- noid indumentum outside, densely (sub)sericeous inside. Staminate inflorescences in pairs, erect; peduncle 6-12 cm long, hirtellous; spathe 4-9 cm long, white to pinkish, hirtellous to subhirsute and with very dense arachnoid indumentum outside, ? densely to sparsely (sub)sericeous inside; spikes 8- 40, (0.8-)2-8 X 0.2-0.5 cm, with stipes 0.2-0.5 mm long and sparsely puberulous to hirtellous or gla- brous; rachis (sparsely) hairy. Staminate flowers with pedicels to 0.2 mm long; perianth tubular, 1-1.5 mm long, puberulous or also with sparse arachnoid indu- mentum below the apex, the apex almost plane, gla- brous; filaments flat; anthers 0.5-0.7 mm long, ap- pendiculate, detached at anthesis, reattached to the margins of the aperture by the appendages at anthesis (?). Pistillate inflorescences in pairs, erect, often sub- tended by caducous bracts, 1-2 cm long; peduncle 2- 6(-9) cm long, puberulous to hirtellous; spathe 4-6 cm long, the color and indumentum as in the stami- nate inflorescence; spikes 4-9(-20), 2-5.5 X 0.4-0.8 cm, to 1O X 1-1.5 cm in fruit and ? curved and often broadened, sessile (or with short stipes). Pistillate flowers: perianth ca. 1.5 mm long, with arachnoid in- dumentum below the apex outside, also below the style channel inside, the apex + convex, muriculate; style short; stigma comose. Fruit ellipsoid to oblon- goid, 1.5-2 mm long, tuberculate.
Distribution (Fig. 8.7). Eastern slopes and foot- hills of the Andes, disjunctly in Colombia (Putu- mayo) and adjacent Ecuador (Sucumbios) and from Ecuador (Morona-Santiago) to Peru (Madre de
Dios), mostly in wet premontane forest, at 400-1200 (-1500) m.
Specimens examined. COLOMBIA. PUTUMAYO: Nr. Mocoa, 18 Feb 1995 (Y fl-fr), Franco et al. 4684 (BG, COL, HUA, US), (d), Franco et al. 4685 (BG, COL, HUA, US); Mun. Mocoa, between La Campucana & San Antonio, 1000 m, 2 May 1994 (5), Franco et al. 5512 (BG), (Y fl-f), Franco et al. 5516 (BG).
ECUADOR. MORONA-SANTIAGO: 10 km W of Rio Za- mora & 2 km S of Rio Bomboiza, nr. Misi6n Salesiana, 800 m, 13 Mar 1986 (cT fl-fr), Baker 6724 (BG, QAME, QCNE); rd. Zamora-Gualaquiza, between Yantzatza & Los Encuen- tros, ca. 900 m, 3 Jan 1991 (5), Berg etal. 1656 (BG, LOJA, QCA); rd. Zamora-Gualaquiza, Estaci6n Exp. El Padmi, ca. 5 km N of Los Encuentros, ca. 900 m, 4 Jan 1991 (Y fl-fr), Berg et al. 1658 (BG, LOJA, NY, QCA); rd. Gualaquiza- San Juan Bosco, nr. Gualaquiza, 16 Feb 1994 (5), Berg et al. 1691 (BG, QCNE). SUCUMBiOS: Lumbaqui, 1000 m, 13 Aug 1975 (Y fl), Little et al. 1681 (MO, Q, QAME, QCNE). ZAMORA-CHINCHIPE: Nr. Zamora, ca. 1000 m, 4 Jan 1991 (5), Berg et al. 1661 (BG, GB, LOJA, NY, QCA), (Y fl-fr), Berg et al. 1662 (BG, LOJA, QCA); Pachicutza, 70 km NE of Zamora, 1000 m, 14 Sep 1975 (? fl-fr), Little et al. 347 (COL, LOJA, Q, QAME); Cant6n Nangaritza, Rfo Nangar- itza, Pachicutza, 900 m, 12 Dec 1990 ( Y fl), Neill et al. 9704 (BG, MO, QCA, QCNE); Cant6n Nangaritza, Yanzatza, Rio Yacuambi, 17 Oct 1991 (5), Palacios 8178 (BG, QCNE).
PERU. HUANUCO: Rd. Tingo Maria-Aucayacu, Puma- huasi, 650 m, 25 Nov 1997 (9 fl-fr), Berg et al. 1742 (BG, COL, MOL); rd. Tingo Maria-Aucayacu, nr. Milagro, 650 m, 25 Nov 1997 (5), Berg et al. 1743 (BG, COL, MOL); 10 km S of Nuevo San Martin, 60 km S of Shumanza, 550 m, 16 Jul 1982 (9 fl-fr), Gentry et al. 37566 (BG, MO); Prov. Leoncio Prado, Dtto. Rupa Rupa, E of Tingo Maria, Cerro Quemado, 700-800 m, 13 Sep 1978 (9 fl), Schunke V 10595 (BG, MO). JUNIN: Prov. Chanchamayo, Catarata del Tirol, 3 Feb 1999 (9 fr), Nuiiiez I. 87 (BG). MADRE DE
Dios: Manu National Park, Yomybato vill., Aug 1997 (5), Yu et al. 1173 (BG). PASCO: Valley of Rfo Palcazu, 300-600 m, 7 Dec 1984 (5), Hartshorn et al. 2694 (F, MO); Prov. Oxapampa, Gran Pajonal, trail Chequitavo- Shumahuani, 1200 m, 24 Sep 1983 (9 fl), D. N. Smith 5225 (BG, MO). SAN MARTiN: Prov. Lamas, rd. Lamas-Pa- mashto, ca. 800 m, 4 Dec 1997 (5), Berg et al. 1768 (BG, COL, MOL); Prov. San Martin, rd. Tarapoto-Yurimaguas, ca. km 13, ca. 700 m, 5 Dec 1997 (5), Berg et al. 1773 (BG, COL, MOL); Prov. Lamas, rd. Tarapoto-Moyobamba, ca. km 62, ca. 1000 m, 8 Dec 1997 (9 fl-fr), Berg et al. 1788 (BG, COL, MOL); nr. Moyobamba, 600-700 m, 18 Dec 1971 (5), Ferreyra 17878 (US); Rio Huallaga valley, between Puerto Pinzana & Punta Arenas (Shumanza), 4 Feb 1984 (9 fl-fr), Gentry et al. 44925 (BG, MO, NY); Prov. Lamas, rd. Tarapoto-Moyobamba, km 55, 700 m, 10 Oct 1984 (d), Maas et al. 6035 (BG, U); Prov. Moyobamba, Dtto. Calzada, Morro Calzada, 1050-1350 m, 11 Nov 196 (9 fl-fr), Sdnchez V et al. 8611 (BG). UCAYALI: Rd. Pu- callpa-Tingo Maria, km 135, ca. 400 m, 26 Nov 1997 (5), Berg et al. 1749 (BG, COL, MOL).
124 FLORA NEOTROPICA
Cecropia montana is probably closely related to C. angustifolia. It has also inflorescences with short peduncles and spikes and the lamina is also (usually) reflexed in the bud. It can be distinguished from C. angustifolia by the dense arachnoid indumentum on the stipules (and spathes), by more numerous lateral veins in the free part of the midsegment of the lamina, usually 30-50 pairs, whereas C. angustifolia (in the non-villous type) normally has at most 30 pairs, and usually also by the long white hairs intermixed with the brown indumentum of the trichilia (a feature not found in all specimens of C. montana, and rarely in C. angustifolia, e.g., in Berg 1233 and Franco et al. 5515). Moreover, the length of the stipules can be used to distinguish the two species (especially in the northern part of the range of distribution of C. mon- tana): to 12 cm long in C. montana and often >12 cm long in C. angustifolia. Intermediates between the two species have not been encountered. Cecropia montana is ecologically different from C. angusti- folia, occurring at lower elevations.
34. Cecropia multisecta P. Franco & C. C. Berg, Cal- dasia 23(1): 77, t. 1. 2001. Type. Colombia. An- tioquia: Mun. Anon', rd. Anorf-Medellin, between Vrda. El Roble & Anon', 1600 m, 8 Mar 1995 (Y), Franco et al. 5573 (holotype: COL; isotypes: BG, COL, HUA). Fig. 35
Tree, to 13 m tall. Leafy twigs 1-4.5 cm thick, dark green to brown or white to grayish, sparsely to densely puberulous to hirtellous to subhispid(ulous), also with rather sparse to dense arachnoid indumen- tum and sparse to dense brown pluricellular hairs. Lamina coriaceous, ca. 25 X 25 cm to 65 X 65 cm, the segments 10-12, (ob)lanceolate, the incisions down to the petiole; apices obtuse to acute to shortly acuminate; upper surface smooth, sparsely hirtellous to strigillose (on the veins); lower surface with ? dense brown pluricellular hairs on the veins or also sparsely puberulous on the main veins, with arach- noid indumentum in the areoles; lateral veins in the free part of the midsegment ca. 20-25, submarginally loop-connected, unbranched; petiole ca. 25-55 cm long, with dense arachnoid indumentum; trichilia sep- arate or fused, the brown indumentum intermixed with rather short whitish to brownish (unicellular) hairs; stipules ca. 15-27 cm long, pink to yellowish, sparsely to densely hirtellous to subsericeous and with dense brown pluricellular hairs outside, glabrous or toward the apex subsericeous inside. Staminate in- florescences in pairs, the peduncle erect, the spikes erect to ? spreading; peduncle 5-6 cm long, hirtel-
lous to puberulous and with dense to rather sparse arachnoid indumentum; spathe ca. 12-13 cm long, pink to greenish, + densely puberulous to hirtellous outside, glabrous inside; spikes ca. 9-12, ca. 10-12 X 0.5 cm, with stipes 0.2-0.3 mm long and with brown pluricellular hairs; rachis short-hairy. Stami- nate flowers: perianth tubular, 1.8-2.2 mm long, sparsely and minutely puberulous below the apex, the apex slightly convex; filaments flat; anthers 0.8-1 mm long, appendiculate. Pistillate inflorescences in pairs, erect to pendulous; peduncle 3-6 cm long, hirtellous to puberulous, and with + dense arachnoid indumen- tum; spathe ca. 14-15 cm long, pink, densely puber- ulous to subsericeous outside, glabrous inside; spikes 2-3(-6), ca. 11-12 X 1-1.3 cm, to 23 X 2 cm in fruit, sessile; rachis short-hairy. Pistillateflowers: perianth 1-1.5 mm long, with arachnoid indumentum below the apex outside, also in the lower part of the style channel inside, the apex convex, punctate to muricu- late or around the aperture minutely puberulous; style rather short, minutely puberulous; stigma comose. Fruit ellipsoid to oblongoid, ca. 2.5-3 mm long, tu- berculate.
Distribution (Fig. 11.3). Colombia (Antioquia), in forest, at 1300-1600 m.
Specimens examined. COLOMBIA. ANTIOQUIA: Mun. San Rafael, nr. campamento Versalles, 1350 m, 23 Mar 1994 (Y fl-fr), Franco et al. 4574 (BG, HUA), (d), Franco et al. 4575 (BG, HUA); Mun. Anorf, rd. Anori-Medellin, between Vrda. El Roble & Anorf, ca. 1600 m, 8 Mar 1995 (Y fl-fr), Franco et al. 5569 (BG, COL, HUA), (d), Franco et al. 5572 (BG, COL, HUA).
This species shows affinities to Cecropia plicata, from which it clearly differs by the incisions of the lamina extending down to the petiole.
35. Cecropia mutisiana Mildbraed, in Cuatrecasas, Trab. Mus. Ci. Nat. (Madrid), Bot. Ser. 26: 29. 1933. Type. Colombia. Cundinamarca: La Esper- anza, 1500 m, 18 Apr 1932 (?), Cuatrecasas 3082 (holotype: MA-n.v., photograph in COL).
Cecropia tolimensis Cuatrecasas, Revista Acad. Colomb. Ci. Exact. 6(22/23): 282. 1945. Type. Colombia. To- lima: Between Fresno & Mariquita, 1150 m, 7 May 1940 (d), Cuatrecasas 9397 (holotype: COL; iso- type: F).
SYSTEMATIC TREATMENT 125
1998~~~~~~~~
FIG. 35. Cecropia multisecta. 1. Lamina, reduced. 2. Apex of lamina and venation. 3. Stipules and bases of petioles with trichilia. 4. Pair of staminate inflorescences at anthesis and base of petiole with trichilium. 5. Staminate flower and stamen (Franco et al. 4575). 6. Pair of pistillate inflorescence at anthesis and base of petiole with trichilium (Franco et al. 4574). 7. Pistillate flower. 8. Fruit (Franco et al. 5569). (From Caldasia 23: 78. 2001.)
126 FLORA NEOTROPICA
Tree, to 12(-20) m tall. Leafy twigs 2.5-4 cm thick, hispidulous to puberulous with straight to un- cinate hairs. Lamina chartaceous to subcoriaceous, ca. 35 X 35 cm to 75 X 75 cm, the segments (9-)10- 12(-13), the free parts of the upper segments oblan- ceolate to (sub)obovate to elliptic, the incisions down to ca. 7/10-8/10; apices obtuse to rounded; upper sur- face scabrous to scabridulous, hispidulous or (ini- tially) also with arachnoid indumentum; lower surface puberulous to hirtellous to subtomentose on the veins, with arachnoid indumentum in the areoles or extend- ing to the main veins; lateral veins in the free part of the midsegment 10-21, submarginally (to margin- ally), loop-connected (the lower ones often faintly so), unbranched or branched; petiole ca. 20-60 cm long, puberulous to subhispidulous or (initially) also with arachnoid indumentum; trichilia fused, the upper rim often irregularly lobate, the brown indumentum in- termixed with short white hairs; stipules 6-23 cm long, dark red to red-brown, hirtellous to puberulous or also with arachnoid indumentum outside, (partly) white sericeous inside. Staminate inflorescences in pairs, the peduncle erect and the spikes pendulous (or spreading); peduncle 5-10 cm long, hispidulous; spathe 8-18 cm long, (red-)brown, sparsely to rather densely hirtellous to puberulous outside, densely yel- lowish subsericeous inside; spikes ca. 15-25, 4.5-14 X 0.2-0.3 cm, with stipes 0.4-1 cm long and sparsely hairy; rachis hairy. Staminate flowers: perianth tu- bular, ca. 1 mm long, sparsely and minutely puberu- lous below the apex, the apex plane; filaments flat; anthers ca. 0.4 mm long, appendiculate, detached at anthesis, reattached to the margins of the aperture by the appendages (?). Pistillate inflorescences in pairs or solitary, erect, soon deflexed to pendulous; pedun- cle 3-7 cm long, hispidulous or also with arachnoid indumentum; spathe 10-13 cm long, the color and indumentum as in the staminate inflorescence; spikes (2-)4-5, 6-12 X 0.4-0.6 cm, to 31 X 1.3 cm in fruit, sessile; rachis hairy. Pistillate flowers: perianth ca. 1.5-2 mm long, with arachnoid indumentum below the apex outside, also in the style channel inside, the apex convex, minutely puberulous to muriculate; style short; stigmas comose. Fruit ellipsoid, ca. 1.5 mm long, smooth, dark brown.
Distribution (Fig. 8.3). Colombia, upper Magda- lena valley, in forest and secondary growth, normally at 800-1800 m, but also found as low as 500 m, or in Caldas even down to 200 m.
Representative specimens examined. COLOMBIA. CALDAS: Nr. La Dorada, 1-20 Feb 1946 (9 fl), Duque- Jaramillo 2590 (COL, NY). CAUCA: Rd. Ricaurte-Guadu- alejo, 1165-1270 m, 19 Jul 1993 (i), Agudelo et al. 3100
(COL); rd. Guadualejo-Inza, 1270-1810 m, 19 Jul 1993 (9 fl-fr), Agudelo et al. 3114 (COL). CUNDINAMARCA: S of Silvania, 7 km on rd. to Tibacuy from km 37 of old rd. to Fusagasuga, 1500 m, 15 Jun 1972 (6), Barclay et al. 3506 (COL, US); Mun. San Bemardo, Vrda. Portones, 1650 m, 28 Jul 1981 (d fl-fr), Diaz P 3263 (COL, MO); Mun. Pueblo Nuevo, rd. to San Antonio, 15 Mar 1986 (9 fl-fr), Franco et al. 2409 (COL); Zipac6n, El Ocaso, rd. to Cartagena La Mesa, 14 Mar 1988 (d), Franco et al. 2441 (BG, COL); Mun. La Magola, 1600 m, 11 Apr 1988 (6), Franco et al. 2483 (COL, MO), (9 fl-fr), Franco et al. 2485 (COL, MO); Mun. Guaduas, Alto La Mona, 1250 m, 18 May 1990 (9 fl- fr), Franco et al. 2973 (COL), (6), Franco et al. 2974 (COL); Silvania, at km 62, 1520 m, 12 Mar 1993 (6), Franco et al. 4344 (COL); Mun. Melgar, Rio Sumapaz, nr. La Nariz del Diablo, 12 Mar 1993 (9 fl), Franco et al. 4345 (COL, MO); Mun. Tena, rd. Bogota-La Mesa, 1650 m, 6 Feb 1995 (6), Franco et al. 4644 (BG, COL; HUA, MO, US); Mun. Guaduas, 10 km from Guaduas, 12 Feb 1985 (9 fl), Galeano et al. 512 (COL), (6), Galeano et al. 513 (COL); Mun. San Francisco, Vrda. El Pifion, 25 Feb 1985 (9 fl-fr), Galeano et al. 552 (COL); rd. Caquera-Puente Quetame, km 68, Puente Quebrada Blanca, 13 Mar 1985 (9 f-fr), Galeano et al. 645a (COL); La Esperanza, RR to Gi- rardot, 1300 m, 23 Jul 1934 (9 fr), Garcia-Barriga 4692 (COL, F, UDBC, US); rd. Fusagasuga-Silvania, 1680 m, 21 Mar 1970 (9 fl-fr), Idrobo 6319 (COL), (6), Idrobo 6320 (COL); Mun. Bituima, 1400 m, 6 Mar 1972 (6 + 9 fr), Pinz6n-Rodrfguez s.n. (COL, US); nr. Anolaima, ca. 1500 m, 25 Jul 1963 (9 fl-fr), Soejarto et al. 112 (GH, K). HUILA: Mun. Garz6n, Vrda. Las Quebraditas, 21-26 Feb 1947 (9), Bermtidez 34923 (F); between Gigante & Rioloro, 19 Mar 1940 (9), Cuatrecasas 8334 (COL, F); Guadalupe, below Gabinete, 1700 m, 16 Oct 1993 (6), Franco et al. 4505 (BG), ( 9 fr), Franco et al. 4506 (BG); Caucho, E of Palacio, ca. 20 km NE of Santa Ana, ca. 1800 m, 24 Feb 1944 (6), Little 7343 (COL, P, US); 15 km NE of Algeciras, ca. 1550 m, 1 Apr 1944 (9 fr), Little 7562 (COL, F, US); 3 km SW of Acevedo, ca. 1450 m, 15 Aug 1944 (9), Little 8478 (COL, F, US); Mun. Colombia, Cerro Bellavista, 1340 m, 15 Apr 1989 (9 fl-fr), Llanos et al. 1372 (COL); between Gigante & Rioloro, 19 Mar 1940 (9 fl-fr), Perez Arbeldez & Cuatrecasas 8334 (COL); beyond Guadalupe, 1000-1300 m, 20 Mar 1940 (9), Perez Arbeldez & Cuatrecasas 8402 (F); Pitalito, Calamo, 1300 m, 30 Dec 1942 (d), Schultes et al. 5128 (GH, NY). QUINDIO: Mun. Quimbaya, Vrda. Pa- lermo, 23 Nov 1989 (d), Velez et al. 724 (COL). TOLIMA: Mun. Mariquita, rd. Mariquita-Honda, 17 May 1990 (9 fl), Franco et al. 2970 (COL, MO), 2973 (COL); without precise locality, 1000 m, 6 Jan 1882 (6), Lehmann 2341 (G).
The species is probably related to Cecropia obtu- sifolia. The general aspect of Franco at al. 4345, col- lected at 510 m, is that of C. peltata, but the flowers match those of C. mutisiana, although the stigma is relatively large due to long papillae.
Tree, to 12(-25) m tall. Leafy twigs 1-5 cm thick, green, hispidulous with curved hairs. Lamina charta- ceous to (sub)coriaceous, ca. 25 X 25 cm to 60 X 60 cm, the segments 6-8(-9), the free parts of the upper segments usually obovate to subobovate, sometimes to subspathulate, the incisions down to 5/10-7/10(-8/ 10); apices obtuse to subacute (short-acuminate); up- per surface ? scabrous or smooth, (very) sparsely to rather densely hispidulous and with ? dense arach- noid indumentum; lower surface puberulous to pu- bescent on the veins, with arachnoid indumentum in the areoles, extending to the main veins; lateral veins in the free part of the midsegment 9-15 pairs, sub- marginally loop-connected, unbranched or faintly branched; petiole 15-50 cm long, puberulous to pu- bescent and with ? dense (persistent) arachnoid indumentum; trichilia fused, the brown indumentum intermixed with short and/or rather long white hairs; stipules 7-12(-20) cm long, dark to bright red or grayish when covered by dense arachnoid indumen- tum, puberulous to hirtellous to strigillose and usually also with sparse to dense arachnoid indumentum out- side, (sub)sericeous inside. Staminate inflorescences in pairs, the peduncle erect and the spikes pendulous; peduncle 4-8.5 cm long, hispidulous or partly pubes- cent; spathe 8-17 cm long, red-brown to yellowish or whitish, puberulous to hirtellous and with sparse to dense arachnoid indumentum outside, glabrous in- side; spikes 15-26, 4-13 X 0.2-0.3 cm, with stipes 0.5-1.5 cm long and densely minutely puberulous; rachis hairy. Staminate flowers: perianth 1-1.5 mm long, minutely puberulous with straight hairs or also with arachnoid indumentum below the margin, the apex plane to slightly convex or truncate, muriculate, the aperture often surrounded by a rim; filaments slightly swollen; anthers 0.5-0.7 mm long, appendi- culate, detached at anthesis, reattached to the margins of the aperture by the appendages (?). Pistillate info- rescences in pairs, erect; peduncle 6.5-8(-14) cm long, hispidulous or partly pubescent; spathe 8-12 cm long, the color and indumentum as in the staminate inflorescence; spikes 4, 3-10 X ca. 8 cm, to 16 X 1.5 cm in fruit, sessile or with stipes to 0.8 cm long and
minutely puberulous or also with arachnoid indumen- tum; rachis hairy. Pistillate flowers: perianth ca. 2 mm long, with arachnoid indumentum below the apex outside, also below the style channel inside, the apex convex, densely muriculate; style rather long, straight; stigma comose. Fruit oblongoid to ellipsoid, ca. 2.5 mm long, ? tuberculate.
Distribution (Fig. 8.4). Guianas and the lower Amazon basin, in forest and secondary growth, at low elevations.
Representative specimens examined. GUYANA. Ru- pununi Distr., between Kuyuwini Landing & Kassikaityu R., 18 Oct 1992 (Y fl-fr), Jansen-Jacobs et al. 2997 (BG, U); Mazaruni R., Kamarang, 21 Aug 1977 (Y fl-fr), Maas et al. 2594 (NY, U); S of Temehri, 13 Sep 1979 (Y fr), Maas et al. 3525 (K, NY, U); Rupununi Distr., foothills NW of Ka- nuku Mtns., nr. Moco-Moco, 27 Oct 1979 (c), Maas et al. 3848 (MO, NY, U); base of Mt. Makarapan, Makarapan Cr., 17 Sep 1988 (Y fl-fr), Maas et al. 7501 (K, NY, U); Marudi Mtns., Mazoa Hill, 13 Nov 1982 (d), Stoffers et al. 339 (BG, K, NY, S, U).
FRENCH GUIANA. Charvein, 20 Nov 1913 (6), Be- noist 279 (P); Saul, Monts La Fumee, 1982 (9 fl-fr), Boom et al. 2086 (NY), 19 Oct 1982 (d), Boom et al. 2162 (NY); nr. Cayenne, 27 Jun 1921 (9 fl), Broadway 582 (GH, NY, US); Lower Oyapock basin, Savane Roche du Quatorze Juil- let, 19 Apr 1991 (st), Cremers 12330 (CAY); Region Paul Isnard, Camp Voltaire, 28 Oct 1991 (9 fl), Cremers et al. 12490 (BG); Regina region, Mt. Tortue, 11 km WNW of Approuague R., 18 Jun 1988 (6), Feuillet 10313 (BG); Cay- enne, Montabo, Mar 1978 (9 fr), Grenand 1542 (U); Piste de St. Elie, 7 Jan 1986 (9 fl-fr), Lauri 187 (BG); nr. Saul, 11 Aug 1987 (9 fl), Mori et al. 18706 (NY), 15 Sep 1989 (6), Mori et al. 20907 (BG, NY); Crique de la Folie, 23 Jul 1965 (9 fl-fr), Oldeman 1486 (P); piste de St. Elie, 19 Jul 1979 (J), Prevost 706 (P, U); Cayenne, Centre ORSTOM, 17 Aug 1979 (9 fl), Prevost 732 (P, U); Oyapock R., Zidock, 11 Sep 1980 (9 fl), Prevost et al. 1009 (P, U); Acarouany, 1857 (d), Sagot 515 (BM, G, P, S, U, US); Saint Laurent de Maroni, 12 Aug 1948 (d), Service des Eaux et Forets 4070 (P).
BRAZIL. AMAPA: Mun. Mazagao, Rio Jari, Morros do Felipe, 10 Oct 1987 (9 fr), H. T Beck et al. 32 (BG, INPA, NY); Rio Oiapoque, Boa Esperanqa, opposite mouth of Ca- mopi R., 18 Aug 1960 (d), Egler 47658 (COL, GH, K, MG,
128 FLORA NEOTROPICA
NY, S, US); Rio Oiapoque, between Oiapoque & Santo An- tonio, 26 Jul 1960 (9 fl-fr), Irwin et al. 47159 (BG, K, MG, NY, US); Rio Araguari, ca. 1?40'N, 51?56'W, 28 Aug 1961 (9 fl), Pires et al. 50533 (MG, MO, NY, U, US); P6rto Platon, 13 Oct 1976 (d), Ribeiro 1502 (INPA, MG, MO, NY, RB, U); Rio Oiapoque, Cachoeira Camaraua, 3 km S of mouth of Camopi R., 29 Sep 1960 (9 fl-fr), Westra 48523 (BG, MG, NY). MARANHAO: Mun. Carutapera, Gurupiuna, 3 Nov 1986 (st), Balee et al. 2783 (BG, NY). PARA: Mun. Altamira, Igarape Ipixuna, 4 Nov 1985 (st), Balee et al. 1830 (BG); Serra dos Carajas, Serra Norte, ca. 20 km N of AMZA Exploration Camp, 16 Oct 1977 (9 fl-fr), Berg et al. 585 (A, AAU, COL, MG, NY, RB, U); Belem, grounds of IAN, 7 Nov 1962 (9 fl-fr), Cuatrecasas 26654 (P, US); Rio Jar- amacarii, 1 Jun 1957 (9 fl), Egler 437 (MG); Belem, Santa Izabel, Oct 1906 (9 fl-fr), Goeldi (MG) 7727 (BM, G, MG, S, U); Tucuruf, 20 Oct 1983 (9 fl-fr), E. Lima et al. 64 (INPA, NY); rd. Bel6m-Braslfia, km 92, 24 Sep 1959 (9 fr), M. Kuhlmann et al. 284 (MG, SP, US); Mun. Altamei- rim, Monte Dourado, 10 Oct 1986 (d), Pires et al. 1425 (BG, MG); rd. BR.22, nr. Cachoeira, 27 Oct 1965 (9 fl-fr), Prance et al. 1700 (GH, K, MICH, MO, NY, S); rd. BR.163, Cuiaba-Santar6m, km 1223, nr. Igarape Kazuo, 18 Nov 1977 (d), Prance et al. 25548 (K, MG, NY, U, US), (9 fl- fr), Prance et al. 25549 (K, MG, MO, NY, U, US); rd. Cu- iaba-Santarem (BR.163), km 1300, nr. Igarape Jose Preto, 23 Nov 1977 (st), Prance et al. 25708 (MG, NY, U); Rio Jarn, Monte Dourado, 11 Oct 1968 (9 fl-fr), N. T Silva 1178 (NY, U, US), 30 Oct 1968 (9 fl-fr), N. T Silva 1340 (NY, U): Mun. Almeirim, 18 Sep 1984 (d), N. T Silva 5393 (INPA, MG); rd. Altamira-Maraba, km 19, 30 Aug 1974 (st), N. Smith B.38 (INPA); Serra de Carajas, AMZA camp, 8 Jun 1982 (9 fl-fr), Sperling et al. 5966 (MG, NY).
The species appears to be closely related to the allopatric Cecropia ficifolia. The two species have many features in commmon. Cecropia obtusa differs from C. ficifolia, e.g., in the larger number of spikes in the staminate inflorescence, the shorter peduncle of the pistillate inflorescence, and the shorter stipules. If the lamina is deeply incised (5/10), then the free part of the midsegment is ? abruptly narrowed and thus + distinctly spathulate, whereas usually obovate to subobovate and rarely subspathulate in C. obtusa. In C. obtusa, the petiole is covered with persistent arach- noid indumentum. When such indumentum occurs in C. ficifolia (as in Ecuador and Colombia), it is (often) in combination with hirsute indumentum. Whereas C. ficifolia is a weedy species, at least in part of its range of distribution, C. obtusa is associated with forest habitats.
The following collection is difficult to interpret: Serra do Navio, Rio Amapari, 12 Nov 1954 (9 fr), Cowan 38280 (NY, US). The label indicates that it is made from a tree 5 m tall, unlikely for this species to be already in fruit; the inflorescences are separate from the partial leaf, which has narrow segments with
up to ca. 25 pairs of lateral veins in the free part of the midsegment, and the arachnoid indumentum is dense in the petiole but sparse on the lamina and ab- sent in the areoles beneath. It is difficult to relate the leaves to the juvenile state of C. obtusa.
The name Cecropia obtusa was based on four col- lections: the present lectotype, Bonpland 1295 from Cuba (= C. schreberiana subsp. antillarum), Gardner 3981 (= C. saxatilis), and Ruiz & Pavon s.n. (= C. polystachya).
Vernacular names. Guyana: congo pump, trum- pet tree. French Guiana: ama'i (Wayapi); bois canon, kulegle (Wayana). Brazil: ama-'y-tuwir (Ka'apor, Maranhao); imbauba branca (Maranhao); ama'i-hete (Tupi, Para); imbauiba branca (Para).
37. Cecropia obtusifolia Bertoloni, Fl. Guatimal. 39. 1840; Woodson & Schery, Ann. Missouri Bot. Gard. 47: 175, t. 61. 1960; Burger, Fieldiana, Bot. 40: 125, t. 23. 1977. Type. Guatemala. Without locality, J. Veldsquez s.n. (BOLO-n.v., photo- graphs in BM, K, MICH, MO). Fig. 36
Cecropia commutata Schott ex Miquel, in Martius Fl. Bras. 4(1): 148. 1853. Type. Material cultivated in Hortus Schonbrunn, from material collected in Mex- ico by Schott in 1852 (holotype: M, photographs in MICH, MO).
Cecropia panamensis Hemsley, Biol. Centr. Amer. Bot. 3: 151. 1883. Type. Panama. Nr. Panama City, Mar 1847 (5), Seemann 499 (holotype: K).
Ambaiba costaricensis Kuntze, Revis. Gen. P1. 2: 623. 1891. Type. Costa Rica. Not designated, nor traced.
Ambaiba hemsleyana Kuntze, Revis. Gen. P1. 2: 623. 1891. Type not designated, nor traced.
Cecropia radlkoferiana V. Richter, Biblioth. Bot. 43: 17. 1897. Type. Colombia. Choc6: Rio San Juan, Triana 865 (P, mixed collection, the pistillate inflorescence designated as the lectotype, Berg & Franco Rosselli, Fl. Ecuador 48: 41. 1993; isolectotype: K).
Cecropia levyana Bureau & V. Richter, Biblioth. Bot. 43: 20, t. 5, 1897. Type. Nicaragua. Chontales, Jun 1870 (5), Levy 473 (holotype: P, fragment in F; isotypes: G, P).
Cecropia mexicana Hemsley var. macrostachya Donnell Smith, Bot. Gaz. (Crawfordsville) 27: 442. 1899. Syntypes: Costa Rica. Laguna de Buenos Aires, Feb 1892 (9 fl-fr), Pittier 6666 (US); Cartago: Atirro, Mar 1894 (9), Donnell Smith 4934 (GH, K, US), Rio Poas, 1200 m, Mar 1896 (9), Donnell Smith
SYSTEMATIC TREATMENT 129
(< :1~~~~~~~~~~~~~~~~~~/ FIG. 3s 1.
,7
4 ~ ~ ~ ~ 1
.4~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~. .4
4~~~~~~~~~~~~~~~~~~~~~.
FIG. 36. Cecropia obtusifolia. 1. Lamina, reduced. 2. Apex of lamina and venation. 3. Leafy twig with stipules, young leaf, and base of petiole with trichilium (Berg 1242). 4. Staminate inflorescence and base of petiole with trichilium
(Berg 1540). 5. Staminate flower and stamen (Berg 1541). 6. Pistillate inflorescence with spathe and base of petiole with trichilium. 7. Pistillate inflorescence at anthesis and base of petiole with trichilium (Berg 1242). 8. Pistillate flower. 9. Fruit
6771 (K, US); La Concepci6n, Llanuras de Santa Clara, Feb 1896 (Y), Donnell Smith 6774 (GH, K, US).
Cecropia maxonii Pittier, Contr. U.S. Natl. Herb. 18: 228. 1917. Type. Panama. Chiriquf: Nr. El Boquete, 1000-1300 m, 2-8 Mar 1911 (d), Maxon 5132 (ho- lotype: US).
Cecropia amphichlora Standley & L. 0. Williams, Ceiba 3: 111. 1952. Type. Honduras. Cortes: La Cumbre, Sierra de Omoa, 30 Nov 1950, Molina R. 3455 (US, mixed collection, the leaf here designated as lecto- type; the d inflorescence is C. peltata).
Cecropia dabeibana Cuatrecasas, Revista Acad. Col- omb. Ci. Exact. 6(21): 67. 1944. Type. Colombia. Antioquia: Between Dabeiba & Llanitos, Rio Sucio, 25 Mar-I Feb 1942 (Y), Metcalf & Cuatrecasas 30168 (holotype: COL; isotypes: F, US).
Cecropia burriada Cuatrecasas, Revista Acad. Colomb. Ci. Exact. 6(22/23): 274. 1945. Cecropia obtusifolia Bertoloni subsp. burriada (Cuatrecasas) C. C. Berg & P. Franco, Fl. Ecuador 48: 41. 1993. Type. Colom- bia. Valle: Rio Calima, La Trojita, 26 Feb 1944 ( ), Cuatrecasas 16505 (holotype: COL; isotypes: F, US, VALLE).
thick, green or partly purplish, hirtellous to subhis- pidulous with straight and/or uncinate hairs. Lamina chartaceous to subcoriaceous, ca. (15 X 15 cm to) 30 X 30 cm to 90 X 90 cm, the segments (8-)10-13 or 7-1 0, the free parts of the upper segments subobovate to oblanceolate (to elliptic or obovate), the incisions down to ca. 5/10-8/10; apices obtuse to rounded or acuminate; upper surface ? scabrous, hispidulous; lower surface puberulous to hirtellous to subtomen- tose on the veins, with arachnoid indumentum in the areoles, also on the reticulum or (almost) lacking; lat- eral veins in the free part of the midsegment (10-) 15-25(-50), marginally to submarginally loop- connected, often branched; petiole (lO-)20-80 cm long, puberulous to hirtellous and/or with arachnoid indumentum, or subglabrous; trichilia fused, the brown indumentum intermixed with short white hairs; stipules 5-15 cm long, (maroon-)red or yellowish, (sparsely) puberulous to hirtellous to hirsute to sub- sericeous or also with arachnoid indumentum outside, sericeous to subvillous or to subglabrous inside. Sta- minate inflorescences in pairs (or solitary), the pe- duncle erect to pendulous, the spikes erect to spread- ing to pendulous; peduncle 4-18 cm long, (minutely) puberulous to subglabrous; spathe 5-20 cm long, red
to red-brown to pink or to purple or yellowish with reddish stripes and spots (or whitish), (sparsely) hir- tellous to subhirsute, often also with sparse (or dense) arachnoid indumentum, or subglabrous (and then sometimes with a bluish waxy layer) outside, gla- brous or sericeous inside; spikes 8-18(-24), 4-25 X 0.2-0.4(-0.8) cm, sessile or with stipes to 2.5(-10) cm long and puberulous; rachis sparsely hairy or gla- brous. Staminateflowers: perianth tubular, 1-1.5 mm long, usually with short hairs below the apex, some- times glabrous, the apex plane, the aperture often sur- rounded by a rim; filaments flat; anthers 0.5-0.7 mm long, appendiculate, detached at anthesis, reattached to the margins of the aperture by the appendages. Pis- tillate inflorescences in pairs (or solitary), erect or at least the spikes pendulous; peduncle 3-24 cm long, (minutely) puberulous to subglabrous; spathe 12-22 cm long, the color and indumentum as in the stami- nate inflorescence; spikes 2-4(-6), (4-)8-25 X 0.4- 0.7 cm, to 35(-55) X 1.5 cm in fruit, sessile or with stipes to 1.5(4) cm long and puberulous; rachis hairy. Pistillate flowers: perianth 1.5-2 mm long, with arachnoid indumentum below the apex, also in the lower part of the style channel inside, the apex convex, muriculate to minutely puberulous or smooth; style rather short; stigma penicillate. Fruit ellipsoid, 2-2.5 mm long, smooth, dark brown.
Distribution (Fig. 8.5). From Mexico to the Pa- cific coastal region, southward to Ecuador, in forest and secondary growth, at elevations to 1300(-1650) m.
Representative specimens examined. MEXICO. CHIAPAS: 5 mi N of Tapiula, 10 Jul 1966 (d), Baird 1029 (US); Mun. Villa Corzo, 10-12 km SW of Colonia Agr6n- omos Mexicanos, Cerro Tres Picos, 1000 m, 8 Feb 1972 (Y fr), Breedlove 23943 (MO); Mun. Huixtla, 6-8 km NE of Huixtla, 27 Dec 1972 (6), Breedlove et al. 30939 (MO); Mun. Ococingo, Centro Arqueol6gico Bonampak, 19 Feb 1982 (6), Castillo 265 (MO); Escuintla, Esperanza, 13 Feb 1945 (6), Matuda 16411 (EAP, MICH, NY); Mun. Angel Albino Corzo, Rio Cuztepeques, 26 Mar 1968 (? fl-fr), Shi- lom Ton 3894 (LL, MICH); Mun. Tuxtla Chico, Curva de Cahoacan, rd. to Talsiman, 3 Jun 1992 (6), Vazquez 176 (MEXU); Mun. Tapachula, El Porvenir, 21 Nov 1984 (6), Ventura et al. 729 (MEXU); Mun. Cacahoatan, Cuatimoc, 14 Nov 1985 (6), Ventura et al. 2705 (MO). GUERRERO:
Nr. Puerto Marquez, 10 Aug 1966 (6), Baird 1041 (US); Puerto Marquez, 20 May 1979 (st), L6pez F 788 (MEXU); Mun. Atoyac, 27 km NE of Atoyac de Alvarez, rd. to Par- aiso, 28 Mar 1983 (Y fl-fr), Soto N. et al. 5064 (MEXU, MO). HIDALGO: 4 km SE of San Bartolo, Tutitepec, 3 Jun 1972 (Y fl-fr), Gimate L. 642 (AAU, COL, EAP, MICH, PMA, US); Dtto. Jacala, Cantil de Espejo, nr. km 340-341 on hwy. below Chapulhuacan, 30 Jan 1948 (Y fl-fr), Moore
SYSTEMATIC TREATMENT 131
et al. 3642 (A, MICH, TEX). JALISCO: 7 km W of Villa de Purificaci6n, 10 Dec 1988 (st), Bullock 2022 (MEXU); 0.1 km SW of rd. Barra de Navidad-Puerto Vallarta, on rd. to Manzanilla, 24 Dec 1983 (Y fl), Lott et al. 2213 (MEXU); rd. to Mazatlan, ca. 19 mi NW of Tepic, 15 Nov 1959 (d), McVaugh et al. 717 (MICH); 15-18 rd.-mi SW of Autlan, 9 Apr 1951 (6), MceVaugh 11957 (MICH); Santa Cruz de Val- larta, 10 Dec 1926 (c), Mexia 1275 (A, BM, G, MICH, MO, NY, US). MEXICO: 3 km S of San Miguel Amatepec, 20 Jan 1973 (c3'), Gonzalez M. et al. 5180 (MEXU); Temascaltepec, Anonas, 3 Jul 1933 (Y fl-fr), Hinton 3568 (G, GH, K, NY). NAYARIT: Ca. 12 km E of San Blas, 27 Apr 1962 (d + Y fl-fr), Barr et al. 62-305 (MEXU); rd. Puerto Vallarta-Tepic, km 103, 22 Apr 1980 (i), Rooden 751 (U), (9 fl-fr), Rooden 752 (U); 9.3 km W of Jalcocatan, rd. to Miramar, 4 Feb 1989 (G), Tellez V et al. 11733 (MO). OAXACA: Rd. Puerto Escondido-Oaxaca, 8 mi N of San Gabriel, ca. 1000 m, 9 May 1965 (9 fl-fr), Breedlove 9912 (MICH); Dtto. Tuxtepec, Presa Temazcal, 8 Dec 1985 (G), Cortes et al. 111 (MEXU); Dtto. Tuxtepec, rd. Esmeralda-Rio Manea, 1 1.5-13.5 mi S of Esmeralda, 19 Jan 1987 (6), Croat 63278 (BG, MO); Mun. Soyaltepec, 4 km S of Hidroelectrica Te- mascal, 20 May 1986 (9 fl-fr), Gereau et al. 2223 (BG, MEXU, MO); Mun. Temascal, Cortina de la Presa M. Aleman, 21 Mar 1964 (6), Gonzalez Q. 575 (MEXU); nr. Rio Soyaloapan, 21-22 Sep 1971 (9 fl-fr), Graham 1395 (MICH); Mun. Juxtlahuaca, 1 km NE of Tilapa, 1200 m, 25 Jan 1985 (9 fl), Lopez G. et al. 262 (MEXU); 5 km N of Pochutla, 2 Mar 1985 (9 fl-fr), L6pez G. 351 (MEXU); Dtto. Choapam, Yaveo, trail of El Chorro, 27 Mar 1938 (9 fl-fr), Mexia 9225 (G, GH, K, MO, NY, U, US); San Felipe Usila, 9 Mar 1929 (6), Sabino s.n. (NY); Chichihua, 17 km N of Lazaro Cardenas, 14 Mar 1983 (6), Tenorio et al. 3450 (MEXU). PUEBLA: Mun. Oaonahua, rd. Teteles-Mazatepec, Atotocoyan, 24 Mar 1976 (6), Mdrquez 717 (NY); Tuza- mapa de Galeana, 1 Apr 1980 (6), Espadas et al. 96 (MEXU); nr. La Ceiba, 19 Apr 1959 (9 fl-fr), Go'mez- Pompa 167 (MEXU); rd. Mexico-Tuxpan, km 247.3,30 Apr 1962 (st), Sarukdn et al. 2090 (MEXU). SAN Luis POTOSi:
3 mi N of Tamazunchale, 13 Aug 1966 (d), Baird 1042 (US); 8 mi NE of Xilitla, 1 Apr 1961 (9 fl), King 4426 (MICH, NY, TEX, US); Mun. Xilitla, El Joba, 6 May 1959 (9 fr), Rzedowski 10595 (US); Ubero, May 1937 (9 fl-fr), LI. Williams 9431 (P), Jun 1937 (9 fl-fr), LI. Williams 9466 (K, P). SINALOA: Mazatlan, Jun 1923 (6), Ortega 5086 (US); El Habal, 1923 (9 fl-fr), Ortega 5194 (US); nr. Col- omas, 20 Jul 1897 (st), Rose 1776 (US). TABASCO: Without locality, 1840 (6), Linden s.n. (G, P). VERACRUZ: 51 mi S of Tantoycua, 13 Jun 1966 (9 fl-fr), Baird 1004 (US); San Andres Tuxtla, Sihuapan, 9 Jan 1973 (6), Calzada 927 (BM, K, MO); between Conejos & Huatusco, 18 Nov 1956 (9 fl- fr), Cox 561 (MEXU); Estaci6n de Biologia Tropical Los Tuxtlas, 22 May 1981 (9 fl), Gentry et al. 32257 (LL, MEXU, MO); rd. Martinez de la Torre-Misantla, km 3, 13 Jun 1963 (9 fl-fr), G6mez-Pompa 913 (A, MEXU); 2 km NE of Putla, 14 Mai 1981 (6), Lorence et al. 3436 (MEXU); Mun. Catemaco, 7 km NE of Sontecomapan, 1 Nov 1981 (9 fl), Nee 22573 (LL); Dtto. Tuxtepec, Ejido Benito Juarez, 30 Aug 1967 (9 fl), Pennington et al. 9073 (NY); Mun.
Catemaco, rd. Catemaco-Montepfo, km 3,15 Mar 1978 (J), Perino 3275 (NY); Cerro de Cuauhtlapan, between Cuauht- lapan & Tuxpango, 30 Apr 1967 (i), Rosas R. 317 (A, K, P, U); 3.1 km W of Santiago Tuxtla, 10 May 1965 (9 fr), M. Sousa 2361 (MEXU); Mun. Hidalgotitlan, Rio Las Cue- vas, campamento La Laguna, 6 Mar 1984 (9 fl-fr), K. Taylor 394 (NY, RB, U); Mun. Hidalgotitlan, Cedillo rumbo a Paso Noral, 17 Jun 1974 (9 fl-fr), Valdivia 800 (K, MO); Mun. Minantitlin, 2 km N of Uxpanapa, 14 Mar 1982 (c), Wendt et al. 3684 (MEXU); Fortunia, Coatzacoalcos R., Mar 1937 (9 fl), LI. Williams 8665 (MICH, P).
GUATEMALA. ALTA VERAPAZ: Cubilqiiitz, Feb 1901 (9 fl-fr), Tuerckheim in herb. Donnell Smith 7666 (GH, K, NY, US), Feb 1901, Tuerckheim in herb. Donnell Smith 7890 (US). ESCUINTLA: Ca. 10 km NE of Escuintla, 1100 m, 2 Aug 1966 (i), Baird 1000 (US); Escuintla, Mar 1890 (9 fl-fr), Donnell Smith 2024 (G, GH, K, NY, P, US); 5 mi N of Escuintla, 31 Jan 1970 (i), Harmon 1828 (MO). IZABAL:
Nr. Izabal, 23 Apr 1966 (6), G. C. Jones et al. 3034 (NY); S of Rio Dulce, 8 Aug 1975 (6), LeDoux et al. 2122 (NY). PETEN: Santa Teresa, Subfn R., 14 Apr 1933 (9 fl-fr), Lun- dell 2891 (MICH, NY, TEX); San Luis, 11 Jul 1959 (9 fl- fr), Lundell 16355 (LL, NY); Laguna Petexbatun, 1 Apr 1964 (9 fl-fr), Lundell 18191 (LL). RETALHULEU: Retal- huleu, Finca Concepci6n Pital, 15 Jul 1966 (d), Baird 1031 (US); San Andres, Apr 1892 (st), Donnell Smith 1499 (K, US); Telahuleu, 12 Jan 1907 (9 fl), Kellerman 6039 (US). SANTA RoSA: Malpais, ca. 1300 m, Sep 1893 (6), Heyde & Lux in Donnell Smith 6238 (G, GH, K, NY, US). SUCH- ITEPEQUEZ: Between Escuintla & Mazatenango, 11 Jan 1978 (9 fl), Harriman 14663 (MO).
BELIZE. Stann Cr. Distr., Cocksomb basin, Jaguar Re- serve, 21 May 1990 (9 fl), Balick et al. 2630 (BG, NY); Toledo Distr., Southern Maya Mtns., Bladen Nature Reserva, 8 May 1996 (d), Davidse et al. 35776 (BG, MO); El Cayo Distr., S of Belmopan, 14-21 Jun 1973 (9 fl-fr), Croat 24563 (GH, MO), 14-21 Jun 1973 (6), Croat 24887 (MO); Toledo Distr., nr. San Antonio, 1 Apr 1969 (9 fl), Lazor et al. 1710a, (9 fr), Lazor et al. 1710d (MO, NY); El Cayo Distr., Valentin, Jun-Jul 1936 (9 fl-fr), Lundell 6373 (MICH), (9 fl-fr), Lundell 6374 (MICH); Middlesex, 2 Oct 1929 (9 fl-fr), Schipp 341 (A, BM, G, K, MICH, MO, NY, S), 1928 (6), Stevenson 14 (US); Maya Mtns., Humming- bird Rd., 13 Aug 1992 (9 fl-fr), R. D. Worthington 21363 (L).
EL SALVADOR. AHUACHAPAN: El Impossible Re- serve, Campana, 1400 m, 24 Jan 1998 (6), Monro et al. 2058 (BG); San Benito, 5 Feb 1990 (6), Sermeno 80 (MO), 23 Dec 1992 (6), Sandoval et al. 942 (MO). LA LIBERTAD:
Cafetal de Miguel Edaudo Araujo, 1000 m, 7 Jul 1988 (9), Monro 2289 (BG). SAN SALVADOR: In or nr. San Salvador, 9 Nov 1985 (6), Berendson 311 (MO), Feb 1922 (9 fl), Calderon 104 (GH, NY, US), 1922 (6), Calderon 1264 (GH, NY, US); San Miguel Tepexontes, 1923 (6), Calderon 1840 (GH, US); San Salvador, 20 Dec 1921-4 Jan 1922 (6), Standley 19614 (GH, NY, US), (9 fl), Standley 22665 (GH, NY), (9 fr), Standley 23181 (US).
HONDURAS. ATLANTIDA: Pico Bonito, Quebrada Grande, ca. 10 km SW of La Ceiba, 8 May 1993 (9 fl-fr),
132 FLORA NEOTROPICA
Evans 1551 (EAP, MO); nr. Tela, 14 Dec 1927-15 Mar 1928, (6), Standley 56637 (A); La Ceiba, 6 Aug 1938 (6), Yuncker et al. 8844 (GH, NY). COMAYAGUA: El Achote, above plains of Siguatepegue, 1350 m, 26 Jul 1936 (st), Yuncker et al. 6127 (NY). CORTES: 1 km SW of Omoa, 13 Dec 1970 (9 fl-fr), Harnon et al. 5198 (MO). OLANCHO: Between Las Marias & La Colonia, 4 Feb 1982 (d), Black- more et al. 1655 (BG, BM, MEXU, MO). TEGUCIGALPA:
Mont. de la Flor, 1937 (st), Hagen et al. 1264 (NY). NICARAGUA. JINOTEGA: Rd. Jinotega-Wiwilf, NE of
Jinotega, 3 Mar 1980 (9 fl), Araquistain et al. 1718 (MO). RiO SAN JUAN: Rio Santa Cruz, 30 Sep 1982 (9 fr), Ara- quistain 3244 (MO); San Juan del Norte, Jan 1896 (9 fl-fr + 6), C. L. Smith 57 (EAP, GH, MO, NY, US). ZELAYA: Nueva Guinea, 11 Aug 1982 (9 fr), Araquistain 3033 (BG, MO); Slilma Sia, 6 Mar 1971 (9 fl-fr), Little 25073 (MO, NY, US); S of Rio Wawa, 60 km NW of Puerto Cabezas, 11 Mar 1971 (6), Little 25077 (MO, US); Mun. Siuna, Santa Rosa, 6 Sep 1982 (9 fl-fr), Ortiz 96 (BG, MO); rd. to Co- loniaYolaina, Colonia La Esperanza, 11-12 Feb 1978 ( 9 fl- fr), Stevens 6359 (BG, MO); rd. to Colonia Manantiales, 0.8 km S of ridge of Serranias de Yolaina, 13-14 Feb 1978 (6), Stevens 6454 (BG, MO); rd. Rio Blanco-Rio Copalar, ca. 29 km E of Rio Branco, 14 Feb 1979 (9 fl-fr), Stevens 12193 (BG, MEXU, MO).
COSTA RICA. ALAJUELA: 6 km E of Ciudad Quesada, 22 May 1968 (9 fl-fr), Burger et al. 5201 (GH, S, US). CARTAGO: Turrialba, 8 Mar 1966 (d), Blum 2216 (MO), (9 fl-fr), Blum 2217 (MO); Atirro, Mar 1894 (9 fl-fr), Don- nell Smith 4934 (GH, US); Volcdn Arenal, 17 Sep 1972 (9 fl-fr), Lent 2948 (U); 9 km SE of San Jose, 1500 m, 19 Dec 1976 (6), Lent 4016 (AAU, MO), (9), Lent 4017 (NY). GUANACASTE: NE of Sabalito, rd. to La Uni6n, 1-18 Jan 1967 (9 fl), Burger et al. 4549 (BM, EAP, G, MO); Pan- American Hwy., Rio Congo, 6 Jan 1964 (6), Molina R. 13571 (EAP, NY). HEREDIA: Finca La Selva, Rio Puerto Viejo, 16 Feb 1981 (9 fl), Folsom 8979 (MO); Finca La Selva, Puerto Viejo de Sarapiqui, 23 Jul 1979 (9 fl), Sperry 1011 (MO); nr. Rio Sarapiquf, 25 Jul 1990 (6), Wiemann 311 (NY); Rio Virilla, 10 km S of Heredia, 1000 m, 25 Mar 1949 (9 fr), L. 0. Williams 16034 (EAP). LIMON: Pta. Ca- huita, 1 Feb 1980 (9 fl-fr), Berg 949 (U); between Siquirres & Rio Pacuare, 20-22 Dec 1919 (9 fl-fr), Burger et al. 6934 (GB, LL, U); 29 km W of Tortiguero, 31 Aug 1979 (9 fl- fr), Davidson et al. 8990A (MO); Parque Tortuguero, Esta- ci6n Cuatro Esquinas, 3 Dec 1987 (9 fl-fr), Robles 1446 (BG, MO); Cerro Coronel, E of Laguna Danto, 15-20 Sep 1986 (9 fl-fr), Stevens 24592 (MO, NY). PUNTARENAS: Golfo Dulce area, nr. Jalaca farm, 8 Jun 1949 (9 fl-fr), Allen 5302 (EAP, US); above Palmar Norte, trail to Buenos Aires, 17 Feb 1951 (6 fl-fr), Allen 5937 (EAP); Esquinas Station, 29 Aug 1952 (9 fl-fr), Allen 6545 (EAP); ca. 17 km N of San Vito, 30 Mar 1978 (9 fl-fr), Davidson 7165 (MO, NY); Monteverde, Lower Tigre Reserve, 1150-1250 m, 29 Jan 1991 (9 fl), Haber et al. 10486 (BG); Canias Gordas, 1100 m, Nov 1899 (9 fl-fr), Pittier 11077 (G, US); 4 mi W of Rinc6n de Osa, 8 Aug 1967 (9 fr), Raven 21644 (NY). SAN JOsE: El General valley, nr. San Isidro El General, 28 Feb 1966 (9 fr), Molina R. et al. 18102 (EAP, NY, US), 2 Mar
1966 (6), Molina R. et al. 18220 (EAP, NY, US); Finca Los Cusingos, 8 mi SE of San Isidro del General, 22 Dec 1948 (6 fl), Norby 133 (EAP); nr. El General, Jan 1939 (9 fl), Skutch 3986 (A, K, MO, NY, S, US); nr. Santo Domingo, 1100 m, 23 Dec 1974 (6 fl-fr), J. Taylor 17540 (NY).
PANAMA. BOCAS DEL TORO: 1 mi W of Almirante, 15 Oct 1965 (6), Blum 1383 (MO); Rio Changuinola, 0.5 mi upstream from mouth, 19 Apr 1969 (st), Lazoretal. 2653 (MO); 10-15 mi S of mouth of Rio Changuinola, 18 Dec 1966 (6), Lewis et al. 868 (MO); Isla Bastimentos, 1-2 km N of Trouble Hole, 14 Feb 1989 (6), Peterson et al. 6858 (MO); nr. Chiriqui Lagoon, Water Valley, 14 Sep 1940 (6), Wedel 771 (GH, MO, US), 9 Nov 1940 (d + 9 fr), Wedel 1559 (GH, MO). CHIRIQUi: Cerro Horqueta, ca. 2300 m, 24 Jul 1966 (6), Blum et al. 2607 (MO); Burica Peninsula, above Brazo Seco, nr. Costa Rican border, 3 Mar 1973 (6), Croat 22562 (MO); Boquete, ca. 1300 m, 1 Jul 1938 (9 fr), Davidson 862 (A). COL6N: Nr. Rio Piedras, rd. to Puerto Bello, 16 Jul 1966 (6), Blum et al. 2496 (MO), (9 fl-fr), Blum et al. 2531 (MO); 3 mi NW of Gamboa, 10 Jul 1966 (9 fl-fr), Haines 467 (LL, MO); Gatun, Dec 1857 (6), Hayes 671 (NY); Las Cruces Trail, 8 Feb 1935 (6), Hunter et al. 462 (GH, MO); nr. Fort Sherman, 5 Jul 1967 (6), Stimson 5251 (COL, NY, PMA, US, SCZ); 7 mi N of Gam- boa, 11 Mar 1966 (9 fl-fr), Tyson 3526 (MO, PMA); Salud, 27 Jul 1971 (9 fl-fr), Lao et al. 169 (MO); 4 km NW of Salamanca, 13 km NE of Buenos Aires, 30 Dec 1973 (d), Nee 9084 (MO, PMA). DARIEN: Nr. Pinogana, 4 Mar 1947 (9 fl-fr), Allen 4311 (EAP, MO); Parque Nacional Darien, Cruce de Mono, 5 Nov 1989 (st), Fisher 54 (BG); 16-19 km SE of Jaque, 20 Jan 1981 (9 fl), Garwood 1130 (BG, MO, PMA); 23 km SE of Jaque, 24 Jan 1981 (6), Garwood 1144 (BG, MO, PMA), (9 fl-fr), Garwood 1147 (MO); nr. Cana, 23 Jun 1959 (d), Stern et al. 490 (GH, MO, US); E of Santa Fe, 18 Jul 1966 (6), Tyson et al. 4852 (MO, SCZ), (9 fl- fr), Tyson et al. 4853 (MO, SCZ); Rio Cocalito, 10 Feb 1982 (9 fl-fr), Whitefoord et al. 125 (BM, MO, PMA, U), (6), Whitefoord et al. 133 (BM, MO, PMA). PANAMA: Cerro Jefe, 30 Aug 1977 (9 fl-fr), Berg et al. 391 (BG); beyond Goofy Lake, rd. to Cerro Jefe, 4 Jan 1968 (6), Correa et al. 575 (MO, PMA); rd. to Cerro Campana, 14 Mar 1968 (9 fl-fr), Correa et al. 850 (MO, PMA); Barro Colorado Is., 15 Apr 1968 (6), Croat 4796 (MO, NY, US), 5 Aug 1970 (9 fl-fr), Croat 11716 (MO, SCZ), 30 Jan 1970 (9 fl), Foster 1484 (PMA); Cerro Azul, Feb 1968 (9 fr), G6mez-Pompa et al. 3413 (MEXU, MO, PMA); 5-10 km NE of Altos de Pacora, 8 Mar 1975 (9 fl-fr), Mori et al. 4989 (BG, MO); Cerro Campana, above Su Lin Motel, 25 Mar 1969 (6), Porter 4194 (MO); rd. to Cerro Azul, 5 Mar 1972 (9 fl-fr), Tyson 6170 (PMA); above El Valle, rd. to La Mesa, 11 Feb 1974 (6), Tyson 6932 (PMA). SAN BLAS: Nusagandi, 16 Aug 1989 (9 fl), Fisher 21 (BG), 26 Jul 1986 (6), Mc- Donagh et al. 405 (BM, MICH, MO); El Llano-Carti rd., continental divide, 25 Aug 1984 (9 fl-fr), Nevers 3754 (PMA); El Llano-Carti rd., km 19, Nusagandi, 26 Aug 1984 (9 fl), Nevers et al. 3769 (BG, MO, PMA). VERAGUAS: Nr. Santa Fe, Rfo Santa Marfa, 26 Mar 1947 (9 fl-fr), Allen 4422 (MO).
COLOMBIA. ANTIOQUIA: Mun. Turbo, rd. Tap6n del
SYSTEMATIC TREATMENT 133
Darien, between Rfo Le6n & Lomas Aisladas, km 37, 25 Aug 1984 (d), Brand 1083 (JAUM); Mun. Anori, Vrda. El Carmen, 18-20.5 km NW of Anorn, 19 Nov 1989 (2 fl-fr), Callejas et al. 8793 (BG, HUA); between Villa Arteaga & Chigorod6, Lomitas, 1 Oct 1961 (2 fl-fr), Cuatrecasas et al. 26114 (COL, P, US); Mun. Mutata, Rio Mutata, 2 May 1987 (2 fl-fr), Fonnegra et al. 2070 (BG, HUA); Mun. San Francisco, Cgto. Aquitania, Rfo Venado, 1150 m, 4 Apr 1992 (2 fr), Fonnegra et al. 4221 (BG, MO); Mun. San Rafael, Campamento Versalles, 1350 m, 23 Mar 1994 (2 fr), Franco et al. 4573 (BG, COL, HUA); Valdivia, 1250 m, 27 Mar 1994 (2 fl-fr), Franco et al. 4596 (BG HUA), (d), Franco et al. 4597 (BG, HUA); Taraza, Cgto. El Doce, Finca Las Mercedes, 28 Mar 1994 (2 fr), Franco et al. 4599 (BG, HUA); Mun. Frontino, Cgto. Nutibara, rd. Nutibara-La Blanquita, Murri region, 4 Mar 1995 (2 fl-fr), Franco et al. 5558 (BG, COL, HUA, LP, MO, US); Mun. Anorf, Vrda. El Roble, 1100 m, 8 Mar 1995 (2 fl-fr), Franco et al. 5567 (BG, COL, HUA, LP, MO, US), 1600 m, 8 Mar 1995 (d), Franco et al. 5571 (BG, COL, HUA, LP, MO, US); Mun. Urrao, Parque Nacional Natural Las Orquideas, Quebrada La Agudelo, 1300-1380 m, 4 Apr 1992 (d), Muntoz et al. 534 (JAUM); Mun. San Luis, Quebrada La Cristalina, 21 May 1987 (d), Ramirez et al. 909 (JAUM), 29 Jul 1987 (d),
Ramfrez et al. 1212 (JAUM); rd. Uraba-Chigorod6-Mala- g6n, Cafio Malag6n, 22 Mar 1986 (2 fl-fr), Renterfa et al. 4713 (JAUM, MO); Mun. Frontino, upper Rfo Cuevas, Cgto. Nutibara, 1630 m, 15 Jan 1987 (2 fl), D. Sdnchez S. et al. 990 (BG); Mun. San Carlos, Cgto. El Jordan, Trocha Las Planes, 10 Apr 1991 (2 fr), Veldsquez et al. 299 (HUA). CAUCA: Rio Micay, El Chachajo, 27 Feb 1943 (2 fl), Cua- trecasas 14266 (F); Mun. El Tambo, Pepital, 1700 m, 12 Jan 1970 (2 fl), Idrobo 6316 (COL); Mun. Guapi, Parque Nat- ural Gorgona, rd. Playa Blanca-Gorgonilla, 6 Sep 1987 (6), Lozano C. et al. 5652 (COL). CHOCO: Bete, Apr 1977 (2 fl-fr), Acero et al. 89 (UDBC); Rio Atrato, nr. Quibd6, 23 Jan 1949 (2 fl), Araque M. et al. 159 (COL, MEDEL, US); ca. 20 km SE of Quibd6, 8 Jul 1986 (d), Berg 1540 (BG); rd. Quibd6-Bolivar, ca. km 30, 9 Jul 1986 (2 fl-fr), Berg 1549 (BG); Mun. Acandi, Vrda. Coquital, El Paramo, 26 May 1989 (2 fl-fr), Betancur et al. 1256 (BG, HUA); Mun. Riosucio, Cerros del Cuchillo, rd. Cuchillo Negro-Cumbre Noroeste, 20 Jan 1988 (2 fl-fr), Cdrdenas 1115 (JAUM, MO); Rio San Juan, Quebrada del Tarapal, 30 May 1946 (2 fr), Cuatrecasas 21510 (F); Cataugad6, nr. Quibd6, rd. to Munquirri, 30 Mar 1958 (2 fl-fr), Cuatrecasas et al. 24050 (COL, US); Rio Atrato, Yuto, 2 Apr 1958 (d), Cuatrecasas et al. 24164 (COL, US); Rio Truando, between La Nueva & La Esperanza, 6 Feb 1967 (2 fl-fr), Duke 9883 (US); Rio Salada, Camp Teresita, 14 Jun 1967 (6), Duke 12204 (NY, US); Rio Munguid6, nr. Altagracia, 5 May 1975 (2 fr), For- ero et al. 1554 (COL, MO); Rio Tamana, tributary of Rio San Juan, below Santa Rosa, 10 Apr 1979 (6 fl-fr), Forero et al. 4969 (BG, COL); Mun. Bahia Solano, nr. Bahia So- lano, 4 Apr 1990 (2 fl), Franco et al. 3011 (COL); Mun. Yuto, rd. Yuto-Quibd6, 12 Apr 1990 (2 fl), Franco 3055 (COL); Quibd6, 14 Apr 1990 (2 fl), Franco et al. 3062 (COL); Rfo Baudo, opposite Quebrada Caimanerita, 11 Feb-29 Mar 1967 (2 fl-fr), Fuchs et al. 21921 (COL, G, K,
NY, P, S, US); Rfo Baud6, between Puerto Pizarro & La Sierpe, 28 Feb 1967 (9 fl-fr), Fuchs et al. 22112 (COL, K, NY, US); rd. Yuto-Certegui, 17 Aug 1976 (9 fl-fr), Gentry et al. 11791 (BG, COL); Rfo Yuto, between Llor6 & La Vuelta, 18 Jan 1979 (6), Gentry et al. 24357 (BG); Mun. San Jose del Palmar, nr. San Jose del Palmar, 11-15 Nov 1985 (9 fl-fr), Lozano C. et al. 4900 (COL); Mun. Riosucio, Cgto. Truand6, confluence of Rfo Chintad6 & Rio Salad6, 24 Oct 1956 (9 fl), Romero-Castanteda 6140 (COL, MEDEL); Nuqui, Alto de Buey, 1000 m, 26 Jun 1940 (9 fl- fr), Sneidern A.71 (A, COL, K, LL, MEDEL, MICH, NY, S, US), (9 fl-fr), Sneidern A.73 (A, COL, K, LL, MICH, NY, S), 28 Jun 1940 (i), Sneidern A.74 (A, MICH, NY, S, US); Rio Jurad6, 4 Oct 1940 (d), Sneidern A.245 (MICH); Parque Nacional Natural Los Katios, Cristales, 19 May 1982 (i), Zuluaga R. 62 (COL). CORDOBA: Tierralta, Rio Sinu, 26 Jul 1988 (9 fr), Cuadros V 4173 (MO). NARINO: Olaya Herrera, Vrda. Naidizales, mouth of Rio Satinga, 5 Mar 1991 (9 fl-fr), Carmona 132 (COL, MEDEL); Mun. Barbacoas, Cgto. Junin, El Diviso, 12 Dec 1993 (9 fl-fr), Franco et al. 5178 (BG); Mun. Barbacoas, Llorente, 12 Dec 1993 (9 fl- fr), Franco et al. 5181 (BG). RISARALDA: Mun. Pueblo Rico, Rio Taiba, 12 Mar 1986 (5), Bernal et al. 952 (COL); Mun. Mistrat6, rd. Mistrat6-Puerto de Oro, Cgto. Jeguades, 26 May 1991 (9 fr), Franco et al. 3442 (COL, MO); Mun. Mistrat6, Cgto. Rio Mistrat6, 11 Sep 1991 (9 fl-fr), Franco et al. 3508 (COL, MO); Mun. Mistrat6, Cgto. Puerto de Oro, 28 Sep 1991 (6), Franco et al. 3605 (COL); Mun. Pueblo Rico, Vrda. Piunda, 26 Apr 1991 (d), Gonzdlez 2419 (COL). SANTANDER: Mun. Puerto Wilches, between La G6- mez & km 80 of Atlantico RR, 23 Apr 1960 (5), Romero- Castaneda 8409 (COL). VALLE: Mun. Buenaventura, Cgto. Bocas del Tiger, Quebrada Mondoya, 27 Oct 1979 (5), Cuadros V 937 (BG, COL, MO); Rio Anchicaya, Quebrada del Retiro, 19 Dec 1942 (5), Cuatrecasas 13689 (COL, F); Rio Naya, Puerto Merizalde, 20-23 Feb 1943 (d), Cuatre- casas 13969 (F, US, VALLE), (9 fl-fr), Cuatrecasas 13979 (F); Rio Yurumangui, El Papayo, 5 Feb 1944 (9), Cuatre- casas 15988 (F, US), (5), Cuatrecasas 16000 (F, US); Rio Cajambre, Quebrada de Ord6fiez, 1 May 1944 (9 fl-fr), Cuatrecasas 17268 (F, VALLE); Quebrada de Guapecito, 16 May 1944 (5), Cuatrecasas 17681 (COL, F); Rio Calima, Quebrada de L6pez, 23 Sep 1961 (9 fl-fr), Cuatrecasas et al. 26042 (COL, P, US); Mun. Restrepo, campamento Rio Azul, Al Chanco, 18 Mar 1986 (9 fr), Devia 1267 (BG, MO); Mun. Trujillo, rd. Trujillo-Naranjal, km 43, Rio San- quinini, 1050 m, 11 Oct 1993 (5), Franco et al. 4489 (BG); Mun. Buenaventura, rd. to Bajo Calima, Bahia Malaga, 16 Mar 1994 (5), Franco et al. 4537 (BG), (9 fl-fr), Franco et al. 4538 (BG), (9 fl-fr), Franco et al. 4539 (BG), (5), Franco et al. 4541 (BG), (5), Franco et al. 4544 (BG), (9 fl-fr), Franco et al. 4546 (BG); Bajo Calima, ca. 15 km N of Buenaventura, 15 Feb 1983 (9 fr), Gentry et al. 40290 (BG, COL, JAUM, MO); Bajo Calima, Cart6n Colombia forest, 1 Jun 1982 (9 fl), Murphy et al. 531 (NY, TULV, US); Mun. Buenaventura, San Isidro, 15 Nov-6 Dec 1979 (5 fl-fr), Rooden et al. 257 (BG, MO, NY, U), (5 fl), Rooden et al. 516 (BG, COL, U).
