BACTERIOLOGICAL REVIEWS, Sept. 1972, p. 263-290 Copyright @ 1972 American Society for Microbiology Vol. 36, No. 3 Printed in U.SA. Cell Biology of the Mycoplasmas JACK MANILOFF AND HAROLD J. MOROWITZ Department of Microbiology and Department of Radiation Biology and Biophysics, The University of Rochester, Rochester, New York 14642, and Department of Molecular Biophysics and Biochemistry, Yale University, New Haven, Connecticut 06520 INTRODUCTION ........................... 263 CYTOLOGY .................................. 264 Cell Size ........................... 264 Cell Morphology ........................... 265 Cell Growth .265 SUBCELLULAR ORGANIZATION ....... .................... 268 Cell Membrane ................................ 269 Biochemical properties ........................... 269 Biophysical properties ........................... 272 Reaggregation studies ........................... 275 Extracellular Aspects ........................... 276 Extracellular structures ........................... 276 Extracellular products ........................... 277 Chromosome ........................... 278 Genome structure ........................... 278 Replication ................................ 279 Genetics ................................. 279 Ribosomes and RNA ........................... 280 Ribosomes and rRNA ........................... 280 tRNA ........................... 282 mRNA ........................... 282 Plasmids ........................... 283 Other Cytoplasmic Organelles ...... ..................... 283 SUMMARY ........................... 284 LITERATURE CITED ........................... 284 INTRODUCTION The term mycoplasma is the name generally used for a group of microorganisms (order Mycoplasmatales), earlier called the pleuro- pneumonia-like organisms, or PPLO. This group lacks cell walls and includes the genus Mycoplasma (which requires sterol), the genus Acholeplasma (which does not require sterol), and the T strains (which require urea). A re- cent thermophilic, acidophilic isolate has been proposed as a new genus, Thermoplasma (29), but since this group has not been extensively characterized, it will not be considered further here. The extensive data concerning the myco- plasmas, which have been reported over the past few years, is now sufficient to allow us to contruct an overview of the cellular and mole- cular biology of these organisms. Therefore, this paper will be critical rather than exhaus- tive, since more encyclopedic reviews are available (e.g., 57, 126, 156, 166) and it is now recognized that many earlier published results reflect serious preparation artifacts. Several generalizations can be derived from a critique of the existing literature in terms of current criteria of validity, particularly the necessity for monitoring the viability of the culture being studied to make sure that bio- chemical and cytological data are collected on exponentially growing cells and not on the debris found in older, dead cultures. The first generalization is that the myco- plasmas are a sufficiently heterogeneous group of organisms that various species cannot be expected to be described by a single type of morphology. In fact, since the mycoplasmas are a group of small procaryotic cells bounded only by a single lipoprotein ("unit") mem- brane, they may well represent a group of mi- croorganisms as diverse as the bacteria. The second generalization is that, under defined culture conditions, each species has a charac- teristic morphology. In cytological prepara- tions, the normal plasticity (polymorphism) arising from the absence of a rigid surface en- velope should not be confused with pleomorph- ism, which would necessarily imply the exist- 263 Downloaded from https://journals.asm.org/journal/br on 24 December 2021 by 39.111.40.240.
Transcript
BACTERIOLOGICAL REVIEWS, Sept. 1972, p. 263-290Copyright @ 1972 American Society for Microbiology
Vol. 36, No. 3Printed in U.SA.
Cell Biology of the MycoplasmasJACK MANILOFF AND HAROLD J. MOROWITZ
Department of Microbiology and Department of Radiation Biology and Biophysics, The University ofRochester, Rochester, New York 14642, and Department of Molecular Biophysics and Biochemistry, Yale
INTRODUCTIONThe term mycoplasma is the name generally
used for a group of microorganisms (orderMycoplasmatales), earlier called the pleuro-pneumonia-like organisms, or PPLO. Thisgroup lacks cell walls and includes the genusMycoplasma (which requires sterol), the genusAcholeplasma (which does not require sterol),and the T strains (which require urea). A re-cent thermophilic, acidophilic isolate has beenproposed as a new genus, Thermoplasma (29),but since this group has not been extensivelycharacterized, it will not be considered furtherhere.The extensive data concerning the myco-
plasmas, which have been reported over thepast few years, is now sufficient to allow us tocontruct an overview of the cellular and mole-cular biology of these organisms. Therefore,this paper will be critical rather than exhaus-tive, since more encyclopedic reviews areavailable (e.g., 57, 126, 156, 166) and it is nowrecognized that many earlier published resultsreflect serious preparation artifacts.
Several generalizations can be derived froma critique of the existing literature in terms ofcurrent criteria of validity, particularly thenecessity for monitoring the viability of theculture being studied to make sure that bio-chemical and cytological data are collected onexponentially growing cells and not on thedebris found in older, dead cultures.The first generalization is that the myco-
plasmas are a sufficiently heterogeneous groupof organisms that various species cannot beexpected to be described by a single type ofmorphology. In fact, since the mycoplasmasare a group of small procaryotic cells boundedonly by a single lipoprotein ("unit") mem-brane, they may well represent a group of mi-croorganisms as diverse as the bacteria. Thesecond generalization is that, under definedculture conditions, each species has a charac-teristic morphology. In cytological prepara-tions, the normal plasticity (polymorphism)arising from the absence of a rigid surface en-velope should not be confused with pleomorph-ism, which would necessarily imply the exist-
263
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/b
r on
24
Dec
embe
r 20
21 b
y 39
.111
.40.
240.
MANILOFF AND MOROWITZ
ence of more than one distinct cellular formduring the organism's life cycle. The thirdgeneralization is that the smallest knownviable mycoplasmas (single cells capable offorming a clone) are about 0.33 Atm in diameter(85). These are the smallest reported livingcells. It would be very interesting if smallercells are found some day, but the smallestmycoplasms are within an order of magnitudeof the 0.1 gm theoretical minimal cell (100,101). Recent reevaluation of the theoreticalminimal cell (discussed below) raises this pos-tulated value to about 0.15 Am. The fourthgeneralization is that most mycoplasmas repli-cate by a binary division (i.e., one cell formstwo daughter cells), while some filamentousspecies are probably able to break into a smallnumber of viable filamentous daughter cells.More elaborate life cycles have previously beenpostulated for the mycoplasmas. In the mostwidely known of these it was assumed that anindividual cell enlarged giving rise to a "largebody." Within this structure "elementary bod-ies" formed. These "elementary bodies" werebelieved to be membrane-bounded virus-likeparticles of dimensions between 0.10 and 0.15Am. The large cell was assumed to rupture,releasing the "elementary bodies" each ofwhich proceeded to grow into cells which couldthen either replicate by binary fission or ma-ture into "large bodies" (40, 41, 42, 74, 107).No currently acceptable data support such atheory and no viable "elementary body" hasever been isolated. The fifth generalization isthat the biochemistry of the mycoplasmas issimilar to the biochemistry of other cellularsystems; thus the unique molecular propertiesin terrestrial biology that biologists had origi-nally hoped to find in these small cells havenot been found (e.g., 100, 101, 115, 116). Thepossible significance of this point will be dis-cussed below.
CYTOLOGYThe lack of viability data in many references
makes it impossible to know the growth phaseof most of the cultures that have been studied.This is important because, as mycoplasma cul-tures age, there are morphological changes (13,68, 87), loss of enzyme and transport activities(126), decrease in membrane fluidity (152),increase in osmotic fragility (152), and resist-ance to lytic viral infection (79).
It should be noted that turbidity measure-ments (unless explicity shown otherwise) arenot an adequate viability assay since absorb-ance has been shown to be a nonlinear func-tion of the number of viable cells (86, 164).
"Good turbidity" generally corresponds to sta-tionary-phase cultures (69, 70, 86). In the dis-cussion below, emphasis will be given to thosestudies that clearly involve exponentiallygrowing cells.
Cell SizeThe volume of individual mycoplasma cells
is equivalent to spheres in the size range of0.33 to 1.0 gm diameter. The sizing involves asimplification because, for example, the 0.9-,um spherical Acholeplasma laidlawii cell isseen only in dying cultures; in growing cul-tures the cell is usually a 0.5-,gm by 2-,gm fila-ment, which has the same volume as the 0.9-,um sphere (87). Some degenerating cells havebeen reported to contain or release smallbodies which have been called "elementarybodies," but no evidence exists that these areviable particles (2, 87).The 0.33-Am cell (85) is about three times
larger than the early reports of 0.1-um clone-forming units, and the reasons for the erro-neous size estimates must be considered. Oneof the techniques used to estimate cell size waspassage through known pore-size filters (74,109). Razin (126) has reviewed studies indi-cating that the plastic mycoplasmas can besqueezed through filter pores smaller than theactual cell size, and Lemcke (77) has recentlyexamined the filtration conditions that allowmycoplasmas to pass through filter poressmaller than the cell diameter. It has also beenreported that staphylococcal L-forms withdiameters greater than 0.6 lim, when grown onfilters, are able to pass through filter-pore sizesas small as 0.05 gm (99). In experiments on A.laidlawii cells, which are 0.5 Am by 2 um, theauthors have found (unpublished data) that,with positive pressure (using a Swinney hypo-dermic adapter), 10% of the titer can be re-covered in a 0.45-Mim pore-size filtrate. How-ever, no titer was recovered when the filtrationwas done with negative pressure (using a wateraspirator). Hence, filtration efficiency of themycoplasmas depends on the conditions andcannot be used as a size measure, as can bedone with rigid virus particles.The small size of the mycoplasmas puts
them close to the 0.25-am limit of resolution ofoptical phase-contrast microscopy (e.g., 87). Inaddition, the optical measurements are com-plicated by: (i) the "halo" artifact produced bythe phase-contrast optics (9) and (ii) the na-ture of the phase-contrast image formation inwhich the size of a circular object appearssmaller than the true object (21, 145). Hence,it must be concluded that measurements in
264 BACTrERIOL. REV.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/b
r on
24
Dec
embe
r 20
21 b
y 39
.111
.40.
240.
CELL BIOLOGY OF MYCOPLASMAS
this size range, based on optical microscopy,are inaccurate and, in a comparison with lightand electron micrographs, it was not alwayspossible to decide whether a light microscopyimage represented a single cell or a clump ofcells (87). Bredt (15-17) has been able to solvethis problem by using time-lapse photographyto follow the growth of the individual viableunits in his samples.
Sizing by electron microscopy, because ofthe difficulties described above, lacks the im-portant correlative data that can be obtainedin other systems by light microscopy observa-tions of living cells. Razin (126) has discussedthe problems inherent in electron microscopicsize measurements. In general these are: (i) forthin sections, dependence of the size on theplane of section; (2) for negative stain prepara-
tions, morphological distortions caused by theheavy metal stains (to be discussed furtherbelow); and (3) for electron microscopy in gen-
eral, the difficulty in knowing whether the par-ticle observed represents a viable cell. The lastproblem can be approached by studying cellsgrowing on electron microscope grids (85, 91,105), in which case each microcolony must rep-resent a viable clone.
Cell MorphologyThe limitations of light and electron micros-
copy observations discussed above must bekept in mind in evaluating reports on cellmorphology. In addition, it has been shownthat mycoplasma shape can be vastly alteredand deformed by the physical and chemicalagents used in the preparations. For example,the pear-shaped Mycoplasma gallisepticumforms: (i) spheres, if centrifuged before fixation(93), (ii) filaments, if exposed to hypotonic so-
lutions (11), (iii) aberrant cells having many
blebs, if grown on a surface (91), and (iv) a va-
riety of distorted shapes, if unfixed cells are
negatively stained with phosphotungstic acid(10, 93). Phosphotungstic acid has also beenreported to alter the spherical shape of unfixedcells of other mycoplasmas (141, 142), pro-
ducing bizarre forms with many protrusionsand filaments, and causing the detachment ofsmall protrusions giving rise to what could befalsely interpreted as small cellular particles.Presumed enzymatic deoxyribonucleic acid(DNA) degradation during centrifugation ofunfixed cells has also been reported in M.mycoides (63) and M. gallisepticum (143), re-
flecting certain cytological alterations duringcell harvesting. Filament production in hypo-tonic solutions has also been reported for M.pulmonis (141). One should distinguish be-
tween filaments as a morphological form re-sulting from growth which has been clearlydemonstrated in phase microscopy, and fila-mentous protrusions resulting from effects ofphysical and chemical agents, both of whichmay appear in a given species.A detailed review of all of the published
papers on morphological forms seen in myco-plasma cultures would indeed be an extensivetask. From our survey of those reports showingthe best cytological preservation, the morpho-types may tentatively be grouped into the fol-lowing categories: (i) coccoid cells, e.g., M.pulmonis (64, 141) and Caprine PPLO strain14 (J. Maniloff, 1968. Bacteriol. Proc., p. 78),the latter forms streptococcal-like chains (Fig.1); (ii) coccoid cells with membrane tubules,which frequently appear as tubule-connecteddiplococci (Fig. 2), e.g., M. arthritidis H39 (85),originally erroneously classified as a M. hom-inis and possibly M. felis (13); (iii) filamentouscells, sometimes showing branching (Fig. 3),e.g., M. orale (3), M. hominis (98), A. laidlawii(87, 129), and M. mycoides (41, 49); (iv) fila-mentous cells with terminal structures, theonly known example of which is M. pneumo-niae (12, 68); and (v) pear-shaped cells withterminal structures, the only known examplesof which are the blebs of the strains of M. gal-lisepticum (Fig. 4; references la, 10, 91, 93,105, 182). The morphology of the T strains hasstill not been adequately described. This clas-sification does not imply that a species has aunique shape under all growth conditions, butrather asserts that under given growth condi-tions the morphological character is distinctenough to be classified into one of a smallnumber of forms.These morphologies characterize the expo-
nentially growing mycoplasmas. There is noevidence that mycoplasmas can retain theirviability if taken up in an intracellular form;they can be phagocytized by animal cells (64,182, 183). On the other hand, the studies ofplant and insect parasites presumed to bemycoplasmas have demonstrated intracellularparticles, and it is not possible at the presenttime to generalize about their possible mor-phological forms (e.g., 22, 51-53, 83, 84, 94,112, 118, 119, 157).
Cell GrowthWhen colony-forming units are assayed,
mycoplasma cultures follow typical bacterialgrowth curves with exponential, stationary,and death phases (43, 45, 69, 70, 80, 86, 129,180). Mycoplasma colonies have been shown toform from single cells for M. gallisepticum
265VOL. 36, 1972
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/b
r on
24
Dec
embe
r 20
21 b
y 39
.111
.40.
240.
