Journal of General Microbiology (1982), 128, 2877-2892. Printed in Great Britain 2877 Cell Cycle Dynamics Inferred from the Static Properties of Cells in Balanced Growth By A. L. KOCH’* AND M. L. HIGGINS2 Program on Microbiology, Department of Biology, Indiana University, Bloomington, Indiana 47405, U.S.A. Department of Microbiology and Immunology, Temple University School of Medicine, Philadelphia, Pennsylvania 19140, U . S . A . (Received 15 April 1981 ; revised 9 April 1982) The duration of a morphological phase of the cell cycle is reflected in the steady state distribution of the sizes of cells in that phase. Relationships presented here provide a method for estimating the timing and variability of any cell cycle phase. It is shown that the mean size of cells initiating and finishing any phase can be estimated from (1) the frequency of cells exhibiting the distinguishing morphological or autoradiographic features of the phase ; (2) the mean size of cells in the phase; and (3) their coefficient of variation. The calculations are based on a submodel of the Koch-Schaechter Growth Controlled Model which assumes that (i) the distribution of division sizes is Gaussian; (ii) there is no correlation in division sizes between successive generations; and (iii) every cell division gives rise to two daughter cells of equal size. The calculations should be useful for a wider range of models, however, because the extra- polation factors are not sensitive to the chosen model. Criteria are proposed to allow the user to check the method’s applicability for any experimental case. The method also provides a more efficient test of the dependence of growth on cell size than does the Collins-Richmond method. This is because the method uses the mean and coefficient of variation of the size of the total population, in conjunction with those of the cells in a final phase of the cell cycle, to test potential growth laws. For Escherichia coli populations studied by electron microscopy, an exponential growth model provided much better agreement than did a linear growth model. The computer simulations were used to generate rules for three types of cell phases : those that end at cell division, those that start at cell division, and those totally contained within a single cell cycle. For the last type, additional criteria are proposed to establish if the phase is well enough contained for the formulae and graphs to be used. The most useful rule emerging from these computer studies is that the fraction of the cell cycle time occupied by a phase is the product of the frequency of the phase and the ratio of the mean size of cells in that phase to the mean size of all cells in the population. A further advantage of the techniques presented here is that they use the ‘extant’ distributions that were actually measured, and not hypothesized distributions nor the special distributions needed for the Collins-Richmond method that can only be calculated from the observed distributions of dividing or newborn cells on the basis of an assumed growth law. INTRODUCTION Charting the events of the cell cycle is especially difficult with prokaryotes. This is largely because light microscopy has insufficient resolution and electron microscopy (with or without autoradiography) precludes following a single cell in time. Synchronization methods devised so far have poor resolution and are subject to severe criticism. An alternative approach, proposed here, is to study fixed populations of cells taken from steady state cultures in balanced growth 0022-1287/82/0000-9910 $02.00 0 1982 SGM
Transcript
Downloaded from www.microbiologyresearch.org by
IP: 54.39.17.49
On: Tue, 13 Feb 2018 04:05:58
Journal of General Microbiology (1982), 128, 2877-2892. Printed in Great Britain 2877
Cell Cycle Dynamics Inferred from the Static Properties of Cells in Balanced Growth
By A. L. KOCH’* A N D M . L. H I G G I N S 2 Program on Microbiology, Department of Biology, Indiana University, Bloomington,
Indiana 47405, U.S.A. Department of Microbiology and Immunology, Temple University School of Medicine,
Philadelphia, Pennsylvania 19140, U.S .A.
(Received 15 April 1981 ; revised 9 April 1982)
The duration of a morphological phase of the cell cycle is reflected in the steady state distribution of the sizes of cells in that phase. Relationships presented here provide a method for estimating the timing and variability of any cell cycle phase. It is shown that the mean size of cells initiating and finishing any phase can be estimated from (1) the frequency of cells exhibiting the distinguishing morphological or autoradiographic features of the phase ; (2) the mean size of cells in the phase; and (3) their coefficient of variation. The calculations are based on a submodel of the Koch-Schaechter Growth Controlled Model which assumes that (i) the distribution of division sizes is Gaussian; (ii) there is no correlation in division sizes between successive generations; and (iii) every cell division gives rise to two daughter cells of equal size. The calculations should be useful for a wider range of models, however, because the extra- polation factors are not sensitive to the chosen model. Criteria are proposed to allow the user to check the method’s applicability for any experimental case.
The method also provides a more efficient test of the dependence of growth on cell size than does the Collins-Richmond method. This is because the method uses the mean and coefficient of variation of the size of the total population, in conjunction with those of the cells in a final phase of the cell cycle, to test potential growth laws. For Escherichia coli populations studied by electron microscopy, an exponential growth model provided much better agreement than did a linear growth model.
The computer simulations were used to generate rules for three types of cell phases : those that end at cell division, those that start at cell division, and those totally contained within a single cell cycle. For the last type, additional criteria are proposed to establish if the phase is well enough contained for the formulae and graphs to be used. The most useful rule emerging from these computer studies is that the fraction of the cell cycle time occupied by a phase is the product of the frequency of the phase and the ratio of the mean size of cells in that phase to the mean size of all cells in the population.
A further advantage of the techniques presented here is that they use the ‘extant’ distributions that were actually measured, and not hypothesized distributions nor the special distributions needed for the Collins-Richmond method that can only be calculated from the observed distributions of dividing or newborn cells on the basis of an assumed growth law.
INTRODUCTION
Charting the events of the cell cycle is especially difficult with prokaryotes. This is largely because light microscopy has insufficient resolution and electron microscopy (with or without autoradiography) precludes following a single cell in time. Synchronization methods devised so far have poor resolution and are subject to severe criticism. An alternative approach, proposed here, is to study fixed populations of cells taken from steady state cultures in balanced growth
0022-1287/82/0000-9910 $02.00 0 1982 SGM
Downloaded from www.microbiologyresearch.org by
IP: 54.39.17.49
On: Tue, 13 Feb 2018 04:05:58
2878 A . L . KOCH A N D M . L . H I G G I N S
2
E l 0
Cell mass, rn 2
0 On’25 0.50 0.75 1.0
Age, a
Fig. 1. The canonical size distribution and the analysis of an idealized cellular phase. The cell size (rn) on a linear scale with the birth size chosen at 1 and a division size at 2 is shown at the top. A second scale for cell age (a) is given as the abscissa; it is related to the upper scale by the relationship, rn = 2“, where a goes from 0 to 1. The ordinate is the probability that the cell size will be in the range rn to rn + drn. The cross-hatched region is a hypothetical phase (designated by X because it is entirely internal to the cycle) that initiates at size x, = 1.7, corresponding to age 0.765, and finishes at size x,- = 1.9, corresponding to age 0.926. The cumulative frequency of the entire distribution is 1 and that of phase Xis E = 0.124. For the Xphase, X = 1.796 and qx = 0.0321. From these values of E, Y, q, together with the mean size of the entire population,t = 1.386, x, and xfcould be deduced from the relationships: xf - x, = (In 2)S(qs,Z + 1 ) X 2 / i and In x,-/x, = (In 2)EY/li:
and make careful measurement of the dimensions, morphology, and autoradiographic patterns amongst a large number of cells. If a phase can be defined on the basis of morphology or labelling, then the end points of the phase can be calculated. Thus, from (1) the percentage of cells showing a certain character, (2) their mean, and (3) the coefficient of variation of the distribution in dimensions of cells belonging to this phase, the distribution of cell dimensions as cells enter or leave the phase can be inferred. These size relationships may be converted into time relationships, if desired.
