Ž . Brain Research 843 1999 95–104 www.elsevier.comrlocaterbres Research report Ž . Cellular localization of dopamine-releasing protein DARP in rat C6 glioma and primary mesencephalic cell cultures Sean Smith a, ) , Victor D. Ramirez a,b a Department of Neuroscience, UniÕersity of Illinois at Urbana-Champaign, 524 Burrill Hall, MIP, 407s Goodwin AÕenue, Urbana, IL 61801, USA b Department of Molecular and IntegratiÕe Physiology, UniÕersity of Illinois at Urbana-Champaign, Urbana, IL 61801, USA Accepted 20 July 1999 Abstract Ž . Dopamine-releasing protein DARP is a multisubunit protein shown to have dramatic effects on development, recovery, and function Ž . of the rat catecholaminergic CA system. This study details efforts to determine if glial cells are responsible for the production of DARP Ž . Ž . in the central nervous system CNS . Enzyme-linked immunosorbent assays ELISA , Western blotting, and immunocytochemical techniques were employed to measure DARP levels and identify DARP immunoreactive proteins in rat C6 glioma cells and medium, respectively. ELISA analysis of serum-free C6 culture media revealed a maximal concentration of DARP by culture day 1. However, ELISA analysis of C6 cultures grown in F-12Krserum medium revealed that maximal levels of DARP were detected on culture day 6 with a 108% increase in DARP immunoreactivity from culture day 1. These values were determined using a polyclonal antibody Ž . Ž . generated against DARP-36aa anti-DARP-36aa , a synthetic peptide with dopamine DA releasing activity, and anti-DARP B9–B10, a monoclonal antibody generated against partially purified DARP. Western blot analysis revealed that anti-DARP B9–B10 recognized proteins of approximately 60, 50, and 45 kDa in C6 cell homogenates while anti-DARP-36aa had immunoreactivity with the 60-kDa protein alone. Immunocytochemical studies demonstrated that anti-DARP-36aa and anti-DARP B9–B10 had strong immunoreactivity with proteins throughout the cytosol and in several processes of C6 cells. These results reveal that DARP is detected in glioma cells and secreted in a time-dependent fashion during culture. Primary rat mesencephalic cultures were also examined using immunocytochemistry. Ž . Incubation with DARP antibodies and antisera against glial fibrillary acidic protein GFAP revealed that DARP and GFAP immunoreactivity co-localized in primary mesencephalic cultures. However, the majority of DARP immunoreactivity was localized to cells without GFAP staining. These findings reveal that DARP is detected in astrocytes although the majority of DARP immunoreactivity is found in non-astrocyte type cells. q 1999 Elsevier Science B.V. All rights reserved. Ž . Keywords: Dopamine-releasing protein DARP ; Rat C6 glioma cell; ELISA; Immunocytochemistry; Neurotrophic factor 1. Introduction Neurotrophic factors have classically been defined as signaling molecules that originate from innervated tissues and govern neuronal survival as well as enhance neuronal w x differentiation and growth 2,7,15,22 . However, it has been reported that neurons not only acquire trophic support from their innervated targets, but also receive trophic w x support from afferent neurons 8,10 , autocrine mecha- w x nisms 1,12 , and glial cells. Specifically, the expression of Ž . brain-derived neurotrophic factor BDNF , neurotrophin-3 Ž . Ž . NT-3 , neurotrophin-4r5 NT-4r5 , and glial cell-line Ž . derived neurotrophic factor GDNF mRNA has been ob- ) Corresponding author. Fax: q1-217-333-1133; E-mail: [email protected]served in glial cultures and localized in glial cells of the rat w x brain 17,19,24,26,27,29–32 . In addition, immunocyto- chemical studies have shown co-localization of NT-3 and Ž . NT-4r5 with glial fibrillary acidic protein GFAP posi- wx tive cells in the rat brain 9 . These studies provide evi- dence for functional interactions between neurotrophic fac- tors and glial cells. wx In 1988, Chang and Ramirez 4 demonstrated that a Ž partially purified protein dopamine-releasing protein Ž .. DARP from the rat adrenal gland released dopamine Ž . DA from in vitro striatal tissue in a dose-dependent manner. Immunopurification of this protein from primary mesencephalic cell culture revealed that DARP is a multi- subunit protein of 200 kDa. Microsequencing the N- terminus of purified DARP enabled this laboratory to Ž . synthesize a 36 amino acid peptide fragment DARP-36aa . 0006-8993r99r$ - see front matter q 1999 Elsevier Science B.V. All rights reserved. Ž . PII: S0006-8993 99 01895-8
Transcript
Ž .Brain Research 843 1999 95–104www.elsevier.comrlocaterbres
Research report
Ž .Cellular localization of dopamine-releasing protein DARP in rat C6 gliomaand primary mesencephalic cell cultures
Sean Smith a,), Victor D. Ramirez a,b
a Department of Neuroscience, UniÕersity of Illinois at Urbana-Champaign, 524 Burrill Hall, MIP, 407s Goodwin AÕenue, Urbana, IL 61801, USAb Department of Molecular and IntegratiÕe Physiology, UniÕersity of Illinois at Urbana-Champaign, Urbana, IL 61801, USA
Accepted 20 July 1999
Abstract
Ž .Dopamine-releasing protein DARP is a multisubunit protein shown to have dramatic effects on development, recovery, and functionŽ .of the rat catecholaminergic CA system. This study details efforts to determine if glial cells are responsible for the production of DARP
Ž . Ž .in the central nervous system CNS . Enzyme-linked immunosorbent assays ELISA , Western blotting, and immunocytochemicaltechniques were employed to measure DARP levels and identify DARP immunoreactive proteins in rat C6 glioma cells and medium,respectively. ELISA analysis of serum-free C6 culture media revealed a maximal concentration of DARP by culture day 1. However,ELISA analysis of C6 cultures grown in F-12Krserum medium revealed that maximal levels of DARP were detected on culture day 6with a 108% increase in DARP immunoreactivity from culture day 1. These values were determined using a polyclonal antibody
Ž . Ž .generated against DARP-36aa anti-DARP-36aa , a synthetic peptide with dopamine DA releasing activity, and anti-DARP B9–B10, amonoclonal antibody generated against partially purified DARP. Western blot analysis revealed that anti-DARP B9–B10 recognizedproteins of approximately 60, 50, and 45 kDa in C6 cell homogenates while anti-DARP-36aa had immunoreactivity with the 60-kDaprotein alone. Immunocytochemical studies demonstrated that anti-DARP-36aa and anti-DARP B9–B10 had strong immunoreactivitywith proteins throughout the cytosol and in several processes of C6 cells. These results reveal that DARP is detected in glioma cells andsecreted in a time-dependent fashion during culture. Primary rat mesencephalic cultures were also examined using immunocytochemistry.