MANILOFF AND MOROWITZ
(105) and M. arthritidis H39 (unpublished re-sults of the present authors, using same experi-mental protocol as reference 105). In the expo-nential phase at 37 C, most cultures have adoubling time in the range of 1 to 6 hr (86,166). The length of the stationary phase variesamong the species (86, 129) but is frequentlyshorter than 24 hr. The subsequent death ratesare quite rapid; the titer drops by a factor of102 to 104 per hr (80, 86).The few studies of macromolecular nucleic
acid synthesis during culture developmentshow growth patterns similar to those found inbacterial cultures (58). When stationary-phaseM. gallisepticum cells are transferred to freshmedium, there is an exponential increase incell titer but a lag in DNA synthesis, resultingin a decrease in DNA per cell. ExponentialDNA synthesis begins and continues while thecell number begins to level off as the culturereaches stationary phase, resulting in an in-crease in the DNA per cell (122). These data,together with the multi-hit ultraviolet inacti-vation studies (discussed below), indicate thatstationary-phase cells may be multinucleate.The ribonucleic acid (RNA)/DNA ratio in ex-ponential-phase M. gallisepticum is reportedto be 4.5 (73) and for stationary-phase cells tobe 2.0 (110).The problem of replication in the myco-
plasmas can now be restated in somewhat dif-ferent terms. The fundamental act of replica-tion in these cells and all other procaryotes isthe replication of the DNA genome, a singlemacromolecule which usually occurs in a loopconfiguration. The two DNA structures mustthen be packaged into separate nuclear bodies.If the cell then divides into two nearly equalhalves, each containing one nuclear body, theprocess is designated binary fission. An une-qual cytoplasmic division at this stage wouldappear as a budding process. Alternatively,several nuclear divisions and cellular elonga-tion followed by cell division would constitutereplication by filament formation. In any case,there is a primary process of nuclear divisionfollowed by secondary cell division. The de-tails of the cell division have not been clarifiedin all mycoplasma strains.The growth of exponential-phase cells has
been followed by the beautiful cinemato-graphic studies of Bredt (15-17) and is inagreement with the interpretations of well-preserved cells from growing cultures exam-ined by electron microscopy. Binary fission isthe characteristic mode of replication and hasbeen observed for M. gallisepticum (91, 105),M. hominis (17), M. pulmonis (W. Bredt, 1972,
Med. Mikrobiol. Immunol.; 157:169), M. pneu-moniae (15, 16), M. arthritidis (85), and Cap-rine PPLO strain 14 (J. Maniloff, 1967, J. CellBiol. 35:87A). Bredt (1972, Med. Mikrobiol.Immunol. 157:169) has also found that some ofthe filamentous cells (e.g., M. hominis), inaddition to binary divisions, can elongate andconstrict at a couple of points giving rise toseveral viable filamentous cells.There are reports of cell numbers doubling
in cultures which are supposed to be synchro-nized, of M. orale (43), M. pneumoniae (45),and two avian PPLO strains (71). These areexpected for binary division as the mode ofcell replication, but this agreement may befortuitous in view of the difficulties in ob-taining high degrees of synchrony, the problemof reproducibly counting mycoplasma colonies,and, in the case of the M. pneumoniae study,the long time interval used makes it difficultto decide whether there are indeed plateaus inthe growth curve.As was mentioned above, there are no cur-
rently acceptable data supporting the exist-ence of "elementary bodies" or an endomy-celial-endofilamentous theory of mycoplasmagrowth. The experiments that were interpretedto suggest such forms appear, in hindsight, tohave suffered from the various artifacts dis-cussed above relating to sizing, preparation formicroscopy (including fragmentation of cellsduring preparation), and morphologicalchanges in drying cultures. Three furtherpoints should be noted here. (i) If "elementarybodies" exist, it should be possible because ofthe nature of phase-contrast light microscopyto observe colonies growing from no preex-isting image, since the hypothetical "elemen-tary body" would be below the limit of resolu-tion. No such observations have been reported.(ii) The report of "elementary body" formationin stationary-phase cultures (129) cannot re-flect a growth phenomenon since the viabilityof such cultures is constant or slightly de-creasing. (iii) One further argument may beoffered against the logical possibility of a stageof the mycoplasma life cycle of a size of 0.15,um or less. We know that the smallest myco-plasma genomes are of the order of 5 x 108daltons (102). Assuming minimal hydration (1g of water/g of DNA), such a molecule cannotbe packed into a membrane-bounded sphere ofless than 0.15 ,um external diameter. Any othernecessary cellular apparatus would raise thisminimum. Conservation of information de-mands that the cell retain its genome over allstages of the life cycle. Therefore, reportedviable mycoplasma units of less than around
266 BACTrERIOL. REV.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/b
r on
24
Dec
embe
r 20
21 b
y 39
.111
.40.
240.
CELL BIOLOGY OF MYCOPLASMAS
,-Aa0.m..r.42*C_*.w
FIG. 1. Micrographs of sections of Caprine PPLO strain 14 logarithmic phase cells; glutaraldehyde fixed,Epon embedded, and uranyl stained (87). The cells form streptococcal-like chains. The cytological elementsseen are the cell membrane, fibrous nuclear material, and ribosome granules. x81,000.
VOL. 36, 1972 267
: ~. , c4
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/b
r on
24
Dec
embe
r 20
21 b
y 39
.111
.40.
240.
MANILOFF AND MOROWITZ
0.2 Asm diameter are not possible on the basisof current knowledge.
SUBCELLULAR ORGANIZATIONThe view of mycoplasma cytology has
changed greatly since Morowitz and Tourtel-lotte (107) showed a schematic representationof a typical PPLO cell as a round bodybounded by a lipoprotein membrane and con-taining disorganized DNA, ribosomes, and sol-uble RNA. The known subcellular structuresnow consist of the cell membrane, possibleextracellular structures, the chromosome, ribo-somes and macromolecular RNA, plasmids,
and other cytoplasmic organelles.Some mycoplasmas have been found to have
special cytological features that relate to theirinteraction with animal cells: the capsularsubstance around M. mycoides (49), the ter-minal bleb on M. gallisepticum (182, 183), andthe terminal knoblike structures of M. pneu-moniae (28). These mycoplasmas, perhapscoincidentally, are among the more pathogenicspecies. The M. neurolyticum exotoxin shouldprobably be included as a special case; the cellmust certainly have an interesting method ofexporting a toxin with a molecular weight over200,000 (172).
1.E
f'
FIG. 2. Section of M. arthritidis H39, formerly named M. hominis H39 (85). Elongated coccoid cells and adividing cell, with the coccoid cells connected by a membrane tubule, are seen. x81,000. Inset: Detail of partof a membrane tubule between two cells. x162,000.
268 BACTERIOL. REV.
A 5%MT
ajVk
Iq ., - r,,6.,
I IN.
I
0 w11
.'. .. f-&C.-AL -.1--rio.-
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/b
r on
24
Dec
embe
r 20
21 b
y 39
.111
.40.
240.
CELL BIOLOGY OF MYCOPLASMAS
.4.-VI..%. A.
t.t.1 olip,. .1
;t ...;".1
I.
cells (87). Large cytoplasmic granular particles can be seen in the fila-
All electron microscopy studies that havebeen done confirm the absence of any mem-
braneous organelle, except for the boundingcell membrane (see references above and 124).This, plus the absence of other observable sur-
face envelope layers, has allowed the prepara-tion of membranes for further study by gentleprocedures (e.g., osmotic shock), and thesefractions are relatively uncontaminated, i.e.,they are of a single membrane type and haveno adhering extracellular components.Biochemical properties. The chemical
composition of mycoplasma membranes is
reported as 50 to 59% protein, 32 to 40% lipid,0.5 to 2% carbohydrate, 2 to 5% RNA, andabout 1% DNA (124, 127). The RNA and DNAare regarded as cytoplasmic contaminants andcan be removed by washing and nucleasetreatment (124, 126, 127). The carbohydratecomponent may be significantly underesti-mated, since Engelman and Morowitz (36)have shown that the analytical method useddoes not detect glucosamine and galactosa-mine, which are present in rather largeamounts (about 5%) in A. laidlawii mem-
branes. Morowitz and Terry (106) indicatedthat the hexosamine is in long-chain polysac-charides. The glycolipid and protein mem-
FIG. 3. Section of A. laidlawii Bmentous cells (arrows). x40,500.
Cell Membrane
VOL. 36, 1972 269
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/b
r on
24
Dec
embe
r 20
21 b
y 39
.111
.40.
240.
MANILOFF AND MOROWITZ
FIG. 4. Sections of exponentially growing M. gallisepticum A5969 (105). Both one-bleb daughter cells andtwo-bleb predivision cells are seen. The subcellular organelles include the cell membrane, nuclear material,ribosomes, bleb, and infra-bleb region. x81,000.
brane components specify the mycoplasmaantigenic properties (5, 65, 138, 139).Polyacrylamide gel electrophoresis of the
urea-phenol-acetic acid-solubilized membraneproteins reveals 20 to 30 bands (125, 131, 148,171). Since not all of the proteins enter thegels, the actual number of membrane proteins
may be larger. The band patterns tend to besimilar for strains within a species and thespecies differences in gel patterns correspondto known serological and nucleic acid ho-mology differences. The membrane amino acidcompositions of the mycoplasmas that havebeen studied are summarized in Table 1. Cor-
270 BAC rRIOL. REV.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/b
r on
24
Dec
embe
r 20
21 b
y 39
.111
.40.
240.
CELL BIOLOGY OF MYCOPLASMAS
TABLE 1. Amino acid composition of membranes
|M. gallsp- A. Iaidlawii A. laidlawii A. lazidlawii Erythro- B. mega-Amino acid ticuma A (mole %) Bc (mole %) Bd (mole %) cytese terium(mole %) (mole %) (mole %)
a Cells grown on Tryptose medium (110).'Cells grown on peptone-yeast extract medium (24).c Cells grown on Tryptose medium (36).d Cells grown on beef-heart infusion medium (106).eAverage of four preparations cited by Rosenberg and Guidotti (144).' Data for Bacillus megaterium protoplast membranes (181).
responding data for erythrocyte and Bacillusmegaterium (as a representative bacterium)membranes are included for comparison. Themycoplasma values are not appreciably dif-ferent from the erythrocyte and bacterial data,but the latter do show some variation inacidic, basic, and uncharged residues, and theerythrocytes have a 4- to 10-fold higher cys-teine content.The biochemistry of the membrane lipids
has been recently reviewed by Smith (165, 166)and Razin (126, 127) and hence only certaintopics will be briefly summarized here. Most ofthe mycoplasmas seem to require fatty acidsfor growth (166); in addition, the species of thegenus Mycoplasma require cholesterol, andseveral Mycoplasma strains also require glyc-erol (126, 127). The major membrane lipid con-stituents are phospholipids and glycolipids. Inthe Mycoplasma species, cholesterol makes up12 to 30% of the total lipid (4). Although theAcholeplasma do not require cholesterol, ifgrown in a cholesterol-containing medium theyincorporate it as 3 to 4% of their total lipid (4).A. laidlawii also contains four major carot-enoid pigments (166) which comprise at most1% of the lipid (126, 127).In a study of nongrowing A. laidlawii, Ka-
hane and Razin (66) showed that, at least in
these cells, membrane lipid and proteinsyntheses are not synchronized. Unfortunately,because of the state of their cells, the data onturnover rates of proteins and lipids cannot beapplied to actively growing cells.When A. laidlawii is grown in media supple-
mented with various fatty acids, includingnon-biologically occurring compounds, theexogenous fatty acid is incorporated into themembrane and can comprise 60 to 80 mole percent of the total fatty acids (81, 114). This al-lows the lipids to be varied, to examine therelationship of the lipid composition to mem-brane properties. These data indicate therange of fatty acids that can be incorporatedand constitute a functional cell membrane,and also show the relative lack of specificityrequired for the membrane lipid components.Smith (166) has reviewed the reports of the
various biochemical activities which have beenshown to be associated with the membrane:redox enzymes (including quinones and cyto-chromes in some species), adenosine triphos-phatase, nucleases, cholesterol esterase, andseveral lipid synthetases. In addition, a mem-brane lysophospholipase (177) and peptidase(25) have been found. Rottem and Razin (147)found that the A. laidlawii adenosine triphos-phatase hydrolyzes other nucleotide triphos-
VOL. 36, 1972 271
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/b
r on
24
Dec
embe
r 20
21 b
y 39
.111
.40.
240.
MANILOFF AND MOROWITZ
phates and hence is probably a nonspecificmembrane nucleotide triphosphatase (nucleo-tide monophosphates were not hydrolyzed). Inaddition, the triphosphatase was not K+- orNa+-activated and was not affected by oua-bain. The pattern of membrane-bound versussoluble enzymes varies among the myco-plasmas (120, 121); e.g., while adenosine tri-phosphatase activity is localized in membranesubcellular fractions, NADH2 oxidase activityhas been found in the soluble fractions of theMycoplasma examined but in the membranefraction of A. Iaidlawii.
Several membrane transport processes havebeen observed in the mycoplasmas. Rottemand Razin (147) showed that K+ and Na+transport in A. laidlawii are not coupled andare independent of the adenosine triphospha-tase activity in the cell. Using the same spe-cies, Cho and Morowitz (23) found that K+transport is energy-dependent and, since it isinhibited by sulfhydryl blocking agents, isprobably mediated by cysteine-containing pro-teins. Membrane amino acid transport sys-tems, resembling microbial permease systems,have been demonstrated for methionine up-take by M. hominis and histidine uptake byM. fermentans (133). The methionine systemwas highly specific, but the histidine systemwas less specific and could be competitivelyinhibited by lysine and arginine. Two energy-dependent noninducible sugar transport sys-tems have been found in M. gallisepticum(149), one for D-glucose and a-methyl- D-glu-coside and one for D-mannose and D-fructose.An inducible a-glucosidase has also been dem-onstrated in A. Iaidlawii (158), but whetherthis activity or a coordinate permease aremembrane-associated has not been shown yet.A number of recent studies indicate that the
mycoplasma cell membrane is also involved indetermining the cells' antibiotic susceptibility.In a study of one Acholeplasma and six Myco-plasma species (39, 155), it has been shownthat resistance to chloramphenicol, dihydro-streptomycin, and tetracycline appears to in-volve a reduced permeability to the antibiot-ics, rather than an alteration in a ribosomalprotein. Also, in a study of A. laidlawii sensi-tivity to viral infections, Liss and Maniloff (79)found that cells selected for streptomycin ornovobiocin resistance were also virus-resistant.These examples indicate the significance ofthe cell membrane in considerations of anti-biotic susceptibility, but there is no reason tobelieve that there is a common mechanism forthese diverse phenomena.
Biophysical properties. Electron micros-
copy studies show the mycoplasma to bebounded by a membraneous structure having a"unit" membrane appearance (20, 31, 127).The generally reported thickness of the mem-brane is 7.5 to 1.0 nm (126), but in fact themembranes studied seem to fall into two sizeranges, 7 to 8 nm and 1.0 to 1.2 nm (20, 31).More accurate measurements are needed be-fore it can be decided whether this representsa significant difference among the myco-plasmas. In addition, consideration must begiven to the ionic conditions, in view of Bern-stein-Ziv's observation (11) that the M. galli-septicum membrane thickness is increased inhypotonic solutions.An interesting membrane arrangement has
been described in M. salivarium (75) wheremany cells on the surface of colonies were sur-rounded by membrane lamellae, giving amyelin-like appearance. Similar observationshave recently been made in virus-infected A.laidlawii cultures (J. Maniloff and A. Liss,unpublished data).