ECUADOR. AZUAY: Azuay/Guayas/Cafiar-border, E of
134 FLORA NEOTROPICA
Jesus Maria, 12 Apr 1980 (5), Gentry et al. 28521 (MO). CARCHI: Nr. Tobar Donoso, 19-28 Jun 1992 (9 fl-fr), Tipaz et al. 1460, 1479 (BG, QCNE). EL ORO: Nr. Piedras, 11 Jun 1943 (9 fl-fr), Little 6610 (F, Q, US). ESMERALDAS:
Reserva Etnica Awa, Alto Tambo, Rio Mira, 22 Mar 1993 (9 f), Aulestia et al. 1385 (BG, QCNE); Rfo Cayapa, Zapalo Grande, 19-25 Oct 1983 (9 fl), Barfod et al. 48433 (QCNE); Reserva Indigena Awa, Mataje, 6 Oct 1993 (9 fl- fr), H. Beck et al. 2237 (QCNE); rd. Quininde-Esmeraldas, km 5, 13 Sep 1977 (5), Berg et al. 430 (BG, COL, MO, QCA, U); rd. Vuelta Larga-Coronel C. Concha, 14 Sep 1977 (9 fl-fr), Berg et al. 432 (BG, COL, QCA, U); nr. Lita, 9 Feb 1981 (5), Berg 1254 (AAU, BG, COL, MO, QCA, U); rd. Quito-San Lorenzo, km 319, Ventanas, 16 Jul 1964 (9 fl-fr), Jativa et al. 802 (NY, S, US); Rio Onzole, Estero Chontaduro, 14 Jul 1966 (9 fl-fr), Jativa et al. 1099 (MO, NY, S, US); Rfo Santiago, nr. Sucre, 10 Aug 1967 (9 fl-fr), Jdtiva et al. 2217 (S); 2 km S of San Lorenzo, 23 Apr 1943 (9 fl-fr), Little 6361 (F, Q, US); Rio San Miguel, 40 km S of Borb6n, Rio Cayapas, 13 Sep 1965 (9 fl), Little et al. 21052 (Q, QAME, QCNE); Anchayacu, Eloy Alfaro, May- ronga, 28 Oct 1993 (9 fl-fr), Pennington et al. 14530 (BG); ca. 20 km S of Esmeraldas, 10 Apr 1967 (d), Sparre 15514 (S); Reserva Ecol6gica Cotacachi-Cayapas, Luis Vargas Tor- res, Rfo Santiago, 28-31 Oct 1993 (9 fl-fr), Tirado et al. 695 (BG, QCNE). GUAYAS: Rio Congo, between Empalme & Pichincha, 27 Feb 1955 (9 fl-fr), Asplund 15548 (S). IMBABURA: Nr. Guadalupe, RR Ibarra-San Lorenzo, 7 Feb 1981 (9 fl-fr), Berg 1242 (BG, COL, MO, QCA, U). Los Rios: Rio Palenque Biological Station, rd. Quevedo-Santo Domingo de los Colorados, km 56, 19 Jun 1974 (d), Dodson et al. 5601 (QCA, US), 2 Oct 1976 (9 fl-fr), Dodson et al. 6237 (MO, US). MANABi: Parque Nacional Machalilla, Cerro el Pechiche, ca. 15 km NW of Machalilla, 22-24 Apr 1992 (st), Josse et al. 785 (BG). PICHINCHA: Rd. Quito- Puerto Quito, km 113, 10 km N of the rd., Reserva Forestal ENDESA, 8 Jul 1984 (9 fl), Jaramillo 6714 (AAU, GB, MO, NY, QCA); trail Santo Domingo de los Colorados- Quininde, km 43-68, 7 Apr 1943 (5), Little 6185 (F, Q, US); Cant6n Pedro Vicente Maldonado, Reserva Rio Pitzara, Nov 1997 (9 fl), Pennington et al. 16239 (BG).
Cecropia obtusifolia is very variable in many characters. The trees tend to have tall trunks with rel-
atively small, ? globose crowns. There is a tendency
to monocauly as branches may not be formed. Ac-
cording to Alvarez-Buylla & Martinez-Ramos (1992),
trees may grow to 20-35 m tall and become elements
of the forest canopy in Veracruz (Mexico). Indications
that this could happen in other parts of the species range are lacking, and on the basis of observations, it
is very unlikely that it would happen in the southern
part of the range. The lamina varies considerably in
shape due to differences in the extension of the inci-
sions. One can recognize two types of leaves with
regard to the number of leaf segments: the most com-
mon type usually with (8-)10-13 lamina segments
and in the free part of the midsegment >20 pairs of lateral veins. The other is a less common one, most typically found in the Pacific coastal region of Colom- bia and Ecuador (as in Cuatrecasas 13689, Franco et al. 3011 and 5178, Pennington et al. 14530, and Rooden et al. 516), with 7-10 segments and in the free part of the midsegment ca. 15 pairs of lateral veins. The latter type might have links with C. sub- integra. Arachnoid indumentum is always present on the lower surface the lamina of material from Mexico. Specimens without or with very scarce arachnoid in- dumentum have been collected in Guatemala. The fre- quency of material without or with very scarce arach- noid indumentum on the leaves increases through Central America to the Pacific coastal region of Co- lombia. Specimens with arachnoid indumentum on the leaves appear to be absent in Ecuador. The differ- ences in the presence of arachnoid indumentum has led to the distinction of two subspecies, subsp. obtu- sifolia, comprising the material with arachnoid in- dumentum, and subsp. burriada, comprising that without arachnoid indumentum. We decided not to maintain this distinction, as the geographical separa- tion of the two proved not to be clear enough. The two can be indicated as the "obtusifolia-type" and the "burriada-type" respectively. The number of lateral veins varies considerably. Leaves with >25 pairs of lateral veins in the free part of the midsegment show a rather regular venation pattern, and the lateral veins are usually unbranched. Material with numerous lat- eral veins in the midsegment appears to be most com- mon in northern Choc6 and Panama and is found most frequently in the "burriada-type." Throughout the range of distribution, one can find a type with short spikes, often sessile or with short stipes and another type with long spikes, and often also long stipes. This difference in the length of the spikes and stipes occurs most pronouncedly in the "burriada-type." Material with erect inflorescences with short (and relatively thick) spikes can be indicated as the "alvarezii-type." These conspicuously different inflorescences can be found in the same population. Cecropia obtusifolia is very closely related to C. subintegra and (less closely) also to C. heterochroma and to C. mutisiana. It shows similarities to the Amazonian species C. putumayonis and C. utcubambana in the shape of the trees and the frequent lack of arachnoid indumentum on the lower surface of the lamina, but features of the staminate flowers indicate that they are not closely related. The trees are usually inhabited with large ant colonies. The fruiting spikes are reported to be favorite food of toucans. The species has been introduced in Hawaii, where it has become naturalized (Wetterer, 1997).
The plant is used medicinally, mostly to treat di- abetes. Bark fibers are used to make hammocks.
38. Cecropia pachystachya Trecul, Ann. Sci. Nat. Bot., ser. 3, 8: 80. 1847; Berg & Carauta, Albertoa 1(1): 10. 1986. Ambaiba pachystachya (Trecul) Kuntze, Revis. Gen. P1. 2: 624. 1891. Syntypes. Brazil. Minas Gerais: Without locality, Claussen 158 (P), and Ceara: Crato, Nov 1838 (9), Gardner 1845, here designated as lectotype (P; isolecto- types: G, K, P).
Cecropia adenopus Martius ex Miquel, in Martius, Fl. Bras. 4(1): 147, t. 50, 1. 1853. Ambaiba adenopus (Miquel) Kuntze, Revis. Gen. P1. 2: 624. 1891. Syn- types. Brazil. Minas Gerais: Rio Francisco, nr. Sal- gado, Martius 2834 (lectotype, here designated: M) and Brazil. Amazonas. Rio Solimoes, nr. Manaquiri, Martius s.n. (not traced). The two collections belong to different species without doubt.
Cecropia cinerea Miquel, in Martius, Fl. Bras. 4(1): 142. 1853. Ambaiba cinerea (Miquel) Kuntze, Revis. Gen. P1. 2: 624. 1891. Type. Brazil. "Southern," without locality, Pohl s.n. (W, destroyed), photo- graph in MO).
Cecropia lyratiloba Miquel, in Martius, Fl. Bras. 4(1): 144. 1853. Ambaiba lyratiloba (Miquel) Kuntze, Re- vis. Gen. P1. 2: 624. 1891, as "lyratiflora." Cecropia adenopus Miquel var. lyratiloba (Miquel) Hassler, Annuaire Conserv. Jard. Bot. Geneve 21: 130. 1919. Brazil. Minas Gerais: Rio Sao Marcos, Pohl s.n. (W, destroyed, fragment in F).
Cecropia cyrtostachya Miquel, in Martius, Fl. Bras. 4(1): 146, t. 49, 1. 1853. Ambaiba cyrtostachya (Miquel) Kuntze, Revis. Gen. P1. 2: 624. 1891. Type. Brazil. Goias: Arraias, Mar 1840 (Y), Gardner 3982 p.p. (lectotype: K; p. 138).
Cecropia digitata Tenore ex Miquel, in Martius, Fl. Bras. 4(1): 149. 1853. Type. Material cultivated in Hortus Schonbrunn, collected in Brazil (holotype: M-n.v.). Cecropia glauca Rojas, Cat. Hist. Nat. Corrientes 80. 1897. Type not indicated nor traced.
Cecropia adenopus Miquel var. macrophylla Hassler,
Annuaire Conserv. Jard. Bot. Geneve 21: 130. 1919. Syntypes: Paraguay. Upper Rio Apa region, Nov 1901 (c), Hassler 7924 (BM, G), (Y), Hassler 7924a (G).
Cecropia adenopus Miquel var. Iata Snethlage, Notizbl. Bot. Gart. Berlin-Dahlem 8: 359. 1923. Syntypes. Brazil. Without locality, Sellow 2157/2146 (B, de- stroyed); Rio de Janeiro: Mun. Rio de Janeiro, Cor- covado, 1891-1892 (i), Glaziou 18500 (B, de- stroyed; LE).
Cecropia adenopus Miquel var. oblonga Snethlage, No- tizbl. Bot. Gart. Berlin-Dahlem 8: 359. 1923. Syn- types. Brazil. Rio de Janeiro. Mun. Rio de Janeiro, Floresta de Tijuca, 1891-1892 (6), Glaziou 18501 (B, destroyed; C, G, LE, P), (Y), Glaziou 18502 (B, destroyed; C, G, K, LE, NY, P), Glaziou 18506 (B, destroyed; NY); Mun. Rio de Janeiro: Gavea, 1891- 1892 (d + Y), Glaziou 18504 (B, destroyed; BM, C, G, K, LE, P), Glaziou 18507 (B, destroyed).
Cecropia catarinensis Cuatrecasas, Brittonia 11: 171. 1959. Type. Brazil. Santa Catarina: Laguna, 22 Dec 1951 (6), Reitz & Klein 177 (holotype: US; isotype: US).
Cecropia lyratiloba Miquel var. nana Andrade & Car- auta, Bradea 3(22): 164. 1981. Type. Brazil. Rio de Janeiro: Recreio dos Bandeirantes, Reserva Biol6- gica de Jacarapagua, 7 Sep 1979 (d), Andrade 15 (holotype: GUA; isotypes: F, K, R, RB, U).
Tree, to 12 m tall. Leafy twigs 1-4 cm thick, green (or whitish), hispidulous to puberulous, sometimes also with arachnoid indumentum. Lamina subcoria- ceous to chartaceous, ca. 20 X 20 cm to 60 X 60 cm, the segments 9-13, the free parts of the upper seg- ments oblanceolate to subobovate, often ? lobate to sinuate, the incisions down to (5/10-)7/10-9/10, sometimes down to 2 cm from the petiole; apices short-acuminate to subobtuse; upper surface scabrous, hispidulous or partly hirtellous; lower surface puber- ulous to subhirtellous to subtomentose (with hairs of different length), with arachnoid indumentum in the areoles, on the smaller veins or also on the main veins, sometimes only on the main veins and the margin; lateral veins in the free part of the midseg- ment 10-20 pairs, submarginally to marginally loop- connected, branched; petiole 10-55 cm long, puber- ulous to hirtellous, mostly also with sparse to dense arachnoid indumentum; trichilia fused, the brown in- dumentum intermixed with white or sometimes
136 FLORA NEOTROPICA
brownish dense rather short (unicellular) hairs, dis- tinctly longer than the brown trichomes, sometimes rather white long hairs; stipules (8-)10-22 cm long, white to pale green, subsericeous to pilose and with ? dense arachnoid indumentum outside, glabrous or hairy inside. Staminate inflorescences in pairs, the pe- duncle erect or deflexed, the spikes pendulous; pe- duncle 5-12(-17) cm long, hirtellous to subhirsute to subtomentose or minutely puberulous, mostly also with arachnoid indumentum; spathe 3-18 cm long, white to pale green, subsericeous to pilose and with ? dense arachnoid indumentum outside, glabrous in- side; spikes 5-20, (1-)2.5-21 x 0.5-1 cm, with stipes 0.1-0.6 cm long and sparsely to densely hairy or gla- brous hairy; rachis hairy. Staminate flowers: perianth tubular, 1.5-2 mm long, glabrous, the apex plane; fil- aments flat; anthers 0.5-0.7 mm long, appendiculate, detached at anthesis, reattached to the margins of the aperture by the appendages. Pistillate inflorescences mostly in pairs, erect, pendulous in fruit; peduncle (2-)4-15 cm long, indumentum similar to that of the staminate inflorescence; spathe 3-10 cm long, the color and indumentum as in the staminate inflores- cence; spikes 3-6(-8), often 4, 4-8 X ca. 0.4-0.5 cm, to 15(-18) X ca. 1 cm in fruit, sessile or with stipes to 0.2 cm long and puberulous; rachis hairy. Pistillate flowers: perianth ca. 1.5 mm long, with arachnoid in- dumentum below and on the apex outside, also in the style channel inside, the apex almost plane; style short; stigma peltate. Fruit oblongoid, 2-2.2 cm long, tuberculate, dark brown.
Distribution (Fig. 18.1). Brazil, from the southern fringes of the Amazon basin, through central and east- ern parts of the country, extending to Paraguay and northern Argentina, in forest, cerrado, and restinga areas, common in secondary growth, at elevations to 1200 m.
Representative specimens examined. BRAZIL. AMA-
ZONAS: Mun. Humaita, ca. 20 km S of Humaita, Rio Ma- deira, 13 Mar 1975 (6), Gottsberger et al. 156-13375 (U). BAHIA: Marati, 6 May 1966 (Y fl-fr), Belem et al. 2074 (U); rd. Canavieiras-Una, km 30, 16 Jul 1981 (6), Berg et al. 1149 (BG, U); Mun. Santa Cruz de Cabralia, 16 km W of P6rto Seguro, 10 Mar 1983 (d), Brito etal. 228 (MG); Mun. Jacobina, Serra do Tombador, ca. 8 km SW of Jacobina, 27 Oct 1995 (6), Carvalho 6139 (NY); Mun. I1heus, rd. Ilheus- Itabuna, km 22, 12 May 1981 (Y fl-fr), Hage et al. 662 (GUA); rd. to Abaira, ca. 8 km N of Rio de Contas (town), 18 Jan 1972 (6), Harley et al. 15223 (AAU, NY, SPF, U); Mun. Rio de Contas, Pico das Almas, 1500 m, 24 Dec 1988 (9 fl-fr), Harley et al. 27357 (SPF); rd. Ilheus-Una, ca. km 50, 15 Feb 1992 (9 fl-fr), Hind et al. 40 (BG, K); Mun.
Uruquca, rd. Uru,uca-Serra Grande, km 28-30, 4 Nov 1978 (9 fl), Mori et al. 11060 (NY, U); between Len,ois & Rio
Mucugezinho, nr. Morro do Pai Inacio, ca. 1000 m, 20 Dec 1984 (6), Stannard et al. (CFCR) 7271 (GUA, SPF). CEARA: Fortaleza, nr. Rio Coco, 22 Jan 1968 (9 fl-fr), Car- auta 544 (US), (5), Carauta 545 (GUA, U, US); Campos da Cruz, 28 Jan 1968 (5), Carauta 551 (GUA, U, US); between Ipiabina & Sao Benedicto, 21 Jul 1995 (5), A. Fer- nandes et al. s.n. (GUA); Serra de Araripe, Oct 1838 (9 fr), Gardner 1845 (BM, G). DISTRITO FEDERAL: Reserva Ecol-
6gia do IBGE, 15 May 1978 (9 fl-fr), Heringer et al. 491 (IBGE, NY, US); Rio Sao Bartolomeu, rd. Brasilia-Unaf, 11 Dec 1979 (d), Heringer et al. 2902 (MG, MO, NY, US); Paranoa, ca. 5 km E of Brasilia, 1000 m, 11 Jul 1966 (9 fr), Irwin et al. 18150 (COL, G, GH, MO, NY, RB, S, US). ESPiRITO SANTO: Vila Velha, Rodovia do Sol, 30 Aug 1980 (9), Andrade 114 (NY); Domingos Martins, 31 Aug 1980 (9 fl), Andrade 122 (G); Mun. Santa Teresa, Estac,o Biol- 6gica de Santa Lucia, 7 Nov 1985 (5), Boudet Fernandes 1620 (GUA, MO, NY, US); Mun. Cariacica, Rio da Vit6ria, 5 Aug 1983 (5), Hatschbach 46710 (BG, US); Mun. Lin- hares, Reserva Florestal da Cia. Vale do Rio Doce, 27 Sep 1978 (9 fl-fr), Martinelli 5011 (RB); between Brejo Grande & Aracruz, 27 Oct 1992 (9 fl-fr), 0. J. Pereira et al. 4014 (GUA). GoIAs: Serra dos Pirineus, ca. 10 km NE of Co- rumba de Goias, 15 May 1973 (9 fl-fr), W R. Anderson 10370 (MO, NY, U, US); Mun. Goiania, Rio Meia Ponte, 5 Feb 1966 (9 fl-fr), Carauta 301 (GUA, K, NY); Mun. Lu- ziana, Rio Bartolomeu, 15 Aug 1980 (5), Hatschbach 43138 (AAU, C, NY, U); Rio Sao Marcos, Cristalina, 29 Jan 1980 (9 fl-fr), Heringer et al. 17630 (K, MG, MO, NY, US); Rio da Prata, ca. 6 km S of Posse, 6 Apr 1966 (5), Irwin et al. 14415 (F, MG, MO, NY, US); 2 km SE of Piranhas, 23 Jun 1966 (9 fl), Irwin et al. 17688 (GH, MO, NY, RB, US); nr. Sao Gabriel, ca. 50 km N of Planaltina, rd. to Veadeiros, 1000 m, 21 Jul 1966 (9 fl-fr), Irwin et al. 18307 (F, GH, MO, NY, RB, US); Rio Javaes, 27 Jul 1978 (9 fl-fr), Pires et al. 16258 (MG, NY, U); Serra Caiap6, 50 km S of Caia- ponia, 23 Oct 1964 (9 fl-fr), Prance et al. 59603 (GH, K, NY, S, U, US). MARANHAO: Mun. Lago Verde, rd. Alto
Alegre-Lago Verde, km 9, 26 Mar 1985 (st), Anderson et al. 2184 (MG); Is. of Sao Luiz, Feb-Mar 1939 (? fl-fr), Froes 11634 (A, G, K, MICH, MO, NY, S, U); Nova Es- peranca, Alto Turiaqu, 5 Dec 1978 (5), Jangoux et al. 256 (MG, U), (9 fl-fr), Jangoux et al. 257 (MG, U); Rio Alper- catas, Parque Estudial de Mirador, 28 Sep 1988 (9 fr), Nob- erto et al. 215 (MG). MATO GROSSO: 6 km S of Xavantina, 19 Sep 1967 (9 fr), Argent et al. 6426 (NY, P, U); 47 km S of Xavantina, rd. to Aragarcas, 9 Nov 1968 (5), Harley et al. 10968 (E, NY, P, RB, U); Mun. Rio Verde, Serra da Pi- menteira, 28 Aug 1973 (5), Hatschbach 32454 (BG, C); nr. Alto Gar9as, 22 Jul 1974 (9 fr), Hatschbach 34702 (MO, U, US); Serra do Roncador, ca. 86 km N of Xavantina, 3 Jun 1966 (9 fr), Irwin et al. 16554 (GH, MO, NY, RB, US); 40 km S of BR.070, rd. to Pocone, Rio Bento G6mes, 20 Jan 1989 (9 fr), Krapovickas et al. 43065 (C, K); Cuiaba, 22 Jun 1902 (5), Malme 11.1811 (S); Sarar6, RADBRASIL, 22 Aug 1978 (5), Pires et al. 16644 (MG, U); Garapu air- strip, 1 Oct 1964 (9 fr), Prance et al. 59208 (K, NY, S, U). MATO GROSSO DO SUL: Rio Aquidauana, rd. to Rochedo, 24 Jan 1979 (9 fl-fr), Carauta et al. 3077 (F); Mun. Co-
SYSTEMATIC TREATMENT 137
rumba, Abobral, 11 Jun 1994 (6), Hatschbach et al. 60929 (NY); Mun. Ivinhema, Riberao Lib6rio, 1 Apr 1986 (6), Pastore et al. 84 (RB); Nhecolandia region, ca. 100 km SE of Corumbai, 30 Oct 1985 (Y fl-fr), Ratter et al. 5081 (NY). MINAS GERAIS: Mun. Januaria, nr. Hospital Sao Vincente, 3 Feb 1970 (9 fl-fr), Carauta 1062 (GUA, NY, RB, U, US); Mun. Januaria, nr. Ponte Boa Vista, 14 Feb 1972 (st), Car- auta 1474 (F, GUA, RB), (d), Carauta 1475 (F, LE, RB, U); Curvelo, 3 Mar 1972 (9 fl-fr), Carauta 1502 (F, GUA, RB), (6), Carauta 1503 (F, LE, RB); Mun. Araguari, Rio Aguarari, BR.050, 10 Jul 1979 (9 fl-fr), Hatschbach 42244 (AAU, MO, NY, U); Mun. Marlieria, Parque Estadual do Rio Doce, 18 Sep 1975 (9 fl-fr), Heringer et al. 15037 (BG, MO, US); Paracuta, 29 Jan 1980 (9 fl-fr), Heringer et al. 17634 (K, MG, NY); between Grao & Mogol, Ribeirao das Mortes, 10 Dec 1989 (6), Pirani et al. (CFCR) 12431 (GUA, SPF); Caldas, 13 Dec 1864 (9 fl-fr), Regnell L415 (C, G, K, LE, P, S, US). PARA: Upper Rio Tapaj6s, Rio Cururu, Missao Cururu, 6 Feb 1974 (9 fl-fr), W R. Ander- son 10556 (BG, US); Rio Xingu, Ilha Belo-Horizonte, 12 Oct 1986(d fl-fr), Dias et al. 219 (MG); Upper Rio Tapaj6s, Rio Cururui, Lago Boiossu, 24 Jul 1959 (9 fl-fr), Egler 1018 (MG). PARAiBA: Mun. Rio Tinto, 23 Feb 1989 (6), Agra 682 (K). PARANA: Mun. Terra Rica, Rio Paranapanema, 25 Jul 1992 (6), Barros 2585 (SP); Morungava, 21 Jan 1915 (9 fl), Dusen 16456 (F, GH, MO, NY, P, S); Mun. Senges, Fazenda Morungava, Rio Funi'l, 10 Oct 1958 (6), Hatsch- bach 5103 (US); Mun. Paranagua, Rio Guaraguacu, 26 Oct 1960, (d), Hatschbach 7333 (US); Mun. Foz do Iguacu, Parque Nacional, 20 Feb 1962 (9 fr), Hatschbach 9920 (US); Mun. Antonina, Rio Xaxim, 15 Feb 1979 (9 fl-fr), Hatschbach 41973 (C, GB, INPA, MG, MICH, U); Mun. Paranagua, Ponta do Poco, 15 May 1982 (9 fl-fr), Hatsch- bach 44931 (MO, U, US); Parque Nacional do Iguacu, Rio Floriano, 1200 m, 3 Dec 1966 (6), Lindeman et al. 3537 (NY, U). PERNAMBUCO: Mun. Caruaru, Brejo dos Cavalos, 21 Jan 1972 (6), Carauta 1467 (U), (9 fl-fr), Carauta 1468 (U); Rio Formoso, Horto Florestal de Saltinho (d + 9 fl), Falcdo et al. 1155 (RB); Recife, Mata de Dois Irmaos, 28 Sep 1988 (d), Guedes 1581 (UEC); Itapessuma, 9 Oct 1887 (9 fl-fr), Ridley et al. s.n. (BM). PIAUh: Bananeiros, Serra Negra, Parque Nacional, 16 Sep 1977 (6), Barroso 282 (RB); Floriana, Fazenda Agua B6a, 12 Dec 1973 (d), Ra- malho 316 (RB); Urucuf, 4 Jul 1925 (st), Snethlage 647 (F). RIO DE JANEIRO: Macae, rd. to Lagoa Comprida, 7 Apr 1982 (d), Araujo et al. 4918 (NY); Mun. Angra dos Reis, Ilha Grande, 4 Jan 1990 (9 fl-fr), Araujo 9055 (GUA); nr. Cabo Frio, 16 Nov 1980 (9 fl-fr), Berg et al. 1135 (BG, NY, RB, U); Mun. Rio de Janeiro, Restinga de Itapeba, 23 Oct 1963 (d), Carauta 201 (COL, U, US); Mun. Rio de Janeiro, Reserva Biol6gica de Jacarepagua, 9 Sep 1964 (st), Carauta 225 (K, MG, NY); Itatiaia, nr. Hotel Donati, 7 Jan 1966 (9 fl), Carauta 296 (GUA, RB, US); Mun. Nova Ig- ua,u, Vila de Mesquita, base of Macico de Gericin6, 19 Nov 1967 (9 fl-fr), Carauta et al. 491 (NY, RB, U, US); Mun. Silva Jardim, nr. Lagoa Jutunaiba, 2 Apr 1976 (6), Carauta 1899 (RB, U); Resende, P6rto Real, Rio Paraiba do Sul, 22 Oct 1981 (d), Carauta et al. 3893 (GUA, K, NY); Barra do Piraf, between Dorandia & Sao Jose do Turvo, 23 Feb 1987
(5), Carauta et al. 5418 (GUA); Mun. Rio de Janeiro, Res- tinga de Jacarepagua, 3 Nov 1962 (Y fl-fr), Cuatrecasas et al. 26644 (P, US), (5), Cuatrecasas et al. 26646 (P, US); Barra Mansa, Fazenda Paraizo, 18 Jan 1961 (Y fl-fr), Duarte 5485 (RB, U); Morro do Pan da Fome, 19 Feb 1981 (? fl- fr), Lira 409 (C, G, LE); Recreio dos Bandeirantes, 3 Jan 1962 (d + Y fl-fr), Strang 343 (K, MG, NY). Rio GRANDE DO SUL: Sao Leopoldo, Jan 1942 (5), Leite 2300 (A, GH); P6rto Alegre, nr. Gl6ria, 16 Dec 1901 (5), Malme II.869 (S); Sao Leopoldo, Campus da Unisinos, 27 Mar 1991 (Y fl-fr), Quadros et al. 1677 (GUA), (5), Quadros et al. 1678 (GUA); Porto Alegre, Morro da Caloria, 16 Oct 1945 (Y fl), Rambo 29298 (S); nr. Osorio, 25 Jan 1958 (Y fl-fr), Rambo 63578 (S). SANTA CATARINA: Tijucas, 20 Jan 1983 (Y fl), Carauta et al. 4463 (GUA); Sao Leopoldo, 7 Apr 1943 ( fl-fr), Reitz 521 (GUA); Sombrio, 19 Nov 1944 (Y fl), Reitz 853 (S); nr. Tijucas, 8 Feb 1980 (d), Reitz 7903 (GUA); Vila Velha, Sombrio, 1 Nov 1959 (d), Reitz et al. 9339 (GUA); Mun. Ararangua, Sombrio, 27 Feb 1952 (Y fl), L. B. Smith et al. 5897 (US); Tubarao, Jan 1889 (Y fl-fr), Ule 1106 (P, US). SiO PAULO: Campinas, 10 Jul 1968 (5), Carauta et al. 620 (SPF, U, US), ( fl), Carauta et al. 621 (G, U, US), 622 (GUA, U, US); Mun. Iguape, 5 km WSW of Iguape, 18 Feb 1965 (Y fl-fr), Eiten et al. 6196 (K, MO, NY, US); Mun. Canan6ia, rd. Canan6ia-Pariquera-aqd, km 19, 6 Sep 1994 (Y fl-fr), V E Ferreira et al. 67 (SP); Ilha do Cardosa, Res- tinga de Itacuruqc, 8 Oct 1980 (Y fl), Forero et al. 8682 (COL, SP); nr. Jundiaf, 5 Aug 1951 (Y fl-fr), W. Hoehne (SPF) 13234 (F, SP, SPF, US), (5), W Hoehne (SPF) 13237 (SPF); Sao Jose dos Campos, Jacaref, 30 Aug 1949 (9 fl), M. Kuhlmann et al. 2006 (K, NY, SPF, US); Barra Bonita, Rio Jacar&Guaqu, 18 Jul 1991 (5), Romaniuc et al. 1199 (SP); Rinopolis, 19 Jul 1991 (9 fl-fr), Romaniuc et al. 1207 (SP); Mun. Teodoro Sampaio, Reserva do Morro do Diabo, 2 Dec 1986 (9 fl-fr), Tamashiro et al. 18821 (UEC), 4 Dec 1986 (5), Tamashiro et al. 18844 (RB, UEC). SERGIPE: Mun. Santa Luzia de Itanhi, nr. Santa Luzia de Itahi, 27 Nov 1993 ( 9 fr), Amorin et al. 1475 (NY); between Areia Branca & Itabaiana, Serra de Itabaiana, 25 Jan 1991 (9 fl-fr), Barros 2469 (SP).
PARAGUAY. Dtto. Paraguari, Cerro Palacios, 2 Mar 1988 (9 fl), Boswaldo 1363 (BG, MO, NY, TEX); Dtto. Itapua, between Capitan Meza & Puerto Triunfo, Sep 1981 (d), Ferndndez C. 3709 (G, MO, NY); Dtto. Alto Parana, N. of Hemandarias, nr. Pto. Curupayty, Sep 1980 (5), Fer- ndndez C. 4138 (COL, G, MO, NY), (9 fl-fr), Ferndndez C. 4139 (G, NY); Dtto. Canendiyd, between Las Palomas & Salto del Guaira, Sep 1981 (9 fr), Ferndndez C. 4171 (COL, G, MO, NY), (5), Fernandez C. 4172 (G, MO, NY); Dtto. Paraguari, Ybycui National park, 19-28 Mar 1980 (9 fr), Hartshorn 2490 (MO, NY); nr. Villa Rica, 10 Feb 1929 (9 fr), J0rgensen 3836 (A, C, F, MO, NY, S, US); Depto. Mi- siones, Santiago, Estancia La Soledad, 20 Apr 1961 (d), Pedersen 5923 (L); Dtto. Cordillera, Tobaty, 22 Mar 1975 (9 fr), Schinini 10866 (G); Dtto. Amambay, 35 km SE of Bella Vista, 24 Aug 1980 (9 fl-fr), Schinini et al. 20584 (G); Dtto. Guaira, Colonia Independencia, 23 Dec 1986 (9), Schinini et al. 25365 (G, K, MO); Dtto. Caaguazu, Caa- guazu, 26 Jan 1951 (9 fl-fr), Sapper et al. 2178 (P); Dtto.
138 FLORA NEOTROPICA
Central, Memby, 18 Nov 1981 (d), Vavrek et al. 451 (MO, US); Dtto. San Pedro, Primavera, 15 Jul 1957 (d + 9 fr), Woolston 835 (C, K, NY, S).
ARGENTINA. CHAco: Las Palmas, 9 Sep 1917 (d), J0rgensen 2158 (GH, MO, US). CORRIENTES: Dtto. Itu- zaing6, Esteros del Ibera, Laguna Isip6, 15 Nov 1976 (d), Arbo et al. 1463 (C, F, MO); Dtto. General Paz, Ita Ibate, 1 Jan 1946 (d), Ibarrola 4060 (BM, K, S); Dtto. Ituzaing6, Puerto San Antonio, Isla Apipe Grande, 8 Dec 1973 (9 fl- fr + d), Krapovickas et al. 23862 (G, RB); Dtto. Capital, Corrientes, 15 Dec 1986 (9 fl-fr), Krapovickas 40699 (A, F, G, GB, INPA, LPB, MICH, MO, NY, SPF, TEX, USZ), 2 Jan 1987 (d), Krapovickas 40700 (A, C, G, GB, INPA, K, LPB, MICH, MO, NY, SPF, TEX, USZ); Estancia Santa Teresa, 13 Jan 1951 (d + 9 fl), Pedersen 945 (C, G, GH, K, MO, NY, P, S, US); Dtto. Concepci6n, Carambola, 24 Nov 1982 (9 fl-fr), Pedersen 13463 (A, C, L, MO, NY). MISIONES: Nr. Puerto Le6n, 12-21 Jul 1914 (9 fr), Curran 714 (US); Posadas, La Granja, 15 Nov 1907 (d + 9 fl), Ekman 2013 (F, GH, NY, S); Dtto. Gral. Belgrano, Rd. 101, km 5, 13 Dec 1983 (9), Hunziker et al. 11026 (TEX); San- tiago, Estancia La Soledad, 20 Apr 1961 (d), Pedersen 59231 (C, MO); Santa Ana, 4 Jan 1913 (9 fl-fr), Rodr(guez 240 (BM, F); Dtto. Cainguas, rt. 14, km 252, 4 Jan 1950 (9
fl-fr), Schwindt 2983 (LL, US).
Cecropia pachystachya is very common and wide-
spread south of the Amazon basin. It is very variable in the margin of the segments, being lobed or not, in the density of the arachnoid indumentum, and in the size of the trees. Leaves with dense arachnoid indu- mentum are common in the southern and eastern part of the species range, e.g., in Santa Catarina and in Paraguay. The material with dense arachnoid indu- mentum on petioles, stipules, and spathes is quite sim- ilar to that of C. metensis. Small trees are found in the restingas along the coast. Carauta 1061 from Mi- nas Gerais is distinct in the incisions of the lamina down to near the petiole; in that respect it is similar to C. concolor, but the indumentum of the trichilia and the lobation of the lamina segments indicate that this collection belongs to C. pachystachya. Some col- lections from Para (Dias et al. 219 and Egler 1018)
also show similarities to C. concolor in the incisions of the lamina.
In K the collections Gardner 3981 and 3982 are represented by sheets with material correctly referred to Cecropia pachystachya, but also by sheets with ma-
terial referred to C. obtusa (and syntype for that name) or sheets with leaves of C. saxatilis, on a Gar- dener 3982 sheet combined with a pistillate inflores- cence (with sessile spikes) of C. pachystachya and on a Gardner 3981 sheet combined with a pistillate in- florescence (with stipitate spikes), possibly belonging to C. glaziovii or C. palmata. Although it is not quite clear from the description and plate, Miquel has prob-
ably based C. cyrtostachya on a leaf of C. saxatilis and a pistillate inflorescence of C. pachystachya, the latter is presently designated as the lectotype element. The collections Glaziou 18500-18507 are partly mixed, comprising both material belonging to C. gla- ziovii and to C. pachystachya.
The species is probably introduced in the Cook Islands (W R. Philipson 10145, US) and Guam (J. B. Thompson 390, US). The collections examined have well-developed trichilia. Whether material introduced in Indonesia and Malaysia belong to this species or to C. peltata is not clear.
The species is or has been in cultivation in Cuba, Harvard Tropical Garden, Soledad, Cienfuegos (20 Mar 1927, J. G. Jack 4355, A); Philippines, Lamao (P. J. Wester 48, A), and Los Banos (28 May 1914 [ Y ], E. Quisumbing 7933, MICH); Burundi, Bujum- bura, University campus (6, M. Reekmans 9622, MO); Zaire, Eala (6, Corbusier Boland 2061, MO; J. Leemans 83 P; 6 Oct 1945 [Y], J. Leonard 76, P); see also the discussion of C. peltata. Indications that this species has become naturalized in Zaire are lack- ing.
Vernacular names. Brazil: imbauba vermelha (Bahia); torem (Ceara); embauiba branca (Pernam- buco); pau de formiga, torem (Piauf); ambahu, ambaiba-tinga, ambai, ambati, arvore da preguica, ibaifba, imbauiba branca, imbaubao, pau de lixa (Rio de Janeiro; also for C. glaziovii and C. hololeuca, see Duarte, 1959). Paraguay: amba'i, amba'y or ambay.
Cecropia bureauiana V. Richter, Biblioth. Bot. 43: 19. 1897. Type. French Guiana. Without locality, 1845 (6), Melinon s.n. (holotype: P).
Tree, to 20 m tall. Leafy twigs 2-5 cm thick, green, hispidulous with uncinate hairs, also partly hirtellous. Lamina (sub)coriaceous, ca. 20 X 20 cm to 60 X 60 cm (to 100 x 100 cm), the segments 7-11, the free parts of the upper segments (sub)obovate to elliptic to oblong or to ovate, the incisions down to 5/10-8/ 10; apices obtuse; upper surface scabrous to scabri-
SYSTEMATIC TREATMENT 139
dulous, hispidulous; lower surface minutely puberu- lous (with straight to uncinate hairs) on the veins, with arachnoid indumentum in the areoles and on the mar- gin; lateral veins in the free part of the midsegment ca. 15-20 pairs, marginally loop-connected, often branched; petiole 20-60(-90) cm long, minutely pu- berulous; trichilia fused, the brown indumentum in- termixed with short white to brownish (unicellular) hairs; stipules 7-15 cm long, pale to dark (brownish) red, often subpersistent, sparsely hirtellous, some- times also with sparse arachnoid indumentum outside, densely to sparsely hairy or glabrous inside. Stami- nate inflorescences in pairs, pendulous; peduncle 6- 18 cm long, minutely puberulous or also with sparse arachnoid indumentum; spathe 10-17 cm long, white or sometimes reddish, with dense arachnoid indu- ment, also sparsely hirtellous outside, mostly gla- brous or sometimes sparsely hairy inside; spikes 4- 6(-11), (3-)8-15 X 0.8-1.8 cm, with stipes 1-2.5 cm long and minutely puberulous; rachis hairy. Staminate flowers: perianth tubular, 2.5-4.5 mm long, with dense arachnoid indumentum below the apex, the apex convex and glabrous; filaments ? swollen, often basally connate; anthers 1-2 mm long, appendiculate, detached at anthesis, remaining attached to the fila- ment by stretched spiral thickenings. Pistillate info- rescences usually in pairs, pendulous; peduncle 20- 40 cm long, hispidulous, or also with arachnoid in- dumentum; spathe 12-16 cm long, the color and in- dumentum as in the staminate inflorescence; spikes 4, 6-14 X 0.6-0.8 cm, to 20 X 1.5 cm in fruit, sessile; rachis hairy. Pistillate flowers: perianth ca. 2 mm long, with arachnoid indumentum below the apex out- side, also in the lower part of the style channel inside, the apex convex and glabrous; style long, ? S-shaped to straight, minutely puberulous; stigma penicillate to comose. Fruit oblongoid, ca. 3 mm long, almost smooth.
Distribution (Fig. 18.2). From the lower and mid- dle Amazon basin through northern French Guiana to northeastern Suriname and to northeastern Brazil and northeastern Bolivia, in non-inundated forest, in some regions in savanna forest, common in secondary growth, at low elevations.
(F, U). FRENCH GUIANA. Nr. Sinnamary, 29 Nov 1977 (Y
fl-fr), Berg 784 (K, NY, U, US); Rorota Lake, 6 Oct 1978 (Y fl), Grenand 1690 (P, U); Oyapock R., above Saut Mou- touci, 15 May 1970 ( fl-fr), Oldeman T 737 (NY, U); Yar- oupi R., nr. Saut Tainoua, 18 Apr 1970 (Y fl-fr), Oldeman 3118 (U); nr. Cayenne, between Cabassou & Rnmire, 11 Sep
1978 (Y fl), Prevost 292 (P, U); Cayenne, Centre ORSTOM, 18 Aug 1979 (5), Prevost 731 (P, U, US); Piste de St. Elie, 28 Oct 1984 (st), Riera 650 (BG, P).
BRAZIL. AMAZONAS: Manaus, campus INPA, 31 Aug 1973 (Y fl), Berg 252 (F, K, MO, RB, U, US); rd. Manaus- Caracarai, km 20, 21 Sep 1973 (st), Berg 284 (U); Manaus, campus INPA, 8 Nov 1979 (Y fr), D. Coelho (INPA) 90393 (INPA); Rio Branco, Caracarai, 8 Jan 1924 (Y fl-fr), Kuhl- mann 1085 (RB 20199); Manaus, Pensador, Aug 1910 (d), Ule 8837 (G, K, MG, US). CEARA: Nr. Lagoa de Messejana, 20 Jan 1968 (5), Carauta 538 (GUA, U); Crato, Chapada do Araripe, 20 Jan 1968 (st), Carauta 553 (GUA, U), (5), Carauta 554 (G, GUA, U); Serra de Maranguabe, 14 Sep 1908 (5), Ducke (MG) 1690 (MG); Mun. Pacatuba, Itai- tinga, 7 Aug 1957 (5), Ducke 2615 (MG); Guaraciaba, 19 Jul 1979 (5), Jordy Filho 53 (RB). MARANHAO: Mun. Lago Verde, rd. Alto Alegre-Lago Verde, km 9, 22 Mar 1985 (5), A. B. Anderson et al. 2073 (MG, NY); Mun. Moncfio, Rio Turiacu, nr. Urutawy, 12 Feb 1985 (5), Balee etal. 809 (K); Mun. Buriti Bravo, Rio Itapecuru, Serro dos Papagaios, 23 Feb 1983 (Y fl), Rosa et al. 659 (BG); rd. BR.136, km 15, 10 km from Araguana, 8 Dec 1978 (Y fl-fr), Rosa et al. 2901 (MG, NY, U); Mun. Buriti Bravo, Rio Itapecuru, 23 Feb 1983 (Y f-fr), J. U. Santos et al. 659 (MG); Sao Luiz, Andiroba, 8 Aug 1980 (st), M. G. Silva 5724 (MG). MATO GROSSO: Nr. Base Camp, ca. 270 km N of Xavantina, 12049'S, 51046'W, 26 Oct 1968 (5), Harley 10838 (E, K, NY, P, RB, U); Mun. Vila Bela da Sma. Trinidade, rd. Vila Bela-Rio Verde, Fazenda Pousada do Guapore, 18 Aug 1997 ( fr), Hatschbach et al. 66999 (BG). PARA: Mun. Barcar- ena, Itupanema, 9 Dec 1985 (Y fl-fr), Amorozo 240 (MG); Belem, 16 Sep 1942 (5), Archer 7645 (NY, US); Serra dos Carajas, Serra Norte, ca. 12 km E of AMZA exploration camp, 16 Oct 1977 (Y fl-fr), Berg et al. 586 (K, MG, MO, NY, U); Belem, grounds of IAN, 7 Nov 1962 (Y fl), Cua- trecasas 26653 (P, US), ( 9 fl-fr), Cuatrecasas 26655, 26658 (P, US); Alto Curuadna, 16 Oct 1962 (9 fl), Duarte 7189 (INPA, RB); E of Faro, Igarape do Cauhy, 28 Jan 1910 (9 fl-fr), Ducke (MG) 10564 (MG); Maraj6, Pacoval, Sep 1896 (5), Huber (MG) 494 (MG); Capitao Poco, rd. Matuttii- Piri, 13 Jan 1974 (9 fl-fr), Oliveira 6139 (MG); Tucuruf, Lago do Cagancho, 13 Oct 1983 (5), Revilla et al. 8642 (INPA, NY); Maraj6, Soure, Teso, 29 Nov 1906 (9 fl-fr), R. S. Rodrigues (MG) 7841 (G, MG), 1 Dec 1906 (d), R. S. Rodrigues (MG) 7842 (G, MG). PARAiBA: Areia, Pica dos Postes, Mata de Pau Ferro, 4 Dec 1980 (5), Fevereiro et al. M.169 (K); Mamanguape, 14 Jan 1970 (5), Carauta 965 (GUA, U); Joao Pessoa, Miramar, 18 Jan 1970 (5), Carauta 969A (C, G, GUA, U, US), (9 fl-fr), Carauta 969B (GUA, NY, US). PERNAMBUCO: Recife, Dois Irmaos, 23 Jan 1972 (9 fl-fr), Carauta et al. 1471 (GUA, U); Mun. Caruaru, Brejo dos Cavalos, 21 Jan 1972 (5), Strang s.n. (GUA). ROND6NIA: 1 km S of Riberao, rd. Abuna-Guajara-Mirfm, 27 Jul 1968 (9 fl-fr), Prance et al. 6574 (INPA, K, MG, MO, NY, S, U, US). RORAIMA: Foothills of Serra Tepe- quem, Igarape Paparu, 13 Feb 1967 (9 fl-fr), Prance et al. 4361 (INPA, K, MG, MO, NY, U, US); Ilha de Maracd, SEMA Ecological Station, 27 Feb 1987 (9 fl-fr), Ratter et al. 5503 (BG, E, INPA, K, NY). SERGIPE: Mun. Areia
140 FLORA NEOTROPICA
Branca, Estaqao Ecol6gica da Serra de Itabaiana, 23 Jan 1992 (fl), Carauta et al. 6485 (GUA); Itabaiana, ca. 41 km W of Aracaju, 5 km W of Ar6ia Branca, BR.235, 22 Jan 1992 (d ), Thomas et al. 8903 (NY).
BOLIVIA. SANTA CRUZ: Prov. Velasco, Parque Na- cional Noel Kempff M., E of campamento Los Fierros, 18 May 1995 (d), Abbott 16816 (BG); Prov. Velasco, campa- mento El Refugio, 17 May 1994 (d fl), Guilln et al. 1314 (USZ); Prov. Velasco, Parque Nacional Noel Kempff M., between Moira & Chore, 14 Jul 1996 (Y dl-fr), Jardim et al. 3067 (BG); Prov. Velasco, Parque Nacional Noel Kempff M., campamento Los Fierros, 5 May 1994 (d), Quevedo et al. 2572 (USZ); Prov. Velasco, Parque Nacional Noel Kempff M., between Los Fierros & Moira, 2 Jun 1993 (d), Saldias et al. 2914 (USZ), (Y fl), Saldias et al. 2915 (USZ).
Cecropia palmata is closely related to C. glaziovii. These species appear to be allopatric. Some evident differences between these species are listed under C. glaziovii. Considering the similarities in the staminate flowers, these two species might be related to C. saxa- tilis.
An occasional monoecious tree has been encoun- tered in this species. Cavalcante 1392 is unusual in that the staminate inflorescences have short (7-10 cm), slender (ca. 0.3 cm diam.) spikes with short (0.4- 0.5 cm) stipes; the perianths lack the characteristic arachnoid indumentum and the stamens are small (and the anthers probably not detached in the ususal way). These inflorescences resemble those of Cecro- pia concolor.
The species is and has been in cultivation in the garden of Museo Goeldi, Belem, represented by Ca- valcante 1391, 1392, 1394, 1395, and 1396 (MG), made in 1961.
40. Cecropia pastasana Diels, Notizbl. Bot. Gart. Berlin-Dahlem 15: 368. 1941. Type. Ecuador. Pas- taza: Mera, ca. 1000 m, 16 Nov 1938 (d), Schultze-Rhonhof 2996 (holotype: B, destroyed, duplicates not traced; here replaced by: Ecuador. Morona-Santiago: Between Plan del Milagro & Limon (Gral. Plaza], ca. 1500 m, 30 Jan 1981 [d ], Berg 1236 [neotype: QCA; isoneotypes: AAU, BG, COL, GB, MO, NY, U]). Fig. 37
Cecropia sucrensis Cuatrecasas, Revista Acad. Colomb. Ci. Exact. 6(22/23): 297. 1945. Type. Colombia. Ca- queta: Rio Hacha, Sucre, 1000-1040 m, 7 Apr 1940 ( Y fl-fr), Cuatrecasas 9204 (holotype: COL; isotype: F).
Tree, to 25 m tall. Leafy twigs (I-)2-5(-8) cm thick, (dark) red-brown or greenish, sometimes with a thin bluish waxy layer, sparsely puberulous to stri- gillose to subglabrous, usually with ? dense brown pluricellular hairs; pith scarce and whitish, sometimes copious and brown. Lamina chartaceous to subcor- iaceous, ca. (20 X 20 cm to) 35 X 35 cm to 85 X 85 cm, the segments 7-9(-10), the free parts of upper segments ovate or elliptic to obovate, the incisions in the upper part of the lamina down to 3/10-6/10 or down to 8/10, in the lower part less deeply incised, entire or lobate; apices obtuse to rounded to acute or to subacuminate; upper surface scabridulous to al- most smooth, puberulous to hispidulous to hirtellous to strigillose and initially with ? dense to sparse to arachnoid indumentum; lower surface hirtellous to subtomentose to tomentellous or to minutely puber- ulous on the veins, with arachnoid indumentum in the areoles, rather sparse and short, or along the margin only; lateral veins 7-12 or 12-16 pairs, submarginally loop-connected, some or most of them branched; pet- iole (I0-)20-75 cm long, with dense to sparse brown pluricellular hairs or also minutely puberulous, oc- casionally with a thin bluish waxy layer; trichilia on the abaxially bulging (subscrotiform) base of the pet- iole, fused or separate, the brown indumentum inter- mixed with short to rather long whitish to brownish (unicellular) hairs; stipules ca. 10-25, caducous, dull dark red, red-brown, pinkish, or yellowish green, with dense brown pluricellular hairs or also minutely appressed-puberulous outside, densely to sparsely hirtellous to subsericeous inside. Staminate inflores- cences in pairs, often subtended by caducous bracts, to 8 cm long, the peduncle erect and the spikes pen- dulous; peduncle 4.5-9 cm long, densely to sparsely subhispid to hirtellous to subhirsute or (sub)glabrous; spathe 15-32 cm long, dull red (to pinkish) or yel- lowish green (with longitudinal red stripes), with dense brown pluricellular hairs or also minutely pu- berulous outside, (sparsely) hairy inside; spikes 10- 25, (6-)12-28 X 0.3-0.5 cm, sessile or with stipes to 0.3 cm long and sparsely tomentose; rachis hairy. Sta- minate flowers: perianth tubular, 0.8-1.5 mm long, glabrous, the apex slightly convex to plane; filaments flat; anthers 0.5-0.6 mm long, appendiculate, at an- thesis detached, remaining attached to the filament by 2 filiform connections between the connective and the upper margin of the filament (?). Pistillate inflores- cences in pairs or solitary, often subtended by cadu- cous bracts, to 8 cm long, the peduncle erect to de- flexed and the spikes erect to ? pendulous; peduncle 4-10 cm long, densely to sparsely hirtellous to hirsute and with dense brown pluricellular hairs or (sub)-
SYSTEMATIC TREATMENT 141
I ( -~~~~~~~~~~~~~~~~2
9 6
FIG. 37. Cecropia pastasana. 1. Lamina and stipules. 2. Apex of lamina and venation. 3. Pair of staminate inflores- cences with spathes and base of petiole with trichilium. 4. Staminate inflorescence at anthesis and base of petiole with trichilium (Berg 1236). 5. Staminate flower. 6. Stamen and filament after detachment of anther (Franco et al. 4691). 7. Pair of pistillate inflorescences with spathes. 8. Pistillate inflorescence at anthesis and base of petiole with trichilium. 9. Pistillate flower. 10. Fruit (Berg et al. 1761).
142 FLORA NEOTROPICA
glabrous; spathe 10-17 cm long, the color and indu- mentum as in the staminate inflorescence; spikes (2-) 4-6(-7), 10-16 X ca. 0.4-0.8 cm, to 28 X 1.3 cm in fruit, sessile to subsessile (or with stipes to 0.8 cm long), glabrous. Pistillateflowers: perianth 1.5-2 mm long, with arachnoid indumentum below the apex out- side, also in the style channel inside, the apex slightly convex, smooth or sparsely granulate style long, straight, minutely puberulous; stigma comose. Fruit ellipsoid to oblongoid, ca. 1.5 mm long, smooth.
Distribution (Fig. 18.8). From Colombia (Ca- queta) to Peru (Pasco), in premontane to montane for- est, at ca. 900-2300 m.
Specimens examined. COLOMBIA. CAQUETA: Mun. Florencia, rd. to Gabinete, ca. 1300 m, 18 Oct 1993 (d), Franco et al. 4512 (BG, COL). PUTUMAYO: Rd. Mocoa- Pasto, La Teboida, 1300 m, 19 Feb 1995 (5), Franco et al. 4691 (BG, COL, HUA, US), (Y fl-fr), Franco et al. 4692 (BG, COL, HUA, US); Mocoa, Vrda. La Campucana, Finca La Mariposa, 1350 m, 26 Apr 1994 (5), Franco et al. 5343 (BG), (Y fl-fr), Franco et al. 5344 (BG).
ECUADOR. MORONA-SANTIAGO: Rd. Gualaquiza- San Juan Bosco, km 17, 1300 m, 16 Feb 1994 (5), Berg 1693 (BG, QCNE). PASTAZA: Mera, 25 Mar 1940 (Y fl), Lugo 118 (S). TUNGURAHUA: Rd. Bafios-Puyo, between Rfo Verde & Rio Negro, ca. 1400 m, 10 Jan 1981 (9 fl), Berg 1201 (AAU, BG, COL, QCA, U). ZAMORA-
CHINCHIPE: Rd. Zamora-Gualaquiza, between Namirez & Yantzatza, ca. 900 m, 3 Jan 1991 (5), Berg et al. 1655 (BG, GB, LOJA, NY, QCA); rd. Loja-Zamora, between Sabanilla & Zamora, ca. 1300 m, 4 Jan 1991 (9 fl), Berg et al. 1664 (BG, LOJA, QCA).
PERU. AMAZONAS: Prov. Bongara, 4-8 km E of Po- macocha, rd. to Rioja, 2100-2200 m, 9 Feb 1984 (9 fl-fr), Gentry et al. 45194 (BG, K, MO, NY). HUANUCO: Rd. Huanuco-Tingo Maria, below Chinchao, ca. 1800 m, 23 Nov 1997 (9 fl-fr), Berg et al. 1733 (BG, COL, MOL), (5), Berg et al. 1736 (BG, COL, MOL), 24 Nov 1997 (5), Berg et al. 1738 (BG, COL, MOL); rd. Huanuco-Tingo Maria, N of Carpish pass, 54 km NE of Huanuco, ca. 2300 m, 6 Dec 1981 (d), Plowman et al. 11177 (BG, NY, UMS). JUNiN:
Prov. Chanchamayo, rd. to Mina Pichita, 1900 m, 2 Feb 1999 (5), Nuiiez 65 (BG). PASCO: Rd. Oxapampa-Llaupi- Cerro de Pasco, ca. km 15, ca. 1750 m, 29 Nov 1997 (9 fl- fr), Berg et al. 1761 (BG, COL, MOL); rd. Oxapampa-Pu- ente Paucartambo, nr. Oxapampa, ca, 1600 m, 30 Nov 1997 (5), Berg et al. 1763 (BG, COL, MOL). SAN MARTiN: Prov. Rioja, rd. Rioja-Pomacocha, ca. km 50, 900 m, 8 Dec 1997 (9 fl-fr), Berg et al. 1794 (BG, COL, MOL).
Considering the similarities in many characters and similarities in the variation patterns, this species appears to be very closely related to Cecropia andina. It can be distinguished by the consistent presence of trichilia on a subscrotiform base of the petiole, often also by the presence of indumentum (including often
dense brown pluricellular hairs!) on the leafy twigs, petioles, and stipules, the more or less clearly whitish upper surface of the lamina, and only occasional pres- ence of a waxy layer on leafy twigs and petioles. In the northern part of the range of this species, almost coinciding with the range of C. andina, this species occurs at elevations to ca. 1500 m, below the eleva- tional range of C. andina (1500-2300 m). In the southern part of the distribution range, C. pastasana can be found at elevations to 2300 m. In the northern part of the species range, the arachnoid indumentum on the upper surface is rather thin and the leaves look whitish only if seen from some distance, whereas in the southern part of the species range (Huainuco and Pasco) they are more often clearly whitish above. In the northern part of the species range, the incisions of the lamina are mostly down to 5/10 or 6/10 and the number of pairs of lateral veins in the free part of the midsegment is 7-12, but in the southern part of the range the incisions are often deeper, down to 8/ 10, and the number of pairs of lateral veins in the free part of the midsegment is 12-16.
Cecropia pastasana is the white- or whitish- leaved species occurring on the eastern slopes of the Andes, in the northern part of its range (southern Co- lombia and Ecuador), at elevations below that of the white-leaved C. telenitida, and in the southern part (Peru) below that of the white-leaved C. albicans. Ants are usually present, but they are not always of the genus Azteca.
41. Cecropia peltata Linnaeus, Syst. Nat., ed. 10. 1286. 1759; Woodson & Schery, Ann. Missouri Bot. Gard. 47: 173, t. 60. 1960; Velasquez, Acta Bot. Venez. 6: 51, t. 11. 1971; Burger, Fieldiana, Bot. 40: 126, t. 23. 1977; Berg in Gorts-van Rijn, Fl. Guianas, Ser. A, Fasc. 11: 101. 1992. Ambaiba peltata (Linnaeus) Kuntze, Revis. Gen. PI. 2: 623. 1891. Type. Jamaica. Without locality, Anonymous s.n. (holotype: LINN. 1159.2). Fig. 38
Cecropia surinamensis Miquel, in Martius, Fl. Bras. 4(1): 143, t. 46, 1. 1853; Berg, Acta Amaz6nica 8(2): 165. 1978. Amaiba surinamensis (Miquel) Kuntze, Revis. Gen. P1. 2: 624. 1891. Type. Suriname. With- out locality, (5), Focke s.n. (holotype: U).
Cecropia digitata Tenore ex Miquel var. grisea Miquel in Martius, Fl. Bras. 4(1): 149. 1853. Type. Material
SYSTEMATIC TREATMENT 143
UliA l
FIG. 38. Cecropia peltata. 1. Lamina, reduced. 2. Apex of lamina and venation. 3. Stipules and young leaf. 4. Staminate inflorescences with spathes and bases of petioles with trichilia (Berg 978). 5. Staminate flower (Franco et al.
5561) and stamen (Franco et al. 4646). 6. Pistillate inflorescences, one with spathe and the other at anthesis, and bases of
petioles with trichilia (Stevens 3341). 7. Pistillate flower. 8. Fruit (Franco et al. 5566).
144 FLORA NEOTROPICA
cultivated in Hortus Schonbrunn, from material col- lected in Mexico (holotype: M, photograph in MICH).
Cecropia propinqua Miquel, in Martius, Fl. Bras. 4(1): 149. 1853. Type. Material cultivated in Hortus Schonbrunn, from material collected in Mexico by Schott in 1852 (holotype: M-n.v., photographs in F, MO).
Cecropia scabrifolia V. Richter, Biblioth. Bot. 43: 15, t. 2. 1897. Type. Nicaragua. Grenada, Jun 1869 (5), L,vy 52 (holotype: P; isotypes: C, G, P, fragment ex C in F).
Cecropia arachnoidea Pittier, Contr. U.S. Natl. Herb. 18: 226. 1917. Type. Panama. Panama: Nr. Matachin, 23 Jun 1911 (Y), Pittier 4056 (holotype: US; isotype: US).
Cecropia asperrima Pittier, Contr. U.S. Herb. Natl. 18: 227. 1917. Syntypes. Nicaragua. Without locality, 1853-1856 (d + Y), C. Wright s.n. (US, 5 + Y; GH, 5).
Cecropia hondurensis Standley, Publ. Field Mus. Nat. Hist., Bot. Ser. 4: 302. 1929. Type. Honduras. Atlan- tida: Nr. Tela, 14 Dec 1927-15 Mar 1928 (?), Stan- dley 54528 (holotype: F; isotype: US).
Cecropia goodspeedii Cuatrecasas, Revista Acad. Col- omb. Ci. Exact. 6(21): 67. 1944; Not. Fl. Colomb. 6 (Trab. Com. Bot. etc. Colomb): 41. 1944. Type. Co- lombia. Antioquia: Rio Cauca, Puerto Valdivia, 17- 20 Feb 1942 (Y), Cuatrecasas et al. 30079 (holo- type: COL; isotypes: F, US).