Ž .Incubation with DARP antibodies and antisera against glial fibrillary acidic protein GFAP revealed that DARP and GFAPimmunoreactivity co-localized in primary mesencephalic cultures. However, the majority of DARP immunoreactivity was localized tocells without GFAP staining. These findings reveal that DARP is detected in astrocytes although the majority of DARP immunoreactivityis found in non-astrocyte type cells. q 1999 Elsevier Science B.V. All rights reserved.
Ž .Keywords: Dopamine-releasing protein DARP ; Rat C6 glioma cell; ELISA; Immunocytochemistry; Neurotrophic factor
1. Introduction
Neurotrophic factors have classically been defined assignaling molecules that originate from innervated tissuesand govern neuronal survival as well as enhance neuronal
w xdifferentiation and growth 2,7,15,22 . However, it hasbeen reported that neurons not only acquire trophic supportfrom their innervated targets, but also receive trophic
w xsupport from afferent neurons 8,10 , autocrine mecha-w xnisms 1,12 , and glial cells. Specifically, the expression of
served in glial cultures and localized in glial cells of the ratw xbrain 17,19,24,26,27,29–32 . In addition, immunocyto-
chemical studies have shown co-localization of NT-3 andŽ .NT-4r5 with glial fibrillary acidic protein GFAP posi-
w xtive cells in the rat brain 9 . These studies provide evi-dence for functional interactions between neurotrophic fac-tors and glial cells.
w xIn 1988, Chang and Ramirez 4 demonstrated that aŽpartially purified protein dopamine-releasing protein
Ž ..DARP from the rat adrenal gland released dopamineŽ .DA from in vitro striatal tissue in a dose-dependentmanner. Immunopurification of this protein from primarymesencephalic cell culture revealed that DARP is a multi-subunit protein of 200 kDa. Microsequencing the N-terminus of purified DARP enabled this laboratory to
Ž .synthesize a 36 amino acid peptide fragment DARP-36aa .
0006-8993r99r$ - see front matter q 1999 Elsevier Science B.V. All rights reserved.Ž .PII: S0006-8993 99 01895-8
( )S. Smith, V.D. RamirezrBrain Research 843 1999 95–10496
Interestingly, this partial sequence information indicatesthat DARP has no sequence homology with known neu-
w xrotrophic factors 23 . It has been reported that DARP-36aaincreased striatal DA content and release while eliminatingamphetamine-induced rotational behavior resulting from
Ž .1-methyl-4-phenylpyridinium MPPq induced unilateralw xlesions of the nigrostriatal dopaminergic system 21 .
This laboratory has demonstrated that DARP has aŽ .dramatic effect on catecholaminergic CA systems during
w xdevelopment. Kuhananthan et al. 13 revealed that theadministration of monoclonal antibodies raised against par-tially purified DARP decreased DA levels in the striatum
Ž .while increasing norepinephrine NE and decreasingŽ .epinephrine Epi levels in the hypothalamus of neonatal
rats. Also, immunoneutralization of DARP in the rat cen-Ž . Ž .tral nervous system CNS on embryonic day 17 E 17
w xselectively alters DA levels in the mesencephalon 16 .Immunocytochemical studies employing anti-DARP
B9–B10 demonstrated that DARP maps in the rat CNScoincide in density as well as anatomical distribution with
Ž .well-known tyrosine hydroxylase TH maps for CA neu-w xrons 6 . Finally, it was reported that patients suffering
from idiopathic Parkinson’s disease had higher values ofDARP-like activity in their serum as compared to age-
w xmatched controls 14 . These findings led to the postulationof the hypothesis that DARP is produced in glia cellsunder a negative control of DA, so that when levels of DAdecrease in the extracellular fluid, a higher secretion ofDARP is triggered in order to compensate for the lack ofDA. For this communication, we asked the question ifDARP is produced in and released from glial cells as afirst study to prove or deny the aforementioned hypothesis.
In this study, enzyme-linked immunosorbent assaysŽ .ELISA , Western blotting, and immunocytochemical tech-niques were employed to measure DARP levels and iden-tify proteins, respectively, in rat C6 glioma cells andprimary mesencephalic cell cultures. Our results demon-strate that C6 cell proteins and media have strong immuno-reactivity with both monoclonal antibodies generated
Ž .against partially purified DARP entire protein and poly-clonal antibodies generated against DARP-36aa. Whileimmunocytochemical analysis of primary mesencephaliccultures also demonstrated that DARP was detected inastrocytes although the majority of DARP staining waslocalized to cells without GFAP immunoreactivity.
2. Materials and methods
2.1. C6 glioma cell cultures
w x 2C6 glioma cells 3 were cultured in 75 cm tissueculture flasks or on coverslips in 24-well culture platesŽ .Corning . These cells were plated in Ham’s F-12K medium
Ž .supplemented with L-glutamine 2 mM , sodium bicarbon-Ž . Ž .ate 1.5 grl , 15% horse serum HS and 2.5% fetal
Ž .bovine serum FBS . Cultures were allowed to adhere for24 h in F-12Krserum medium. Following this period,selected cultures were washed 3= with serum-free Opti-
Ž .MEM medium Gibco supplemented with 0.5% bovineŽ . Ž .serum albumin BSA and CaCl 300 mgrl . Following2
this wash C6 cells were cultured in 15 ml of serum-freeOptiMEM medium. Cultures were kept at 378C in a hu-midified atmosphere containing 94% air and 6% CO .2
A 0.5-ml sample of medium was removed from eachculture for ELISA analysis every 24 h. In addition, culturedensity was quantified every 24 h using a Nikon phasecontrast microscope and a calibrated reticle with a fieldsize of 0.1156 mm2. Fresh OptiMEM medium was supple-mented to replace the aliquot taken for ELISA. Thisprocedure was implemented 24 h after the introduction of
Ž .serum-free medium day 1 and terminated 96 h afterwardsŽ .day 4 .