In freeze-etching studies of A. laidlawii cells(Fig. 5) and membranes (173, 175), the mem-brane is split along an internal plane exposing7.5- to 12.5-nm particles. Since these are notremoved by lipid solvents (J. Das and J. Mani-loff, 1971, Biophys. Soc. Abstr., p. 291a), theyprobably represent proteins within the mem-brane. The outer and inner (cytoplasmic)membrane surfaces had a rough-textured ap-pearance (173). Similar results have been ob-served for M. gallisepticum (Fig. 6), CaprinePPLO strain 14, and M. arthritidis (J. Mani-loff and R. Steere, unpublished data).
It is extremely difficult to evaluate the ob-servations of surface spikes in negativelystained preparations (26, 59, 60, 142). Most ofthese experiments involve the negativestaining of unfixed (sometimes disrupted) cellsand, hence, there is concern about the produc-tion of staining artifacts. On the other hand,there must be specific types of surface sites inthe mycoplasmas in order to explain their in-teraction with mammalian cells (167). M. galli-septicum, for example, binds to mammaliancells by the bleb end of the mycoplasma cell(182, 183). Therefore the ends of this myco-plasma must have the binding site constitu-ents. There have been, as yet, no data local-izing the surface spikes on the bleb structures.Therefore there are two unknowns: the statusof the spikes and the structure of the surface-binding sites.The electrical properties of M. gallisepticum
and A. laidlawii membranes have been meas-ured (20, 154) and found to be similar to those
272 BACTERIOL. REV.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/b
r on
24
Dec
embe
r 20
21 b
y 39
.111
.40.
240.
CELL BIOLOGY OF MYCOPLASMAS 273
.4I;
A~~~~
~ ~ ~ ~ ~ ~ vl
jr
-~~~J
}
.. A. A
FIG. 5. Micrograph of freeze-etched A. laidlawii B cells (90). The arrow shows the shadowing direction.The cells are filamentous, as seen in sections (Fig. 3). The fracture plane through the filamentous cell has(starting from the part of the cell at the lower left) travelled along the inside face of the membrane, throughthe cytoplasm, and through an internal membrane plane, exposing particles on a convex surface. x81,000.
VOL. 36, 1972
:!AQkx..
!. -i, j:,- .!r j.., 19
./ i.-. .,:..
.: ....
W- 1.
:1. z ,
'. :., .0
i,. --k "I V
?Ariz.
-. -.j - :", "I...- 1. .
..
.i
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/b
r on
24
Dec
embe
r 20
21 b
y 39
.111
.40.
240.
MANILOFF AND MOROWITZ
- q v**a.vp 'A
IV1W
a -j,..
I.s
. I~~~~~."il~~
1 .
,,_ W, .HI..v.tFIG. 6. Micrograph of freeze-etched M. gallisepticum A5969 (90). The arrow shows the shadowing direc-
tion. The cell morphology is the same as seen in sections (Fig. 4). The dividing two-bleb cell has been frac-tured along an internal membrane plane, showing particles on a convex surface. x81,000.
of other cell membranes. The cells have a neg-ative surface charge of about -0.01 coul/m2,which gives rise to a very small surface con-ductivity (about 0.003 mho/m), and have amembrane capacitance of about 0.9 ,Af/cm2.Mycoplasmas are much more osmotically
stable than bacterial protoplasts (127). It isinteresting to note that, using cells grown sothat their membranes are enriched with regardto specific fatty acids (discussed above), boththe rate of glycerol permeability (82) and theosmotic fragility (137, 146) follow the order
linoleic (18:2) > oleic (18: 1 cis) > eladic (18: 1trans). This would argue against leakiness sta-bilizing the cell, since then high leakinessshould correspond to low osmotic fragility. Allof these studies have involved A. laidlawiiwhich is much more osmotically fragile thanother Acholeplasma species (J. Maniloff and J.Robbins, unpublished data) and the Myco-plasma (123).
Lenard and Singer (78) reported that theoptical rotatory dispersion spectrum of A. laid-lawii membranes showed a minimum at 235
274 BACTERIOL. REV.
ffl. ."I,-hsso... r And:- -D
ownl
oade
d fr
om h
ttps:
//jou
rnal
s.as
m.o
rg/jo
urna
l/br
on 2
4 D
ecem
ber
2021
by
39.1
11.4
0.24
0.
CELL BIOLOGY OF MYCOPLASMAS
nm which indicated the presence of some a-
helical structure in the membrane proteins.Similar data were obtained by Choules andBjorklund (24) who also observed spectral pat-terns, characteristic of a-structure, in circulardichroism and infrared studies. Choules andBjorklund have summarized the A. Iaidlawiimembrane protein conformation, based on thethree physical chemical studies, as 23 to 31%a(helix), 30 to 57% ,B(pleated sheet), and 13 to45% random coil. The a content is in agree-
ment with other membranes (78), but no sig-nificant amount of , structure has been re-
ported in other membranes (179). Since all thedifferent membranes have the same spectralpatterns, the structure differences arise fromthe interpretations of the small band shiftsthat occur between the membranes and thepolypeptides of known conformation that are
used as standards. The various problems thatmight give rise to band shifts unrelated to theprotein structure (such as turbidity artifactsand spectra perturbation by other chromo-phores) have been discussed by Choules andBjorklund (24) and Wallach and Gordon (179).Except for the quarter to third of the proteinthat is a-helix, there is no agreement on theconfiguration of the rest of the membrane pro-tein structure.
Lipids in the mycoplasma membrane un-
dergo a melting type of thermal phase transi-tion from highly ordered fatty acid chains to a
liquid paraffin-line configuration. The exist-ence of this thermal phase transition in A.laidlawii cells, membranes, and lipids was firstdemonstrated by Steim et al. (168; see also 95,140) using differential thermal calorimetry.The phase transition has also been measuredby electron paramagnetic resonance (175) andX-ray diffraction studies (34, 35). From thesestudies, it has been concluded that most of themembrane lipid is in a bilayer arrangement.Since the transition temperature is a functionof the lipid composition, both can be varied bysupplementing the medium with a specificlipid (168) or by changing the growth tempera-ture (95). Melchior et al. (95) concluded thatthe cells try to regulate their lipid compositionso that the transition temperature is slightlybelow the growth temperature-i.e., the cellsattempt to grow on the disorder side of theorder-disorder phase transition. Rottem et al.(152), using electron paramagnetic resonance
studies, also noted an increase in membrane"fluidity" when the cells were grown at lowertemperatures-i.e., when "fluidity" is meas-
ured at 37 C, cells grown at 37 C will be justpast their lipid transition temperature, but
cells grown at 15 C will be well beyond theirtransition temperature and hence will showmore "fluidity."The membrane electron paramagnetic reso-
nance studies also show an increase in thefreedom of motion of the probe molecule as itis moved away from the polar end of the fattyacids (152), which would be expected for alipid bilayer structure. Tourtellotte, Branton,and Keith (175) found a decrease in the probemobility in membranes as compared with lipidextracts, an indication that membrane lipidmobility may be influenced by proteins. Theprotein-lipid interaction is also shown by theinaccessibility of some of the membrane pro-tein to proteolytic attack (106), to binding anuclear magnetic resonance probe (96), and tobinding a fluorescent probe (97).Reaggregation studies. Razin, Morowitz,
and Terry (134) showed that A. Iaidlawiimembrane could be solubilized by detergentsand that, if the detergent was removed, mem-brane reaggregates were obtained which hadmost of the original reduced nicotinamideadenine dinucleotide oxidase activity, a "unit"membrane appearance, and consisted of lipidand protein. Similar results have been foundfor membranes of M. gallisepticum (135, 151)and M. mycoides (135). An extensive review ofthe solubilization and reconstitution studies isin press (128).Most data have been accumulated for the
interaction of A. laidlawii membrane and so-dium dodecyl sulfate (SDS), and these havebeen reviewed by Engelman (33). The SDSsolubilizes the native membrane to form sepa-rate protein-SDS and lipid-SDS complexes,and not lipoprotein membrane subunits, ashad been thought earlier. Auborn, Eyring, andChoules (6) have recently shown that the solu-bilization kinetics are complex, half-order withrespect to both membranes and SDS micellsand second-order with respect to SDS mon-omers. The soluble components have molec-ular weights in the range of 20,000 to 49,000.Kahane and Razin (67) have shown that, if toolow SDS concentrations (10 mM) are used, thenative membrane is not completely solubilizedand this partially disaggregated membrane canbe subsequently recovered and erroneouslythought to be reaggregated membrane. Whenthe SDS is removed by dialysis, a membranereaggregate is formed, showing that no preex-isting structure is needed for this assemblyprocess. The lipid-protein ratio in the reaggre-gate is a function of the Mg2+ concentrationduring dialysis (33). Kahane and Razin (67)have shown that, at 20 mM Mg2+, the reaggre-
VOL. 36, 1972 275
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/b
r on
24
Dec
embe
r 20
21 b
y 39
.111
.40.
240.
MANILOFF AND MOROWITZ
gate lipid-protein ratio is the same as in thenative membrane.A variety of techniques have been used to
examine the reaggregated membrane structurefor comparison with the native membrane.Electron paramagnetic resonance studies haveshown that the "fluidity" of the reaggregatewas similar to that of the native membrane(152). X-ray diffraction patterns of the reaggre-gate resembled that of the native membrane,and a thermal phase transition was observed(96). These data indicate that a lipid bilayerstructure has been formed in the reaggregatedmembrane similar to that in the native mem-brane. However, since the 0.415-nm lipid X-ray reflection is about twice as broad for thereaggregate as compared with the nativemembrane, the ordered region giving rise tothe reflection must be smaller in the reaggre-gate than in the native membrane. Freeze-etching examination of the reaggregated mem-brane has revealed the absence of particles onthe fracture face (173). These particles appearto be proteins (discussed above) and indicate abasic structural difference between the reag-gregated and native membranes. Confirmingthis, the nuclear magnetic resonance (96) andfluorescence probes (97) show many protein-binding sites exposed in the reaggregate thatwere not available in the native membrane.Hence, in the reaggregate the membrane pro-teins are incorrectly reassembled into thestructure; either they have not been able toreassume the correct conformation or theyhave not been able to interact with the lipidscorrectly so as to make their binding sites in-accessible to the probes.The lack of specificity in the reassembly
process has been demonstrated by: the produc-tion of hybrid membranes, reaggregates con-taining lipids and proteins from both A. laid-lawii and M. gallisepticum (130), reaggregatescontaining M. pneumoniae glycolipids and A.laidlawii proteins (27, 136, 139), and reaggre-gates of A. laidlawii membranes which haveincorporated extraneous proteins such as peni-cillinase and bovinetserum albumin (150).The lack of lipid specificity (discussed
above) as far as membrane structure is con-cerned should be noted and may reflect thesignificance of lipid as a common denominatorin establishing the membrane structure. Thelipid composition sets the membrane "fluidi-ty" and the lipid bilayer structure seems toreassemble spontaneously into a thermody-namically stable structure. By contrast, mem-brane protein specificity is higher (e.g.,without the appropriate permease, substrates
cannot be utilized), and protein reassemblyinto membranes seems to be a more compli-cated architectural problem.
Extracellular AspectsExtracellular structures. In considering
extracellular structures, it seems reasonable toquestion the possible existence of surface enve-lopes external to the cell membrane and ofsurface appendages. The problem of the latterstructure arises because of the motility of someof the mycoplasmas (15, 18; W. Bredt, 1972,Med. Mikrobiol. Immunol. 157:169). However,since no surface appendages have been ob-served by electron microscopy the motility isreferred to as gliding.
Since the mycoplasmas are defined as cellsbounded only by a cell membrane, the ques-tion of other surface envelopes is a serious one.With one possible exception (to be discussed),electron microscopy shows no external surfaceenvelopes, and electrical measurements of thesurface conductivity of unfixed cells confirmsthe absence of surface structures other thanthe membrane (20). The possible exception isthe mycoplasma prototype strain Mycoplasmamycoides. In their biochemical studies of thegalactan produced by M. mycoides, Plackett,Buttery, and Cottew (117) suggested that thegalactan might form a capsular or slime-layersubstance, although they could not see anysuch structure by electron microscopy. How-ever, Gourlay and Thrower (49) showed thatgrowing M. mycoides cells were within a muci-nous homogenous matrix, which if precipitatedby specific antibody was frequently seenaround the cells as a capsular envelope. Theyalso mentioned the possibility that the viru-lence of particular M. mycoides strains may berelated to the amount of capsule. In a study ofCaprine PPLO strain 14 (a M. mycoides),Maniloff (Bacteriol. Proc., p. 78, 1968) showedthat, in older, dying cultures, cells were fre-quently surrounded by an amorphous surfaceenvelope-compare Fig. 1 of exponentiallygrowing cells with Fig. 7 of the same culture inearly death phase. Future considerations ofmycoplasma taxonomy should include the sig-nificance of M. mycoides capsular material.The osmotic stability of the mycoplasmas
suggests that consideration should be given topossible special properties or components ofthe surface envelope of these organisms. Theabsence of a bacterial type cell wall has beeninferred from penicillin resistance and the lackof wall-specific biochemical components (166).A possible exception is the presence of poly-
276 BACTERIOL. REV.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/b
r on
24
Dec
embe
r 20
21 b
y 39
.111
.40.
240.
CELL BIOLOGY OF MYCOPLASMAS
AS,.
* * . P% *
F. . s t t @ ,i. S * .;<,, e {.
z t s
>. ^ a,, ,, 4 !. 4
*' sIL-; as b 8 t ;F 4
S:, a, .s , s- o"'-; in + \
< "@ Add
An'I'd x -;
N t_.. oil 5,> N
- do _
FIG. 7. Section of late-stationary-phase Caprine PPLO strain 14 cells. Cells are seen either swollen ordensely packed with particles. An extracellular material surrounds these cells. Compare with Fig. 1, which isof the same culture sampled in log phase. x60,750.
hexosamine, which is about 5% of the dryweight of the A. laidlawii membrane (46).Extracellular products. Smith (166) re-
cently reviewed the mycoplasma extracellularproducts: neurotoxins, hemolysins, and exoen-zymes. However, the cytological mechanismfor the elaboration of these products is notknown. Thomas (172) has reviewed the data onthe two known mycoplasma neurotoxins, thoseof M. neurolyticum (an exotoxin) and M. galli-septicum. The M. neurolyticum toxin is a
thermolabile protein. Thomas (172) postulatedthat the pathogenicity of M. arthritidis andM. pulmonis may also be due to toxin produc-tion. Some mycoplasmas have alpha or betahemolytic activity, depending on the cultureconditions and the erythrocyte species beingused. Hemolysis seems to be due to peroxide,but it has not been possible to demonstrate arelationship between peroxides and pathogen-icity. The exoenzymes have been found to beresponsible for some of the effects of myco-
VOL. 36, 1972 277
At
41
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/b
r on
24
Dec
embe
r 20
21 b
y 39
.111
.40.
240.
MANILOFF AND MOROWITZ
plasmas on animal cells and appear to includenucleases and enzymes that can alter erythro-cyte antigenic determinants (see also 167).