Tree, to 15(-25) m tall. Leafy twigs 1.5-4 cm
thick, green, hispidulous with curved to uncinate hairs. Lamina chartaceous to subcoriaceous; ca. (10 X 10 cm to) 20 X 20 cm to 60 X 60 cm, the segments (7-)8-10(-1 1), the free parts upper segments oblong or elliptic, the incisions down to ca. 5/10-7/10; apices short-acuminate to rounded; upper surface scabrous, hispidulous; lower surface minutely puberulous, in- termixed with rather sparse longer hairs, with arach- noid indumentum in the areoles, also on the reticulum or extending to the main veins; lateral veins in the free part of the midsegment (8-)10-15(-17) pairs, marginally (or submarginally) loop-connected, usu- ally branched; petiole (10-)20-50 cm long, puberu- lous or also with arachnoid indumentum; trichilia fused, the brown indumentum intermixed with short (or sometimes with rather long) white hairs; stipules 3-10(-12) cm long, pinkish or reddish, sometimes subpersistent, strigose to hirtellous and often also with arachnoid indumentum outside, sparsely to densely (sub)sericeous inside. Staminate infiores-
cences erect to deflexed; peduncle 2-12.5 cm long, sparsely to densely puberulous to hirtellous to sub- hispid(ulous); spathe 2.5-7 cm long, pinkish (to pur- plish), greenish or whitish, puberulous to hirtellous and often also with arachnoid indumentum outside, (sub)sericeous inside; spikes ca. (10-)15-25(-60), 0.5-6 X 0.2-0.25 cm, with stipes to 0.8 cm long and hairy; rachis hairy. Staminate flowers: perianth tu- bular, 1-1.5 mm long, with short and stiff hairs or sometimes short arachnoid indumentum below the apex, the apex plane, smooth; filaments flat; anthers ca. 0.5-0.6 mm long, appendiculate, detached at an- thesis, reattached to the margin of the aperture by the appendages at anthesis. Pistillate inflorescences in pairs, erect to pendulous; peduncle 3-10(-16) cm long, sparsely to densely puberulous to hirtellous to hirsute or to subhispid, often also with arachnoid in- dumentum; spathe 3.5-6 cm long, the color and in- dumentum as in the staminate inflorescence; spikes 3-4(-5), 1-4 X 0.3-0.4 cm, to 9 X 1.1 cm in fruit, sessile or with stipes to 0.5 cm long and hairy; rachis hairy. Pistillate flowers basally connate; perianth 1- 1.5 mm long, with arachnoid indumentum below and on the apex outside, absent (or in and below the style channel) inside, the apex flat to slightly convex; style short; stigma peltate. Fruit ovoid to ellipsoid, ca. 2 mm long, tuberculate, dark brown.
Distribution (Fig. 18.3). From southemn Mexico to northern South America (Colombia, Venezuela, Guyana, Suriname, and northern fringe of the Brazil- ian part of the Amazon basin), also in Jamaica and Trinidad, in wet to deciduous forest areas, commonly in secondary growth, roadsides, pastures, etc., at el- evations to 1000 m, sometimes to 1500 m, or (in Co- lombia: Santander) to 2000 m.
Representative specimens examined. MEXICO. CAMPECHE: 2 mi S of Ruinas Edzna, 27 Aug 1966 (d), Baird 1017 (US); Ruinas Edzna, 27 Aug 1966 (d), Baird 1018 (US); 30 km E of Escarcega, rd. Escarcega-Chetumal, 12 Jul 1983 (dc), Cabrera 5099 (MO). CHIAPAS: Mun. Fron- tera Comalpa, Hwy 190, 3 mi S of Rio San Gregorio, 29 Apr 1965 (d), Breedlove 9822 (CAS, LL, MICH, US); Mun. Ocozocoautla, 45 km N of Ocococoautla, 17 Oct 1971 (d), Breedlove et al. 20725 (MEXU); Mun. Catzadja, E of Ba- jadas Grandes, 11 Oct 1972 (d), Breedlove 28781 (CAS); nr. Judrez, NW of Pichucalco, 31 Aug 1965 (d), Chavelas et al. 2463 (MEXU); Mun. Palenque, betweeen Salto de Agua & Ruinas Palenque, 30 Jul 1967 (9 fr), Clarke s.n. (CAS, US); Mun. Ocosingo, Boca de Chajul, 27 Apr 1993 (9 fl-fr), Dminguez V 698 (MEXU); Pueblo Nuevo, Solis- tahuacdn, 1260 m, 11 Aug 1967 (d), G6mez-Pompa 2568 (MEXU); Mun. Ocosingo, nr. San Javier, 23 May 1991 (6), Gonzalez-Espinosa et al. 1486 (MEXU); 5.6 km E of Chiapa de Corzo, 15 Jul 1966 (6), Laughlin 1282 (CAS); Mun.
SYSTEMATIC TREATMENT 145
Tuxtla Gutierrez, El Zapotal, SE of Tutxla Gutierrez, 19 Aug 1988 (Y fr), Palacios E. 66 (CAS); nr. Ocozocoautla, 12 Nov 1988 (Y fl-fr), Reyes G. et al. 1266 (BM); nr. Simojovel de Allende, 1000 m, 10 Oct 1967 (Y fr), Shilom Ton 3075 (CAS). OAXACA: Hwy. 185, 4 km N of Matfas Romero, 24 Jul 1958 (Y fl), King 715 (LL, MICH, TEX, US); Ubero, Apr 1937 (d), Ll. Williams 9264 (MICH, US). QUINTANA
Roo: 26 mi S of Felipe Carrillo Puerto, 5 Jul 1966 (6), Baird 1024 (US); 1 E of Cancun, 12 Sep 1982 (6), Cabrera et al. 3483 (MO); 30 km E of Escarcega, 12 Jul 1983 (6), Cabrera 5099 (MEXU); Mun. Oth6n P. Blanco, 9 May 1982 (6), Chan et al. 1432 (MEXU); Coba, 10 Aug 1975 (6), Lopez F 537 (MEXU); nr. Felipe Carrillo Puerto, 7 Jul 1989 ( fl), Serralta P. 34 (MEXU); Cancun, nr. airport, 9 May 1980 (6), Tellez et al. 2189 (MEXU, MO); 17 km N of Felipe Carrillo Puerto, 11 Jun 1980 (6), Tellez et al. 2510 (MEXU); nr. Andr6s Quintana Roo, 4 Sep 1980 (d), Tellez et al. 3352 (MEXU); 2 km S of Tres Garanitas, 5 Sep 1980 (Y fl), Tellez et al. 3425 (MEXU). TABASCO: Nr. airport of Villahermosa, 14 Oct 1988 (Y fl-fr), Ascencio R. 140 (MEXU); 10-40 km W of Huimanguillo, 20-29 Mar 1963 (6), Barlow 30 (BM, MICH); Carretera del Golfo, km 24, 17 Aug 1977 (Y fl-fr), Cdrdenas 186 (MEXU); Balancan, La Palma, 1-6 Jan 1939 (6), Matuda 3240 (GH, MEXU, MICH); Lomas de San Sebastian, 27 Feb 1889 (6), Rovi- rosa 383 (K, NY). VERACRUZ: Isla Villa Azueta, 24 Dec 1968 (? fr), Hernandez et al. 488 (LL, MEXU, NY); Co- atzacoalcos, Isthmus of Tehuantepec, Jan 1895 (6), C. L. Smith 1040 (EAP, GH, MO, NY, US). YUCATAN: Chichan- kanab, (d), Gaumer et al. 1459 (BM, C, GH, MO, US); nr. Piste, May-Aug 1938 (6), Lundell et al. 7548 (MICH); Xcoochle, ruins of Uxmal, 16 Sep 1865 (d), Schott 639 (BM, MO, US); Chichen Itza, 29 Jun 1932 (6), Steere 1700 (MICH).
GUATEMALA. ALTA VERAPAZ: Nr. Sepacuite, Jun 1905 (Y fl-fr), Cook et al. 336 (US); Chamiquin, Apr 1889 (6), Donnell Smith 1675 (US). IZABAL: Rio Sauce, 0-2 km N of Lake Izabal, 21 May 1966 (d fl-fr), G. C. Jones et al. 3465 (EAP, LL, MICH, NY, US); Morales, 10 Mar 1907 (6), Kellerman 6416 (MO, US); Sinai, ca. 30 km SW of Santo Tomas de Castillo, 15 Mar 1988 (6), Marshall et al. 394 (BG, MO, NY); Rio Motagua, nr. Quirigua, 6 Jun 1905 (6), Pittier 385 (US); Quebradas, 19-22 May 1919 (9 fl), Pittier 8626 (NY, US); nr. Quirigua, 15-31 May 1922 (d), Standley 24143 (GH, NY, US). PETEN: Lake Peten Itza, be- tween Remate & San Jose, 25 May 1960 (9 fl-fr), Contreras 1007 (LL, NY, S); Tikal, 5 Aug 1960 (9 fl-fr), Contreras 1390 (LL, NY, S); Macanche, 21 May 1966 (9 fl), Contreras 5832 (LL, US), 22 May 1966 (6), Contreras 5845 (EAP, GH, LL, TEX); Santa Teresa, Rio Subin, 13 Apr 1933 (d), Lundell 2902 (GH, MICH, NY, S); La Libertad, 25 Apr 1933 (9 fl), Lundell 3006 (MICH, NY), (6), Lundell 3016 (MICH, NY, S, US); Lake Zoty, 18 May 1933 (9 fl-fr), Lundell 3295 (MICH, NY, S), (d), Lundell 3296 (MICH, NY); Tikal National Park, Tikal, 8 May 1959 (6), Lundell 15972 (LL, NY); Santa Elena, rd. to Sacpuy, 9 Sep 1971 (9 fr), Tun Ortfz 1899 (EAP, G, MO, S). ZACAPA: Quirigua, 18 Nov 1970 (9 fl-fr), Harmon et al. 4932 (GH, MO); 8 mi
E of Lobo, 8 Jul 1975 (9 fl-fr), Watkins et al. 726 (LL, MO, NY).
BELIZE. Cayo Distr., 36 mi W of Belize, 3 Jul 1966 (9 fl-fr), Baird 1023 (US); Belize Distr., Northern Hwy., mi 42.5, 27 May-6 June 1973 (6), Croat 23990 (MO); Belize Distr., Northern Hwy., mi 42.5, N of Maskall R., 7 Jun 1973 (9 fl), Dwyer 11017 (GH, MO); Corozal Distr., 1931-1932 (d), Gentle 455 (MICH, NY, S); Maskall, 15 May 1934 (9 fl), 8 Mar 1935 (9 fl-fr), Gentle 1566 (A, MICH, MO, NY), 29 May 1935 (6), Gentle 1664 (GH, K, MICH, MO, NY); Corozal Distr., Alfonsville, 8 Jul 1933 (6), Lundell 4813 (MICH, NY, S), (9 fl-fr), Lundell 4814 (MICH, NY); Or- ange Walk Distr., between Orange Walk & Benque Viejo, 20 Aug 1980 (9 fl-fr), Sutton et al. 152 (BM, MO); Altun Ha, 24 Aug 1970 (9 fl-fr), Wiley 572 (MO, NY).
HONDURAS. ATLANTIDA: San Jose de Texiguat, 30 km NE of Tela, 20 Jun 1989 (9 fr), Galo 3 (BM); nr. La Ceiba, 6 Aug 1938 (9 fl-fr), Yuncker et al. 8844 (BM, G, MO, S, US). COL6N: Rfo Guaimoreto, 4.5 mi NE of Tru- jillo, 20 Feb 1981 (9 fr), Saunders 1035 (BM, NY, TEX). COPAN: Santa Rosa de Copan, 10 May 1987 (6), Blackmore et al. 3797 (BM, MO). GRACIAS A DIos: Laguna Ibans, 28 May 1985 (juv), Knees 2812 (BM). ISLAS DE LA BAHiA:
Isla de Roatan, 30 Mar 1984 (9 fl), Nelson et al. 8737 (MO, NY). MORAZAN: Rio Jicarito, drainage of Rio Yeguare, 22 Jun 1948 (6), Glassman 1694 (EAP, NY); El Zamorano, 6 Jul 1948 (6), Molina R. 427 (EAP), 15 Nov 1945 (d), Val- erio R. 3593 (EAP, GH, US). OLANCHO: Between Campa- mento & border Olancho/Morazan, rd. Tegucigalpa-Juti- calpa, ca. km 100-101, 27 Oct 1996 (6), Berg 1810 (EAP); 6 km SE of Catacamas, 16 May 1987 (9 fl-fr), Ortega U. 361 (EAP, MO). SANTA BARBARA: Mt. Bella Vista, above San Pedro Sula, 4 Oct 1969 (9 fl-fr), Barkley et al. 39574 (GH); San Pedro Sula, Jul 1887 (9 fr + 6), Thieme 194 in herb. Donnell Smith 5494 (G, GH, K, US). YORO: Subirana, ca. 1000 m, Sep 1937 (st), Hagen et al. 1063 (NY). SWAN
ISLANDS: Great Swan, 15-24 Aug 1971 (st), Proctor 32563 (IJ).
NICARAGUA. BOACO: 4 km NW of Camoapa, 1 Feb 1979 (d), Grijalva et al. 16 (MO, U). CHINANDEGA: Chin- andega, 13 Jan 1903 (9 fl-fr), Baker 2007 (A, G, GH, K, MO, NY, P, US); Ameya, 19-21 Jun 1923 (6), Maxon etal. 7193 (US). CHONTALES: Rd. Juigalpa-Puerto Diaz, 11 Jul 1980 (6), Guzmdn et al. 378 (MO, U); Cerro La Bateca, 8 km S of Cuapa, 21 Sep 1983 (d), Nee 28268 (MO, NY); 3.6 km E of La Gateada, rd. to Nueva Guinea, 17 Jul 1977 (d), Stevens 2856 (BG, BM, MO); ca. 2.8 km N of Cuapa, 3 Sep 1977 (6), Stevens 3641 (BG, BM, MO); rd. Juigalpa- La Libertad, ca. km 17.4, 23-25 Sep 1977 (9 fl-fr), Stevens 4145 (BG, MEXU, MO); 4.9 km E of Puerto Diaz, rd. to Juigalpa, 5 Jun 1980 (d), Stevens et al. 17423 (BG, MO). ESTELi: Isiquf, 14 Jun 1984 (9 fl-fr), Laguna 400 (BG, MO). GRANADA: Volcan Mombacho, 2 Jul 1975 (9 fl-fr), Atwood et al. 70 (MO, NY), 28 Oct 1976 (9 fr), Gomez s.n. (MO); between Santa Rosa & Santa Margarita, 19 Mar 1982 (6), Moreno 15989 (BG, MO). LE6N: Volcan Casita, 900- 1400 m, 14 Jun 1984 (9 fr), Moreno 24261 (BG, MO). MADRIZ: Santa Teresa, Cerro Meino, 3.5 km SE of Somoto, 23 Sep 1980 (6 fl-fr), Moreno 2796 (BG, MO). MANAGUA:
146 FLORA NEOTROPICA
El Charco, Rfo El Carmen, 2 km W of Salamina, 7 Sep 1981 (i), Moreno 10791 (MO); Hwy. 12, km 24, WSW of sum- mit of Sierra de Managua, 7 Jul 1977 (9 fl), Stevens 2703 (BG, BM, MO); ca. 2.3 km from Hwy. 12, rd. to Sierra de Managua, 29 Jul 1977 (9 fl-fr), Stevens 2924 (BG, MO). MASAYA: Laguna de Apoyo, 24 Oct 1980 (d), Moreno 3990 (MO); Parque Nacional Volcan Masaya, 13 Dec 1977 (9), Neill 3094 (MO, U), 21 May 1980 (9 fr), Stevens et al. 17212 (MO). MATAGALPA: 20 km NE of El Tuma, be- tween Rio Wasaka & Rio Bulbul, 9 Oct 1982 (9 fr), Grijalva et al. 1355 (MO); rd. Muy Muy-Equipulas, ca. km 19, 30 Jul 1978 (c), Stevens 9556 (MO), (9 fl-fr), Stevens 9557 (MO). NUEVA SEGOVIA: 8 km NE of Jicaro, 2 Aug 1980 (d), Moreno 1707 (BG, MO); ca. 0.5-4 km N of San Fer- nando, 13 Aug 1977 (9 fl-fr), Stevens 3341 (BG, MO); ca. 1.6 km W of Murra, rd. to El Jicaro, 27 Jun 1980 (c), Ste- vens et al. 17599 (BG, MO). Rio RIVAS: Volcan Maderas, above Balgiue, Isla de Ometepe, 15 Sep 1983 (9 fr), Nee et al. 28120 (BG, MO, NY); nr. Moyogalpa, 16 Sep 1983 (9 fl-fr), Nee et al. 28174 (BG, MO, NY); Isla Ometepe, Volcan Maderas, 27 Apr 1984 (9 fl), Robleto 401 (BG, MO). ZE- LAYA: Between El Muelle de los Bueyes & Villa Somoza, 6 Apr 1961 (9 fl), Bunting et al. 1052 (NY, S); Puerto Ca- bezas, 28 Feb 1971 (9 fr), Little 25025 (MO, US); Mun. Siuna, El Hormiguero, 30 Jun 1984 (6), Ort(z 1979 (MO); Corn Is., 4-6 Apr 1981 (9 fr), Stevens 19926 (MEXU, MO).
COSTA RICA. ALAJUELA: Alajuela, Mar 1896 (6 + 9 fl), Donnell Smith 6772 (BM, G, GH, K, NY, US); nr. Los Chiles, Rio Frio, 7 Aug 1949 (9 fr), Holm et al. 950 (A, G, MO, P). GUANACASTE: Pozo Azul, 14 May 1992 (9 fl), Biesmeijer et al. 188 (U); 6 km SE of Santa Cruz, 24 Jul 1964 (9 fl-fr), Ferreyra 15939 (USM); Inter-American Hwy., Rio Lagartos, s.d., Khan et al. 1155 (BM); Nicoya, Mar 1900 (9 fl-fr), Tonduz 13870 (BM, K, NY, P, US). PUNTARENAS: Rio Terraba, nr. Palmar Norte, 27 Aug 1952 (d fl-fr), Allen 6543 (EAP); Monteverde, rd. Santa Elena- San Luis-Lagarto, 10 Jul 1990 (9 fl-fr), Haber 9970 (MO); nr. Cascajal, 25 km ESE of Puntarenas, 3 Jul 1949 (6), Holm et al. 234 (G, MO, P, U); between Lagarto & Botuca, 24 Aug 1965 (9 fl), A. Jimenez M. 3473 (NY, PMA, US); Rin- c6n de Osa, 11 Feb 1974 (6), Liesner 2066 (BG, MO); Santo Domingo de Golfo Dulce, Mar 1896 (6), Tonduz 10055 in herb. Donnell Smith 7164 (US). SAN JosE: Be- tween Santa Ana & Villa Col6n, 21 Jul 1971 (9 fr), Burger et al. 7611 (BM, MO, U).
PANAMA. CHIRIQUi: Puerto Armuelles, 23 Jul 1967 (d), Hladik 497 (P); 6 mi W of David, 29 Aug 1962 (9 fr), Tyson 929 (MO); 6 mi N of David, 25 Apr 1969 (9 fl-fr), Lazor 2824 (MO); 10 mi N of David, 25 Apr 1969 (9 fl), Lazor 2825 (MO, PMA). COCLE: El Cope, 1 Sep 1977 (6), Berg et al. 397 (BG), (9 fl-fr), Berg et al. 398 (BG); 0.5 km N of El Cope, 10 Sep 1977 (6), Folsom 5226 (MO, PMA, U). COL6N: Nr. RIo Piedras, rd. to Puerto Bello, 16 Jul 1966 (6), Blum et al. 2515 (MO), (9 fr), Blum et al. 2529 (MO); Coco Solo, 13 Aug 1967 (9 fr), Elias et al. 1606 (GH, MICH, MO); Salud, 3 Aug 1971 (9), Lao et al. 203 (MO, PMA); Col6n Hwy., bridge crossing Rio Chagres, 1 May 1969 (9 fl), Lazor et al. 3066 (MO); 3 km E of Buena Vista, 28 Oct 1973 (9 fr), Nee 7652 (MO); Fort Sherman, 28 Jun
1967 (9 fl-fr), Stimson 5230 (NY); Gatun Railway Station, 5 Mar 1966 (6), Tyson 3524 (MO). DARIEN: Rfo Pirre, 2-5 mi above El Real, 23 Jun 1962 (9 fr), Duke 4904 (GH, MO); 23 km SE of Jaque, 23 Jan 1981 ( 9 fr), Garwood 1162 (MO, PMA, U); nr. El Real, 22 May 1969 (9 fl), Lazor et al. 3439 (MO); Rio Tuquesa, mining camp Charco Chiva, 5 Jul 1975 (9 fl-fr), Mori 6999 (BG, MO). Los SANTOS: 12 mi S of Macaracas, 22 Feb 1966 (9 fl-fr), Blum et al. 2176 (MO). PANAMA: Barro Colorado Is., 27 Apr 1968 (6), Croat 5131 (MO), 25 Sep 1968 (9 fl), Croat 6452 (MO); rd. to Cerro Azul, 4 mi from Inter-American Hwy., 26 Jul 1970 (d), Croat 11496 (MO); Summit Gardens, 6 Aug 1970 (9 fl-fr), Croat 11749 (MO); El Llano, 14-19 Oct 1962 (9 fl-fr), Duke 5846 (US); San Miguelito, 9 Aug 1963 (9 fr), Duke et al. 6645 (BG); San Jose Is., 9 Oct 1944 (d), Johnston 78 (GH, US), 15 Oct 1944 (9 fl-fr), Johnston 154 (MO), (6), Johnston 155 (GH, US), 12 Nov 1944 (9 fl-fr), Johnston 471 (GH); La Capitana, Chepo, 9 Jun 1971 (6), Lao 122 (MO); rd. to Cerro Campana, 1-3 mi from Pan-American Hwy., 3 Aug 1983 (6), J. S. Miller et al. 953 (MO); 6 km S of Arraijan, 28 Aug 1973 (6), Nee 6659 (MO); 2 km W of Gamboa, 21 Oct 1973 (9 fl-fr), Nee 7533 (LL, MO, US); rd. to Cerro Azul, 23 Apr 1972 (6), Tyson 6205 (PMA). SAN BLAS: Nusagandi, 18 Aug 1989 (9 fl-fr), Fisher 26 (BG), (6), Fisher 27 (BG); rd. Suedi-Kariadi, Rio Ailin- gandi, 10 Dec 1989 (9 fl-fr), Herrera et al. 644 (MO); El Llano-Carti rd., continental divide, 25 Aug 1984 (9 fl-fr), Nevers 3753 (BG, MO). VERAGUAS: Ca. 20 mi ESE of Gua- bala, 3 Jul 1966 (9 fl-fr), Blum et al. 2443 (MO); Islas Con- treras, Isla Brincaneo, 20 Jul 1984 (9 fl), Churchill 5720 (BG, MO).
JAMAICA. Parish of St. Andrew, grounds of University of the West Indies, 2 Jul 1966 (9 fl-fr), Anderson et al. 3000 (GH, LL, MICH, MO, US); Parish of St. Andrew, rd. Gordon Town-Guava Ridge, 3 Jul 1966 (9 fl-fr), Anderson et al. 3001 (MICH, US), (6), Anderson et al. 3010 (GH, MICH, US); Parish of Portland, rd. Section-Hardware gap, 3 Jul 1966 (6), Anderson et al. 3020 (MICH, US); Cockpit Coun- try, Troy, 13-18 Sep 1906 (9 fl-fr), Britton et al. 508 (NY); nr. Nigril, 9-12 Mar 1908 (9 fl-fr), Britton et al. 2048 (NY); Nary Is., 26 Jul 1897 (9 fl), Fredholm 3266 (NY); Parish of St. Ann, ca. 1 km N of Hollymount, Mt. Diablo, 6-7 Aug 1965 (9 fr), Hespenheide et al. 874 (GH, LL, MICH, MO); Parish of St. James, Irwin, 30 Apr 1952 (6), Johnson et al. 22J181 (IJ); Parish of St. Elizabeth, Giddy Hall, Apr 1926 (9 fl), Maxwell s.n. (BM); Arntully, 26 Sep 1927 (9 fl-fr), Orcutt 3030 (G, GH, K, S, US); Parish of St. Mary, nr. Hampstead, 1954 (9 fl-fr), Proctor 8692 (IJ).
GRENADINES. Becquia, (d fl), D. Joseph (in H. H. Smith et al.) 13100 (K).
BARBADOS. St. John, Bath Woods, Mar 1940 (6), Gooding 331 (BM).
TRINIDAD. Roxborough-Bloody Bay rd., 1 Mar 1981 (juv), Baksh et al. 321 (NY); Caura-Royal rd., 15 Feb 1979 (6), Berg 978 (U) & 979 (AAU, BG, U); Arima-Blanchis- seuse rd., ca. km 6 km NNW of Arima, 15 Feb 1979 (9 fl- fr), Berg 984 (BG, U); Aripo Savanna, 19 Aug 1951 (st), Richardson 683 (NY); Maraval, 1891-1892 (st), Warming s.n. (C).
SYSTEMATIC TREATMENT 147
COLOMBIA. ANTIOQUIA: Mun. Taraza, Vrda. Bana- blanco, source of Rfo Puri, 24 Apr 1993 (d), Callejas et al. 10879 (HUA); Rfo Sucio, nr. Uramita, 4 Oct 1961 (9 fl-fr), Cuatrecasas et al. 26213 (COL, US), (d), Cuatrecasas et al. 26214 (P, US); Rfo Cauca, between Puerto Valdivia & Valdivia, 550-1 100 m, 19 Feb 1942 (6), Cuatrecasas et al. 30096 (COL, F, US); 1 km W of Turbo, 31 Mar 1962 (9 fl- fr), Feddema 2112 (MICH, US); Mun. San Luis, Rfo Claro, 25 Mar 1994 (i), Franco et al. 4583 (BG, HUA), (d), Franco et al. 4584 (BG, HUA); Mun. Frontino, nr. Cgto. Nutibara, 1200 m, 5 Mar 1995 (c), Franco et al. 5561 (BG, COL, HUA, LP, MO, NY, QCA); Mun. Anorf, Vrda. El Ro- ble, 1400 m, 8 Mar 1995 (9 fl-fr), Franco et al. 5566 (BG, COL, HUA, LP, MO, US); between Villa Artega & Rio Mu- tata, 20 Nov 1948 (d), Johnson et al. 18C240 (MEDEL); Mun. Jeric6, 1200 m, 5 Jul 1983 (c), Ortiz S. et al. 110 (MEDEL); Vrda. La Rubiela, rd. Segovia-Antioquia, 19 Jul 1979 (9 fl-fr), Renterfa et al. 1688 (COL, NY); Mun. Fre- donia, Cgto. Marsella, 14 Apr 1992 (9 fl-fr), D. Sdnchez S. 1760 (MEDEL); Mun. Zaragoza, Cgto. de Providencia, 9 Feb 1971 (6), Soejarto et al. 2662 (COL, GH); nr. Medellfn, 10 Sep 1927 (9 fl-fr), Toro 586 (NY); Mun. Caucasia, rd. to Nechf, 17 km from rd. Caucasia-Planeta Rica, 22 Mar 1987 (9 fl-fr), Zarucchi et al. 4915 (BG, COL, MO). AR-
CHIPELAGO DE SAN ANDRES Y PROVIDENCIA: Isla de Prov- idencia, Mar-Jul 1977 (6), Freeman 35 (MEDEL). ATLANTICO: Nr. Barranquilla, Aug 1928 (9 fl-fr), Hno. Elias 578 (US), Jul 1934 (9 fr), Hno. Elias 1230 (F, MICH, US). BOLiVAR: Mun. Turbaco, 8 Jun 1982 (6), Cuadros V 1372 (MO), (9 fl), Cuadros V 1383 (MO); nr. Turbaco, 29 Jul 1953 ( 9 fr), Dugand et al. 3357 (COL, F); nr. San Martfn de Loba, Apr-May 1916 (6), Curran 65 (GH, S, US); Mun. San Juan Nepomuceno, Loma de Los Colorados, ca. 2 km S of San Juan, 7 May 1987 (st), Gentry et al. 57488 (BG, MO); Torrercilla, nr. Turbaco, 7-19 Nov 1926 (st), Killip et al. 14243 (GH, NY, US); Mun. Cartagena, 7 km SW of Arroyo Grande, 31 Jul 1985 (9 fl-fr), Zarucchi et al. 3884 (BG, MO, NY). CALDAS: Nr. La Dorada, 1-2 Feb 1946 (6), Duque-Jaramillo 2590 (NY); Quebrada Yeguas, 20 km N of Honda, 7 Mar 1977 (9 fl-fr), Gentry et al. 18194 (COL, MO); Pan-American Hwy., nr. Marmato turnoff, NE of Rfo Sucio, 27 Mar 1980 (6), Gentry 28801 (AAU, MO, U, US). CESAR: Rd. La Paz-Manaure, 26 Apr 1987 (6), Cuadros V et al. 3402 (BG); Sierra de Perija, E of Manaure, El Podrido, 1550-1600 m, 15 Nov 1959 (6), Cuatrecasas 25356 (US); rd. San Martfn-Ocaina, Quebrada del Gobernador, 27 Sep 1969 (d), Cuatrecasas et al. 27965 (COL, NY, US), (9 fl- fr), Cuatrecasas et al. 27968 (COL, NY, US); 5 km W of Manaure, 13 Jan 1988 (st), Gentry et al. 60723 (MO). CHOcO: Cerros del Cuchillo, Rosa Marfa, 16 Oct 1987 (9 fl), Cdrdenas 679 (JAUM, MO); Mun. Riosucio, Cerros del Cuchillo, rd. Cuchillo Negro-Punta de las Flores, 18 May 1988 (9 fl-fr), Cardenas 1934 (JAUM, MO); Mun. Pizarro, rd. Pie de Pep6-Berrecul, km 21-24, (9 fl-fr), Espina 1920 (COL, MO); nr. Bahfa Solano, 4 Apr 1990 (9 fl-fr), Franco et al. 3001 (COL); Rfo San Juan, above Istmina, 14 Aug 1976 (6), Gentry et al. 17664 (AAU, BG, MO, US); Mun. Acandf, Cgto. de Ungufa, Rfo Cuti, 27 Jul 1957 (6), Romero-Castanteda 6449 (COL, MO, NY); Rio Jurad6, 4
Oct 1940 (6), Sneidern A.246 (MICH, NY, S, UA); Parque Nacional Natural Los Katios, Sautata, 3 May 1983 (9 fl), Zuluaga R. 637 (COL). C6RDOBA: Planeta Rica, rd. Mon- teria-Planeta, El Charco, 24 Oct 1987 (9 fl-fr), Leguizamo 493 (MO), (6), Leguizamo 494 (MO). CUNDINAMARCA: 9 km E of Melgar, on rd to Fusagasuga, 8 Jul 1972 (9 fl), Barclay et al. 3557 (COL, US); Mun. Melgar, Rio Sumapaz, nr. La Nariz del Diablo, 12 Mar 1993 (9 fl), Franco et al. 4346 (COL, MO), (d), Franco et al. 4347 (BG, COL). GUA- JIRA: Between Cuestecita & Carraipia, Rio Cesar, 30 Nov 1959 (d), Cuatrecasas et al. 25522 (COL), (9 fl-fr), Cua- trecasas et al. 25535 (COL, US); 15 km S of Mingueo, trail to Pueblo Viejo, 21 Aug 1986 (st), Gentry et al. 55425 (MO); Serrania La Macuira, Cerro Manzano, 5 Mar 1963 (juv), Saravia T 2419 (COL, US). HUILA: Between Gigante & Potrerillo, 1 Nov 1926 (9), Juzepczuk 6710 (LE); nr. Neiva, 7 Apr 1944 (9 fl-fr), Little et al. 7595 (COL, F, P); between Neiva & Campoalegre, 19 Mar 1940 (9 fr), Perez Arbelaez et al. 8301 (COL, F). MAGDALENA: Rd. to Rio- hacha, km 62, 17 Apr 1982 (9 fl), Cuadros V 1352 (COL, MO); Sierra Nevada de Santa Marta, Rio Donachuf, below Donachui, 1230-1350 m, 24 Sep 1959 (9 fl-fr), Cuatrecasas et al. 24419 (COL, US); Sierra de Perija, Manaure, Rio Manaure, 4 Nov 1959 (d), Cuatrecasas et al. 25020 (COL); Rio Buritaca, Alto de Mira, 1100 m, 20 Jul 1989 (6>), Mad- riniin et al. 406 (MO); Santa Marta, 6 Apr 1899 (d), H. H. Smith 2105 (BM, COL, E, G, GH, K, LE, LL, MO, NY, P, S, U, US); 13 km SE of Santa Marta, 8 May 1977 (6), White et al. 438 (COL, HUA, MO, NY). NORTE DE SANTANDER:
Between Gramalote & Peralonso, 24 Jul 1940 (9 fr), Cua- trecasas 10140 (COL, F); Rio Pamplonita, between Cdcuta & Pamplona, 25 Jul 1940 (9 fr), Cuatrecasas 10178 (COL, F). SANTANDER: 5 km S of Bucaramanga, 14 Jul 1968 (9 fl-fr), Barkley 38C212 (GH); Mun. Piedecuesta, Cgto. Pes- cadero, Vrda. Mesetas, Rio Umpala, 12 Jul 1996 (9 fl-fr), Cadena et al. 3008 (COL); RIo Chicamocha, between Cap- itanejo & Enciso, 1200-1300 m, 18 Jul 1940 (9 fr), Cua- trecasas 9843 (COL, F); Rio Pienta, between El Limite & Encino, 2000 m, 4 Aug 1940 (9 fl), Cuatrecasas 10437 (COL, F), (6), Cuatrecasas 10438 (COL, F); Mun. Pie de Cuesta, La Mesa de los Santos, 1460-1700 m, 16 May 1969 (6), Garcia-Barriga 19686 (COL, NY); between Puerto Wilches & Puerto Santos, 29 Nov 1926 (6), Killip et al. 14892 (A, GH, NY, US); Mesa de los Santos, 1000-1500 m, 11-15 Dec 1926 (9 fl-fr), Killip et al. 15010 (A, GH, NY, US); 12 mi SE of Barranca Bermeja, 15 Sep 1954 (9 fl-fr), Romero-Castaneda 4847 (COL, US). TOLIMA: Be- tween Mariquita & Guayabal, 7 May 1940 (9 fl-fr), Cua- trecasas 9410 (COL, F); Mun. Mariquita, rd. Mariquita- Honda, 17 May 1990 (9 fl), Franco et al. 2968 (COL, MO); rd. Espinal-Ibague, nr. Rio Coello, 6 Feb 1955 (6), Franco et al. 4646 (BG, COL); Mun. Piedras, rd. Piedras-Doima, km 5, 7 Feb 1995 (9 fl-fr), Franco et al. 4649 (COL); 11 km NE of Melgar, Rfo Sumapaz, 8 Jan 1974 (9 fl-fr), Gentry et al. 8958 (COL, GH, MO); between Colache & Coyaima, 19 Oct 1926 (6), Juzepczuk 6928 (LE). VALLE: Buenaven- tura, 16 Mar 1994 (9 fl-fr), Franco et al. 4536 (BG); rd. to Bajo Calima, Bahia Malaga, 16 Mar 1994 (6), Franco et al. 4542 (BG, TULV), (6), Franco et al. 4543 (BG, TULV).
148 FLORA NEOTROPICA
VENEZUELA. AMAZONAS: Depto. Atabapo, Rfo Ocamo, nr. Raudal Arata, Jan 1990 (d), Ferndndez 6533 (BG). ANZOATEGUI: Cantaura, 19 Apr 1950 (d), F D. Smith 141 (US). ARAGUA: Parque Nacional Henri Pittier, Rancho Grande, 11 Aug 1953 (Y fr), Little 15451 (NY, VEN), (6), Little 15452 (NY, VEN); nr. Maracay, 2 Apr 1926 (6), Pittier 12143 (G, NY, US); rd. San Casimiro- Camatagua, Aug 1966 (6), Veldsquez 86 (US, VEN); Ran- cho Grande, Aug 1966 (Y fl-fr), Veldsquez 94 (US), (6), Vela'squez 95 (US, VEN); Guamites, 12 May 1938 (Y fr), Ll. Williams 10074 (F, VEN). BARINAS: Mun. Pedreza, 40 km SE of Ciudad Bolivia, 24 Apr 1953 (Y fl), Little 15119 (VEN), 25 Apr 1953 (d), Little 15120 (VEN); Mun. Cruz Paredes, Dtto. Obispos, 20 km NW of Barrancas, 26 Mar 1932 (6), Marcano Berti et al. 2963 (VEN); rd. Merida- Barinas, between La Yuca & Barinitas, 21 May 1980 (6), Marcano Berti et al. 109-980 (G, U, VEN); rd. M6rida- Barinas, between La Yuca & Barinitas, 11 May 1980 (Y fl- fr), Marcano Berti et al. 113-980 (G, U, US); rd. Barinas- San Crist6bal, Canagua, 21 May 1980 (Y fl-fr), Marcano Berti et al. 114-980 (G, U, US, VEN); rd. Barinas-San Cris- tobal, between Socop6 & Capitanejo, 22 May 1988 (6), Marcano Berti 115-980 (G, U, VEN); rd. Barinas-Acarigua, Dec 1966 (9 fl-fr), Veldsquez 102 (US, VEN). BOLiVAR:
Dtto. Cedefno, nr. Corozal, 13 Apr 1986 (6), Boom et al. 6493 (NY); Dtto. Roscio, San Martin de Turumban, Aug 1979 (9 fl-fr), Delascio et al. 7845 (VEN); Mun. Raul Leoni, 86 km E of San Francisco, Jun 1989 (9 fl-fr), Del- gado 241 (BG, MO, NY, VEN); 20-35 km SW of El Man- teco, rd. to San Pedro de las Dos Bocas, 1-3 Aug 1978 (9 fl-fr), Liesner et al. 5815 (MO, U, VEN); Reserva Forestal Imataca, nr. confluence of Rio Botanamo & Rio Corumo, 19-20 Jan 1983 (9 fl), Stergios et al. 5372 (US); Rio Asa, above Raudal Cotua, S of La Paragua, 1 Aug 1960 (9 fl-fr), Steyermark 86723 (NY, US); Rio Caura, Salto Para, 15 Jan 1977 (6), Steyermark et al. 113020 (F, K, MO, U, VEN); Rio Uairen, Gran Sabana, Santa Elena, Feb 1946 (6), Ta- mayo 2981 (F, G, S, VEN); Reserva Forestal del Imataca, Aug 1967 (9 fl-fr), Veldsquez 252 (US); El Palmar, 25 Apr 1940 (9 fl-fr), Ll. Williams 12893 (F, K, S, US, VEN). Co- JEDES: Dtto. Giradot, Hato Piniero, rd. to Charco Azul, 6 Jul 1990 (9 fl-fr), Aristeguieta et al. 1482 (VEN); El Mapuey, 26 Sep 1975 (9 fr), Delascio et al. 3446 (VEN). COLON: San Crist6bal, 5 Jan-22 Feb 1923 (d), Broadway 259 (GH, NY, US). DELTA AMACURO: Rd. Tucupita-Los Giiires, 17 Apr 1973 (9), Agostini et al. 1624 (U); Lower Rio Orinoco, Manoa, May 1896 (6), Rusby et al. 99 (A, BM, F, G, K, MO, NY, US, VEN); Rio Cuyubini, between La Paloma & Moron, 20 Nov 1960 (d), Steyermark 87693 (VEN). Dis- TRITO FEDERAL: Rd. Caracas-La Guaira, 22 Mar 1922 (d),
rd. El Sombrero-Calabozo, 8 Aug 1966 (Y fl-fr), Veldsquez 87 (US, VEN), (6), Veldsquez 88 (US, VEN). MERIDA: Be- tween Ejido & Mesa de los Indios, 1200 m, 23 Apr 1953 (6), Bernardi 416 (NY); Dtto. Justo Briceilo, between La Panamaricana & Las Virtudes, 11 May 1978 (6), Bunting et al. 6341 (U); rd. Merida-Tovar, between Ejido & Lagun- illas, 850 m, 10 Apr 1969 (6), Oberwinkler et al. 15607 (VEN); nr. Merida, Feb 1968 (6), Veldsquez 263 (US, VEN). MIRANDA: Dtto. Paez, rd. San Juan-Montevideo, 7 Sep 1977 (Y fr), A. Gonzales et al. 1405 (MO, VEN); Alto de Guayabo, rd. Caracas-Cua, km 17, 17 Apr 1924 (Y fl), Pittier 11511 (A, G, K, NY, P, US, VEN); Parque Nacional de Guatopo, Santa Cruz, between Santa Teresa & Altagracia de Orituco, 23 Nov 1961 (Y fl-fr, Steyermark 89976 (NY, US, VEN); El Guapo, Aug 1965 (? fl-fr), Veldsquez 78 (US, VEN), (6), Veldsquez 79 (US, VEN); Parque Nacional Gua- topo, Oct 1966 (6), Veldsquez 97 (US, VEN); Panaquire, Jul 1966 (Y fl-fr), Velasquez 247 (US, VEN). MONAGAS: Rd. Jusepin-Caicara de Maturfn, 22 Aug 1970 (6), Ariste- guieta et al. 7531 (VEN); Reserva Forestal de Guarapiche, 22 Feb 1978 (Y fl), Castillo 711 (NY, VEN); Dtto. Caripe, Cerro de Los Rastrojos, Quebrada Las Majaguas, 13 May 1982 (Y fr), 0. Huber 6348 (NY, U); 2 km SSW of Jusepin, 5 Mar 1967 (d), Pursell et al. 8241 (VEN); Caicaro, 12 Jun 1940 ( fr), Ll. Williams 13300 (F, US). NUEVA ESPARTA: Isla de Margarita, Jul 1984 (Y fr), Delascio et al. 12225 (MO, VEN); Isla Margarita, Cerro Copey, 25 Apr 1983 (Y fl-fr), Sugden 1182 (K, MO, VEN), 2 May 1983 (d), Sugden 1204 (K, MO). PORTUGUESA: Dtto. Guanare, grounds of UNELLEZ, 21 Jan 1982 (9 fr), Aymard et al. 811 (MO); Dtto. Sucre, Caserfo Villa Rosa, 20 km E of Buscucuy, 1200-1500 m, 16 Jun 1985 (9 fl), Aymard et al. 3655 (US); Dtto. Ospino, El Corozo, Rfo Guache, 22 May 1983 (d), Delgado 20 (MYF). SUCRE: Penfnsula de Paria, Cerro Pa- tao, 6 Sep 1984 (d) Milliken et al. 159 (MO); Penfnsula de Paria, Cerro Patao, NE of Giuiria, 23 Jul 1962 (9 fl-fr), Stey- ermark et al. 91272 (US, VEN); rd. Rfo Caribe-Tunapui, Feb 1967 (9 fl-fr), Veldsquez 98 (US), VEN). TACHIRA: 7 km W of Rubio, 18 Mar 1981 (d), Liesner et al. 10737 (MO, U, VEN); Rfo Frio, between jct. of Rio Quinimari & Rfo Frio, between (village) Rfo Negro & La Laguna, 30 Jul 1979 (d), Steyermark et al. 119082 (MO, U, VEN); Quebrada Colorada, between San Juan de Col6n & Las Cruces, at La Pocina, 14 Nov 1979 (6), Steyermark et al. 120308 (MO, NY, U). YARACUY: Rfo Yurubf, 13 Sep 1974 (6), Delascio et al. 2578 (VEN); Salom, 18 Feb 1954 (9 fl), Little 16215 (VEN). ZULIA: Dtto. Col6n, between Casigua El Cubo & km 8 on rd to Palmira, 28 Apr 1979 (9 fl), Bunting et al. 7323 (U); Dtto. Bolfvar, between Quir6s-El Pensado & Cerro Socope, 5-8 Aug 1980 (9 fl-fr), Bunting 9853 (NY, U, VEN); Dtto. Perija, nr. Estaci6n Hidrol6gica Aricuaisa- Pie de Monte, 1-3 1982 (9 fl-fr), Bunting et al. 11458 (NY, U); Mun. Santa Barbara, Dtto. Col6n, 17 Feb 1981 (9 fl- fr), Marcano Berti et al. 146-981 (G, U, VEN); Rio del Pal- mar, San Martin, 15 Oct 1922 (9 fr), Pittier 10518 (G, NY, US); Dtto. Mara, 2.5 km W of Corpozulia Campamento Cur- ichuano, 3 Jun 1980 (9 fl-fr), Steyermark et al. 123221 (MO, NY, U, VEN); Sierra de Perija, rd. La Matera-El Par-
SYSTEMATIC TREATMENT 149
amito, San Jose de Los Altos, 8 Jul 1975 (9 fr), Zambrano 222 (VEN).
GUYANA. Demerara R., 7 Apr 1987 (d), Boom 7190 (BG, NY, U); nr. Bartica, 30 Jan 1951 (6), Fanshawe 3033 = FD 6363 (K, NY, U); Essequibo Ils-W. Demerara Region, Naamryck Canal, ca. 3.5 km SW of Parika, 14 Apr 1989 (9
fr), Gillespie 995 (BG, U); U. Takatu-U. Essequibo Region, ca. 15 km S of Sand Creek Vill., 21 Jun 1989 (9 fl-fr), Gillespie et al. 1729 (BG, NY); Potaro-Siparuni region, Pak- araima Mtns., E slope of Malakwalai-Tipu, 1 100 m, 10 Jul 1994 (d), Henkel et al. 5542 (BG); Kanuku Mtns., Mai- paima, 17 Nov 1987 (9 fl-fr), Jansen-Jacobs et al. 971 (BG, K, MO, NY, U); Rupununi Distr., Kusad Mtns., 27 Sep 1992 (9 fl-fr), Jansen-Jacobs et al. 2658 (BG, MO, NY, U); Ber- bice R., S of New Dageraad, 6 Oct 1981 (9 fl), Maas et al. 5571 (F, K, MO, NY, U); Kanuku Mtns., Moku-moku Cr. 31 Mar-16 Apr 1938 (9 fl-fr), A. C. Smith 3426 (A, F, G, K, MO, NY, P, S, U, US).
SURINAME. Wilhelmina Mtns., Kayser Airstrip, 4.5 km above confluence of Zuid R. & Lucie R., 29 Sep 1963 (9 fl), Irwin et al. 57681 (US); Waneweg, 30 km S of Paramaribo, 9 Dec 1960 (d), Kramer et al. 2317 (NY, U); Coppename R., Nature Reserve Voltzberg, 25 Feb 1977 (9 fl-fr), Lindeman & Mennega et al. 150 (K, MG, NY, RB, U); Sipaliwini savanna, nr. Brazilian frontier, 25 Sep 1970 (st), Oldenburger et al. 1362 (U); Zanderij, 3 Jul 1916 (d), Samuels 506 (F, GH, K, NY, P); Afobaka rd. 10 km S of Paranam, 1 May 1970 (6), Teunissen (LBB) 12739 (U).
BRAZIL. PARA: Parque Nacional do Tumucumaque, Rio Paru do Oeste, Missao Tiriyo, 16 Feb 1970 (d), Cav- alcante 2403 (NY, U, US), 20 Feb 1970 (9 fl-fr), Cavalcante 2475 (NY, U, US). RORAIMA: Ilha de Maraca, 12-26 Jul 1986 (st), Campbell et al. 15832 (BG); Ilha de Maraca, SEMA Ecological Reserve, 7 Jul 1987 (6), Milliken 408 (BG, E, K).
In Jamaica, some collections have trichilia, others have vestigial ones, and in several collections they are absent. Material without trichilia or vestigial ones can be distinguished from Cecropia schreberiana var. an- tillarum (found in other Caribbean islands) by the scarce and white pith in the intemodes and the ab- sence of relatively long and soft hairs on the veins of the lamina beneath. Trichilia are present in the col- lection from the Swan Islands and in all collections made elsewhere. Several collections, e.g., Jansen- Jacobs et al. 971 and 1260 from Guyana, and Fer- ndndez 6533 from Venezuela (Amazonas), have long white hairs in the trichilia; they resemble C. metensis in this respect, but lack the dense arachnoid indumen- tum on the petiole.
The species has been introduced into several coun- tries in Africa, Asia, and the Pacific, and also to the Bermuda Islands (Brown et al. 1683, NY). It has be- come naturalized in several countries in West Africa, at least in Cameroun (R. Letouzey 12474 and 14828,
P; S. A. Thompson et al. 1398, MO), Ghana (H. H. Schmidt et al. 2032, MO), Ivory Coast (E. Merklen s.n., US; Leeuwenberg 4504, MO; Miege & Ake Assi 3960, G, P; Tere HGT 2155, G), and Senegal (Berhaut 984, P); also French Polynesia (Makatea, Raiatea, Rapa, Tahaa, and Tahiti, according to Flor- ence [1997: 24-26, fig. 1]), for the first time collected in 1927; and Malaysia (Mat Asri FRI 25549, A; see also Putz & Holbrook, 1988). In Cameroun, it is com- peting with Musanga cecropioides (McKey, 1988a). The species has been or is still present in some bo- tanical gardens: China (Guangzhou, according to Liao [1991, 1992], and Xishuangbanna), Gabon (Libre- ville, 1900, C. Chalot 48, P), Singapore (1930, C. X. Furtado s.n., A), Indonesia (Bogor, 1952, Anonymous s.n., A), and Taiwan (Shiaping Tropical Botanical Garden, Chishan, according to Liao [1991, 1995]; in- troduced from El Salvador and with trichilia, see Liao, 1991: 131, fig. 5, photo. 1; and Liao, 1995: 134, fig. 5, photo. 1). For the material in the Botanical Garden of Bogor, Brazil was indicated as the prove- nance. According to Putz & Holbrook (1988) the ma- terial in Malaysia is introduced from Indonesia (Bo- gor). If the annotation on the Bogor material about the provenance is correct, the material should belong to Cecropia pachystachya, but the collections exam- ined do not show some of the characteristic features of that species. Moreover, C. pachystachya has never been encountered without trichilia. Thus, the infor- mation about origin or the identity of this material is dubious. In material from Africa trichilia are absent or poorly developed. In French Polynesia they may often be lacking. In Malaysian material examined by Putz & Holbrook (1988), 80% of the specimens had fully (fused) developed trichilia, 8% poorly (separate) developed ones, and 12% none. The intriguing ques- tion is whether all this material has as its origin Ja- maica, the only area in which C. peltata is found with and without trichilia, or whether loss of the ability to form (well-developed) trichilia was developed after introduction elsewhere (in two species?). The latter case would confirm the hypothesis of dissolution of the mutualism between Cecropia and Azteca due to the absence of the latter partner and manifest in the absence of trichilia (Janzen, 1973).
42. Cecropia pittieri Robinson, in Stewart, Proc. Ca- lif. Acad. Sci., ser. 4, 1: 389. 1912; Burger, Fiel- diana, Bot. 40: 127. 1977. Type. Costa Rica. Co- cos Is., Jan 1902 (d), Pittier 16237 (holotype: GH).
Tree, to 20 m tall. Leafy twigs 2.5-4 cm thick, (sub)hispudulous with straight hairs. Lamina charta- ceous, ca. 30 X 30 cm to 60 X 60 cm, the segments 10-11, the incisions down to 2/10-3/10; apices rounded to subacuminate; upper surface scabrous, hispidulous; lower surface hirtellous to subhirsute (to subtomentose) on the veins, with arachnoid indumen- tum in the areoles and on the reticulum; lateral veins in the free part of the midsegment ca. 10-12 pairs, submarginally to marginally loop-connected, often branched; petiole 20-50 cm long, with arachnoid in- dumentum, at base and apex hirtellous to subhirsute; trichilia lacking; stipules 8-17 cm long, color un- known, hirtellous to subhirsute and with arachnoid indumentum outside, sericeous inside. Staminate in- florescences in pairs (?), the peduncle erect and the spikes pendulous (?); peduncle 8-9 cm long, puber- ulous to hirtellous and with arachnoid indumentum; spathe ca. 15 cm long, color unknown, hirtellous to strigose to hirsute outside, sparsely hairy inside; spikes ca. 10-12, ca. 10-12 X ca. 0.3 cm, with stipes 0.6-0.8 cm long and sparsely hairy; rachis hairy. Sta- minate flowers: perianth tubular, 1-1.5 mm long, sparsely minutely puberulous below the apex, the apex almost plane; anthers 0.5-0.6 mm long, de- tached at anthesis (?). Pistillate inflorescences in pairs, pendulous; peduncle 5-10 cm long, sparsely puberulous to hirtellous and also with arachnoid in- dumentum; spathe not seen; spikes 4, to 21 X 1-1.2 cm in fruit, sessile; rachis glabrous. Pistillate flowers: perianth ca. 2 mm long, with arachnoid indumentum below the apex outside, the apex convex, muriculate; stigma comose. Fruit ellipsoid to ovoid, ca. 2 mm long, slightly tuberculate, dark brown.
Distribution (Fig. 8.5). Endemic of Cocos Island.
Specimens examined. COSTA RICA. Cocos ISLAND:
Bahfa de Chatham, 13 Aug 1973 (Y fr), Carrasquilla 362 (MO, NY), 13 Aug 1973 (9 fl-fr), Dressler 4460 (NY, PMA, US), 8 Mar 1964 (9 fl-fr), Fournier 311 (NY), 13 Apr 1965 (9 fl-fr), Jimenez 3184 (COL, MO, U); Bahfa de Wafer, 12 Oct 1994 (9 fl-fr), Queseda 1064 (BG), Bahia de Chatham, 1905-1906 (juv), Stewart 291 (GH).
This species is interesting because of its limited distribution and the absence of trichilia. It is not in- habited by ants and is probably related to Cecropia longipes.
43. Cecropia plicata Cuatrecasas, Revista Acad. Col- omb. Ci. Exact. 6(22/23): 280, t. 1. 1945. Type. Colombia. Valle: Rfo Sanquinini, La Laguna, 1250-1400 m, 10-20 Dec 1943 (9), Cuatrecasas 15469 (holotype: COL; isotypes: F, US).
Cecropia diguensis Cuatrecasas, Revista Acad. Colomb. Ci. Exact. 9(36/37): 332. 1956. Type. Colombia. Valle: Rio San Juan, below Queremal, 1300-1500 m, 24 Mar 1947 (d), Cuatrecasas 23967 (holotype: US; isotype: F).
Tree, to 20(-30) m tall. Leafy twigs 2-4 cm thick, dark green to brown or white sparsely puberulous, also with ? dense pluricellular hairs and ? dense arachnoid indumentum. Lamina coriaceous and often ? plicate, ca. 30 X 30 cm to 75 X 75 cm, the seg- ments 8-10(-l 1), the free parts of the upper ones el- liptic to oblong to subobovate, the incisions down to 6/10-8/1 0; apices rounded (or acuminate); upper sur- face smooth, sparsely strigillose (on the main veins) or subglabrous; lower surface sparsely minutely pu- berulous and with ? dense brown pluricellular hairs, with arachnoid indumentum lacking or sparse (in minute patches) in the small areoles (surrounded by thick veinlets); lateral veins in the free part of the midsegment 15-24 pairs, submarginally loop- connected, unbranched or some branched; petiole ca. 25-70 cm long, sparsely minutely puberulous and with ? dense brown pluricellular hairs and rather sparse arachnoid indumentum; trichilia fused (or sep- arate), the brown indumentum intermixed with short whitish hairs; stipules 12-33 cm long, dark red-brown to purplish, puberulous to hirtellous and with ? dense brown pluricellular trichomes outside, glabrous or (sub)sericeous inside. Staminate inflorescences solitary or in in pairs, erect; peduncle 5-13 cm long, dark red to red-brown, (sparsely) puberulous or to hir- tellous, also with brown pluricellular hairs and arach- noid indumentum; spathe 12-21 cm long, dark red to red-brown, ? sparsely puberulous to strigose to hir- tellous and with dense brown pluricellular hairs outside, glabrous or in the upper part (sub)sericeous inside; spikes 5-9, 11-18 X 0.7-1.5 cm, with stipes 0.5-2.5 cm long, red, and appressed-puberulous or glabrous; rachis hairy. Staminate flowers short- pedicellate; perianth tubular, 2-2.5 mm long, puber- ulous (or also with arachnoid indumentum) below the apex and on the pedicel, the apex convex to plane, hispidulous; filaments swollen; anthers 0.6-0.8 mm long, appendiculate, detached at anthesis, reattached to the margins of the aperture by the appendages after anthesis. Pistillate inflorescences in pairs or solitary, erect, deflexed in fruit; peduncle 4-8 cm long, dark red or partly whitish, puberulous to subhirsute, also
SYSTEMATIC TREATMENT 151
with brown pluricellular hairs and arachnoid indu- mentum; spathe 9-11 cm long, the color and indu- mentum as in the staminate inflorescence; spikes 2- 3(4), 4.5-11 X 0.6-1.2 cm, to 25 X 3 cm in fruit, (sub)sessile; rachis hairy. Pistillate flowers: perianth 2-3 mm long, with arachnoid indumentum below the apex outside, absent inside, the apex convex, muri- culate to hispidulous; style rather long, muriculate; stigma comose. Fruit subovoid, ca. 3 mm long, smooth, pale brown.
Distribution (Fig. 11.1). Colombia, from Antio- quia to Valle, in (sub)montane forest, at 1000-2300 M.
Representative specimens examined. COLOMBIA. ANTIOQUIA: Don Matfas, 2100 m, 27 Mar 1994 (d), Franco et al. 4592 (BG, HUA); nr. Yarumal, 2050 m, 27 Mar 1994 (Y fl-fr), Franco et al. 4593 (BG, HUA); Mun. Urrao, Parque Nacional Natural Las Orquideas, Quebrada La Agu- delo, 1300-1380 m, 6 Jul 1992 (st), Ram[rez et al. 4151, 4184, 4242 (JAUM). CHocO: Mun. San Jose del Palmar, Vrda. El Sinai, 1650 m, 19 Mar 1994 (d), Franco et al. 4563 (BG, HUA); Mun. San Jose del Palmar, ca. 1 km E of San Jose del Palmar, 1250 m, 19 Mar 1994 (? fl-fr), Franco et al. 4570 (BG, HUA); Mun. Frontino, Cgto. Nutibara, rd. Nutibara-La Blanquita, Murri region, 1900 m, 4 Mar 1995 (Y fl-fr), Franco et al. 5549 (BG, COL, HUA), 1500 m, 4 Mar 1995 (d), Franco et al. 5554 (BG, COL, HUA). RIs- ARALDA: Mun. Pueblo Rico, rd. to Cerro Monctezuma, 1350 m, 15 Mar 1986 (Y fl-fr), Bernal et al. 997 (COL, MO, NY). VALLE: Mun. Darien, Lago Calima, 1400 m, 15 1994 ( fl-fr), Franco et al. 4529 (BG, TULV).
This species appears to be very closely related to Cecropia maxima, from which it differs in general in the smaller dimensions of several parts (such as lam- ina, stipules, spikes of pistillate inflorescences) and the denser indumentum of brown pluricellular hairs. Cecropia plicata can be distinguished from C. maxi- ma by the smaller number of segments of the lamina (8-10 vs. 10-15) and by the arachnoid indumentum on the lamina beneath, this being confined to the ar- eoles and not present on the main veins. Staminate specimens may be consistently distinct in the pres- ence/length of the stipes of the spikes. The nature of the differences are such that they could justify sepa- ration only at the subspecific level. The two taxa ap- pear to be allopatric. Cecropia plicata also resembles C. insignis, particularly specimens with 8 lamina seg- ments. Such specimens differ from those of C. plicata in the presence of arachnoid indumentum on the smaller veins and at least initially also on the main veins on the lamina beneath. Pistillate specimens can be distinguished by the difference in the number of spikes: usually 2-3 in C. plicata and mostly 5 in C.
insignis. Overlap of the elevational ranges of the two species has not yet been found in Colombia. Ants are often present.
Cecropia nivea Poeppig ex Klotzsch, Linnaea 20: 532. 1847 (Oct). Type. Peru. Without locality (or Huan- uco: Cuchero), (5), Poeppig s.n. or 1593? (holotype: B, destroyed, photographs in F, MO; isotype?: LE).
Cecropia pinnatiloba Klotzsch, Linnaea 20: 533. 1847 (Oct); Miquel in Martius, FH. Bras. 4(1): 153. 1853. Type. Peru. Without locality, (d + Y), Ruiz & Pavon s.n. (syntypes: B, destroyed, photograph in F; Y, K).
Cecropia leucophaea Poeppig ex Miquel, in Martius, Fl. Bras. 4(1): 151, t. 50, f. 1. 1853. Type. Peru. Huan- uco: Cuchero, (5), Poeppig s.n. or 1593 (W, de- stroyed, fragment in F, photographs in F, MO).
Cecropia klotzschiana Miquel, in Martius, Fl. Bras. 4(1): 152. 1853, as synonym of C. scabra Klotzsch.
Cecropia boliviana Cuatrecasas, Revista Acad. Colomb. Ci. Exact. 9(36/37): 335. 1956. Type. Bolivia. Santa Cruz: Prov. Sara, Buenavista, 3 Oct 1925 (Y), Stein- bach 7267 (holotype: F; isotypes: A, G, GH, K, MO, S).
Tree, to 20 m tall. Leafy twigs 3-6 cm thick, green to red-brown with conspicuous lenticels, hispidulous. Lamina subcoriaceous, ca. 40 X 40 cm to 85 x 85 cm (to 100 X 100 cm), the segments 9-11(-12), the free parts of the upper segments elliptic to oblong to subobovate, the incisions down to ca. 5/10-7/10, the larger lobes (especially in subjuvenile material) pin- nately lobed (with the lobes pointing upward in fresh material); apices obtuse; upper surface smooth (to
152 FLORA NEOTROPICA
I 1~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~I
[VQw1 I
10 7 ~
FIG. 39. Cecropia polystachya. 1. Lamina, reduced (Berg 1696). 2. Apex of lamina and venation (Fisher 178). 3. Stipules and bases of petioles with trichilia (Nee 37514). 4. Stipules, young leaf, and base of petiole with trichilium (Berg et al. 1641). 5. Pair of staminate inflorescences with spathes and base of petiole with trichilium (Berg et al. 1638). 6. Staminate inflorescence at anthesis and base of petiole with trichilium. 7. Staminate flower and stamen (Berg 1696). 8. Pair of pistillate inflorescences with spathes and base of petiole with trichilium (Berg et al. 1641). 9. Pistillate inflorescence after anthesis and base of petiole with trichilium (D. N. Smith et al. 3123). 10. Pistillate flower and fruit (Beck 3793).
SYSTEMATIC TREATMENT 153
scabridulous), the "umbilicus" white-hirsute; lower surface on the main veins sparsely hirtellous to pu- berulous to hispidulous (mainly with uncinate to curved hairs) and also with sparse brown pluricellular hairs, with arachnoid indumentum in the areoles and on part of the reticulum; lateral veins in the free part of the midsegment 15-25 pairs, submarginally loop- connected, most of them branched; petiole ca. 35-70 cm long, grayish, sparsely to densely hirtellous to subhispidulous, with brown pluricellular hairs, and with ? dense arachnoid indumentum, + conspicu- ously lenticellate (especially in the lower part); tri- chilia fused, the brown indument intermixed with long white hairs; stipules 20-35(-45) cm long, ca- ducous, red-brown to dark red (or greenish), but often ? grayish due to the indumentum, with ? dense arachnoid indumentum, brown pluricellular hairs, and simple whitish hairs outside, (sub)glabrous inside; terminal buds often curved. Staminate inflorescences in pairs, erect to subpendulous; peduncle 8-14 cm long, hirsute to hispidulous to puberulous, and with arachnoid indumentum; spathe ca. 10-25 cm long, red-brown to dark red or greenish to pinkish, but ? grayish due to the indumentum, + dense arachnoid indumentum), with sparse brown pluricellular hairs, and scattered long white unicellular hairs outside, gla- brous inside; spikes ca. 15-45, 7-18 X 0.2-0.4 cm, with stipes to 1.5 cm long and glabrous; rachis sparsely hairy. Staminateflowers: perianth tubular, 1- 1.5 mm long, glabrous, the apex ? distinctly 2-lobed; filaments flat; anthers ca. 0.6-0.8 mm long, appen- diculate or not, remaining attached to the filament at anthesis. Pistillate inflorescences in pairs, erect, later on the peduncle curved downward and the spikes curved upward; peduncle 6-18(-26) cm long, with indumentum like that of the staminate inflorescence; spathe ca. 8-12 cm long, the color and indumentum as in the staminate inflorescence; spikes (2-)4-8, 5- 11 X ca. 0.4-0.8 cm, to 18 X 1.2 cm in fruit, some- times initially tortuose, subsessile; rachis sparsely hairy. Pistillate flowers: perianth ca. 1.5 mm long, with arachnoid indument below the apex outside, and often also below the style channel inside, the apex convex, muriculate, the aperture slit-shaped; style short; stigma penicillate. Fruit oblongoid, ca. 2 mm long, slightly tuberculate.
Distribution (Fig. 18.4). From Peru to Bolivia and Brazil (Acre and Rondonia), in forest and secondary growth, at elevations to 1400(-1800) m.
Representative specimens examined. PERU. Cuzco: Prov. La Convenci6n, Illapani, 1 Feb 1969 (d), Chavez A. 463, in herb. Vargas C. 21274 (US); Prov. La Convenci6n, Quiteni, 6 Jan 1976 (Y fl-fr), Chavez A. 3389 (MO); Prov.