2.2. Primary mesencephalic cell cultures
Primary mesencephalic cell cultures were prepared fromŽ . Ž .postnatal day 3 P3 rat pups Sprague–Dawley . The
mesencephalon of P3 rat pups was dissected and pooledtogether in a sterile petri dish containing Mg2qrCa2q-freeTyrodes Buffer. Following dissection, mesencephalic tis-sue was diced into 1–3 mm cubes and washed 2= with 20volumes of Mg2qrCa2q-free Tyrodes Buffer. Cells werethen pelleted by centrifugation at 325=g for 5 min. Theresulting pellet was then incubated in 2 ml of tryp-sinrEDTA at 378C for 15 min. Enzymatic digestion wasstopped with the addition of 2= volume of DMEM con-taining 10% FBS. Tissue was then mechanically triturated
Žusing a sterile Pasteur pipette attached to a Pi-Pump 20.passes . Cell suspensions in DMEM containing 10% FBS
and 1% Penstrep were seeded onto coverslips in 24-wellŽ .cell culture plates Corning . Cultures were maintained at
378C in a humidified atmosphere containing 94% air and6% CO with two media changes per week.2
2.3. Western blotting
C6 glioma cells, harvested on culture day 4, werewashed 3= with 10 volumes of 0.1 M phosphate-buffered
Ž .saline PBS containing 0.1 mM bacitracin and 0.5 mMŽ4-2-aminoethyl benzenesulfonylfluoride-HCl AEBSF, a
.protease inhibitor; Boehringer-Mannheim and pelleted byŽ .centrifugation 5 min at 325=g . Cells were then homog-
Ž .enized in 10 ml of ice-cold PBS buffer listed above bysonication. Cell homogenate was centrifuged for 30 min at
Ž .8000=g 48C to remove large cellular debris. Then cellŽ .homogenate samples were heated 958C in the presence of
5% b-mercaptoethanol. Unpurified C6 glioma cell super-natant was then loaded on a 4%–20% gradient sodium
Ždodecyl sulfate-polyacrylamide gel electrophoresis SDS-. Ž .PAGE minigel Biorad . The separated proteins were then
electrophoretically transferred to a nitrocellulose mem-
( )S. Smith, V.D. RamirezrBrain Research 843 1999 95–104 97
brane. The membrane was incubated in 2% BSA in 0.1 MŽ .PBS containing either anti-DARP B9–B10 1:50 or anti-
Ž . ŽDARP-36aa 1:100 . A secondary antibody anti-mouse.IgM or anti-rat IgG, Sigma conjugated to horseradish
Ž . X Ž .peroxidase HRP and a 3,3 -diamiodenzidine DAB sub-Ž .strate system with NiCl 0.05% enhancement was used2
to visualize immunoreactive proteins. Non-specific bindingof the membrane was blocked with 2% BSA in PBS. Allwashings between steps were performed with 0.1 M PBSŽ .pH 7.4 . Pre-adsorption of anti-DARP-36aa was accom-plished by incubating 8 ug of DARP-36aa per 8 ug ofanti-DARP-36aa. This solution was incubated overnight at48C followed by a centrifugation at 14,000=g for 20 min.The resulting supernatant was used as a primary antibodyin control experiments.
2.4. Immunocytochemistry
C6 cells grown on coverslips for 2 days were firstwashed with 0.1 M PBS then fixed for 20 min in 4%paraformaldehyde in 0.1 M PBS. These cells were blockedand permeabilized by a 1-h incubation in 0.1 M PBS with
Ž .0.2% Triton X-100 and 2% goat serum GS . This wasŽ .followed by an incubation with anti-DARP B9–B10 1:25
Ž .or anti-DARP-36aa 1:100 diluted in 2% GS in PBS for16 h at 48C. Anti-rat IgG or anti-mouse IgM conjugated
Ž . Ž .either to HRP or indocarbocyanine Cy3 Jackson, 1:1000in 2% GS in PBS was applied for 2 h at room temperatureŽ .RT . Cell labeling was visualized through an incubation
Žwith a DAB solution DAB 16 mgr30 ml, 0.05% nickel.chloride, 30 ul H O in 0.5 M Tris for 5 min at RT.2 2
Staining specificity was determined by substitution of thefollowing for the primary antibody: 2% GS in 0.1 M PBSor anti-DARP-36aa pre-adsorbed with DARP-36aa follow-ing the same procedure detailed in Section 2.3.
Primary mesencephalon cultures cells were grown oncoverslips for 13 days to ensure that cultures were conflu-ent. Mesencephalic cells were first washed with 0.1 MPBS then fixed for 20 min in 4% paraformaldehyde in 0.1M PBS. These cells were blocked and permeablized by a1-h incubation in 0.1 M PBS with 0.2% Triton X-100 and2% GS. This was followed by an incubation with anti-
Ž . Ž .DARP B9–B10 1:50 or anti-DARP-36aa 1:100 andŽ .antisera against GFAP Cemicon, 1:1000 diluted in 2%
GS in PBS for 16 h at 48C. Anti-rat IgG or anti-mouseIgM conjugated either to HRP or Cy3 and anti-rabbit IgG
Ž . Ž .conjugated to fluorescein FITC Jackson, 1:500 in 2%GS in PBS was applied for 2 h at RT. Immunofluorescencecell labeling was visualized using a Nikon microscopeequipped with fluorescence optics. Staining specificity wasdetermined by substitution of 2% GS in 0.1 M PBS for theprimary antibody.
( )2.5. Enzyme-linked immunosorbent assay ELISA
C6 cell culture media was used as an antigen in theseŽ .assays. Cell media samples were diluted 1:1000 with 0.1
M PBS to a final protein concentration of 0.8 ug per well.A total of 50 ul of diluted sample was incubated in Nunc96-well ELISA plates for 2 h at RT. Non-specific bindingwas blocked through an incubation with 2% BSA in PBS
Žfor 16 h at 48C. Primary antibodies anti-DARP B9–B10.1:50 or anti-DARP-36aa 1:500 were incubated for 2 h at
RT. HRP conjugated anti-mouse IgM or anti-rat IgGŽ .1:1000 in 2% BSA in PBS was incubated for 2 h at RT.
X X Ž .A Turbo 3,3 , 5,5 -tetramethyl benzidine TMB microwellŽ .substrate system KPL was employed as a chromogen.
Plates were read on a Dynatech plate reader at a wave-length of 450 nm. All washings between steps were per-
Ž .formed with 0.1 M PBS see Fig. 1 .
2.6. Antibody production and purification
As part of an earlier study, anti-DARP-36aa was gener-ated in female Sprague–Dawley rats. Female rats were
Ž .injected with 100 ug of DARP-36aa in a 1:1 vrv emulsi-Ž .fication with complete Freund adjuvant Sigma . A second
immunization of 50 ug DARP-36aa and incomplete Freundadjuvant was administered 17 days after the initial injec-tion. Finally, an intraperitoneal injection of DARP-36aaŽ .50 ug was given 17 days after the second immunization.