ChromosomeThe mycoplasma DNA comprises 4 to 7% of
the dry weight of the cell (132); the variationprobably reflects species differences and alsovariations in the DNA per cell at different cul-ture phases, as discussed above. The adenine-thymine and guanine-cytosine (GC) ratios areabout 1.0, in agreement with the Watson-CrickDNA double-helix structure (166). The myco-plasma genetic information is in a single chro-mosome, which is a circular DNA molecule(102).Genome structure. The base ratios of the
mycoplasmas span the range of 23 to 41% GC(113). Within this range the Acholeplasma spe-cies are 30 to 36% GC, the Mycoplasma spe-cies (except for M. pneumoniae) are 23 to 36%GC, and M. pneumoniae is 39 to 41% GC. Theseven T strains that have been studied are allabout 28% GC (7). For comparison, the GCcontent of bacteria ranges from 25 to 75%.The variation in base composition within a
chromosome was shown by Ryan and Mo-rowitz (153) who were able, using Mycoplasmasp. (Kid) with 25% GC, to isolate a DNA frac-tion representing 1.4% of the total genomewhich was enriched in ribosomal RNA (rRNA)and transfer RNA (tRNA) cistrons and was46% GC.The published values of the DNA molecular
weights of mycoplasma chromosomes aresummarized in Table 2. The most extensivestudies were done by Bak et al. (8), usingmeasurements of the DNA renaturation reac-
tion rate constants to calculate the molecularweights. The other values come from a varietyof techniques and, unfortunately, adequatedata are not generally available to evaluatesome of the reported numbers. For example,the molecular weight of M. gallisepticummeasured by Riggs (143) is frequently quotedto be 10 x 108 daltons. However, the 10 x 108daltons measured (by radioautography, theKleinschmidt technique and chemical analy-sis) is the DNA per growing cell and, to correctfor exponential growth, it must be multipliedby 0.69 to obtain the DNA per newly formedcell. Therefore, 6.9 x 108 daltons is the DNAcontent of a daughter M. gallisepticum cell,and the true chromosome size (depending on
ploidy and the temporal relationship betweencell division and DNA replication) will be less
TABLE 2. Molecular weights of Mycoplasmachromosomes (x 108)
a Bak et al. (8).b By Kleinschmidt technique (108).c By Kleinschmidt technique (102).d By Kleinschmidt technique (14).e Calculated from data of Riggs (143).' M. meleagridis data from Allen (1).' By Kleinschmidt technique (153).h By radioautography (143).'By radioautography (102).By sucrose gradient centrifugation (32).
than or equal to this value. The sizes of theMycoplasma chromosomes are in the range of4.2 to 6.9 x 108 daltons, whereas those of theAcholeplasma chromosomes are 7.6 to 11.0 x108 daltons. As noted by Bak et al. (8), theAcholeplasma genomes are about twice aslarge as those of the Mycoplasma. Thesmallest reported bacterial genome is 8 x 108daltons and values range as high as several bil-lion daltons (100).The early reports of 108 daltons or less of
DNA per cell (103, 110), although now consid-ered too small, did stimulate interest in cellswith only a limited amount of genetic informa-tion. The mycoplasma genomes, while shownto be larger, still only code for about 600 to1,000 cistrons per cell (102, 108), and thislimits the ultimate complexity. This was dem-onstrated by the experiments of Ryan andMorowitz (153), using Mycoplasma sp. (Kid),which showed the genome of this cell to haveonly one cistron each for the 16S and 23SrRNA and only about 44 cistrons for tRNAmolecules.Those chromosomes which have been spread
out and examined by radioautography or the
278 BACTERIOL. REV.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/b
r on
24
Dec
embe
r 20
21 b
y 39
.111
.40.
240.
CELL BIOLOGY OF MYCOPLASMAS
Kleinschmidt technique have shown a circulartopology; they are M. agalactiae bovis (108),M. arthritidis H39 (14), M. gallisepticum (143),and Mycoplasma sp. (Kid) (153). Double Y's or
replication forks with two arms of equal lengthwere observed in the chromosomes of M. aga-lactiae bovis (102), M. arthritidis H39 (14), M.gallisepticum (143), and A. laidlawii (102).With respect to topology, chromosone replica-tion in the mycoplasmas involves DNA circlessimilar to those shown for Escherichia coli (19).
Replication. Smith (160) has shown thatDNA replication in A. kaidlawii is semiconserv-ative and proceeds unidirectionally from atmost a few growing point regions. The analysisof this datum and of the experiments on thy-mineless death in these organisms (162) sug-
gests that DNA replication continuesthroughout the cells' generation time. A smallaccount of nonconservative repair replicationhas been found in normally growing A. laid-lawii (162, 163) and is believed to occur in A.laidlawii after thymine starvation (162); a sim-ilar type of replication has been demonstratedin irradiated cells.The DNA growing point region is believed to
be a membrane-associated complex in A. laid-lawii and M. gallisepticum. Using osmoticallyshocked A. laidlawii lysates on sucrose gra-dients (159, 161), a subcellular fraction was iso-lated which contained a twofold enrichment ofpulse-labeled (i.e., newly synthesized) DNA.Since the DNA could be released by Pronaseor detergents, it was suggested that thegrowing point is localized at the cell mem-
brane. In M. gallisepticum preparations (122),lysed by freeze-thaw cycles, a low-speed sub-cellular fraction has been isolated which has a
four- to fivefold enrichment and contains 15%of the total cellular DNA and 80% of the pulse-labeled DNA. These experiments have shownthat this is the DNA growing region and thatthe subcellular fraction is enriched in the M.gallisepticum terminal structures (see Fig. 4),suggesting that this structure is the DNA rep-lication complex site.
Ultraviolet and X-ray irradiation experi-ments of mycoplasma cultures show multihitor multitarget kinetics; i.e., the survival curves
show an initial shoulder, followed by exponen-
tial inactivation (30, 38, 43, 44, 104). The mul-tiple-hit character of the inactivation curvesmay be due to cell clumping or polyploidy. InA. laidlawii it has been shown that both theultraviolet sensitivity and the number of lethalevents to kill a cell vary with the cell's growthphase (30). The multiorder survival kinetics
can be altered by the experimental conditions,giving rise to first-order kinetics. Furness et al.(44) noted the inconsistency in titer dependingon whether their irradiated cells were grown inbroth or on plates; hence the counting methodcan affect the type of survival curve obtained.The ultraviolet repair replication studies
reported thus far have all examined A. laid-lawii. Folsome (38), Smith and Hanawalt (163),and Das et al. (30) have all shown that thecells are capable of photoreversal of some ofthe DNA damage. Das et al. (30) found thatthis is maximal in middle to late exponential-phase cells. Smith and Hanawalt (163) andDas et al. (30) also demonstrated that the cellshave a dark-repair capability by the excision-repair mechanism. Smith and Hanawalt (163)estimated that the size of the region excised,the region around the radiation-induced py-rimidine dimer, is about 150 to 600 nucleo-tides. Das et al. (30) found that the single-strand DNA regions, between the breaks, wereabout 1/23 of the total strand length, and 60% ofthe breaks were repaired by 2 hr after irradia-tion. Hence, at least this one mycoplasma hasthe genetic information and enzymatic capa-bilities for both dark and light repair of itsDNA.The multiorder ultraviolet survival curves of
A. laidlawii have suggested that these cellsmight be multinucleate (30). A similar conclu-sion was reached for M. gallisepticum, basedon the difference in rates of DNA synthesis indifferent phase cultures (discussed above).However, in M. arthritidis H39, Bode andMorowitz (14) found the DNA per cell to be1.2 to 1.5 times the genome size, in agreementwith the expectation that an exponentiallygrowing cell with only one copy of its chromo-some should contain about 1.4 times its ge-nome. From so few cases, no generalizationcan be made, but it may be expected thatsmall coccoid mycoplasmas, like M. arthritidisH39, will have only one genome per cell, andthe larger mycoplasmas, especially the fila-mentous ones, will be multinucleate.Genetics. The stability of mycoplasma mu-
tants has not been carefully examined, butSteinberg et al. (169) noted leakiness in theirtemperature-sensitive mutants, and Liss andManiloff (unpublished data) have found thatnaladixic acid-resistant A. laidlawii reverts tosensitivity about a month after selection; theselection may, in fact, be for a slower-growth-rate cell, which will be naturally selectedagainst by a back mutation. It must be con-cluded that there are few stable mycoplasma
VOL. 36, 1972 279
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/b
r on
24
Dec
embe
r 20
21 b
y 39
.111
.40.
240.
MANILOFF AND MOROWITZ
mutants for which the mechanism of the muta-tional alteration is known.Mycoplasma mutation frequencies for anti-
biotic resistance and for carbohydrate utiliza-tion have been measured (86). Mutants (stablevariants) have been selected for antibiotic re-sistance (76), temperature sensitivity using themutagens nitrosoguanidine (169, 170), amino-purine or bromodeoxyuridine (C. E. Folsomeand J. Folsome, Bacteriol. Proc., p. 30, 1966),and constitutivity for the inductible glucosi-dase enzyme using the mutagen nitrosoguan-idine (158). The antibiotic-resistant mutantsthat have been studied, which involved anti-biotics known to interfere with protein syn-thesis, turned out to be resistant due to a re-duced antibiotic uptake rather than a specificribosomal protein alteration, as had been ex-pected (39, 155).
Unsuccessful attempts at DNA transforma-tion of A. Iaidlawii have been reported by Fol-some (38) and Smith (166). Folsome (38) re-ported that the cells were able to bind DNA sothat it was not accessible to deoxyribonucleaseattack, but no recombinants could be found.The data are completely negative for bothDNA-mediated (transformation) and cell-me-diated (conjugation) transfer of genetic infor-mation.The discovery of mycoplasma viruses (47)
opens the possiblity of other mechanisms ofgene transfer, in particular transduction andtransfection. Most of the known mycoplasmaviruses are bullet-shaped particles (47, 50, 79)and are believed to contain single-strandedDNA (A. Liss and J. Maniloff, unpublisheddata); hence, transduction by these virusesmay be quite rare. A better possibility fortransduction may be the few enveloped spher-ical mycoplasma viruses (48). Experimentshave shown that transfection can occur, i.e., A.laidlawii can be infected with mycoplasmavirus DNA, undergo a lytic infection, and pro-duce mature viruses (A. Liss and J. Maniloff,unpublished data).
Ribosomes and RNARNA represents 8 to 17% of the mycoplasma
cell mass (132). For M. gallisepticum, theRNA-DNA ratio is 4.5 for exponentiallygrowing cells (73) and 2.0 for cells in early sta-tionary phase (110). The RNA species are 22S,16S, and 5S rRNA, 4S tRNA, and unstablemessenger RNA (mRNA). The RNA is in-volved with protein synthesis, which Tour-tellotte (174) has shown follows transcriptionand translation mechanisms similar to those ofother procaryotes. Kirk and Morowitz (73)
found that about 86% of the RNA is present asrRNA and the rest as soluble RNA. Kirk (72)estimated that mRNA represents about 4.5%of the total RNA.Ribosomes and rRNA. Ribosomes are seen
as cytoplasmic granules in electron micro-graphs of the mycoplasmas. The 70S particlesize is 1.7 nm, determined from the opticalanalysis of M. gallisepticum micrographs (89).The ribosome composition in M. hominis is61% RNA and 39% protein (61), and in M. gal-lisepticum is 55% RNA and 45% protein (73).The so-called "70S particle" in M. hominis
has a sedimentation coefficient (corrected tostandard conditions) of 71S and dissociatesinto 53S and 33S subunits (61), and in M. gal-lisepticum has a corrected value of 74S anddissociates into 56S and 36S subunits (73).Johnson and Horowitz (61) have reported thatM. hominis 70S ribosomes require higher Mg2+concentrations for stability than those of E.coli. Polysomes with corrected sedimentationcoefficients of 110S and 135S have been re-ported in M. gallisepticum (73). Figure 8 showsa polysome distribution from M. gallisepticumwith polysomes of uncorrected sedimentationcoefficients at 98S (corresponding to the cor-rected 110S particle), 122S (the 135S particle),143S, 163S, and 182S (J. Maniloff, unpub-lished data).
CN C- C, C c 00Q0oCo eDt 0C CO LA"n
I I I I I
FIG. 8. Schlieren pattern of polyribosome distri-bution of M. gallisepticum A5969. The numbers arethe uncorrected sedimentation coefficients of eachpeak. The cells were broken in a French pressure cell(J. Maniloff, unpublished data).
280 BACTERIOL. REV.
I
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/b
r on
24
Dec
embe
r 20
21 b
y 39
.111
.40.
240.
CELL BIOLOGY OF MYCOPLASMAS
Johnson and Horowitz (61) examined M.hominis ribosomal proteins by polyacrylamidedisc-gel electrophoresis. They observed 18 pro-tein bands for the 30S subunit, as compared to15 bands for E. coli 30S particles, and 21bands for the 50S subunit as compared to 20bands for E. coli 50S particles. Hence, myco-plasma ribosomes are as complex as eubac-terial ribosomes both with regard to the num-bers of proteins and RNA species. Johnson andHorowitz (61) also noted that about half theproteins from both M. hominis and E. colishowed similar electrophoresis mobilities andhalf were different.