Paucartambo, rd. Pilcopata-Atalaya, halfway, 24 Oct 1984 (Y fl), Maas et al. 6161 U); Prov. La Convenci6n, rd. Cuzco-Quillabamba, ca. 150 km, 1100-1300 m, 14 Nov 1987 (Y fl), Ntiuez 8578 (MO); Prov. La Convenci6n, Rfo Manguriari, 2 Feb 1991 (9 fr), Nuinez et al. 12793 (BG). HUANUCO: Nr. Tingo Marfa, 23 Jul 1940 (6), Asplund 12446 (S); rd. Huanuco-Tingo Marfa, below Chinchao, ca. 1800 m, 23 Nov 1997 (6), Berg et al. 1735 (BG, COL, MOL); rd. Huanuco-Tingo Marfa, nr. Tres Estrellas, ca. 1 100 m, 24 Nov 1997 (9 fl-fr), Berg et al. 1740 (BG, COL, MOL); ca. 90 km N of Tingo Marfa, rd. to Tocachi, 4 Feb 1984 (9 fl), Gentry et al. 44934 (NY); Prov. Pachitea, ca. 26 km S of Puerto Inca, 18 Aug 1988 (6), Johann et al. 4/5- 18888 (BG), 15 Nov 1988 (9 fl-fr), Johann 2/32-151188 (BG); ca. 90 km N of Tingo Marfa, rd. to Tocachi, 4 Feb 1984 (9 fl-fr), Gentry et al. 44934 (MO, NY). JUNiN: Prov. Tarma, nr. La Merced, 22 Jan 1946 (9 fl), Ferreyra 398 (USM); between San Ram6n & La Merced, 27 Feb 1982 (9
fl-fr), Gentry et al. 35775 (BG, USM); Prov. Chanchamayo, Catarata del Tirol, 3 Feb 1999 (d), Nuiiiez L 85 (BG), (9 fl- fr), Ntiuiez I. 86 (BG); Prov. Satipo, Dtto. Satipo, Zapallal, 31 May 1982 (6), Reynel 480 (MOL); nr. San Ram6n, Funda La Genova, 1300 m, 16 Apr 1989 (9 fl), Reynel et al. 4016 (MO), 9 Sep 1989 (d), Reynel et al. 4417 (MO). LORETO: Prov. Alto Amazonas, rd. to San Ram6n, 22 Sep 1954 (d), Ferreyra 10186 (US, USM). MADRE DE Dios: Prov. Manu, nr. Salvaci6n, 31 Jul 1988 (d), Berg etal. 1612 (BG, Centro de Medicina Andina, Cuzco); Prov. Tahua- manu, Rfo Tahuamanu, 16 Nov 1973 (9 fl), ChdvezA. 1721 (MO); Prov. Tambopata, 15 km ENE of Puerto Maldonado, 1991 (9 fl-fr), Fisher 178 (BG); Parque Nacional Manu, Rfo Manu, Tayakome, 29 Sep 1986 (d), Foster et al. 11526 (US); between Shintuya & Salvaci6n, 31 Oct 1978 (9 fl), Gentry et al. 27376 (BG); Prov. Tambopata, Reserva Tam- bopata, 22 Aug 1990 (6), Reynel et al. 5396 (BG, MO); 39 km SW of Puerto Maldonado, above confluence of Rfo La Torre & Rfo Tambopata, 6 Oct 1985 (9 fl), S. E Smith et al. 785 (NY, US); Prov. Tambopata, Las Piedras, Albergue Cuzco Amaz6nico, 26 Feb 1991 (9 fl-fr), Timand 1539 (BG, MO); Prov. Tambopata, Las Piedras, Quebrada Gamitana, 6 Mar 1991 (6), Timand et al. 1595 (BG, MO, NY). PASCO: Nr. Pozuzo, 5 Feb 1983 (st), Gentry et al. 40079 (BG); Prov. Oxapampa, Cedropampa, 12 km S of Villa Rica, 1350 m, 18 Jan 1983 (9 fl-fr), D. N. Smith et al. 3123 (BG, MO). SAN MARTiN: Prov. Lamas, rd. Lamas-Pamashto, ca. halfway, 4 Dec 1997 (9 fl-fr), Berg et al. 1769 (BG, COL, MOL), (6), Berg et al. 1770 (BG, COL, MOL); Prov. San Martfn, nr. Tarapoto, 31 Aug 1968 (9 fl-fr), Ferreyra 17423 (US, USM); rd. Tarapoto-Juanjui, km 34, 7 Oct 1984 (d), Maas et al. 5981 (BG, U); Prov. Mariscal Caceres, Tocache Nuevo, 23 Jul 1974 (6), Schunke V 7694 (PMA, U). UCAYALI: Nueva Requena, 11 km from Monte Verde, at km 33 on rd. Pucallpa-Lima, 11 Aug 1988 (d), Berg et al. 1638 (BG, COL, MOL); San Miquel de Semuya, S of Campo Verde, 12 Aug 1988 (9 fl-fr), Berg et al. 1641 (BG, COL, K, MOL); jct. of Rio Pachitea & Rio Yuyapichis, SE of Pu- callpa, 13 Oct 1985 (9 fl), Morawetz et al. 18-131085 (BG), 21 Oct 1985 (d), Morawetz etal. 11-211085 (BG); Semuya, 15 Mar 1988 (9 fl-fr), Rfos T 1801 (K).
154 FLORA NEOTROPICA
BRAZIL. ACRE: Between Igarape do Cuyubim & Igar- ape Jocamin, Fazenda Bom Sossego, 27 Sep-7 Oct 1985 (st), Campbell et al. 9720 (BG); Rio Branco, 18 Jun 1980 (5), L. Coelho et al. 1795 (INPA, NY); nr. Sena Madureira, 27 Sep 1980 (Y fl-fr), C. A. Cid Ferreira et al. 2542A (INPA, MG, NY); nr. mouth of Rio Macaua, 5 Aug 1933 (5), Kru- koff 5307 (A, BM, F, G, K, MICH, MO, NY, S, U, US). ROND6NIA: Ji-Parana, Ouro Preto, 23 Jun 1981 (Y fl), Fearnside 1019 (INPA); rd. Ariquemes-Ouro Preto, km 136, 21 Oct 1979 (d), Vieira et al. 554 (INPA, NY, U).
BOLIVIA. BENI: Prov. Ballividn, rd. Caranavi-San Borja, Serranfa del Pil6n Lajas, 30 Oct 1989 (5), D. N. Smith 13772 (BG, MO); between Trinidad & Misiones Guarayos, Sep 1926 (? fl), Werdermann 2588 (LPB, MO, S); Prov. Ballivian, Rio Beni, nr. Rurrenabaque, 15 Sep 1989 (Y fl), D. E. Williams 987 (BG). COCHABAMBA: Prov. Cha- pare, Villa Tunari, 30 Dec 1982 (Y fl), Ferndndez C. et al. 7907 (G, NY); Prov. Chapare, Villa Tunari, 24 Nov 1981 (Y fl-fr), Va'zquez Avila 425 (U). LA PAZ: Prov. Larecaja, rd. Caranavi-Guanay, km 26.8, 28 Nov 1980 (Y fl-fr), Beck et al. 3793 (BG, LPB); Prov. Nor Yungas, rd. Chuspipata-Ya- losa, km 14, 1150 m, 6 Aug 1988 (5), Beck 13913 (BG, LPB, MO); Prov. Sud Yungas, nr. Sapecho, 26 Feb 1994 (Y fl-fr), Berg 1696 (BG, LPB); Prov. Sud Yungas, Santa Ana de los Mosetenes, 23 Mar 1990 (9 fl-fr), Hinojosa et al. 1104 (LPB); Prov. Sud Yungas, Rio Bopi, San Bartolome, 1-22 Jul 1939 (9 fl-fr), Krukoff 10305 (A, G, K, MICH, MO, NY, S, US), (5), Krukoff 10465 (A, F, G, K, MICH, MO, NY, S, U); Prov. SudYungas, nr. Sapecho, 23 Oct 1993 (5), Seidel 7394 (BG, LPB); Prov. Nor Yungas, 0.3 km N of Yolosa on rd. to Caranavi, 1200 m, 6 Oct 1984 (d), Sol- omon et al. 12471 (AAU, BG); Prov. Nor Yungas, rd. Yo- losa-Chuspipata, km 5.4, 1280 m, 6 May 1990 (5), Solo- mon et al. 18965 (BG, MO). PANDO: Prov. Nicolas Suarez, nr. Cobija, 7 Jan 1983 (9 fl-fr), Ferna'ndez C. et al. 8052 (G, NY); Prov. Nicolas Sudrez, Campo Ana, 14 Jan 1983 (5), Ferndndez C. etal. 8241 (G, NY); Prov. Nicolas Suarez, San Luis, 15 Aug 1990 (st), G. Gonzdlez 12 (BG, LPB); nr. Cobija, Oct 1911 (5), Ule 9311 (MG). SANTA CRUZ: Prov. Andres Ibafiez, San Jose, 13 Nov 1994 (9 fl-fr), Mostacedo et al. 2530 (NY, USZ); Prov. Andres Ibainez, Buena Vista, 18 Dec 1994 (9 fl-fr), Mostacedo et al. 2610 (NY, USZ); Prov. Andres lbafiez, W of Santa Cruz, 16 Jan 1988 (9 fl), Nee et al. 35924 (BG, LPB, NY, USZ); Prov. Andres Ibdfiez, Botanical Garden, E of Santa Cruz, 4 Jan 1989 (d fl), Nee 37513 (LPB, NY), (5), Nee 37514 (BG, LPB, MO, NY, TEX), (9 fr), Nee 37515 (BG, LPB, NY); Prov. Andres Ibafiez, SW of Santa Cruz, Villa Fatima, 4 Nov 1990 (9 fl), Nee 39708 (BG, LPB, NY, USZ); Prov. Ichilo, 0-2 km SW of El Carmen, Parque Nacional Ambor6, 9 Nov 1990 (d), Nee 39842 (LPB, NY, USZ); Prov. Ichilo, Parque Nacional Ambor6, Rio Saguayo, nr. mouth of Quebrada Yapoje, 15 Jun 1991 (9 fl-fr), Nee 41051 (LPB, MO, NY, USZ); Prov. Ichilo, 4 km WSW of Hondo, 14 Feb 1994 (9 fl-fr), Nee et al. 44967 (NY, USZ); Nuflo de Chdvez, Reservas Vida Sil- vestre Rios Blanco y Negro, 14 Apr 1992 (9 fl-fr), Saldias et al. 2409 (USZ); between Angostura & Samaipata, 30 Oct 1980 (9 fl), Zuloaga et al. 1552 (U).
Young trees of this species usually have pinnately lobed segments, especially in the upper part of the lamina; the lobes point upward. In adult (flowering) trees, pinnately lobed leaf segments are less common or the lobes are less pronounced. At the southern limit of the species distribution in Santa Cruz (Bolivia), near the transition of semideciduous subtropical forest to chaco scrub forest, the tree shape is usually con- spicuously different from the normal (lowland Ce- cropia) tree shape in being more robust and usually with distinct umbrella-shaped crowns, short and thick trunks, and relatively short internodes. Moreover, in this area the species is predominantly represented by a green morph (with whitish stipules). The common red morph, with reddish-greyish stipules occurring exclusively elsewhere in the range of the species, is far less common in the region indicated. The append- ages at the bases of the thecae varies from well- developed (to 0.1 mm long) to absent. Cecropia po- lystachya is essentially a lowland species, but in Huainuco and Junin, Peru, it can be found at elevations to 1800 m. Material of this species collected by Ruiz and Pavon comprises specimens with the segments of the lamina distinctly lobed and leaves with (sub)entire segments. Both Trecul (1847) and Klotzsch (1847) described the two morphs as distinct species. Ruiz and Pav6n collected both staminate and pistillate in- florescences of this species. Some sheets contain both types of inflorescences (like those on which Klotzsch described as C. pinnatiloba and C. ruiziana), others only one of them. The names Cecropia leucophaea and C. nivea are probably based on the same collec- tion made by Poeppig.
The species is (or has been) in cultivation in sev- eral places in Brazil: Minas Gerais (Vicosa) and Sao Paulo (Campinas, and Piracicaba), and it may have become naturalized in Mogi-Guacu.
BRAZIL. SAO PAULO: Campinas, Dtto. Barao Geraldo, Cidada Universitaria, 16 Oct 1978 (6), Gabrielli et al. 8758 (UEC), (d + ), Gabrielli et al. 8759 (UEC), Mun. Mogi- Guaqu, Reserva Biol6gica de Mogi-Guaqu, 19 Nov 1992 (6), Godoi et al. 269 (SP), (9), Godoi et al. 270 (SP).
It has also been planted in the Botanical Gardens of Durban (South Africa: 1967, A. P Mills 22, A) and Singapore (1967, Sidek bin Kiah S.60, A, C; 1979, J. E Maxwell 79-59, AAU). The species is planted as an ornamental tree along an avenue in Santa Cruz de la Sierra (Bolivia: 1994, Mostacedo et al. 2391, NY).
Vernacular names and use. Peru: toroc (Cuzco); yungul (Junin). Brazil: imbauba branca (Rondonia). Bolivia: ambaibo blanco (Beni, Santa Cruz).
SYSTEMATIC TREATMENT 155
The fruiting spikes are sold in markets in Santa Cruz de la Sierra.
45. Cecropia purpurascens C. C. Berg, Acta Ama- zonica 7: 185. 1977, Acta Amazonica 8(2): 179. 1978. Type. Brazil. Amazonas: Nr. Manaus, Berg 264 (holotype: INPA; isotypes: MO, NY, P, RB, U, US) = P 18808 (isotypes: COL, K, MG, P, VEN, Z). Fig. 40
Tree, to 15 m tall. Leafy twigs 1-5 cm thick, green, hispidulous to hirtellous, with curved to uncinate hairs. Lamina (sub)coriaceous, ca. 20 X 20 cm to 65 X 65 cm, the segments (5-)6-7, the free parts of the upper segments ovate, the incisions down to 4/10-5/ 10; apices obtuse; upper surface scabrous, hispidu- lous; lower surface minutely puberulous on the veins, on the larger veins also sparsely hirtellous with un- cinate hairs to subtomentose, with arachnoid indu- mentum in the areoles or sometimes also on the main veins; lateral veins in the free part of the midsegment 8-10 pairs, submarginally loop-connected, usually unbranched; petiole 20-50 cm long, puberulous to hirtellous, or sometimes also with arachnoid indu- mentum; trichilia fused, the brown indumentum in- termixed with short white hairs; stipules 8-16 cm long, pale red-brown, hirsute outside, subsericeous in- side. Staminate inflorescences in pairs, with the pe- duncle erect and the spikes pendulous; peduncle 6- 9(-15) cm long, hispidulous to puberulous; spathe 12-15 cm long, grayish red to orange-red, densely hirtellous outside, glabrous inside; spikes 15-20, 7- 13 X 0.3-0.4 cm, with stipes 1.2-1.5 cm long and minutely puberulous. Staminate flowers connate; per- ianth tubular, ca. 1.5 mm long, glabrous, the apex plane; filaments ? swollen; anthers ca. 0.5 mm long, appendiculate, detached at anthesis (?), reattached to the margins of the aperture by the appendages (?). Pistillate inflorescences in pairs, erect, deflexed in fruit; peduncle 5-10 cm long, hispidulous to puber- ulous; spathe 8-12 cm long, the color and indumen- tum as in the staminate inflorescence; spikes 4, 8-10 X 0.4 cm, to 13 X 1 cm in fruit; subsessile or stipes to 0.5 cm long and minutely puberulous; rachis hairy. Pistillate flowers: perianth ca. 2 mm long, with arach- noid indumentum below the apex outside, also in the style channel inside, the apex truncate, muriculate; style long, ? S-shaped, minutely puberulous; stigma comose. Fruit oblongoid, ca. 2.5 mm long, ? tuber- ulate.
Distribution (Fig. 10. 1). The middle Amazon ba- sin, near and north of Manaus, in non-inundated for- est.
Representative specimens examined. BRAZIL. AMA-
ZONAS: Rd. BR.174, Reserva Campina-INPA, 7 Aug 1979 (9 fl-fr), Benson 10345 (BM, K, NY, RB, UEC); San Al- berto, confluence of Rio Negro & Rio Branco, 25 Aug 1924 (9 fl-fr), Bequaert 3 (GH); rd. Manaus-Caracarai, km 65, 21 Sep 1973 (6), Berg 281 (K, NY, U); Distr. Agropecuario, 2?24-25'S, 59043-45'W, 18 Nov 1988 (9 fl-fr), Boom et al. 8533 (BG, MO, NY); Mun. Manaus, Reserva Florestal Ducke, 28 Sep 1962 (d), Duarte 7188 (RB), 21 Jun 1994 (9 fl-fr), Hopkins et al. 1422 (BG, INPA); Distr. Agrope- cuario, 2?24-25'S, 59043-45'W, 15 Nov 1988 (6), Mori et al. 19817 (BG, INPA, NY); Mun. Manaus, ca. 80 km N of Manaus, Distr. Agropecuario da SUFRAMA, 25 Jun 192 (9
fl-fr), Nee 42895 (INPA, K); Rio Negro, Camanau, 11 Jul 1995 (9 fl), Oliveira 2771 (SPF); Manaus, Igarape do Binda, 26 Jun 1961 (9 fl-fr), Rodrigues et al. 2090 (INPA); rd. Manaus-Itacoatiara, km 66, 18 Aug 1970 (9 fl-fr), Ro- drigues 8908 (INPA, MO).
This species is exceptional in the connate peri- anths of the staminate flowers. It shows affinities to Cecropia ficifolia and C. obtusa.
Cecropia trilobata Cuatrecasas, Revista Acad. Colomb. Ci. Exact. 9(36/37): 336. 1956. Type. Colombia. Ca- queta: 97 km NW of Las Guacamayas, ca. 1200 m, 24 Apr 1944 (6), Little 7766 (holotype: F; isotypes: COL, US).
Tree, to 10(-20) m tall. Leafy twigs ca. 1.5-3 cm thick, green, puberulous. Lamina chartaceous, ca. 25 X 25 cm to 60 X 60 cm, with 8-10 radiating primary veins, only in the upper part distinctly incised down to 3/10-5/10, separating into 3 lobes, the lower part faintly lobed to subentire; apices acuminate; upper surface ? scabrous, hispidulous and with very sparse arachnoid indumentum, lower surface on the veins puberulous, with arachnoid indumentum usually con- fined to the margin, sometimes also present on the main veins; lateral veins in the free part of the mid- segment 7-11 pairs, submarginally loop-connected, most of them branched; petiole ca. 25-50 cm long, with dense arachnoid indumentum; trichilia fused, the brown pluricellular hairs intermixed with (rather) sparse short hairs; stipules ca. 5-15(-20) cm long, bright to dark red, puberulous outside, sparsely to
156 FLORA NEOTROPICA
-K>~~~~~~~~~~~~~~~2
6 '3
FIG. 40. Cecropia purpurascens. 1. Leaf and stipules. 2. Apex of lamina and venation (Berg s.n.). 3. Pair of staminate inflorescences with spathes and base of petiole with trichilium (Berg 281). 4. Staminate inflorescence at anthesis and base of petiole with trichilium (Mori et al. 19817). 5. Staminate flower and stamen (Berg 281). 6. Pistillate inflorescence at anthesis and base of petiole with trichilium (Pereira et al. 2303.2165). 7. Pistillate flower. 8. Fruit with persistent style (Berg et al. P 18808).
SYSTEMATIC TREATMENT 157
J~~~~~~~~~~~~~~~~~~~~~
10 cm. i TnS ij j
FIG. 41. Cecropia putumayonis. 1. Leaf, stipules and base of petiole with trichilium. 2. Apex of lamina and venation (Gentry et al. 37903). 3. Stipules, staminate inflorescence with spathe, open prostoma, and bases of petioles with trichilia (Gudino 586). 4. Staminate inflorescences with spathes and bases of petioles with trichilia (Palacios 1892). 5. Staminate inflorescence at anthesis and base of petiole with trichilium (Lugo 4825). 6. Staminate flower. 7. Stamen (Franco et al. 4509). 8. Pistillate inflorescence in fruit and base of petiole with trichilium (Berg et al. 1093). 9. Pistillate flower. 10. Pistil. 11. Fruit (Franco et al 4678)
158 FLORA NEOTROPICA
densely sericeous inside. Staminate inflorescences in pairs, pendulous; peduncle ca. 15-25 cm long, min- utely puberulous and with sparse to dense arachnoid indumentum; spathe ca. 25-35 cm long, yellowish brown, puberulous to hirtellous and usually also with sparse to dense arachnoid indumentum outside, gla- brous inside; spikes 4-5, 20-26 X ca. 0.4 cm, sessile; rachis hairy. Staminate flowers: perianth tubular, 1- 1.2 mm long, with short straight hairs on the margin of the apex and lower down ? dense arachnoid in- dumentum, the apex plane; anthers 1-1.2 mm long, not appendiculate, remaining attached to the filament by a bundle of stretched spiral thickenings of tra- cheary elements at anthesis. Pistillate inflorescences in pairs, pendulous; peduncle 25-45 cm long, min- utely puberulous and with sparse to dense arachnoid indumentum; spathe ca. 30-35 cm, whitish, puberu- lous to hirtellous and usually also with dense (to sparse) arachnoid indumentum outside, almost gla- brous inside; spikes 4, ca. 30 X 0.5 cm, to ca. 50 X
1.2 cm in fruit, sessile; rachis hairy. Pistillateflowers: perianth ca. 2-2.5 mm long, with arachnoid indu- mentum below the apex outside, also in the upper part of the style channel inside, the apex convex, finely muriculate; style long, S-shaped, sparsely minutely puberulous; stigma penicillate to comose. Fruit ob- longoid, ca. 1.8 mm long, smooth.
Distribution (Fig. 18.5). From Amazonian Co- lombia (Caqueta, Meta, and Putumayo) to Peru (Ama- zonas and Huanuco), mostly in forest in non- inundated places, at elevations to 1000(-1350) m.
Representative specimens examined. COLOMBIA. CAQUETA: Mun. Florencia, rd. to Gabinete, 850 m, 18 Oct 1993 (d), Franco et al. 4509 (BG), 1000 m, 18 Oct 1993 (d), Franco et al. 4514 (BG); trail Las Guacamayas-Ramos, km 59, 97 km NW of Las Guacamayas, ca. 1200 m, 24 Apr 1944 (Y fl-fr), Little 7765 (COL, F, US). META: Mun. Vi- llavicencio, rd. Villavicencio-Guayabetal, 900-1100 m, 13 Feb 1995 (Y fl-fr), Franco et al. 4678 (BG, COL, HUA, NY, US); Mun. Cubarral, Vrda. Agua Claras, 19 Nov 1995 (Y fl-fr), Morales et al. 647 & 648 (COL). PUTUMAYO: Rio San Miguel, between Rio Bermeja & Rio Conejo, 13 Dec 1940 (d), Cuatrecasas 11053 (COL); Mun. Mocoa, rd. Mo- coa-Villa Garzon, El Pepino, 18 Feb 1995 (Y fl-fr), Franco et al. 4689 (BG, COL); Vrda. La Campucana, Finca La Mar- iposa, 1350 m, 28 Apr 1994 (st), Franco et al. 5464 (BG); Mocoa, between La Campucana & San Antonio, 1000 m, 2 May 1994 (6 fl-fr), Franco et al. 5513 (BG).
ECUADOR. MORONA-SANTIAGO: Rio Blanco, be-
tween Macas & Sucua, 26 Jan 1981 (Y fr), Berg 1215 (AAU, BG, COL, GB, K, MO, NY, QCA, TUR, U); rd. Gualaquiza- San Juan Bosco, km 15, 1200 m, 16 Feb 1994 (d fr), Berg et al. 1694 (BG, QCNE); Rio Cuyes, between Bomboiza & Gualaquiza, 1 Nov 1982 (d), Palacios 1447 (BG, QAME,
QCNE). NAPO: Tena, 14 Oct 1939 (Y fl-fr), Asplund 9301 (S); rd. Coca-Auca, ca. km 50, 28 Feb 1980 ( fl-fr), Berg et al. 1093 (AAU, BG, COL, NY, QCA, TUR, U); Reserva Biol6gica Jatun Sacha, 8 km E of Misahualli, 22 Oct 1988 (Y fl-fr), Cer6n et al. 5429 (MO, QAME, QCNE); Cant6n Archidona, rd. Hollin-Loreto, Rfo Huataraco, 23-30 Aug 1989 (st), Ceron et al. 7633 (MO, QCNE); San Jose de Pay- amino, 40 km W of Coca, 3 Apr 1984 (st), Irvine 728 (F); Codo Alto, between Santa Rosa & Cascada de San Rafael, ca. 1000 m, 14 Sep 1990 (d), Jaramillo etal. 12711 (QCA); Parque Nacional Yasuni, rd. Pompeya Sur-Iro, km 76, 13 Aug 1994 (Y fl), Jaramillo et al. 16961 (QCA); rd. Coca- Tiputini, 5 Aug 1975 (d), Little et al. 61 (Q, QAME, QCNE); Puyopungu, 15 Sep 1976 (6), Lugo 4825 (BG, GB); Reserva Biol6gica Jatun Sacha, 1-15 Sep 1987 (d), Palacios 1892 (BG, NY, QAME, QCNE); Cant6n El Chaco, Rio Quijos, Codo Sinclair, 16-20 Sep 1990 (d), Palacios 5741 (BG, QCNE); Afiangu, nr. mouth of Rio Afiangu, Parque Nacional Yasuni, 30 Jun-9 Jul 1982 (d), SEF 10245 (AAU, NY, QCA, U). PASTAZA: Between Mera & Moravia, ca. 1000 m, 17 Dec 1955 (9 fl-fr), Asplund 18877 (S); Can- t6n Pastaza, Pozo petrolero Corrientes, 1-31 Aug 1990 (6), Gudino 519 (BG, MO, QCNE); 1 km S of Mera, ca. 1000 m, 6 Mar 1985 (9 fl-fr), Neill et al. 6003 (MO, QAME). SUCUMBiOS: Rd. Lago Agrio-Tarapoa, 1 Aug 1975 (6), Little et al. 6 (Q, QAME, QCNE); Lumbaqui, ca. 1000 m, 13 Aug 1975 (9 fl-fr), Little et al. 169, 178 (Q, QAME, QCNE). ZAMORA-CHINCHIPE: Rd. Zamora-Gualaquiza, between Yantzatza & Cumbaratza, 4 Jan 1991 (d fl-fr), Berg et al. 1660 (BG, LOJA, QCA).
PERU. AMAZONAS: Rfo Cenepa, nr. Huampami, ca. 5 km E of Chdvez Valdivia, 11 Jul 1978 (6), Ancuash 1020 (BG), 25 Jul 1978 (d), Ancuash 1094 (BG, MO); Rio Ce- nepa, Quebrada Huampani, ca. 5 km E of Chavez Valdivia, 11 Jul 1978 (9 fl-fr), Ancuash 1020 (BG); Prov. Bagua, Dtto. Imaza, Rio Marafi6n, Kampaenza, 7 Oct 1994 (d), N. Jaramillo et al. 517 (MO); Prov. Bagua, Yamayakat, 11 Oct 1995 (9 fl-fr), N. Jaramillo et al. 851 (MO); Rio Cenepa, Quebrada Huampani, 15 Feb 1973 (9 fl-fr), Kayap 368 (F, GH, MO, USM); Prov. Bagua, Dtto. Imaza, Rio Marafi6n, Kampaenza, Feb 1995 (9 fl-fr), R. Vdsquez et al. 19532 (MO); Prov. Bagua, Rio Marafi6n, opposite Quebrada Mir- and, 16 Sep 1962 (9 fl-fr), Wurdack 2025 (F, NY, US). LOR-
ETO: Mouth of Rio Santiago, (9 fl-fr), Tessmann 4033 (NY). SAN MARTiN: Prov. San Martin, rd. Tarapoto-Yuri- maguas, ca. km 22, ca. 900 m, 5 Dec 1997 (9 fr), Berg et al. 1774 (BG, COL, MOL), km 20-27 km, 21 Jul 1982 (6), Gentry et al. 37903 (BG, MO, USM).
This species shows clear affinties to Cecropia ut- cubambana. They are similar in their usual habit: slender trees with relatively small and narrow crowns and inflorescences with very long peduncles. The lat- ter species has more deeply incised laminas, but such a difference may occur in the same species (as in C. andina and C. pastasana). However, the staminate flowers are distinctly different. This species occasion- ally shows another habit: large trees with thick stems
SYSTEMATIC TREATMENT 159
and broad crowns (Franco et al. 4678), collected at the limit of its geographical and elevational range.
47. Cecropia reticulata Cuatrecasas, Revista Acad. Colombia Ci. Exact. 6(22/23): 279. 1945; Berg & Franco Rosselli, Fl. Ecuador 48: 47. 1993. Type. Colombia. Valle: Rio Digua, Piedra de Moler, 900-1180 m, 19-28 Aug 1943 (Y), Cuatrecasas 15117 (holotype: COL; isotypes: F, US). Fig. 42
Cecropia bracteata Cuatrecasas, Revista Acad. Colomb. Ci. Exact. 9(36/37): 331. 1956. Type. Colombia. Valle: Quebrada de San Juan, below Queremal, 1350 m, 8 Nov 1946 (Y), Cuatrecasas 22763 (holotype: US; isotypes: F, VALLE).
Cecropia reticulata Cuatrecasas forma alboreticulata Cuatrecasas, Revista Acad. Colomb. Ci. Exact. 9(36/ 37): 329. 1956. Type. Colombia. Valle: Rio Dfgua, between Queremal & La Elsa, 1160-1200 m, 29 Mar 1947 (Y), Cuatrecacas 24008 (holotype: US; iso- type: F).
Cecropia scutata Cuatrecasas, Revista Acad. Colomb. Ci. Exact. 9(36/37): 334. 1956. Type. Colombia. An- tioquia, nr. Dabeiba, 21 Mar 1948 (Y), W Johnson & E A. Barkley 127 (holotype: US; isotypes: COL, F, MEDEL, VALLE).
Tree, to 20 m tall. Leafy twigs 1.5-2 cm thick, grey to blackish, with dense to sparse (filiform) brown pluricellular hairs, glabrescent. Lamina chartaceous, ca. 40 X 40 cm to 75 X 75 cm (to 90 X 90 cm), the segments 7-9(-10), the free parts of upper segments ovate to elliptic, the incisions down to 3/10-5/10(-7/ 10) in the upper part of the lamina, down to ca. 3/10 or less in the lower part of the lamina; apices obtuse to rounded; upper surface scabridulous to almost smooth, minutely hispidulous or partly puberulous, sometimes ? bullate; lower surface puberulous and also with brown pluricellular hairs, with (sometimes very sparse) arachnoid indumentum in the areoles and on the smaller veins, often (initially) also on the main veins, or absent; lateral veins in the free part of the midsegment 10-15 pairs, submarginally loop- connected, unbranched (or rarely branched); petiole ca. 20-65(-80) cm long, sparsely puberulous and with sparse to dense arachnoid indumentum, mostly on an abaxially bulging (subscrotiform) base of the petiole; trichilia fused, large, mostly extended with lateral as- cending lobes, the brown indumentum intermixed with dense long (curved to straight) white hairs; stip- ules ca. 10-20(-25) cm long, red-brown (to whitish),
caducous, densely sericeous and/or with dense arach- noid indumentum outside, very sparsely hairy to gla- brous inside. Staminate inflorescences in pairs, sub- tended by caducous (to subpersistent) bracts, to 7.5 cm long, the peduncle erect and the spikes erect to ? spreading; peduncle 2-7 cm long, tomentose to stri- gose, often also with sparse arachnoid indumentum, or subglabrous; spathe 8-14 cm long, brown to red- brown, with dense brown pluricellular hairs and often also with (sparse to rather dense) arachnoid indumen- tum and/or sparsely sericeous outside, (sub)glabrous inside; spikes 4-12, 6-12 X 0.4-0.8 cm, sessile; ra- chis hairy or glabrous. Staminate flowers sessile or with pedicels to 0.6 mm long; perianth tubular, 1.2- 1.5 mm long, glabrous, the apex convex to plane; fil- aments flat; anthers 0.6-0.8 cm long, appendiculate, detached at anthesis (?), reattached to the margins of the aperture by the appendages (?). Pistillate inflores- cences in pairs, erect, subtended by caducous (to sub- persistent) bracts, to 3 cm long; peduncle 3-5 cm long, densely hirtellous to hispid; spathe 5-9 cm long, the color and indumentum as in the staminate inflo- rescence; spikes 4-5, 4-7 X ca. 0.5 cm, to 1 cm diam. in fruit, sessile; rachis hairy. Pistillate flowers: peri- anth 1.5-2 mm long, with arachnoid indumentum be- low the apex outside, also in the lower part of the style channel inside, the apex convex, finely granulate to muriculate; style short; stigma penicillate to comose. Fruit ellipsoid, ca. 2 mm long, ? tuberculate.
Distribution (Fig. 18.5). Western Ecuador and Colombia and adjacent Panama, in forest and second- ary growth, at elevations to ca. 1800 m, mostly above 1000 m.
Representative specimens examined. PANAMA. DARIEN: Cerro Pirre, 1200 m, 16 Nov 1989 (Y fr), Fisher 57 (BG); N of Cerro Pirre, between Cerro Pirre & Rancho Pastico, 1200-1400, 14 Nov 1977 (st), Folsom et al. 6351 (BG); Summit of Cerro Pirre, 1000-1400 m, 29 Dec 1972 (d), Gentry et al. 7022 (BG, MO).
COLOMBIA. ANTIOQUIA: Mun. Anorn, Vrda. El Car- men, km 12-16 NW of Anorn, 940-1100 m, 18 Nov 1989 (6), Callejas et al. 8757 (BG, HUA, NY); Yarumal, 1725 m, 27 Mar 1994 (6), Franco et al. 4595 (BG HUA); Mun. Frontino, Cgto. Nutibara, rd. Nutibara-La Blanquita, Murri region, 1650-1700 m, 4 Mar 1995 ( fl-fr), Franco et al. 5551 (BG, COL, HUA, LP, US), (6), Franco et al. 5552 (BG, COL, HUA, LP, US), 1250 m, 4 Mar 1995 (6), Franco et al. 5557 (BG, COL, HUA, LP); Mun. Frontino, Cgto. Nutibara, source of Rio Cuevas, 1630 m, 15 Jan 1984 (Y fl- fr), D. Sdnchez S. et al. 990 (BG, MEDEL). CAUCA: W of Rio San Joaquin, 1400-1500 m, 29-30 Jun 1922 (Y fl), Kil- lip 7873 (GH, NY, US). CHoCO: Mun. San Jose del Palmar, Rio Torito, Finca Los Guadales, 1 Mar 1980 (Y fl), Forero et al. 6480 (COL, US); Mun. San Jose del Palmar, Vrda. El
160 FLORA NEOTROPICA
~~~~~ 1~~~~~~I0
FIG. 42. Cecropia reticulata. 1. Leaf, reduced. 2. Apex of lamina and venation. 3. Stipules and young leaf. 4. Base of petiole with trichilium. 5. Pair of staminate inflorescences with subtending bracts (Berg 1244). 6. Staminate inflorescence with spathe (Dodson et al. 12747). 7. Staminate inflorescence at anthesis (Berg 1244). 8. Staminate flower and stamen (Franco et al. 4696). 9. Pair of pistillate inflorescences with spathes (0llgaard et al. 57707). 10. Pistillate inflorescence at anthesis and base of petiole with trichilium (Dodson 6668). 11. Staminate flower. 12. Fruit (Franco et al. 5551).
SYSTEMATIC TREATMENT 161
Sinai, 1800 m, 19 Mar 1994 (Y fl-fr), Franco et al. 4560 (BG, HUA). NARINO: Mun. Barbacoas, El Diviso, 7 Oct 1993 (st), Franco et al. 4462 (BG); Mun. Ricaurte, rd. Ricaurte-Altaquer, 1300 m, 8 Oct 1994 (6), Franco et al. 4474 (BG); nr. Altaquer, 1150 m, 21 Feb 1995 (6), Franco et al. 4696 (BG, COL, HUA, LP, US); Mun. Ricaurte, Res- guardo Indigena Nulpe Medio, Quebrada La Conga, 8 Jan 1996 ( fl-fr), M. S. Gonzdlez et al. 1619 (COL); Mun. Nar- bacoas, Resguardo El Sabalo, Rio Cangapi, 21 Aug 1995 (6), M. S. Gonzalez et al. 8200 (COL). RISARALDA: Mun. Mistrat6, rd. San Antonio de Chami-Mistrat6, Quebrada Sutu, 27 Apr 1992 (9 fl-fr), Alonso et al. 10193 (COL). VALLE: Rio San Juan, below Queremal, 1300-1700 m, 19 Mar 1947 (6), Cuatrecasas 23883 (US, VALLE); Mun. Cal- ima, Vrda. Campo Alegre, 1000 m, 4 Oct 1983 (6), Devia 396 (TULV).
ECUADOR. AZUAY: Nr. Molleturo, 25-27 Jan 1991 (6), L. Ortiz 203 (QCA, QCNE). CARCHI: Cerro Golon- drinas, Upper Rio Pablo drainage, 1730-1760 m, 24 Apr 1993 (st), Boyle et al. 1730 (BG); San Marcos de los Coai- queres, trail Chical-Tobar Donoso, 10 Feb 1985 (9 fl-fr), 0llgaard et al. 57707 (AAU, QCA); Chical, 1700-1900 m, 20-29 Jul 1991 (9 fl-fr), Quelal et al. 273 (BG, QCNE); Cant6n Tulcan, Reserva Indigena Awa, San Marcos, 25 km NW of Chical, 1500 m, 16-30 Nov 1990 (6), Rubio et al. 978 (BG, QCNE); Tobar Donoso, 19-28 Jun 1992 (6), Ti- paz et al. 1480 (BG, QCNE). COTOPAXI: Rd. Quevedo- Latacunga, nr. El Palmar, 17 Feb 1981 (6), Berg 1280 (AAU, BG, COL, GB, MO, QCA, U); rd. Quevedo-Lata- cunga, km 52-53, Tenefuerte, Rio Pilal6, 750-1300 m, 21 Feb 1982 (d), Dodson et al. 12724 (MO, QCNE); rd. Pilal6- Quevedo, between Macuchi & La Mana, 29 Jul 1980 (6), Holm-Nielsen et al. 24705 (AAU); Rio Guapara, 20 km NW of El Coraz6n, 24 Jun 1967 (6), Sparre 17358 (S). EL ORO:
Between La Vega & Rivera, Rio Chilola, 19 Nov 1994 (9 fl), Cornejo et al. 3578 (BG); rd. Pifias-Santa Rosa, km 12.5, 7 Oct 1979 (6), Dodson et al. 8959 (MO, QCNE), (9 fr), Dodson et al. 8960 (MO, QCNE). ESMERALDAS: Mun. Quininde, Bilsa Biological Station, Mache Mtns., 35 km W of Quininde, Fila de Bilsa, 7 km E of San Jose de Bilsa, ca. 80 km SW of Esmeraldas, 4 Oct 1994 (9 fl), J. L. Clark et al. 140 (BG, QCNE), 30 Jan 1991 (6), Gentry et al. 72939 (QCNE). IMBABURA: Nr. Lita, 7 Feb 1981 (6), Berg 1244 (AAU, BG, COL, GB, K, MO, QCA, U, WIS). Los Rios: Rio Palenque Biological Station, rd. Quevedo-Santo Do- mingo de los Colorados, km 56, 2 Oct 1976 (9 fl-fr), Dod- son 6451 (F, MO, QCA, US), 5-14 Mar 1977 (9 fl-fr), Dod- son 6668 (AAU, MO, QCA). Los RiOS/PICHINCHA: El Centinela, at crest of Montanias de Ila, rd. Patricia Pilar-24 de Mayo, km 12, 27 Nov 1978 (6), Dodson 7304 (BG, MO, QCNE, U). MANABf: Machalilla National Park, San Sebas- tian, 20 Jan 1991 (6), Gentry et al. 72460 (MO, QCNE). PICHINCHA: Nr. Santo Domingo de los Colorados, Hda. Gloria Maria, 18 May 1955 (9 fl-fr), Asplund 16402 (S); rd. Tandapi-Santo Domingo de los Colorados, km 20, 13 Sep 1977 (6), Berg et al. 422 (AAU, BG, COL, MO, QCA, U); Rio Guajalito, ca. 4 km N of Palmeras, km 59 on rd. Quito-San Juan-Chiriboga-Empalme, 1800 m, 15 Dec 1990 (6), Berg et al. 1647 (BG, QCA); rd. Nono-Rio Yacuambi,
5-10 km above Nanegalito, 1700 m, 21 Jul 1980 (d), Holm- Nielsen et al. 24410 (AAU); Rio Guajalito, ca. 4 km N of Palmeras, km 59 on rd. Quito-San Juan-Chiriboga-Em- palme, 1800 m, 29 Feb 1992 (d), Jaramillo et al. 14657
(NY, QCA).
The arachnoid indumentum on various parts varies from dense to almost lacking, the latter state found in particular in the northern part of the distribution range (Antioquia). Moreover, in the northern part of the spe- cies range, the incisions are often less deep (down to ca. 3/10) and the characteristic shape of the trichilia on a subscrotiform base of the petiole wanting. This northern type has been described as Cecropia scutata
and might represent a distinct infraspecific entity, oc- curring in Antioquia at elevations higher than the more typical one with the lamina more deeply incised (down to ca. 5/10), subscrotiform bases of the peti- oles, and trichilia with ascending lateral lobes.
From some distance the leaves (and therefore the trees) often look whitish in spite of the absence of arachnoid indumentum on the upper surface of the lanmina. The bracts subtending the peduncle are con- spicuously present in fresh material, but as they are mostly caducous, usually absent in herbarium mate- rial.
Vernacular name. Colombia: cosedera (Narifio).
48. Cecropia sararensis Cuatrecasas, Revista Acad. Colomb. Ci. Exact. 6(22/23): 292. 1945. Type. Co- lombia. Norte de Santander: Sarare region, Rfo Cubugon, El Indio, 13 Nov 1941 (9), Cuatrecasas 13098 (holotype: COL, isotype: F).
Cecropia cobariana Cuatrecasas, Revista Acad. Col- omb. Ci. Exact. 6(22/23): 278. 1945. Type. Colom- bia. Norte de Santander: Sarare Region, confluence of Rio Cubug6n and Rio Cobaria, El Banco, 15 Nov 1941 (9), Cuatrecasas 13197 (holotype: COL; iso- type: F).
Cecropia libradensis Cuatrecasas, Revista Acad. Col- omb. Ci. Exact. 9(36/37): 337. 1956; Velasquez, Acta Bot. Venez. 6: 43, t. 7. 1971. Type. Colombia. Norte de Santander: Sarare Region, Quebrada de la China, Santa Librada, El Reposo, 800 m, 20 Nov 1941 (d), Cuatrecasas 13355(holotype: F; isotype: COL).
Tree, to 15 m. Leafy twigs 1.5-4 cm thick, green to brownish with conspicuous lenticels, sparsely to rather densely hispidulous with curved hairs to gla- brous. Lamina chartaceous to subcoriaceous, ca. 25 X 25 cm to 70 X 70 cm, the segments 8-10(-13), the free parts elliptic to oblong, the incisions down to 6/ 10-8/10; apices subacute to subacuminate or to ob- tuse; upper surface smooth or ? scabrous, subgla-
162 FLORA NEOTROPICA
brous to hispidulous; lower surface sparsely to rather densely (minutely) puberulous and with longer unci- nate hairs to subglabrous on the veins, with arachnoid indumentum in the areoles and on the smaller veins; lateral veins in the free part of the midsegment 11- 18 or 18-24 pairs, marginally loop-connected, un- branched or (often only the lower ones) branched; pet- iole 15-45 cm long, sparsely (to densely) puberulous to subglabrous, sometimes with sparse arachnoid in- dumentum, the base of the petiole sometimes ? bulg- ing (subscrotiform); trichilia fused, the brown indu- mentum intermixed with short white to brownish hairs; stipules 7-18 cm long, greenish to yellowish brown or red(dish), with only brown pluricellular trichomes or also sparsely strigillose or sparsely to densely whitish to brownish hirtellous to subhirsute to sericeous, sometimes also with sparse (to ? dense) arachnoid indumentum outside, (densely) yellowish sericeous to villous inside. Staminate inflorescences in pairs, (sometimes?) subtended by caducous bracts, to 3 cm long, the peduncle erect to deflexed and the spikes spreading to pendulous; peduncle 4-17 cm long, (sub)glabrous or sparsely puberulous, hirtellous or subhirsute, sometimes in the lower part with sparse arachnoid indumentum; spathe 9-19 cm long, green- ish, yellowish (brown), red(dish) or whitish, with only brown pluricellular trichomes or also sparsely to rather densely whitish to brownish hirtellous to sub- villous, sometimes sparse to ? dense arachnoid in- dumentum outside, densely whitish to yellowish to brownish sericeous to villous (or subglabrous) inside; spikes 4-18, 6-17 X 0.3-0.5 cm, with stipes 0.2-1 (-2) cm long and sparsely hirtellous or minutely pu- berulous; rachis glabrous. Staminateflowers: perianth tubular, 1-1.5 mm long, glabrous or sparsely puber- ulous below the apex, the apex plane; filaments flat; anthers ca. 0.6 mm long, appendiculate, detached and reattached to the margins of the aperture by the ap- pendages. Pistillate inflorescences in pairs or solitary, erect to (at least in fruit) pendulous, (sometimes?) subtended by caducous bracts, to 2 cm long; peduncle 7-19 cm long, subglabrous or sparsely (to densely) puberulous to hirtellous to subhirsute, sometimes in the lower part with sparse arachnoid indumentum; spathe 8-15 cm long, the color and indumentum as in the staminate inflorescence; spikes (3 or) 4, 6-13 X ca. 0.5 cm, to 33 X 1 cm in fruit, (sub)sessile or with stipes to 0.8 cm long and glabrous; rachis (sub)glabrous. Pistillate flowers free or connate at the base; perianth ca. 1.5 mm long, with arachnoid in- dumentum below the apex or to near the aperture out- side, absent (or present in the style channel) inside, the apex plane, muriculate or smooth; stigma truncate
and penicillate to peltate. Fruit oblongoid to ellipsoid, ca. 2 mm long, smooth, blackish.
Distribution (Fig. 18.6). Andean region of Vene- zuela and in Colombia in northern part of the eastern slopes of the eastern Cordillera, southward to Meta, in forest and secondary growth, at 300-1400 m, in Venezuela to 2300 m.
Specimens examined. COLOMBIA: ARAUCA: Mun. Fortul, Vrda. Palmarito, 26 Jan 1995 (d), Franco et al. 4632 (BG, COL, HUA, US), (Y fl-fr), Franco et al. 4633 (BG, COL, HUA, US); Mun. Tame, rd. to Sacama, 26 Jan 1995 (Y fl-fr), Franco et al. 4634 (BG, COL, HUA, US). BOYACA: Mun. Pajerito, between Corinto & Penia del Gallo, 1200 m, 29 Jan 1995 (Y fl-fr), Franco et al. 4642 (COL, HUA, MO, US). CASANARE: Rd. to Sacama, 29 Jan 1995 (6), Franco et al. 4635 (COL), (Y fl-fr), Franco et al. 4636 (COL); Sacama, 29 Jan 1995 (d), Franco et al. 4637 (BG, COL). CUNDINAMARCA: Rd. Guayabetal-Villavicencio, 1035-1150 m, 29 May 1993 (d), Franco et al. 4459 (BG, COL). META: Above Rio Negro, ca. 20 km W of Villavi- cencio, 1100 m, 23 Feb 1972 (Y fl-fr), A. S. Barclay et al. 3197 (COL, US); nr. Villavicencio, 10 Feb 1995 (Y fl), Franco et al. 4655 (BG, COL), (6), Franco et al. 4656 (BG, COL), (d), Franco et al. 4663 (BG, COL); rd. Villavicen- cio-Guayabetal, 900-1 00 m, 13 Feb 1995 ( Y fl-fr), Franco et al. 4672 (COL), (6), Franco et al. 4673 (BG, COL), (d), Franco et al. 4674 (BG, COL), (Y fl-fr), Franco et al. 4675 (BG, COL); Llanos de San Martin, Rio Humadea, Guamal, 10 Sep 1958 (Y fl-fr), R. Jaramillo-Mejia et al. 1017 (COL). NORTE DE SANTANDER: Mun. Toledo, Cgto. San Bernardo, 1250 m, 23 Jan 1995 (d), Franco et al. 4623 (BG, COL); Mun. Toledo, rd. La Mesa-Samore, 1150 m, 23 Jan 1995 (d), Franco et al. 4626 (BG, COL), (Y fl-fr), Franco et al. 4627 (BG, COL), (d), Franco et al. 4628 (BG, COL); Mun. Toledo, Samore, 25 Jan 1995 (Y fl-fr), Franco et al. 4629 (BG, COL); nr. Chinacota, 1400 m, 18 Mar 1927 (6), Killip et al. 20852 (A, GH, NY, US).
VENEZUELA. BARINAS: Mun. Pedraza, Alto del Aguada, 1250-1300 m, 22 Feb 1955 (Y fr), Bernardi 2031 (NY); rd. Barinitas-El Cacao, 7 Jul 1981 (Y fl-fr), Marcano Berti et al. 277-981 (U, US); rd. Barinas-San Crist6bal, El Corozo, Dec 1966 (d), Veldsquez 101 (VEN). FALC6N:
Cerro Galicia, Dtto. Petit, 15 Dec 1977 (juv), T Ruiz et al. 2806 (VEN). LARA: Dtto. Jim6nez, Parque Nacional Ya- cambu, Quebrada Negra, 24 Oct 1982 (d), Davidse 20921 (MO, VEN). MERIDA: Dtto. Tovar, rd. Tovar-Zea, 900- 1000 m, 1 Jul 1954 (Y fl-fr), Bernardi 1313 (NY); between Estanquez & La Coloradas, 12 May 1982 (Y fl-fr), Marcano Berti et al. 982-013 (G, U, US, VEN); between Merida & Santo Domingo, 500-2500 m, Feb 1968 (d), Velasquez 261 (US, VEN), (Y fl-fr), Velasquez 262 (US, VEN). PORTU-
GUESA: Concepci6n, 10 Jan 1967 (d), R. E Smith V526 (VEN). TACHIRA: 10 km E of La Fundaci6n, nr. Represa Dorada, 600-1000 m, 1-13 Mar 1981 (9 fl), Liesner et al. 10437 (MO, VEN); Cerro Las Minas, 18-20 km SE of Santa Ana, 1150-1250 m (9), Steyermnark et al. 119050 (MO, U). TRUJILLO: Dtto. Bocon6, La Morita, above Rio Saguras, S
SYSTEMATIC TREATMENT 163
of Campo Elias, 2300 m, 4 Jun 1988 (Y fl), Dorr et al. 5367 (BG, NY).
The stigma varies from penicillate to peltate, and linked to this difference, the arachnoid indumentum varies from present only below the apex of the peri- anth to present near the aperture of the perianth. The indumentum of the stipules and spathes varies from brown to white subhirsute to subvillous to subgla- brous (only with brown pluricellular hairs present) to (sometimes) with ? dense arachnoid indumentum. A mixture of the various types of indumentum is most common in the middle part of the range of this spe- cies. The number and the length of the spikes of the staminate inflorescence varies from numerous and rel- atively short to few and relatively long. In the northern part of the range (Venezuela), the material with more numerous lateral veins (18-24 pairs) in the free part of the midsegment, with spathes being (sub)glabrous outside, with staminate inflorescences with few and long spikes, and with comose(-penicillate) stigmas appears to be predominant. In this part of the range, Cecropia sararensis reaches to elevations of 2300 m. The type with these features includes the type of C. libradensis. Cecropia sararensis, as it is delimited in the present treatment, is rather heterogeneous and may prove to comprise two entities, parallel to the two subspecies of C. schreberiana, showing differ- ences rather similar to those between the two types circumscribed above. However, with the material and data available at present, it is not possible to find ac- ceptable delimitations of these types. Moreover, C. sararensis as a whole can barely be distinguished from C. peltata and C. metensis, the neighboring taxa of the C. peltata group. Cecropia sararensis can be distinguished from C. metensis by the absence of dense white arachnoid indumentum on the stipules, spathes, and (also) the petioles, and from the former by subglabrous stipules, spathes and leafy twigs or by the relatively long brownish hairs on the stipules (and spathes).
49. Cecropia saxatilis Snethlage, Notizbl. Bot. Gart. Berlin-Dahlem 8: 360. 1923; Berg & Carauta, Al- bertoa 1(1): 10. 1986. Type. Brazil. Syntypes. Bra- zil, Piuai, Serra Branca, Jan 1907 (d), Ule 7174 (B, destroyed, K, only an inflorescence), (?) Ule 7174 (B, destroyed), specimen with pistillate in- florescence(s) at K here designated as lectotype (K; isolectotype: G).
Tree, to 10 m tall. Leafy twigs 2-4 cm thick, hir- tellous with curved to uncinate hairs or also with
sparse to rather dense arachnoid indumentum. Lamina subcoriaceous to coriaceous, ca. 15 X 15 cm to 50 X 50 cm, the segments 6-10, the free parts of the upper ones (broadly) obovate, the incisions down to ca. 5/ 10-7/10; apices rounded; upper surface scabrous to almost smooth, sparsely to densely hispidulous or subvelutinous, sometimes with (rather dense) arach- noid indumentum; lower surface on the (main) veins subtomentose to hirtellous, with arachnoid indumen- tum almost confined to the areoles or extended to the main veins; lateral veins in the free part of the mid- segment 8-10 pairs, submarginally to marginally loop-connected, branched; petiole 10-35 cm long, reddish, puberulous and with sparse to dense arach- noid indumentum; trichilia fused, the brown indu- mentum intermixed with short and rather long (? bristle-like) white hairs; stipules 8-15 cm long, reddish, with ? dense arachnoid indumentum and hirtellous outside, sparsely hairy inside. Staminate in- florescences in pairs, erect or the peduncle erect and the spikes pendulous (?); peduncle 3-9 cm long, sub- hispidulous to hirtellous and with arachnoid indu- mentum; spathe 8-18 cm long, red to purplish, sparsely puberulous to densely hirtellous and with sparse to dense arachnoid indumentum outside, gla- brous inside; spikes 4-8, 7-17 X 0.3-0.8 cm, with stipes 0.4-1.5 cm long and puberulous and/or with arachnoid indumentum; rachis (sub)glabrous. Stami- nate flowers: perianth tubular, ca. 1.5 mm long, with dense arachnoid indumentum below the apex, the apex muriculate to smooth, the aperture surrounded by a low rim; filaments flat; anthers 0.8-1.2 mm long, appendiculate, detached at anthesis but remaining at- tached to the filament by stretched spiral thickenings. Pistillate inflorescences in pairs, erect to pendulous (?); peduncle 4-23 cm long, subhispidulous to hirtel- lous and with arachnoid indumentum or glabrous; spathe 6-8 cm, the color and indumentum as in the staminate inflorescence; spikes 4, 6-15 X 0.5-0.8 cm, to 28 X 1.5 cm in fruit, (sub)sessile; rachis glabrous. Pistillate flowers: perianth ca. 1.5 mm long, with arachnoid indumentum below the apex outside, also in the style channel inside, the apex slightly convex, muriculate to smooth; style short; stigma penicillate. Fruit oblongoid to subovoid, ca. 2.5 mm long, tuber- culate, dark brown.
Distribution (Fig. 8.6). Central Brazil and eastern Bolivia (Santa Cruz), in cerrado areas, usually on sandstone or limestone rocks, in Bolivia in grassy sa- vanna, at elevations to 1400 m.
Representative specimens examined. BRAZIL. BA- HIA: Espigao Mestre, 34 km W of Barreiras, 2 Mar 1972
164 FLORA NEOTROPICA
(9 fl-fr), Anderson et al. 36458 (BG, NY, US); Mun. Gentio do Ouro, nr. Santo Inacio, rd. to Xique-Xique, Serra do Acu- rua, 27 Nov 1992 (d), Arbo et al. 5346 (BG, K, SPF); nr. Barreiras, 3 Jun 1991 (9 fl-fr), Brito et al. 333 (GUA, NY), (d), Brito et al. 336 (NY); Mun. Oliveira dos Brejinhos, Serra de Agua Quente, 16 Apr 1999 (9 fl-fr), Forza et al. 1242 (NY); valley of Rio das Ondas, Espigao Mestre, ca. 5 km NW of Barreiras, 4 Mar 1971 ( 9 fl-fr), Irwin etal. 31509 (BG, NY, UEC, US). DISTRITO FEDERAL: Rd. Campo Grande-Cuiaba, BR.163, 30 Jan 1979 (d + 9 fl), Heringer et al. 997 (NY, UEC, US). GoiAs: Chapada dos Veadeiros, 6-7 km E of Alto Paraiso, ca. 1400 m, 7 Mar 1973 (9 fr), Anderson et al. 6552 (MO, NY, U, US); Chapada dos Vead- eiros, S of Terezina, ca. 1000 m, 19 Mar 1973 (9 fr), An- derson 7495 (MO, NY, U, US); Vila Boa de Goias, 22 Jan 1969 (9 fr), Carauta 723 (GUA, U); Mun. Alto Paraiso de Goias, rd. Terra-Nova Roma, 6 km E of Alto Paraiso de Goias, 1200 m, 13 Mar 1995 (9 fl-fr), Cavalcanti etal. 1294 (SPF); Campos Anayas, Mar 1840 (9 fl-fr), Gardner 3981 (BM, K) Serra Dourada, 21 Jan 1966 (5), Heringer 1096 (US); Serra Dourada, 20 km SE of Goias Velho, 16 Jan 1966 (5), Irwin et al. 11799 (G, NY, RB, S, SP, US); Serra Geral do Parana, ca. 20 km S of Sao Joao de Alianqa, ca. 1000 m, 17 Mar 1971 (9 fl-fr), Irwin et al. 32099 (BG, MOL, NY, QCA, US); Mun. Alto Paraiso, rd. to Cavalcanti, Chapada dos Veadeiros, 1200-1300 m, 13 Feb 1990 (9 fl), Hatsch- bach et al. 53946 (SP); Serra Dourada, 21 Jan 1966 (d), Heringer 10961 (NY), 22 Jan 1978 (5 + 9 fl-fr), Rizzini (RB) 205206 (GUA, RB). MATO GROSSO: Chapada dos Guimaraes, nr. Portao do Inferno, 27 Jan 1989 (9 fl-fr), Carauta 5776 (GUA, NY), (d), Carauta 5777 (GUA, NY); Mun. Rondon6polis, rd. BR.163, km 15, 12 Nov 1975 (d), Hatschbach 37457 (U); Campo Grande-Cuiaba, 30 Jan 1979 (9 fl-fr), Heringer et al. 997 (GUA); nr. Diamantina, 23 Apr 1983 (9 fl-fr), Moraes et al. 279 (MG); jct. of rds. to Cuiaba, Santarem & Porto Velho, 4 Feb 1979 (9 fl), M. G. Silva et al. 4441 (MG, NY, U). MINAS GERAIS: 15 km W of Janu-
aria, 20 Apr 1973 (9 fl-fr), Anderson 9258 (MO, NY, U, US); Mun. Januaria, Dtto. de Fabiao, 24 Jan 1997 (5), Lom- bardi et al. 1757 (BHCB), 16 Feb 1998 (9 fl), Lombardi et al. 2214 (BG, BHCB). PUAi: Sao Raimundo Nonato, Serra da Capivara, 13 Dec 1978 (st), Freire s.n. (GUA).
BOLIVIA. SANTA CRUZ: Prov. Velasco, rd. Santa Rosa de la Roca-Concepci6n, km 45, 9 Dec 1994 (9 fl-fr), Guil-
ln et al. 2686 (USZ); Prov. Velasco, Estancia Flor de Oro, W side of Rio Guapor6 (= Rio Itenez), 22 Jun 1991 (d), Nee 41225 (LPB, MO, NY), (9 fl-fr), Nee 41226 (K, LPB, MO, NY, USZ); Prov. Velasco, Parque Nacional Noel Kempff M., Pampa Toledo, 1 Jul 1993 (9 fl-fr), Saldias et al. 2891 (USZ).
The leaves of Cecropia saxatilis resemble those of
C. obtusa, but the lateral veins in the free parts of the
leaf segments are branched, whereas they are mostly
unbranched in C. obtusa. The pistillate inflorescences
of the two species are also quite similar. However, the
staminate flowers are very different. Those of C. ob-
tusa have anthers of the most common type, but those
of C. saxatilis have anthers of the less usual type with long anthers remaining after abscission attached to the filament by spiral thickenings, as found in C. gla- ziovii, C. palmata, and C. sciadophylla. Anderson 9258 differs from all other collections by the rela- tively narrow and subobovate free parts of the lamina segments in which the lateral veins are not much branched.
50. Cecropia schreberiana Miquel, in Martius, Fl. Bras. 4(1): 150. 1853. Type. Probably from one of the French islands of the Lesser Antilles, certainly not from "Guyane" as indicated on the label, (d),
Schreber s.n. (holotype: U; isotype: M, photo- graph in F). Fig. 43
Tree, to 15 m tall. Leafy twigs 1.5-3 cm thick, green, glabrous, sparsely puberulous (sometimes only on the stipular scars) or densely hirtellous to subhir- sute to subvillous or to hispidulous; internodes partly filled with brown pith. Lamina subcoriaceous to cor- iaceous, sometimes ? plicate, ca. 25 X 25 cm to 50 X 50 cm (to 70 X 70 cm), the segments 8-10, the free parts of the upper ones ovate to elliptic to oblong, the incisions down to 4/10-7/10; apices rounded to subacute; upper surface smooth and (sub)glabrous or ? scabrous and hispidulous to (densely) puberulous to hirtellous, often also with sparse to dense arachnoid indumentum; lower surface glabrous, with rather dense brown pluricellular trichomes or hirtellous to subhirsute on the veins, with arachnoid indumentum in the areoles and on smaller veins, at least initially often extending to the main veins; lateral veins in the free part of the midsegment 10-16 pairs, marginally loop-connected, usually branched; petiole 10-35 (-45) cm long, glabrous or hirsute to hirtellous or to subvillous, often only at the base, often also with sparse to dense arachnoid indumentum; trichilia ab- sent, occasionally present, but then usually poorly de- veloped; stipules 7-20 cm long, green, glabrous or sparsely to densely (sub)hirsute to (sub)strigose, often also with arachnoid indumentum outside, sparsely to rather densely (sub)sericeous (or subglabrous) inside. Staminate inflorescences solitary or in pairs, the pe- duncle erect and the spikes erect to pendulous (?); peduncle 4-10(-13) cm long, glabrous or hirtellous to subhirsute, often also with arachnoid indumentum; spathe 4-8(-10) cm long, green or grayish green, gla- brous or hirtellous to subhirsute to strigose, often also with arachnoid indumentum outside, glabrous inside;
SYSTEMATIC TREATMENT 165
L~~~~~~~
FIG. 43. Cecropia schreberiana subsp. schreberiana. 1. Leafy twig with staminate inflorescences with spathes. 2.
Staminate inflorescence and part of spike (Wagner 566). 3. Staminate flower and stamen (Berg 1526). 4. Pistillate inflores-
cence at anthesis (Beard 180). 5. Pistillate flower and style. 6. Fruit (Berg 1533). C. schreberiana subsp. antillarum. 7.
Pistillate flower (Maas et al. 6420). (By T. Schipper and Hendrieke Berg.)
166 FLORA NEOTROPICA
spikes ca. 10-20, 1.5-6(-9.5) X 0.2-0.3 cm, with sti- pes 0.3-1.5 cm long and glabrous or hairy; rachis hairy. Staminate flowers: perianth tubular, ca. 1-1.2 mm long, glabrous, the apex convex to almost plane; filaments flat; anthers ca. 0.5 mm long, appendiculate, detached and reattached to the margins of the aperture by the appendages at anthesis. Pistillate inflores- cences often solitary, erect; peduncle 2-8(-1 1) cm long, glabrous or hirtellous to subhirsute, often also with arachnoid indumentum; spathe 3-8 cm long, the color and indumentum as in the staminate inflores- cence; spikes 4-6, 1.5-6 X ca. 0.5 cm, to 13 X 1.3 cm in fruit, sessile or stipes to 0.4 cm long, glabrous or hairy. Pistillate flowers: perianth ca. 1.5-2 mm long, with arachnoid indumentum below the apex or extending to near the aperture outside, absent inside, the apex slightly convex to plane, muriculate to min- utely puberulous or smooth; style short; stigma ligu- late to subpeltate to peltate. Fruit ellipsoid to oblon- goid, ca. 1.5-2.5 mm long, tuberculate, dark brown.
In the presence of ample brown pith in the inter- nodes, the absent or poorly developed trichilia, and the smooth leafy twigs and upper surface of the lam- ina (in subsp. schreberiana), this species shows sim- ilarities to several Andean species. The presence of peltate stigmas and arachnoid indumentum on the apex of pistillate flowers in subsp. antillarum indicate affinities to the C. peltata-group, in particular to C. sararensis, which show parallel variation in the in- dumentum and features of the staminate flowers. Sub- juvenile specimens of C. schreberiana can hardly or cannot be distinguished from material of C. peltata without trichilia (from Jamaica). Even if trichilia are sometimes more or less developed, then they do not contain Mullerian bodies (Rickson, 1977), or only ab- normal (?) ones (Wheeler, 1942: 73). In many studies on the genus in the Caribbean, e.g., those by Janzen (1973) and Rickson (1977), this species was not re- garded as distinct from C. peltata, in spite of the fact that Snethlage (1923) established several names for this species and that Wheeler (1942) noted that the Cecropia species in Puerto Rico did not belong to C. peltata. Two subspecies can be recognized. The tran- sition of characters is gradual and intermediates occur in Hispaniola and Puerto Rico. Juvenile and subju- venile specimens of the subspecies are quite similar. Characters of the subjuvenile state can be retained until flowering starts.
Key to the subspecies
1. Lamina with the upper surface smooth, glabrous, or only with arachnoid indumentum; lower surface with only arachnoid indumentum
on the main veins, this often initially covering the main veins .................. a. subsp. schreberiana
1. Lamina with the upper surface ? scabrous, hispidulous to hirtellous; lower surface hirtellous to subhirsute or often also with arachnoid indumentum on the main veins ............................................ b. subsp. antillarum
Leafy twigs (usually) glabrous or only with brown arachnoid trichomes or white hairs at the stipular scars. Lamina with the upper surface (usually) smooth, glabrous or only with arachnoid indumen- tum, lower surface only with arachnoid indumentum, initially often extending to the main veins. Perianth of the pistillate flower without arachnoid indumentum at the apex; stigma ligulate to subpeltate.
Distribution (Fig. 18.3). Lesser Antilles, Puerto Rico, and Hispaniola, in forest and secondary growth, at elevations to 1100 m.
Representative specimens examined. HAITI. Tortue Is., nr. La Vallee, 28 Dec 1928-9 Jan 1929 (?), Leonard et al. 11496 (A, GH, US); between Plaisance & Limbe, 28 Aug 1903 (5), Nash 889 (NY, U).