Blood samples were collected via jugular venous punc-ture and sera were screened for anti-DARP-36aa titer usingan ELISA with DARP-36aa used as an antigen. Sera withstrong immunoreactivity with DARP-36aa was treated with5% sodium dextran sulfater11.1% calcium chloride toremove lipoproteins. Following lipoprotein removal, im-munoglobulin was precipitated with 50% ammonium sul-fate and resuspended in 0.1 M PBS. The preparation of
w xanti-DARP B9–B10 has been reported previously 13 .
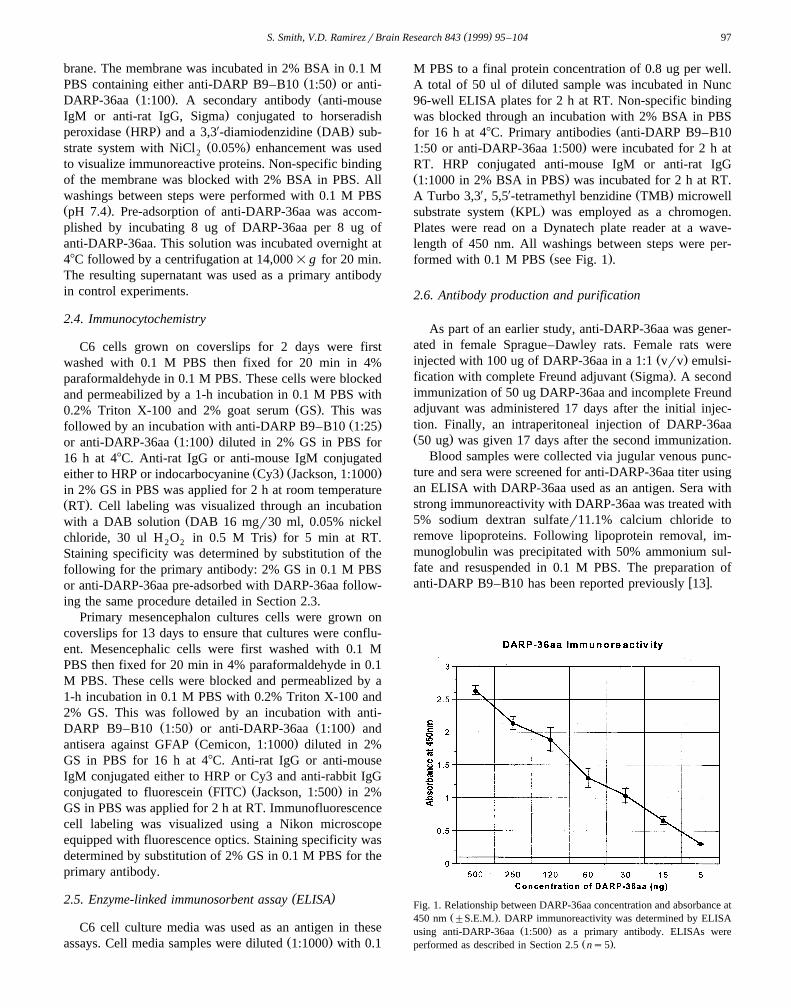
Fig. 1. Relationship between DARP-36aa concentration and absorbance atŽ .450 nm "S.E.M. . DARP immunoreactivity was determined by ELISA
Ž .using anti-DARP-36aa 1:500 as a primary antibody. ELISAs wereŽ .performed as described in Section 2.5 ns5 .
( )S. Smith, V.D. RamirezrBrain Research 843 1999 95–10498
2.7. Statistical analysis
All data values were expressed as means"S.E.M. Aone-way ANOVA and an unpaired t-test were employed toanalyze data generated in this study. In all cases, a p value-0.05 was considered significant.
3. Results
3.1. Detection of DARP in C6 cell culture medium
To determine if C6 glioma cells release DARP, C6 cellculture medium was collected and sampled by ELISA. A0.5-ml sample of culture medium was collected for ELISAanalysis every 24 h for a total of 4 days. Utilizing anti-DARP-36aa, we detected approximately 95 ng of DARP in0.5 ml of serum-free medium by culture day 1. Intrigu-ingly, maximal levels were detected 24 h after introduction
Ž .of serum-free OptiMEM medium day 1 . This concentra-tion of DARP remained constant through culture day 4Ž .Fig. 2 . Replacement of anti-DARP-36aa with 2% BSA inPBS eliminated observed immunoreactivity in culturemedium.
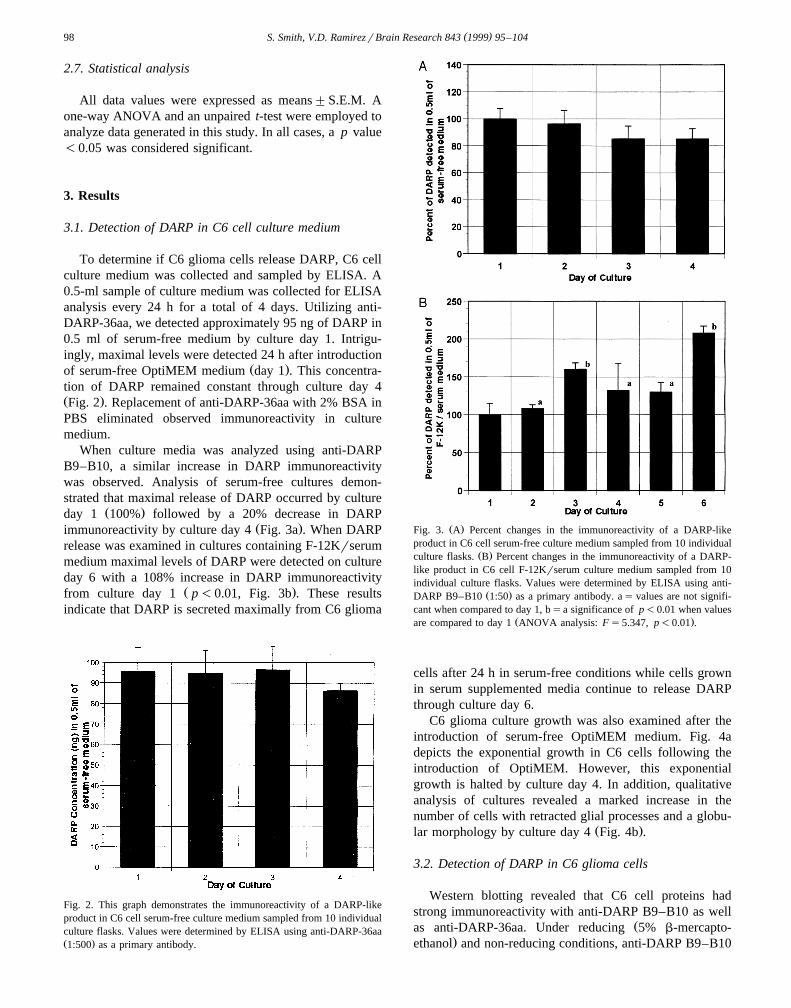
When culture media was analyzed using anti-DARPB9–B10, a similar increase in DARP immunoreactivitywas observed. Analysis of serum-free cultures demon-strated that maximal release of DARP occurred by culture
Ž .day 1 100% followed by a 20% decrease in DARPŽ .immunoreactivity by culture day 4 Fig. 3a . When DARP
release was examined in cultures containing F-12Krserummedium maximal levels of DARP were detected on cultureday 6 with a 108% increase in DARP immunoreactivity
Ž .from culture day 1 p-0.01, Fig. 3b . These resultsindicate that DARP is secreted maximally from C6 glioma
Fig. 2. This graph demonstrates the immunoreactivity of a DARP-likeproduct in C6 cell serum-free culture medium sampled from 10 individualculture flasks. Values were determined by ELISA using anti-DARP-36aaŽ .1:500 as a primary antibody.