Kirk and Morowitz (73) found two rRNAspecies in M. gallisepticum ribosomes, at 16Sand 22S. The first corresponds to E. coli 16SrRNA (from the 30S subunit), but the seconddefinitely sediments slower than the E. coli23S rRNA (from the 50S subunit). Similar re-sults are cited by Tourtellotte (174) for M.pneumoniae which has rRNA at 16S andslightly lower than 23S. In M. hominis,Johnson and Horowitz (61) reported the 16Sand 22S rRNA, but also were able to isolatethe 5S rRNA in the ratio of one 5S moleculefor each 16S and 22S pair, so mycoplasma ri-bosomes have the same three rRNA species asthe eubacteria. The difference between themycoplasma 22S rRNA and the bacterial 23SrRNA is not due to nuclease action duringpreparation (61) so must reflect either slightsize or conformational differences in the twoRNA species.Kirk and Morowitz (73) reported that 63% of
the M. gallisepticum rRNA was 22S RNA.This is in agreement with the observation thatsedimentation patterns of M. gallisepticumshow more 50S than 30S particles (73; see Fig.8). The sedimentation pattern of M. hominisrRNA also shows more 22S RNA than 16SRNA (61). The reason why these amounts areunequal is not known.Table 3 summarizes the reported nucleoside
compositions of mycoplasma ribosomes. Dataon E. coli ribosomes and on the DNA composi-tions have been included for comparison. Themycoplasma rRNA compositions are all sim-ilar and not too different from that of E. coli;the difference between the mycoplasma and E.coli data is seen to be the small decrease inguanosine and small increases in adenosineand uridine, making the mycoplasma GClower than that of E. coli. The M. hominis andE. coli 16S and 22 to 23S RNA species are alsosimilar to each other and to the bulk rRNAcomposition. Kirk and Morowitz (73) cite sim-ilar results for M. gallisepticum 16S and 22S
rRNA. However, large differences in both as-pects are seen for the 5S RNA species. Al-though earlier workers were unable to findminor bases (54), Johnson and Horowitz (61)also found that M. hominis 16S and 22S rRNAcontained the modified nucleosides pseudouri-dine, dihydrouridine, ribosylthymidine, 3-methyluridine, 5-methylcytidine, inosine, 1-methyladenosine, 6-N-methyladenosine, 6,6-N-dimethyladenosine, 2-N-methylguanosine,and 7-methylguanosine. Hence, at least M.hominis rRNA does contain a number of modi-fied nucleosides and, although they are notquantitatively as large as in E. coli, they aresignificant. In 16S rRNA, 1.02 mole% of theM. hominis bases are modified and 1.85 mole%of those in E. coli; in 22 to 23S rRNA, 0.58mole% are modified in M. hominis and 1.12mole% in E. coli.Thre is a significant difference between the
percent GC of the mycoplasma DNA and thepercent GC of the rRNA (Table 3). This indi-cates the heterogeneity in percent GC of dif-ferent cistrons and, since the mycoplasmarRNA GC is close to that of most procaryotes,probably reflects the evolutionary conservationof the rRNA base sequences. Ryan and Mo-rowitz (153) showed that, although Myco-plasma sp. (Kid) DNA is 25% GC, a DNA frac-tion containing the rRNA cistrons was 46%GC, close to the rRNA 48% GC, and they wereable to use the per cent GC heterogeneity inMycoplasma sp. (Kid) to partially purify therRNA cistrons. They were able to show thatthe Mycoplasma sp. (Kid) genome containsonly one cistron each for its 16S and 22SrRNA. This compares with E. coli which hasfive cistrons each for 16S and 23S rRNA.Ribosome superstructures have been re-
ported in the form of helices in M. gallisep-ticum (92) and as tetrads in T strains (F. T.Black, 1972, Med. Mikrobiol. Immunol. 157:170). The helices have been shown to ariseduring preparation (93) and are not polysomecondensations (88). They have been shown tobe self-assembly structures of 70S ribosomes,with 10 particles per 3 turns as the repeat, andthere are indications that the helices are stabi-lized by interactions between 50S particles (88,89).Mycoplasma protein synthesis, both in vivo
and in vitro, is sensitive to those antibioticsknown to be inhibitory through their action on70S ribosomes: chloramphenicol, puromycin,erythromycin, and tetracycline (39, 174, 176).Fraterrigo and Perlman (39) have shown thatribosomes from a tetracycline-resistant A. laid-lawii were antibiotic-sensitive, indicating that
VOL. 36, 19^72 281
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/b
r on
24
Dec
embe
r 20
21 b
y 39
.111
.40.
240.
TABLE 3. Nucleoside composition of Mycoplasma rRNA
aNeimark (113).b Johnson and Horowitz (61).c Walker (178).d Ryan and Morowitz (153).'Kirk and Morowitz (73).
the mutation has not been in a ribosome pro-tein gene. Maniloff (88) was able to show thatchloramphenicol and lincomycin, which bindto the 50S part of the bacterial ribosomes, donot allow ribosome helices to form in M. galli-septicum, indicating that these antibioticsblock the interaction sites that stabilize thehelix. Conversely, cycloheximide, which blockseucaryotic protein synthesis involving 80S ri-bosomes, has no effect on mycoplasmas (174,176).tBNA. Mycoplasma tRNA has been shown
to sediment at 4S, along with E. coli tRNA(55, 61), and to have a thermal denaturationcurve similar to E. coli tRNA, indicating sim-ilar secondary structures (55).Some M. hominis tRNA species (61) and
Mycoplasma sp. (Kid) tRNA species (55) canbe charged by some E. coli aminoacyl-tRNAsynthetases, and Mycoplasma sp. (Kid) tRNAcan also substitute for E. coli tRNA in in vitrocell-free protein synthesis. Redundancy hasbeen observed in the tRNA isoaccepting spe-cies: A. Iaidlawii was found to have only asingle tRNA each for tyrosine, phenylalanine,alanine, glycine, and cysteine; two tRNA spe-cies for arginine, methionine, and isoleucine;and three tRNA species for valine, serine, andleucine (37, 55). Mycoplasma sp. (Kid) hasthree isoleucine tRNA species (62). The Myco-plasma sp. (Kid) tRNA redundancy is inter-esting in view of the data of Ryan and Mo-rowitz (153) showing that these cells only haveabout 44 tRNA cistrons, three of which are forIle-tRNA species and two for Met-tRNA spe-cies.
One of the two Met-tRNA species has beenfound to fMet-tRNA in M. gallisepticum (55),Mycoplasma sp. (Kid) (55), A. kaidlawii A (37),and A. laidlawii B (55). This suggests that, likeother procaryotes, the mycoplasmas initiatetheir peptide chains with formylmethionine.
Studies have been carried out on the basecompositions of bulk tRNA of several myco-plasmas and isolated Mycoplasma sp. (Kid)Ile-tRNA (Table 4). In addition to these, M.gallisepticum tRNA has been found to containisopentenyladenosine (55), and M. mycoidestRNA contains pseudouridine, 4-thiouridine,ribothymidine, and dihydrouridine (178). Al-though these studies are still in a relativelyearly stage, it is clear that mycoplasma tRNAspecies contain a variety of minor bases. Theabsence of ribothymidine in two Mycoplasmaspecies (Table 4) indicates that these tRNAspecies may differ significantly from eubac-terial tRNA. The mycoplasma tRNA percentGC is (Table 4): 54% for A. laidlawii, 56% forM. hominis, 54% for Mycoplasma sp. (Kid),50% for Mycoplasma sp. (Kid) Ile-tRNA, 55%for M. gallisepticum (73), and 61% for E. coli.These values are quite high compared with theDNA percent GC of these mycoplasmas(Table 3: 25 to 35%). The tRNA base ratio, aswell as the rRNA base ratio, seems to havebeen conserved during the evolution of theseorganisms.mRNA. Using actinomycin D, which inhibits
RNA synthesis, Tourtelotte (174) was able tofollow mRNA decay in seven mycoplasmas bythe decay in protein synthesis. There was avariation in the rate of cessation of protein
282 MANILOFF AND MOROWITZ BACTRIOL. ]REV.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/b
r on
24
Dec
embe
r 20
21 b
y 39
.111
.40.
240.
CELL BIOLOGY OF MYCOPLASMAS
TABLE 4. Nucleoside composition of Mycoplasma tRNA
Nucleosides per tRNA moleculeNucleoside | uloimetN Mylcoplam°
Nucleoside ~~Acholeplasma M. hominisb Mycoplasma MycplasmaIl Eschercikaidkawiia sp. (Kid)c S RNA(Kddle s rChia
a Feldman and Falter (37).'Calculated from data of Johnson and Horowitz (61).c Calculated from data of Hayashi, Fisher, and S511 (55), assuming 80 nucleotides/tRNA.d Johnson, Hayashi, and S6ll (62).
synthesis, but in all cases it was complete in15 min. From these data, the mRNA half-lifewas calculated to be less than 4 min, which isclose to the 2-min mRNA half-life which Kirk(72) reported for M. gallisepticum.
PlasmidsPlasmids are extranuclear genetic structures
which can reproduce autonomously (56). Thefinding that many mycoplasmas carry viruses
(48, 79) makes it fairly certain that there are
mycoplasma plasmids.In electron microscopy studies of M. arthri-
tidis DNA, Morowitz (102) reported, in addi-tion to the large pieces of DNA, small circlesof less than 20 x 106 molecular weight. Also inM. arthritidis, Haller and Lynn (Bacteriol.Proc., p. 68, 1968) found a satellite DNA bandin cesium chloride gradients. It is not knownwhether this is a plasmid; the one reportedeffort to isolate a virus from M. arthritidis was
unsuccessful (79), but one has been isolated inmore recent experiments (A. Liss and J. Mani-loff, unpublished data). There is also the possi-bility of nonviral plasmids.A virus was isolated from A. kaidlawii B (79)
and, in other studies of these cells, in additionto the cells' chromosomal DNA, a low-molec-
ular-weight DNA peak was identified in neu-tral and sucrose gradients (30, 32). This peak,which is 35% of the total DNA in the cell, isbelieved to be viral plasmids (90).
Other Cytoplasmic OrganellesCytoplasmic granules or structures have
been described in M. gallisepticum, A. laidla-wii, and M. pneumoniae. The structures ap-pear morphologically different in each case.The pear-shaped M. gallisepticum cells (Fig.
4, 6) have a terminal bleb structure, connectedto the cell by an infra-bleb region (91, 93, 105).The bleb is hemispherical, about 80 by 125mm, and the base, its attachment to the infra-bleb, is a flat, circular plate. The infra-blebregion is about 200 nm in diameter and ap-pears to be two concentric spherical regions.M. gallisepticum binds to mammalian cells bythe bleb end of the cell (182, 183); hence, theterminal structures must have some sort ofbinding sites. During the life cycle of the cell,these structures are replicated and form oppo-site poles of the cell at division (91, 105).These structures are found in the subcellularfraction containing the DNA growing region(122) and appear to be involved in DNA repli-cation. Munkres and Wachtel (111) showed
283VOL. 36, 1972
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/b
r on
24
Dec
embe
r 20
21 b
y 39
.111
.40.
240.
MANILOFF AND MOROWITZ
that the cells' adenosine triphosphatase ac-tivity is localized along the inside of the mem-brane bounding the bleb and infra-bleb region,and acid phosphatase is localized only in theinfra-bleb region. There is a suggestion thatacid phosphatase activity may be different inthe one-bleb daughter cell versus the two-blebpredivision cell. Maniloff (90) has shown thatthe central core of the infra-bleb region andthe periphery of the bleb are rich in basic pro-tein and, with tellurite used as an electronacceptor, redox activity was localized in theinfra-bleb region. In summary, the M. gallisep-ticum terminal structures seem to be the sitesof a variety of enzymatic processes and to beinvolved, at least as the probable site of DNAsynthesis, in cellular regulation.Each filamentous A. Iaidlawii cell seems to
contain a granular region (Fig. 3) which parti-tions the filament (87). It is spherical and aswide as the cell, about 0.2 to 0.3 ,m. There areno data on the possible function of this mate-rial.
Biberfeld and Biberfeld (12) described theknoblike structures on the ends of M. pneumo-niae filaments; these consist of a denselystained rod surrounded by cytoplasm andending with a platelike structure. The onlydata on the functional aspect of these terminalstructures is the recent observation of Collierand Clyde (28) that they are the regions of in-teraction between M. pneumoniae cells andhuman fetal trachea organ culture cells.
SUMMARYThe basic points of this review of myco-
plasma cell biology were presented at the be-ginning as a set of generalizations: (i) differ-ence of morphology among species, with spe-cies characteristic cell structure; (ii) 0.33 ,um(instead of 0.1 jim) as the smallest viable my-coplasmas; (iii) cell replication primarily bybinary division; and (iv) a biochemistry similarto that of other procaryotic cells. The "elemen-tary bodies" with their promise of novel modesof cell growth and division have proven to benonexistent.Regarding the last generalization, myco-
plasma biology is seen to be qualitatively thesame as that of other procaryotes; the only dif-ferences are small quantitative ones arisingbecause of the limited cell size and genome.All the complexity of molecular biology is seenin the mycoplasmas, and this must mean thatthe subtle sophistications seen in the biochem-ical structures and mechanisms are not evolu-tionary additions but constitute the funda-mental requirements of all biological systems.
The linear dimensions of the smallest myco-plasmas are about two times that of the theo-retical minimal 0.15-Aim cell; in other words,there are living cells only 10 times more mas-sive than the most restrictive cell that can bepostulated, assuming conventional terrestrialbiology. Mycoplasma are indeed near thelimits imposed by the required apparatus ofmolecular biology. If appreciably smaller cellsare ever found to exist, the finding will chal-lenge our ideas of what constitutes the neces-sary biochemical processes for life.
ACKNOWLEDGMENTS
We thank the many members of our research groups whohave contributed to our information and ideas concerningthe mycoplasmas. Support for this work has come from thePublic Health Service National Institutes of Health, theNational Aeronautics and Space Administration, the U.S.Atomic Energy Commission, and the University of Roch-ester Atomic Energy Project (Report No. UR-3490-54). J.M.is the recipient of Public Health Service Research CareerDevelopment Award A117480.We thank our friend and colleague, Mark Tourtellotte,
for many valuable discussions, guidance in experimentalproblems, and his advice in planning this review.
LITERATURE CITED
1. Allen, T. C. 1971. Base composition and ge-nome size of Mycoplasma meleagridis deoxy-ribonucleic acid. J. Gen. Microbiol. 69:285-286.
la. Allen, T. C., J. 0. Stevens, E. R. Florance,and R. 0. Hampton. 1970. Ultrastructure ofMycoplasma gallisepticum isolate 1056. J.Ultrastruct. Res. 33:318-331.
2. Anderson, D. L., M. E. Pollock, and L. F.Brower. 1965. Morphology of Mycoplasmalaidlawii type A. II. Effect of glucose ongrowth and cellular morphology. J. Bacteriol.90:1768-1777.
3. Anderson, D. R., and M. F. Barile. 1966. Ultra-structure of Mycoplasma orale isolated frompatients with leukemia. J. Nat. Cancer Inst.36:161-167.
4. Argaman, M., and S. Razin. 1965. Cholesteroland cholesterol esters in mycoplasma. J. Gen.Microbiol. 38:153-168.
5. Argaman, M., and S. Razin. 1969. Antigenicproperties of mycoplasma organisms andmembranes. J. Gen. Microbiol. 55:45-58.
6. Auborn, J. J., E. M. Eyring, and G. L. Choules.1971. Kinetics of sodium dodecyl sulfate solu-bilization of Mycoplasma laidlawii plasmamembranes. Proc. Nat. Acad. Sci. U.S.A. 68:1996-1998.
7. Bak, A. L., and F. T. Black. 1968. DNA basecomposition of human T strain mycoplasmas.Nature (London) 219:1044-1045.
8. Bak, A. L., F. T. Black, C. Christiansen, and E.A. Freundt. 1969. Genome size of myco-plasmal DNA. Nature (London) 224:1209-1210.
9. Barer, R. 1959. Phase, interference and polar-
284 BACTERIOL. REV.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/b
r on
24
Dec
embe
r 20
21 b
y 39
.111
.40.
240.
CELL BIOLOGY OF MYCOPLASMAS
izing microscopy, p. 169-272. In R. C. Mel-lors (Ed.), Analytical cytology. McGraw-HillBook Co., New York.
10. Bernstein-Ziv, R. 1969. Cell division in Myco-plasma gallisepticum. Can. J. Microbiol. 15:1125-1128.
11. Bernstein-Ziv, R. 1971. The effect of hypotonicsolutions on the morphology of cells of Myco-plasma gallisepticum. Can. J. Microbiol. 17:1203-1205.
12. Biberfeld, G., and P. Biberfeld. 1970. Ultra-structural features of Mycoplasma pneumo-niae. J. Bacteriol. 102:855-861.