PUERTO RICO. Caribbean National Forest, Luquillo Mtns., 27 Sep 1989 (5), Acevedo R. 2956 (NY, US); Maun- abo, Cuchilla de las Panduras, 24 Feb 1991 (? fl-fr), Axelrod et al. 2062 (UPRRP); Jayuya, Toro Negro Forest Reserve, 17 Jan 1992 (? fl-fr), Axelrod et al. 3772 (UPRRP); Carri- bean National Park, El Junque, 31 Mar 1985 (5), Berg 1526 (AAU, BG, U); between Arecibo & Rio Abajo Forest, 2 Apr 1985 (st), Berg 1527 (BG); Rio Abajo Forest, 2 Apr 1985 (Y fl-fr), Berg 1528 (BG); Marici Forest, 3 Apr 1985 (Y fl- fr), Berg 1533 (BG, U); Caribbean National Forest, rd. to summit of El Yunque, 4 Jan 1987 (Y fl-fr), Boom 6894 (NY, UPRRP); Sierra de Luquillo, between Mameyes & Juncos, Sep-Oct 1966 (5), Byer 66-960 (UPRRP); Mun. Cayey, Bosque Nacional Carite, 5 Apr 1992 (5), Escobar et al. 9935 (UPRRP); Luquillo National Forest, El Yunque, 1 Mar 1985 (5), Gentry et al. 50369 (BG, NY); Mun. Patillas, Reserva Forestal Carite, 19 Nov 1981 (5), Hansen et al. 9158 (US); Rio Piedras, Finca Bueno Consejo, May 1914 (5), Bro. Hioram s.n. (NY, US); Cerro Maravilla area, 30 Jan 1979 ( fl), Liogier et al. 28274 (NY, US); Quebradillas, 26 Feb 1986 (st), Liogier et al. 35988 (NY); Caribbean Na- tional Forest, 25 Jun 1950 (5), Little 13103 (NY, US), (9 fr), Little 13014 (US); Las Piedras, 22 May 1989 (9 fl), Loiz et al. 37 (UPRRP); Mun. Cayey, rd. to Barrio Farrall6n, 9 Feb 1991 (5), G. S. Miller et al. 6000 (UPRRP); Mun. Rio Grande, rd. to El Verde, 9 Dec 1988 (st), Ortega 128
SYSTEMATIC TREATMENT 167
(UPRRP); Mun. Caguas, Sierra de Cacey, Carite Forest Re- serve, 2 Feb 1986 (6), Proctor 41467 (IJ); Maricoa Insular Forest, 25 Jun 1965 (9 fl), Stimson 1337 (MICH); Mun. Orocovis, rd. 564, km 7.3, 18 Aug 1966 (9 fl-fr), Stimson 4057 (MICH); Mun. Aibonito, rd. 162, between rd. 1 & rd. 716, 8 Apr 1989 (9 fl-fr), Taylor et al. 8823 (UPRRP); Sa- bana rd., nr. Palmer, 2 Feb 1968 (9 fl-fr), Wagner 1321 (A, U) Barranquitas, 30 Jul 1979 (9 fl-fr), Woodbury et al. s.n. (NY, US); El Yunque, 23 Aug 1979 (9 fl-fr + juv), Wood- bury s.n. (NY, US).
MONTSERRAT. Olveston area, 19 Mar-16 Apr 1979 (9 fl-fr), Howard et al. 18982 (A, BM, NY, US); Gorge of Gage's, Soufriere stream, 12 Feb 1959 (6), Proctor 19153 (A, BM, IJ, US); Fergus Mtns., 30 Jan 1907 (6), Shafer 342 (NY, U, US).
SABA. The Mountain, 24 Jul 1906 (st), Boldingh 2158 (U); top of The Mountain, 11 Aug 1953 (st), Stoffers 4230 (A, K, NY, U).
ST. EUSTATIUS. The Quill, 7 Jun 1906 (st), Boldingh 254 (U); Bottom of the Quill, 17 Jun 1906 (st), Boldingh 904 (U), 21 Jun 1906 (st), Boldingh 955 (U).
ST. KITT'S. Molyneaux Estate, 8 Sep-5 Oct 1901 (st), Britton et al. 572 (NY).
GUADELOUPE. Bains-Jaunes, 1892-1893 (? fl-fr), Duss 2860 (NY); Bains-Jaunes, Mar 1905 (st), Duss 4207 (NY); Marie Galante, Les Balisiers distr., 1 Jun 1960 (d), Proctor 21135 (A).
DOMINICA. Morne Colla Anglais, 10-23 Aug 1938 (st), Hodge 632 (GH, NY, U, US); nr. Calibishie, 23 Apr 1940 (9 fl-fr), Hodge et al. 3168 (GH); La Chaudiere, 10- 14 Mar 1940 (? fl-fr), Hodge et al. 3663 (BM, GH, US); St. Joseph Parish, Layou Park Estate, 22 Nov 1964 (9 fl), Nicholson 2037 (BM, GH, U, US); Laudat, 24 Apr 1988 (9 fl-fr), Ramage s.n. (BM, K); without locality, Mar 1942 (d), D. Taylor 125 (GH); rd. to Trou Cochon, 9 Oct 1983 (6), Whitefoord 3958 (BM, US); Baiac, 18 Sep 1984 (d), White- foord 4135 (A, BM, NY, US).
MARTINIQUE. Mome-Rouge, Fonds St. Denis, 1883, Duss 1405 (NY, US); Bois de Caleluze (?), Dec 1867 (st), Hahn 210 (BM, G, K); La Chapelle, Apr 1873 (? fl-fr), Hahn s.n. (G); Pitons Boucher, Lacroix et Dumauze, ca. 1000 m, 13 Apr 1968 (6), Oldeman et al. M.58 (P); Morne Mitan, 4 Apr 1980 (st), Sastre 6912 (U); Piton Dumauze, 11 Apr 1980 (9 fl-fr), Sastre 6922 (U); La Soufriere, between Baines Jaunes & Savane a Mulets, 1000-1100 m, 17 May 1980 (st), Sastre 6939 (U).
ST. VINCENT. Lodge R., Jan 1890 (d), Eggers 6829 (A, P, US); Chateaulair R., 16-25 Apr 1947 (st), Morton 5419 (US); Cumberland R., 2-3 May 1947 (st), Morton 5571 (US); Parish of St. Andrew, Upper Buccament R. valley, 7 Mar 1965 (6), Proctor 26152 (IJ).
ST. LUCIA. Quiless, 14 Dec 1943 (6), Beard 180 (A, K, NY, U, US); SE of Piton Troumass6e, 11-12 May 1958 (? fl-fr), Proctor 17949 (A, BM, IJ, US); Piton Flore, (d + 9 fl-fr), Sturrock 714 (A).
Brokaw (1998) presented an account of this sub-
species, particularly in relation to its role as a colo- nizer, e.g., after hurricanes.
Vernacular names. Dominica and St. Lucia: bois canot. Montserrat: trumpeter tree. St. Vincent: trum- pet. French Antilles: bois trompette.
50b. Cecropia schreberiana Miquel subsp. antil- larum (Snethlage) C. C. Berg & P. Franco, comb. et stat. nov. Fig. 43.7
Cecropia sericea Snethlage, Notizbl. Bot. Gart. Berlin- Dahlem 8: 368. 1923. Type. Haiti. Nr. Terreneufe, Buch 99 (holotype: B, destroyed).
Leafy twigs hirtellous to hirsute to subvillous or to hispidulous. Lamina with the upper surface + scabrous, hispidulous to hirtellous; lower surface hir- tellous to subhirsute on the main veins and initially often also with arachnoid indumentum. Perianth of the pistillate flower with arachnoid hairs on the apex to near the aperture; stigma subpeltate to peltate (or ligulate).
Distribution (Fig. 18.3). Cuba, Hispaniola, Puerto Rico, and the Virgin Islands, in forest and secondary growth, at low elevations.
Representative specimens examined. CUBA. CAMA-
GUEY: Nr. La Gloria, 8 Feb 1909 (st), Shafer 325 (NY, US). LAS VILLAS: Trinidad Mtns., Tope de Collantes, 17 Jul 1957 (9 fl-fr), (Bro. Alain) Liogier 6432 (US), (i), (Bro. Alain) Liogier 6433 (US). MATANZAS: Valley of the St. Augustine R., 2 Sep 1903 (9 fr), Britton et al. 258 (NY); San Miquel de los Banios, Apr 1951 (9 fl), Dahlgren 51/102 (US); be- tween Mantanzas & Varadero, 21 Jun 1988 (9 fl), Fernandez C. et al. 10690 (NY). ORIENTE: Bayate, nr. Rio Jagua, 5 Nov 1915 (st), Ekman 6548 (NY, S); Los Pifiales, SE of Paso Estancia, 1-2 May 1909 (9 fl-fr), Shafer 1726 (A, NY, US); Monteverde, Jul 1859 (9 fl-fr), Wright 1440 (G, GH, K, P, S). PINAR DEL Rio: Nueva Gerona, May 1904 (6), Curtiss s.n. (NY); Vinales, 11 Dec 1930 (9 fr), Killip 13571 (US); nr. Cabafias, 20 May 1900 (st), Palmer et al. 768 (US); nr. Pinar del Rio, 19 May 1941 (9), Rutten-Pekelharing 436 (U). SANTA CLARA: Cienfuegos Distr., Cieneguita, 23 Mar 1895 (9 fl-fr), Combs 111 (GH, NY, P); Cienfuegos, Sole- dad, 30 Mar 1926 (9 fl), Jack 4485 (S); Cienfuegos, Sole- dad, Limones, 1 1 Aug 1927 (6), Jack 5271 (P, US); Cien- fuegos, Soledad, 8 Oct 1927 (9 fl-fr), Jack 5558 (S); Cienfuegos, Soledad, Belmonte, 17 Jul 1929 (9 fl-fr), Jack 7481 (A, NY, P, US).
HAITI. Massif de la Selle, Papette, 28 Nov 1927 (9 fl- fr), Ekman 9376 (NY, S, US); Soliette R., Fonds Verrettes, 21 Mar 1942 (6), Holdridge 1061 (MICH, NY, US); nr. Petionville, 15-29 Jun 1920 (9 fl-fr), Leonard 4875 (NY, US); nr. St. Michel de I'Atalaye, 3 Jan 1926 (9 fl-fr), Leon- ard 8532 (NY, US); Dept. du Nord, nr. Dondon, 7 Jan 1926 (6), Leonard 8655 (US); Dept. de l'Artibonite, 18 Feb 1926
168 FLORA NEOTROPICA
(st), Leonard 9990 (US), 23 Feb 1926 (d), Leonard 10026b (GH, US); Tortue Is., nr. La Vallee, 28 Dec-9 Jan 1929 (9 fr), Leonard 11496 (A, GH); nr. Basse Terre, 23 Mar 1929 (d6), Leonard et al. 13998 (US), 21-29 Mar 1929 (9 fl-fr), Leonard et al. 14084 (A, GH, NY, US); Dept. Sud, Bouzy, Dabo, 20 Jun 1980 (st, juv), Sastre et al. 6950 (P).
DOMINICAN REPUBLIC. BARAHONA: 3-4 km N of Paraiso, on rd. to Las Auyamas, nr. Nizaito, 11 Apr 1985 (? fl), Gentry et al. 50717 (BG, MO, NY); El Arroyo, nr. Barahona, 25 Mar 1985 (d), Maas et al. 6414 (BG, U); La Filipina, 6.3 km from Ocean Hwy., 25 Mar 1985 (9 fl), Maas et al. 6420 (BG, U). DISTIUTo NACIONAL: Santo Do- mingo, 30 Apr 1929 (9 fl-fr), Ekman 12324 (GH, S, US); Santo Domingo, Jardin Botdnico Nacional, 2 Jun 1980 (9 fl-fr), Mejfa et al. 6732 (NY), (6), Mejia et al. 6733 (NY); Santo Domingo, 24-25 Mar 1923 (6), Rose et al. 4138 (NY, US), 8 Jul 1981 (9 fl-fr), Zanoni et al. 15282 (NY). MONTE
CRISTI: Sabaneta Distr., La Ceiba, 17 Oct 1930 (? fl-fr), Valeur 480 (C, G, MICH, US). MONTE PLATA: Rd. Ya- masa-Maim6n, Rio Verde, 28 Mar 1987 (6), Zanoni et al. 38796 (NY). SAN CRISTOBAL: Rd. Cambita Garabita (de San Crist6bal)-Arroyo Marfa, 4 Oct 1987 (6), Zanoni et al. 40487 (NY).
PUERTO RICO. Yabucoa, Punta Guayanes, old rd. from S end of Palmas del Mar, 12 May 1991 (9 fl-fr), Ax- elrod et al. 2427 (NY, UPRRP), 18 Aug 1991 (9 fl-fr), Ax- elrod et al. 2787 (NY, UPRRP); Vieques, Monte Pirata, 23 Jul 1992 (9 fl), Axelrod et al. 4921 (NY, UPRRP); Mun. Isabela, Bo. Arenales Altos, 8 Nov 1993 (9), Nee 44158 (NY); Bayam6n 31 Mar 1885 (d), Sintenis 250b (G, GH, K, NY, P, S, US); nr. San. German, 9 Mar 1886 (9 fl-fr), Sintenis 3971 (BM, G, GH, K, NY, P); nr. Penluelas, Talba- boa, 8 Aug 1886 (6), Sintenis 4805 (BM, G, GH, K, NY, P, S, US); Vieques Is., between Isabel Segunda & Martineau, 31 Jan 1914 (d), Shafer 2643 (NY, US); S of Cayay, 15 Mar 1932 (6), J. S. Miller 1604 (US).
ST. CROIX. Caledonia Gut, 8 Jul 1873 (9 fl-fr), Eggers s.n. (C); Jolly Hill, 29 Jan 1906, Raunkiaer s.n. (C); Lebanon Hill, 24 Jun 1896 (9 fl-fr), Ricksecker 449 (NY); without locality, (9), West s.n. (C).
ST. JOHN. Cruz Bay, Center Line rd., 1 Jul 1989 (9 fl), Acevedo 2845 (NY, UPRRP, US).
TORTOLA. Rd. to High Bush, 13-17 Feb 1913 (6), Britton et al. 715 (NY, US).
Due to the (sub)peltate stigmas, the white arach- noid indumentum also largely covering the apex of the perianth pistillate flower, and some features of the indumentum of the vegetative parts, material referred to this subspecies looks quite similar to Cecropia pel- tata without trichilia. This applies even more so to material from the Virgin Islands and Vieques Island, as the lamina has a less coriaceous texture than else- where. Cecropia schreberiana subsp. antillarum has been introduced in Madagascar and known under the vernacular name "Taigisy" or "Taingisa." According to Capuron 23568 (P) the (sub)species is planted in (Est) Faragangana. Material collected by Capuron ( 15
Oct 1964) has staminate inflorescences. Another col- lection in P, collected by Rakotoarivelo Andrianjar- afeno (under number 459, 20 Oct 1961) is made near Vangaindrana has pistillate inflorescences. The label data of the latter collection suggest that this (sub)species is becoming naturalized in Madagascar.
51. Cecropia sciadophylla Martius, Flora 24, Beibl. 2: 93. 1841; Velasquez, Acta Bot. Venez. 6: 60, t. 14; Berg, Acta Amazonica 8(2): 181. 1978. Am- baiba sciadophylla (Martius) Kuntze, Revis. Gen. P1. 2: 624. 1891. Type. Colombia. Amazonas: Rfo Caqueta', Puerto Mirania, Martius 630(1) (holo- type: M, fragment in U).
Cecropia inchuensis Cuatrecasas, Revista Acad. Col- omb. Ci. Exact. 9(36/37): 329. 1956. Type. Peru: Ucayali: Prov. Coronel Portillo, Inchua, 2 Aug 1946, Soukup 3018 (F, mixed collection consisting of a pis- tillate inflorescence of uncertain identity [possibly C. ficifolia, see Soukup 3017] and separate leaf parts, the latter element(s) designated as lectotype, Berg & Franco Rosselli, Fl. Ecuador 48: 48. 1993, F).
Tree, to 30 m tall. Leafy twigs 1.5-5 cm thick, green to brownish, glabrous or puberulous. Lamina coriaceous, ca. 20 X 20 cm to 100 x 100 cm, the segments (7-)10-15, lanceolate, the incisions down to the petiole and the segments usually with petiolules 1-7 cm long; apices acuminate to acute; upper surface
SYSTEMATIC TREATMENT 169
smooth and glabrous; lower surface with sparse brown pluricellular hairs on the (main) veins, with short and sparse arachnoid indumentum in the are- oles; lateral veins in the midsegment (25-)30-40(-45) pairs, submarginally loop-connected, unbranched; petiole ca. 20-90 cm long, glabrous; trichilia absent, sometimes a few long white hairs instead of trichilia; stipules 13-50 cm long, red-brown (or greenish), gla- brous or sparsely puberulous outside, (densely) to- mentose to subsericeous inside. Staminate inflores- cences in pairs, erect; peduncle 7-15 cm long, minutely puberulous; spathe 8-18 cm, red-brown (or greenish to yellowish), puberulous outside, glabrous or with sparse arachnoid indumentum inside; spikes ca. 8-15, 7-12 X 0.3-0.8 cm, with stipes to 4 cm long and subglabrous; rachis hairy. Staminate flow- ers: perianth tubular, ca. 1.5-2.5 mm long, with dense arachnoid indumentum below the apex, the apex slightly convex to plane muriculate; filaments ? swollen, sometimes (?) connate at the base; anthers ca. 1-1.5 mm long, appendiculate, detached, remain- ing attached to the filament by a bundle of stretched spiral thickenings of tracheary elements at anthesis. Pistillate inflorescences in pairs or solitary, erect to pendulous; peduncle 3-10 cm long, minutely puber- ulous; spathe 8-12 cm long, the color and indumen- tum as in the staminate inflorescence; spikes 3-6 (-10), (2-)5-10 X 0.8-1 cm, to 26 X 1.5(-2.5) cm in fruit, (sub)sessile or with stipes to 0.5 cm long and glabrous; rachis hairy with stiff short hairs and/or arachnoid indumentum. Pistillate flowers: perianth ca. 2-3 mm long, with arachnoid indumentum below the apex outside, also in the lower part of the style channel (or absent?) inside, the apex convex to almost plane, muriculate; style rather long, straight, with short hairs; stigma penicillate. Fruit oblongoid to su- bobovoid, 2-3 mm long, tuberculate.
Distribution (Fig. 1 1.1). Amazon basin, the Gui- ana region (French Guiana to eastern Venezuela), and the llanos region in Colombia. In primary upland and gallery forest and in secondary growth, in non- inundated places, at elevations to ca. 1300 m.
Representative specimens examined. COLOMBIA. AMAZONAS: Rfo Igara-Parand, La Chorrera, 9 May 1975 (st), Idrobo 8086 (COL); Santa Izabel, Reserva Indfgena Miratia, 5 Nov 1984 (Y fr), La Rotta et al. 543 (COL); Mun. Leticia, Parque Nacional Natural Amacayacu, nr. Quebrada de Agua Pudre, 21-24 Mar 1992 (st), Rudas et al. 3558 (COL, MO); Rio Igara-Parana, nr. La Chorrera, 4-10 Jun 1942 (st), Schultes 3964 (F, G); Trapecio Amaz6nico, Rio Loretoyacu, Oct 1946 (Y fl-fr), Schultes 8446 (F). AMA-
ZONAS/VAUPES: Rio Apaporis, between Rio Pacoa & Rio Kananari, Soratama, 1-15 Dec 1951 (Y fl-fr), Garcia-
Barriga 14012 (COL, US), 20 Aug 1951 (9 fr), Schultes et al. 13679 (COL, U, US), 15 Sep 1951 (6), Schultes et al. 13990 (COL, GH, U, US). CAQUETA: Mun. Florencia, rd. to Gabinete, 17 Oct 1993 (9 fl-fr), Franco et al. 4507 (BG, COL); nr. Las Guacamayas, 27 Apr 1944 (9 fl-fr), Little 7749 (US), (st), Little 7750B (US); Solano, Rfo Caqueta, 8 km SE of Tres Esquinas, below mouth of Rfo Ortequeza, 13 Mar 1945 (6), Little et al. 9785 (COL, GH, NY, US); Ar- aracuara, 25 Jan 1993 (9 fl-fr), Vester et al. 725 (BG). CAS- ANARE: Hato Corozal, Vrda. Las Tapias, 30 Oct 1981 (st), Mahecha et al. 3572 (UDBC). META: Mun. Puerto L6pez, rd. to Puerto Gaitan, 10 Feb 1995 (d fl-fr), Franco et al. 4659 (BG, COL, HUA, MO, US); nr. La Macarena, 11 Feb 1995 (9 fl-fr), Franco et al. 4666 (BG, COL, HUA, MO, US); Cord. La Macarena, between Rio Guejar & Canlo Gua- payita, Canlo Yerli, 20-28 Dec 1950 ( 9 fr), Idrobo et al. 792 (COL, GH, U, US); Mun. Puerto L6pez, SE of Cabuyaro, nr. Laguna de Yurimena, 16 Sep 1958 (6), Jaramillo M. 1237 (COL); Serrania de Menegua, 17 Sep 1958 (6), Jar- amillo M. et al. 1261 (COL, NY); Mun. San Juan Arama, Rio Siejas, 1971 (9 fr), Mahecha s.n. (UDBC 8093); Mun. Vista Hermosa, cacerfo Pifialito, Nov 1973 (st), Mosquera s.n. (UDBC 10855); Sierra de La Macarena, Serrania Cha- musa, Parque Nacional Natural Tinigua, Jan 1991 (6), Ste- venson 226 (COL). PUTUMAYO: Mocoa, Jul 1989 (st), Ma- hecha et al. 5779 (UDBC). VAUPES: Miti, 10 Sep 1956 (9 fr), A. S. Barclay et al. 623 (COL, GH, NY); nr. Miraflores, 8 Feb 1944 (9 fr), Gutierrez V et al. 769 (COL, GH, MEDEL).
VENEZUELA. AMAZONAS: Alto Orinoco, Raudal de los Guaharibos, Salto Salas, 18 Aug 1951 (6), Croizat 501 (F, NY, US); Rio Orinoco, 5 Jan 1952 (st), Cruxent 158 (NY); Dtto. Atabapo, Rio Ventuari, Salto Yureba, 24 Oct-4 Nov 1981 (9 fl-fr), Delascio et al. 11050 (VEN); Rio Ca- siquiare, RIo Padamo, Jan-Feb 1969 (st), Farinias et al. 689 (MO, NY, VEN); 14 km NE of San Carlos de Rio Negro, 24 Jan 1980 (9 fr), Liesner 8664 (MO, NY, VEN); Tayari, 7 May 1975 (9 fl-fr), Lissot 75/35 (VEN); San Carlos de Rio Negro, 17-18 Apr 1970 (6), Steyermark et al. 102718 (NY, US, VEN). BOLiVAR: Uairen region, Aprada Tepui, 1200 m, 19 Aug 1953 (d), Bernardi 826 (NY); rd. Pica Caicara del Orinoco-San Juan de Manapiare, Rio Suapure, Mar 1975 (9 fl-fr), Delascio et al. 2782 (VEN); Dtto. Ced- eno, Rio Tabaro, Research Station Dedmai, 11 Aug 1992 (st), Knab et al. 196 (MICH); Rio Caura, rd. Campamento Las Pavas-El Play6n, 11 May 1982 (6'), Morillo et al. 9154 (MO, VEN); Rio Canaracuni, 13-26 Apr 1988 (9 fl-fr), Stergios 11784 (BG, NY); Quebrada O-paru-ma, below Santa Teresita de Kavanayen, 915-1065 m, 25 Nov 1944 (9 fr), Steyermark 60540 (F, NY, VEN); Chimanta Massif, Chimanta-tepui, above Rfo Tirica, 1000-1400 m, 15 May 1953 (9 fr), Steyermark 75387 (F, NY, VEN); nr. Venezuela/ Brazil border, NE of Serrania Piua-soi, 5-6 Jan 1962 (6), Steyermark 90614 (US); rd. El Dorado-Brazilian border, km 88, Sep 1967 (9 fr), Velasquez 258 (US, VEN); nr. El Pal- mar, 25 Nov 1967 (9 fl-fr), Wessels Boer 2102 (U). DELTA AMACURO: Rio Grande, ENE of El Palmar, nr. Bolivar bor- der, 19 Feb 1964 (9 fl-fr), Marcano Berti 89 (GH, K, NY, P, U, VEN); Sierra Imataca, Rimacuro, upstream from San
170 FLORA NEOTROPICA
Victor, past Rio Matapaima, up to Salto Quebradero, 2 Nov 1960 (5), Steyermark 87270 (BG, NY).
GUYANA. Northwest Distr., nr. Port Kaituma, Aug 1967 (st), Davis 247 (NY); Essequibo R., Rockstone, 15 Jul- 1 Aug 1921 (st), Gleason 663 (GH, K, NY, US); Mazaruni Station, 19 Jun 1942 (Y fl), Fanshawe 731 FD 3467 (K); Northwest Distr., Wauna Mission, 11 May 1961 (5), Harris Y29 (K); Demerara R., Kamini R., Pokorero Cr., Santa, 21 Apr 1923 (st), Hohenkerk 115A (K); Essequibo R., Gunn's, 8 Aug 1989 (? fl-fr), Jansen-Jacobs et al. 1542 (BG, NY, U); Pakaraima Mtns., N of Paruima Mission, 19 Oct 1981 (9 fl-fr), Maas et al. 5859 (K, NY, U); Boerasirie area, 5- 25 mi W of Georgetown, 25 Oct 1955 (9 fr), Little 16954 (US); Soesdyke Hwy., 7 Nov 1982 (9 fl), Persaud 243 (U); Marudi Mtns., nr. Aishalton, 12 Nov 1982 (9 fl-fr), Stoffers et al. 317 (INPA, NY, U).
SURINAME. Sectie 0 (tree 531), 20 Oct 1915 (9 fl), BW s.n. (U, US); S of Onverwacht, W of Paramaribo-Zan- derij rd., 3 Jul 1977 (st), Heyde 710 (U); Wilhelmina Mtns., Lucie R., below confluence with Oost R., 10 Sep 1963 (5), Irwin et al. 55577 (COL, K, MG, MO, NY, S, U, US); Cop- pename R., nr. Raleigh Falls, 13 Sep 1933 (st), Lanjouw 824 (RB, U); Mt. Nassau, 7 Mar 1949 (st), Lanjouw et al. 2485 (U); Lely Mtns., 29 Sep 1975 (st), Lindeman & Stoffers et al. 549 (K, NY, U); Mapane region, Jul 1970 (d), Roberts (LBB) 12791 (U); between Lucie R. & Wilhelmina Mtns., 17 Jul 1964 (5), Schulz (LPB) 10109 (COL, U); Zanderij, Jan 1943 (3 fl-fr), Stahel (Woodherb. Suriname) 179 (A, K, MO, NY, S, U).
FRENCH GUIANA. Rd. St. Laurent-Cayenne, km 11, 1 Aug 1953 (st), BAFOG 39M (U); Crique Margot region, 9 Oct 1956 (st), BAFOG 7561 (NY, P, U); Charvein, 23 Dec 1913 (st), Benoist 442 (P); Mont Chauve, 24 Apr 1997 (5), Cremers et al. 15193 (BG); Taluwen, 28 Jan 1997 (st), Fleury 1065 (BG); Lower Oyapock R., Armontabo Cr., 24 Feb 1981 (st), Granville 4397 (P, U); Bassin de l'Oyapock, Roche Touatou, 20 May 1995 (5), Granville et al. 12995 (BG); Piste de St. Elie, Station ECEREX, 2 Nov 1990 (9 fl-fr), Hoff 6866 (BG); Crique Plomb, 12 Jul 1992 (st), Loubry 1766 (CAY); Sail, 8 Nov 1990 (9 fl-fr), Mori et al. 21555 (NY); piste de St. Elie, 21 Sep 1979 (9 fl-fr), Prevost 801 (P, U), 7 Nov 1984 (9 fl-fr), Riera 905 (BG); basin of Approuage R., Noutagues Station, 11 Nov 1993 (9 fl), Riera et al. 1943 (US).
ECUADOR. MORONA-SANTIAGO: Puerto Morona, Rfo Morona, 30 Sep-2 Oct 1975 (9 fl-fr), Little et al. 572 (LOJA, Q); Cordillera de Cutucu, 5-10 km E of Logrofno, 1200-1500 m, 7-9 Oct 1975 (5), Little et al. 654 (LOJA, QAME, QCNE); 56 km SE of Taisha, 23-25 Sep 1976 (st), Ortega U. 205 (Q); Pozo petrolero Garza, ca. 35 km NE of Montalvo, 2-12 Jul 1989 (9 fl), Zak et al. 4756 (MO, QCNE). NAPO: Tena, 13 Oct 1939 (9 fl-fr), Asplund 9296 (S), 18 Oct 1939 (5), Asplund 9419 (S); Parque Nacional Yasunf, Maxus rd., km 40, 19 Sep 1994 (st), Aulestia et al. 2826 (QCNE); Parque Nacional Yasunf, Pozo petrolero Amo 2, 14 Jan 1988 (9 fl), Ceron 3332 (AAU, BG, MO, QAME); Parque Nacional Yasunf, Pozo petrolero Daimi 2, 14 Jan 1988 (9 fl), Cer6n et al. 3860 (BG, QCNE); rd. Auca-Coca, km 22, 30 Apr 1982 (st), Chaguaro 36 (QCA, QCNE); con-
fluence of Rio Quiwado & Rio Tiwaeno, 11 Apr 1981 (9 fr), Davis et al. 921 (QCA, U); San Jose de Payamino, 20 Apr 1982 (st), Irvine 103 (F, QCA, QCNE); La Joya de los Sachas, Maxus rd., km 1-5, 1-28 Sep 1992 (9 fl-fr), Gri- jalva et al. 68 (NY, QCNE); Campamento Cosapo, 2 Aug 1975 (5l), Little et al. 19 (Q, QAME, QCNE); Rio Napo, 15 km W of Coca, 18-20 Apr 1985 (st), Neill et al. 6335 (BG, QAME, QCNE); Reserva Biol6gica Jatun Sacha, 8 km E of Misahualli, 1-15 Sep 1987 (9 fr), Palacios 1890 (BG, GB, K, MO, NY, QAME, QCNE); Aniangu, Parque Nacional Ya- suni, 30 May-21 Jun 1982 (5), SEF 8644 (AAU, MO, NY, QCA), (9), SEF 9227 (AAU, MO, NY, QCA). PASTAZA:
Cant6n Pastaza, Pozo petrolero Danta 2, 50 km SSE of Cur- aray, 1-20 Oct 1990 (9 fl-fr), Espinoza et al. 379 (BG, GB, NY, QCNE); Cant6n Pastaza, Pozo Petrolero Namoyacu, 30 km S of Curaray, 13-30 Nov 1990 (9 fr), Espinoza et al. 585 (BG, MO); Cant6n Arajuno, 25 km NW of Pozo Villano 2, 3-14 Sep 1998 (9 fl), E. Freire et al. 3260 (BG). Suc- UMBiOS: Rio Cuyabeno, ca. 0O10'S, 75?55'W, 19 Feb 1980 (st), Berg et al. 1067 (BG, MO, QCA, U); Cant6n Gonzalo Pizarro, 30 km NW of Lago Agrio, 1050 m, 31 Mar 1990 (st), Cer6n et al. 9417 (BG); Sacha Lodge, 3 km NW of Aniangu, 5-13 Jun 1995 (5), J. L. Clark et al. 1004 (QCNE); Rio Aguarico, between Dureno & Poblaci6n Cofanes, 10 Jul 1980 (9 fl), Jaramillo et al. 3001 (AAU, QCA); Reserva Faunistica Cuyabeno, nr. Laguna Grande, 1 Apr 1990 (st), Valencia et al. 67779 (QCA). ZAMORA-CHINCHIPE: 27 km NE of Zamora, 1200-1500 m, 17 Sep 1975 (5), Little et al. 430 (COL, LOJA, Q, QAME, QCNE).
PERU. AMAZONAS: Rio Cenepa, nr. Huampami, ca. 5 km E of Chavez Valdivia, 11 Jul 1978 (5), Berlin 2041 (MO); Rio Santiago valley, 65 km N of Pinglo, nr. Caterpiza, 20 Nov 1979 (9 fl-fr), Huashikat 1358 (MO); Rio Cenepa, Quebrada Yutui entsa, 22 Jan 1973 (9 fr), Kayap 213 (GH, MO, USM); Prov. Bagua, Dtto. Imazam Rio Marafi6n, Ya- mayakat, Feb 1995 (5), R. Vdsquez et al. 19527 (MO), (st), R. Vasquez et al. 19807 (MO). Cuzco: Prov. Paucartambo, rd. Pilcopata-Patria, ca. km 5, 1 Aug 1988 (9 fl-fr), Berg et al. 1621 (BG, Centro de Medicina Andina, Cuzco); Prov. Paucartambo, between Atalaya & Pilcopata, 16 Nov 1964 (9 fl-fr), Vargas C. 15760 (US). HUANUCO: Tingo Maria, 23 Aug 1940 (9 fr), Asplund 13311 (S); Prov. Pachitea, ca. 26 km S of Puerto Inca, 3 Nov 1988 (9 fl), Johann 1/3- 31188 (BG); Prov. Leoncio Prado, Dtto. Jose Crespo, Cas- tillo, Carretera Marginal de la Selva, km 11, Aucayacu, 29 Mar 1968 (st), Rodriguez del A. 23 (MOL); Prov. Pachitea, ca. 26 km S of Puerto Inca, 7 Oct 1988 (st), Wallnoefer 19- 71088 (BG); Prov. Puero Inca, Dtto. Yuyapichis, DANTAS, 16-31 Oct 1990 (d), Tello 506 (G, NY), 16-31 Apr 1991 (9 fl), Tello 1841 (G, NY). JUNiN: Puerto Bermddez, 14- 17 Jul 1929 (5), Killip et al. 26499 (NY, US). LORETO: Rio Corrientes, at Ecuador border, 4 Apr 1977 (9 fr), Gen- try et al. 18998 (F, MO); Prov. Requena, Jenaro Herrera, 23 Feb 1987 (st), Gentry et al. 56424 (MO); Prov. Alto Ama- zonas, Puranchim, Rio Sinchiyacu, 30 Mar-I Apr 1987 (d), Lewis et al. 13413 (MO); Prov. Maynas, Rio Ampiyacu, Puca Urquillo, 5 Apr 1977 (9 fr), Plowman et al. 6664 (F, GH, K, US); nr. Indiana, Rio Napo, 1924 (5), Tessmann 3718 (G, NY, US); mouth of Rio Santiago, 1924 (5), Tess-
SYSTEMATIC TREATMENT 171
mann 4717 (G, F, NY); Prov. Alto Amazonas, Capahuari Norte, 7 Jun 1981 (5), R. Vasquez et al. 1995 (BG, MO); Prov. Maynas, Mishana, Rio Nanay, 7 Oct 1982 (Y fr), R. Vasquez et al. 3276 (BG, F, MO); Prov. Maynas, Puerto Almendras, 2 Jan 1985 (Y fl-fr), R. Vdsquez et al. 6074 (BG, MO); Rio Amazonas, Caballo-Cocha, 6 Aug 1929 (st), Ll. Williams 2130 (F, NY, US). MADRE DE DIos: Parque Na- cional Manu, Cocha Cashu Station, 7 Nov 1986 (st), Foster et al. 12189 (BG, LPB, MOL); Prov. Tambopata, jct. of Rio La Torre & Rio Tambopata, 26 May 1987 (Y fl-fr), Gentry et al. 57660 (BG, MO, NY); Prov. Manu, Parque Nacional Manu, Cocha Cashu, 20 Jul 1986 (Y fl-fr), Nuinez 5461 (BG, MO); Prov. Tambopata, Reserva Tambopata, 14 Aug 1990 (6), Reynel et al. 5131 (BG, MO). PASCO: Pichis valley, Puerto Bermudez, Dec 1980 (5), Foster et al. 8035 (U); Prov. Oxapampa, Iscozacin, 15 Jun 1982 (5), D. N. Smith 1875 (MO, MOL, NY, USM); Prov. Oxapampa, Palcazu val- ley, Cabeza de Mono, 5-6 km W of Iscozacin, 17-20 Apr 1983 (? fr), D. N. Smith 3827 (AAU, MO). PUNO: Rfo Can- damo, nr. mouth of Rio Guacamayo, 25 May 1992 (st), Gen- try et al. 77156 (MO). UCAYALI: Prov. Coronel Portillo, Dtto. Campo Verde, rd. to Nueva Requena, km 11, 22 May 1988 (Y fl-fr), Arana 74 (K); Pucallpa, Rio Uyayali, 8 Apr 1953 (5), Ferreyra 9035 (USM), (Y fl-fr), Ferreyra 9042 (USM); Prov. Coronel Portillo, nr. Neshuya, between Pu- callpa & Tingo Maria, 20 Dec 1968 (5), Ferreyra 17649 (US, USM); Prov. Coronel Portillo, rd. Pucallpa-Tingo Maria, km, 86, Bosque Nacional de von Humboldt, 8 Aug, 1980 (5), Gentry et al. 29516 (MO); Prov. Coronel Portillo, Bosque Nacional de von Humboldt, 24 Jun 1981 (Y fr), Young et al. 1052 (BG, MO, NY, TEX).
BRAZIL. ACRE: Upper Rio Moa, Fazenda Arizona, 24-30 Sep 1984 (st), Campbell etal. 8162 (BG); Mun. Bras- ileia, Colocaqco Sao Jose, 25 km N of km 4 of rd. Brasileia- Assis Brasil, 26 May 1991 (5), Daly et al. 6722 (BG, INPA, NY); Mun. Xapuri, Rio Acre, 3 hr. downstream from Xap- uri, 5 Nov 1991 (Y fl-fr), Daly et al. 7143 (BG, NY); Mun. Marechal Taumaturgo, Rio Jurua, Fazenda Paraguay, 3 Apr 1993 (d), Daly et al. 7727 (NY); Hwy. Abuna-Rio Branco, km 242-246, nr. Campinas, 20 Jul 1968 (Y fl-fr), Forero et al. 6415 (GH, INPA, K, MG, MO, NY, S, U, US); nr. mouth of Rio Macaua, 9 Aug 1933 (9 fl-fr), Krukoff 5389 (A, BM, F, G, GB, K, MICH, MO, NY, S, U, US); Serra de Moa, 21 Apr 1971 (9 fr), Prance et al. 12171 (F, INPA, NY, U, US), 29 Apr 1971 (5), Prance et al. 12603 (F, INPA, MICH, MO, NY, P, S, U). AMAPA: Reserva INCRA Rio Falsino, 15-20 Aug 1985 (st), Campbell et al. 10334 (BG); Mun. Macand, rd. Serra do Navio (vill.)-Agua Branca, km 8, 4 Jan 1985 (9 fl-fr), Mori et al. 17671 (NY); Mun. Mazagao, Morro do Felipe V, 1 Oct 1986 (juv), Pires et al. 1393 (BG); Rio Ar- aguari, 101 tN, 52?8'W, 28 Sep 1961 (9 fl-fr), Pires et al. 51301 (MG, MO, NY, U, US); Mun. Macapa, rd. BR.210, 147 km NW of P6rto Grande, 30 Dec 1984 (9 fr), Rabelo 3112 (BG, NY); 2 km SE of Clevelandia, 2 Aug 1960 (9 fl-fr), Westra 47315 (BG, MG). AMAZONAS: Mun. Presi- dente Figueiredo, Rio Uatuma, Sep 1986 (9 fl-fr), Amaral 2082 (INPA); Rio Jurua, Lago do Curape, Saracura, 22 Aug 1975 (st), D. Coelho et al. (INPA) 52385 (INPA); Benjamin Constant, Mapatjirana, 16 Oct 1956 (9 fl-fr), Drees 5596
(INPA, U); Mun. Manaus, Reserva Florestal Ducke, 28 Sep 1962 (6), Duarte 7187 (F, G, INPA, RB); nr. mouth of Rio Embira, 13 Jun 1933 (6), Krukoff 4789 (A, BM, F, G, K, MICH, MO, NY, S, U, US); Mun. Humaita, nr. Tres Casas, 14 Sep-11 Oct 1934 (st), Krukoff 6237 (A, BM, F, G, K, MICH, MO, NY, RB, S, U, US); Mun. Sao Paulo de Oli- venqa, nr. Palmares, 11 Sep-26 Oct 1936 (Y), Krukoff 8083 (A, BM, F, G, K, MICH, MO, NY, P, S, U, US); Cano Tu- cano, Rio Cauaburf, 12 Nov 1965 (6), Maguire et al. 60161 (GH, K, MG, MO, NY, S, US); Distr. Agropecuario, ca. 2?24-25'S, 59?40-45'W, 15 Nov 1988 (6), Mori et al. 19821 (BG, INPA, NY); Mun. Manaus, rd. Manaus-Itacoa- tiara, km 26, Reserva Florestal Ducke, 6 Dec 1976 (6), Nas- cimento et al. (INPA) 66334; Mun. Manaus, ca. 80 km N of Manaus, Distr. Agropecuario da SUFRAMA, 19 May 1992 (Y fl-fr), Nee 42752 (INPA, K, NY); rd. Manaus-Itacoatiara, km 45, 29 Feb 1968 (Y fl), Rodrigues et al. 8439 (INPA, MO); Rio Jurua, Cachoeira, May 1901 (Y fl), Ule 5512 (MG). MARANHAO: Mun. Monqao, Rio Turiaqu, 4 km NW of Urutawy, 5 Feb 1985 (st), Balee et al. 408 (NY); Mun. Carutepara, Gurupiuna, 4 Nov 1986 (st), Bale et al. 2801 (BG, NY). MATO GROSSO: Aripuana, 29 May 1976 (Y fl- fr), Monteiro et al. 1126 (INPA); Rio Peixoto de Azevedo, 15 Aug 1980 (st), Werner 24/80 (MG). PARA: Ariramba, 1 Jun 1957 (6), Egler 439 (MG); Peixe Boi, Jul 1907 (6), Goeldi (MG) 8232 (BM, G, MG); Rio Tapaj6s, Boa Vista, 28 Jul 1933 (9 fl-fr), Kauffinann 603 (F); Tucuruf, Cagan- cho, Jan 1981 (st), Lisboa et al. 2271 (MG); rd. Belem- Brasilia, km 93, 5 Sep 1959 (6), Kuhlmann et al. 202 (GUA, MG, US); nr. Belem, Sep-Oct (st), Pires 51868 (NY, US); nr. Cachoeira, rd. BR.22, km 96, 29 Oct 1965 (9 fl), Prance et al. 1778 (GH, K, MO, NY, S, U, US); Mun. Almeirim, Monte Dourado, rd. to Sul de Pacanari, 20 Nov 1978 (6 fl), M. R. Santos 352 (MG); Rio Jarn, Monte Dourado, 15 Oct 1968 (6), N. T Silva 1226 (NY, U, US). ROND6NIA: Mun. Santa Barbara, rd. BR.364, km 120, 23 May 1982 (9 fr), Teixeira et al. 672 (INPA, NY). RORAIMA: Mun. Alto Ale- gre, Upper Rio Uaikas, Maiongong village, Sep 1994 (st), Milliken 2101 (BG); Serra dos Surucucus, 2042-47'N, 63?33-36'W, NW of Mission Station, 21 Feb 1969 (9 fl- fr), Prance et al. 10152 (INPA, NY, U, US).
BOLIVIA. BENI: Prov. Moxos, 130 km S of San Ig- nacio, 18 Oct 1991 (st), Del Aguila et al. 196 (BOLV, LPB); Prov. Vaca Dfez, rd. Riberalta-Cobija, km 6.7, 28 Sep 1993 (9 fr), Michel et al. 2437 (BG); Prov. Ballivian, Serranfa del Pil6n Lajas, 18 May 1989 (9 fl-fr), D. N. Smith 13218 (BG, BOLV, LPB, MO, USZ); 3 km E of Riberalta, rd. to Guayaramirim, 7 Jun 1982 (d), Solomon 7982 (LPB, MO, NY, U); Tumapasa, 11 Jan 1902 (st), Williams 400 (BM, K, NY, US). COCHABAMBA: Prov. Carrasco, Estaci6n Valle del Sajta UMSS, 29 Oct 1991 (st), Atahuachi et al. 24 (BOLV), Galarza et al. 22 (BOLV, LPB); Prov. Carrasco, rd. Santa Cruz-Villa Tunari, km 240, 8-9 Jul 1989 (9 fr), D. N. Smith et al. 13643 (BG, BOLV, LPB, MO, USZ). LA PAZ: Prov. Abel Iturralde, Alto Madidi, 21 May 1990 (st), Gentry et al. 70264 (MO). PANDO: Prov. Nicolas Suarez, Bella Flor, 10 Oct 1989 (9 fr), Beck et al. 19266 (BG, COL); Prov. Nicolas Suarez, Bella Flor, 10 Oct 1989 (9 fl-fr), Beck et al. 19266 (COL, LPB); Prov. Manupiri, 12 km W of Conquista, 4 Oct
172 FLORA NEOTROPICA
1991 (st), Beck et al. 20092 (BG, LPB); Prov. Nicolas Sua- rez, Cocamita, 4 Sep 1987 (J), Buchanan-Smith 3 (BG, K, LPB); Prov. Nicolis Suarez, Mukden, Jun-Dec 1979 (6), Izawa 25 (MO); Prov. Manuripe, 35 km N of Puerto Amer- icana, 3 Jun 1994 (Y fl-fr), Jardim et al. 871 (USZ); Prov. Madre de Dios, Concesi6n de Mobil, 20-25 Aug 1992 (Y fr), Killeen 4407 (BG, USZ); Prov. Nicolas Suarez, Cobija, 19 Oct 1978 (Y fr), Meneces 786 (BOLV, INPA); Rio Ma- deira, 6 km above Abuna, 11 Jul 1968 (Y fr), Prance et al. 5857 (GH, INPA, K, MG, MO, NY, P, S, U, US).
This species is rather uniform. The most apparent variation is found in the length of the petiolate parts of the segments of the lamina, varying from nearly absent to 6 cm long. Cecropia sciadophylla is the most common and widespread forest species through- out the Amazon basin and the Guianas. It usually out- numbers the other forest species. However, in the area in which this species co-occurs with C. herthae (Ec- uador, Colombia), the two species often occur in about equal proportions. These two species can be easily confused. They have the same general habit, but C. herthae does not have petiolulate bases of the lamina segments, the upper surface of the lamina is ? scabrous, and it has trichilia, although these are rather inconspicuous.
Vernacular names. Colombia: yemeoba (Mirania, Amazonas); zmuikaz (Amazonas); guarumo blanco (Casanare). Venezuela: yagrumo morado (Ama- zonas); sarasara-yek (Bolivar); yagrumo montaniero (Delta Amacuro). Guyana: congo pump, wanasoro (Arawak). Suriname: manbospapaja. French Guiana: diapapaYe, makamakalu (Wayana). Ecuador: su, yan- tan (Morona-Santiago); dundu (Quichua, Napo); hu- arumo, maguimehue (Huaorani, Napo); mangineowe (Haorani, Napo); su or suu (Shuar, Napo); yantan (Napo). Peru: yanat (Huambisa, Amazonas), setico colorado (Amazonas); torog (Cuzco); cetico colorado (Huanuco); Imuisai (Huitoto, Loreto); setico de al- tura, suu (Jivaro, Loreto); torototza (Machiguenga, Madre de Dios); setico colorado, setico roja, tacona, tacona ojo de venado (Pasco); setico colorado (Uca- yali). Brazil: imbauba gigante, imbauiba ver- melha (Acre); torem do igapo (Amapa'); tour6m da folha grande (Amazonas); ama'y-ata, ama'y-puku (Ka'apor, Maranhao); sakaka (Maiangong, Roraima). Bolivia: ambaibo blanco (Cochabamba); ambaibo colorado, toronoma (Pando).
52. Cecropia silvae C. C. Berg, Acta Bot. Neerl. 21: 655. 1972; Acta Amazonica 8(2): 180. 1978. Type. Brazil. Para: Rio Jarn, Monte Dourado, Planalto A, 11 Sep 1968 (9), N. T Silva 956 (holotype: U; isotypes: MG, NY, US).
Tree, to 30 m tall. Leafy twigs 2-4 cm thick, hir- tellous to subhispid. Lamina subcoriaceous, ca. 30 X 30 cm to 70 X 70 cm, the segments (12-)15-17, ob- lanceolate, the incisions down to near the petiole; api- ces obtuse to subacute to short-acuminate; upper sur- face scabrous to scabridulous, hispidulous, often also with sparse arachnoid indumentum; lower surface minutely puberulous to pubescent on the main veins, with arachnoid indumentum in the areoles, the smaller veins, or also on the main veins; lateral veins in the midsegment ca. 40-45 pairs, submarginally loop-connected, unbranched; petiole ca. 25-55 cm long, puberulous to subhirsute; trichilia fused, the brown indumentum intermixed with rather long stiff to soft white hairs; stipules 5-6 cm long, hirtellous to subhirsute outside, sericeous inside. Staminate inflo- rescences: peduncle ca. 9 cm long, hirtellous and with sparse arachnoid indumentum; spathe whitish, hirtel- lous and with dense arachnoid indumentum outside, glabrous inside; spikes ca. 40-50, 4-8 X ca. 0.3 cm, with stipes 0.2-0.3 cm long and with dense brown pluricellular trichomes; rachis hairy. Staminateflow- ers: perianth tubular, ca. 1-1.5 mm long, glabrous, the apex convex to plane; filaments flat; anthers ca. 0.5-0.6 mm long, appendiculate, detached at anthesis (?), reattached to the margins of the aperture by the appendages (?). Pistillate inflorescences in pairs (?), erect (?); peduncle 3-7 cm long, hirtellous; spathe ca. 6-7 cm long, the color and indumentum as in the staminate inflorescence; spikes 4, ca. 5-6 x ca. 0.4 cm, to 9 X 1.5 cm in fruit, sessile; rachis hairy. Pis- tillate flowers: perianth ca. 2 mm long, with arach- noid indumentum below the apex and on the apex to near the aperture outside, also below the style channel inside, the apex convex, puberulous on the margins of the aperture; style rather long; stigma penicillate. Fruit oblongoid, ca. 2.5 mm long, tuberculate, dark brown.
Distribution (Fig. 11.2). Brazil, northern Para, ex- tending to southern Suriname and French Guiana.
Specimens examined. SURINAME. Sipaliwini sa- vanne, 22 Jan 1970 (st), Oldenburger et al. 1112 (U).
FRENCH GUIANA. Piste St. Laurent-Apatou, km 46, 25 Feb 1984 (st, juv), Prevost 1727 (BM, INPA, NY, P, U).
BRAZIL. AMAPA: Mun. Mazagao, Morro do Felipe V, 1 Oct 1986 (st, juv), Pires et al. 1394 (BG). PARA: Rio Jacamacaru, 1 Jun 1957 (st), Egler 445 (MG); Rio Jari re- gion, rd. Planalto A.-Tinguelim, 24 Aug 1970 (6), Silva 3284 (IAN).
This species is distinguished by the narrow seg- ments of the lamina with numerous lateral veins in the free part of the midsegment. This species shows morphological affinities to Cecropia ulei. Due to the scarcity of material, there are some gaps in the de-
SYSTEMATIC TREATMENT 173
scription. The collections from Amapa' and French Guiana are made from juvenile trees, and their iden- tity is not certain.
53. Cecropia strigosa Trecul, Ann. Sci. Nat. Bot., Ser 3, 8: 82. 1847 (Aug). Type. Peru. Without locality, (d + Y), Ruiz & Pav6n s.n. (FI-W, photograph seen, mixed collection, the sheet with the stami- nate inflorescence here designated as the lecto- type; isolectotype: G).
Tree, to 10(-18) m tall. Leafy twigs (I.5-)3-1-0 cm thick, green or to dark brown, puberulous to hirtellous with straight hairs, sometimes densely hirtellous to brownish (or whitish) hirsute (or glabrous and bluish by a waxy layer); intemodes occasionally with ample brown pith. Lamina chartaceous to subcoriaceous, ca. 30 X 30 cm to 90 X 90 cm (to 140 X 140 cm), the segments (6-)8-10(-12), the free parts of upper seg- ments elliptic to subobovate to oblong, the incisions down to 5/10-9/10; apices short-acuminate (to sub- acute or to obtuse); upper surface smooth or ? scab- rous, often + bullate, sparsely to densely hispidulous (on the main veins); lower surface minutely puberu- lous (with uncinate to curved hairs) on the veins, often with sparse longer (uncinate to straight) hairs espe- cially on the main veins, or sometimes the main veins hirtellous to brownish (sub)hirsute, with dense arach- noid indumentum in the areoles and also on the smaller veins, sometimes extending to the main veins, occasionally almost absent; lateral veins in the free part of the midsegment 8-17 pairs, submarginally loop-connected, many of them branched, the lower ones usually not distinctly loop-connected; petiole (20-)35-70(-105) cm long, rather sparsely puberu- lous to hirtellous with straight hairs or sometimes densely brownish or whitish hirsute; trichilia fused or separate on the usually ? bulging (subscrotiform) base of the petiole, the brown indumentum intermixed with long white hairs or also with sparse to dense arachnoid indumentum; stipules (8-)10-35 cm long,
usually green to yellowish, sometimes partly reddish, or occasionally to red-brown, often subpersistent, (rather) sparsely puberulous to hirtellous or some- times densely brownish- (or whitish-)hirsute, occa- sionally partly setose with irritating hairs outside, gla- brous or sometimes (densely) hairy inside; terminal bud ? inflated. Staminate inflorescences in pairs (or solitary), often subtended by white, caducous bracts, to 1 cm, sometimes to 8 cm long, the peduncle erect to spreading and the spikes (sub)pendulous; peduncle 4-8(-10) cm long, puberulous to hirtellous; spathe 10-23 cm long, (pale) green to yellowish, (rather) sparsely puberulous to hirtellous, or sometimes densely brownish to whitish hirsute outside, glabrous or hairy inside; spikes ca. (15-)25-100, (3-)5-22 X
ca. 0.2-0.3 cm, with stipes (0.5-)1-2.5 cm long and minutely puberulous; rachis (sub)glabrous or short- hairy. Staminate flowers: perianth short-tubular to cup-shaped, 0.8-1.2 mm long, 2-lobed, glabrous; fila- ments slightly swollen; anthers ca. 0.5-0.7 mm long, not appendiculate, at anthesis not detached. Pistillate inflorescences in pairs, erect, soon pendulous, often subtended by whitish, caducous bracts, to 8(-15) cm long; peduncle 4-16(-21) cm long, (sparsely) puber- ulous to hirtellous to subhirsute or also with arachnoid indumentum, sometimes densely brownish hirsute, occasionally partly setose with irritating hairs; spathe 8-18(-23) cm long, the color and indumentum as in the staminate inflorescence; spikes 3-7(-9), occasion- ally tortuose, 6-15(-33) X 0.3-0.6 cm, to 25(-40) X 1(-1.5) cm in fruit, (sub)sessile or with stipes to 0.9 cm long and densely hairy; rachis (sub)glabrous. Pis- tillate flowers: perianth, ca. (0.6-)l.2-1.5(-2.3) mm long, with arachnoid indument below the apex out- side, also in the lower part of the style channel inside, the apex convex with a collar-shaped rim around the aperture, punctate to muriculate; style short; stigma comose. Fruit ellipsoid to oblongoid, 1.5-2 mm long, almost smooth.
Distribution (Fig. 18.2). From northern Peru to Bolivia, in forest and secondary growth, at 400-1900 m.
Representative specimens examined. PERU. Cuzco: Prov. Paucartambo, between Atalaya & Pilcopata, 31 Jul 1988 (d), Berg et al. 1617 (BG, COL, Centro de Medicina Andina, Cuzco); Prov. La Convenci6n, Quillabamba, Salas- pampa, rd. to Kitem, Rio Urubamba, 1100 m, 28 Oct 1986 (Y fl), Nuiiez et al. 6304 (MO); Prov. La Convenci6n, Quil- labamba, Mandor, 1200 m, 16 Oct 1987 (Y fl), Nuihez et al. 8230 (BG, MO); Prov. Urubamba, rd. Cuzco-Machupicchu, km 112, 1050-1300 m, 18 Nov 1987 (d), Nuiiez 8616 (MO); Prov. Quispicanchis, Puente Inambari, 600 m, 8 Dec 1966 (d), Vargas C. 18459 (US). HUANCAVELICA: Prov. Tayacaja, nr. Marabamba, 1700 m, 25 Apr 1964 (Y fl-fr),
174 FLORA NEOTROPICA
Tovar 4798 (US). HUANUCO: Rd. Huanuco-Tingo Maria, below Chinchoa, ca. 1800 m, 23 Nov 1997 (d), Berg et al. 1734 (BG, COL, MOL); rd. Huanuco-Tingo Maria, nr. Santa Catalina, ca. 1400 m, 24 Nov 1997 (Y fl-fr), Berg et al. 1739 (BG, COL, MOL); rd. Tingo Maria-Aucayacu, nr. Pumahuasi, 25 Nov 1997 (6), Berg etal. 1745 (BG); 69 km NE of Tingo Maria, on rd. to Tocache, 16 Jul 1982 (Y fl- fr), Gentry et al. 37616 (BG); Prov. Puerto Inca, Dtto. Yu- yapichis, DANTAS, 14 Apr 1989 (Y fl-fr), Kroell S. 251 (BG, G), 16-31 Mar 1991 (Y fl), Tello 1117 (G, NY), (6), Tello 1118 (G, NY). JUNiN: Rd. Tarma-San Ram6n, ca. km 75, ca. 1800 m, 28 Nov 1997 (Y fl-fr), Berg et al. 1754 (BG, COL, MOL), (6), Berg et al. 1755 (BG, COL, MOL); La Merced, 29 May-4 Jun 1929 (st), Killip et al. 24047 (F, NY, US); Prov. Chanchamayo, rd. to Mina Pichita, 1880 m, 2 Feb 1999 (Y fl), Ntiunez 68 (BG), 3 Feb 1999 (d), Niniez 75 (BG); nr. San Ram6n, La Florencia, 1800 m, 13 Sep 1989 (Y), Reynel et al. 4692 (MO). MADRE DE DIos: Prov. Manu, Parque Nacional Manu, Rio Manu, Rio Sotileja, 2 Oct 1986 (d), Foster et al. 11570 (BG), (Y fl), Foster 11876 (BG, LPB, MOL, US). PASCO: Rd. Puente Paucartambo- Oxapampa, ca. 1000 m, 28 Nov 1998 (6), Berg 1756 (BG, COL, MOL); 3.7 km N of Puente Paucartambo, 900 m, 30 Jan 1983 (9 fl-fr), Gentry et al. 39840 (BG, MO, MOL, TEX); Iscozacin, 1 Apr 1986 (st, juv), Salick 7390 (NY). SAN MARTiN: Prov. Lamas, rd. Lamas-Pamashto, ca. 700 m, 4 Dec 1997 (9 fl-fr), Berg et al. 1764 (BG, COL, MOL); Prov. San Martin, rd. Tarapoto-Yurimaguas, ca. km 12, ca. 700 m, 5 Dec 1997 (6), Berg etal. 1772 (BG, COL, MOL); Prov. Moyobamba, rd. Moyobamba-Jepelacio, ca. km 8, ca. 1100 m, 8 Dec 1997 (9 fl-fr), Berg et al. 1790 (BG, COL, MOL); San Roque, 1350-1500 m, Jan-Feb 1930 (9 fl-fr), LI. Williams 2373 (US). UCAYALI: Rd. Tingo Maria-Pu- callpa, km 82, nr. Boquer6n, 26 Nov 1997 (9 fl), Berg et al. 1746 (BG, COL, MOL), (6), Berg et al. 1747 (BG, COL, MOL); rd. Tingo Maria-Pucallpa, ca. km 125, 26 Nov 1997 (9 fl-fr), Berg et al. 1748 (BG, COL, MOL); Prov. Coronel Portillo, San Alejandro, between Pucallpa & Tingo Maria, km 115, 8 Apr 1953 (9 fl), Ferreyra 9036 (US); rd. Pu- callpa-Tingo MarIa, km 90-130, Bosque Nacional Alexan- der von Humboldt, (9 fl-fr), Gentry et al. 41409 (BG, NY).
BOLIVIA. BENI: Prov. Ballivian, rd. Yucumo-Caran- avi, km 13, 900 m, 4 Mar 1993 (d), Berg 1712 (BG, COL, LPB); Prov. Ballivian, Serrania Pil6n Lajas, rd. Caranavi- San Borja, 25 km from Yucumo, 850-900 m, 16-26 Feb 1990 (9 fl-fr), D. N. Smith et al. 13989 (USZ, MO), 14013 (BG, MO). COCHABAMBA: Rd. Cochabamba-Villa Tunari, ca. km 95, 1800 m, 6 Dec 1995 (9 fl-fr), Berg et al. 1723
(BG, BOLV, COL, LPB), (6), Berg et al. 1725 (BG, BOLV, COL, LPB). LA PAZ: Prov. Sud Yungas, rd. Caranavi-Yu- cumo, nr. La Cascada, ca. 2.5 km SSW of Quiquibey, 900 m, 4 Mar 1993 (9 fl-fr), Berg 1711 (BG, COL, LPB), (9 fl-fr), Berg 1714 (AAU, BG, COL, LPB, MO, NY).
The trichilia are either fused or separate, even on the same tree. In the southern part of the species range, the trichilia are more commonly separate than in the northern part of the range. In the southern part of the range, reddish-colored parts are lacking, but in
the northern part the presence of a red dye in the "um- bilicus" and the basal parts of the main veins, or also in the stipules, is not uncommon. In the southern part of the species range, the incisions of the lamina are mostly down to 7/10, but in the northern part, the incisions are often down to 8/10 or even 9/10, the segments are often narrower (obovate to oblong), and the lateral veins of the midsegment are more numer- ous. The base of the petiole is mostly ? scrotiform, but in particular in the northern part of the species range sometimes or often hardly so. The terminal buds are ? conspicuously inflated. The spathe of sta- minate inflorescences often remains hanging on one or two of the longest spikes; the tips of these spikes are curved into the calyptrate apex of the spathe. The spikes of the pistillate inflorescences are usually to 15 cm long at anthesis, but in Ucayali (Peru) they can be to 33 cm long. The number of lamina segments is usually 9-10, but in Huanuco (Peru) individuals with 10-12 segments can be found. Material with densely hirsute leafy twigs, stipules, petioles, veins of the lam- ina beneath and/or spathes can be found in Huanuco, Ucayali and San Martfn (Peru). This species is prob- ably related to Cecropia marginalis and to C. mem- branacea.
Vernacular names. Peru: tacona blanco (San Martin); shiari (Ucayali).
54. Cecropia subintegra Cuatrecasas, Rev. Acad. Colomb. Ci. Exact. 6(22/23): 279. 1945. Type. Co- lombia. Valle: Rio Digua, Piedra de Moler, 1000 m, 25 Aug 1953 (9), Cuatrecasas 15111 (holo- type: COL; isotypes: F, US).
Tree, to 20 m tall. Leafy twigs 1-3 cm thick, green, puberulous to subhispidulous with straight to curved to uncinate hairs. Lamina chartaceous to subcoria- ceous, ca. 20 X 20 cm to 50 X 50 cm, 3-lobed, the main radiating veins 7-10, the incisions down to ca. 3/10-4/10 in the upper part of the lamina and the lower part shallowly lobate to subentire; apices acu- minate to acute; upper surface smooth, sparsely pu- berulous; lower surface minutely puberulous and with sparse longer uncinate hairs on the veins, with arach- noid indumentum absent or confined to the margin; lateral veins in the free part of the midsegment 8-10 pairs, submarginally, unbranched or some (lower ones) branched; petiole ca. 10-40 cm long, minutely puberulous or also with sparse to rather dense arach- noid indumentum; trichilia fused, the brown indu- mentum intermixed with short white hairs; stipules 4-12 cm long, (purplish-)red or green with red stripes,
SYSTEMATIC TREATMENT 175
(sparsely) puberulous and with brown pluricellular hairs or also with arachnoid indumentum outside, hairy to subglabrous inside. Staminate inflorescences solitary, erect; peduncle 2.5-3.5 cm long, puberulous; spathe ca. 10-15 cm long, (purplish) red, sparsely pu- berulous and with dense brown pluricellular hairs out- side, glabrous or sparsely hairy inside; spikes ca. 7- 12, ca. 9-12 X 0.3 cm, with stipes 0.3-0.5 cm long and puberulous; rachis sparsely hairy. Staminateflow- ers: perianth tubular, 1-1.5 mm long, with short arachnoid indumentum below the apex, the apex con- vex, smooth to slightly tuberculate, the aperture sur- rounded by a rim; filaments flat; anthers 0.5-0.6 mm long, appendiculate, detached at anthesis, reattached to the margins of the aperture by the appendages (?). Pistillate inflorescences in pairs or solitary, erect to pendulous; peduncle 3-24 cm long, puberulous to subhispidulous; spathe 12-20 cm long, the color and indumentum as in the staminate inflorescence; spikes 4(-6), ca. 9-12 X 0.5-0.6 cm, to 32 X 0.8-1 cm in fruit, sessile or with stipes to 1 cm long and puberu- lous; rachis hairy. Pistillate flowers: perianth 1.5-2 mm long, with arachnoid indumentum below the apex outside, also in the style channel inside, the apex con- vex to conical, muriculate; style rather long, straight; stigma comose. Fruit ellipsoid, 2-2.5 mm long, smooth, dark brown.
Distribution (Fig. 18.8). Pacific coastal region of Colombia (Valle and Nariiio) and Ecuador (Carchi), in forest, at 800-1300 m.
Specimens examined. COLOMBIA. NARINO: Mun. Barbacoas, Cgto. Altaquer, Vrda. El Barro, 1325 m, Franco et al. 4909 (BG); Mun. Barbacoas, El Diviso, 850 m, 7 Oct 1993 (Y fl-fr), Franco et al. 4463 (BG); Mun. Barbacoas, Cgto. Altaquer, rd. El Barro-Junin, El Mirador, 1150 m, 12 Dec 1993 (6), Franco et al. 5174 (BG); Mun. Barbacoas, rd. Altaquer-El Diviso, nr. Reserva Colibries, 1300 m, 10 Mar 1995 (9 fl-fr), Lozano C. et al. 6906 (COL); Mun. Barbacoas, Cgto. Junin, rd. Junfn-Barbacoas, ca. 950-1100 m, 14 Mar 1995 (d), Lozano C. et al. 6987 (COL); Ricaurte, ca. 1300 m, 30 Mar 1941 (9 fr), Sneidern A.485 (A, MICH, NY, S, TEX, U, US). VALLE: Mun. Trujillo, rd. Trujillo- Naranjal, km 20.4, Rio Sanquinini, 1200 m, 11 Oct 1993 (d), Franco et al. 4482 (BG, TULV).
ECUADOR. CARACHI: N of Lita, N of Rio Mira & E of Rio Baboso, 1 Jul 1994 (st), Boyle et al. 3304 (BG).
The shallowly incised leaves are found only on lateral branches. Before branches are produced, the leaves on the trunk are more or less deeply incised and often larger. In the monocaul state, the leaves are similar to the normal C. obtusifolia. Subjuvenile traits seem to reappear in the leaves of the branches, not only in the first leaves formed, but persistently. This
submontane species shows close affinities to Cecropia obtusifolia, in particular to the type with relatively few segments of the lamina. Cecropia subintegra may prove to be distinct only at the level of subspecies.