The simplest possible case is shown in Fig. 1. It would apply if all cells arose at division with the same size, grew exponentially in mass and all divided in exactly the same time at exactly the same size. For this ‘canonical’ case, size (m) and age (a) are directly related by m = 2a, and a scale for each is indicated on the abscissa. If a particular phase of the cell cycle began at a particular age (or size) and ended at given age (or size) (for example, as indicated by the cross-hatched region of Fig. l), then the age when cells begin and end this phase could be inferred from the extremes of sizes of cells exhibiting the characteristics of the phase. The same endpoints of the phase can also be deduced from (1) the percentage of cells exhibiting the phase (the area of cross- hatching), (2) their mean size, and (3) the coefficient of variation. For the hypothetical phase corresponding to the cross-hatched area of Fig. 1, such a calculation is given in the legend. Evidently, the latter method is preferable to searching for extremes of range because of biological and statistical artefacts. We propose here methods to allow the same type of interpretation to be made for the less regular cell cycle of actual bacteria. Because in the actual case, as contrasted with the ‘canonical’ case of Fig. 1 , there is variability in the size at cell division, the size distribution of the total population and any particular phase is broader.
As a second example, consider two histograms of the volumes of rod-shaped bacteria taken from a single population of bacteria in balanced growth; one concerns cells with a central constriction, the other concerns all cells in the sample population. Many examples of this kind have been published by the Amsterdam workers (Woldringh et al., 1977; Koppes et al., 1978; Trueba, 1982). Evidently, constriction is a criterion for a cell cycle phase that precedes and terminates with the actual cell division event. From data about the cell lengths or volumes of those cells showing constriction, we might wish to calculate the distribution of lengths or
Downloaded from www.microbiologyresearch.org by
IP: 54.39.17.49
On: Tue, 13 Feb 2018 04:05:58
Charting the prokaryotic cell cycle 2879
volumes of cells in the final act of cell division; i.e., to calculate the extant distribution of mother cells [t$,(m), in the terminology of Painter & Marr (1968)l. The fraction of cells actually dividing at any instant is infinitesimal, while typically, 10-20% of cells are classified as showing constriction, so the actually observed phase of constricting cells ends with division, but does occupy a significant portion of the cell cycle. The actual critical event of leaving the constriction phase (i.e., dividing) defining the 4e(m) distribution occurs with a larger mean size, but a smaller standard deviation (and coefficient of variation) than characterizes the population of cells showing constrict ion.
This problem of calculating the variability of size at division from the variability of size of any observable phase which ends the cell cycle is related to a common problem treated in elementary statistics. The problem arises when data are grouped into classes. By grouping, the computation of means and statistical measure of variations is greatly simplified : however, corrections must be applied (see, for example, Sokal & Rohlf, 1969). The most often used is Sheppard’s correction for the second moment, applied by the formula:
s = [sg2 - h2/12]’
where s is the corrected standard deviation, sg is the standard deviation of the grouped measurements, and h is the grouping interval. If the class measure was the midpoint, the mean resulting from the grouped data is an unbiased estimate of the population mean and may be very accurate if enough measurements have been taken. If the class interval designates one or the other boundary of the group, then the mean must be corrected by 1/2 the grouping interval.
We may draw the analogy between the grouping interval and the range of sizes of a phase of the cell cycle. The frequencies of such a group, and the means and variabilities of size within the group, are experimentally measurable quantities. What we wish to know about a cell cycle phase is the mean size for entry and exit from that phase and its variability. Because the class intervals that we need to consider are sometimes broad and the distributions far from normal, it is not at all evident that a Sheppard-like correction would be appropriate. However, the analogy with Sheppard’s problem of grouping data turns out to be an appropriate one. It will be shown below that a formula like equation (1) can be used to calculate the standard deviation of the size of cells as they move through a phase boundary, the only change necessary being to replace the usual factor 12 by a factor, a, whose numerical values are given for different cases below. By computer modelling, we have found that the coefficient of variation of phase size, together with the value of a suitable for the position in the cycle where the phase occurs in the cycle, can lead to an estimate of the standard deviation of the size at transition into or out of phase with little error. The results of computer calculation also yield factors needed to find the initiating mean size of the phase, and the finishing mean size of the phase. These correction factors (and a) vary depending on whether the phase is near the beginning, middle, or the end of the cell division cycle.
A N A L Y S I S OF P O P U L A T I O N D I S T R I B U T I O N D A T A
Nomenclature of cell cycle phases Basically, the method presented here was developed to apply when synthesis of protoplasm is
continuous, as it probably is for most bacteria. The method will be presented, therefore, in the terminology used in prokaryote biology. Although as explicitly presented, exponential growth is assumed, growth laws between the limits of linear growth and exponential growth could be handled quite similarly. If there are quiescent periods or periods of net degradation in the cycle as can occur in eukaryotic cells, our method would require considerable revisions.
In this paper, we have adopted a set of symbols as consistent as possible with those of our previous work (Koch & Schaechter, 1962; Koch, 1977) and of others. The symbols are defined in Table 1. Briefly, capital letters are used to identify both the phase and the duration of that phase, lower case letters for cell sizes, and capital Greek letters for the frequency of cells in a given phase. Superscript bars indicate means of phases, and the subscripts i andfindicate the mean
Downloaded from www.microbiologyresearch.org by
IP: 54.39.17.49
On: Tue, 13 Feb 2018 04:05:58
2880 A L . KOCH A N D M . L . H I G G I N S
Table I Symbols
Capital letter Small letters
Capital Greek letters
Subscripts i
f
Statistical measures Superscript bar
Y l 3 B z
41-
Notation for cell cycle parameters
Meaning Examples S, C, E, T, X s, c, e, t , .Y
Name & duration of cell phase Size of cell Fraction of total population
in extant phase z, r, H , T, 3
Mean initiating size of coefficient
Mean finishing size of coefficient of variation of phase
of variation of phase
XI? 9x1
-yr, 4 x 1
Mean Skewness, kurtosis of total extant
Coefficient of variation of extant population
X-phase cells
Phases of the cell cycle
Cell division
Normalization of the total cycle
B + C + D = T = 1
e, = d, = t, = 2 P + T + d = T = l
3 = b = t = l 1 1 1
Cell division
Special definitions Current size of a cell = rn Age of cell = a 4 s = q e = 4 Sheppard’s correction factor = OL
Mean phase sizes relative to mean population size = R = ?/i
initiating and finishing sizes of phases. The statistical measures are designated by q for coefficient of variation, y1 for measure of skewness, and /I2 for the measure of kurtosis. Note that the value of f12 for a normal population is 3.
Certain established phases of the cell cycle are indicated in Table 1. These include B, the gap between cell division and the initiation of DNA synthesis, C, the period of DNA synthesis, and D, the gap between the termination of DNA synthesis and cell division. In addition, the total cell cycle is indicated by T. Without lack of generality, ?and 6 have been set to 1 and $to 2. We also use S for phases that start with cell division and E for phases that end with cell division.