13. Boatman, E. S., and G. E. Kenny. 1970. Three-dimensional morphology, ultrastructure, andreplication of Mycoplasma felis. J. Bacteriol.101:262-277.
14. Bode, H. R., and H. J. Morowitz. 1967. Sizeand structure of the Mycoplasma hominisH39 chromosome. J. Mol. Biol. 23:191-199.
15. Bredt, W. 1968. Motility and multiplication ofMycoplasma pneumoniae. Pathol. Microbiol.32:321-326.
16. Bredt, W. 1968. PhasenkontrastmikroskopischeUntersuchungen zur Morphologie und Ver-mehrung von Mycoplasma pneumoniae anGlas. Zentralbl. Bakteriol. Parasitenk. Abt. IOrig. 208:549-562.
17. Bredt, W. 1970. Experimentelle Untersu-chungen uiber Morphologie und Vermehrungder beim Menschen vorkommenden Myco-plasmen unter besonderer Beruicksichtigungvon Mycoplasma hominis. Z. Med. Mikro-biol. Immunol. 155:248-274.
18. Bredt, W., K. H. H6fling, H. H. Heunert, andB. Milthaler. 1970. Messungen an beweg-lichen Zellen von Mycoplasma pneumoniae.Z. Med. Mikrobiol. Immunol. 156:39-43.
19. Cairns, J. 1963. The chromosome of Escherichiacoli. Cold Spring Harbor Symp. Quant. Biol.28:43-46.
20. Carstensen, E. L., J. Maniloff, and C. W. Ei-nolf. 1971. Electrical properties and ultra-structure of Mycoplasma membranes. Bio-phys. J. 11:572-581.
21. Charman, W. N. 1963. Some experiments con-cerning the limitations and errors in sizemeasurement of small objects by visual mi-croscopy. J. Roy. Opt. Soc. 82:81-94.
22. Chen, T.-A., and R. R. Granados. 1970. Plant-pathogenic mycoplasma-like organism: main-tenance in vitro and transmission to Zeamays L. Science 167:1633-1636.
23. Cho, H. W., and H. J. Morowitz. 1969. Charac-terization of the plasma membrane of Myco-plasma laidlawii. Biochim. Biophys. Acta183:295-303.
24. Choules, G. L., and R. F. Bjorklund. 1970. Evi-dence of # structure in Mycoplasma mem-branes: circular dichroism, optical rotatorydispersion, and infrared studies. Biochem-istry 9:4759-4767.
25. Choules, G. L., and W. R. Gray, 1971. Pepti-dase activity in the membranes of Myco-
26. Chu, H. P., and R. W. Horne. 1967. Electronmicroscopy of Mycoplasma gallisepticum andMycoplasma mycoides using the negativestaining technique and their comparison withMyxovirus. Ann. N.Y. Acad. Sci. 143:190-203.
27. Cole, R. M., J. Popkin, B. Prescott, R. M.Chanock, and S. Razin. 1971. Electron mi-croscopy of solubilized Acholeplasma laid-lawii membrane proteins reaggregated withMycoplasma pneumoniae glycolipids.Biochim. Biophys. Acta 233:76-83.
28. Collier, A. M., and W. A. Clyde. 1971. Relation-ships between Mycoplasma pneumoniae andhuman respiratory epithelium. Infect. Immu-nity 3:694-701.
29. Darland, G., T. D. Brock, W. Samsonoff, and S.F. Conti. 1970. A thermophilic, acidophilicmycoplasma isolated from a coal refuse pile.Science 170:1416-1418.
30. Das, J., J. Maniloff, and S. B. Bhattacharjee.1972. Dark and light repair in ultraviolet-ir-radiated Acholeplasma laidlawii. Biochim.Biophys. Acta 259:189-197.
31. Domermuth, C. H., M. H. Nielsen, E. A.Freundt, and A. Birch-Andersen. 1964. Ultra-structure of Mycoplasma species. J. Bacte-riol. 88:727-744.
32. Dugle, D. L., and J. R. Dugle. 1971. Presence oftwo DNA populations in Mycoplasma laid-lawii. Can. J. Microbiol. 17:433-434.
33. Engelman, D. M. 1968. Solubilization and ag-gregation properties of membrane compo-nents from Mycoplasma laidlawii, p. 203-228.In L. Bolis and B. A. Pethica (ed.), Mem-brane models and the formation of biologicalmembranes. John Wiley, New York.
34. Engelman, D. M. 1970. X-ray diffractionstudies of phase transitions in the membraneof Mycoplasma laidlawii. J. Mol. Biol. 47:115-117.
35. Engelman, D. M. 1971. Lipid bilayer structurein the membrane of Mycoplasma laidlawii. J.Mol. Biol. 58:153-165.
36. Engelman, D. M., and H. J. Morowitz. 1968.Characterization of the plasma membrane ofMycoplasma laidlawii. IV. Structure andcomposition of membrane and aggregatedcomponents. Biochim. Biophys. Acta 150:385-396.
37. Feldmann, H., and H. Falter. 1971. Transferribonucleic acid from Mycoplasma laidlawiiA. Eur. J. Biochem. 18:573-581.
38. Folsome, C. E. 1968. Deoxyribonucleatebinding and transformation in Mycoplasmalaidlawii. J. Gen. Microbiol. 50:43-53.
39. Fraterrigo, C. C., and D. Perlman. 1971. Tetra-cycline inhibition of Mycoplasma ribosomalprotein synthesis. J. Antibiot. 24:185-188.
40. Freundt, E. A. 1958. The Mycoplasmataceae.Munksgaard, Copenhagen.
41. Freundt, E. A. 1969. Cellular morphology and
285VOL. 36, 1972
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/b
r on
24
Dec
embe
r 20
21 b
y 39
.111
.40.
240.
286 MANILOFF AND MOROWITZ
mode of replication of the mycoplasmas, p.281-315. In L. Hayflick (ed.). The mycoplas-matales and the L-phase of bacteria. Ap-pleton-Century-Crofts. New York.
42. Freundt, E. A. 1970. Morphology and ultra-structure of the mycoplasmas, p. 29-81. In J.T. Sharp (ed.). The role of mycoplasmas andL forms of bacteria in disease. Charles CThomas, Springfield, Ill.
43. Furness, G. 1968. Analysis of the growth cycleof Mycoplasma orale by synchronized divi-sion and by ultraviolet irradiation. J. Infect.Dis. 118:436-442.
44. Furness, G., F. J. Pipes, and M. J. McMurtrey.1968. Susceptibility of human mycoplasmatato ultraviolet and x-irradiations. J. Infect.Dis. 118:1-6.
45. Furness, G., F. J. Pipes, and M. J. McMurtrey.1968. Analysis of the life-cycle of Myco-plasma pneumoniae by synchronized divisionand by ultraviolet and x-irradiations. J. In-fect. Dis. 118:7-13.
46. Gilliam, J. M., and H. J. Morowitz. 1972. Char-acterization of the plasma membrane ofMycoplasma laidlawii. IX. Isolation and char-acterization of the membrane polyhexosa-mine. Biochim. Biophys. Acta, in press.
47. Gourlay, R. N. 1970. Isolation of a virus in-fecting a strain of Mycoplasma laidlawii.Nature (London) 225:1165.
48. Gourlay, R. N. 1971. Mycoplasmatales virus-laidlawii 2, a new virus isolated from Achole-plasma laidlawii. J. Gen. Virol. 12:65-67.
49. Gourlay, R. N., and K. J. Thrower. 1968. Mor-phology of Mycoplasma mycoides thread-phase growth. J. Gen. Microbiol. 54:155-159.
50. Gourlay, R. N., J. Bruce, and D. J. Garwes.1971. Characterization of mycoplasmatalesvirus laidlawii 1. Nature New Biol. 229:118-119.
51. Gourret, J.-P. 1970. Ultrastructure et micro-ecologie des mycoplasmes de phloeme dantrois maladies des petales verts. Etude deslesions cellulaires. J. Microscopie 9:807-822.
52. Granados, R. R. 1969. Electron microscopy ofplants and insect vectors infected with thecorn stunt disease agent. Contrib. BoyceThompson Inst. 24:173-188.
53. Granados, R. R., K. Maramorosch, and E. Shi-kata. 1968. Mycoplasma: suspected etiologicagent of corn stunt. Proc. Nat. Acad. Sci.U.S.A. 60:841-844.
54. Hall, R. H., A. Mittelman, J. Horoszewicz, andJ. T. Grace. 1967. The RNA of mycoplasma880. Ann. N.Y. Acad. Sci. 143:799-800.
55. Hayashi, H., H. Fisher, and D. S611. 1969.Transfer ribonucleic acid from Mycoplasma.Biochemistry 8:3680-3686.
56. Hayes, W. 1968. The genetics of bacteria andtheir viruses, 2nd edition. John Wiley, NewYork.
57. Hayflick, L. (ed.). 1969. The mycoplasmatalesand the L-phase of bacteria. Appleton-Cen-tury-Crofts, New York.
BACTERIOL. REV.
58. Herbert, D. 1961. The chemical composition ofmicroorganisms as a function of their envi-ronment. Symp. Soc. Gen. Microbiol. 11:391-416.
59. Horne, R. W. 1970. The ultrastructure of myco-plasma and mycoplasma-like organisms.Micron 2:19-38.
60. Hummeler, K., N. Tomassini, and L. Hayflick.1965. Ultrastructure of a mycoplasma (Ne-groni) isolated from human leukemia. J. Bac-teriol. 90:517-523.
61. Johnson, J. D., and J. Horowitz. 1971. Charac-terization of ribosomes and RNAs from My-coplasma hominis. Biochim. Biophys. Acta247:262-279.
62. Johnson, L., H. Hayashi, and D. S611. 1970. Iso-lation and properties of a transfer ribonucleicacid deficient in ribothymidine. Biochemistry9:2823-2831.
63. Jones, A. S., and R. T. Walker. 1963. Isolationand analysis of the deoxyribonucleic acid ofMycoplasma mycoides var. capri. Nature(London) 198:588-589.
64. Jones, T. C., and J. G. Hirsch. 1971. The inter-action in vitro of Mycoplasma pulmonis withmouse peritoneal macrophages and L-cells. J.Exp. Med. 133:231-259.
65. Kahane, I., and S. Razin. 1969. Immunologicalanalysis of mycoplasma membranes. J. Bac-teriol. 100:187-194.
66. Kahane, I., and S. Razin. 1969. Synthesis andturnover of membrane protein and lipid inMycoplasma laidlawii. Biochim. Biophys.Acta 183:79-89.
67. Kahane, I., and S. Razin. 1971. Characteri-zation of the mycoplasma membrane pro-teins. I. Reaggregation of solubilized mem-brane proteins of Acholeplasma laidlawii.Biochim. Biophys. Acta 249:159-168.
68. Kammer, G. M., J. D. Pollack, and A. S.Klainer. 1970. Scanning-beam electron mi-croscopy of Mycoplasma pneumoniae. J. Bac-teriol. 104:499-502.
69. Keller, R., and H. E. Morton. 1954. The growthof pleuropneumonia-like organisms of humanorigin. J. Bacteriol. 67:129-134.
70. Kelton, W. H. 1960. Growth-curve studies ofpleuropneumonia-like organisms. Ann. N.Y.Acad. Sci. 79:422-429.
71. Kelton, W. H. 1962. Synchronized division ofavian pleuropneumonia-like organisms. J.Bacteriol. 83:948-955.
72. Kirk, R. G. 1967. RNA of Mycoplasma gallisep-ticum. Ph.D. thesis, Yale University. Univ.Microfilms, Ann Arbor, Mich. (Diss. Abstr.67-8385).
73. Kirk, R. G., and H. J. Morowitz. 1969. Ribonu-cleic acids of Mycoplasma gallisepticumstrain A5969. Amer. J. Vet. Res. 30:287-293.
74. Klieneberger-Nobel, E. 1962. Pleuropneumonia-like organisms (PPLO): Mycoplasmataceae.Academic Press Inc., New York.
75. Knudson, D. L., and R. MacLeod. 1970. Myco-plasma pneumoniae and Mycoplasma sali-
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/b
r on
24
Dec
embe
r 20
21 b
y 39
.111
.40.
240.
CELL BIOLOGY OF MYCOPLASMAS
varium: electron microscopy of colony growthin agar. J. Bacteriol. 101:609-617.
76. Koostra, W. L., J. N. Adams, and P. F. Smith.1966. In vitro selection and identification ofantibiotic-resistant Mycoplasma mutants. J.Bacteriol. 91:2386-2387.
77. Lemcke, R. M. 1971. Sizing small organisms.Nature (London) 229:492-493.
78. Lenard, J., and S. J. Singer. 1966. Protein con-
formation in cell membrane preparations as
studied by optical rotatory dispersion andcircular dichroism. Proc. Nat. Acad. Sci.U.S.A. 56:1828-1835.
79. Liss, A., and J. Maniloff. 1971. Isolation ofmycoplasmatales viruses and characterizationof MVL1, MVL52, and MVG51. Science 173:725-727.
80. Low, I. E., and M. D. Eaton. 1965. Replicationof Mycoplasma pneumoniae in broth culture.J. Bacteriol. 89:725-728.
81. McElhaney, R. N., and M. E. Tourtellotte.1969. Mycoplasma membrane lipids: varia-tions in fatty acid composition. Science 164:433-434.
82. McElhaney, R. N., J. DeGier, and L. L. M. VanDeenen. 1970. The effect of alterations infatty acid composition and cholesterol con-
tent on the permeability of Mycoplasma laid-lawii B cells and derived liposomes. Biochim.Biophys. Acta 219:245-247.
83. Maillet, P. L. 1970. Infection simultanee pardes particules de type PLT(Rickettsiales) etde type PPLO (Mycoplasmatales) chez un
insecte vecteur de la phyllodie du trefle, Eus-celis lineolatus brulle (Homoptera, Jassidae).J. Microsc. 9:827-832.
84. Maillet, P. L., and J. Gouranton. 1971. Etudedu cycle biologique du mycoplasme de laphyllodie du trefle dans l'insecte vecteurEuscelis lineolatus brulle (Homoptera, Jassi-dae). J. Microsc. 11:143-162.
85. Maniloff, J. 1969. Electron microscopy of smallcells: Mycoplasma hominis. J. Bacteriol. 100:1402-1408.
86. Maniloff, J. 1969. Use of blood agar plates tostudy Mycoplasma physiology. Microbios. 1:125-135.
87. Maniloff, J. 1970. Ultrastructure of Myco-plasma laidlawii during culture development.J. Bacteriol. 102:561-572.
88. Maniloff, J. 1970. Structure of ribosome helicesof Mycoplasma gallisepticum. Proc. Int.Congr. Electron Microscopy, 7th, Grenoble 3:71-72.
89. Maniloff, J. 1971. Analysis of the helical ribo-some structures of Mycoplasma gallisep-ticum. Proc. Nat. Acad. Sci. U.S.A. 68:43-47.
90. Maniloff, J. 1972. Cytology of the myco-plasmas. Ciba Symposium on PathogenicMycoplasmas, in press.
91. Maniloff, J., and H. J. Morowitz. 1967. Ultra-structure and life cycle of Mycoplasma galli-septicum. A5969. Ann. N.Y. Acad. Sci. 143:59-65.