55. Cecropia tacuna C. C. Berg & P. Franco, Novon 6: 248. t. 13, 1996. Type. Peru. Pasco: Prov. Ox- apampa, 5 km SE of Oxapampa, 1850 m, 25 May 1983 (Y fl-fr), D. N. Smith 4179 (holotype: MO; isotypes: BG, K). Fig. 44
Tree, to 20(-30) m tall. Leafy twigs 4-8 cm thick, green to bluish gray, densely villous and with filiform brown pluricellular hairs (or only with these pluri- cellular hairs). Lamina (sub)coriaceous, ca. 50 X 50 cm to 100 X 100 cm, the segments 10-12 or 13-18 (but on new shoots down to 8), the free parts of the segments oblanceolate to oblong, the incisions down to ca. 6/10-9/10; apices subacuminate to acute; upper surface scabrous to scabridulous, sparsely to rather densely hispidulous to subhispid or to hirtellous or largely hirsute to subvillous and with sparse to dense filiform brown pluricellular hairs, "umbilicus" prom- inent and villous; lower surface rather sparsely sub- villous and with filiform brown pluricellular hairs or also sparse white arachnoid indument on the main veins, varying to pilose (to subtomentose) on the lesser veins, with arachnoid indumentum in the are- oles and on the reticulum; lateral veins in the free part of the midsegment 15-25 or 25-35 pairs, most of them unbranched or the lower ones branched, sub- marginally ( ? faintly) loop-connected; petiole 40-80 cm long, ? densely (sub)villous and with filiform brown pluricellular hairs and sparse arachnoid indu- ment or sparsely villous in the upper part, glabrescent or (sub)glabrous; trichilia absent; stipules 15-35 cm long, green (or red to grayish), caducous, densely vil- lous outside, sparsely to densely hairy inside. Stami- nate inforescences in pairs, erect to spreading with the spikes curved upward, often subtended by cadu- cous bracts, to 13 cm long; peduncle 5-12 cm long, ? densely white-villous (at least in the upper part) and also or only with filiform brown pluricellular hairs, often also with sparse white arachnoid indu- mentum or (sub)glabrous; spathe 14-20 cm long, green (or purple), with dense filiform brown pluricel- lular hairs and sparsely villous (glabrescent) outside, hairy inside; spikes ca. 10-25, 12-20 X 0.3-0.5 cm, sessile or with stipes to 1 cm long and with brown pluricellular hairs or also sparsely hirtellous; rachis hairy with stiff hairs and arachnoid indumentum. Sta- minate flowers: perianth tubular, ca. 1.2-1.8 mm long, with short arachnoid indumentum below the
176 FLORA NEOTROPICA
FIG. 44. Cecropia tacuna. 1. Lamina, reduced. 2. Apex of lamina and venation. 3. Stipules and young leaf (Berg et al. 1635). 4. Staminate inflorescence with spathe. 5. Staminate flower. 6. Stamen and filament after detachment of anther (Berg et al. 1634). 6. Pistillate inflorescence at anthesis. 7. Pistillate flower and fruit. 8. Pistillate flower. 9. Fruit (Berg et al. 1635). (From Novon 6: 249. 1996, modified.)
SYSTEMATIC TREATMENT 177
apex, the apex plane to slightly convex, sparsely to densely muriculate; filaments swollen; anthers ca. 0.6-0.8 mm long, not appendiculate, remaining at- tached at anthesis. Pistillate inflorescences in pairs, erect to spreading and with the spikes curved upward, often subtended by caducous bracts, to 13 cm long; peduncle 5-12 cm long, with indumentum similar to that of the staminate inflorescence; spathe 9-14 cm long, the color and indumentum as in the staminate inflorescence; spikes 3-7, 6-13 X ca. 0.5 cm, to 22 X 1.5 cm in fruit, (sub)sessile; rachis hairy with stiff hairs and arachnoid indumentum. Pistillate flowers: perianth, ca. 1.5-2 mm long, with arachnoid indu- mentum below the apex outside, also in the lower part of the style channel inside, the apex plane to slightly convex, muriculate; style long, slightly curved, mur- iculate; stigma comose. Fruit oblongoid, ca. 1.5 mm long, finely tuberculate.
Distribution (Fig. 8.1). Peru, from Pasco to Cuzco, in cloud forest, at (800-)1800-2500(-2700) m.
Specimens examined. PERU. AYACUCHO: Between Huanta & Rfo Apurimac, 750-1000 m, 7-17 Nov 1929 (juv), Killip et al. 23117 (NY, US). Cusco: Prov. Paucar- tambo, rd. Pilcopata-Paucartambo, ca. 2000 m, 2 Aug 1988 (d), Berg et al. 1634 (BG, Centro de Medicina Andina, Cusco), 2200-2300 m, 2 Aug 1988 (Y fl-fr), Berg et al. 1635 (BG, COL, Centro de Medicina Andina, Cusco); Machu Pic- chu, 19 Apr 1957 (st), Ellenberg 880 (U); Prov. Paucar- tambo, rd. Paucartambo-Pilcopata, km 132, 2260-2290 m, 27 Jun 1978 (st), Gentry et al. 23564 (BG); Prov. Urubamba, Machu Picchu, 2000 m, 4 Jul 1972 (5), Mueller 2861 (LZ); Prov. Urubamba, between Pampacahua & Cedrobamba, Ma- chu Picchu, 2800 m, 8 Jul 1989 (st), Nuhez et al. 11111 (MO); Prov. Urubamba, nr. Machu Picchu, Rio Mandor, 2055 m, 2 Jun 1982 (5), Peyton et al. 374 (MO); Prov. Paucartambo, Kosfiipata, 2700 m, 4 Jul 1972 (5), Vargas C. 15490 (US). HUANUCO: Rd. Huanuco-Tingo Maria, Car- pish, ca. 2500 m, 23 Nov 1997 (Y fl-fr), Berg et al. 1731 (BG, COL, MOL), 2400 m, 10 Mar 1982 (5), Gentry et al. 36153 (BG, MO, USM). PASCO: Prov. Oxapampa, Rio Bo- queria, ca. 26 km from Oxapampa, via Rio Yamaquizu, 2040 m, 3 Jun 1982 (d), D. N. Smith etal. 1832 (BG, MO, USM); Prov. Oxapampa, 5 km SE of Oxapampa, 1850 m, 9 Apr 1983 (5), D. N. Smith 3663 (BG, MO, MOL); Prov. Oxa- pampa, Huancabamba, Rio Yanachaga, 2280 m, 26 May 1983 (Y fl), D. N. Smith et al. 4196 (BG, MO); Prov. Oxa- pampa, Oxapampa-Villa Rica rd., 29 Sep 1983 (D fl-fr), D. N. Smith et al. 5340 (BG, MO); Prov. Oxapampa, RIo San Albert valley, E of Oxapampa, 2300 m, 4 Jul 1984 (st), D. N. Smith et al. 7608 (BG, MO, USM); Prov. Oxapampa, Yanachaga via Rio San Daniel, 2500 m, 17 Jul 1984 (st), D. N. Smith et al. 7848 (BG, MO, USM); Prov. Oxapampa, Rio Alberto valley, E of Oxapampa, slopes of Cord. Yana- chaga, 2400 m, 23 Jul 1984 (st), D. N. Smith et al. 7974
(BG, MO, USM); Prov. Oxapampa, Palmazu, 2100 m, 28 Sep 1984 (Y fl-fr), D. N. Smith 8555 (BG, MO). SAN MARTiN: Prov. Rioja, rd. Pedro Ruiz-Moyobamba, 2100 m, 7-9 Aug 1983 (d), D. N. Smith et al. 4686 (MO).
This species is probably related to the Bolivian montane species, Cecropia elongata. It shows simi- larities to the villous forms of C. angustifalia (see page 62) in the indumentum. In the collections from Huainuco (Peru), the leafy twigs are subglabrous, bearing only brown pluricellular hairs, the peduncles are glabrous, and the petioles are glabrous or sparsely villous only in the upper part. Moreover, these col- lections differ in the less deeply incised laminas (down to 6/10-7/10), fewer incisions (10-12), and fewer lateral veins in the free part of the midsegment (15-25 pairs), features matching those of subjuvenile leaves of material from Cusco and Pasco. Ants are absent.
Vernacular name. Peru: tacuna (Pasco).
56. Cecropia telealba Cuatrecasas, Revista Acad. Colomb. Ci. Exact. 6(22/23): 293 (1945). Type. Colombia. Valle: Rio Cali, Quebradahonda, above La Glorieta, rd. to Miralindo, 2100-2250 m, 31 Oct-I Nov 1044 (d), Cuatrecasas 18406 (holo- type: COL; isotypes: F, US).
Cecropia telealba Cuatrecasas var. hirsuta Cuatrecasas, Revista Acad. Colomb. Ci. Exact. 6(22/23): 294 (1945). Type. Colombia. Valle: Rio Calima, El Cairo, 1650-1750 m, 6-7 Jan 1943 (Y), Cuatrecasas 13947 (holotype: COL; isotype: US).
Cecropia alborugosa Cuatrecasas, Revista Acad. Col- omb. Ci. Exact. 9(36/37): 330. 1956. Type. Colom- bia. Valle: Rio Digua, Quebrada del San Juan, be- tween Paraguita & Queremal, 1570-1740 m, 17 Mar 1947 (d), Cuatrecasas 23841 (holotype: US; iso- types: F, VALLE).
Tree, to 20(-30) m tall. Leafy twigs 1.5-5 cm thick, green, hirsute, sometimes only at the stipular scars; intemodes partly to entirely (?) filled with orange-brown pith. Lamina coriaceous, ca. 20 X 20 cm to 90 X 90 cm, the segments 8-10, the free parts of the upper segments obovate or elliptic, the inci- sions down to 7/10-8/10; apices acute to obtuse; up- per surface smooth, often ? rugose, very sparsely strigose and with dense to + sparse arachnoid indu- mentum (or subglabrous), lower surface sparsely to densely hirsute to hirtellous on the main veins, hir-
178 FLORA NEOTROPICA
tellous to puberulous on the smaller veins, with arach- noid indumentum in the areoles or extending to the smaller veins or also to the main veins (not distinctly in two layers as in C. telenitida); lateral veins in the free part of the midsegment ca. 15-25(-30?) pairs, submarginally loop-connected, unbranched or some- times branched; petiole 15-65 cm long, sparsely sub- villous in the upper and lower part or also with arach- noid indumentum; trichilia fused (or separated), the brown indumentum intermixed with short (or also long) white hairs; stipules (6-) 13-32 cm long, reddish to purplish or pale green to yellowish, hirsute to sub- villous (to rather sparsely strigose to hirtellous to sub- sericeous outside, sparsely hairy inside. Staminate in- florescences in pairs or solitary, erect; peduncle 5-7 cm long, subvillous, often only in the upper and lower part; spathe 9-17 cm long, pink to reddish to purplish or pale green to yellowish, subvillous outside, gla- brous inside; spikes 8-12, 5-12 X 0.5-0.9 cm, with stipes 0.5-1.2 cm long and sparsely hairy or glabrous; rachis hairy. Staminate flowers with pedicels to 1.5 cm long and minutely puberulous; perianth tubular, ca. 1.2-2 mm long, minutely puberulous below the apex, the apex plane to slightly convex; filaments flat; anthers 0.6-0.8 mm long, appendiculate, detached at anthesis (?), reattached to the margins of the aperture by the appendages (?). Pistillate inflorescences in pairs or solitary, erect, later on deflexed, subtended by caducous bracts, to 4 cm long; peduncle 3-11 cm long, subvillous, often only in the upper and lower part or subglabrous; spathe 8-13 cm long, the color and indumentum as in the staminate inflorescence; spikes l-3(-4), (1.4-)5-9 X 0.4-0.6 cm, to 32 X 2.5 cm in fruit, sessile or with stipes to 1 cm long and subvillous; rachis hairy. Pistillate flowers: perianth ca. 2-3 mm long, with arachnoid indumentum below the apex outside, also in the lower part of the style channel inside, the apex convex, muriculate to min- utely puberulous (only in the lower part); style long, straight, minutely puberulous; stigma penicillate. Fruit oblongoid, 3.5-4 mm long, finely tuberculate.
Distribution (Fig. 10.2). Colombia (Caldas to Cauca), in (cloud) forest, in the western Cordillera at 1300-2500 m.
Representative specimens examined. COLOMBIA. CALDAS: Rio Boquia, Salento, 1600-1900 m, 22 Jul 1922 (st), Killip et al. 8827 (GH). CAUCA: Between Santander de Quilichao & Rfo Ovejas, 1500 m, 3 Feb 1965 (d), Cuatre- casas et al. 26894 (VALLE- under the same number a col- lection from Valle); Sevilla, Las Brisas, 2000 m, 10 Apr 1970 (Y fl-fr), Cuatrecasas et al. 28946 (US); between San Antonio & Rio Ortega, 2000-2200 m, 1-2 Jul 1922 (st), Pennell et al. 8042 (GH). QUINDIO: Without locality, 1200
m, Mar 1876 (st), Andre 2574 (K, NY); Mun. Circasia, Vrda. Barcelona Alta, Finca Buenos Aires, 1690 m, 23 Mar 1991 (6), Agudelo et al. 1043 (COL); Mun. Armenia, Vrda. Las Colillas, Finca Las Mercedes, 1700 m, 12 Jun 1992 (9 fl- fr), Wlez et al. 2926 (COL). RISARALDA: Nr. Termales 2060-2130 m, 13 Aug 1995 (9 fr), Wijninga 541 (BG, U). VALLE: Rio Cali, Rio Pichind6, Alto de las Brisas, 2050- 2100 m, 29 Oct 1944 (9 fl-fr), Cuatrecasas 18353 (COL, F, US); Rio Pichinde, Los Carpatos, 2250-2350 m, 24 Jul 1946 (9 fr), Cuatrecasas 21691 (F, US, VALLE); El Tabor, above Las Brisas, 1970-2100 m, 19 Oct 1946 (9 fl), Cua- trecasas 22304 (F, US); above La Carbonera, between Las Brisas & Alban, 2000 m, 24 Oct 1946 (st), Cuatrecasas 22504 (F); nr. Las Mesitas, above Villa Colombia, El Placer, 1800 m, 4 Feb 1965 (6), Cuatrecasas et al. 26894 (TULV, US); Mun. Trujillo, rd. Trujillo-Naranjal, km 9, 1800 m, 1 1 Oct 1993 (9 fl-fr), Franco et al. 4479 (BG, COL); Mun. San Pedro, rd. Buenos Aires-San Pedro, km 15, 1900 m, 13 Oct 1994 (6), Franco 4490 (BG); Mun. Yotoco, El Canay, rd. Medio Canoa-Darien, 1500 m, 15 Mar 1994 (6), Franco et al. 4528 (BG); Mun. Loboguerrero, rd. to Buenaventura, 1350 m, 15 May 1994 (9 fl-fr), Franco et al. 4530 (BG, TULV); Santa Helena, above Topacio, edge of Los Faral- lones de Cali National Park, 1920 m, 12 Dec 1985 (9 fl-fr), Gentry et al. 53202 (MO); E of Bitaco, 1500 m, 16 Nov 1963 (9 fr), Hutchinson et al. 3039 (F, G, K, MICH, NY, P); Mun. Anserma Nuevo, rd. Anserma-San Jos6 del Palmar, 1860 m, 12 Nov 1985 (6), Lozano C. et al. 4978 (COL); Mun. Darien, Alto Calima, 1970 (6), Mahecha s.n. (UDBC 5789).
Cecropia telealba shows affinities to C. gabrielis, from which it can be distinguished by the (usually) dense arachnoid indumentum on the upper surface of the lamina and the consistent presence of trichilia.
Cecropia gabrielis (normally) occurs at lower eleva-
tions than C. telealba. It is the conspicuous white-
leaved species on the mountain slopes bordering the
Cauca valley.
57. Cecropia telenitida Cuatrecasas, Revista Acad.
Colomb. Ci. Exact. 6(22/23): 295. 1945; Velas- quez, Acta Bot. Venez. 6: 62, t. 15. 1971. Type. Colombia. Norte de Santander: Mun. Toledo, Rio Samaria, 2000-2100 m, 30 Oct 1941 (9), Cua-
trecasas, Schultes & E. Smith 12781 (holotype: COL; isotype: F). Fig. 45
inflorescence at anthesis. 8. Pistillate flower and style. 9. Fruit (Berg et al. 1666). (From Fl. Ecuador 48: 11l. 1993, modified.)
180 FLORA NEOTROPICA
Cecropia teleincana Cuatrecasas, Revista Acad. Col- omb. Ci. Exact. 6(22/23): 296. 1945. Type. Colom- bia. Antioquia: Alto del Toyo, between Antioquia & Canias Gordas, 2200 m, 2 Mar 1942 (d), Cuatreca- sas 13548A (holotype: COL; isotype: F).
Cecropia santanderensis Cuatrecasas, Revista Acad. Colomb. Ci. Exact. 6(22/23): 296. 1945; Velasquez, Acta Bot. Venez. 6: 58, t. 13. 1971. Type. Colombia. Norte de Santander: Rfo Chitaga, Ventanas, Alto del Loro, 1820 m, 17 Oct 1941 (Y), Cuatrecasas et al. 12346 (holotype: COL; isotype: F).
Cecropia angelica C. C. Berg & P. Franco, Fl. Ecuador 48: 9, t. 1. 1993. Type. Ecuador. Morona-Santiago: Angel Sabio, between Lim6n (Gral. Plaza) & Gual- aco, ca. 2250 m, 28 Jan 1981 (Y), Berg 1229 (ho- lotype: QCA; isotypes: AAU, BG, COL, NY, U).
Tree, to 15(-30) m tall. Leafy twigs 2-8 cm thick, green to bluish by a waxy layer, glabrous or (initially) with dense brown pluricellular trichomes, hirtellous, villous to hirsute, or puberulous with straight white (unicellular) hairs; intemodes with scarce pith. Lam- ina (sub)coriaceous, ca. 30 X 30 cm to 90 X 90 cm (to 100 X 100 cm), the segments 7-10(-1 1), the free parts of the upper segments ovate to elliptic to oblong to (sub)obovate, the incisions down to 4/10-9/10; api- ces rounded to short-acuminate; upper surface smooth or scabridulous (to scabrous), very sparsely to densely puberulous (to hirtellous or subvelutinous) or hispidulous and with sparse to dense arachnoid in- dumentum, often slightly bullate, lower surface hirtellous to subtomentose (or puberulous), espe- cially on the main veins hirsute to villous, or main veins subglabrous, with very short arachnoid indu- mentum in the areoles or also on the reticulum and usually very sparse to rather dense longer arachnoid indumentum on the main veins and supported by the other indumentum on the smaller veins (the arach- noid indumentum thus in two distinct layers); lateral veins in the free part of the midsegment 7-16 pairs, submarginally loop-connected, unbranched or branched, the smaller veins often prominent; petiole ca. 25-90 cm long, glabrous, villous in the upper or also the lower part, also with arachnoid indumentum and/or dense brown pluricellular trichomes (some- times at the base dense, and suggesting the occur- rence of trichilia, but Muillerian bodies lacking), or puberulous; trichilia absent or present, weakly developed without Mullerian bodies or with Muller- ian bodies and separate or fused (and then sometimes lobate), the brown indumentum intermixed with short or also long white hairs; stipules (12-)20-55 cm long, green to orange to pink to reddish to dark maroon, with sparse to dense dark brown pluricellu- lar hairs and sparsely minutely puberulous to tomen-
tellous or often also sparsely to densely hirsute to villous to sericeous, subglabrous, subhirsute or hirtellous outside, densely to sparsely subvillous to subsericeous or subglabrous inside. Staminate inflo- rescences solitary or in pairs, (usually/always?) sub- tended by caducous bracts, to 8 cm long, the pedun- cle erect and the spikes spreading to subpendulous; peduncle 2-11 cm long, glabrous or in the upper part sparsely to densely (sub)villous or puberulous to hir- tellous to subhirsute; spathe 9-24 cm long, red- brown to orange to pinkish or yellowish brown or yellowish, rather sparsely to densely (sub)villous to hirsute or also with sparse arachnoid indumentum outside, sparsely hairy or glabrous inside; spikes 5- 12(-16), 6-25 X ca. 0.5-1 cm, sessile or with stipes to 1 cm long and hairy; rachis hairy. Staminate flow- ers: perianth tubular, 1-2(-2.5) mm long, (sparsely) puberulous or glabrous below the apex, the apex slightly convex, glabrous; filaments swollen; anthers 0.6-0.8 mm long, detached at anthesis (?). Pistillate inflorescences solitary or in pairs, erect to deflexed, (usually/always?) subtended by caducous bracts, to 8 cm long; peduncle 4-12(-15) cm long, glabrous or sparsely villous or ? densely hirtellous to puberu- lous; spathe 6-13 cm long, the color and indumen- tum as in the staminate inflorescence; spikes (I-)4- 5(-6), purplish or reddish, 3-12 X 0.7-1.6 cm, to 25 X 2.5 cm in fruit, subsessile (or with to 2 cm long stipes); rachis subglabrous. Pistillate flowers: peri- anth 2-2.5 mm long, with arachnoid indumentum be- low the apex outside, also below the style channel inside (or the style channel minutely puberulous), the apex conical to convex, hispidulous near the slit- shaped aperture; style rather long, puberulous; stigma comose to subpeltate. Fruit oblongoid to sub- obovoid, 2-2.5 mm long, tuberculate or smooth.
Distribution (Fig. 18.7). The Andean region of Venezuela to southern Colombia, in the central and eastern cordilleras, and from southern Ecuador to northern Peru, in cloud forest, as Podocarpus forest in Ecuador, at (1200-)1400-2600 m.
Representative specimens examined. COLOMBIA. ANTIOQUIA: Mun. Medellin, Cgto. Santa Elena, 12 km SE of Medellin, 2200 m, 21 Jan 1994 (d), Callejas et al. 11110 (HUA, COL); Cordillera Central, between Palmitas & Alto (or Boquer6n de San Crist6bal), 2200-2250 m, 5 Oct 1961 (d), Cuatrecasas 26247 (COL, P, US), (9 fl-fr), Cuatre- casas 26248 (COL, P, US); El Retiro, Los Alpes, 15 Jan 1953 (st), Hno. Daniel 4492 (US); Mun. Medellfn, Santa Elena, old rd. to Rionegro, 1700 m, 24 Mar 1994 (d), Franco et al. 4579 (BG, HUA), (9 fl-fr), Franco et al. 4581 (BG, HUA); Yarumal, 2150 m, 28 Mar 1994 (d), Franco et al. 4602 (BG, HUA); Mun. Guarne, Parque Ecol6gico Pie-
SYSTEMATIC TREATMENT 181
dras Blancas, 2350 m, 9 Feb 1994 (J), Rolddn et al. 2161 (COL, HUA). BOYACA: Mun. Pajarito, between Corinto & Penia del Gallo, 1800 m, 29 Jan 1995 (d fl-fr), Franco et al. 4640 (BG, COL). CAQUETA: Mun. Florencia, below Gabi- nete, km 17, 2200 m, 15 Oct 1993 (6), Franco et al. 4494 (BG, COL), (cd), Franco et al. 4498 (BG). CASANARE: Mun. Sacama, Vrda. Guivarin, 1750 m, 27 Jan 1995 (Y fl- fr), Franco et al. 4639 (BG, COL, NY). CAUCA: Moscopan region, Rfo San Jose, Aguabonita, 2280 m, 30 Jan 1947 (Y fl-fr), Cuatrecasas 23543 (US, VALLE) & Cuatrecasas 23553 (VALLE). CESAR: Sierra de Perija, E of Manaure, San Antonio, 1700 m, 15 Nov 1959 (i), Cuatrecasas et al. 25346 (COL, US); Mun. Manaure, rd. Manaure-El Cinco, 1950 m, 14 Nov 1993 (Y fl-fr), Franco et al. 4520 (BG), (c), Franco et al. 4521 (BG). CUNDINAMARCA: Mun. Funza, Cerro de Rosales, ca. 2600 m, 26 Oct 1961 (6), Cuatrecasas et al. 26508 (COL, P, US), (Y fl), Cuatrecasas et al. 26510 (COL, P, US); Mun. Granada, 2225 m, 12 Mar 1993 (? fl-fr), Franco et al. 4342 (BG, COL); Mun. Tena, rd. Bogota-La Mesa, Vrda. Laguneta, 1900 m, 6 Feb 1995 (6), Franco et al. 4643 (BG, COL); rd. San Miguel-La Aguadita, 2-3 km below Quebrada Aguabonita, 2100 m, 12 Aug 1970 (9 fl-fr), Idrobo 6321 (COL); Mun. Pacho, 1975 (d), Mahecha 1764 (UDBC); Mun. Pasca, Vrda. El Pedre- gal, 2550-2600 m, 27 Jun 1987 (9 fl-fr), G. Morales et al. 1116 (COL). HUILA: Mun. La Plata, Vrda. Agua Bonita, Finca Merenberg, 1200-1300 m, 15 Jul 1975 (6), Dfaz P et al. 575 (COL); Finca Merenberg, 2300 m, 23 Dec 1975 (d), Gaulin 5 (COL); Finca Merenberg, Cauca border, E of Leticia, 2300 m, 8 Jul 1984 (9 fr), Gentry et al. 47757 (JAUM, MO); 30 km NW of Palermo, ca. 2700 m, 9 Oct 1944 (9 fr), Little 8780 (COL, US); Rfo Villalobos, nr. Rio Suazita, 1400 m, Jan 1943 (9 fl-fr), Schultes et al. 5172 (F, GH); confluence of Rio Villalobos & Rio Cachos, 1400 m, Jan 1943 (9 fl-fr), Schultes et al. 5205 (COL, F, GH). META: Parque Chingaza, rd. from San Juanito, 17 Sep 1997 (st), E. Acero D. 43 (COL); Cordillera Oriental, between Hda. El Pato & camp Los Cueros, ca. 2150 m, 12 Jan 1944 (9 fr), Little 8088 (COL, US). NORTE DE SANTANDER: Ocafia region, between La Maria & Jurisdicciones, 2200 m, 25 Sep 1969 (6), Cuatrecasas et al. 27940 (US); Ocaiia Region, nr. Apasica, Quebrada del Guarumal, 1900-2000 m, 26 Sep 1969 (9 fl-fr), Cuatrecasas et al. 27949 (COL, NY, P, US); Mun. Toledo, 1850 m, 24 Jan 1995 (9 fl), Franco et al. 4624 (BG, COL). QUINDIO: Mun. Pijao, Vrda. Car- cineros, Finca La Esperanza, 1920 m, 14 Aug 1992 (9 fl- fr), Velez et al. 2997 (COL). PUTUMAYO: 1.5 km E of Si- bundoy, 2200 m, 3 Oct 1963 (9 fl-fr), Bristol 1441 (COL, GH, US); rd. Mocoa-Pitalito, between Suacira & Santo Do- mingo, 2000 m, 5 May 1994 ( 9 fr), Franco etal. 5540 (BG); nr. Sibundoy, 2250 m, 18 Feb 1942 (9 fr), Schultes 3275 (F, GH). SANTANDER: Rio Surata, above Surata, 2000-2300 m, 5-6 Jan 1927 (st), Killip et al. 16667 (GH, NY, US). TOLIMA: Rio Claro, E of Nevada del Huila, ca. 2700 m, 30 Sep 1944 (d), Little 8747 (COL, F, US).
VENEZUELA. LARA: Huymucaro, rd. to Barbacoas, 1600 m, 1 Dec 1967 (st), R. E Smith V910 (VEN); Dtto. Jimenez, 10-15 km SSE of Sarare, 1750-1800 m, 7 Aug 1970 (6), Steyermark et al. 103564 (K, NY, U, US, VEN);
Dtto. Moran, Mun. Barbacoas, nr. Hato Arriba, 1870 m, 12 Aug 1983 (9 fl), Trujillo et al. 18553 (MO). MERIDA: Filo del Sai-Sai-El Carrizal, 1850 m, 24 Feb 1955 (9 fl-fr), Ber- nardi 2072 (NY); above Las Cuadras, Quebrada Molino, N of Torondoy, 1820-2255 m, 27 Mar 1944 (st, juv), Steyer- mark 55817a (F); Rio Capaz, above La Azulita, 2100-2400 m, 1 Sep 1966 (d), Steyermark et al. 97107 (NY, US, VEN). PORTUGUESA: Dtto. Sucre, La Divisoria de la Concepci6n, 1700 m, 24 Oct 1985 (st), Werif 7587 (BG, MO). TACHIRA: Dtto. Junin, between Rubio & Delicias, 2200 m, 19 Oct 1984 (st), Bono 4317 (MO); Rio Copas, between Las Copas & San Vincente de la Revancha, S of Alquitrana, SW of Santa Ana, 2000-2400 m, 22 Jan 1988 (9 fl-fr), Steyermark et al. 101275 (MO, NY, VEN); Quebrada Agua Azul, S of El Re- poso, 14 km SE of Delicias, 2150-2300 m, 22-23 Jul 1979 (9 fl-fr), Steyermark et al. 118640 (MO, U, VEN). ZULIA: Sierra de Perijd, between Misi6n de Los Angeles de Tukuku & Pishikakaro, 1400-1850 m, 1-3 Apr 1972 (9 fl-fr), Stey- ermark et al. 105784 (F, MO, NY, US, VEN).
ECUADOR. LOJA: San Jose de Yanacocha, ca. 1 km W of jct. of rd. Zamora-Loja & Rfo Jipiro, nr. border Loja- Zamora-Chinchipe, nr. Rio Jipiro, 2300 m, 5 Jan 1991 (d), Berg et al. 1665 (BG, GB, LOJA, QCA), ca. 2450 m, 5 Jan 1991 (9 fl-fr), Berg etal. 1666 (BG, LOJA, QCA); rd. Loja- Zamora, 7 km SE of Loja, 2300-2400 m, 3 Jul 1947 (9 fl- fr), R. Espinoza 1575 (NY). LOJA/ZAMORA-CHINCHIPE: Rd. Loja-Zamora, ca. km 14, 2500-2820 m, 29 Dec 1988 (9 fl-fr), Jorgensen et al. 65761 (AAU, BG), QCA). MORONA-SANTIAGO: Angel Sabio, between Lim6n & Gualaceo, ca. 2250 m, 28 Jan 1981 (9 fl-fr), Berg 1232 (BG, COL, QCA, U). ZAMORA-CHINCHIPE: 11 km E of Loja, 2600 m, 5 Sep 1975 (9 fl-fr), Little et al. 248 (COL, LOJA, Q, QAME, QCNE).
PERU. SAN MARTiN: Prov. Moyobamba, Rioja, Ven- ceremos, 1985 (9 fl), Lao M. 21 (K); Prov. Mariscal Ca- ceres, Rio Abiseo National Park, N-side of river across from La Playa base camp, 2600 m, 1 Sep 1985 (6), Young 1544 (F, MO).
The species is in cultivation in the Botanical Gar- den "Jose Celestino Mutis" in Bogota (9 Jun 1993 [d], Franco et al. 4461 [BG, COL]).
The type material of Cecropia teleincana and C. santandarensis, or more generally, the material col- lected in Antioquia and Santander (Colombia), differs (somewhat) from the other collections of the species in the relatively short stipules (to 25 cm), in the hir- sute rather than villous indumentum, and in the rela- tively long peduncle (to 15 cm long) and relatively short spikes of the pistillate inflorescence. Moreover, specimens with this hirsute indumentum (such as Cal- lejas et al. 11110 and Franco et al. 4579) appear to be rather often (?) inhabited by ants, in contrast to the type with villous indumentum as common in other parts of the species range, for which the presence of ants has never been recorded. The material from Ec- uador and Peru shows some differences in the length
182 FLORA NEOTROPICA
of the hairs on the leafy twigs and longer spikes in the staminate inflorescences. The variation found is not unusual in Cecropia species and does not justify recognition of infraspecific taxa. The longest spikes sometimes perforate the apex of the spathe of the pis- tillate inflorescences before anthesis.
Tree, to 4(-8) m tall, usually unbranched, the leafy part of the stem 3-5 cm thick, green, hispid to setose, sometimes also with sparse arachnoid indumentum. Lamina chartaceous to subcoriaceous, ca. (20 X 20 cm to) 30 X 30 cm to 80 X 80 cm, the segments 12- 15, oblanceolate, the incisions down to 1.5-5 cm from the petiole; apices acute to acuminate (to ob- tuse); upper surface ? scabrous, hispidulous to hir- tellous, initially also with sparse arachnoid indumen- tum; lower surface hirtellous on the veins, with arachnoid indumentum in the areoles, sometimes in- tially also on the main veins; lateral veins in the free part of the midsegment ca. 20-25 pairs, submargin- ally loop-connected, unbranched or some branched; petiole 40-75 cm long, hispidulous to subsetose, sometimes also with sparse arachnoid indumentum; trichilia fused, the brown indumentum intermixed with rather long white hairs; stipules 8-13 cm long, red-brown to grayish, hirtellous and with arachnoid indumentum outside, subsericeous inside. Staminate inflorescences mostly in pairs, the peduncle erect and the spikes pendulous; peduncle 4-10 cm long, hispi- dulous to subsetose; spathe 8-12 cm long, red-brown to grayish, hirtellous to subhispid outside, glabrous inside; spikes 2-5, often 4, 6-13 X 0.6 cm, with stipes ca. 0.5 cm long and only with brown pluricel- lular trichomes; rachis hairy. Staminate flowers: per- ianth tubular, 2-2.5 mm long, glabrous, the apex al- most plane, the aperture surrounded by a rim; filaments ? swollen; anthers 0.5-0.7 mm long, ap- pendiculate, detached at anthesis(?), reattached to the margins of the aperture by the appendages (?). Pistil- late inflorescences solitary or in pairs, erect; pedun- cle 7-10 cm long, hispidulous to subsetose; spathe 7- 9 cm long, the color and indumentum as in the sta-
minate inflorescence; spikes 1-3, often 2, 6-8 X ca. 0.7 cm, to 15 X 1.1 cm in fruit, sessile or with stipes to 0.3 cm long; rachis hairy. Pistillate flowers: peri- anth ca. 2.5 mm long, with arachnoid indumentum below the apex outside, also below the style channel inside, the apex convex, hispidulous; style rather long, slightly curved; stigma truncate and comose to sublingulate. Fruit oblongoid to ellipsoid, 3-3.5 mm long, ? tuberculate.
Distribution (Fig. 11.2). Central Amazonian Bra- zil, mostly in secondary growth.
Specimens examined. BRAZIL. AMAZONAS: Mun. Manaus, rd. BR. 174, km 45, Aug 1978 (Y fl), Benson 8465A (UEC, US), 7 Aug 1979 (Y fl), Benson 10341 (INPA, UEC); Manaus, INPA, 9 Sep 1973 (Y), Berg 275 (K, MO, U); Rio Cuieras, 15 Sep 1977 (d), Berg 277 (U), (Y fr), Berg 278 (F, U); Manaus-P6rto Velho Hwy., km 160, 23 Mar 1974 (Y fl-fr), Campbell et al. P20862 (INPA, U); Rio Negro, nr. Rio Arara, SIDERAMA, 2 May 1973 (Y fl-fr), Loureiro et al. (INPA) 37937 (INPA); Mun. Manaus, Distr. Agropecu- ario da SUFRAM, 27 km E of rd. BR. 174, km 64, Fazenda Esteio, 19 Mar 1981 ( fl-fr), Nascimento et al. 1103.371 (BG, INPA); Rio Madeira, Mun. Humaitd, rd. Humaita-La- brea, km 50, 29 Nov 1966 (d), Prance et al. 3460 (F, INPA, NY, U, US); Manaus-P6rto Velho Hwy. BR.319, km 268, 28 km S of Igap6 A,u, 17 Mar 1974 (? fl-fr), Prance et al. 20649 (INPA, K, MG, MO, NY, S, U, US). PARA: Santarem, Serra Diamantina, 14 Dec 1966 (? fr), Cavalcante et al. 1757 (MG); Faro, Castanhal da B6a Vista, 1 Feb 1910 (6), Ducke (MG) 10632 (MG); Serra do Humaitd, Rio Tapajoz, Boinu, 8 Apr 1924 (Y fl-fr), Kuhlmann 1941 (RB 19881); Rio Trombetas, Monte Branco, 6 Oct 1982 (6), Revilla et al. 6976 (INPA).
This species is distinguished by its predominantly monocaul habit and the small size of the trees. It shows some affinities to Cecropia silvae.
59. Cecropia utcubambana Cuatrecasas, Phytologia 52: 157. 1982; Berg & Franco Rosselli, Fl. Ec- uador 48: 51, t. 9. 1993. Type. Peru, Amazonas: Prov. Bongara, Rio Utcubamba, 4 km below Cam-
pamento Ingenio, 1250 m, 28 Jan 1964 (Y), Hutchinson & Wright 3844 (holotype: UC-n.v.; isotypes: G, K, LE, MICH, MO, NY, P, US).
Fig. 46
Cecropia puberula C. C. Berg & P. Franco, Novon 6: 248, t. 12. 1996. Type Peru. Ucayali: San Miguel de Semuya, S of Camp Verde (at km 33 on rd. Pucallpa- Lima), 12 Aug 1988 (d), Berg et al. 1639 (holotype: MOL; isotypes: AAU, BG, COL, MO, NY, U).
SYSTEMATIC TREATMENT 183
199'1
FIG. 46. Cecropia utcubambana. 1. Lamina, reduced (Berg et al. 1640). 2. Apex of lamina and venation (Berg et al. 1639). 3. Stipules and bases of petioles with trichilia (Berg et al. 1640). 4. Staminate inflorescences, one with spathe, the other at anthesis, and bases of petioles with trichilia. 5. Staminate flower. 6. Stamen after detachment of anther (Berg et al. 1639). 7. Pistillate inflorescence at anthesis and base of petiole with trichilium (Berg et al. 1640). 8. Pistillate flower. 9. Fruit (Berg et al. 1623). (From Novon 6: 247. 1996, modified.)
184 FLORA NEOTROPICA
Tree, to 15(-25) m tall. Leafy twigs 1.5-3.5(-5) cm thick, green (or partly bluish), puberulous to sub- hispidulous with curved to uncinate hairs. Lamina chartaceous to subcoriaceous, ca. (10 X 10 cm to) 25 x 25 cm to 60 X 60 cm, the segments 7-9(-1 1), the free part of upper segments oblong to subobovate or to obovate, the incisions down to 5/10-6(-7/1)0; api- ces obtuse to short-acuminate; upper surface scabrous to scabridulous (to almost smooth), puberulous to his- pidulous (to hirtellous); lower surface minutely pu- berulous, mostly also with longer uncinate to straight hairs on the veins, with arachnoid indumentum in the areoles or (almost) confined to the margin; lateral veins in the free part of the midsegment 9-16 pairs, submarginally loop-connected, some or most of them branched; petiole 15-50 cm long, green or red-brown, minutely puberulous; trichilia fused, the brown in- dumentum intermixed with sparse, (rather) short, white hairs; stipules 4-12 cm long, red-brown to brown, dark red to purplish, or greenish, minutely pu- berulous or on the ribs sparsely hirtellous outside, sparsely to densely sericeous with white (or brown- ish) hairs inside. Staminate inflorescences in pairs, pendulous; peduncle (6-)10-21 cm long, puberulous and with very sparse arachnoid hairs; spathe 7-26 (-32) cm long, yellowish brown to greenish, dark red to orange, or grayish, puberulous to sparsely strigil- lose to subhirtellous or also with sparse to dense arachnoid indumentum outside, (almost) glabrous in- side; spikes 4-7(-15), (4-)10-22(-3 1) X 0.2-0.5 (-0.8) cm, sessile (or with stipes 0.8-1.8 cm long and hairy); rachis glabrous or sparsely hairy. Staminate flowers: perianth tubular, 0.8-1.5 mm long, with short hairs just below or also on the margin of the apex, the apex slightly convex to plane; filaments flat; anthers 0.4-0.7 mm long, not appendiculate, detached and re- maining attached by 2 filiform connections between the connective and the upper margin of the filament. Pistillate inflorescences in pairs, pendulous; peduncle 15-50 cm long, puberulous to subhispidulous; spathe 9-20 cm long, the color and indumentum as in the staminate inflorescence; spikes 4-5(-6), 10-20 X 0.8-1 cm, to 35 X 1.2 cm in fruit, (sub)sessile; rachis hairy to subglabrous. Pistillate flowers: perianth ca. 2 mm long, with arachnoid indumentum below the apex outside, also below the style channel inside, the apex sparsely to densely muriculate; style rather short; stigma penicillate. Fruit ellipsoid, 2-2.5 mm long, smooth, the upper and lower part dark brown, the middle part pale brown.
Distribution (Fig. 11.3). Southern Ecuador to southern Peru (Madre de Dios), in forest (margins) and secondary growth, at elevations to 2000 m.
Representative specimens examined. ECUADOR. MORONA-SANTIAGO: Between Plan del Milagro & Lim6n (Gral. Plaza), 1500 m, 30 Jan 1981 (i), Berg 1235 (AAU, BG, COL, MO, QCA, U). ZAMORA-CHINCHIPE: Rd. Loja- Zamora, between Sabanilla & Zamora, ca. 1250 m, 3 Jan 1991 (? fl-fr), Berg et al. 1654 (BG, LOJA, QCA); rd. Val- ladolid-Rio Palanda, ca. km 6-8, ca. 1500 m, 31 Jan 1985 (Y fl-fr), Harling et al. 21363 (BG, GB, QCA); El Padmi, 5 Jun 1995 (Y fl-fr), Merino et al. 4514 (BG, LOJA).
PERU. AMAZONAS: Prov. Bongara, Dtto. Sipabamba, above Quebrada Fortuna, on trail to Subida Alba, ca. 1350 m, 5 May 1981 (st), Young et al. 341 (BG, NY). Cuzco: Prov. Paucartambo, between Pilcopata & Atalaya, 29 Jul 1988 (Y), Berg et al. 1604 (BG, COL, Centro de Medicina Andina, Cuzco); Prov. Paucartambo, between San Jorge & Salazar, Rio Tono, ca. 600 m, 1 Aug 1988 (Y), Berg et al. 1623 (BG, Centro de Medicina Andina, Cuzco). HUANUCO: Rd. Huhnuco-Tingo Maria, nr. Tambillo Grande (km 515), 24 Nov 1997 (Y fl-fr), Berg et al. 1741 (BG, COL, MOL); nr. Tingo Maria, Cueva de las Lechuzas, 25 Nov 1997 (c), Berg et al. 1744 (BG, COL, MOL); Prov. Puerto Inca, Dtto. Yuyapichis, DANTAS, 18 Jul 1989 (Y fl-fr), Kroell S. 508 (BG, G); Prov. Pachitea, ca. 26 km S of Puerto Inca, 21 Sep 1988 (? fr), Morawetz et al. 15-21988 (BG); Prov. Leoncio Prado, Dtto. Rupa Rupa, E of Tingo Maria, Cerro Quemado, 10 Sep 1978 (9 fl-fr), Schunke V 10593 (BG); Prov. Puerto Inca, Dtto. Yuyapichis, DANTAS, 1-15 Oct 1990 (9 fl-fr), Tello 242 (G), 16-30 Jun 1991 (i), Tello 2639 (G). LOR- ETO: Purancanchim, Rio Sinchiuyacu, 22-27 Nov 1986 (9 fr), Lewis et al. 12181 (U). MADRE DE Dios: Prov. Tam- bopata, Cuzco Amaz6nico, 13 Dec 1989 (st), Gentry et al. 68643 (BG, MO), 14 Jul 1991 (9 fl-fr), Fisher 210, 211 (BG), 14 Jun 1989 (9 fl), Niiuez et al. 10727 (BG). PANDO: Prov. Manupiri, Conquista, 23 Aug 2000 (st), Balcdzar 2177 (USZ). PASCO: Rd. Oxapampa-Llaupi-Cerro de Pasco, ca. 17 km, nr. Maria Teresa, ca. 1650 m, 29 Nov 1997 (d), Berg et al. 1760 (BG, COL, MOL); Prov. Oxapampa, Panjil, 12 km from Puerto Inca, 27 Sep 1982 (9 fl-fr), D. N. Smith et al. 2422 (BG). SAN MARTiN: Rio Huallaga, Cerros de Es- toraqui, W of Shapaja, trail to Tarapoto, 2-8 km, 4-7 Aug 1937 (9 fl-fr), Belshaw 3224 (GH, MICH, NY, US); Prov. San Martfn, rd. Tarapoto-Yurimaguas, ca. km 27, 5 Dec 1997 (9 fr), Berg et al. 1777 (BG, COL, MOL), ca. km 39, 5 Dec 1997 (6), Berg et al. 1780 (BG, COL, MOL); Prov. San Martin, rd. Tarapoto-Chazuta, km 24, 6 Dec 1997 (c), Berg et al. 1783 (BG); Prov. Lamas, rd. Tarapoto-Moyob- amba, ca. km 60, 8 Dec 1997 (9 fr), Berg et al. 1787 (BG); Prov. Rioja, rd. Rioja-Pomacocha, ca. 50 km, 8 Dec 1997 (9 fl-fr), Berg et al. 1795 (BG, COL, MOL). UCAYALI: San Miguel de Semuya, S of Campo Verde (at km 33 on rd. Pucallpa-Lima), 12 Aug 1988 (9 fl-fr), Berg et al. 1640 (BG, COL, K, MO, MOL); rd. Pucallpa-Tingo Maria, ca. km 149, 26 Nov 1997 (d), Berg et al. 1751 (BG, COL, MOL); rd. Pucallpa-Tingo Maria, ca. km 179, nr. Boquer6n, 26 Nov 1997 (9 fl-fr), Berg et al. 1752 (BG, COL, MOL).
Cecropia utcubambana and C. puberula, consid-
ered to be morphologically, ecologically, and geo- graphically distinct (Berg & Franco Rosselli, 1996),
SYSTEMATIC TREATMENT 185
had to be merged as recent collecting reduced the dif- ferences considerably. In the northern part of the dis- tribution range, the species is submontane and the arachnoid indumentum is confined to the margin of the lamina. In the southern part of the range, the spe- cies is a lowland element, and it often has arachnoid indumentum in the areoles of the lamina beneath. The species has a green morph and a red one (with red to red-brown stipules, spathes, petioles, and peduncles). This species shows strong affinities to C. latiloba. It differs from the latter in the indumentum of the stip- ules, the indumentum on the veins at the lower surface of the lamina, and the lateral veins being submargin- ally loop-connected instead of marginally. It is clearly distinct from C. latiloba in its ecology, as it is a true upland species. In its habit, the species resembles C. putumayonis, as briefly discussed under the latter. The pistillate inflorescences with long peduncles (and spikes) of one of the Ruiz & Pavon s.n., on sheets at B and MA, indicated as C. peltata and in G without name, might belong to C. puberula. Matching leaves have not been found among the Ruiz & Pavon col- lections.
Vernacular name. Peru: kabeari (Machiguenga, Madre de Dios).
60. Cecropia velutinella Diels, Notizbl. Bot. Gart. Berlin-Dalhem 15: 367. 1941. Type. Ecuador. Pas- taza: Mera, ca. 1100 m, 2 Dec 1938 ( Y ), Schultze- Rhonhof 3068 (holotype: B, destroyed, duplicates not traced), herewith replaced by: Ecuador. Morona-Santiago: Rd. Gualaquiza-Don Bosco, km 19, ca. 1500 m, 16 Feb 1994 (Y), Berg et al. 1695 (neotype: QCNE; isoneotype: BG). Fig. 47
Tree, to 25 m tall. Leafy twigs 5-13 cm thick, dark brown to black, white-hirtellous and with dark brown to black pluricellular hairs. Lamina subcoriaceous, ca. 50 X 50 cm to 130 X 130 cm, the segments 14-20 (-24), lanceolate to linear, the incisions down to the petiole; apices acute; upper surface smooth or scabri- dulous, minutely puberulous and initially with sparse brown pluricellular hairs; lower surface with very dense arachnoid indumentum, on the main veins (in- itially) also with brown pluricellular hairs; lateral veins in the midsegment 50-80 pairs, submarginally loop-connected, mostly unbranched; petiole 30-100 cm long, with dense arachnoid indumentum and with brown pluricellular hairs, the upper part (pale brown) hirsute to subvillous; trichilia fused, the dark brown indumentum intermixed with short grayish hairs; stip- ules 25-60 cm long, white to brownish, with dense
arachnoid indumentum, intermixed with brown plur- icellular hairs outside, dark red, with very short hairs on the ribs or glabrous inside. Staminate inflores- cences in pairs, the peduncle erect and the spikes pendulous (?); peduncle 7-14 cm long, brown- puberulous to-subhispidulous, toward the apex hir- tellous to setulose, also with arachnoid indumentum and dense brown pluricellular hairs; spathe not seen; spikes ca. 8-10, 12-20 X ca. 0.3 cm, with stipes 0.7-1 cm long and sparsely hirtellous and with dense arach- noid indumentum; rachis glabrous. Staminate flow- ers: perianth tubular, 1-1.5 mm long, with dense arachnoid indumentum below the apex, the apex mur- iculate; filaments flat; anthers ca. 0.6 mm long, at an- thesis detached, reattached to the margins of the ap- erture by the appendages (?). Pistillate inflorescences solitary, erect to deflexed; peduncle 7-14 cm long, dark green, with sparse arachnoid indumentum and toward the base and apex brownish-subhispidulous; spathes not seen; spikes 4-5, ca. 25-45 X 1-1.5 cm in fruit, + curved; rachis subglabrous. Pistillate flow- ers: perianth ca. 1.5-2 mm long, with arachnoid in- dumentum below the apex outside, also the lower part of the style channel inside, the apex almost plane, muriculate; style rather long, straight to slightly curved, muriculate; stigma comose. Fruit ellipsoid to subovoid, ca. 2 mm long, slightly tuberculate, dark brown.
Distribution (Fig. 18.4). Ecuador, from Pastaza to Morona-Santiago, and in Peru (San Martin), in (sub)montane forest, at 1200-1700 m.
Specimens examined. ECUADOR. MORONA-
SANTIAGO: Rd. Lim6n (Gral. Plaza)-Cuenca, ca. km 15, Plan del Milagro, ca. 1700 m, 30 Jan 1981 (juv), Berg 1234 (AAU, BG, GB, MO, QCA, U); rd. Gualaquiza-Don Bosco, km 19, ca. 1500 m, 16 Feb 1994 (juv), Berg et al. 1695A (BG, COL); Cord. de Cutucu, 5-10 km from Logronio, 1200-1500 m, 7-9 Oct 1975 (d), Little et al. 651 (COL, LOJA, QAME, QCNE).
PERU. SAN MARTiN: Prov. Rioja, rd. Rioja-Pomaco- cha, km 402. ca. 1350 m, 9 Dec 1997 (Y fr), Berg et al. 1804 (BG, COL, MOL).
This spectacular submontane species can be easily recognized by the numerous narrow leaf segments with more lateral veins than in any other Cecropia species. Mullerian bodies are abundantly produced, but the trees are probably rarely inhabited by ants. The absence of ants may be related to the fact that the stems of young trees are covered by densely brown- hirsute indumentum and (sub)persistent stipules cov- ered with very dense and matted, white-arachnoid in- dumentum mixed with filiform brown-pluricellular hairs.
186 FLORA NEOTROPICA
i:
FIG. 47. Cecropia velutinella. 1. Lamina, reduced. 2. Apex of lamina and venation (Berg 1234). 3. Stipules and young leaf. 4. "Umbilicus" of lamina. 5. Interior of leafy twig (Berg et al. J695A). 6. Pistillate inflorescence after anthesis. 7. Pistillate flower. 8. Fruit with remnant of style (Berg et al. 1695).
SYSTEMATIC TREATMENT 187
Vernacular names. Ecuador: su (Shuar, Morona- Santiago); guarumo blanco (Pastaza).
61. Cecropia virgusa Cuatrecasas, Revista Acad. Colomb. Ci. Exact. 6(22/23): 286. 1945; 7(27): t. 5.2. 1947. Type. Colombia. Valle: Between El Aguacate & Quebrada de La Yuca, 8 Feb 1944 (d), Cuatrecasas 16093 (holotype: COL; isotype: F).
Tree, to 15 m tall. Leafy twigs 2-4 cm thick, green to purplish brown with conspicuous lenticels, sparsely puberulous to hispidulous. Lamina subcoriaceous, ca. 30 X 30 cm to 80 X 80 cm, the segments (6-)7-8, the free parts of upper segments ovate, the incisions down to 3/10-6/10; apices subacuminate to rounded; upper surface almost smooth to scabridulous, sparsely hispidulous, on the main veins and the "umbilicus" sparsely hirtellous to subhirsute; lower surface sparsely puberulous with brown pluricellular hairs on the veins, with arachnoid indumentum in the areoles and extending to the main veins; lateral veins in the free part of the midsegment ca. 10-15 pairs, submar- ginally loop-connected, the lower ones usually branched; petiole 15-40 cm long, sparsely puberulous to subhispidulous and with rather dense arachnoid in- dumentum; trichilia separate, sunken into 2 pockets with slit-shaped apertures; stipules (7-)10-26 cm long, reddish to brownish or to purplish, ? curved, sparsely hirtellous and with dense arachnoid indu- mentum and with brown pluricellular trichomes out- side, densely pale yellow-sericeous inside. Staminate inflorescences in pairs, erect or + deflexed; peduncle 7-11 cm long, hispidulous or also with sparse arach- noid hairs; spathe 7-12 cm long, reddish to brownish or to purplish, sparsely hirtellous to strigose and with dense arachnoid indumentum and brown pluricellular trichomes outside, densely yellowish-sericeous in- side; spikes 2-4, 4-13 X 0.3-0.7 cm, sessile; rachis hairy. Staminateflowers: perianth tubular, 1-1.5 mm long, with stiff hairs on the ribs below the apex, the apex plane to slightly convex, glabrous or muriculate; anthers ca. 0.4-0.5 mm long, appendiculate, detached at anthesis (?). Pistillate inflorescences in pairs, pen- dulous (?), subtended by caducous bracts, 0.5-5 cm long; peduncle 5.5-15 cm long, hispidulous or also with sparse arachnoid hairs; spathe 6-8 cm long, the color and indumentum as in the staminate inflores- cence; spikes 2-4, 3.5-6 X 0.4-0.5 cm, to 11 X 1.5 cm in fruit, sessile; rachis hairy with straight hairs or also arachnoid indumentum. Pistillate flowers: peri- anth 2-2.5 mm long, with arachnoid indumentum be- low the apex outside, absent inside, the apex slightly
convex, muriculate, the aperture wide and round, sur- rounded by a rim; style rather short the apex slightly convex, muriculate, with a wide round aperture sur- rounded by a rim; stigma comose. Fruit ellipsoid to ovoid, ca. 2 mm long, tuberculate.
Distribution (Fig. 8.1). From northwestern Ec- uador to eastern Panama, in wet forest or secondary growth, at low elevations.
Specimens examined. PANAMA: DARIEN: Cruce de Mono, Parque Nacional Darien, 5 Nov 1989 (st), Fisher 55 (BG).
COLOMBIA. CAUCA: Rio Dagua, Juntas, 21 Sep 1882 (i), Lehmann 1969 (G). CHOC6: Ca. 20 km SE of Quibd6, 8 Jul 1986 (Y fl), Berg 1542 (BG, COL); nr. Tutunendo, ca. 25 km ENE of Quibd6, 9 Jul 1986 (i), Berg 1545 (BG, COL); Serrania de Baud6, rd. Las Animas-Pato, Rio Pato, 18 Apr 1983 (6), Croat 56100 (BG, JAUM, MO); between Quibd6 & Tutunendo, 4 Apr 1958 (Y fl-fr), Cuatrecasas et al. 24221 (COL, US); rd. Quibd6-Yuto, nr. Yuto, 12 Apr 1990 (Y fl), Franco et al. 3057 (COL); rd. Yuto-Llor6, 18 Jan 1979 (f), Gentry et al 24376 (BG, COL, MO); between Andagoya & Condoto, 9 Sep 1949 (Y fl-fr), Idrobo 1949 (COL); Bahia Solano, Quebrada Jellita, 22 Feb 1939 (st), Killip et al. 33551 (US); Novita, Mar 1853 (st), Triana 1865 (K). NARINO: Mun. Barbacoas, nr. Guayacana, 12 Dec 1993 (d), Franco et al. 5180 (BG, COL); rd. Junin-Barbacoas, 7 Jan 1989 (Y fl), Gentry et al. 64565 (PSO); Mun. Tumaco, Resguardo Indigena Alto Albi, 30 May 1992 (st), M. S. Gon- za'lez 115 (COL); La Guayacana, 3 Jun 1986 (6), Leon et al. 1245 (U). VALLE: Rio Calima, La Trojita, 19 Feb-10 Mar 1944 (Y fl), Cuatrecasas 16716 (MO, VALLE); Rio Cajambre, Barco, 21-30 Apr 1944 (6), Cuatrecasas 17266 (F, US, VALLE); Rio Digua, C6rdoba, 14 Nov 1945 (Y fl), Cuatrecasas 19859 (F, US); RIo Calima, between Qubradas de Aguaclara & La Brea, 21 May 1946 (Y fl-fr), Cuatre- casas 21170 (F, US, VALLE); Rio Calima, nr. La Brea, 25 May 1946 (st), Cuatrecasas 21314 (F, US); Rio Calima, Quebrada de L6pez, 23 Sep 1961 (Y fl-fr), Cuatrecasas et al. 26043 (COL, P, US); 16 km NW of Buenaventura, 25 Apr 1987 (st), Faber-Langendoen et al. 256 (MO); nr. Bue- naventura, 15 Mar 1994 (? fl-fr), Franco et al. 4534 (COL, TULV); Rio Calima, ca. 6 km N of Buenaventura, 13 Dec 1981 (Y fl), Gentry 35641 (BG, COL, JAUM, MO, TULV); Bajo Calima, Canalete, 15 Jun 1968 (Y fl), Gentry et al. 62936 (BG); Rio Dagua, 10 May 1922 (juv), Killip 5352 (NY, US); Mun. Buanaventura, 1970 (st), Mahecha s.n. (UDBC 11162); Bajo Calima, 1970 (Y fl-fr), Mahecha s.n. (UDBC 11673); Bajo Calima, Concesi6n Pulpapel/Buena- ventura, 2 Jun 1987 (9 fl), Monsalve B. 1501 (BG, MO); rd. San Isidro-Juanchaco, 15 Nov-6 Dec 1979 (9 fl-fr), Rooden et al. 515 (BG, COL, U).
ECUADOR. ESMERALDAS: Between Mataje & Molina, 5 Sep 1991 (9 fl), Jaramillo et al. 13742 (COL, GB, NY, QCA); Reserva Ecol6gica Cotacachi-Cayapas, Luis Vargas Torres, Rio Santiago, 8-14 Dec 1993 (9 fl-fr), Tirado et al. 735 (QCNE); Reserva Ecol6gica Cotacachi-Cayapas, Rio
Cecropia virgusa has an exceptional type of tri- chilium. The two trichilia are hidden in lateral (de- pressions) pockets with a slit-shaped opening at the base of the petiole. The pockets develop as grooves departing from the base of the petiole and become longer and deeper; this occurs before trichilia are formed. Tannin is not found in the developing peri- carps, as usual in the genus. Ants are usually present.
Vernacular names. Colombia: llarumo (Choco); cosedera teu, tud teu (Awapit, Nariiio); virgusa (Valle).
EXCLUDED NAME Cecropia scabra Martius, Flora 24(Beibl. 2): 95.
1841. Type. Brazil. Para': Rio Amazonas, Martius 630(2) (M), a mixed collection consisting of a leaf of Pourouma, almost certainly a juvenile one of P. guianensis Aublet subsp. guianensis, and a pistil- late inflorescence, possibly of Cecropia ulei. The leaf is here designated as the lectotype.
Cecropiajavitensis Pittier, in Pittier, Luces de Febres, Badillo & Lasser, Cat. Fl. Venez. 1: 258. 1945.
Cecropia nigra Pittier, in Pittier, Luces de Febres, Badillo & Lasser, Cat. Fl. Venez. 1: 258. 1945.
Cecropia solanoensis Pittier, in Pittier, Luces de Fe- bres, Badillo & Lasser, Cat. Fl. Venez. 1: 258. 1945.
Cecropia paludosa Warburg ex Glaziou, Bull. Soc. Bot. France 59(Mem. 3): 645. 1913.
Cecropia stenostachya Warburg, in Karsten & Schenck, Vegetationsbilder 4(1): t. 1-2. 1906.
ACKNOWLEDGMENTS This study has been supported by The Research
Council of Norway (NFR) through its program Con- servation of Biological Diversity, The Norwegian Council for Science and Humanities (NAVF), and The Meltzer Foundation of the University of Bergen, mak- ing (joint) fieldwork and visits to herbaria possible. The Olaf Grolle Olsen Foundation of the University of Bergen financed most of the illustrations prepared for the present contribution. Studies by the second author have been supported by the University of Ber- gen for a stay of some months (1987) to study Ce- cropia, to Conciencias (Colombia) to carry out further research in Bergen and Utrecht (Aug-Dec 1994), Red Latinoamericana de Botainica (RLB) for a stay at Universidad de la Plata (Argentina), with special rec- ognition to J. V. Crisci and J. J. Morone. Universidad Nacional de Colombia, particulary Instituto de Cien- cias Naturales (ICN) supported and provided facili- ties. Students of Universidad Nacional de Colombia (Bogota' and Medellin) assisted in fieldwork. D. W. Davidson is thanked for her willingness to write a chapter on the Cecropia-ant relations and for her comments on the introductory part of this contribu- tion, and D. W. Yu (Harvard University) for infor- mation about names and use of species by the Ma- chiguenga people in Madre de Dios (Peru). The University of Bergen and the Norwegian Arboretum (at Milde) are acknowledged for funding and facilities provided to work together during an early and a later phase of the present study. Fieldwork was essential for the preparation of the present monograph. It started in 1973 and finished in 1998 and in that period numerous colleagues and institutions have provided logistical and administrative support to one or both of us, which is gratefully acknowledged. Some of them have to be named: C. D. Adams (then at Port of Spain), S. G. Beck (La Paz), F. Borchsenius (then at Quito), R. Callejas (Medellin), J. P. P. Carauta (Rio de Janeiro), W. Devia (Tulua'), R. L. Dressler (then in Panama), G. Hatschbach (Curitiba), L. Holm-Nielsen (then at Quito), J. Jaramillo (Quito), B. Klitgaard
LITERATURE CITED 189
(then at Loja), S. Lamotte (then at Iquitos), L. Leoni (Carangola), P. Lozano (Loja), B. Merino (Loja), G. T. Prance (then at Manaus), Luzmila Quifiones (Vi- Ilavicencio), W. Steiner (then at Sapecho), J. Rios T. (Lima), Centro de Medicina Andina (Cuzco), Her- bario Forestal Nacional (Cochabamba), Museu Goeldi (Belem), and Universidad Nacional Agraria de la Selva (Tingo Marla). The directors and curators of the following herbaria allowed examination of the collections or, in a few cases, sent material on loan: A, AAU, B, BG, BHCB, BM, BOLV, C, CAY, COL, CRVD, CUVC, EAP, F, G, GB, GFJP, GH, GUA, HUA, IAN, IJ, INPA, JAUM, K, L, LE, LL, LOJA, LPB, MEDEL, MEXU, MG, MICH, MOL, MY,
OXF, P, PMA, PSO, Q, QAME, QCA, QCNE, RB, S, SCZ, SP, TEX, TULV, TUR, U, UDBC, UPRRP, US, USM, USZ, VALLE, VEN, and WIS. The distri- bution maps have for the greater part been prepared by Alvaro Neira (Bogota') and the illustrations have been made by Hendrieke Berg (Voss). The abstract has been translated into Spanish by Alvaro Neira (Bogota) and into Portuguese by J. P. P. Carauta (Rio de Janeiro). The reviewers, W. C. Burger (Field Mu- seum, Chicago), J. Kallunki (The New York Botanical Garden), S. Mori (The New York Botanical Garden), and an anonymous one, are owed thanks for their comments leading to the improvement of the present contribution.
LITERATURE CITED
Literature cited by Davidson is marked with an asterisk (*), that cited by both by Berg and Franco Rosselli and by Davidson are marked with two aster- isks (**). Adams, E. S. 1990a. Boundary disputes in the territorial
ant Azteca trigona: effects of asymmetries in colony size. Anim. Behav. 39: 31-328.*
. 1990b. Interaction between the ants Zacrypto- cerus maculatus and Azteca trigona: interspecific parasitism of information. Biotropica 22: 200- 206.*
. 1994. Territory defense by the ant Azteca tri- gona: maintenance of an arboreal ant mosaic. Oec- ologia 97: 202-208.*
Adanson, M. 1763. Familles des plantes 2. Vincent, Paris.
Agrawal, A. 1998. Leaf damage and associated cues in- duced aggressive ant recruitment in a neotropical ant- plant. Ecology 79: 2100-2112.
Akkermans, R. W. A. P. & C. C. Berg. 1982. New spe- cies and combinations in Coussapoa (Cecropiaceae). Proc. Kon. Ned. Akad. Wetensch., Ser. C, 85: 441- 471.
Allemao, F. F. 1860. Algunas consideraoes, e alguns factos novos, concernentes a estructura da flor, e fruto de Embauba- Cecropia peltata. Revista Braz. 3: 8- 11.
Allen, P. H. 1956. The rain forests of Golfo Dulce. Uni- versity of Florida Press, Gainesville.
Alvarez-Buylla, E. R. & M. Martinez-Ramos. 1990. Seed bank versus seed rain in the regeneration of a tropical pioneer tree. Oecologia 84: 314-325.
& . 1992. Demography and allometry of Cecropia obtusifolia, a neotropical pioneer tree-an evaluation of the climax-pioneer paradigm for tropi- cal forests. J. Ecol. 80: 275-290.
Andrade, J. Cardoso de. 1980. Necta6rios em Cecropia lyratiloba Miq. Resumos da 32a. Reuniao Anual da SBPC, Rio de Janeiro.
. 1981. Biologia da Cecropia lyratiloba Miq. var. nana Andr. & Car. (Moraceae) na Restinga do Re- creio dos Bandeirantes. Thesis, Universidade Federal do Rio de Janeiro.**
. 1982. Corpusculos de Muller: uma possivel defensa das Embauibas mirmecofitas contra preda,c de suas flores. Atas Soc. Biol. Rio de Janeiro 23: 1-2.