The symbol X is used to denote any phase of the cell cycle of interest. Consequently, this symbol will change its meaning in the course of the paper. At times, it will be set equal to well established prokaryote phases, B, C or D and at others to that phase where the cells show terminal constriction. But this list is not inclusive; the whole point of the paper is that the method of data analysis developed here may be applied to measurements based on any classification scheme, even those that have not been yet discovered.
The correction factors presented here have been calculated on the assumption that cell division splits the cytoplasm precisely in two. This is a great simplification from the mathematical and numerical point of view. In a number of bacteria, the ratio of a daughter’s size to its mother’s size has a coefficient of variation less than 10% (e.g., Trueba, 1982). A special symbol q, bearing no subscript, is defined for the coefficient of variation of the size of cell distribution at division; i.e., the coefficient of variation of the #,(m) distribution. Therefore, the coefficient of variation of the division size distribution and the birth size are equal and qsi = qef - qk = q4f = q.
The caveat of the current paper is that the rules and graphs presented here only apply to those phases that lie entirely within a single T-period and do not extend into earlier or later cell cycles.
-
Downloaded from www.microbiologyresearch.org by
IP: 54.39.17.49
On: Tue, 13 Feb 2018 04:05:58
Charting the prokaryotic cell cycle
The Koch-Schaechter method for cell cycle statistics
It can be assumed that cell division is the normal response of the cell when it has succeeded in growing sufficiently. It is more controversial, but not unreasonable, to assume that under constant conditions the mass of an individual cell grows approximately exponentially with time ; i.e., the rate of growth of an individual cell is directly proportional to its own mass or its content of ribosomes. If a cell divided precisely when it achieved a critical size, then the frequency distribution of cell sizes in a population of cells in asynchronous balanced growth would be as indicated in Fig. 1. There are four times as many cells in the smallest size class as in the largest for the canonical distribution. One factor of twofold arises simply because one cell divides into two cells. The second factor of two arises because the population distribution represents a census of the culture at a given instant of time, but classified on a basis of cell size. Obviously it takes twice as long for a small, newborn cell to increase its size a unit amount than it does a large cell that is just about to divide and has twice the mass, number of ribosomes, etc., needed to carry out the necessary anabolism. This means that the smallest cells will spend twice as long in a size category and will, therefore, be twice as highly represented as the largest cells in the population distribution. Because the birth size has been defined as 1 and the division size as 2, this distribution can be described by:
288 1
8(m) = 2/m2 dm; 1 < rn < 2
Of course, actual populations of prokaryotes do show a small degree of variation in the size that they attain as they divide, This variability causes a further widening of the population size distribution above that shown in Fig. 1 (see Fig. 3, later). The degree of broadening of the distribution depends on variability of the size at cell division.
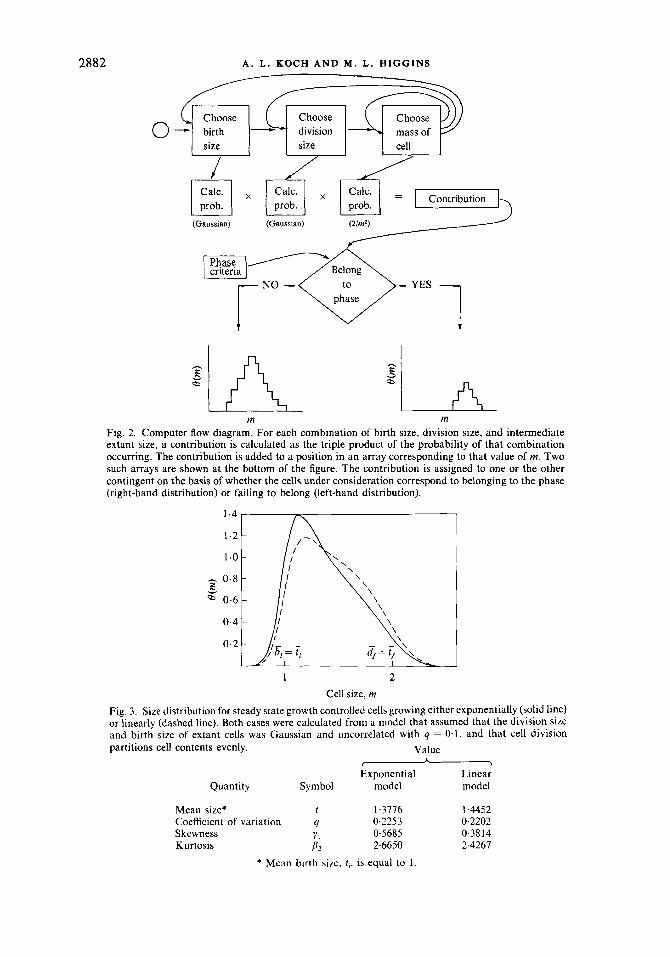
For the calculations presented below, we have assumed a particular sub-model of the general growth controlled (GC) model of Koch & Schaechter (1962). This specific model is designated SGC (standard growth control). The program computes a cell size distribution by summing many distributions each like Fig. 1. These sub-distributions differ in their minimum and maximum size. Figure 2 is a flow chart for the program. First, the computer successively chooses birth sizes varying systematically from three standard deviations below the mean to three standard deviations above the mean. For each choice, it makes weighted contributions to the population distribution with the corresponding probability value calculated from the normal distribution. For each choice of birth size, the computer considers a range of division sizes from three standard deviations below the mean to three standard deviations above the population mean and weights them as it did the birth sizes; i.e., according to the value drawn from a Gaussian table of probabilities. In this way the computer has taken into account the variation of birth and division sizes. For each combination of birth and division size, there are bacteria that arose at various times before the instant of sampling and have grown to some intermediate size. Their contribution to the frequency distribution depends on the inverse square law of equation (2), as depicted in Fig, 1. The computer adds these contributions into appropriate memories indicated on the right-hand side of Fig. 2 to form the cell size histogram. In this way, the distribution shown by a solid line in Fig. 3 was generated. These calculations were laborious and involved consideration of 601 birth sizes, 601 division sizes and 300 potential sizes of cells, giving rise to a total of about lo8 contributions for each distribution.
The distribution of cell sizes if increase in cell size occurs according to a linear growth law instead of an exponential law, is shown by a dashed line. The computer program in this case was operated as indicated above, except that the inverse square weighting factor was replaced with a negative exponential weighting factor. For linear growth, size and age are interchangeable. It is well known that for the canonical case, the age distribution is a negative exponential (Koch & Schaechter, 1962). This relationship was originally proposed by Euler in 1775 (Koch & Blumberg, 1976). The calculations presented in this paper supersede earlier calculations and have increased precision because finer intervals of birth and division sizes were used than in previous work (Koch, 1966, 1977). In both this paper and Koch (1977), a range of sizes extending three standard deviations below and above the mean were considered.
Downloaded from www.microbiologyresearch.org by
IP: 54.39.17.49
On: Tue, 13 Feb 2018 04:05:58
2882
0
A . L . KOCH A N D M . L . H I G G I N S
size size cell
prob. prob.. prob. (Gaussian) (Gaussian)
mf-0 NO -
1 m m
Fig. 2. Computer flow diagram. For each combination of birth size, division size, and intermediate extant size, a contribution is calculated as the triple product of the probability of that combination occurring. The contribution is added to a position in an array corresponding to that value of m. Two such arrays are shown at the bottom of the figure. The contribution is assigned to one or the other contingent on the basis of whether the cells under consideration correspond to belonging to the phase (right-hand distribution)
h
5 rn
or failing to belong (left-hand distribution).