Ž .Fig. 3. A Percent changes in the immunoreactivity of a DARP-likeproduct in C6 cell serum-free culture medium sampled from 10 individual
Ž .culture flasks. B Percent changes in the immunoreactivity of a DARP-like product in C6 cell F-12Krserum culture medium sampled from 10individual culture flasks. Values were determined by ELISA using anti-
Ž .DARP B9–B10 1:50 as a primary antibody. as values are not signifi-cant when compared to day 1, bsa significance of p-0.01 when values
Ž .are compared to day 1 ANOVA analysis: Fs5.347, p-0.01 .
cells after 24 h in serum-free conditions while cells grownin serum supplemented media continue to release DARPthrough culture day 6.
C6 glioma culture growth was also examined after theintroduction of serum-free OptiMEM medium. Fig. 4adepicts the exponential growth in C6 cells following theintroduction of OptiMEM. However, this exponentialgrowth is halted by culture day 4. In addition, qualitativeanalysis of cultures revealed a marked increase in thenumber of cells with retracted glial processes and a globu-
Ž .lar morphology by culture day 4 Fig. 4b .
3.2. Detection of DARP in C6 glioma cells
Western blotting revealed that C6 cell proteins hadstrong immunoreactivity with anti-DARP B9–B10 as well
Žas anti-DARP-36aa. Under reducing 5% b-mercapto-.ethanol and non-reducing conditions, anti-DARP B9–B10
( )S. Smith, V.D. RamirezrBrain Research 843 1999 95–104 99
Ž . Ž .Fig. 4. A Summary of C6 glioma culture growth "S.E.M. in serum-free OptiMEM medium sampled from 10 individual culture flasks. Day
Ž 6 . 2s0 corresponds to the number of cells seeded 1.0=10 per 75 cmŽ .culture flask. B Phase contrast photomicrograph of culture density on
day 4. Scale bar: 30 mm.
recognized proteins of approximately 60, 50, and 45 kDain C6 cell homogenates. Anti-DARP B9–B10 had strongimmunoreactivity with C6 cell homogenates cultured ineither F-12Krserum medium or serum-free OptiMEM
Ž .medium Fig. 5, lanes 2 and 3, respectively . In controlexperiments, immunoreactivity was eliminated when anti-
ŽDARP B9-B10 was replaced with 2% BSA data not.shown . A positive control is shown in Fig. 5, lane 5,
corresponding to partially purified DARP obtained fromprimary mesencephalic cell culture medium as reported
w xpreviously 13,23 . Also, note that lane 4 in Fig. 5 contain-ing the synthetic peptide DARP-36aa is not recognized bythe anti-DARP B9–B10 monoclonal antibody. In addition,the original production of anti-DARP B9–B10 screenedfor individual clones that blocked DARP-evoked DA re-
w xlease in an in vitro superfusion assay 13,23 . Also, thislaboratory has demonstrated that anti-DARP B9–B10 im-
Fig. 5. This figure demonstrates that C6 cell proteins grown in eitherF-12Krserum medium or serum-free OptiMEM medium has strong
Ž .immunoreactivity with anti-DARP B9–B10 1:50 . Lane 1: 10 ul ofŽ .standard Biorad broad range . Lane 2: 35 ug of C6 cell homogenate
cultured in F-12Krserum media. Lane 3: 35 ug of C6 cell homogenatecultured in serum-free OptiMEM media. Lane 4: 100 ng of DARP-36aa.Lane 5: 10 ug of partially purified DARP isolated from primary rat
w xmesencephalic cell cultures 23 .
munoreactivity is eliminated upon preabsorption with par-w xtially purified DARP 6 . Based on these results, we are
confident that the immunoreactivity observed in these ex-periments was due to specific interactions between anti-DARP B9–B10 and endogenous DARP.
Interestingly, anti-DARP-36aa was found to have strongimmunoreactivity only with the 60-kDa protein in C6 cell
Žhomogenates grown in F-12Krserum medium Fig. 6, lane.1 . However, anti-DARP-36aa had greatly reduced im-
munoreactivity with the 60-kDa subunit in C6 cell ho-Ž .mogenates grown in serum-free medium Fig. 6, lane 2 .
As expected, anti-DARP-36aa had an intense reaction withŽ .the synthetic peptide DARP-36aa Fig. 6, lane 3 . Anti-
Fig. 6. This figure demonstrates that C6 cell proteins grown in F-12Krserum medium have strong immunoreactivity with anti-DARP-36aaŽ .1:100 . Lane 1: 35 ug of C6 cell homogenate cultured in F-12Krserummedia. Lane 2: 35 ug of C6 cell homogenate cultured in serum-freeOptiMEM media. Lane 3: 100 ng of DARP-36aa. Lane 4: 35 ug of C6cell homogenate cultured in F-12Krserum media. Lane 5: 35 ug of C6cell homogenate cultured in serum-free OptiMEM media. Lane 6: 100 ng
Ž .of DARP-36aa. Lanes 1–3 were incubated with anti-DARP-36aa 1:100while lanes 4–6 represent control blots incubated with anti-DARP-36aapre-adsorbed with DARP-36aa.