. 1984a. Observa,oes preliminares sobre a ecoe- tologia de quatro cole6pteros (Chrysomelidae, Te- nebrionidae, Curculionidae) que dependem da Embauba (Cecropia lyratiloba var. nana-Cecropi- aceae); na Restinga do Recreio dos Bandeirantes, Rio de Janeiro. Revista Bras. Entomol. 28: 99-108.**
. 1984b. Desempenho e distribuicao de Azteca alfari (Formicidae) em Cecropia lyratiloba var. nana (Cecropiaceae) na Restinga do Recreio dos Bandeir- antes, Rio de Janeiro- RJ. Atas Soc. Biol. Rio de Janeiro 24: 11-15.
. 1984c. Nectar em Cecropia lyratiloba var. nana Andr. & Car. (Cecropiaceae). Rodriguesia 36(58): 81-84.
& J. P. P. Carauta. 1979. Associac,ao Cecropia- Azteca na restinga do Recreio dos Bandeirantes, RJ. Bradea 3(5): 31-33.
& . 1981. Cecropia lyratiloba var. nana, nova variedad. Bradea 3(22): 163-168.
& . 1982. The Cecropia-Azteca associ- ation: a case of mutualism? Biotropica 14: 15.**
Ayala, E, J. Wetterer, J. Longino & D. Hartl. 1996. Molecular phylogeny of Azteca ants (Hymenoptera: Formicidae) and the colonization of Cecropia trees. Molec. Phylogenet. Evol. 5: 423-428.*
190 FLORA NEOTROPICA
Bailey, I. W. 1922. Notes on neotropical ant-plants I. Cecropia angulata sp. nov. Bot. Gaz. (Crawfords- ville) 74: 369-391.**
Baird, J. N. 1967. Observation on the Azteca-Cecropia interaction (Hym. Formicidae-Moraceae). Thesis, University of Kansas.**
Barrere, P. 1741. Essai sur l'histoire naturelle de la France equinoxiale. Paris.
Barth, 0. M. 1974. 0 p6len de algumas especies de Ce- cropia (Moraceae). Leandria 3/4(no. 4/5): 115-117.
Beattie, A. J. 1985. The evolutionary ecology of ant- plant mutualisms. Cambridge University Press, Cam- bridge.*
Becerra, J. & D. Venable. 1989. Extrafloral nectaries: a defense against ant-Homoptera mutualisms? Oikos 55: 276-279.*
Belin-Depoux, M., P. Solano, C. Lubrano, J. Robin, P. Chouteau & M. Touzet. 1997. Myrmecophilous function of Cecropia obtusa Trecul (Cecropiaceae) in French Guiana. Acta Bot. Gallica 144: 289-313.*
Benson, W. W. 1985. Amazon ant-plants. Pp. 239-266. In G. T. Prance & T. E. Lovejoy, Key environments. Amazonia. Pergamon Press, Oxford.**
Berg, C. C. 1972. A new species of Cecropia (Mora- ceae). Acta Bot. Neerl. 21: 655-656.
- . 1977a. Cecropia purpurascens, a new species for Brazilian Amazonia. Acta Amazonica 7: 185- 188. - . 1977b. Abscission of anthers in Cecropia Loefl. Brief communication. Acta Bot. Neerl. 26: 417-419.
. 1978a. Cecropiaceae a new family of the Urti- cales. Taxon 27: 39-44.
* 1978b. Especies de Cecropia da Amaz6nia Brasileira. Acta Amaz6nica 8: 149-182.
. 1980. Moraceae. Pp. 111-120 in A. L. Stoffers (ed.), Flora of the Netherlands Antilles. Vol. 2. Utrecht.
-. 1981. An exceptional new species of Cecropia (Moraceae) from Ecuador. Nord. J. Bot. 1: 485-487.
. 1985. Studies on the Flora of the Guianas 15: a new species of Cecropia (Cecropiaceae) from French Guiana. Bull. Mus. Natl. Hist. Nat. Paris, Ser. 4, 7(sect. B, Adansonia, no. 3): 255-258.
- - . 1989. Systematics of Urticales. Pp. 193-220 in P. R. Crane & S. Blackmore (eds.), Evolution, sys- tematics, and fossil history of the Hamamelidae 2, 'Higher' Hamamelidae. Clarendon Press, Oxford.
* . 1990. Differentiation of flowers and inflores- cences of Urticales in relation to their protection against breeding insects and to pollination. Sommer- feltia 11: 13-34.
. 1992. Cecropiaceae. Pp. 93-124 in A. R. A. Gorts-van Rijn, Flora of the Guianas, Ser. A, Fasc. 11. Koeltz Scientific, Koenigstein.
. 1998. Cecropiaceae. Pp. 174-190 in P. E. Berry, B. K. Holst & K. Yatskievych, Flora of the Venezue- lan Guayana 4. Missouri Botanical Garden, St. Louis.
. 2000. Cecropiaceae. Pp. 191-249 in Flora de Venezuela. Fundaci6n Instituto Botanica de Vene- zuela "Dr. Tobias Lasser," Caracas.
. 2001. A new species of Cecropia (Cecropi- aceae) from Colombia. Caldasia 23: 77-79.
- . 2002. An account on the Cecropia species (Ce- cropiaceae) of Peru. Caldasia 24: 229-238.
& J. P. P. Carauta. 1986. Cecropia (Cecropi- aceae) no Brasil, ao sul da Bacia Amaz6nica. Alber- toa 1(1): 3-17; reprint Albertoa 4(16): 213-221.
& P. Franco Rosselli. 1993. Cecropiaceae. P. 48 in G. Harling & L. Andersson, Fl. Ecuador. Berlings, Arlov.
& - . 1996. New taxa and combinations in Moraceae and Cecropiaceae from Central and South America. Novon 6: 230-252.
, R. W. A. P. Akkermans & E. H. van Heusden. 1990. Cecropiaceae: Coussapoa and Pourouma, with an introduction to the family. Fl. Neotrop. Monogr. 7. New York Botanical Garden, Bronx.
Bertoloni, A. 1840. Fl. Guatimal. Emygdio ab Ulmo, Bologna.
Bonsen, K. & B. J. H. ter Welle. 1983. Comparative wood and leaf anatomy of the Cecropiaceae (Urti- cales). Bull. Natl. Hist. Nat. Paris, Ser. 4, 5 (Sect. B, Adansonia, no. 2): 151-177.
Brandao, C. R. F. 1991. Adendos ao catalogo abreviado das formigas da regiao neotropical (Hymenoptera: Formicidae). Revista Bras. Ent. 35: 319X412.*
Brokaw, N. V. L. 1985. Gap-phase regeneration in a tropical forest. Ecology 66: 682-687.
.1986. Seed dispersal, gap colonization, and the case of Cecropia insignis. Pp. 323-331 in A. Estrada & T. H. Fleming (eds.), Frugivores and seed disper- sal. W. Junk, Dordrecht.
. 1987. Gap-phase regeneration of pioneer tree species in tropical forest. J. Ecol. 75: 9-19.
. 1998. Cecropia schreberiana in the Luquillo Mountains of Puerto Rico. Bot. Rev. (Lancaster) 64: 91-120.
Brooks, D. 1979. Testing hypotheses of evolutionary re- lationships among parasites: the digeneans of croc- odiles. Amer. Zool. 19: 1225-1238.*
Brown, W. L. 1973. A comparison of the Hylean and Congo-West African rain forest ant fauna. Pp. 161- 185 in B. Meggers, E. Ayensu & W. Duckworth (eds.), Tropical forest ecosystems in Africa and South America, a comparative review. Smithsonian Institute Press, Washington, DC.*
Browne, P. 1756. Civil and natural history of Jamaica. London.
Burger, W. C. 1977. Flora Costaricensis: Moraceae. Fieldiana, Bot. 40: 95-215.
Burret, K. E. 1924. Vermischte Diagnosen I. Notizbl. Bot. Gart. Berlin-Dahlem 9: 49-50.
Buscalioni, L. & J. Huber. 1900. Eine neue theorie der Ameisenpflanzen. Beih. Bot. Centralbl. 9: 85-88.
Bush, M. B. & P. A. Colinvaux. 1988. A 7000-year pol- len record from the Amazon lowlands, Ecuador. Ve- getatio 76: 141-154.
, D. R. Piperno & P. A. Colinvaux. 1989. A 6,000 year history of Amazonian maize cultivation. Nature 340: 303-305.
LITERATURE CITED 191
, P. A. Colinvaux, C. Wiemann, D. R. Piperno & K.-B. Liu. 1990. Late Pleistocene temperature de- pression and vegetation change in Ecuadorian Ama- zonia. Quatern. Res. 34: 330-345.
Carrol, C. R. 1979. A comparative study of two ant fau- nas: the stem-nesting ant communities of Liberia, West Africa, and Costa Rica, Central America. Amer. Naturalist 113: 551-561.*
. 1983. Azteca (hormiga, Azteca, Azteca ants, Cecropia ants). Pp. 691-693. In D. H. Janzen (ed.), Costa Rican natural history. University of Chicago Press, Chicago.*
Charles-Dominique, P. 1986. Inter-relations between frugivorous vertebrates and pioneer plants: Cecropia, birds and bats in French Guiana. Pp. 119-135 in A. Estrada & T. H. Fleming (eds.), Frugivores and seed dispersal. W. Junk, Dordrecht.
Chew, Wee-Lek. 1963. Flora malesianae precursores- XXXIV. A revision of the genus Poikilospermum (Urticaceae). Gard. Bull. Straits Settlem. 20: 1-103.
Clark, D. B. & D. A. Clark. 1990. Distribution and ef- fects on tree growth of lianas and woody hemiepi- phytes in a Costa Rican tropical wet forest. J. Trop. Ecol. 6: 321-331.
Coley, P. D. 1986. Costs and benefits of defense by tan- nins in a neotropical tree. Oecologia 70: 238-241.
, J. Bryant & F. Chapin. 1985. Resource avail- ability and plant antiherbivore defense. Science 230: 895-899.*
Colinvaux, P. A. 1993. Pleistocene biogeography and di- versity in tropical forests of South America. Pp. 473- 499 in P. Goldblatt (ed.), Biological relationships be- tween Africa and South America. Yale University Press, New Haven.
, M. Frost, I. Frost, K.-B. Liu & M. Steinitz- Kannan. 1988a. Three pollen diagrams of forest dis- turbance in the western Amazon basin. Rev. Palaeo- bot. Palynol. 55: 73-81.
, K. Olson & K.-B. Liu. 1988b. Late-glacial and Holocene pollen diagrams from two endorheic lakes of the inter-Andean plateau of Ecuador. Rev. Palaeo- bot. Palynol. 55: 83-99.
Corner, E. J. H. 1962. The classification of Moraceae. Gard. Bull. Straits Settlem. 19: 187-252.
Cronquist, A. 1981. An integrated system of classifi- cation of flowering plants. Columbia University Press, New York.
Cuatrecasas, J. 1944. Notas a la Flora de Colombia VI. Revista Acad. Colomb. Ci. Exact. 6(21): 32-67.
. 1945. Notas a la flora de Colombia, VII. Revista Acad. Colomb. Ci. Exact. 6(22/23): 274-299.
. 1947. Vistazo a la vegetaci6n natural del bajo Calima. Revista Acad. Colomb. Ci. Exact. 7(27): 306-312.
. 1949. Moraceae. In R. E. Schultes, Plantae Col- ombianae XII. Bot. Mus. Leafl. 14: 24-27.
1951. Cecropia kavanayensis. In J. A. Steyer- mark, Botanical exploration in Venezuela-I: Mora- ceae. Fieldiana, Bot. 28: 210.
. 1956. Notas a la flora de Colombia, XIV. Revista Acad. Colomb. Ci. Exact. 9(36/37): 325-341.
. 1959. Studies on South American plants. V. Brittonia 11: 163-172.
. 1967. Cecropia auyantepuiana. In J. A. Steyer- mark, Flora del Auyan-tepui. Acta Bot. Venez. 2: 203.
. 1971. Miscellaneous notes on the neotropical flora. Phytologia 20: 465-481.
. 1976. Cecropia steyermarkii. In J. A. Steyer- mark, La vegetaci6n de la Cima del Macizo de Jaua. Bol. Soc. Venez. Ci. Nat. 32: 321.
. 1982. Miscellaneous notes on the neotropical flora, XIV. Phytologia 52: 157-159.
Davidson, D. W. 1997. The role of resource imbalances in the evolutionary ecology of tropical arboreal ants. J. Linn. Soc., Biol. 61: 153-181.*
Davidson, D. W. (in press). Ecological stoichiometry of ants in a New World rain forest. Oecologia.
& W. W. Epstein. 1989. Epiphyte associations with ants. Pp. 200-233 in U. Litthe (ed.), Vascular plants as epiphytes. Springer-Verlag, New York.*
& B. L. Fisher. 1991. Symbiosis of ants with Cecropia as a function of light regime. Pp. 289-309 in C. R. Huxley & D. F. Cutler (eds.), Ant-plant in- teractions. Oxford University Press, Oxford.**
& D. McKey. 1993. The evolutionary ecology of symbiotic ant-plant relationships. J. Hymenopt. Res. 2: 13-183.*
, J. Longino & R. R. Snelling. 1988. Pruning of host plant neighbors by ants: an experimental ap- proach. Ecology 69: 801-808.*
, R. B. Foster, R. R. Snelling & P. W. Lozada. 1991. Variable composition of some tropical ant- plant symbioses. Pp. 145-162 in P. Price, T. Lewin- sohn, G. Fermandes & W. Benson (eds.), Herbivory: tropical and temperate perspectives. John Wiley, New York.**
, S. C. Cook, R. R. Snelling & T. H. Chua. 2003. Explaining the abundance of ants in lowland tropical rainforest canopies. Science 300: 969-972.*
Didham, R. K. & J. H. Lawton. 1999. Edge structure determines the magnitude of changes in microclimate and vegetation structure in tropical forest fragments. Biotropica 31: 17-30.
Diels, L. 1941. Neue Arten aus Ecuador. IV: Moraceae. Notizbl. Bot. Gart. Berlin-Dahlem 15: 366-368.
Donnell Smith, J. 1899. Undescribed plants from Gua- temala and other Central American Republics. XXI. Bot. Gaz. (Crawfordsville) 27: 434-443.
Downhower, J. F. 1975. The distribution of ants on Ce- cropia leaves. Biotropica 7: 59-62.
Duarte, A. P. 1959. Contribucao para o conhecimento do genero Cecropia na cidade do Rio de Janeiro. Rodriguesia 22(33/34): 177-186.
Eiseman, E. 1961. Favorite foods of neotropical birds: flying termites and Cecropia catkins. Auk 78: 636- 637.
Engler, G. H. A. 1889. Moraceae. Pp. 66-98 in G. H. A. Engler & K. Prantl, Naturlichen Pflanzenfamilien 3(1). Engelmann, Leipzig.
192 FLORA NEOTROPICA
Federle, W., U. Maschwitz, B. Fiala, M. Riederer & B. Holldobler. 1997. Slippery ant-plants and skillful climbers: selection and protection of specific ant part- ners by epicuticular wax blooms in Macaranga (Eu- phorbiaceae). Oecologia 112: 217-224.**
Ferguson, B., D. Boucher, M. Pizzi & C. Rivera. 1995. Recruitment and decay of a pulse of Cecropia in Ni- caraguan rain forest damaged by Hurricane Joan: re- lation to mutualism with Azteca ants. Biotropica 27: 455-460.*
Fiala, B. & U. Maschwitz. 1992a. Domatia as most im- portant adaptations in the evolution of myrmecophy- tes in the paleotropical tree genus Macaranga (Eu- phorbiaceae). PI. Syst. Evol. 180: 53-64.
& . 1992b. Food bodies and their signif- icance for obligate ant-association in the tree genus Macaranga (Euphorbiaceae). J. Linn. Soc., Bot. 110: 61-65.
, & Tho Yow Pong. 1991. The associa- tion between Macaranga trees and ants in South-east Asia. Pp. 263-270 in C. R. Huxley & D. F. Cutler (eds.), Ant-plant interactions. Oxford University Press, Oxford.
, H. Grunsky, U. Maschwitz & K. F. Linsemair. 1994. Diversity of ant-plant interactions: protective efficacy in Macaranga species with different degrees of ant association. Oecologia 97: 186-192.*
Fiebrig, K. 1909. Cecropia peltata und ihre Verhaltnis zu Azteca alfari, zu Atta sexdens, und andern Insek- ten, mit einer Notiz uber Ameisendoruen bei Acacia cavenia. Biol. Centralbl. 29: 1-16, 33-55, 65-77.**
Fleming, T. H. 1988. The short-tailed fruit bat. Univer- sity of Chicago Press, Chicago.
& C. F. Williams. 1990. Phenology, seed dis- persal, and recruitment in Cecropia peltata (Mora- ceae) in Costa Rican tropical dry forest. J. Trop. Ecol. 6: 163-178.
Florence, J. 1997. Cecropiaceae. Flore de la Polyn6sie franqaise 1: 24-26, t. 1.
Folgarait, P. J. & D. W. Davidson. 1994. Antiherbivore defences of myrmecophytic Cecropia under different light regimes. Oikos 71: 305-320.
& . 1995. Myrmecophytic Cecropia: an- tiherbivore defenses under different nutrient treat- ments. Oecologia 104: 189-206.**
, H. Johnson & D. Davidson. 1994. Responses of Cecropia to experimental removal of Mullerian bodies. Funct. Ecol. 8: 22-28.*
Franco Rosselli, P. & C. C. Berg. 1997. Distributional patterns of Cecropia (Cecropiaceae): a panbiographic analysis. Caldasia 19: 285-296.
Galiano-Sanchez, W. 1976. Aspectos taxon6micos, fi- tosociol6gicos y aplicados del genero Cecropia en el valle de Kcosniipata y el Manu. Thesis, Universidad Nacional de "San Antonio Abad" del Cusco.
Garay-Arroyo, A. & E. R. Alvarez-Buylla. 1997. Iso- zyme variation in a tropical pioneer tree species (Ce- cropia obtusifolia, Moraceae) with high contents of secondary compounds. Biotropica 29: 280-290.
Garcia-Barriga, H. 1974. Flora medicinal de Colombia. Botanica medica 1. Universidad Nacional de Colom- bia, Bogota.
Gottsberger, G. 1978. Seed dispersal by fish in the in- undated regions of Humaita, Amazonia. Biotropica 10: 170-183.
Guariguata, M. R. 1990. Landslide disturbance and for- est regeneration in the upper Luquillo Mountains of Puerto Rico. J. Ecol. 78: 814-832.
Halle, F. & R. A. A. Oldeman. 1970. Essai sur l'architecture et la dynamique de croissance des ar- bres tropicaux. Masson, Paris.
& P. B. Tomlinson. 1978. Tropical trees and forests-an architectural analysis. Springer- Verlag, Berlin.**
Harada, A. Y. & W. W. Benson. 1988. Especies de Az- teca (Hymenoptera, Formicidae) especializadas em Cecropia spp. (Moraceae): distribucao e considera- qoes ecol6gicas. Revista Bras. Entomol. 32: 423-435.
Harper, J. 1989. The value of a leaf. Oecologia 80: 53- 58.*
Hemsley, W. B. 1883. Biologia centrali-americana, Bot- any 3. R. H. Porter, London.
Hodges, S. & M. Arnold. 1994. Columbines: a geo- graphically widespread species flock. PNAS 91: 5129-5132.*
Holldobler, B. & E. Wilson. 1990. Host tree selection by the neotropical ant Paraponera clavata (Hymen- optera: Formicidae). Biotropica 22: 213-214.*
Holthuijzen, A. M. A. & J. H. A. Boerboom. 1982. The Cecropia seedbank in Suriname lowland rain forest. Biotropica 14: 62-68.
Huber, J. 1910. Mattas e madeiras amazonicas. Bol. Mus. Paraense Hist. Nat. 6: 91-225.
Ihering, H. von. 1907. Die Cecropien und ihre Schutz- ameisen. Bot. Jahrb. Syst. 39: 666h714.**
Itioka, T., M. Nomura, Y. Inui, T. Itino & T. Inoui. 2000. Difference in intensity of ant defense among three species of Macaranga myrmecophytes in Southeast Asian dipterocarp forest. Biotropica 32: 3 18-326.
Janzen, D. H. 1969. Allelopathy by myrmecophytes: the ant Azteca as an allelopathic agent of Cecropia. Ecol- ogy 50: 147-153.**
. 1973. Dissolution of mutualism between Ce- cropia and its Azteca ants. Biotropica 5: 15-28.**
. 1980. When is it coevolution? Evolution 34: 611-612.*
& D. McKey. 1977. Note: Musanga cecropioi- des is a Cecropia without its ants. Biotropica 9: 57.
Jolivet, P. 1987. Remarques sur la biocenose des Ce- cropia (Cecropiaceae). Biologie des Coelomera Chevrolat avec la description d'une nouvelle espece du Bresil (Coleoptera, Chrysomelidae, Galerucinae). Bull. Mens. Soc. Linn. Soc. Bot. Lyon 56: 255-276.
. 1989. The Chrysomelidae of Cecropia (Cecro-
LITERATURE CITED 193
piaceae): a strange cohabitation. Entomography 6: 391-395.**
- . 1990a. Fourmis, Cecropia et epiphytes. L'Entomologiste 46: 121-127.
. 1990b. Relative protection of Cecropia trees against leaf-cutting ants in tropical America. Pp. 251-254 in R. K. Vander Meer, K. Jaffe & A. Cedeno (eds.), Applied myrmecology. A world per- spective. Westview Press, Boulder, CO.
& P. Salinas. 1993. Ponte de Coelomera cajen- nensis (Fabricius, 1787) dans la tige des Cecropia (Col. Chrysomelidae). Bull. Soc. Entomol. Franq. 98: 472.*
Judd, W. S., R. W. Sanders & M. J. Donoghue. 1989. Angiosperm family pairs: preliminary phylogenetic analyses. Harvard Pap. Bot. 1: 1-51.
Kemper, S. & J. Satore. 2000. Madidi National Park. National Geographic 197(3): 2-23.
Kempf, W. W. 1972. Catalogo abreviado das formigas da regiao neotropical (Hymenoptera: Formicidae). Stud. Entomol. 15: 2-344.
Klotzsch, J. F. 1847. Beitrage zu einer Flora der Aequinoctial-Gegenden: Cecropia. Linnaea 20: 530- 535.
Kubitzki, K. & A. Ziburski. 1994. Seed dispersal in flood plain forests in Amazonia. Biotropica 26: 30- 43.
Kuntze, 0. 1891. Revisio generum plantarum 2. Arthur Felix, Leipzig.
Lamotte, S. 1990. Fluvial dynamics and succession in the Lower Ucayali River basin, Peruvian Amazonia. Forest Ecol. Managem. 33/34: 141-156.
. 1992. Essai d'interpretation dynamique des v6- getations en milieu tropical inondable. La plaine al- luviale de Haut Amazonie. Thesis, University of Montpellier II.
Leck, C. F. 1972. Observations of birds at Cecropia trees in Puerto Rico. Wilson Bull. 84: 489-500.
Leston, D. 1973. The ant mosaic-tropical tree crops and the limiting of pests and diseases. Pestic. Abstr. 19: 311-341.*
Letourneau, D. 1998. Ants, stem-borers, and fungal pathogens: experimental tests of a fitness advantage in Piper ant-plants. Ecology 79: 593-603.*
Liao, Jih-Ching. 1991. The taxonomic revisions of the family Moraceae in Taiwan. Department of Forestry, Taipei.
. 1995. The taxonomic revisions of the family Moraceae in Taiwan. Ed. 2. Department of Forestry, Taipei.
Liebmann, F. M. 1851. Mexicos og Central-Americas nedleagtige planter (Ordo: Urticaceae) indbefattende familierne: Urticeae, Moreae, Artocarpeae og Ulma- ceae. Kongel. Danske Vidensk. Selsk. Nuturvidensk. Math. Afh., Ser. 5, 2: 285-343.
Linnaeus, C. 1759. Systema Naturae. Ed. 10. Impensis Laurentii Salvii, Stockholm.
Liu, K.-B. & P. A. Colinvaux. 1985. Forest changes in
the Amazon basin during the last glacial maximum. Nature 318: 556-557.
& . 1988. A 5200-year history of Ama- zonian forest. J. Biogeogr. 15: 231-248.
Lobova, T. A., S. A. Mori, F. Blanchard, H. Peckham & P. Charles-Dominique. 2003. Cecropia as a food source for bats in French Guiana and the significance of fruit structure in seed dispersal and longevity. Amer. J. Bot. 890: 388-403.
Loefling, P. 1758. Iter Hispanicum. Impensis Laurentii Salvii, Stockholm.
Longino, J. T. 1986. Ants provide substrate for epi- phytes. Selbyana 9: 100-103.*
. 1989a. Geographic variation and community structure in an ant-plant mutualism: Azteca and Ce- cropia in Costa Rica. Biotropica 21: 126-132.**
. 1989b. Taxonomy of the Cecropia-inhabiting ants in the Azteca alfari species group (Hymenoptera: Formicidae): evidence two broadly sympatric spe- cies. Los Angeles County Mus. Contr. Sci. 412: 1- 16.**
. 1991a. Azteca ants in Cecropia trees: taxonomy, colony structure, and behaviour. Pp. 271-288 in C. R. Huxley & D. F. Cutler (eds.), Ant-plant interactions. Oxford University Press, Oxford.
1991b. Taxonomy of the Cecropia-inhabiting Azteca ants. J. Nat. Hist. 25: 1571-1602.*
Macbride, J. F. 1937. Flora of Peru. Moraceae. Publ. Field Mus. Nat. Hist., Bot. Ser. 13(2.2): 274-331.
Marcgrave, G. 1648. Historiae rerum naturalium Bras- iliae (libri VIII). F. Hackium, Amsterdam.
Marshall, J. J. & F. R. Rickson. 1973. Characterization of the a-D-glucan from the plastids of Cecropia pel- tata as a glycogen-type polysaccharide. Carbohydr. Res. 28: 31-37.**
Martinelli, T., G. Hormiga, R. Dirzo, J. Lambert, S. Messier, J. Rettig & K. Schultz. 1993. Ontological changes in defense against herbivory in Cecropia sp. Pp. 48-50 in N. Greig & J. Blake (eds.), Tropical biology: an ecological approach (OTS 93-1). Org. Trop. Stud., Durham.*
Martinez-Ramos, M. & E. R. Alvarez-Buylla. 1986. Seed dispersal, gap dynamics and tree recruitment: the case of Cecropia obtusifolia at Los Tuxtlas, Mex- ico. Pp. 333-346 in A. Estrada & T. H. Fleming (eds.), Frugivores and seed dispersal. W. Junk, Dor- drecht.
Martius, C. F. P. von. 1841. Herbarium florae brasilien- sis. Flora (Regensburg) 24(Beibl. 2): 1-112.
McKey, D. 1984. Interaction of the ant-plant Leonardoxa africana (Caesalpiniaceae) with its obligate inhabi- tants in a rainforest in Cameroon. Biotropica 16: 81- 99.*
1988a. Cecropia peltata, an introduced neotrop- ical pioneer tree, is replacing Musanga cecropioides in southwesterm Cameroon. Biotropica 20: 262-264.
194 FLORA NEOTROPICA
. 1988b. Promising new directions in the study of ant-plant mutualisms. Pp. 335-355 in W. Greuter & B. Zimmer (eds.), Proceedings of the XIC Interna- tional Botanical Congress. Koeltz Scientific, Koenig- stein.*
& D. W. Davidson. 1993. Ant-plant symbioses in Africa and the Neotropics. Pp. 568-606 in P. Gold- blatt (ed.), Biological relationships between Africa and South America. Yale University Press, New Ha- ven.*
Medellin, R. A. 1994. Seed dispersal of Cecropia ob- tusifolia by two species of opossums in Selva Lacan- dona, Chiapas, Mexico. Biotropica 26: 400-407.
Mildbraed, G. J. W. 1925. Plantae Tessmannianae Pe- ruvianae II. Notizbl. Bot. Gart. Berlin-Dahlem 9: 260-268.
. 1933. Cecropia mutisiana. In J. Cuatrecasas, Plantae colombianae novae. Trab. Mus. Ci. Nat. (Ma- drid), Bot. Ser. 26: 3-31.
Milton, K. 1991. Leaf change and fruit production in six neotropical Moraceae species. J. Ecol. 79: 1-26.
Miquel, F. A. G. 1853. Urticineae. Pp. 78-218, t. 25-70 in C. F. P. von Martius, Flora Brasiliensis 4(1). Ol- denburg, Leipzig.
Mooney, H. & S. Gulmon. 1982. Constraints on leaf structure and functions in reference to herbivory. BioScience 32: 198-206.*
Muller, F. 1876. Ueber das Haarkissen am Blattstiel der Imbauba (Cecropia), das Gemiusebeet der Imbauba- Ameise. Z. Naturwiss. 10: 281-286.
. 1880. Die Imbauba und ihre Beschiitzer. Kos- mos 8: 109-116.**
O'Dowd, D. 1982. Pearl bodies as ant food: an ecolog- ical role for some leaf emergences of tropical plants. Biotropica 14: 40-49.*
Oldeman, R. A. A. 1974. L'architecture de la foret guy- anaise. Un modele de la foret guyanaise. Un cas par- ticulier simple: la vegetation secondaire a Cecropia. Memoires O.R.S.T.O.M. No. 73.
Peeters, C. 1997. Morphologically 'primitive' ants: comparative review of social characters, and the im- portance of queen-worker dimorphism. Pp. 372-391 in J. C. Chloe & B. J. Crispi (eds.), Social behaviour in insects and arachnids. Cambridge University Press, Cambridge.*
Piso, G. 1658. De Indiae utriusque re naturali et medica (libri XIV). Elzevirios, Amsterdam.
Pittier, H. 1917. New and noteworthy plants from Co- lombia and Central America. Four new species of Cecropia. Contr. U.S. Natl. Herb. 18: 226-228.
Plowman, T. 1981. Amazonian Coca. J. Ethnopharma- col. 3: 195-225.
. 1984. The ethnobotany of coca (Erythroxylum spp., Erythroxylaceae). Adv. Econ. Bot. 1: 62-111.
Poorter, L. & S. F. Oberbauer. 1993. Photosynthetic induction responses of two rainforest tree species in relation to light environment. Oecologia 96: 193- 199.
Putz, F. E. 1982. How trees avoid and shed lianas. Bio- tropica 16: 19-23.
. 1984. How trees avoid and shed lianas. Biotro- pica 16: 19-23.
& N. M. Holbrook. 1988. Further observations on the dissolution of mutualism between Cecropia and its ants: the Malaysian case. Oikos 53: 121- 125.**
Reekie, E. G. & F. A. Bazzaz. 1989. Competition and patterns of resource use among seedlings of five trop- ical trees grown at ambient and elevated CO2. Oec- ologia 79: 212-222.
Renner, 0. 1907. Beitrage zur Anatomie und Systematik der Artocarpeen und Conocephaleen insbesondere der Gattung Ficus. Bot. Jahrb. Syst. 39: 319-448.
Rettig, E. 1904. Ameisenpflanzen-Pflanzenameisen. Ein Beitrag zur Kenntnis der von Ameisen bewohnten Pflanzen und der Beziehung zwischen beiden. Beih. Bot. Centralbl. 17: 89-122.**
Richter, A. 1897. Uber die Blattstruktur der Gattung Cecropia inbesondere einiger bisher unbekannter Imbauba-Baume des tropischen Amerika. Biblioth. Bot. 43: 1-25, t. 1-8.
Rickson, F.R. 1971. Glycogen plastids in Mullerian body cells of Cecropia peltata-a higher green plant. Science 173: 344-347.**
. 1973. Review of glycogen plastid differentiation in Mullerian body cells of Cecropia peltata. Ann. New York Acad. Sci. 210: 104-114.**
. 1976. Ultrastructural differentiation of the Mill- lerian body glycogen plastid of Cecropia peltata. Amer. J. Bot. 63: 1272-1279.**
. 1977. Progressive loss of the ant-related traits of Cecropia peltata on selected Caribbean islands. Amer. J. Bot. 64: 585-592.
Robinson, B. L. 1912. Cecropia pittieri. P. 389 in A. Stewart, Notes on the botany of the Cocos Islands. Proc. Calif. Acad. Sci., Ser. 4, 1.
Rocha, C. & H. Bergallo. 1992. Bigger ant colonies reduce herbivory and herbivore residence time on leaves of an ant-plant: Azteca muelleri vs. Coelomera ruficornis on Cecropia pachystachya. Oecologia 91: 249-252.*
Romaniuc-Neto, S. 1999. Cecropioideae (C. C. Berg) Romaniuc-Neto stat. nov. (Moraceae- Urticales). Al- bertoa, n.s. 4: 13-16.
Ruiter, G. de. 1976. Revision of the genera Myrianthus and Musanga (Moraceae). Bull. Jard. Bot. Etat 46: 471-510.
Rusby, H. H. 1907. An enumeration of the plants col- lected in Bolivia by Miquel Bang. Part 4 with de- scriptions of new genera and species. Bull. New York Bot. Gard. 4: 309-470.
. 1910. New species from Bolivia, collected by R. S. Williams. Bull. New York Bot. Gard. 6: 487- 517.
Salo, J., R. Kalliola, I. Hakkinen, Y. Makinen, P. Nie- melii, M. Puhakka & P. Coley. 1986. River dynam-
LITERATURE CITED 195
ics and the diversity of Amazon lowland forest. Na- ture 322: 254-258.*
Schimper, A. F. W. 1888. Die Wechselbeziehungen zwischen Pflanzen und Ameisen im tropischen Amerika. Bot. Mitt. Tropen 1: 1-95.**
1898. Planzen-geographie auf physiologischer Grundlage. Gustav Fischer, Jena.
Schupp, E. 1986. Azteca protection of Cecropia: ant oc- cupation benefits juvenile trees. Oecologia 70: 379- 385.*
& D. Feener Jr. 1991. Phylogeny, lifeform and habitat dependence of ant-defended plants in a Pan- amanian forest. Pp. 175-197 in C. Huxley & D. Cut- ler (eds.), Ant-plant interactions. Oxford University Press, Oxford.*
Setoguchi, H, H. Tobe, H. Ogba & M. Okazaki. 1993. Silicon-accumulating idioblasts in leaves of Cecro- piaceae (Urticales). J. PI. Res. 106: 327-335.
Skutch, A. F. 1971. Life history of the keel-billed tou- can. Auk 88: 381-396.
Snethlage, E. H. 1923. Neue Arten der Gattung Cecro- pia nebst Beitragen zu ihrer Synonymik. Notizbl. Bot. Gart. Berlin-Dahlem 8: 357-369.
. 1924. Neue Cecropien aus Nordbrasilien. No- tizbl. Bot. Gart. Berlin-Dahlem 9: 171-172.
Snow, B. K. & D. W. Snow. 1971. The feeding ecology of tanagers and honeycreepers in Trinidad. Auk 88: 291-322.
Sposito, T. C. & F. A. M. Santos. 2001a. Architectural patterns of eight Cecropia species of Brazil. Flora 196: 215-226.
& . 2001b. Scaling of stem and crown in eight Cecropia (Cecropiaceae) species of Brazil. Amer. J. Bot. 88: 939-949.
Standley, P. C. 1929. Studies of American plants-IL. Publ. Field Mus. Nat. Hist., Bot. Ser. 4: 301-345.
. 1940a. Studies of American plants-X. Publ. Field Mus. Nat. Hist., Bot. Ser. 22: 3-62.
. 1940b. Studies of American plants-X. Publ. Field Mus. Nat. Hist, Bot. Ser. 22: 65-129.
& J. A. Steyermark. 1944. Studies of Central American plants VI. Publ. Field Mus. Nat. Hist., Bot. Ser. 23: 153-191.
& L. 0. Williams. 1952. Plantae Centrali- Americanae IV. Ceiba 3: 101-132.
Stevenson, P. R., M. J. Quifiones & J. A. Ahumada. 2000. Influence of fruit availability on ecological overlap among four neotropical primates at Tinigua National Park, Colombia. Biotropica 32: 533-544.
Strauss-Debenedetti, S. & F. A. Bazzaz. 1991. Plastic- ity and acclimation to light in tropical Moraceae of different successional positions. Oecologia 86: 377- 387.
Terborgh, J. 1983. Five New World primates. A study in comparative ecology. Princeton University Press, Princeton.
Trecul, A. 1847. Sur la famille des Artocarpees. Ann. Sci. Nat. Bot., s6r. 3, 8: 38-157, t. 1-6.
Ule, E. 1900. Verschiedene Beobachtungen vom Gebiet der Baumbewohnenden Utricularia. Ber. Deutsch. Bot. Ges. 18: 249-260. [Cecropia: 255-260.]
Vasconcelos, H. L. & A. B. Casimiro. 1997. Influence of Azteca alfari ants on the exploitation of Cecropia trees by a leaf-cutting ant. Biotropica 29: 84-92.**
Vaizquez-Yanes, C. & A. Orozco-Segovia. 1986. Dis- persal of seed by animals: effect on light controlled dormancy in Cecropia obtusifolia. Pp. 71-77 in A. Estrada & T. H. Fleming (eds.), Frugivores and seed dispersal. W. Junk, Dordrecht.
& H. Smith. 1982. Phytochrome control of seed germination in the tropical rain forest pioneer trees Cecropia obtusifolia and Piper auritum and its eco- logical significance. New Phytol. 92: 477-485.
Velasquez, J. 1971. Contribuci6n al conocimiento de las especies del genero Cecropia L. (Moraceae) "Yagru- mos" de Venezuela. Acta Bot. Venez. 6: 25-64.
Vepsalainen, V. 1999. The avifauna feeding on fruits of Cecropia angustifolia in a premontane rainforest in the western Andes of Colombia. Thesis, University of Helsinki.
Vester, H. 1997. The trees and the forest. The role of tree architecture in the canopy development; a case study in secondary forests (Araracuara, Colombia). Ponsen & Looyen, Wageningen.
Visiani, R. de. 1842. Ort. Bot. Padova. Angelo Sicca, Padova.
Ward, P. 1991. Phylogenetic analysis of ant-plant as- sociations involving pseudomyrmecine ants. Pp. 335- 352 in C. Huxley & D. Cutler (eds.), Ant-plant in- teractions. Oxford University Press, Oxford.*
Welle, B. J. H. ter, K. Bonsen & P. Detienne. 1992. Wood and timber. Pp. 141-191 in A. R. A. Gorts-van Rijn (ed.), Flora of the Guianas, Ser. A., Fasc. 11. Koeltz Scientific, Koenigstein.
Wetterer, J. K. 1997. Ants on Cecropia in Hawaii. Bio- tropica 29: 128-132.**
Wheeler, W. M. 1913. Observations of the Central American Acacia ants. Trans. 2d Congr. Entom. Ox- ford (1912) 2: 109-139.*
. 1942. Studies of neotropical ant-plants and their ants 1. The neotropical ant-plants. Bull. Mus. Comp. Zool. (Harvard) 90: 3-154.**
& J. C. Bequaert. 1929. 2. Amazonian myr- mecophytes and their ants. Zool. Anz. 82: 10-39.
White, P. S. 1983. Corner's rule in eastern deciduous trees: allometry and its implications for the adaptive architecture of trees. Bull. Torrey Bot. Club 110: 203-212.*
Wijmstra, T. A. 1967. A pollen diagram from the Upper Holocene of the Lower Magdalena Valley. Leidse Geol. Meded. 39: 261-267.
Wijninga, V. M. 1996. Paleobotany and palynology of Neogene sediments from the high plains of Bogota'. Ponsen & Loijen, Wageningen.
196 FLORA NEOTROPICA
Willdenow, C. L. 1806. Species plantarum 4. G. C. Nauk, Berlin.
Wilson, E. 0. 1985. Invasion and extinction in the West Indian ant fauna: evidence from the Dominican am- ber. Science 229: 265-267.
Woodson, R. E. & R. W. Schery. 1960. Flora of Panama, Moraceae. Ann. Missouri Bot. Gard. 47:114-178.
Yu, D. & D. W. Davidson. 1997. Experimental studies of species-specificity in Cecropia-ant relationships. Ecol. Monogr. 67: 273-294.*
NUMERICAL LIST OF TAXA
1. Cecropia albicans Trecul 2. C. andina Cuatrecasas 3. C. angulata I. W. Bailey 4. C. angustifolia Trecul 5. C. annulata C. C. Berg & P. Franco 6. C. bullata C. C. Berg & P. Franco 7. C. chlorostachya C. C. Berg & P. Franco 8. C. concolor Willdenow 9. C. distachya Huber
10. C. elongata Rusby 11. C. engleriana Snethlage 12. C. ficifolia Snethlage 13. C. gabrielis Cuatrecasas 14. C. garciae Standley 15. C. glaziovii Snethlage 16. C. goudotiana Trecul 17. C. granvilleana C. C. Berg 18. C. herthae Diels 19. C. heterochroma C. C. Berg & P. Franco 20. C. hispidissima Cuatrecasas 21. C. hololeuca Miquel 22. C. idroboi Cuatrecasas 23. C. insignis Liebmann 24. C. kavanayensis Cuatrecasas 25. C. latiloba Miquel 26. C. litoralis Snethlage 27. C. longipes Pittier 28. C. marginalis Cuatrecasas 29. C. maxima Snethlage 30. C. megastachya Cuatrecasas 31. C. membranacea Trecul 32. C. metensis Cuatrecasas
33. C. montana Snethlage 34. C. multisecta P. Franco & C. C. Berg 35. C. mutisiana Mildbraed 36. C. obtusa Tr6cul 37. C. obtusifolia Bertoloni 38. C. pachystachya Trecul 39. C. palmata Willdenow 40. C. pastasana Diels 41. C. peltata Linnaeus 42. C. pittieri Robinson 43. C. plicata Cuatrecasas 44. C. polystachya Trecul 45. C. purpurascens C. C. Berg 46. C. putumayonis Cuatrecasas 47. C. reticulata Cuatrecasas 48. C. sararensis Cuatrecasas 49. C. saxatilis Snethiage 50. C. schreberiana Miquel
a. subsp. schreberiana b. subsp. antillarum (Snethlage) C. C. Berg & P.
Franco 51. C. sciadophylla Martius 52. C. silvae C. C. Berg 53. C. strigosa Trecul 54. C. subintegra Cuatrecasas 55. C. tacuna C. C. Berg & P. Franco 56. C. telealba Cuatrecasas 57. C. telenitida Cuatrecasas 58. C. ulei Snethlage 59. C. utcubambana Cuatrecasas 60. C. velutinella Diels 61. C. virgusa Cuatrecasas
LIST OF EXSICCATAE
Abbensetts, N. J., 57 (36). Abbott, J. R., 16816 (39). Acero D., E. [et al.], 43 (57), 89 (37). Acevedo R., P. [et al.], 2845 (SOb), 2956 (50a), 7490
(31). Acosta C., S., 1217 (37). Acosta Solis, M., 5848 (6), 12777 (4), 16221 (23). Adalardo-Oliveira, A., 2771 (12).
Agostini, G. et al., 1624 (41). Agra, M. F., 682 (38). Agudelo, C. A. et al., 1043 (56), 1429 (4), 3100 (35),
3114 (35). Aguilar, 0. T., 508 (38). Ake Assi, L. et al., 3960 (41). Alarc6n, R., 88 (11). Albert de Escobar, L., 984 (26).
LIST OF EXSICCATAE 197
Albuquerque, B. W. [et al.], 5 (45), 8 (51), 13 (8), 311 (25).
(15). Amaral, I. L., 2082 (51). Amorin, A.M.A. et al., 1475 (38). Amorozo, M. C., 240 (39). Ancuash, E., 50 (31), 1020 (46), 1020 (46), 1094 (46). Andeer, M., s.n. (38). Anderson, A. B. et al. 2073 (39), 2181 (39). Anderson, A. I. et al., 1450 (8). Anderson, W. R. [et al.], 2184 (38), 3000 (41), 3001 (41),
Andrade, R., 33149 (31). Andre, E., 2574 (56). Andreata, R.H.P. et al., 937 (15). Angeli, C., 20 (15), 239 (38), 326 (15), 344 (15), 345
(15), 347 (15), 48 (15), 50 (21). Antonio, T., 1309 (20). Appun, C. F., 1710 (25). Arana, A., 7 (11), 74 (51). Araque M., J. et al., 159 (37). Araquistain, M. [et al.], 1718 (37), 3033 (37), 3244 (37). Araujo, D. S. Dunn de [et al.], 224 (15), 608 (38), 4918
(38), 5016 (38), 5246 (38), 9055 (38). Arbo, M. M. et al., 1463 (38), 5346 (49). Arbocz, G. et al., 33417 (15). Archer, W. A., 588 (41), 7645 (39). Argent, G. [et al.], 6426 (38), 6705 (38). Aristeguieta, L. et al., 1482 (41), 2308 (41), 7435 (25),
7531 (41). Amason, T. et al., 17016 (41). Arroyo, L. et al., 326 (9). Ascencio R., J., 140 (41). Asplund, E., 7849 (2), 9140 (28), 9198 (12), 9261 (31),
Assungao, P.A.C.L. et al., 3204.2267.2 (45), 3209.3028 (9), 3304.6610 (58).
Atahuachi, M. et al., 23 (31), 24 (51). Atwood, J. T. et al., 70 (41). Aubreville, A., 15 (36), 23 (41), 354 (25). Aulestia, C. et al., 1385 (37). Aulestia, M. et al., 2826 (51). Aviles, S., 61a (37). Axelrod, F. et al., 2062 (SOa), 2427 (SOb), 2787 (SOb),
3772 (SOa), 4921 (SOb). Ayala, F. (et al.), 689 (12), 2901 (31), 3310 (25), 5791
(25), 5930 (25). Aymard C., G. et al., 811 (41), 3655 (41), 4146 (3).
Baker, C. F., 2007 (41). Baker, M. A. [et al.], 6317 (28), 6724 (33). Baksh, Y. S. et al., 321 (41). Balansa, B., 2575 (38). Balcazar, J. [et al.], 1033 (44), 2177 (59). Balee, W. L. [et al.], 408 (51), 809 (8), 1830 (36), 2670
(8), 2783 (36), 2801 (51). Balick, M. et al., 1811 (41), 2183 (41), 2630 (37). Balslev, H. (et al.), 1984 (13), 2280 (31), 84740 (26),
97160 (26), 97161 (26), 97162 (25). Bamps, P., 5018 (38). Bang, M., 2260 (10). Barbosa, C., 1191 (23), 1194 (23), 1334 (31). Barbour, P. J., 5820 (11). Barclay, A. S. [et al.], 623 (51), 2730 (41), 3197 (48),
3506 (35), 3557 (41). Barclay, G. W., 2064 (41). Barfod, A. et al., 48433 (37), 48615 (13). Barkley, F. A. [et al.], 17C675 (13), 38C212 (41), 39574
(41). Barlow, F. D., 30 (41). Barr, R. J. et al., 62-305 (37). Barrera, E., 228 (4). Barringer, K. et al., 1607 (4). Barros, F. de [et al.], 1929 (38), 2382 (15), 2385 (21),
2469 (38), 2585 (38), 2694 (38). Barroso, G. M., 282 (38). Bartolomeu, J. [et al.], 12999 (38), 13000 (38), 15193
(15). Bastos, M., 23 (36). Bausen, E., s.n. (21).
198 FLORA NEOTROPICA
BBS (boschbeheer Suriname), 4 (25), 223 (25). Beard, J. S., 180 (SOa). Beck, H. T. et al., 32 (36), 2237 (37). Beck, S. G. et al., 2504 (8), 3329 (8), 3793 (44), 5292
Begazo, N., 48 (51). Be1anger, G. P., s.n. (50a). Bel6m, R. P. et al., 2074 (38). Belshaw, C. M., 3224 (59). Beltran, W., 13 (4). Benavides, 0. de, 10102 (14), 11138 (6). Bennett, B. C. et al., 3801 (11), 4474 (46). Benoist, R., 279 (36), 442 (51). Benson, W., 7954 (38), 8304 (12), 8465A (58), 10341
Bertero, C. G., s.n. (SOa). Betancur, J. et al., 1255 (27), 1256 (37). Biesmeijer, J. C. et al., 188 (41). Bigay, ? et al., DI.805 (31). Black, G. A., 51-14103 (36). Blackmore, S. et al., 1655 (37), 3797 (41). Blanchet, J. S., 464 (38), s.n. (15). Blanco, C. A., 616 (3), 790 (41). Blum, K. E. [et al.], 1169 (41), 1383 (37), 1861B (37),
Broadway, W. E., 259 (41), 582 (36). Brown, S. et al., 1683 (41). Buchanan-Smith, 3 (51). Buchtien, O., 2118 (4). Bulhoes, M. et al., 226 (38). Bullock, 2022 (37). Bunting, G. S. [et al.], 1052 (41), 3790 (12), 6341 (41),
Chac6n, I. A. et al., 1563 (4), 1816 (4). Chagas, J., 138 (8), 1288 (45), 7280 (8). Chaguaro, ?, 36 (51). Chalot, C., 48 (41). Chan, C. et al., 1432 (41).
200 FLORA NEOTROPICA
Chavelas, J. et al., 2463 (41). Chavez A., R., 389 (44), 409 (53), 463 (44), 1721 (44),
3389 (44). Chickering, A. M., 43 (41). Christensen, ?, 82750 (31), 82794 (31). Churchill, H. W., 5720 (41). Clark, H. L., 6879 (12), 6880 (31). Clark, J. L. et al., 140 (47), 1004 (51), 1121 (28), 1530
Coimbra, C., 71B (11). Colella, M. et al., 1444 (3). Combs, R., 111 (50b). Conant, D. S., 1053 (8). Contreras, E., 1007 (41), 1390 (41), 5832 (41), 5845
(41). Converse, O., s.n. (4). Conzatti, C. [et al.], 3242 (37), 4518 (37). Cook, 0. F., et al., 3 (41), 336 (41), 813 (SOa). Corbusier Boland, ?, 2061 (38). Comejo, X. et al., 3578 (47). Correa A., M. D. et al., 27 (27), 575 (37), 850 (37), 1806
(19). Correa L., G. et al., 384 (9), 392 (9), 407 (9). Cortes, L. et al., 111 (37). Cortes B., R. et al., 1545 (41), 1546 (23). Costa, J. C. da et al., 1202.2406 (51), 1202.2862 (58). Cowan, C. P., 38280 (36). Cox, D. K., 561 (37). Crane, C. L., 54 (41), 499 (41). Cremers, G. [et al.], 12329 (51), 12330 (36), 12490 (36),
13269 (25), 13270 (25), 15193 (51). Croat, T. B. [et al.], 1884 (27), 4796 (37), 5030 (41),
Davis, D. H., 79 (41), 239 (3), 240 (3), 247 (51). Davis, E. W. et al., 921 (51). Del Aguila, M. et al., 90 (11), 168 (31), 196 (51). Delascio C., F. et al., 2578 (41), 2782 (51), 3446 (41),
Dulmen, A. van [et al.], 58 (12), 147 (31), 148 (12). Dumont, M. et al., 41 (35). Duno, R. et al., 174 (25). Duque-Jaramillo, J. M., 2030 (25), 2206 (25), 2590 (35),
(SOa). Dwyer, J. D. [et al.], 147 (41), 553 (37), 3447 (19), 7110
(20), 9885 (37), 11017 (41).
Echavarria, J. A. et al., 149 (27), 249 (41). Eden, M. J., 12A (12). Edwall, G., 6403 (21). Eggers, H.F.A., 6829 (50a), s.n. (SOb). Eggers, L. et al., 3402.5108 (51). Egler, W. A., 413 (39), 437 (36),439 (51),445 (52),1018
(38), 45948 (25), 47658 (36). Eiten, G. et al., 6196 (38). Ekman, E. L., 983 (SOb), 2013 (38), 6548 (SOb), 9376
(SOb), 12324 (SOb). Elcoro, S., 81 (3). Elias, Bro., 578 (41), 1230 (41). Elias, T. S. et al., 1606 (41). Ellenberg H., 880 (55), 2242 (51), 2296 (51), 2385 (31),
Erlanson, C. O., 233 (41). Ernani Diaz, B., 446 (15), 452 (38), 453 (38). Ernst, A., 1756 (4). Ervik, F., 36871 (29). Escobar, L. et al., 9935 (SOb). Escobar M., E., s.n. (4). Espadas, M. et al., 96 (37). Espina, J. [et al.], 695 (37), 1920 (41), 2745 (41). Espinal T., S., 2325 (56), 2746 (31). Espinoza, R., 1575 (57). Espinoza, S. et al., 379 (51), 585 (51). Evans, ?, 1551 (37). Ewan, J. A., 16731 (12), 16732 (11).
Faber-Langendoen, D. et al., 256 (61), 1106 (14). Falcao, J. I. Almeida de et al., 1155 (38). Fanshawe, D. B., 731 (51), 732 (3), 2990 (3), 3033 (41). Farinias, M. [et al.], 349 (31), 378 (51), 501 (11), 502
Fleury, M., 1026 (36), 1063 (36), 1065 (51). "Flora Falc6n (HW, ES)," 460 (4). Flores N., C., 312 (31). Focke, H. C., s.n. (41). Folli, D. A., 310 (21), 1051 (38), 3567 (15). Folsom, J. P. [et al.], 5226 (41), 6161 (19), 6351 (47),
7766 (41), 8979 (37). Fonnegra, R. et al., 2070 (37), 4221 (37). Fonseca, M. L. et al., 1570 (38). Forero, E. [et al.], 116 (57), 1554 (37), 1871 (41), 1890
Fredholm, A., 3266 (41). Freeman, J., 35 (41). Freire, E. et al., 2784 (9), 3260 (51). Freire, F.M.T., s.n. (49). Freire-Fierro, A. et al., 1797 (49). Freitas, ?, 129 (31). Fr6es, R. de Lemos, 11634 (38), 19911 (39). Fromm, E. et al., 471 (8). Fuchs, H. P. et al., 21921 (37), 22112 (37). Fuertes, M., 78 (SOb). Furtado, C. X., s.n. (41). Furtado, P. Pires, 154 (38).
Gabriel, A., s.n. (39). Gabrielli, A. C. et al., 8751 (38), 8752 (38), 8758 (44),
8759 (44), 8759 (15).
LIST OF EXSICCATAE 203
Galarza, M. I. et al., 2 (8), 22 (51). Galeano, G. et al., 512 (35), 513 (35), 552 (35), 562 (35),
Galeano S., W., 3 (8), 9 (9). Galo, M., 3 (41). Garber, P. A., 57 (41), 154 (41). Garcia, H. et al., 60 (8). Garcia-Barriga, H. et al., 4692 (35), 12286 (41), 14012
Gregory, L. E., 67 (50a). Grenand, P., 1542 (36), 1690 (39), 2428 (8). Grewal, M. S., 215 (3). Griffis, J. D. et al., s.n. (SOb). Grijalva, A. et al., 16 (41), 68 (51), 91 (12), 1355 (41). Groll-Meyers, J. van, 105 (SOa). Grubb, P. J. et al., 1627 (28), 1331 (2). Guanchez, F., 786 (9), 787 (12). Gudiiio, E. [et al.], 164 (12), 179 (18), 519 (46), 586 (46),
Guillaumet, J.-L. et al., 5850 (32), 5851 (32). Guillen, R. et al., 1314 (39), 2686 (49), 3170 (25), 4430
(8). Guti6rrez, G. et al., 17C675 (13), s.n. (13). Gutierrez, V. et al., 542 (12), 769 (51). Gutte, P. et al., 1566 (11), 1578 (12), 8161 (44). Guzman, M. et al., 378 (41).
Hage, J. L. [et al.], 662 (15 and 38), 2255 (38). Hagen, C. von et al., 1063 (41), 1264 (37). Hahn, L., 210 (SOa), 342 (37), s.n. (50a). Hahn, R., 6 (31). Haines, ?, 467 (37). Hall, E., 53 (41). Hall6, F., 992 (36). Hamilton, C. et al., 616 (19), 1108 (37). Hansen, B. et al., 9158 (50a). Harley, R. M. [et al.], 10838 (39), 10968 (38), 15223
(38), 18099 (38), 27357 (38), 50510 (38). Harling, G. [et al.], 3809 (12), 21363 (59). Harmon, W. E., 1828 (37), 4932 (41), 5198 (37). Harriman, ?, 14663 (37). Harris, C. G., 1612 (41). Harris, S. A., Y.29 (51), TP.578 (3), TP.583 (3). Hartshorn, G. S. [et al.], 2490 (38), 2694 (33). Harvey, D. R., 5282 (37). Hassler, E., 617a (38), 617b (38), 2278 (38), 7924 (38),
Hermnndez, R. et al., 488 (41). Hermnndez X., E. H., 606 (37), 635 (37). Herrera, H. et al., 644 (41), 1763 (19). Herzog, T., 1508 (8). Hespenheide, H. A. et al., 874 (41). Heyde, E. J. [et al.], 6238 (37). Heyde, N. M., 710 (51). Hill, M., 16 (37). Hill, R. J., 1722 (8).
Hind, D.J.N. et al., 40 (38). Hinojosa, I. et al., 1104 (44). Hinton, G. B., 3568 (37). Hioram, Bro., s.n. (SOa). Hladik, A., 497 (41). Hodge, W. A. [et al.], 632 (SOa), 3168 (SOa), 3663 (SOa),
4403 (50b). Hoehne, F. C. et al., s.n. (38). Hoehne, W. [et al.], 2004 (21), 2005 (21), 3325 (15),
Hoff, M., 6244 (36), 6866 (51). Hohenkerk, L. S., l1SA (51). Holdridge, L. R., 1061 (SOb). Holm, R. W. et al., 234 (41), 950 (41). Holm-Nielsen, L. et al., 20205 (31), 20211 (31), 20426
Holt, E. G. et al., 248 (25), 657 (25). Holton, I. F., 251 (31), s.n. (31). Hopkins, M.J.G. et al., 872 (25), 1422 (45). Homer, C. et al., 73 (25), 360 (25). Hostmann, F.W.R. et al., s.n (36). Howard, R. A. et al., 18982 (50a). Hoyos, J. et al., 822 (41). Huashikat, V., 253 (12), 793 (12), 982 (12), 1189 (9),
Huber, O., 6348 (41), 9235 (24). Hunt, D. R., 5778 (38). Hunter et al., ?, 462 (37). Hunziker, J. H. et al., 11026 (38). Hurtado, F. et al., 2403 (31.note), 2457 (4), 2581 (4). Hutchinson, P. C. et al., 3035 (4), 3009 (4), 3028 (4),
3039 (13 and 57), 3844 (59).
Ibarra M., G. et al., 1467 (37), 2002 (37), 2468 (37). Ibarrola, T. S., 4060 (38). Idrobo, J. M. [et al.], 604 (32), 662 (22), 792 (51), 1092
Jenman, G. S., 4975 (36). Jimenez, S. et al., 358 (10). Jimenez M., A., 2682 (37), 3184 (42), 3473 (41). Johann, M. [et al.], 1/3-31188 (51), JO-2/35 (11), 2/32-
Johnson, G. E. et al., 18C240 (41), 22J181 (41). Johnson, W. et al., 127 (47). Johnston, I. M., 78 (41), 154 (41), 155 (41), 471 (41),
1503 (27), 1758 (23). Joly, A. B., 7349 (15). Jones, G. C. et al., 3034, 3465 (41). Jones, G. N. 11024 (SOa). Jong, W. de, 163 (12). Jordy Filho, S., 53 (39). J0rgensen, P. [et al.], 2158 (38), 3836 (38). J0rgensen, P. M. et al., 65761 (57). Joseph, ?, 13100 (41). Josse, C. et al., 785 (37). Juncosa, A. [et al.], 695 (37), 1134 (37). Juzepczuk, S., 4863 (41), 6411 (12), 6710 (41), 6928
Kapelle, M. et al., MK.519 (4). Kauffmann, E., 603 (51). Kayap, R., 213 (51), 368 (46), 442 (12), 680 (28), 805
(31), 832 (11), 1052 (28), 1099 (11). Kellerman, W. A., 6039 (37), 6416 (41). Kenoyer, L. A., 310 (41). Kernan, C., 1064 (23). Khan, R. et al., 1155 (41). Killeen, T. [et al.],, 1083 (8), 3072 (4), 4407 (51).
Kujikat, A., 132 (11). Kuntze, O., s.n. (37). Kvist, L. P. et al., 48433 (37).
Ladeira, J. et al., 264 (15). Laessoe, T. et al., H.52562 (38). Laguna, A., 399 (41), 400 (41). Lallathin, B. R., 5065 (23). Lamotte, S., 123 (9), 126 (12), 166 (25), 231 (25), 248
(41), 2485 (51), s.n. (25). Lanna Sobrinho, J. de Paula [et al.], 257 (15), 290 (8),
403, (25), 429 (31), 451 (12). Lao, E. A. [et al.], 120 (27), 121 (27), 122 (41), 169 (37),
181 (20), 203 (41), 211 (23). Lao M., R., 21 (57), s.n. (59). La Rotta, C. et al., 543 (51), 723 (37). LaSalle, ? et al., 810704-8 (37). Laughlin, R. M., 1282 (41).
206 FLORA NEOTROPICA
Lauri, ?, 187 (36). Lawesson, J. E. et al., 39398 (12). Lazor, R. L. [et al.], 1710a (37), 1710d (37), 2653 (37),
Lechler, W., 2436 (4 and 53). LeDoux, D. G., et al., 2122 (37). Leemans, J., 83 (38). Leeuwenberg, A.J.M., 4504 (41). Leguizamo, I., 493 (41), 494 (41). Lehmann, F. C., 1969 (61), 2341 (35). Leija, G. et al., 1112 (37), 2720 (37). Leitao Filho, H. F., 20345 (38). Leite, J. E., 2300 (38). Lemonnier, L. G. (herb.), s.n. (36). Lent, R. W., 1031 (23), 1614 (4), 2052 (4), 2119 (23),
Lizot, J., s.n. (11). Llanos H., F. et al., 1372 (35). Lleras, E. [et al.], P.17033 (12), P.17161 (12), P.17161
(12), P.17553 (8). Locke, R. D. et al., 73 (38). Loefgren, A. et al., 1657 (38). Loiz, A. et al., 37 (50a). Lojtnant, B. et al., 13297 (28), 14569 (28). Lombardi, J. A. [et al.], 331 (38), 637 (38), 1757 (49),
2214 (49). Lopes, J. M. et al., 2206.2257 (9). Lopes, D., s.n. (45). L6pez, F., 537 (41), 788 (37). L6pez, G. et al., 262 (37), 351 (37). Lorence, D. H. et al., 3436 (37). Loroca, S., 25 (38). Lott, E. J. et al., 1154 (37), 2213 (37). Loubry, D., 1570 (36), 1570 (39), 1766 (51), 2351 (36). Loureiro, A. et al., 37937 (58), 38041 (8), 48349 (8). Lowe, J., 3999 (8). Lozano, P. et al., 901 (4), 902 (26), 903 (26), 912 (26). Lozano C., G. [et al.], 314 (12), 901 (4), 1078 (4), 2877
Macedo, A., 1503 (38). Macia, M. J. et al., 325 (12), 2491 (51). Maciel, U. N. et al., 2113 (31). Mackenzie, C. A. et al., 2206.3034 (45). Madriiain, S. et al., 406 (41), 407 (41), 458 (41), 458A
(41), 982 (12), 1000 (12). Maestro, A. L. et al., 6 (38). Maguire, B. et al., 34787 (12), 41691 (12), 54150 (25),
Marquete, N., 1590 (15). Marquez, W., 717 (37). Mars, L. et al., 3304.2552 (9). Marshall, N. [et al.], 86 (37), 188 (36), 394 (41). Martin, B.-A. (herb), s.n. (36). Martinelli, G. et al., 5011 (38), 6965 (25), 9908 (21). Martinez C., R. et al., 10849 (38). Martinez-Calderon, G., 3106 (37). Martinez S., E. M., 7004 (37). Martins, H. J. et al., 1301.523 (45). Martins, L.H.P. et al., 17 (45). Martius, C.F.P. von, 630(1) (51), 630(2) (excl. name),
2834 (38), s.n. (25, 37). Marulanda, 0. et al., 1204 (9). Mat Asri, FRI 25549 (41). Mathes, L.A.F., 31C (38), 10084 (38). Mathias, M. E. et al., 3446 (53). Matthes, L., 1 (15). Mattos Silva, L. A. et al., 1155 (38), 2674 (21). Matuda, E., 3240 (41), 16411 (37). Maxon, W. R. [et al.], 5132 (37), 7193 (41). Maxwell, I., s.n. (41). Maxwell, J. F., 79-59 (44). McDaniel, S. et al., 29832 (12). McDonagh, J. F. et al., 79 (20), 405 (37). McDowell, T., 2455 (25). McVaugh, R., 717 (37), 11957 (37). Meave, J. et al., B.265 (37). Meijer Drees, E., 20 (41). Mejia, M. et al., 6732 (SOb), 6733 (SOb). Melendez, E. J. et al., 175 (SOa). M6linon, M., 18 (36), s.n. (39). Mello, M.M.R.F. et al., 565 (38). Mello-Silva, R. et al., 547 (21). Mendonca, R. C., 235 (38). Mendonca Filho, C. V., 28 (38). Meneces, E. [et al.], 45 (11), 786 (51). Merino, B. et al., 4514 (59), 4779 (2).