1 2
Cell size, m Fig. 3. Size distribution for steady state growth controlled cells growing either exponentially (solid line) or linearly (dashed line). Both cases were calculated from a model that assumed that the division size and birth size of extant cells was Gaussian and uncorrelated with q = 0.1, and that cell division partitions cell contents evenly. Value
Exponential Linear Quantity Symbol model model
Mean size* t 1-3776 1.4452
Skewness Y1 0.5685 0.3814 Kurtosis P . 2.6650 2-4267
* Mean birth size, t , , is equal to 1.
Coefficient of variation q 0.2253 0.2202
Downloaded from www.microbiologyresearch.org by
IP: 54.39.17.49
On: Tue, 13 Feb 2018 04:05:58
Charting the prokaryotic cell cycle 2883
This method of treating the cell cycle is consistent with the Collins & Richmond (1962) method. The two approaches are the inverse of each other. As usually used, both methods are deterministic and assume that a cell of a certain size will grow at the same rate as any other cell of the same size, independently of how long an interval has passed since the last cell division, or the interval that will pass until the next cell division.
Relationship to the Collins-Richmond treatment
Collins & Richmond (1962) derived an equation for calculating the growth rate as a function of cell size from the measurement of size distributions of cells taken from balanced, asynchronous cultures. Their equation (Painter & Marr, 1968) depends on knowing three distri- butions : the size distribution of the extant total population, the size distribution of a sample of mothers, and the size distribution of a sample of babies. In practice, the first distribution is measured and the other two are assumed. The approximation has usually been to assume apriori that both are narrow normal distributions. It has been generally assumed that both the distributions of division and birth sizes have the same coefficient of variation. This requires that division creates equal sized daughters. These approximations are needed because the required division and birth distributions have not been measured. For many applications, the exact shape of these distributions is not critical. Examples where this was the case were presented by Harvey et al. (1967). The Collins-Richmond technique yields the average velocity of size increase as a function of cell size. An early study reported that the velocity of size increase was very low for both extremely small cells and extremely large cells (Harvey et al., 1967), but substantially proportional to cell volume for the bulk of the cells.
In their critical review of the mathematics of microbial populations, Painter & Marr (1 968) consider several types of frequency functions for different phases of the cell cycle (see their Table 1). It is to be noted that all the distributions formulated here correspond to those classified as extant size distributions. Powell (1 964) referred to them as 'realized' distributions. This choice means that there is a direct correspondence between the experimentally observed size distributions of cells in E phases, S phase, the total population, and the limiting distributions of cells that are just dividing, +,(m), or are newly arisen, +&n). It is necessary to point out that the distributions needed for the Collins-Richmond technique are quite different from those used here. For this technique to be used on a completely experimental footing, these must be computed from the extant distributions by assuming a growth law. Even if an exponential growth law is assumed, the conversion is quite complicated [see Equation (27) of Powell (1964)l. Otherwise the appropriate sample of cells must be followed in time until they divide and a distribution prepared from the sizes observed just prior to division, +(m), and immediately after division, has occurred, +(rn).
Choice of growth law
The application of the Collins-Richmond technique has led to a variety of interpretations (Collins & Richmond, 1962; Harvey et al., 1967 ; Zusman et al., 1971 ; Cullum & Vincente, 1978 ; Trueba, 1982). However, we suggest that the published evidence is consistent with exponential growth of individual cells, but also indicates that a low percentage of cells are abnormal and grow much more slowly than the remainder. The abnormal cells are usually either very large or very small (Koch, 1980). When only the mid-range of cell sizes are considered, there is little support for a linear model of growth.
For the purposes of analysing cell cycle phases, however, the choice of a growth law is of relatively little importance. Even if the growth in cell size were linear, instead of exponential, little effect can be expected on the magnitude of the correction factors presented below. This, as noted by several authors previously, is because there is very little difference in cell mass, rn, between, for example, a linear and an exponential mathematical function when both functions are constrained to go from m of 1 to rn of 2 as the independent variable (the age of the cell), a, goes from 0 to 1. Thus the greatest difference between the lines m = 1 + a and m = 2" in the region of 0 < a < 1 is only slightly greater than 6%. Consequently, the course of cell enlargement is not much different in the two cases. From the distributions for cell sizes shown in
Downloaded from www.microbiologyresearch.org by
IP: 54.39.17.49
On: Tue, 13 Feb 2018 04:05:58
2884 A . L. KOCH A N D M . L . HIGGINS
*-O r I
0.5 1.0
*-. Age, a
Fig. 4. Linear, bilinear, and exponential growth. Cellular growth curves are shown for the linear and exponential (bold line) case and for those two bilinear cases deviating least from the exponential mode of growth. The residual sums of squares of differences (RSS) between the exponential case and all possible bilinear modes of growth is plotted versus the relative cell age at which the growth rate of the bilinear model doubles. Linear growth is the special case where this doubling occurs at a = 0 or a = 1. The lines marked 0.28 and 0.85 are the bilinear growth curves, in which the growth rates double at these fractions of the cell cycle; these are the cases that most closely approach the case of exponential growth.
Fig. 3, it can be seen that there is really very little difference between the linear and the exponential model. This impression is reinforced from inspection of the statistical parameters of the distributions given in the legend to Fig. 3. The difference between the two models is that it takes a constant time for cells to pass through a size interval for linear growth, while this time is inversely proportional to cell size for exponential growth.
The differences in the statistical parameters of the size distributions provide a test for the linear model versus the exponential one. Although the mean values differ, this is only of value in distinguishing between the growth laws experimentally where there are accurate independent measurements of the mean birth or division size. This is an important point and is a critical advantage of the present method (see below). The small difference in y1 and p2 requires accurate data, unbiased by truncation or special properties of a few abnormal cells. The coefficient of variation, qr, which is the most accurately measurable and accessible quantity, shows almost no difference between linear and exponential growth.
Today, the linear model in the original form, in which the growth rate is assumed to double at the instant of cell division, has been revised to assume that the growth rate doubles at a fixed age in the cell cycle (Kubitschek, 1970, 1981). Such ‘bilinear’ models may approach the simple exponential much more closely than does the linear (Fig. 4). This figure shows a linear (thin line), an exponential (bold line), and two bilinear growth curves. The ordinate values on the curved heavy line marked RSS (residual sums of squares) are the sums of the squares of the differences between cell sizes at the same ages for exponential growth and bilinear growth. Each point on this curve corresponds to the sum for ages ranging from 0 to 1, but for different bilinear models in which the doubling point for the growth rate was shifted systematically from a = 0 to a = 1 . This curve goes through two minima corresponding to the two bilinear growth curves shown (where the doubling point occurs at a = 0.28 and at a = 0.85). In a number of cases in the literature where bilinear growth has been suggested, the data were consistent with growth in which the break occurs at about the age of the second minimum of the RSS curve. Consequently, it is felt that the methods developed below should apply with an extremely small error even if volume growth takes place by a bilinear mechanism.