( )S. Smith, V.D. RamirezrBrain Research 843 1999 95–104100
( )S. Smith, V.D. RamirezrBrain Research 843 1999 95–104 101
DARP-36aa immunoreactivity with the 60-kDa protein andDARP-36aa was completely eliminated in control blotsincubated with anti-DARP-36aa pre-adsorbed with DARP-
Ž .36aa Fig. 6, lanes 4–6 . However, control experimentsrevealed a non-specific immunoreactive protein of 15 kDaŽ .Fig. 6, lanes 2 and 5 . This protein is only present inserum-free cultures and appears to be a product of serum-free culture conditions. Also, this immunoreactive proteinappears to have an affinity for a non-specific rat IgG in theprimary antibody solution. Differences in DARP immuno-reactivity observed between anti-DARP B9–B10 and anti-DARP-36aa can be attributed to the generation of theseantibodies. Anti-DARP-36aa was generated against a syn-thetic peptide corresponding to the first 36 amino acids ofthe 60-kDa subunit of DARP, where as anti-DARP B9-B10
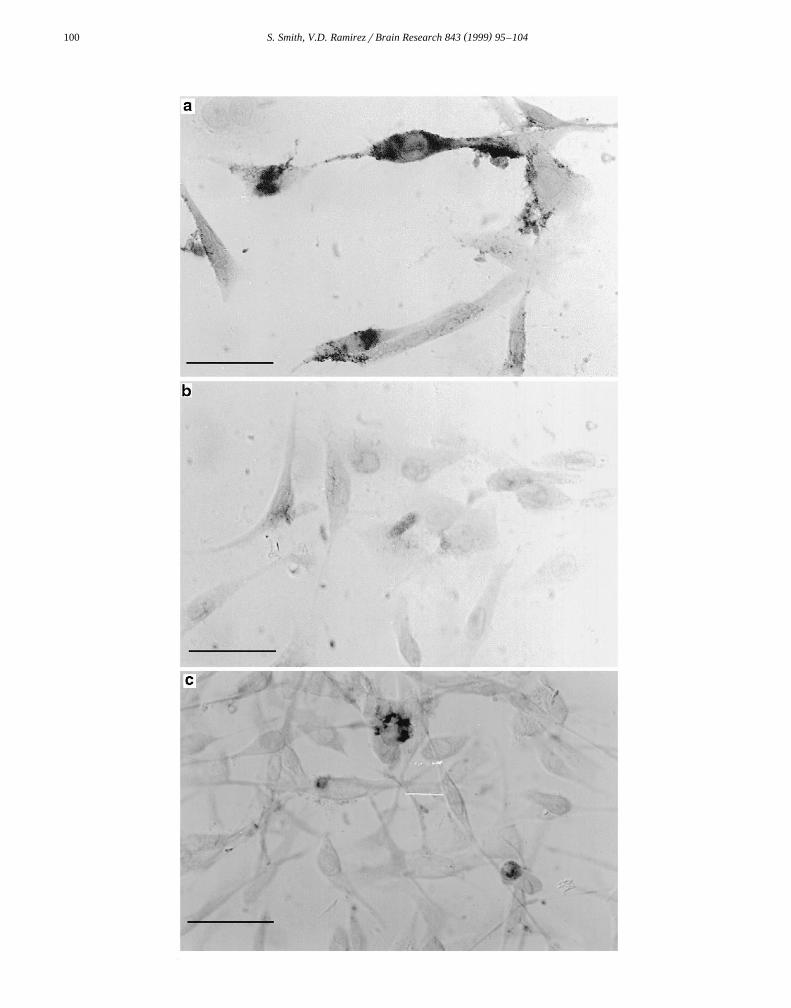
w xwas generated against partially purified DARP 13 .Immunocytochemical localization of DARP in C6
glioma cells is depicted in Fig. 7. Fig. 7a demonstrates thatŽ .anti-DARP-36aa 1:100 had strong immunoreactivity with
proteins throughout the cytosol and in several processes ofthese cells. Specifically, anti-DARP-36aa appears to recog-nize diffuse protein throughout the entire cytosol of thecell. Interestingly, there is an increase in staining intensityaround the nucleus of several cells. Staining intensityclearly decreased but still remained well-defined when theprimary antibody was diluted by a factor of 1:500 or
Ž .1:1000 data not shown . The specificity of staining wasdemonstrated using a pre-adsorbed primary antibody.Anti-DARP-36aa staining was eliminated when cultureswere incubated with anti-DARP-36aa pre-adsorbed with
Ž . Ž .DARP-36aa Fig. 7b . Anti-DARP B9–B10 1:25 alsoŽrecognizes specific proteins within C6 glioma cells Fig.
.7c . Several areas of intense immunoreactivity can beobserved in cultures incubated with anti-DARP B9–B10.These areas of intense staining have a perinuclear localiza-tion in C6 cells. Diffuse staining was also observed allthrough the cytosol. Control experiments revealed that cellstaining is eliminated when primary antibodies are re-placed with 2% GS. Thus, both anti-DARP-36aa andanti-DARP B9–B10 specifically stained proteins withinC6 glioma cells.
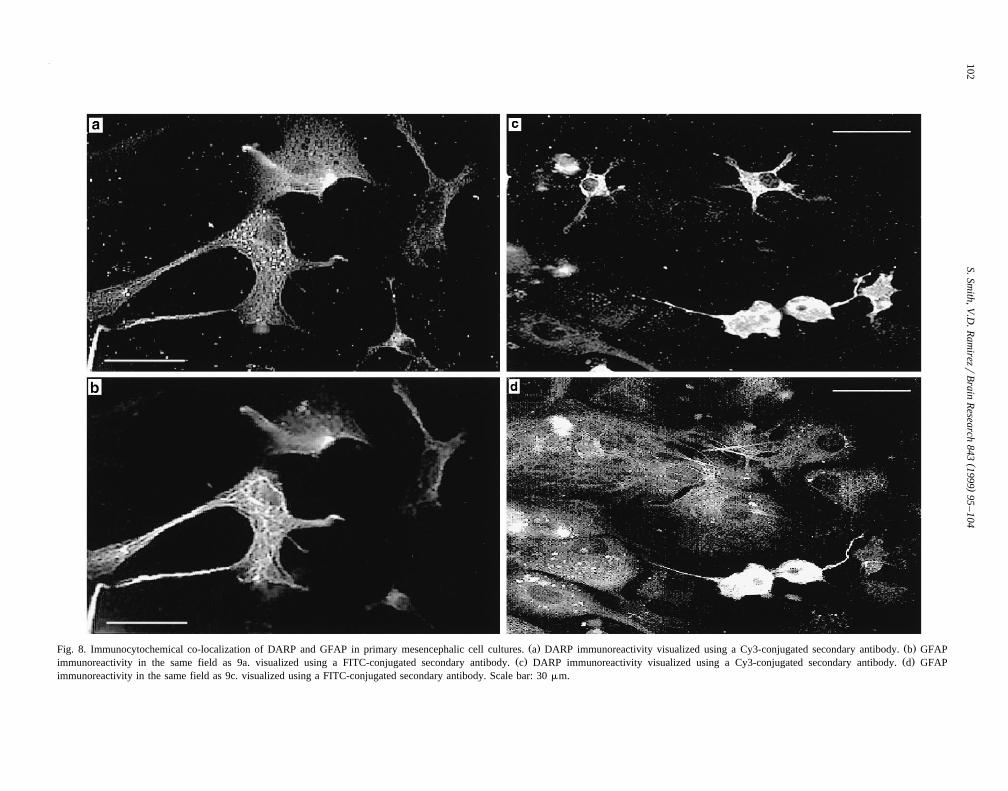
Immunocytochemical labeling for DARP was also de-tected in primary rat mesencephalic cultures using ananti-mouse IgM or anti-rat IgG antibody conjugated to
Ž .HRP and a DAB solution enhanced with NiCl 0.05% or2
anti-mouse IgM conjugated to Cy3. Incubation with eitheranti-DARP-36aa or anti-DARP B9–B10 revealed exten-sive DARP-immunoreactivity with mesencephalic cellsŽ .Fig. 8a and c . In addition, double labeling studies usingDARP antibodies and antisera against GFAP revealed thatDARP and GFAP immunoreactivity co-localized in pri-
Ž .mary mesencephalic cell cultures Fig. 8a–b and c–d .However, the majority of DARP immunoreactivity waslocalized to cells without GFAP staining. Specifically,immunofluorescence with anti-DARP B9–B10 revealedintense staining localized near the nucleus of cells. Incontrol experiments, DARP-immunoreactivity is elimi-nated when primary antibodies were replaced with 2% GSŽ .data not shown . These findings reveal that DARP isdetected in astrocytes although the majority of DARPimmunoreactivity is found in non-astrocyte type cells inprimary mesencephalic cultures.