Merklen, E., s.n. (41). Metcalf, R. et al., 30168 (37). Mexia, Y, 1275 (37), 4349 (15), 4389 (21), 9225 (37). Michel, R. de et al., 2437 (51). Miege, E. et al., 3960 (41). Miller, G. S. [et al.], 1251 (SOb), 1350 (41), 6000 (SOa). Miller, J. S. et al., 953 (41), 1604 (SOb). Milliken, W. [et al.], 159 (41), 388 (25), 408 (41), 2101
(51). Mills, A. P. 22 (44). Miranda, ?, s.n. (39). Molina R., A. [et al.], 427 (41), 3455 (37), 13571 (37),
17928 (37), 18102 (37), 18220 (37). Monsalve B., M., 627 (37), 1501 (61), 1843 (37). Monteiro, L. et al., 122 (38), 123 (38). Monteiro, 0. P. [et al.], 1126 (51), 1278 (45). Moore, et al., 3642 (37). Mora 0., L. E., 2310 (37), 2311 (14), 2310 (37), 3185
(4), 4474 (4). Moraes, M.C.C. et al., 279 (49). Moraes R., M. et al., 263 (31), 800 (8), 982 (31), 1239
(25), 2091 (8), 2286 (8). Morais, H. C., 9201 (21), s.n. (49). Morales, G. et al., 1116 (57). Morales, M. E. et al., 647 (46), 648 (46). Morawetz, W. et al., 11-11988 (9), 11-18988 (51), 15-
Nicholson, D., 2037 (SOa). Noberto, F. et al., 215 (38). Noblick, L. R. et al., 2106 (38). Norby, ?, 133 (37). Noriega, R., s.n. (9). Nuiiez I., 85 (44), 86 (44), 87 (33). Nufiez, P. [et al.], 65 (40), 5461 (51), 5748 (51), 6044
(41). Oliveira, E., 6139 (39). Oliveira, A. A. de, 2771 (45). Oliver, R. L. et al., 1884 (27), 1901 (41). 0llgaard, B. et al., 57100 (12), 57707 (47), 90836 (2). O'Neill, H., 8819 (41). Orcutt, C, R., 3030 (41). Orozco, C. I. et al., 2181 (20). Ortega, J. G., 4761 (37), 5086 (37), 5194 (37). Ortega, R., 128 (SOa). Ortega U., A., 148 (28), 205 (51), 361 (41). Ortiz, L., 203 (47). Ortiz, M., 805 (38). Ortiz S., G. A. [et al.], 96 (37), 110 (41), 1979 (41). Outer, R. W. den, 881 (41).
Pacheco, M. et al., 174 (8). Paixao, J. L. et al., 74 (38). Palacios, E., 66 (41). Palacios, P. et al., 2655 (9)
Palmer, W. et al., 768 (SOb). Pardo, M. de et al., 75 (25). Paredes, R., 680 (19), 681 (19), 924 (19). Pastore, U. et al., 84 (38). Pedersen, T. M., 945 (38), 5923 (38), 13463 (38). Pedralli, G. et al., s.n. (38). Peixoto, A. Luna et al., 359 (38). Pennell, F.W., 3635 (16), 8042 (56). Pennington, R. T. et al., 31 (11). Pennington, T. D. et al., 9073 (37), 9273 (4), 14054 (23),
Pereira, B.A.S. et al., 3448 (38), 3514 (38), 3542 (38). Pereira, D. F. et al., 3 (38). Pereira, E. [et al.], 271 (38), 2838 (38), 3247 (39), 3715
(38), 3944 (15). Pereira, M.J.A. et al., 1301.2629 (9), 2303.1319 (9). Pereira, M.J.G. et al., 2303.2165 (45). Pereira, M.J.R. et al., 2302.593 (51), 1301.3155 (51),
2303.2114 (9), 3304.4133 (51). Pereira, 0. J. et al., 4014 (38). Pereira Neto, M., 337 (38). Perez Arbelaez, E. et al., 5737 (35), 6117 (4), 8135 (4),
8301 (41), 8334 (35), 8371 (57), 8402 (35). Perino, C.-H., 3275 (37). Persaud, C. A., 242 (36), 243 (51), 252 (41). Peterson, P. M. et al., 6858 (37). Peyton, B. et al., 374 (55). Pfeifer, W., 1726 (41). Pfeiffer, H. W. et al., 2619 (SOa). Philipson, W. R. et al., 1623 (12), 1982 (4), 1983 (4),
10145 (38.vs). Phillips, 0. et al., 172 (31), 181 (51), 554 (31). Pic6n N., G. [et al.], 1108 (24), 1484 (41). Pimentel, J. et al., 575 (50b). Pinheiro, P. R., 175 (38). Pinto E., P. et al., 941 (12), 1165 (32), 6268 (31), 6290
(25), 6299 (25). Pinz6n-Rodrfguez, M., s.n. (35). Pipoly, J. et al.,8217 (41), 11288 (41), 11316 (41), 13115
Poeppig, E., 1593 (44), s.n (44). Pohl, J. E., s.n. (21 and 38). Poiteau, P. A., s.n. (39). Polak, M. et al., 333 (3). Porter, D. M. [et al.], 4068 (19), 4194 (37). Pozetti, G. L., s.n. (38). Prance, G. T. et al., 1700 (36), 1778 (51), 3460 (58),
Pulle, A. A., 94 (25). Purpus, C. A., 15387 (37). Pursell, ? et al., 8241 (41).
Quadros, J. et al., 1677 (38), 1678 (38), 1679 (15), 1680 (15).
Quarfn, C. et al., 2446 (38). Quarles van Ufford, L. H., 94 (37). Quelal, C. [et al.], 65 (13), 273 (47). Quevedo, R. et al., 2572 (39). Quisumbing, E., 7933 (38).
Rabelo, B. V., 62 (8), 3112 (51). Ramage, G. A., s.n. (SOa). Ramalho, R. S. [et al.], 316 (38), 1337 (21), 1388 (15),
1825 (15), 3398 (38). Rambo, B., 29298 (38), 53542 (38), 63578 (38). Ramfrez, J. G. et al., 691 (23), 909 (37), 1212 (37), 4151
(43), 4184 (43), 4242 (43), 4327 (37). Ramfrez P., B. R. [et al.], 4167 (4), 6061 (29). Ramos, J. E. et al., 3071 (31). Rangel, 0. et al., 13234 (4). Ratter, J. A. et al., 202 (38), 5081 (38), 5503 (39), 6300V
Ribeiro, R., 21 (21). Richardson, W. D., 683 (41). Ricksecker, A. E., 449 (SOb). Ridley, H. N. et al., s.n. (38). Ridley, ?, s.n. (SOa). Riedel, L. [et al.], 74 (15, 78). Riera, B. J. J.-L. [et al.], 649 (36), 650 (39), 905 (51),
1943 (51). Rimachi Y., M., 593 (12), 8352 (25). Rios, M. et al., 154 (20). Rios T., J., 100 (4), 1801 (44). Rivera, G., 1933 (23), 1934 (4). Rivero, E., 135 (11). Rizzini, C. M., 205206 (49). Rizzo, A. et al., 4642 (49). Roberts, L., 12791 (51). Robinson, J.W.L., 364 (31). Robles, R., 1446 (37). Robleto, W., 401 (41). Rocha, D.M.S., 10370 (38). Rodrigues, R. S., 7841 (39), 7842 (39). Rodrigues, W. [et al.], 403 (12), 1793 (51), 2090 (45),
(25), 1951 (9). Sanchez V., I. et al., 6060 (4), 8611 (33). Sandino, J. C., 168 (23). Sandoval, E. et al., 942 (37). Sandwith, N. Y., 574 (3). Sanoja, E. et al., 3329 (11). Santos, F.A.M. dos et al., 14228 (9). Santos, J. L. dos et al., 1202.4522 (51). Santos, J. U. et al., 659 (39). Santos, M. R., 352 (51). Santos, R. R. de et al., 1503 (38). Santos, T. S., 1950 (21), 2063 (38). Sapper, ? et al., 2178 (38). Saravia T., C., 2419 (41). Sartori, A. et al., 32671 (15). Sarukhan, J. et al., 2090 (37). Sastre, C. [et al.], 2404 (12), 3110 (12), 3116 (12), 6909
(37), 614 (41). Shilom Ton, A., 3075 (41), 3631 (37), 3894 (37). Sidek bin Kiah, S.60 (44). Sieber, F. W., s.n. (8, 39). Silva, A.S.L. da et al., 450 (12). Silva, I. A., 190 (38). Silva, J. M. da et al., 3304.596 (51). Silva, M., 992 (25). Silva, M. F. et al., 173 (12), 501 (12), 1381 (12). Silva, M.F.F. da et al., 15 (8), 16 (8), 67 (36), 388 (25). Silva, M. G. [et al.], 4441 (49), 5724 (39). Silva, N. T., 956 (52), 1178 (36), 1226 (51), 1340 (36),
3284 (52).
LIST OF EXSICCATAE 211
Silveira, M. et al., 712 (12), 815 (12), 1310 (25), 1311 (25), 1327 (31), 1329 (31).
Sim6es, J. M. et al., 73 (15). Simonis, J. E. et al., 134 (38). Sintenis, P., 250b (50b), 3971 (50b), 4805 (50b). Skutch, A. F., 3986 (37). Smith, A. C., 2164 (3), 3426 (41). Smith, C. L., 57 (37), 1040 (41). Smith, D. N. et al., 465 (2), 1832 (55), 1875 (51), 2422
Smith, L. B. et al., 5897 (38). Smith, L. C., 681 (37). Smith, N., B.38 (36). Smith, R. F., V.526 (48), V.910 (57). Smith, S. F. et al. 785 (44), 1608 (44). Sneidem, K. von, A.61 (23), A.71 (37), A.72 (31), A.73
Soares, A. M. et al., 2303.255 (58). Sodiro, A., s.n. (13). Soejarto, D. D., et al., 112 (35), 2661 (23), 2662 (41). Solomon, J. C. [et al.], 3437 (12), 6165 (12), 7982 (51),
Stoffers, A. L. [et al.], 159 (3), 317 (51), 339 (36), 457 (32), 516 (32), 4230 (SOa).
Stork, H. E., 4621 (37). Strang, H. E., 343 (38), 601 (38), s.n. (39). Strelnikow, I., 13 (38), 76 (38), 86 (38). Strube, L. B., s.n. (SOa). Strudwick, J. J. et al., 3187 (25). Sturrock, ?, 714 (50a). Sucre, D. [et al.], 795 (38), 1968 (15), 3641 (38), 4923
(38), 5834 (38). Sugden, A., 1182 (41), 1204 (41). Sutton, ? et al. 152 (41). Swartz, O., s.n. (41). Sylvestre, L. et al., 821 (38).
Tamashiro, J. Y. et al., 688 (15), 1255 (15), 18821 (38), 18844 (38).
Tamayo, F. T., 2981 (41).
212 FLORA NEOTROPICA
Tameirao Neto, E. [et al.], 747 (38), 898 (15), 899 (38). Tate, G.H.H., 250 (24), 394 (37). Taylor, C. M. et al., 8823 (50a). Taylor, D., 125 (50a). Taylor, J. [et al.], 12628 (41), 17540 (37). Taylor, K., 394 (37). Taylor, N., 235 (50b). Taylor, R. J., 4393 (4). Teague, G. W., 397 (38). Teixeira, L.O.A. et al., 672 (51). Telles, ?, 49542 (15). Tellez, 0. et al., 2189 (41), 2510 (41), 3352 (41), 3425
Velasquez, Ma. P. et al., 238 (41), 293 (23). Velez, M. C. et al., 724 (35), 1693 (4), 2926 (56), 2997
(57), 3370 (4). Ventura, E. et al., 729 (37), 2705 (37). Vera Santos, J., 2891 (37). Versteeg, G. M., 418 (25). Vester, H. et al., 501 (9), 725 (51), 726 (9), 727 (12). Vianna, M. C. et al., 341 (38), 342 (38). Vicentini, A. et al., 574 (45). Vickers, W. T., 91 (9), 92 (18). Vieira, C. M. et al., 81 (15), 554 (44). Villanueva, ?, 321 (41). Vinha, S. G. da, 27 (38). Vivar C., F. et al., 2844 (26). Vogl, C., 787 (41). Voorhies, B. et al., 16-23 (41). "V.V.," 5139 (48).
Williams, R. S., 339 (37), 400 (51), 645 (8). Wilson, P., 229 (37). Wood, C. W., 429 (4). Woodbury, R. et al., s.n. (SOa). Woodson, R. E. et al., 1312 (27), 1313 (37). Woodworth, R. H. et al., 644 (37). Woolston, A. L., 835 (38). Woronow, S. et al., 4556 (41), 6302 (25). Worthington, R. D., 21363 (37). Woytkowski, F., 23 (11), 34544 (55). Wright, C., 1440 (SOb), s.n. (41). Wurdack, J. J. [et al.], 1849 (9), 2025 (46), 40997 (9),
41691 (12), 42785 (25), 43287 (12), 43634 (25).
Yanez, A. P. et al., 300 (20), 1473 (37). Young, K. [et al.], 202 (51), 341 (59), 357 (2), 1052 (51),
1544 (57), 4071 (1). Yuncker, T. G. [et al.], 4573 (37), 4573 (41), 4573 (37),
Zambrano, ?, 222 (41). Zanette, V. C. et al., 1438 (15). Zanoni, T. et al., 15282 (SOb), 38796 (SOb), 40487 (SOb). Zappi, D. C. et al., 10335 (38). Zardini, E. M. [et al.], 4211 (38), 5084 (38), 6287 (38),
for the number of myrmecophytes, or true "ant- plants" counted among its species (McKey & David- son, 1993). Based on the proportion of Cecropia spe- cies producing Mullerian bodies in at least some parts of their distribution, myrmecophytes comprise the vast majority (ca. 80%) of species in the genus; most nonmyrmecophytes occur at higher elevations and on islands, where their ants are missing (Wheeler, 1942). Geographically, myrmecophytic Cecropia occur throughout the latitudinal range of the genus, from southern Mexico to northern Argentina. Given that association with ants is so widespread taxonomically and geographically, it is likely that relationships with ants have been highly influential in the evolutionary diversification of Cecropia. Here, I review the benefits which Cecropia-ants and their hosts receive from their symbiotic partnerships. I then discuss how interspe- cific variation in the ant-attractants of Cecropia might have evolved and then influenced relationships with particular ant taxa. Finally, I consider factors influ- encing the diversity of ants, plants, and partnerships in these associations.
BENEFITS TO CECROPIA-ANTS
Myrmecophytes are plants with "biotic defenses," or traits which attract ants, mainly as a defense against herbivores and encroaching vines (Davidson & McKey, 1993). By definition, myrmecophytes have evolved not only to provision ants with food re- sources, but also to grant them residence inside either modified plant parts or preadapted structures. Evolu- tion by plants to attract and house ant colonies can often be in doubt in presumed myrmecophytes with primary domatia, i.e., naturally hollow or hollowed- out stems that are frequently or always occupied by ants, because many of these species have no obvious specializations for housing or feeding associated col- onies (Davidson & McKey, 1993). Such is not the case in Cecropia species, which accommodate ants by widening their naturally hollow stems at fixed de- velopmental stages, predictable from theories of de- fensive investment (see below), and temporally cor- related with the onset of food body production. Moreover, as described above by Berg, myrmecophy- tic Cecropia also produce prostomata, or weakened sites in the stem walls beneath internodal septa. Pros- tomata lie at the distal ends of the internodal grooves that occur even in nonmyrmecophytic Cecropia, per- haps due to pressure exerted by axillary buds (Bailey, 1922). However, their evolutionary enlargement rel-
ative to the internodal groove is thought to be the product of natural selection imposed by mutualistic ants (Schimper, 1888). Both the internodal groove and prostoma are devoid of fibrovascular bundles, collen- chyma, and lignified parenchyma (Schimper, 1888). Colonizing queens of obligate Cecropia-ants recog-
nize and use prostomata as easily excavated sites where stems can be entered without rupturing stem transport tissues and flooding internodes with muci- lage.
The stems of myrmecophytic Cecropia are hollow, lacking the pith that often fills stems of nonmyrme- cophytic congeners. The thin, spongy medullary lin- ing the hollow stems of myrmecophytes is exploited by queens that found their colonies claustrally, i.e., without foraging, and by using resources from di- gested wing muscles to produce their first worker broods. Claustrally founding queens scrape this ma- terial from the internode walls to seal the prostoma and develop their colonies in seclusion. Foundresses of some Cecropia-ants also accumulate and store
medullary tissue near their developing broods. Water contained within this succulent material (and lending it the appearance of "bubblepack" under a stereo- scope), may nourish developing brood and/or enhance humidity in the vicinity of brood.
In addition, as described above by Berg, Cecropia have evolved to supply two types of food rewards for their ants, and at least some ant associates of Cecropia appear to reject foods typically taken by ants with more generalized habits. Mullerian bodies, produced at hairy trichilia, located abaxially on the bases of petioles, are harvested by obligate Cecropia-ants, but
not by opportunistically foraging ants with general- ized diets (Davidson & Fisher, 1991). These food rewards contain considerable lipid and at least some protein (Rickson, 1973, 1976) but are approximately 30% glycogen (Rickson, 1971; Marshall & Rickson, 1973), the form of soluble polysaccharide in which animals store excess glucose. Synthesized and stored in plastids, the glycogen component of Mill- lerian bodies lends credence to the hypothesis that these rewards may be nutritional mimics of herbivor- ous insects or their larvae and thus may attract ants which typically feed on such prey (Janzen, 1969, 1973).
Worker ants often stand guard at trichilia to await the appearances of Miillerian bodies. In several Ce- cropia species for which the diurnal production schedules of these food bodies have been studied in
CECROPIA AND ITS BIOTIC DEFENSES 215
the greenhouse, the rewards were produced in greatest profusion immediately after dark (Davidson & Fisher, 1991), coincidentally or not, the time of day when many predator-wary herbivores emerge from their di- urnal hiding places to settle at feeding sites for the night. The same diurnal schedule of Mullerian body production was demonstrated for two Cecropia spe- cies in a project from an Organization for Tropical Studies field course in Peru, near the junction of the Rio Sucusori with the Rio Napo (pers. obs.), but a field study by Belin-Depoux et al. (1997) in French Guiana showed production peaking three hours before sunset in C. obtusa. Additional field studies of food body production schedules in different species should shed greater light on the adaptive significance of these schedules. Interestingly, both the manual removal of previously emerged Miillerian bodies (Folgarait et al., 1994) and slight downward pressure on petioles (pers. obs.) can induce preemergent bodies to be released early. The significance of pressure changes in these premature emergences is the suggestion that changes in turgor pressure, coincident with shutdown of pho- tosynthesis and transpiration, could trigger their nat- ural release. Fast-growing plants like Cecropia move back toward water balance relatively quickly after dark, and the period immediately after dark may rep- resent the start of the least expensive time (nighttime) to produce these rewards.
Cecropia also produce tiny, lipid-rich and protein- containing pearl bodies ("pearl glands" to botanists) on abaxial leaf surfaces (Rickson, 1976). These bod- ies are not usually apparent in the field, since they are removed rapidly and continuously by obligate ant in- habitants, or in the absence of these, by unspecialized and opportunistic ants. However, pearl bodies are readily found on greenhouse-grown plants and in the field, on plants or plant parts protected from foraging ants. In contrast to trichilia, pearl glands are simple structures. As an alternative or adjunct to extrafloral nectaries, pearl bodies are taxonomically widespread as a biotic defense of myrmecophilic plant species, i.e., those that attract ants with food rewards but not nest sites (e.g., O'Dowd, 1982; Schupp & Feener, 1991). Their occurrence both in nonmyrmecophytic Cecropia and in closely related Musanga and Cous- sapoa suggests that their presence may be plesio- morphic in the genus.
Inside host-plant stems, many obligate Cecropia- ants also tend coccids and mealybugs (Hemiptera: Coccidae and Pseudococcidae), and they may exca- vate pits in external layers of nodal diaphragms to permit coccids to feed on the soft internal layers con- taining strands of conducting tissues. The numbers
and biomasses of Hemiptera are typically low com- pared with those in plants that almost always house ants but have few or no obviously ant-attractive traits (reviewed in Davidson & McKey, 1993). Becerra & Venable (1989) have argued that myrmecophytes should be selected to reduce ant-tended Hemiptera, which not only consume plant resources but also transmit diseases, alter plant metabolism and devel- opment, and are difficult to oppose by chemical de- fenses. By provisioning symbiotic ants with mainly carbohydrate rewards, myrmecophytes may induce the ants to balance their diets by consuming a greater fraction of their protein-rich associates. Interestingly, in insular Cecropia populations lacking ants, at least externally feeding Hemiptera can reach outbreak den- sities and, combined with overgrowth by vines, may even limit the species' distribution (Janzen, 1973). (See also Fiala et al., 1994, for indications that sym- biotic ants may protect myrmecophytic Macaranga [Euphorbiaceae], the Asian equivalent of Cecropia, from damaging infestations of pseudococcids.)
BENEFITS TO CECROPIA
Despite earlier theories postulating no significant effect of ants on plant fitness (Rettig, 1904; von Iher- ing, 1907; Ule, 1906; Fiebrig, 1909; Wheeler, 1913; Andrade & Carauta, 1982; but see Muller, 1880, and Schimper, 1888), symbiotic ants appear often to ben- efit myrmecophytic Cecropia by protecting their host plants against herbivores (Schupp, 1986; Ferguson et al., 1995; Vasconcelos & Casimiro, 1997; Davidson & Yu, unpubl. data), and/or vines and other vegetation that potentially compete with the hosts for light (Jan- zen, 1969; Schupp, 1986; Davidson et al., 1988; but see Putz & Holbrook, 1988). The occurrence of ant- occupied trees with foliar damage cannot be taken as evidence against a beneficial role of ants, since alter- native ant species may occur and differ in the quality of protection proffered. Moreover, rates of leaf pro- duction and investment in biotic defenses may vary across habitats in ways that affect the quality of de- fense. In a long-term, experimental study in Rio Pal- enque, Ecuador (Schupp, 1986), herbivore densities and herbivory rates were lower on ant-inhabited sap- lings of Cecropia vs. obtusifolia than on plants that lacked ants either naturally or after experimental removal. There, Azteca constructor was effective in defending its hosts against foliar herbivory by large- bodied, chewing insects, including leaf beetles (e.g., Coelomera atrocaerulea, Chrysomelidae). Protection was effective in the dry season but not the wet season, when herbivores were more abundant. During the 15-month investigation, saplings defended
216 FLORA NEOTROPICA
by ants against herbivores and encroaching vines grew significantly faster than did those lacking ants. Shorter-term studies concur that obligate Cecropia- ants defend against insect herbivores. Thus, Rocha & Bergallo (1992) showed that colony size was posi- tively related to resistance against herbivory.
Leafcutter ants can also pose significant threats to Cecropia, especially in habitats not subject to fre- quent inundations that kill colonies of these ground- nesting species. Vasconcelos & Casimiro (1997) con- ducted an 18-month, experimental study of such herbivory in central Brazil. Frequency of attack was not related to leaf palatability as assessed in feeding trials, but for three of the four species included in the study, to occupation by colonies of Azteca ants. Thus, although the fastest-growing species (Cecropia dis- tachya) was most likely to be attacked when ants were absent, and to be favored by leafcutter ants in feeding trials, occupied saplings of this species contained larger worker populations than did stems of a slower- growing species, and their leaves also benefited most from ant attendance. (Likewise, in myrmecophytic Asian Macaranga, species suffering most from ant removal are those apparently producing greater quan- tities of ant rewards, which in tum support larger worker populations [Itioka et al., 2000]). The slower- growing species (C. ulei) was least preferred by leaf- cutters in feeding trials and among plants lacking Az- teca, but it was also least likely to be colonized by Azteca and most likely to lose its Azteca colonies. It is possible that slow-growing species invest more in physical and chemical defenses of leaves (see below). Across all four host species, more attacked than un- attacked saplings died during the observation period, though the difference was not significant at small sample sizes, and plants attacked more than once grew more slowly than did those attacked just once or not at all.
Most studies of the effects ants on herbivory of myrmecophytic and myrmecophilic plants have fo- cused on worker protection of leaf blades (e.g., Beat- tie, 1985). However, the location of Miillerian bodies at the bases of petioles in Cecropia attracts workers mainly here and to adjacent stems. The principal value of symbiotic ants (as opposed to that of oppor- tunistic, nonresident species that consume just pearl bodies) could therefore be the protection these sym- bionts afford petioles and stems. (See also Fiala et al., 1994, for a potentially similar situation in Macaranga [Euphorbiaceae].) At least some leafcutter ants (Az- teca laevigata) damage or destroy terminal meristems of Cecropia and remove entire leaves by severing
their petioles (Vasconcelos & Casimiro, 1997). More- over, thick-stemmed (pachycaulous) plants are often colonized by stem-boring insects such as the weevils (Coleoptera, Curculionidae) that are extremely com- mon in Cecropia stems at higher elevations in the tropics (H. Hespenhide, pers. comm. for Costa Rica; pers. obs., for at least 1500-2000 m in the eastern An- des of southern Peru). At lower elevations, leaf beetles (Chrysomelidae) plague young Cecropia throughout their South American distribution (Andrade, 1981, 1984a; Schupp, 1986; Jolivet, 1989; Davidson & Fisher, 1991; Rocha & Bergallo, 1992; Jolivet & Sa- linas, 1993). Some of these beetles (e.g., in the genus Coelomera) lay eggs inside stems, where larvae hatch and emerge to feed on either leaves or their petioles (Andrade, 1981, 1984a; pers. obs.). Where herbivory rates are high, Cecropia seedlings are hard-pressed to stay ahead of herbivory by producing new leaves just as the previous ones are either removed entirely or largely consumed, with only skeletons of veins left behind (pers. obs. along rivers of southeastern Peru). The severing or weakening of petioles can be disas- trous for plants like Cecropia, which invest many re- sources in very large individual leaves. Like leafcutter ants, stem-inhabiting beetles can also damage termi- nal meristems, slowing the vertical height growth nec- essary for Cecropia to overtop other fast-growing competitors in light gaps and other disturbed habitats. For seedlings and saplings with one or very few meri- stems, this type of damage would likely convey sub- stantial material and opportunity costs.
Given the diverse types and high levels of damage attributable to stem-dwelling herbivores, it is possible to conclude that the primary importance of symbiotic ant colonies (vs. opportunists feeding on pearl bodies) is to preclude stem occupancy by herbivorous beetles, perhaps especially in vulnerable young Cecropia seedlings and saplings. In accord with this argument, an unusual Cecropia species, C. hispidissima, illus- trates how protection of leaves and stems may be funded by different ant rewards that are monopolized by different ants. Stem-inhabiting colonies of Pachy- condyla sp. nov. 1 have been found to date only on Cecropia hispidissima in Panama, and their workers apparently harvest Miillerian bodies from beneath the stipules and without ever visiting the leaves (T. Kur- sar, pers. comm.). Nevertheless, atypically large, hard, and purple Millerian bodies of this host would appear to be adapted for use by the large-bodied Pachycon- dyla rather than by tiny Azteca ants. Pachycondyla workers do not exclude other ants, such as those of nonresident Azteca colonies, from frequent opportun-
CECROPIA AND ITS BIOTIC DEFENSES 217
istic foraging on leaves, probably for pearl bodies. In a hypothetical ancestor of myrmecophytic Cecropia, opportunistic ants may have harvested pearl bodies and protected leaves without alleviating selection for ways of ridding the plant of stem-dwelling herbi- vores. Mullerian bodies and prostomata may have evolved under such selection pressures. Moreover, protection of stems from stem-boring insects may start as soon as queens begin to colonize myrmeco- phytic Cecropia; many or most internodes of individ- ual seedlings and saplings are colonized indepen- dently by one or more queens, and queens of some species do feed while developing their first worker broods (see below).
Accounts of other myrmecophytes also suggest that ants may protect against stem-boring insects. Thus, Pheidole ants of myrmecophytic Piper in Cen- tral America have been shown to protect hosts against stem-boring weevils (Letourneau, 1998). Replace- ment of stem-boring plant parasites by ants is thought to have been an early stage in the evolution of myr- mecophytism in other plant taxa with primary dom- atia (e.g., Ward, 1991). Although ants probably often tended Hemiptera within these domatia, the net ef- fects of ants and Hemiptera on plants may have been positive, especially in habitats where abundant light allowed rapid carbon gain to offset losses to Hemip- tera (Davidson & McKey, 1993). Herbivore pressures may also have been more intense in comparatively productive, sunny environments (Davidson & Fisher, 1991; Davidson & McKey, 1993; Davidson & Yu, un- publ. data), reinforcing selection for ant attraction.
Finally, for Cecropia species inhabited by more than one species of ant, the quality of defense may vary with the identity of the partner. Obligate sym- bionts of Cecropia differ in their diurnal foraging schedules, the extent of their activity and aggression, the numbers and sizes of workers, and whether or not workers prune vines and other vegetation (Davidson et al., 1991; Davidson & Fisher, 1991; Longino, 1989b, 1991). Evolution on the part of the plant may produce adaptations that favor one ant species over another, but to some extent, the predominant inhabi- tant may be determined by the outcome of ant-ant competition and be beyond the plant's control (Da- vidson & McKey, 1993; Yu & Davidson, 1997). Poorly defending ants may sometimes gain posses- sion of their hosts by virtue of rapid colony devel- opment, or pleometrosis, i.e., colony founding by two or more queens, whose combined first worker broods should produce larger and more competitive incipient colonies (e.g., Davidson et al., 1991).
INTERSPECIFIC VARIATION IN
DEFENSIVE INVESTMENT
Early ecological studies of Cecropia were under- taken mainly in the Central America lowlands, where species diversity of both the host plants and their ants is relatively low. It is not surprising, then, that rela- tionships between Cecropia and its ants were initially considered rather uniform. That is, all myrmecophytic Cecropia produced Muillerian (and perhaps pearl) bodies, and despite very early reports to the contrary (Wheeler, 1942), all Cecropia were implied to house Azteca ants (Janzen, 1969, 1973; Rickson, 1976, 1977). Not until the early 1990s did research begin to focus on interspecific variation in the ant-attractants of Cecropia and the significance of this variation to ant associates (Longino, 1989a; Davidson et al., 1991; Davidson & Fisher, 1991; Yu & Davidson, 1997). It now appears that there is important and in- teresting interspecific variation in the ontogeny, quantity, and composition of biotic defenses, and that some of this variation both accords well with general theories of plant defensive investment and is impor- tant to ants.
Plant defense theory begins by postulating that de- fenses are costly and are manufactured and used only when benefits, calculated in reduced herbivory, ex- ceed costs. The costliness of biotic defenses in Ce- cropia is perhaps most apparent when ant-attractive traits are lost in the absence of selection imposed by the typical ant associates. It is interesting, then, that populations of C. peltata on Jamaica both occur in the absence of their ants and are polymorphic in their expression of trichilia (see treatment of C. peltata in Berg, above).
Several other observations also provide evidence for a cost of biotic defenses in Cecropia. First, food bodies are produced mainly at stages of leaf devel- opment and whole-plant development when they are apt to do the most good, including when chemical and physical defenses are poorly developed. In the species surveyed to date, production of either Muillerian bod- ies, pearl bodies, or both occurs within days of leaves first beginning to emerge from their stipules, and peaks relatively early in leaf lifetimes (week 1 for a pioneer species, and week 5-6 for a small gap species: Folgarait & Davidson, 1995). Young leaves are es- pecially vulnerable to herbivores (Davidson & Fisher, 1991), probably due to their high foliar nitrogen levels and poorly developed physical and chemical defenses (Folgarait & Davidson, 1995). In addition, the value of a leaf to the plant is greatest at this stage, since most of its productive life span lies ahead (Harper,
218 FLORA NEOTROPICA
"Reclaimable" extrafloral nectar and pearl bodies
Cumulative "No n-reclaimable" tannin & cost of defense lignin or Mullerian bodies
Short B Long
Leaf lifetime FIG. 1. The McKey (1984) model of plant defenses, modified to accommodate the biotic defenses of Cecropia.
Cumulative costs of "reclaimable" biotic defenses (e.g., pearl bodies of Cecropia) are lower than those of "non-reclaimable" defenses (e.g., tannins and lignins, or trichilia and Mullerian bodies), for relatively short-lived leaves and other plant parts. However, they are higher in species for which "non-reclaimable" defenses can be amortized over the life spans of long- lived leaves.
1989). In relation to whole-plant development, tests of leaf palatability to herbivores suggest that Cecropia seedlings are better protected by chemical and phys- ical defenses before than after the plants acquire their ants (Martinelli et al., 1993).
Three final patterns are indicative of a cost to the production of food rewards for ants. First, both Mill- lerian and pearl bodies are responsive to supplies of key resources (Folgarait & Davidson, 1994, 1995). Fertilization (N-P-K) and high light intensity increase Mullerian body production per unit time and per ac- tive lifetime of trichilia. In contrast, pearl body pro- duction is enhanced by nutrients but not by higher light intensities, perhaps because these rewards have higher nitrogen content than do Muillerian bodies, and because an increase in carbon fixation under high light draws nitrogen away from pearl body production and into growth as the first priority. Second, across species, there appears to be a trade-off between in- vestment in biotic vs. chemical and physical defenses (Vasconcelos & Casimiro, 1997). Third, both the rate of Miillerian body production, and the total produc- tion per lifetime of a trichilium, increase in response to their manual removal, simulating harvesting in the presence of ants (Folgarait et al., 1994).
Granting then that defense can be costly, two im- portant theories have related defensive investment ei- ther to resource availability (Coley et al., 1985) or to
leaf life spans (McKey, 1984). The resource avail- ability theory starts with the premise that plant growth rates vary in response to the availability of limiting resources, typically light in tropical forests. Plant spe- cies characteristic of relatively light-rich environ- ments have evolved rapid growth and low levels of defense, since costly defensive investment would ex- act opportunity costs in the form of reduced growth that could lead to overtopping by fast-growing competitors. In contrast, slow-growing species of resource-poor (shaded) habitats would have little to lose by defending (i.e., low opportunity costs) and perhaps much to gain, since low resource availability prevents rapid replacement of lost tissues.
In contrast, McKey's leaf lifetime hypothesis sug- gests that plants should use different kinds of defenses depending on whether leaf lifetimes are short or long (Fig. 1). Relatively fast-growing plants with short- lived leaves should invest mainly in "reclaimable de- fenses," equated by McKey to biotic defenses (ex- trafloral nectar and, presumably also, pearl bodies). These defenses have low initial construction costs but significant maintenance costs, so cumulative costs in- crease linearly through time. As leaves age and ac- quire other defenses such as toughness, the resources invested in biotic defenses can be routed to new leaves at little expense. Therefore, biotic defenses are often produced on the newest, most valuable, and most vul-
CECROPIA AND ITS BIOTIC DEFENSES 219
nerable foliage. In contrast, McKey postulated that comparatively slow-growing plants with long-lived leaves could afford to invest in "non-reclaimable" de- fenses, such as tannins, lignin, fiber, etc. Once laid down, these defenses were (theoretically) permanent and could not be broken down and remobilized to new tissues. Only in plants with long-lived leaves could the high initial construction costs of non-reclaimable defenses be amortized over a sufficiently long period so that their cumulative costs would be exceeded by the cumulative costs of "reclaimable" defenses.
Interspecific differences in investment in the biotic defenses of Cecropia are well explained by these the- ories if a distinction is made between "reclaimable" pearl bodies and less reclaimable Muillerian bodies and trichilia, with the latter having higher initial con- struction costs (Folgarait & Davidson, 1994, 1995). The more light-demanding "pioneer" Cecropia spe- cies, typical of very open (often riparian) habitats, have the relatively rapid growth rates and short-lived leaves characteristic of other tropical rain forest pio- neers. In accord with McKey's theory, such species tend to invest proportionately more heavily in pearl bodies and less heavily in Muillerian bodies (evaluated as dry weight of reward per unit of leaf area) than do related but more shade-tolerant species with slower intrinsic growth rates and longer leaf life spans (Fol- garait & Davidson, 1994, 1995). Greater investment in Muillerian bodies is also consistent with the re- source availability theory of plant defensive invest- ment, because leaf lifetimes tend to be highly corre- lated with (and lower in) light-rich habitats (Mooney & Gulmon, 1982).
Using the McKey (1984) model as originally for- mulated to compare myrmecophytes and myrmeco- philes with plants lacking biotic defenses altogether may also help to explain the distribution of nonmyr- mecophytes within the genus Cecropia. Investment in non-reclaimable tannins, fiber, and lignin would be predicted to replace partially or mainly reclaimable biotic defenses in plants with comparatively long leaf life spans. It is interesting in this context to note that nonmyrmecophytic Cecropia sciadophylla, a rela- tively slow-growing Cecropia species of infertile soils on higher terraces in Amazonia, produced longer- lived leaves than did any of eight myrmecophytic Cecropia species with which it was grown simulta- neously, or which were grown in other experiments in temperate greenhouses (D. Davidson et al., unpubl. data). Because of the correlation between intrinsic plant growth rates and mean leaf life spans (Folgarait & Davidson, 1994, 1995), a decline in myrmecophy- tism with decreasing intrinsic growth rates at high
elevation might also be anticipated, but a lack of ap- propriate ant species could also play a role (Wheeler, 1942). Finally, plant defense theories based on either leaf life spans or resource availability, and developed explicitly for foliar defenses, should also apply to de- fenses of other plant parts, such as stems benefiting from ant occupation; this is particularly true consid- ering that new stems of Cecropia tend to be photo- synthetic.
Interspecific differences in Cecropia reveal an- other pattern that was not previously predicted, but was readily apparent because biotic defenses are eas- ily observable externally (Davidson & Fisher, 1991; Folgarait & Davidson, 1994, 1995). The onset of (bi- otic) defense occurs earlier in seedling development for relatively slow-growing, shade-tolerant Cecropia than for their faster-growing, light-demanding rela- tives-a pattern that could prove typical in other (e.g., chemically defended) genera of tropical trees but is currently poorly explored. Early in seedling ontogeny, leaves are produced without trichilia, and stems both lack prostomata and are too narrow to accommodate ant colonies. In common garden experiments, the pro- duction of active trichilia and the swelling of stems concurrent with the appearance of prostomata, occur earlier in time and earlier in development in relatively slow-growing, shade-tolerant Cecropia species than in their faster-growing relatives.
Some of the most distinctive myrmecophytic Ce- cropia occur in small light gaps, uncharacteristically shaded habitats for this largely "pioneer" genus. Low light slows growth, increasing the probability that Az- teca colonies will fail (Vasconcelos & Casimiro, 1997), yet it also prolongs rates of replacement of leaves lost to herbivores. In this context, some species have evolved very early developmental switches to myrmecophytism. For example, in western Ama- zonia, trichilia first appear on median leaf number six in "Cecropia pungara," though at leaf number 13 in the apparently closely related Cecropia membranacea (Davidson et al., 1991). (Based on gene sequence data [S. Cook and D. Davidson, unpubl. data], "C. pun- gara" appears to be a distinct species but is treated as a "form" by Berg & Franco [this volume, see discus- sion under C. membranacea].) Remarkably, in "Ce- cropia herrerensis" of northeastern Peru, trichilia are produced on the first set of leaves after cotyledons. From the perspective of the ants that colonize their seedling hosts as soon as swollen stems with prosto- mata appear, tiny, slow-growing juveniles of small- gap Cecropia are a far different resource than are tall, rapidly growing pioneer Cecropia of light-rich habi- tats. Although small-gap species may supply more re-
220 FLORA NEOTROPICA
sources to their colonies per area of leaf surface (Fol- garait & Davidson, 1994, 1995), rates of resource supply per plant are the more important factor to de- veloping colonies, and those rates are greater in com- paratively fast-growing pioneer relatives of the small gap species. Therefore, it is not surprising that the early-defending, shade-tolerant Cecropia can have unusual ant associates (Davidson & Fisher, 1991). For example, "C. pungara" is inhabited, not by any of the Azteca species (subfamily Dolichoderinae) so wide- spread on Central and South American Cecropia, but by Pachycondyla luteola (Ponerinae) and Campono- tus balzani (Formicinae), and the typical associate of "Cecropia herrerensis" in northeastern Peru is Cre- matogaster aff. curvispinosa (Myrmicinae).
ANTS SYMBIOTIC WITH CECROPIA
Table 1 presents a list of obligate Cecropia-ants as we presently know them. Most impressive is their tax- onomic diversity. All ants belong to the same insect family, the Formicidae, and the major systematic cat- egories of ants are the subfamilies. The associates of Cecropia represent four ant subfamilies, including all but one of those in which at least some species are known to rely heavily on plant resources. Absent are only the Pseudomyrmecinae, stem-nesters found fre- quently in narrow twigs, where workers can effec- tively use their heads to block small entrances against invasions of competing and predatory ants.
The most common and best-known associates of Cecropia belong to the dolichoderine genus Azteca. This group is endemic to the New World, and its di- versity and abundance are greatest in the lowland tropics. Approximately 63 species and numerous named varieties remain in the genus after reconciling Brandao (1991) and Longino (1991) with Kempf (1972), and 13 of these species are specialists on Ce- cropia (Table 1). The genus includes many free-living species as well as specialized plant-ants associated with myrmecophytes in at least 16 genera and 9 fam- ilies (Davidson & McKey, 1997). Azteca ants tend to have populous colonies, finely divided into many small workers that tend Hemiptera (Coccidae and Pseudococcidae), scavenge, and hunt live prey, (usu- ally insects, and sometimes including competing ants; Carroll, 1983; Adams, 1990a). Based on nitrogen iso- topic ratios, various Azteca species place among the most predatory of arboreal ants (Davidson et al., 2003) and are the most nitrogen-deprived of neotrop- ical ants (Davidson, in press). Abundant carbohy- drates from hemipteran honeydew may fuel both the large worker populations, which are maintained prin- cipally by carbohydrates rather than by protein re- sources (Davidson, 1997), and the infamous aggres-
sion exhibited by these ants (Carroll, 1983). Monogynous (single-queen) colonies are often poly- domous, i.e., occurring as numerous affiliated nests distributed over one or more crowns. The large colony sizes of Azteca are correlated with low population densities, perhaps reflecting high mortality of incipi- ent colonies with comparatively few hemipteran as- sociates and thus limited protein resources. Wilson (1985 & pers. comm.) has argued that low population densities may be correlated with high local extinction rates that could account for the extinction of Azteca from West Indian islands, despite their presence there in fossil ambers from the late Oligocene or early Mio- cene.
Interspecific and intraspecific spatial territoriality have been demonstrated for a number of Azteca spe- cies (Adams, 1990a, 1990b, 1994; see also Leston, 1973, and Carroll, 1979) and may have preadapted members of this genus for defense of myrmecophytic host plants. Cecropia and other myrmecophytes pro- vide carbohydrates which fuel large, aggressive, and territorial worker populations (Davidson & McKey, 1993; Davidson, 1997). Azteca colonies subdivide la- bor by physical subcastes, with (mainly) major work- ers stationed on limbs at key branch intersections, at the bases of plants, and at colony borders (Adams, 1990a, 1990b, 1994). Defending workers frequently exhibit characteristic postures, with mandibles open and gasters upraised, ready to release carbon-based alarm pheromones should enemy ants approach. Rel- atively long-distance recruitment of nestmates (over meters) is apparently mediated in Azteca by trail pher- omones from the Pavan's glands. Signaling food finds and breaches in colony defense, short distance re- cruitment (over several centimeters) occurs in re- sponse to alert/alarm products (cyclic ketones) of the pygidial gland, as well as through worker-to-worker contact and tactile displays. Outcomes of territorial interactions appear to be determined by mutual as- sessment of asymmetries in colony size, mediated in part by recruitment responses that escalate with prox- imity to nests and nestmate reinforcements. With in- creased aggressiveness at or near nest sites, resident advantage can blunt advances of numerically superior colonies. Finally, because the traits summarized here occur in free-living Azteca species, they almost cer- tainly evolved in the context of territorial defense against other ants. Although many of the same traits undoubtedly preadapted Azteca to protect their host plants, arguments that such behaviors of Azteca evolved by selection on Cecropia for "induced de- fenses" (Agrawal, 1998) are currently not convincing because they ignore the basic biology of ants in this genus.
CECROPIA AND ITS BIOTIC DEFENSES 221
TABLE 1 Known or presumed obligate Cecropia-ants.a
Subfamily & Species Geographic Distribution Reference
DOLICHODERINAE Azteca spp. Longino, 1989, 1991 A. alfarib Central America through Amazonia A. aragua Aragua State, Venezuela A. australis W Amazonia (Peru, Bolivia), Amazonas, Brasil A. coeruleipennis S Mexico through NW Costa Rica A. constructor Guatemala through Guyana A. isthmica Panama and Colombia A. lattke Venezuela (coast ranges & Cordillera de Merida) A. merida E slopes, Cordillera de Merida, Venezuela A. muelleri S Brasil A. ovaticepsb Lowland tropics of Central and South America A. petalocephala SW of Coroico, (La Paz), Bolivia A. salti Sierra Nevada de Santa Marta, Colombia A. xanthochroa Mexico through Panama
PONERINAE Pachycondyla luteola W Amazonia Davidson et al., 1991 P prov. dianaec SE Panama W. L. Brown, in ms P ''insignis''c Mid-elevation wet forest, Costa Rica J. Longino 2000d
P unidentata rugulosae Emery 1902
FORMICINAE Camponotus balzani Amazonia Davidson et al. 1989, 1991;
Benson 1985
MYRMICINAE Crematogaster nr. curvispina Descent from E Andean cordillera in SE Peru D. W. Davidson, unpubl. Pheidole sp.
aOther ants may nest opportunistically in Cecropia stems without using prostomata or Miillerian bodies (Longino, 1989a; Davidson & Fisher, 1991). This phenomenon is especially common in plants with obligate associates nesting polydomously, only in growing tips. Polydomy, or fragmentation of single colonies over multiple nest sites (here, disjunct stems of a single host plant) is characteristic of Azteca alfari and A. ovaticeps (Longino, 1989b).
bMembers of the Azteca alfari group thought to have descended from ancestors that nested in live stems (Longino, 1989, 1991). The remaining Azteca species are proposed to be descendants of carton-nesting ancestors.
cThese names are conditional proposals (sensu Art. 15 of the 1985 ICZN) by the late W. L. Brown Jr., and thus not made available here. Their appearance here or in any duplication of this article does not constitute publication.
dSee Longino (2000) for natural history, distribution, etc. The species appears to be a specialist on sapling Cecropia, and colonies mature before saplings grow to tree size. Occupation of saplings by this species may lead to the demise of seedlings by precluding colonization by the more typical Azteca ants.
eThis variant of P. unidentata was synonymized with P. unidentata in an unpublished manuscript by W. L. Brown Jr., but sentiment remains among myrmecologists for considering it to be a specialist on Cecropia saplings.
Among the usually predacious and scavenging Po- nerinae, there appear to have been no fewer than three (and perhaps even four) independent evolutionary colonizations of Cecropia (Table 1). All of the po- nerines specialized to live in Cecropia are species in the genus Pachycondyla, relatively large-bodied ants occurring in both the New and Old World tropics. Most Pachycondyla species are terrestrial, but some
species are arboreal, nesting opportunistically in cav- ities of dead or live stems, or in and around epiphytes and debris, in the upper or lower rainforest canopies. Of the arboreal species, several are known to occupy abandoned intemodes of seedling and sapling Ce- cropia. Among the Pachycondyla specialized to live in Cecropia, Pachycondyla "insignis" appears to be related to P. villosa and P bugabensis, two general-
222 FLORA NEOTROPICA
ized arboreal predators (Longino, 2000). Pachycon- dyla luteola and Pachycondyla (provisionally) "dianae" (W. L. Brown manuscript), are morpholog- ically distinctive from one another and from Pa- chycondyla species nov. 2, as well as geographically disjunct in their distributions. Host-specific on "Ce- cropia pungara," Pachycondyla luteola departs most notably from other ponerines in behavior and colony size, typically just tens to hundreds of workers in the genus as a whole (Peeters, 1997). One might specu- late that hemipteran tending may occur in this species and provide carbohydrates to subsidize the large worker populations. However, the fearsome stings of these aggressive ants-painful over weeks to months-have deterred investigators from opening P luteola nests in all but small seedlings and saplings where Hemiptera are least apt to have colonized. Workers both prune encroaching vines and exhibit ab- solute fidelity to their host-plants, feeding principally on food rewards provided by the plant (Davidson & Fisher, 1991). As is typical of ponerines, queens do not found their colonies claustrally; instead they col- lect, store and use Muillerian bodies in the early es- tablishment of their colonies. Thus, they must leave the prostoma open in order to reach the trichilia, a practice that makes them susceptible to loss of brood to parasitoid wasps (Perilampidae, Davidson & Fisher, 1991). External foraging by nonclaustral queens (frequently multiple, competing queens per colony) may provide some protection to juvenile plants immediately after colonization and prior to col- ony establishment.
Formicines and Myrmicines are also represented among the associates of Cecropia. Camponotus bal- zani (subfamily Formicinae), a host generalist, is ex- tremely timid and largely nocturnal, and seldom ven- tures away from stems to patrol leaves (Davidson & Fisher, 1991). Another formicine, a species of Myr- melachista, is not listed in Table 1, because rather than collecting and feeding on Muillerian bodies, it appears to feed on the abundant queens of Azteca spp. that colonize a variety of Cecropia along the eastern escarpment of the Peruvian Andes (pers. obs.). Within the Myrmicinae, Crematogaster (Table 1) is a genus of ecologically dominant ants that are most diverse in the Old World tropics, where they are frequently ob- ligate plant-ants (Davidson & McKey, 1993). Their possibly late arrival in the New World tropics (Brown, 1973) might account for why small-bodied, stem- nesting and carton-building ants in the endemic neo- tropical genus Azteca were able to diversify and be- come codominant with this ecologically similar genus. The species associated with "Cecropia herrer- ensis" is most closely related to C. curvispinosa, a
generalized cavity nester, widespread in the Neotrop- ics (J. T. Longino, pers. comm.) and found occasion- ally in Cecropia at high elevations (pers. obs.). Al- though colonizing queens have occasionally been found with incipient colonies on "C. herrerensis," sometimes cofounding with conspecific queens, larger colonies are more often resident without their queens, which may either occupy trunks of adjacent Cecropia or live off the host altogether. Nevertheless, documentation of queen colonization and the collec- tion, storage, and use of Muillerian bodies suggests that the ants are specialized associates of Cecropia.
A third myrmicine, in the cosmopolitan genus Pheidole, is listed here on the basis of a single col- lection in Cecropia sp. at relatively high elevation (ca. 1500 m) on the descent from the easterm cordillera of the Andes in southeasterm Peru (D. Davidson, unpubl. data). Although the queen was not located, a large colony occupied numerous interuodes of a relatively tall plant, and was harvesting and storing Mullerian bodies near its brood. Until more colonies of the Phei- dole are located, and queens are shown to recognize and colonize these hosts, the lone record must be re- garded with suspicion. If the ant is eventually con- firmed as an obligate symbiont of Cecropia, the find- ing would be consistent with the pattern of novel associates occurring at the elevational and latitudinal limits of the distributions of myrmecophytes (David- son & McKey, 1993; McKey & Davidson, 1993).
Even among the predominant associates of Ce- cropia in the genus Azteca, there appear to have been multiple evolutionary colonizations of myrmecophy- tic hosts. Based on both morphological and behav- ioral evidence (Longino, 1991), and gene sequence data (Ayala et al., 1996), at least two independent line- ages of Cecropia-ants have been distinguished. One lineage, the Azteca alfari group (A. alfari and A. ova- ticeps), is likely derived from ancestors that nested in live stems. The other, the A. muelleri group (A. muel- leri plus A. aragua, A. australis, A. isthmica, A. pe- talocephala, A. salti, A. xanthochroa, and possibly A. constructor and A. merida), has probably descended from carton-nesters, and species in this lineage still build carton nests within their hosts. Carton nests con- sist of masticated plant material, sometimes mixed with ant or vertebrate feces and soil (reviewed in Lon- gino, 1986; Davidson & Epstein, 1989), all building materials that are available practically anywhere. Al- though the occurrence of stem-nesters may often be limited by the availability of suitable stems, nesting in carton frees ants to locate their nests near abundant food. Thus, the two Azteca lineages may have had very different histories of association with Cecropia. Carton-nesting ancestors of contemporary Cecropia-
CECROPIA AND ITS BIOTIC DEFENSES 223
ants may have initially sought out Cecropia because of its pearl bodies (see above), just as other carton- nesting taxa are sometimes overrepresented on hosts supplying extrafloral nectar (Hoildobler & Wilson, 1990; Davidson & Epstein, 1989). In contrast, stem- nesters would have been restricted to habitats where suitable hollow stems were available; wide stems with weak pith are characteristic of pioneer plants in dis- turbed habitats.
This scenario is consistent with experimental ev- idence sorting out the contributions of host specificity and habitat specificity to the determination of pairings between Cecropia and its ants. Yu and Davidson (1997) cultivated several Cecropia species from seeds in screen tents in Amazonian Peru, until all of the species had expanded their stems and produced pros- tomata. They then placed them out in two habitats, riverine edge and forest gaps (created or augmented) and monitored colonization over 2-3 months. Azteca ovaticeps, a member of the stem-nesting A. alfari group, proved to be habitat-specific (to riverine edge) and not host-specific, but the reverse was true of A. australis of the carton-nesting A. muelleri group. The relatively small thorax and reduced wing musculature of A. ovaticeps queens (Davidson et al., 1991) was consistent with its inability to seek out potential hosts in gaps dispersed throughout the forest. In contrast, with its large thorax, presumably correlated with a greater wing muscle mass, A. australis queens were successful at locating seedlings in both habitats. How- ever, in behavioral assays, queens preferred to colo- nize species with high pearl body production; they also were underrepresented among foundresses on hosts with low pearl body production and successfully produced brood during the experimental period only on the preferred hosts (Yu & Davidson, 1997). (The dearth of pearl bodies themselves is not likely to have caused failure of brood production, since the queens found their colonies claustrally.)
A parsimonious interpretation of the history of the relationships between the two ant lineages and their hosts might therefore suggest that the two were dif- ferently preadapted to use Cecropia, and that aspects of their preadaptations survive to influence their use of these myrmecophytes today. Thus, Azteca australis and its allies occur naturally mainly on hosts with high pearl body production (Davidson & Fisher, 1991; Yu & Davidson, 1997; Folgarait & Davidson, 1994, 1995). In contrast, A. ovaticeps and its sister species, A. alfari, are restricted to large-scale distur- bances throughout their ranges (Longino, 1989, 1991; Davidson & Fisher, 1991). Interestingly, in green- house experiments, A. alfari did not even feed on pearl bodies (Baird 1967).
This overview of Cecropia-ants suggests that Az- teca were likely the original associates of Cecropia. Not only are they the most diverse and widespread among the many symbionts of these ant-plants, but species in the Azteca muelleri group (including A. australis) may have built carton nests on these plants to use pearl bodies, even before Mullerian bodies had evolved. For this and other reasons, the muelleri group may have been first among the Azteca species to inhabit myrmecophytic Cecropia. In seeking out host plants with pearl bodies, queens of carton- nesting Azteca might have initially colonized larger plants, not seedlings, and it is difficult to believe that these foundresses could have initiated their colonies on plants with preexisting colonies of stem-nesting species. On the other hand, queens of stem-nesters may very well have gotten their start on Cecropia by colonizing seedlings that had not yet acquired other ants. As for the Cecropia-ants in other genera, these are almost certainly all derived from stem nesters, and with the exception of Camponotus balzani (wide- spread in the Amazon basin), are all species with lim- ited biogeographic and/or host ranges that suggest rel- atively recent origins of their relationships with Cecropia. A subset of these ant species, in the genera Pachycondyla and Crematogaster, also coexist with much more widespread and common species that are their likely progenitors (Table 1).
The richness and diversity of the ants symbiotic with Cecropia imply that colonization of Cecropia by unspecialized ants over evolutionary time has been a relatively easy transition. At least two factors could have facilitated this transition. First, for many ants with generalized diets, long-lived and secure nest sites, such as those afforded by cavities in live plants, may be more limiting than are food resources. Sec- ond, pachycaulous stems with weak pith probably evolved in Cecropia as a form of minimal support structure for large leaves (Halle et al., 1978; White, 1983; Davidson & McKey, 1993). Once evolved, these large stems were easily coopted as desirable and accessible nest sites for ants of diverse body sizes and colony sizes. In the context of the susceptibility of stems to exploitation by damaging herbivores (see above), such ants may have provided net benefits to their hosts and exerted selection pressures favoring the evolution of other ant-attractive traits.
COEVOLUTION OF ANTS AND CECROPIA
Coevolution consists of coadaptation and cospe- ciation (Brooks, 1979). Coadaption signifies that each species in the partnership has evolved one or more traits in response to selective pressures exerted by the other. There is little question that this has occurred in Cecropia and its ants, but since most Cecropia-ants
224 FLORA NEOTROPICA
are found on more than one host species, and many Cecropia species can be inhabited by more than one ant associate, coadaptation likely was diffuse. In dif- fuse coadaptation, selection pressures responsible for the evolution of a trait in one partner come from a set of associates, rather than from one species in partic- ular (Janzen, 1980).
What traits are indicative of coadaptation? In Ce- cropia, prostomata and trichilia with Mullerian bod- ies are surely characteristics that have evolved due to selection pressures exerted by symbiotic ants, and pearl bodies could have evolved in response to selec- tion imposed (originally) by opportunistic, as well as (later) symbiotic associates. Moreover, once these traits had evolved, there is evidence that they may have been modified by selection pressures generated by particular ant species (Davidson et al., 1991; Da- vidson & Fisher, 1991). Thus, the unusually large and protruding (rather than recessed) prostoma of "Ce- cropia pungara," together with its much larger Mill- lerian bodies, may be interpreted as adaptative re- sponses to its occupation by relatively large-bodied Pachycondyla and Camponotus species. Pachycon- dyla luteola is also unique among well-studied Cecropia-ants in its nutritional dependency on Mill- lerian bodies during the earliest stages of colony founding, and despite this dependency, P luteola queens refuse smaller Mullerian bodies transferred from other species of Cecropia (D. W. Davidson & P. Hererra, unpubl. data). Although composed of gly- cogen, as in congeneric species (F. Rickson, pers. comm.), the Muillerian bodies of "C. pungara" must be distinctive in some other way; based on superficial examination, they are unusually hard and less apt to degrade during storage. The Miillerian bodies of C. hispidissima appear to have evolved with Pachy- condyla prov. dianae, a species even larger-bodied than P luteola, and are larger and harder even than those of "Cecropia pungara." Other Cecropia, for ex- ample "C. grisita" (a name used for material not yet matched with any of the recognized Cecropia species) in southeastern Peru, have especially small Miillerian bodies (Folgarait & Davidson, 1994, 1995) or bodies to which obligate ants from different habitats react abnormally (Davidson & Fisher, 1991, for an Azteca ovaticeps colony placed on Cecropia ficifolia). Among the six common species co-occurring in southeastern Peru, "C. grisita" is the only one that never houses large-bodied ants, e.g., Camponotus bal- zani (Davidson et al., 1991), perhaps because of the unusually narrow stems at the time when seedlings are being colonized.
Other ant-related traits of Cecropia might have originated either through coadaptation with associ-
ated ants or as preadaptations, arising through alter- native selection pressures. For example, among the Cecropia species present in southeastern Peru, C. membranacea and "C. pungara" are distinctive in in- itiating Mullerian body production prior to the broad- ening of stems and the production of prostomata. Since the trait occurs in C. membranacea, on which Pachycondyla luteola queens fail to establish colo- nies, it is possible that it evolved in a different context and merely preadapted "Cecropia pungara" for use by this nonclaustral ant species (Yu & Davidson, 1997). The long, relatively sparse stem hairs of "C. pungara" could either be an adaptation to facilitate movements of its relatively large ant species (Pachy- condyla luteola and Camponotus balzani) or have provided a preadapted nest site where urticating hairs deterred potential predators of ant brood. Rates of production of Muillerian and pearl bodies, or leaves bearing both rewards, could either have coevolved with or been preadapted to colony growth rates of particular ant species. Pearl body production may be low in Cecropia membranacea (Folgarait & David- son, 1995) because its most frequent associate, Azteca ovaticeps, relies only weakly or not at all on these food rewards (Baird, 1967), or because of low nitro- gen levels in the frequently inundated habitats of this species. Glaucous stems, such as those that restrict climbing by all but the most obligate ant associates of some myrmecophytic Macaranga (Federle et al., 1997), occur as a developmental stage (D. W. David- son, unpubl. data) or polymorphism in some Cecropia species. While this trait might have evolved to limit access to all but beneficial ants, it could also have originated as a defense against insect herbivores. (Re- cent evidence [D. W. Davidson et al., unpubl. data] suggests the latter for Cecropia vs. strigosa.) Finally, selection to enhance the integrity of the intemodal septa inside Cecropia stems (thicker and harder in myrmecophytes than in nonmyrmecophytes: see Bai- ley, 1922) could occur either for structural reasons or to allow for competition among multiple foundresses and their incipient colonies. By assuring that multiple colonies of one or more species can develop in iso- lation from one another, the plant should both in- crease its chances for successful establishment of at least one colony, and help to magnify selection among competitors for rapid colony development rates.
Among ants, coadapted traits resulting from as- sociation with one or more Cecropia species would include the recognition and use of both prostomata and Muillerian bodies by queens and workers. In both the introduced (Wetterer, 1997) and native ranges of Cecropia (Davidson & Fisher, 1991), ants that are not
CECROPIA AND ITS BIOTIC DEFENSES 225
obligate Cecropia associates tend not to recognize Miillerian bodies as food. Those in the introduced ranges have also not been observed to use prostomata to nest in Cecropia stems (Putz & Holbrook, 1988; Wetterer, 1997). In contrast, queens of obligate as- sociates in the Azteca alfari group appear to have evolved head shapes that facilitate stem entry at pros- tomata (Longino, 1989b). Facing high risk of preda- tion during colony founding, queens of obligate Cecropia-ants should also have evolved to locate their hosts quickly from a distance, by responding to pos- sible chemical cues elaborated by one or more hosts. At closer range, queen transfer experiments have shown that both Azteca australis and Pachycondyla luteola exhibit strong preferences for host species on which brood production is most successful (Yu & Da- vidson, 1997, and above). The latter species colonizes just "Cecropia pungara," the only species on which this ponerine ant succeeds in establishing colonies, and C. membranacea, believed to be a very close rel- ative (Yu & Davidson, 1997).
Finally, many Cecropia-ants attack and prune vines and other vegetation that contacts their host plants (Janzen, 1969; Davidson et al., 1988). This be- havior, present in the associates of many myrmeco- phytes, reduces access to the plants by competing and predatory ants (Davidson et al., 1988). Occurring among Cecropia-ants in (at least) a number of Azteca species (e.g., Janzen, 1969) and in Pachycondyla lu- teola (Davidson et al., 1988), the behavior may be an aspect of territoriality that evolved either before the relationship with Cecropia, or as longer colony life- times became possible in the long-lived and relatively protective nesting environments provided by myr- mecophytes.
Despite abundant evidence for coadaptation, there is little support for cospeciation or cocladogenesis in either Cecropia or its ants. Thus, although particular associates may be highly species-specific (e.g., Pachycondyla luteola on "Cecropia pungara," Pachy- condyla sp. nov. 1 on Cecropia hispidissima, and Crematogaster aff. curvispina on "Cecropia herrer- ensis"), no substantial radiation of partner lineages appears to have occurred through cocladogenesis. Nevertheless, cospeciation cannot be firmly ruled out, and evidence might yet be found as relationships of Azteca and Cecropia come under greater scrutiny.
EVOLUTIONARY COLONIZATION, HOST SHIFTS, AND HABITAT SHIFTS
De novo or evolutionary colonization is the alter- native to cocladogenesis in magnifying the diversity of Cecropia and its ants over evolutionary time. As
described above, the transition from free-living ants to symbiotic associates of Cecropia has occurred fre- quently and apparently easily, and accounts for much of the diversity in these symbionts (Table 1). More- over, after these species were evolutionarily commit- ted to Cecropia, the combination of habitat shifts in plants and host switches in ants would have further enhanced the diversity of ant-plant partnerships. In turn, the facility with which Cecropia spp. acquired new ant partners may have permitted the genus to attain high species diversity through frequent evolu- tionary habitat shifts, often correlated with changes in growth rates and defensive investment (Davidson & Fisher, 1991; Folgarait & Davidson, 1994, 1995). Although many lowland rain forest Cecropia species require some sort of light gap for establishment, spe- cies differ widely in their light requirements, as well as characteristic soil types. As exemplified by species encountered in southeastern Peru, some are tightly bound to riparian edge (e.g., C. latiloba and C. en- gleriana), others to frequently inundated soils (C. membranacea, "C. pungara," and C. utcubambana), some to less fertile terra firme (C. ficifolia, C. poly- stachya, and "C. grisita"), and still others to high- elevation cloud forests along the eastern Andean cor- dillera (C. angustifolia and C. tacuna). In their comparison of ant-plant relationships in Africa and the Neotropics, McKey & Davidson (1993) argued that a fine-scale habitat mosaic, created by Andean orogeny and rivers meandering widely and at differ- ent elevations over geologic time (Salo et al., 1986), has been profoundly important in magnifying diver- sity within a number of ant-plant taxa, including Ce- cropia, and in the partnerships of habitat-shifting plant species with novel ant taxa. Thus, compared to Africa, where geomorphology is more monotonous, the Neotropics have 3.5-fold greater species richness of plant-ants, despite just 1.3 times as many total ant species.
Yu & Davidson (1997) have attributed to the in- dependent (horizontal) dispersal of symbionts both the predominance of de novo colonization over co- speciation in the Cecropia-ant system and the diver- sity of mechanisms associated with evolutionary col- onization and species-specificity. Thus, rather than being codispersed with their partners, as is the case in many symbiotic associations between mutualists, or between parasites and hosts, colonization of Cecropia seedlings and saplings occurs anew each generation, allowing much opportunity for evolutionary coloni- zation and host shifts that may, in turn, inhibit pair- wise coevolution and cocladogenesis. In this circum- stance, historical coincidences such as coordinated or exclusionary dispersal (to the same or different hab-
226 FLORA NEOTROPICA
itats, respectively) may be as important to, or more important than, competition for partners in setting the stage for future coevolution. Moreover, pairings be- tween Cecropia and its obligate associates are ex- plained largely by the effects of local propagule pools, queen preferences and host locating abilities, and col- ony performances on young seedlings and saplings, i.e., by events early in the life histories of these as- sociations (Yu & Davidson, 1997).
Almost certainly, given the species richness of Cecropia and the widespread distribution of this important neotropical genus, additional ant associ- ates remain to be discovered. Analysis of these new species, together with continued progress in recon- structing the phylogenies of both Azteca and Cecro- pia with molecular and other characters will provide new and informative tests of the ideas developed here.
INDEX OF SCIENTIFIC NAMES 227
INDEX OF SCIENTIFIC NAMES
Synonyms are in italics. Names in [brackets] are nomina nuda or doubtful names. Names between "quo- tation marks" are names for Cecropia material with uncertain or unknown identity and used in studies on ants. Page numbers with an asterisk (*) indicate pages with illustration or maps.