PROCEDURE
The following operational approach is proposed. (1) Measure sufficient cells (at least 500) from a population in a well defined state of balanced growth in order to define accurately the
Downloaded from www.microbiologyresearch.org by
IP: 54.39.17.49
On: Tue, 13 Feb 2018 04:05:58
Charting the prokaryotic cell cycle 2885
1-39 I
C ' 1.37 -
0.6
-- x v
v) m
0.5 3
m
T
h s v)
La
*
.- 0
f L-
1-36 1 0.20 1 2 ' I I I I 1 1
0.05 0.10 0.15 4
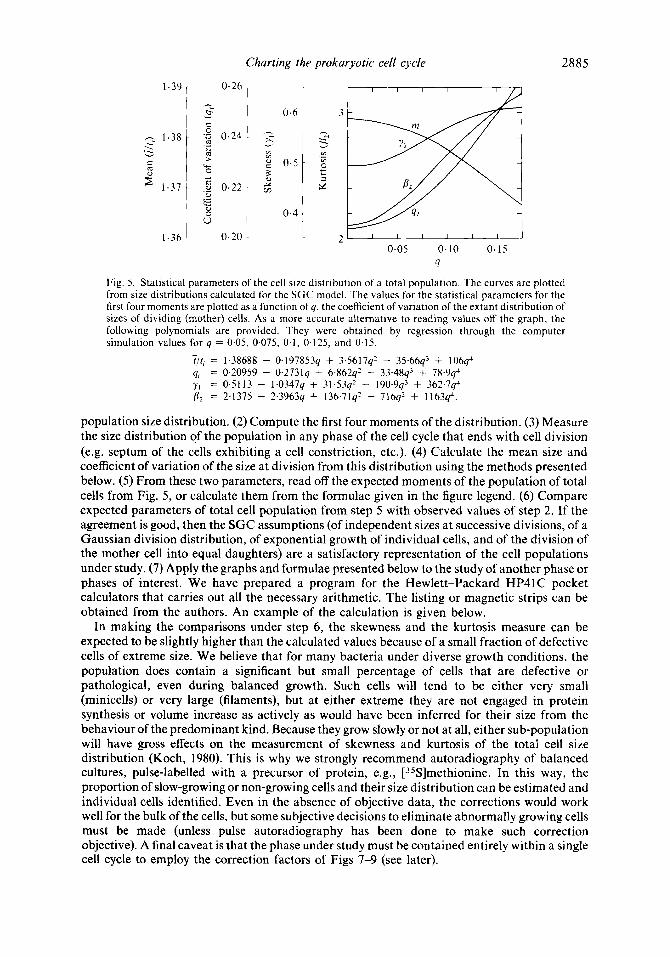
Fig. 5. Statistical parameters of the cell size distribution of a total population. The curves are plotted from size distributions calculated for the SGC model. The values for the statistical parameters for the first four moments are plotted as a function of 4, the coefficient of variation of the extant distribution of sizes of dividing (mother) cells. As a more accurate alternative to reading values off the graph, the following polynomials are provided. They were obtained by regression through the computer simulation values for q = 0.05, 0.075, 0.1, 0.125, and 0.15.
population size distribution. (2) Compute the first four moments of the distribution. (3) Measure the size distribution of the population in any phase of the cell cycle that ends with cell division (e.g. septum of the cells exhibiting a cell constriction, etc.). (4) Calculate the mean size and coefficient of variation of the size at division from this distribution using the methods presented below. (5) From these two parameters, read off the expected moments of the population of total cells from Fig. 5 , or calculate them from the formulae given in the figure legend. (6) Compare expected parameters of total cell population from step 5 with observed values of step 2. If the agreement is good, then the SGC assumptions (of independent sizes at successive divisions, of a Gaussian division distribution, of exponential growth of individual cells, and of the division of the mother cell into equal daughters) are a satisfactory representation of the cell populations under study. (7) Apply the graphs and formulae presented below to the study of another phase or phases of interest. We have prepared a program for the Hewlett-Packard HP41C pocket calculators that carries out all the necessary arithmetic. The listing or magnetic strips can be obtained from the authors. An example of the calculation is given below.
In making the comparisons under step 6, the skewness and the kurtosis measure can be expected to be slightly higher than the calculated values because of a small fraction of defective cells of extreme size. We believe that for many bacteria under diverse growth conditions, the population does contain a significant but small percentage of cells that are defective or pathological, even during balanced growth. Such cells will tend to be either very small (minicells) or very large (filaments), but at either extreme they are not engaged in protein synthesis or volume increase as actively as would have been inferred for their size from the behaviour of the predominant kind. Because they grow slowly or not at all, either sub-population will have gross effects on the measurement of skewness and kurtosis of the total cell size distribution (Koch, 1980). This is why we strongly recommend autoradiography of balanced cultures, pulse-labelled with a precursor of protein, e.g., [35S]methionine. In this way, the proportion of slow-growing or non-growing cells and their size distribution can be estimated and individual cells identified. Even in the absence of objective data, the corrections would work well for the bulk of the cells, but some subjective decisions to eliminate abnormally growing cells must be made (unless pulse autoradiography has been done to make such correction objective). A final caveat is that the phase under study must be contained entirely within a single cell cycle to employ the correction factors of Figs 7-9 (see later).
Downloaded from www.microbiologyresearch.org by
IP: 54.39.17.49
On: Tue, 13 Feb 2018 04:05:58
2886 A . L . KOCH AND M . L . HIGGINS
CORRECTION FACTORS FOR CELL CYCLE PHASES
The computer program used in the present study is a revision of one originally constructed to study the phase of DNA synthesis (Koch, 1977). For the more general purposes of the present paper, the program was reworked into several variants, spanning a range of assumptions as to criteria for entry and exit of cells from the phase of interest. The modified programs calculate, as outlined above, the contributions of cells from birth and division size classes for each intermediary size. But in addition (Fig. 2), the computer now checks each contribution to see if it belongs to the phase under consideration. If a contribution [i.e. the triple product of the probability of the birth size, the division size, and the inverse square probability from equation (2)] belongs to a cohort of cells that satisfy the criteria for inclusion within the phase, the contribution is added into the register for that size of cell in the memory bank indicated on the right-hand side of Fig. 2. In the opposite case, when the cells do not belong to the phase in question, the contribution is added to the registers indicated on the left of Fig. 2. When all combinations have been made and totalled, the computer then calculates a variety of items concerning the distribution of cells within the phase. These include mean, standard deviation, etc., but also include items allowing the generalizations presented in the sections below to be applied.
Phases ending with cell division, E phases
The phases ending the cell cycle will be considered first, since morphological changes are generally observable in cells about to divide and therefore the method of this section is always applicable. E-phases are also considered first because their treatment is a prelude to predicting the properties of the entire population. This permits an essential step in the method-the comparison between the predicted and observed total size distributions in order to decide whether the method is appropriate. If so, then the method can be applied to other kinds of phases.
Distributions of cell sizes were calculated as indicated above for five choices of the coefficient of variation for the dividing population (i.e., q = 0~05,0~07,0-09,0~11 and 0.13). This covers the range of variability in size of division seen in experimental studies with many prokaryotes. For each value of q, the computer was asked to calculate the distribution of the population of E- phase cells assuming that the duration of Econstituted the last 5, 10, 15,20, or 25% of the mean cell cycle time, F. In so doing, we have tacitly assumed that no matter whether a cell is larger or smaller than average at division, it will take just as long for the cell to pass through the terminal phases of the cell cycle.