92. Maniloff, J., H. J. Morowitz, and R. J. Barr-nett. 1965. Studies of the ultrastructure andribosomal arrangements of the pleuropneu-monia-like organism. A5969. J. Cell Biol. 25:139-150.
93. Maniloff, J., J. J. Morowitz, and R. J. Barr-nett. 1965. Ultrastructure and ribosomes ofMycoplasma gallisepticum. J. Bacteriol. 90:193-204.
94. Maramorosch, K., E. Shikata, and R. R. Gran-ados. 1968. Structures resembling myco-
plasma in diseased plants and in insect vec-
tors. Trans. N.Y. Acad. Sci. 30:841-855.95. Melchior, D. L., H. J. Morowitz, J. M. Sturte-
vant, and T. Y. Tsong. 1970. Characterizationof the plasma membrane of Mycoplasmalaidlawii. VI. Phase transitions of membranelipids. Biochim. Biophys. Acta 219:114-122.
96. Metcalfe, J. C., S. M. Metcalfe, and D. M.Engelman. 1971. Structural comparisons ofnative and reaggregated membranes fromMycoplasma laidlawii and erythrocytes by x-
97. Metcalfe, S. M., J. C. Metcalfe, and D. M.Engelman. 1971. Structural comparisons ofnative and reaggregated membranes fromMycoplasma laidlawii and erythrocytes usinga fluorescence probe. Biochim. Biophys. Acta241:422-430.
98. Metz., J., and W. Bredt. 1971. Elektronen-mikroskopische Untersuchungen an Myco-plasma hominis (Stamm W463/69). Z. Med.Mikrobiol. Immunol. 156:368-378.
99. Molander, C. W., H. J. Weinberger, and B. M.Kagan. 1965. Growth of staphylococcal Lforms on and through membrane discs of var-
ious pore sizes. J. Bacteriol. 89:907.100. Morowitz, H. J. 1966. The minimum size of
cells, p. 446-459. In G. E. W. Wolstenholmeand M. O'Connor (ed.), Principles of biomo-lecular organization. Little, Brown and Co.,Boston.
101. Morowitz, H. J. 1967. Biological self-replicatingsystems. Progr. Theoret. Biol. 1:35-58.
102. Morowitz, H. J. 1969. The genome of mycoplas-mas, p. 405-412. In L. Hayflick (ed.), Themycoplasmatales and the L-phase of bacteria.Appleton-Century-Crofts, New York.
103. Morowitz, H. J., and R. C. Cleverdon. 1959. Anextreme example of the coding problem,avian PPLO 5969. Biochim. Biophys. Acta34:578-579.
104. Morowitz, H. J., and R. C. Cleverdon. 1960.The irradiation of the pleuropneumonia-likeorganism A5969 with cobalt-60 gamma rays.Radiat. Res. 13:854-856.
105. Morowitz, H. J., and J. Maniloff. 1966. Anal-ysis of the life cycle of Mycoplasma gallisep-ticum. J. Bacteriol. 91:1638-1644.
106. Morowitz, H. J., and T. M. Terry. 1969. Char-acterization of the plasma membrane ofMycoplasma laidlawii. V. Effects of selective
VOL. 36, 1972 287
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/b
r on
24
Dec
embe
r 20
21 b
y 39
.111
.40.
240.
288 MANILOFF AND MOROWITZ
removal of protein and lipid. Biochim. Bio-phys. Acta 183:276-294.
107. Morowitz, H. J., and M. E. Tourtellotte. 1962.The smallest living cells. Sci. Amer. 206:117-126.
108. Morowitz, H. J., H. R. Bode, and R. G. Kirk.1967. The nucleic acids of Mycoplasma. Ann.N.Y. Acad. Sci. 143:110-114.
109. Morowitz, H. J., M. E. Tourtellotte, and M. E.Pollack. 1963. Use of porous cellulose estermembranes in the primary isolation and sizedetermination of pleuropneumonia-like orga-nisms. J. Bacteriol. 85:134-136.
110. Morowitz, H. J., M. E. Tourtellotte, W. R.Guild, E. Castro, and C. Woese. 1962. Thechemical composition and submicroscopicmorphology of Mycoplasma gallisepticum,avian PPLO 5969. J. Mol. Biol. 4:93-103.
111. Munkres, M. and A. Wachtel. 1967. Histochem-ical localization of phosphatases in Myco-plasma gallisepticum. J. Bacteriol. 93:1096-1103.
112. Murant, A. F., and I. M. Roberts. 1971. Myco-plasma-like bodies associated with Rubusstunt disease. Ann. Appl. Biol. 67:389-393.
113. Neimark, H. C. 1970. Division of mycoplasmasinto subgroups. J. Gen. Microbiol. 63:249-263.
114. Panos, C., and S. Rottem. 1970. Incorporationand elongation of fatty acid isomers by My-coplasma laidlawii A. Biochemistry 9:407-412.
115. Pirie, N. W. 1964. The size of small organisms.Proc. Roy. Soc. B 160:149-166.
116. Pirie, N. W. 1969. Possible limitations to thesize of free-living organisms, p. 3-14. In L.Hayflick (ed.), The mycoplasmatales and theL-phase of bacteria. Appleton-Century-Crofts, New York.
117. Plackett, P., S. H. Buttery, and G. S. Cottew.1963. Carbohydrates of some mycoplasmastrains. Recent Progr. Microbiol. Int. Congr.Microbiol. 8:535-547.
118. Ploaie, P. G. 1971. Particles resembling virusesassociated with mycoplasma-like organism inplants. Rev. Roum. Biol.-Botanique 16:3-6.
119. Ploaie, P., and K. Maramorosch. 1969. Electronmicroscopic demonstration of particles re-sembling mycoplasma or psittacosis-lympho-granuloma-trachoma group in plants infectedwith European yellows-type diseases. Phyto-pathology 59:536-544.
120. Pollack, J. D., S. Razin, and R. C. Cleverdon.1965. Localization of enzymes in Myco-plasma. J. Bacteriol. 90:617-622.
121. Pollack, J. D., S. Razin, M. E. Pollack, and R.C. Cleverdon. 1965. Fractionation of Myco-plasma cells for enzyme localization. Life Sci.4:973-977.
122. Quinlan, D. C., and J. Maniloff. 1972. Partialisolation and characterization of a mem-brane-bound DNA growing-point region inMycoplasma gallisepticum A5969. Manu-script in preparation.
BACTERIOL. REV.
123. Razin, S. 1964. Factors influencing osmotic fra-gility of mycoplasma. J. Gen. Microbiol. 36:451-459.
124. Razin, S. 1967. The cell membrane of myco-plasma. Ann. N.Y. Acad. Sci. 143:115-129.
125. Razin, S. 1968. Mycoplasma taxonomy studiedby electrophoresis of cell proteins. J. Bacte-riol. 96:687-694.
126. Razin, S. 1969. Structure and function in myco-plasma. Annu. Rev. Microbiol. 23:317-356.
127. Razin, S. 1969. The mycoplasma membrane, p.317-348. In L. Hayflick (ed.), The mycoplas-matales and the L-phase of bacteria. Ap-pleton-Century-Crofts, New York.
128. Razin, S. 1972. Reconstitution of biologicalmembranes. Biochim. Biophys. Acta 265:241-296.
129. Razin, S., and B. J. Cosenza. 1966. Growthphases of Mycoplasma in liquid media ob-served with phase-contrast microscope. J.Bacteriol. 91:858-869.
130. Razin, S., and I. Kahane. 1969. Hybridizationof solubilized membrane components fromdifferent mycoplasma species by reaggrega-tion. Nature (London) 223:863-864.
131. Razin, S., and S. Rottem. 1967. Identificationof Mycoplasma and other microorganisms bypolyacrylamide-gel electrophoresis of cellproteins. J. Bacteriol. 94:1807-1810.
132. Razin, S., M. Argaman, and J. Avigan. 1963.Chemical composition of mycoplasma cellsand membranes. J. Gen. Microbiol. 33:477-487.
133. Razin, S., L. Gottfried and S. Rottem. 1968.Amino acid transport in Mycoplasma. J. Bac-teriol. 95:1685-1691.
134. Razin, S. H. J. Morowitz, and T. M. Terry.1965. Membrane subunits of Mycoplasmalaidlawii and their assembly to membranelikestructures. Proc. Nat. Acad. Sci. U.S.A. 54:219-225.
135. Razin, S., Z. Ne'eman, and I. Ohad. 1969.Selective reaggregation of solubilized myco-plasma-membrane proteins and the kineticsof membrane reformation. Biochim. Biophys.Acta 193:277-293.
136. Razin, S., B. Prescott, and R. M. Chanock.1970. Immunogenicity of Mycoplasma pneu-moniae glycolipids: a novel approach to theproduction of antisera to membrane lipids.Proc. Nat. Acad. Sci. U.S.A. 67:590-597.
137. Razin, S., M. E. Tourtellotte, R. N. Mc-Elhaney, and J. D. Pollack. 1966. Influence oflipid components of Mycoplasma laidlawiimembranes on osmotic fragility of cells. J.Bacteriol. 91:609-616.
138. Razin, S., B. Prescott, G. Caldes, W. D. James,and R. M. Chanock. 1970. Role of glycolipidsand phosphatidylglycerol in the serologicalactivity of Mycoplasma pneumoniae. Infect.Immunity 1:408-416.
139. Razin, S., B. Prescott, W. D. James, G. Caldes,J. Valdesuso, and R. M. Chanock. 1971. Pro-duction and properties of antisera to mem-
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/b
r on
24
Dec
embe
r 20
21 b
y 39
.111
.40.
240.
VOL. 36, 1972 CELL BIOLOGY O0
brane glycolipids of Mycoplasma pneumo-
niae. Infect. Immunity 3:420-423.140. Reinert, J. C., and J. M. Steim. 1970. Calori-
metric detection of a membrane-lipid phasetransition in living cells. Science 168:1580-1582.
141. Reuss, K. 1967. Influence of fixation on gross
morphology of Mycoplasma. J. Bacteriol. 93:490-492.
142. Reuss, K., C. Plescher, D. Hfulser, and K. Herz-berg. 1967. Morphologische befunde an fix-ierten und unfixierten Mykoplasmen. Zen-tralbl. Bakteriol. Parasitenk. Abt I Orig. 203:121-136.
143. Riggs, A. D. 1966. Studies on Mycoplasma gal-lisepticum. Ph.D. thesis. California Instituteof Technology. Univ. Microfilms, Ann Arbor,Mich. (Diss. Abstr. 66-6838).
144. Rosenberg, S. A., and G. Guidotti. 1969. Theproteins of the erythrocyte membrane, p. 93-109. In. G. A. Jamieson and T. J. Greenwalt(ed.), Red cell membrane structure and func-tion. J.B. Lippincott, Philadelphia.
145. Ross, K. F. A. 1967. Phase contrast and inter-ference microscopy for cell biologists. St.Martin's Press, New York.
146. Rottem, S., and C. Panos. 1969. The effect oflong chain fatty acid isomers on growth, fattyacid composition and osmotic fragility ofMycoplasma laidlawii A. J. Gen. Microbiol.59:317-328.
147. Rottem, S., and S. Razin. 1966. Adenosine tri-phosphatase activity of mycoplasma mem-
branes. J. Bacteriol. 92:714-722.148. Rottem, S., and S. Razin. 1967. Electrophoretic
patterns of membrane proteins of Myco-plasma. J. Bacteriol. 94:359-364.
149. Rottem, S., and S. Razin. 1969. Sugar transportin Mycoplasma gallisepticum. J. Bacteriol.97:787-792.
150. Rottem, S., A. Kalkstein, and N. Citri. 1971.Incorporation of penicillinase into reaggre-
151. Ro-ttem, S., 0. Stein, and S. Razin. 1968. Reas-sembly of Mycoplasma membranes disaggre-gated by detergents. Arch. Biochem. Biophys.125:46-56.
152. Rottem, S., W. L. Hubbell, L. Hayflick, and H.M. McConnell. 1970. Motion of fatty acidspin labels in the plasma membrane of myco-plasma. Biochim. Biophys. Acta 219:104-113.
153. Ryan, J. L., and H. J. Morowitz. 1969. Partialpurification of native rRNA and tRNA cis-trons from Mycoplasma sp. (Kid). Proc. Nat.Acad. Sci. U.S.A. 63:1282-1289.
154. Schwan, H. P., and H. J. Morowitz. 1962. Elec-trical properties of the membranes of thepleuropneumonia-like organism A5969. Bio-phys. J. 2:395-407.
155. Schwartz, J. L., and D. Perlman. 1971. Anti-biotic resistance mechanisms in Mycoplasmaspecies. J. Antibiot. 24:575-582.
156. Sharp, J. T. (Ed.) 1970. The role of myco-
F MYCOPLASMAS 289
plasmas and L forms of bacteria in disease.Charles C Thomas, Springfield, Ill.
157. Shikata, E., K. Maramorosch, and K. C. Ling.1969. Presumptive mycoplasma etiology ofyellows disease. FAO Plant Protection Bull.17:121-128.
158. Slater, M. L., and C. E. Folsome. 1971. Induc-tion of a-glycosidase in Mycoplasma laidlawiiA. Nature New Biol. 229:117-118.
159. Smith, D. W. 1968. Properties of the growingpoint region in bacteria and DNA replicationin PPLO. Ph.D. thesis. Stanford University,Univ. Microfilms. Ann Arbor, Mich. (Diss.Abstr. 68-11,350).
160. Smith, D. W. 1969. DNA replication in Myco-plasma laidlawii B. Biochim. Biophys. Acta179:408-421.
161. Smith, D. W., and P. C. Hanawalt. 1967. Prop-erties of the growing point region in the bac-terial chromosome. Biochim. Biophys. Acta149:519-531.
162. Smith, D. W., and P. C. Hanawalt. 1968. Ma-cromolecular synthesis and thymineless deathin Mycoplasma laidlawii B. J. Bacteriol. 96:2066-2076.
163. Smith, D. W., and P. C. Hanawalt. 1969. Re-pair replication of DNA in ultraviolet irra-diated Mycoplasma laidlawii B. J. Mol. Biol.46:57-72.
164. Smith, P. F. 1956. Quantitative measurementof the growth of pleuropneumonia-like orga-nisms. Appl. Microbiol. 4:254-259.
165. Smith, P. F. 1969. The lipid chemistry of myco-plasmas, p. 469-489. In L. Hayflick (ed.), Themycoplasmatales and the L-phase of bacteria.Appleton-Century-Crofts, New York.
166. Smith, P. F. 1971. The biology of mycoplasmas.Academic Press Inc., New York.
167. Stanbridge, E. 1971. Mycoplasmas and cell cul-tures. Bacteriol. Rev. 35:206-227.
168. Steim, J. M., M. E. Tourtellotte, J. C. Reinert,R. N. McElhaney, and R. L. Rader. 1969.Calorimetric evidence for the liquid-crystal-line state of lipids in a biomembrane. Proc.Nat. Acad. Sci. U.S.A. 63:104-109.