4. Discussion
In this study we were interested in determining ifDARP is produced in and released from glial cells. Ourresults demonstrate that DARP is detected in and releasedfrom rat C6 glioma cells. ELISA analysis of serum-free C6culture media revealed a maximal concentration of DARPby culture day 1. However, ELISA analysis of C6 culturesgrown in F-12Krserum medium revealed that maximallevels of DARP were detected on culture day 6 with a108% increase in DARP immunoreactivity from cultureday 1. Western blot analysis indicated that proteins of;60, 50, and 45 kDa are recognized by anti-DARPB9–B10 in C6 cell homogenates cultured in serum-freeand serum containing media while anti-DARP-36aa hadimmunoreactivity with the 60-kDa subunit alone. Immuno-cytochemical examination of C6 glioma cultures withDARP antibodies revealed several areas of DARP-immunoreactivity throughout the cytosol with concentratedstaining around the nucleus and in several processes of C6cells. Finally, immunocytochemical analysis of primary ratmesencephalic cultures revealed that DARP is detected inastrocytes although the majority of DARP immunoreactiv-ity was found in non-astrocyte type cells in primary mes-encephalic cultures.
When DARP release was examined in OptiMEMserum-free culture medium, we observed a maximal re-lease of DARP 24 h after OptiMEM introduction. Anestimation of the amount of DARP secreted on culture day1 in serum-free culture medium indicates that approxi-mately 95 ng of DARP was detected in a 0.5-ml mediumsample. This concentration is reasonable since it represents
Ž .about 1.2% of the total protein present 7.6 mg in theŽ .culture media sample see Figs. 1 and 2 . Following this
maximal release, DARP concentration in culture mediumremained constant through culture day 4 while cell numberexponentially increased through culture day 3. Analysis ofC6 cultures grown in F-12Krserum medium revealed a
Ž . Ž .Fig. 7. Immunocytochemical localization of DARP in C6 glioma cells cultured in OptiMEM serum-free medium. a Anti-DARP-36aa 1:100 was used asŽ . Ž . Ž .a primary antibody Ab in this culture. b Control cultures incubated with anti-DARP-36aa pre-adsorbed with DARP-36aa. c Anti-DARP B9–B10
Ž .1:25 was used as a primary Ab in this culture. Scale bar: 30 mm.
()
S.Smith,V
.D.R
amirezr
Brain
Research
8431999
95–
104102
Ž . Ž .Fig. 8. Immunocytochemical co-localization of DARP and GFAP in primary mesencephalic cell cultures. a DARP immunoreactivity visualized using a Cy3-conjugated secondary antibody. b GFAPŽ . Ž .immunoreactivity in the same field as 9a. visualized using a FITC-conjugated secondary antibody. c DARP immunoreactivity visualized using a Cy3-conjugated secondary antibody. d GFAP
immunoreactivity in the same field as 9c. visualized using a FITC-conjugated secondary antibody. Scale bar: 30 mm.
( )S. Smith, V.D. RamirezrBrain Research 843 1999 95–104 103
108% increase in DARP immunoreactivity from cultureŽ .days 1 to 6 Fig. 3b . It has been reported that neurotrophic
factor expression and release from C6 glioma cells isgreatly influenced by culture conditions. Specifically, ithas been concluded that serum-supplemented culturemedium contains factors necessary for stimulation of neu-
w x w xrotrophic factor release 18,31 . Also, Morita et al. 20have demonstrated that serum deprivation results in re-duced viability of C6 glioma cultures, a phenomenon thatwas also observed in our studies by day 4 of culture. Thesestudies and our findings suggest that the stimulation ofDARP synthesis and release from C6 glioma cells isdependent upon a factor contained in culture mediumsupplemented with animal sera. When serum-supple-mented culture medium is replaced with serum-free culturemedium, stimulation of DARP synthesis and release ap-pears to decrease markedly. ELISA examination of DARPsecretion revealed a marked increase in DARP mediumconcentration from culture days 1 to 6 in serum supple-mented cultures while levels of DARP immunoreactivityremained constant through culture day 2 with an insignifi-cant decrease on culture days 3 and 4 in serum-freecultures.