For each of these 25 cases, data from computer simulation were used to compare the fraction of the total population in the terminal phase, (N), the coefficient of variation of sizes in the phase (qe), and in the total population (4). It was found empirically that:
q = (qe2 - H2/14-4)4 (end of cycle) (3) This expression can be accurately used in calculating q over the entire range of durations of the E-phase and over the range of q values. Thus, an equation of the form of equation (l), but with 14.4 replacing the usual 12, can be used to estimate the coefficient of variation of the division size distribution.
Correcting the mean of the class to the size of the dividing cell is not quite as simple, but Fig. 6 gives a graph of appropriate factors. More precisely, the ratio of the mean size at the finishing of an E-phase to the mean size of the phase is given by:
The mean size for initiating an E-phase is given by: e,/Z = 0.9922 + 0.1648q - 0-431H - 0.05qH (end of cycle) (5)
As is evident from the system of nomenclature (Table l), D and Tare also phases that end the cell cycle. Therefore, ef = d, = tp since all are symbols for the mean size at cell division. The
Downloaded from www.microbiologyresearch.org by
IP: 54.39.17.49
On: Tue, 13 Feb 2018 04:05:58
Charting the prokaryotic cell cycle 2887
.- Y
.-
.- A u L W .-
/ End of cycle , . , ; ’ ”
I I 1 I I , l . J \ ’ . ,
0-9 0 10 20 30
2, Cells in class (96) Fig. 6. Factors for estimating the mean initiating and finishing sizes of cell phases that either end or start with a cell division event. These factors are to be multiplied with the mean size of cells exhibiting a morphological or autoradiographic phase. The lines in the upper half give correction factors to compute the mean finishing size of phases that either start or end with cell division. Those in the lower half give correction factors for the mean initiating size for phases that either start or end with cell division. The correction factors for phases terminating with cell division are partially dependent on q (dashed line, q = 0.05; dotted line, q = 0.1 1) and linear interpolation is necessary and sufficient. For phases starting with cell division, the value of q is much less critical and no interpolation is necessary. The equations (4), (9, (7), and (8) (see text) can be used more accurately than the figure for calculating mean initiating and finishing sizes when q is in the range 0.05 to 0.1 1. When q is larger (0.15@25), different regressions are appropriate; these can be obtained from the authors.
mean initiating size of an E-phase, ei, may or may not have any intrinsic relationship to other phases of the cell cycle.
Phases starting with cell division, S phases
In eukaryotic systems, the phase extending from the end of mitosis to the beginning of DNA synthesis is designated G1. It appears to be the important phase in the regulation of the cell cycle (see, e.g. Stubblefield, 1981). Many hormones and growth factors seem to act on cells in this phase. In slow-growing bacterial cells, there can be an interval between cell division and the initiation of DNA synthesis, designated B. In many circumstances, B may be negative and vary widely for individual cells within the same population. There is at least one morphological phase of short duration that seems to be initiated at cell division, or very close to it. This is the band splitting in Streptococcus faeciurn, which in slow growing cultures occurs in most cells immediately after cell fission (M. L. Higgins and A. L. Koch, unpublished observation).
For phases starting with cell division, the same range of values of q was examined as in the previous section. As before, the computer output where the duration of S constituted 5 , 10, 15, 20,25% of the mean cell cycle time, r, was examined to develop an empirical relationship for the interpretation of the simulation data. Over these ranges, the analogue of the Sheppard’s correction formula is:
q = [qs‘ + C2/44I7 (start of cycle) (6)
Because the Sheppard’s correction factor for S-phase is so large, unless the phase under consideration includes a very large fraction of the population, q will be very nearly equal to qs. Thus, the observed variation in the size of a cell in a starting phase is almost equal to the variation in birth size. To compute the mean values at the end of an S-phase, the solid line in the
1
Downloaded from www.microbiologyresearch.org by
IP: 54.39.17.49
On: Tue, 13 Feb 2018 04:05:58
2888 A . L . KOCH AND M . L . H I G G I N S
upper part of Fig. 6 can be consulted or, more accurately, the following regression can be employed.
sfl? = 1.00035 + 0-24526C + 0-1233Z2 (start of cycle) (7) The mean size at initiation of an S-phase can be calculated from the solid line in the lower part of Fig. 6 or is given by:
Of course, in this case, si is an estimate of bi or one-half of df.
Phases contained entirely within the cell cycle, X phases A quite different problem concerns phases of the cell cycle that neither originate nor
terminate with cell division. The phase of D N A synthesis has generally been assumed to be an example of such an X-phase in slow-growing cells. For this class of problem, the computer was given parameters concerned with the assumed duration and position of the phase of interest in the cell cycle, at various fixed times relative to either the start or the end of the cell cycle; the computer was then asked to calculate the distribution of cells in phase X . We carried out computations under two different assumptions : (1) that the duration of the interval between the end of Xand the end of the cell cycle has no variability, and (2) that the interval between division and the beginning of the X phase has no variability. Except for some caveats described below, these two extremes give rise to nearly the same rules for finding the mean initiating and finishing sizes x, and xf, the variability of these sizes (qsxi and qxf) as well as the duration of the phase, X . For the case in which the X phase is identified with the D N A synthesis phase, these two assumptions correspond, respectively, to the assumption that D has no variability and that B has no variability (see Koch, 1977). In these cases, all the variability must reside in the other phase. These two cases, of course, do not exhaust the possibilities. Other reasonable possibilities include cases where there is variation in the duration of the X phase and where the phase starts in each cell at random at a certain mean size with a certain coefficient of variation, independently of the size of that cell at the start or end of its cell cycle.
A condition that must be met to use the graphs and regressions presented in this section is that the phase must not extend outside the duration of a single cell cycle. For example, the method as described would not give accurate results in those cases where the variability in the start of D N A synthesis is so large that D N A synthesis in part of the population starts in a different cell generation from that in which it is completed. The method would be inapplicable even if in most cell cycles the C phase is contained within a single cell cycle.
Consequently, the computational method given below is more likely to be successfully applied if the phase is short and occurs near the middle of the cycle. A rough rule is that Xi?, the mean size of the phase relative to the mean size of cells of the total population, should be somewhere between 0.9 and 1.25.
Figures 7 and 8 show the value of the Sheppard’s correction factor, a, as a function of R = x / t . As above, the value of a from the each simulation was calculated from:
a = P / ( q , * - qxi?) (internal to cycle) (9)
It can be seen that the factor ranges between the limiting values of 44 and 14.4 presented in previous sections, appropriate to the phases that start or end with a cell division event. It is also apparent that a is dependent almost solely on Fir. Thus, for those cases in which the duration of Xis assumed to be the same for all cells in the population, the user can calculate the coefficient of variation and the size of initiating and finishing of the phase from three experimentally accessible quantities: q x , q, and Z/T.
The duration of an Xphase can be calculated from the following empirical relationship, which was discovered by inspection of the computer output: X = E l f . This relationship appears to have no simple mathematical basis. That it is quite accurate, however, can be seen from Fig. 9. The deviation from this simple relationship is less than & 10% as q is varied, as Xis varied from 0 to 0-6, or as the interval between X and either flanking cell division event is varied. The graph
Downloaded from www.microbiologyresearch.org by
IP: 54.39.17.49
On: Tue, 13 Feb 2018 04:05:58
Charting the prokaryotic cell cycle 2889
26
18
16
14
8 50
E g 40
30
c1
... c
8 5 20 cd a a
v)
2 10
d
I I 1 I I I
I I I I I I I 0.8 0.9 1.0 1.1 1.2 1 - 3 0.7 0.8 0.9 1.0 1.1 1.2 1.3 1.4
R = X/7 R = X I 7
Fig. 7 Fig. 8 Fig. 7. Sheppard’s correction factor (a) for phases interior to the cell division cycle. Calculations are for q = 0.1 (solid lines); q = 0.05 (dashed lines). The values of OL are calculated, except for one line, on the assumption that there is no variation in the interval from the end of variable phase X to cell division. This exception (dotted line) applies when q = 0.05 and there is no variation in the time from cell division to the initiation of phase X.