169. Steinberg, P., R. L. Horswood, and R. M.Chanock. 1969. Temperature-sensitive mu-tants of Mycoplasma pneumoniae. I. In vitrobiologic properties. J. Infect. Dis. 120:217-224.
170. Steinberg, P., R. L. Horswood, H. Brunner, andR. Chanock. 1971. Temperature-sensitivemutants of Mycoplasma pneumoniae. II.Response of hamsters. J. Infect. Dis. 124:179-187.
171. Theodore, T. S., J. G. Tully, and R. M. Cole.1971. Polyacrylamide gel identification ofbacterial L-forms and Mycoplasma species ofhuman origin. Appl. Microbiol. 21:272-277.
172. Thomas, L. 1970. The toxic properties of M.neurolyticum and M. gallisepticum, p. 104-109. In J. T. Sharp (ed.), The role of myco-plasmas and L forms of bacteria in disease.Charles C Thomas, Springfield, Ill.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/b
r on
24
Dec
embe
r 20
21 b
y 39
.111
.40.
240.
290 MANILOFF A]
173. Tillack, T. W., R. Carter, and S. Razin. 1970.Native and reformed Mycoplasma laidlawiimembranes compared by freeze-etching.Biochim. Biophys. Acta 219:123-130.
174. Tourtellotte, M. E. 1969. Protein synthesis inmycoplasmas, p. 451-468. In L. Hayflick(ed.), The mycoplasmatales and the L-phaseof bacteria. Appleton-Century-Crofts, NewYork.
175. Tourtellotte, M. E., D. Branton, and A. Keith.1970. Membrane structure: spin labeling andfreeze etching of Mycoplasma laidlawii. Proc.Nat. Acad. Sci. U.S.A. 66:909-916.
176. Tourtellotte, M. E., M. E. Pollack, and R. P.Nalewaik. 1967. Protein synthesis in myco-plasma. Ann. N.Y. Acad. Sci. 143:130-138.
177. Van Golde, L. M. G., R. N. McElhaney, and L.L. Van Deenen. 1971. A membrane-boundlysophospholipase from Mycoplasma laidlawiistrain B. Biochim. Biophys. Acta 231:245-249.
178. Walker, R. T. 1971. Biosynthetic precursors ofsome modified nucleosides in the transfer
N]D MOROWITZ BACTERIOL. REV.
ribonucleic acid of Mycoplasma mycoidesvar. capri. J. Bacteriol. 107:618-622.
179. Wallach, D. F. H., and A. Gordon. 1968. Pro-tein conformations in cellular membranes, p.293-300. In L. Bolis and B. A. Pethica (ed.),Membrane models and the formation of bio-logical membranes. John Wiley, New York.
180. Weibull, C., and B. Lundin. 1963. Morphologyof pleuropneumonia-like organisms and bac-terial L-forms grown in liquid media. J. Bac-teriol. 85:440-445.
181. Yudkin, M. D. 1966. Isolation and analysis ofthe protoplast membrane of Bacillus megate-rium. Biochem. J. 98:923-928.
182. Zucker-Franklin, D., M. Davidson, and L.Thomas. 1966. The interaction of myco-plasmas with mammalian cells. I. HeLa cells,neutrophils and eosinophils. J. Exp. Med.124:521-532.
183. Zucker-Franklin, D., M. Davidson, and L.Thomas. 1966. The interaction of myco-plasmas with mammalian cells. II. Monocytesand lymphocytes. J. Exp. Med. 124:533-542.




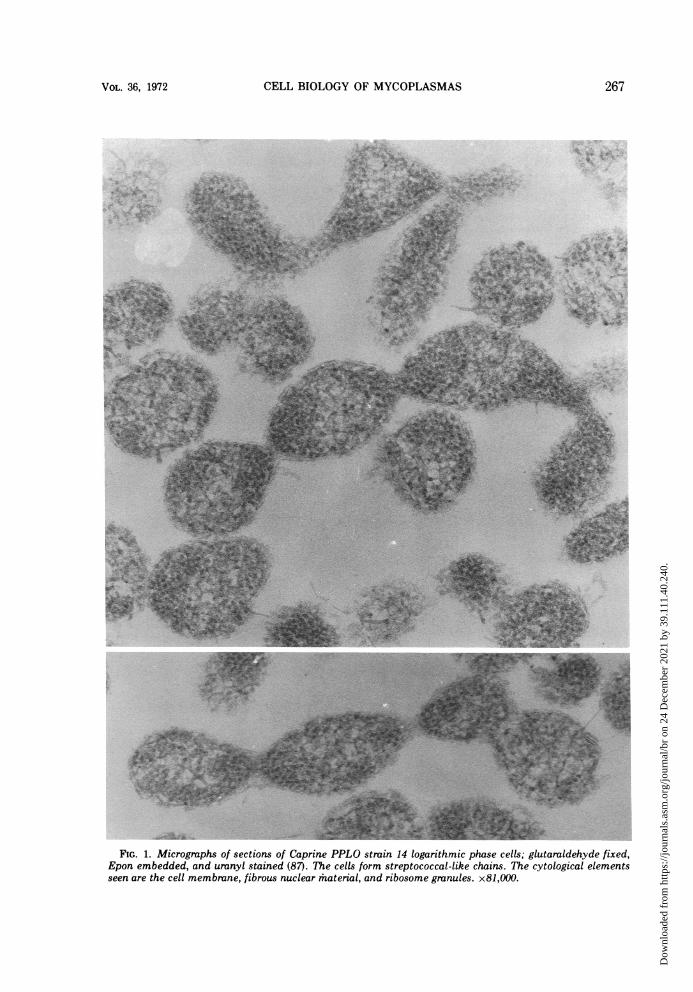
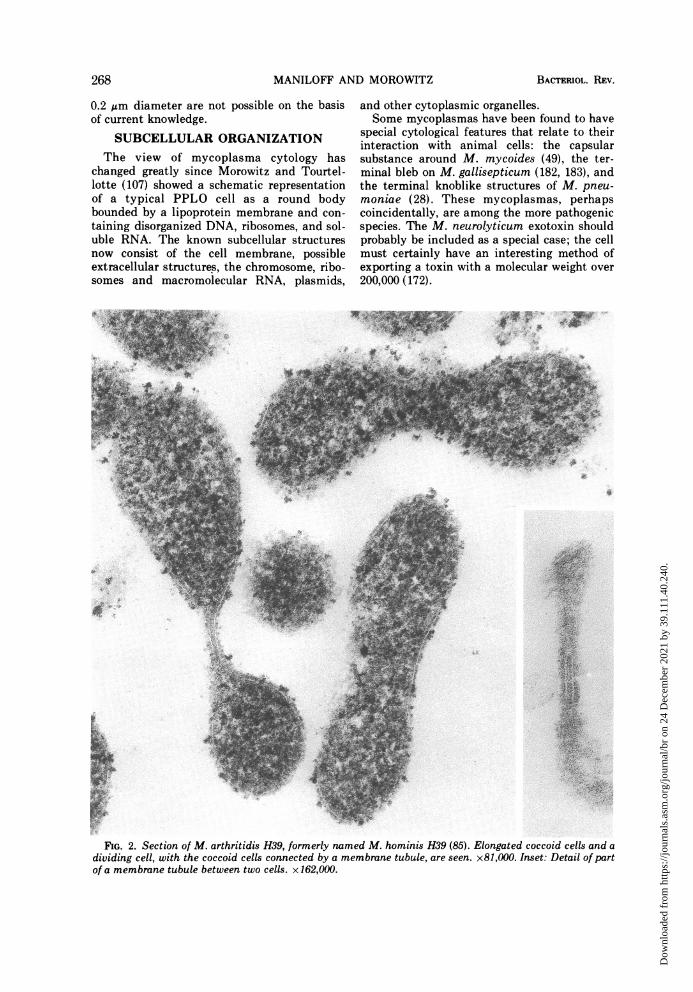
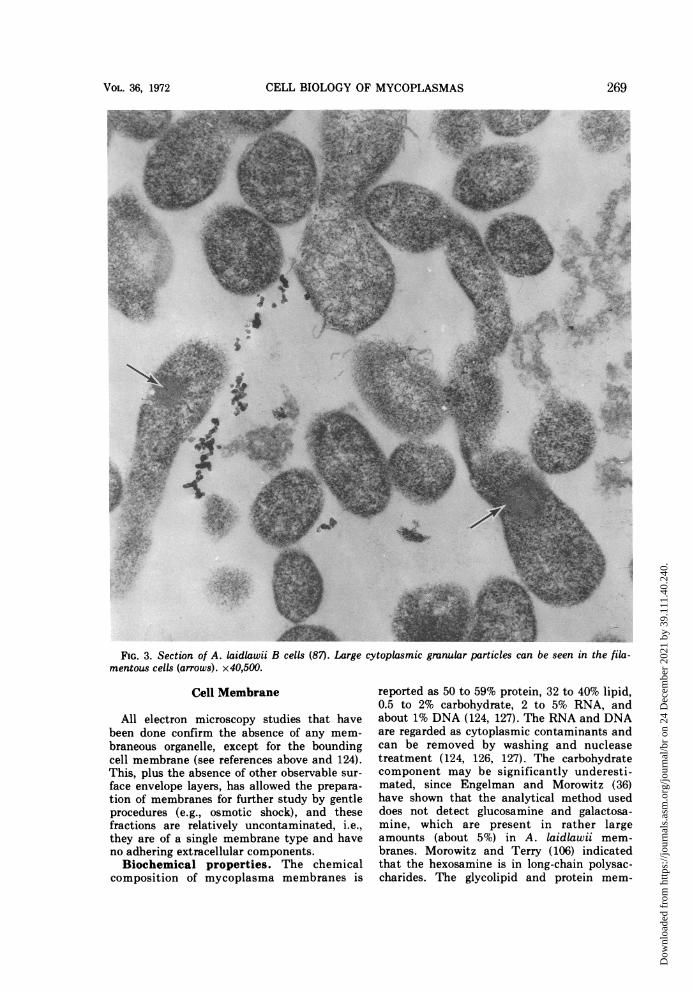
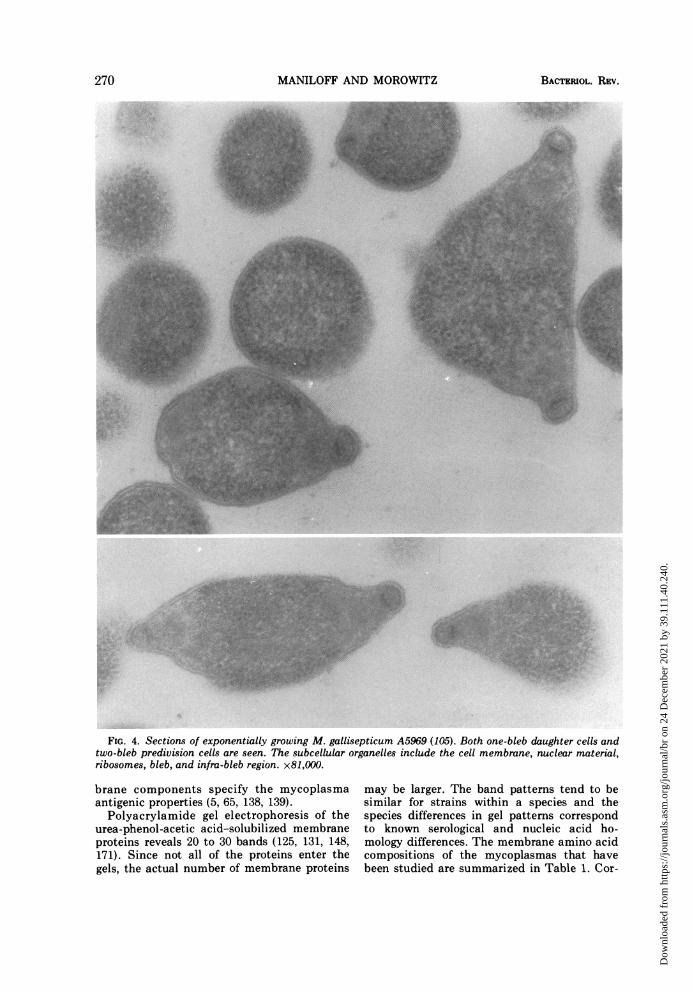









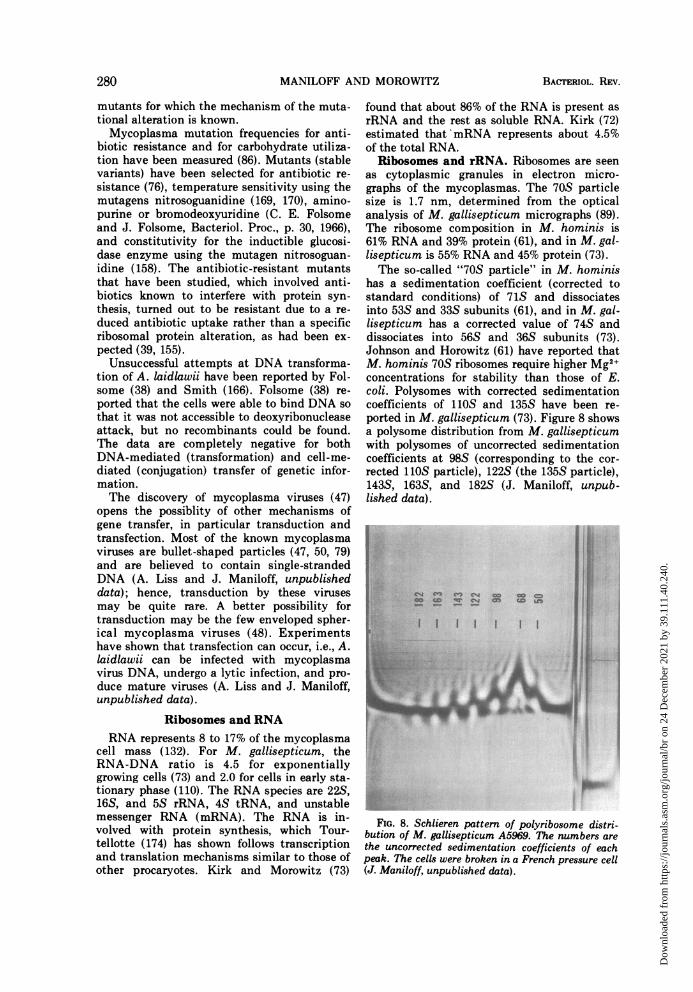


























![6 Molecular Mechanism of Mycoplasma Gliding - A Novel Cell ...miyata/dl/history/miyata_chapter-rev1.pdf · pathogenicity of Mycoplasmas [3, 4, 5]. Interestingly, Mycoplasmas have](https://static.documents.pub/doc/80x56/5f88dce53ed7d44920576775/6-molecular-mechanism-of-mycoplasma-gliding-a-novel-cell-miyatadlhistorymiyatachapter-rev1pdf.jpg)