Fig. 8. Sheppard’s correction factor for short phases. The value of a is given for a phase which can occur at any point in the cell cycle but maintains accurate relationship to the next division (continuous line) or to the previous division (dotted line). The calculations are for q = 0.05 and X = 0.1. For very short phases, R = T//fcan vary from 0.693 to 1.386. The predictions of the model both where the phase X times with no error the next division, and where the previous division times the initiation of the Xphase are in agreement for a considerable part of the range of .Y/i. They differ at lower values of R. This corresponds to cases where there is a long gap before the next cell division. Then in an appreciable fraction of cells, the phase must be started in the previous cell cycle for the cases where the X-phase times the next division.
I I I I I
0.8 0.9 1.0 1.1 1.2 1.3 1.4 R = .?/i
Fig. 9. Error in the approximation X = ZUji . The ordinate is a correction factor which, if multiplied by B.Y//T, yields the duration of the phase under study in units of fractions of a mean generation time. Factors are given for q = 0.05 (dashed line) and 0.1 (solid line) and for X = 0.1 (thin line) and 0.5 (thick line). These calculations are based on the assumption that the X phase times cell division. The origin of each line corresponds to the situation in which the phase terminates with cell division. The lines end at arrow heads when the interval from the end of the phase to cell division is sufficiently long so as to push IO%of the phase Xcells into the preceding cell cycle. Consequently, the factors given here should apply to data if the distribution of cells within the phase versus cell size is monomodal.
Downloaded from www.microbiologyresearch.org by
IP: 54.39.17.49
On: Tue, 13 Feb 2018 04:05:58
2890 A: L . KOCH A N D M. L. HIGGINS
can be employed when higher accuracy is required, but in most cases no correction factor is needed.
A P P L I C A T I O N O F T H E M E T H O D TO Escherichiu coh'
In recent years, extensive and careful electron microscope studies of the properties of balanced populations of E. coli have been reported by the group in Amsterdam. These studies have been based on the measurements of tens of thousands of images of bacteria. At this point, these published data can be used to justify the application of the method to a variety of strains of E. coli grown under a variety of conditions. In two papers (Woldringh et ul., 1977 ; Koppes et al., 1978), data are presented which are adequate to test the applicability of the procedure. These workers measured the distribution of sizes of the sub-population of cells possessing constrictions as well as that of the total population of cells. The data base consists of 14 cell samples where between 632 and 3163 cells were measured and the first two moments of the total and constricting populations were reported. In these two papers, 21 279 cells in total were measured. While the mean size at division varied considerably with the growth conditions and the strain, the coefficient of variation of the distribution of constricting cells was quite constant with a mean value of 0.0853 and with a standard deviation of 0-0197.
As a numerical example of our method, consider the set of 1549 cells of E. coli B/rA growing with a 109 min doubling time reported on the first line of Table 2 of the paper of Koppes et al. (1978). For cells showing constriction, the coefficient of variation, qe, 'was reported as 0.084 and the fraction of cells in the constriction phase, H , as 0.105. When these values were substituted into equation (3), q was found to be 0.0793, slightly less than qe. By substituting q, H , and the reported mean size of constricting cells, Z, of 2.0 pm, into equation (4), the mean size of dividing cells, er, was found to be 2.1 16 pm. Then, substituting q into the equations in the legend to Fig. 5, the moments of the entire population were predicted. These values are: i / t , = 1.380; qr = 0.2175; y1 = 0.5467; p2 = 2.4962. Consequently,i = 1.380 x 2.1 16 pm/2 = 1.47 pm which can be compared with '7 reported by Koppes et al. (1978) of 1.47 pm. Similarly, the value of qt calculated from distribution of constricting cells, 0.2175, can be compared with the value of qr they calculated directly from the distribution of the total population of 0.226.
Similarly, for the remaining 13 cases, we computed q from equation (3) and Zf from equation (4). From the values of q and Zr, the mean size e,of the total cell population, t, was calculated using Fig. 5. On average, the observed mean cell sizes of the 14 populations were only a little smaller (2.3%) than the values calculated from the constricting population. Another way to report the same finding is that the ratio of the observed to calculated mean sizes was 0.097 & 0.03.
A very large, unpublished, data sample was kindly given to us by Frank Trueba. It consisted of 14 populations with a total of 14000 cells. What was of special importance about this set of populations is that the extant distribution of the lengths of constricting cells and the lengths of incipient daughter cells were measured. In this case, the ratio of the observed to calculated mean size was 0.970 f 0.044 or 0.971 & 0.040, depending on whether the dividing cell distribution or newly arisen cell distribution was used in the calculation.
The coefficient of variation for the total population, qt, was also calculated from Fig. 5 for the three sets of populations. For the Woldringh et al. (1977) paper, the ratio of the observed to calculated values of qt was 1.122 0.039; for the Koppes et al. (1978) paper the same ratio was 1.057 f 0.040; and for the Trueba unpublished data, the ratio was 1.045 f 0.0505. Thus, the observed mean of the sizes in the total population is consistently a very little smaller than the calculated mean, and the actual width of the population size distribution is slightly larger than predicted from the constricting subpopulation.
These findings are close enough to support the SGC model for these strains of E. coli and the application of the methods presented here to these strains and culture conditions. The slight discrepancies for both the mean and width of the total population distribution are in a direction such that they would be further enlarged if the calculations were repeated under the assumption of a linear rate of volume increase. This argues against linear growth. However, the deviations
Downloaded from www.microbiologyresearch.org by
IP: 54.39.17.49
On: Tue, 13 Feb 2018 04:05:58
Charting the prokaryotic cell cycle 289 1
are consistent with the assumption that a few cells in the population are large and have a decreased ability to carry out macromolecular biosynthesis, or that cells increase their volume growth rate slightly above the exponential model in the last phases of the cell cycle.
The third and fourth moments of two population distributions of cell sizes of E. coli B/rA cultures in balanced growth at doubling times of 125 and 109 min have been kindly provided by Conrad Woldringh (personal communication). The values of y l , 0.74 and 0.77 from the total population are larger than the values of 0.56 and 0.55 calculated from the observed distribution of constricting cells. The values of kurtotic statistics are 2.89 and 3.15, respectively, larger than the predicted 2.80 and 2-49. Again, the observed deviations from the expected values are consistent with a few, abnormal, slowly growing cells that are predominantly larger.
DISCUSSION
The earliest studies of the cell cycle of prokaryotes were the microscopic studies of interdivision times (Rahn, 193 1-1932). His observations, which have been often confirmed, were that within a single culture under constant conditions Tis highly variable. Typically, T has a coefficient of variation of 20-25%. We first noted (Schaechter et a / . , 1962) that size at division was much better controlled and typically had half this coefficient of variation. This means that relating phases of the cycle to cell size had much greater potential for identifying critical events than relating them to age.
It is curious that 20 years later, cell size as an index of progress through the cell cycle is only now attaining ascendancy over cell age. This is largely because of advances in electron microscopy, in electron microscopic autoradiography, and in computer techniques that facilitate measurement of cellular dimensions from microphotographs. These methods, of course, provide data primarily about cell size and not cell age.
The GC model (Koch & Schaechter, 1962) had as its basic tenet that growth in terms of production of cytoplasm must ultimately control growth. It was proposed there, and further stressed later (Koch, 1977), that a search for the least variable phase of the cell cycle could identify the stage that responds to the cell’s success in synthesizing cytoplasm. This still remains a working hypothesis, since in no single case has a controlling mechanism been dissected out or defined.
An obvious candidate for a phase to be controlled by cell mass is the initiation of DNA synthesis (Cooper & Helmstetter, 1968; Donachie, 1968), but the evidence is clear in certain cases that DNA initiation has a broader variability than does cell division (Koch, 1977). In such cases, it is therefore impossible that chromosome replication has the critical role in the division cycle, although it may have a veto role. This, we believe, is a fair conclusion, even though analysis of the published data of Myxococcus xanthus (Koch, 1977) and the analysis of the data of Woldringh et al. (1977), and Koppes et al. (1978) are consistent with DNA synthesis having qa values similar to, but not smaller than, the population q. Certainly, the size at initiation of DNA synthesis does not seem to have a smaller coefficient of variation than does the size at cell division; i.e., qa is not less than q.
The method developed here provides a highly critical way to test the growth laws of different organisms under different conditions. Although the population distribution contains the required information, the distribution must be very precisely defined (Koch, 1966) to make the errors on the third and fourth moments small enough. We now point out that a comparison of the mean of the total population calculated from the total distribution and the mean calculated from an E phase provide a refined test. Although the sizes of cells vary widely, the standard error of the mean can be quite small since many cells can be measured. Although there are usually fewer E-phase cells, because the variation in size is smaller, the standard error of the calculated mean size of the total population is in most cases comparable. Thus, with adequate sampling, both can be small compared with the 4% difference corresponding to exponential versus linear growth when q = 0.10 (see legend to Fig. 3). Thus the observed mean size of each of the E. coli populations is not significantly different from the mean size calculated from the dividing population when exponential growth is assumed. However, the difference for all populations is
Downloaded from www.microbiologyresearch.org by
IP: 54.39.17.49
On: Tue, 13 Feb 2018 04:05:58
2892 A . L. KOCH A N D M. L. HIGGINS
highly significant when linear growth is assumed. Consequently, we conclude that E. coli grows exponentially and that our new approach can exceed the power of tests based on total population distributions alone.
The purpose of this paper has been to propose a method to aid in the definition of phases of the cell cycle; it should aid in the search for the most precisely regulated phase as well as establishing size and temporal relation in the cycle. Of course, the method should not be applied when the standard growth controlled model (SGC) does not predict the population dynamics adequately, nor if the phase under study overlaps generations. The criteria proposed must be satisfied if one is to use the computational methods, but for those cases where it can be applied, we trust that these graphs and formulae will serve as a Rosetta stone to translate pictures of fixed cells into an understanding of the dynamic processes of the cell cycle.
We thank the consulting staff of the Wrubel Computing Center for aid during the development of the programs presented here. The work was supported by the Indiana University Computing Fund. We thank Dr Lolita Daneo- Moore for suggesting this problem, and Drs Conrad Woldringh and Frank Trueba for providing us with their unpublished data. Experimental work in our laboratories is supported by NSF PCM 79 11241 (A.L.’K.) and NIH A1 10971 (M.L.H.).
R E F E R E N C E S COLLINS, J . F. & RICHMOND, M. H. (1962). Rate of
growth of Bacillus cereus between divisions. Journal of General Microbiology 28, 15-33.
COOPER, S. & HELMSTETTER, C. E. (1968). Chromo- some replication and the division cycle of Escherichia coli Bjr. Journal of Molecular Biology 31, 5 19-540.
CULLUM, J . & VINCENTE, M. (1978). Cell growth and length distribution in Escherichia coli. Journal of Bacteriology 134, 330-337.
DONACHIE, W. D. (1968). Relationship between cell size and the time of initiation of DNA replication. Nature, London 219, 1077-1079.
HARVEY, R. J . , MARR, A. G. & PAINTER, P. R. (1967). Kinetics of growth of individual cells of Escherichia coli and Azotobacter agilis. Journal of Bacteriology 93,
KOCH, A. L. (1966). Distribution of cell size in growing cultures of bacteria and the applicability of the Collins-Richmond principle. Journal of General Microbiology 45, 409-41 7.
KOCH, A. L. (1977). Does the initiation of chromosome replication regulate cell division? Adcances in Microbial Physiology 16, 49-98.
KOCH, A. L. (1980). Does the variability of the cell cycle result from one or many chance events? Nature, London 286, 80-82,
KOCH, A. L. & BLUMBERG, G. (1976). Distribution of bacteria in the velocity gradient centrifuge. Biophysical Journal 16, 389-405.
KOCH, A. L. & SCHAECHTER, M . (1 962). A model for the statistics of the cell division process. Journal uf’ General Microbiology 29, 435-454.
KOPPES, L. J. H., WOLDRINGH, C. L. & NANNINGA, N. (1978). Size variation and correlation of different cell cycle events in slow-growing Escherichia coli. Journal of’ Bacteriology 134, 423433.
605-6 1 7.
KUBITSCHEK, H. E. (1970). Evidence for the generality of linear cell growth. Journal of Theoretical Biology
KUBITSCHEK, H. E. (1981). Bilinear cell growth of Escherichia coli. Journal of Bacteriology 148,730-733.
PAINTER, P. R. & MARR, A. G . (1968). Mathematics of microbial populations. Annual Review of Micro- biology 22, 5 19-548.
POWELL, E. 0. (1964). A note on Koch & Schaechter’s hypothesis about growth and fission of bacteria. Journal of General Microbiology 37, 23 1-249.
RAHN, 0. (1931-1932). A chemical explanation of the variability of growth rate. Journal of General Phj.sio1og.v 15, 257-277.
SCHAECHTER, M., WILLIAMSON, J . P., HOOD, J . R. & KOCH, A. L. (1 962). Growth, cell and nuclear divi- sion in some bacteria. Journal of General Micro- biology 29, 42 1-454.
SOKAL, R. R. & ROHLF, F. J . (1969). Biometry. San Francisco : W . H. Freeman.
STUBBLEFIELD, E. (1981). Cell cycle. In Advanced Cell Biologji, pp, 1005-1025. Edited by L. M. Schwartz & M. M. Azar. New York: Van Nostrand Reinhold.
TRUEBA, F. J . (1982). On the precision and accuracy achieved by Eschrrichia coli cells at fission about their middle. Archit!es qf’ Microbiology 131. 55-59.
WOLDRINGH, c. L., DEJONG, N . A., VAN DEN BERG, W. & KOPPES. L. J. H. (1977). Morphological analysis of the division cycle of two Escherichia coli substrains during slow growth. Journal of Bac- teriology 131, 270-279.
ZUSMAN, D., GOTTLIEB, P. & ROSENBERG, E. (1971). Division cycle of My.~ococcus xanthus. 111. Kinetics of cell growth and protein synthesis. Journal of Bac- teriology 105, 81 1-819.