As mentioned earlier, differences were observed inDARP immunoreactivity between anti-DARP B9–B10 andanti-DARP-36aa. Western blot analysis reveals that anti-DARP B9–B10 recognizes multiple proteins, that corre-
w xspond to the reported size of purified DARP 4,16,23 , inboth cultures grown in serum containing as well as serum-free medium. Fig. 6 demonstrates that anti-DARP-36aaimmunoreactivity with the 60-kDa subunit of DARP isgreatly reduced in cultures grown in serum-free medium.We attribute the differences in DARP immunoreactivity tothe generation of these antibodies. Partial purification re-vealed that DARP is a complex multisubunit protein of200 kDa with subunits of ;60, 50, and 45 kDa. Microse-quencing the N-terminus of the 60-kDa subunit of DARPled to the synthesis of DARP-36aa which enabled thislaboratory to generate antisera against this synthetic pep-
Ž .tide anti-DARP-36aa . The monoclonal antibody, anti-DARP B9–B10, was generated against the partially puri-fied 200 kDa DARP. Lane 4 in Fig. 5 demonstrates thatDARP antibodies have distinct epitope recognition sites.Thus, anti-DARP B9–B10 may recognize a conservedregion within different DARP subunits while anti-DARP-36aa only recognizes a specific region within the first 36amino acids of the 60-kDa subunit. The generation ofmultiple epitope recognition sites would enable anti-DARPB9–B10 to recognize reduced levels of DARP found in C6glioma cells cultured in serum-free medium. Also, serum-free culture conditions have been shown to result in de-
w xcreased viability of C6 cultures 20 . This decrease inculture viability may have resulted in an alteration of theepitope recognized by anti-DARP-36aa leading to the de-creased immunoreactivity in lane 2 of Fig. 6 withouteffecting the epitope recognition site of anti-DARP B9–
B10. Further analysis, including complete sequence infor-mation for DARP, peptide competition ELISA, and BIA-
w xcore analysis 25 , is required to determine the specificepitope recognition site for anti-DARP B9–B10.
The distribution of DARP in the rat brain has beenexamined in the rat CNS using immunocytochemical tech-niques. These studies demonstrated that DARP maps in therat brain coincide with well-known TH maps for CA
w xneurons 6 . However, these studies did not define thespecific cell type that produces DARP. The findings pre-sented in this study reveal that DARP is synthesized andreleased from C6 glioma cells. The detection of DARP inC6 cells, and more importantly in primary rat mesen-cephalic cell cultures, suggests that DARP may play a rolein neuron–glial interactions.
Acknowledgements
We wish to thank Dr. M. Holzwarth for allowing us touse her culture facilities and for her helpful advice on theimmunocytochemistry work. Also, we thank Dr. S. Ro-driguez for her assistance in preparing the polyclonalantibody against DARP-36aa.
References
w x1 A. Acheson, J.C. Conover, J.P. Fandl, T.M. DeChiara, M. Russell,A. Thadani, S.P. Squinto, G.D. Yancopoulos, R.M. Lindsay, ABDNF autocrine loop in adult sensory neurons prevents cell death,
Ž .Nature 372 1995 450–453.w x Ž .2 Y.A. Barde, Trophic factors and neuronal survival, Neuron 2 1989
1525–1534.w x3 P. Benda, J. Lightbody, G. Sato, L. Levine, W. Sweet, Differentiated
Ž .rat glial cell strain in tissue culture, Science 161 1968 370.w x4 G.D. Chang, V.D. Ramirez, A potent dopamine-releasing factor is
present in high concentrations in the rat adrenal gland, Brain Res.Ž .463 1988 385–389.
w x6 W.S. Choi, B.H. Lee, V.D. Ramirez, Immunocytochemical mappingŽ .of a novel dopamine-releasing protein DARP in the rat brain,
Ž .Neuroendocrinology 60 1994 194–204.w x7 P. Ernfors, C. Wetmore, L. Olson, H. Persson, Identification of cells
in rat brain and peripheral tissues expressing mRNA for members ofŽ .the nerve growth factor family, Neuron 5 1990 511–525.
w x8 I. Ferguson, J. Schweitzer, E. Johson, bFGF receptor-mediatedinternalization metabolism and anterograde transport in retinal gan-
Ž .glion cells, J. Neurosci. 10 1990 2176–2189.w x9 W. Friedman, I. Black, D. Kaplan, Distribution of the neurotrophins
brain derived neurotrophic factor, neurotrophin-3, and neurotrophin-4r5 in the postnatal rat brain: an immunocytochemical study,
Ž .Neuroscience 84 1998 101–114.w x10 G. Hageman, M. Kirchoff-Rempe, G. Lewis, S. Fisher, D. Ander-
son, Sequestration of bFGF in the primate retinal interphotoreceptorŽ .matrix, Proc. Natl. Acad. Sci. U.S.A. 88 1991 6706–6710.
w x12 Z. Kokaia, J. Bengzon, M. Metsis, M. Kokaia, H. Persson, O.Lindvall, Coexpression of neurotrophins and their receptors in neu-rons of the central nervous system, Proc. Natl. Acad. Sci. U.S.A. 90Ž .1993 6711–6715.
w x13 S. Kuhananthan, S.D. Miklasz, V.D. Ramirez, Monoclonal antibod-
( )S. Smith, V.D. RamirezrBrain Research 843 1999 95–104104
Ž .ies against a dopamine-releasing protein DARP arrest fetal devel-opment, decrease brain catecholamines, and increase adrenal weight
Ž .of neonatal rats, Mol. Cell. Neurosci. 2 1991 410–417.w x14 S. Kuhananthan, V.D. Ramirez, An ELISA assay to detect
Ž .dopamine-releasing protein DARP and potential clinical applica-Ž .tions, Soc. Neurosci. Abstr. 670.1 1994 .
w x15 R. Levi-Montalcini, The nerve growth factor 35 years later, ScienceŽ .237 1987 1154–1162.
w x Ž16 D.A. Llano, V.D. Ramirez, Isolation of DARP Dopamine-Releasing.Protein from fetal rat brain and effects of DARP immunoneutraliza-
w x17 B. Lu, M. Yokoyama, C. Dreyfus, I. Black, NGF gene expression inŽ .actively growing brain glia, J. Neurosci. 11 1991 318–326.
w x18 D. Monard, R. Lindsay, Y. Schurch-Rathgeb, Problems in theŽ .detection of glial factor activity, Drug Res. 27 1977 477.
w x19 G. Moretto, D. Walker, P. Lanteri, F. Taioli, S. Zaffagini, R. Xu, N.Rizzuto, Expression and regulation of glial-cell-lined-derived neu-
Ž .rotrophic factor GDNF mRNA in human astrocytes in vitro, CellŽ .Tissue Res. 286 1996 257–262.
w x20 K. Morita, K. Ishimura, Y. Tsuruo, D. Wong, Dexamethasoneenhances serum deprivation-induced necrotic death of rat C6 gliomacells through activation of glucocorticoid receptors, Brain Res. 816Ž .1999 309–316.
w x21 S.M. Phelps, V.D. Ramirez, Unilateral deficits induced in rats byMPPq are markedly reduced by an N-terminal peptide fragment of
Ž .dopamine-releasing protein, Brain Res. 666 1994 117–119.w x22 D. Purves, The trophic theory of neural connections, Trends Neu-
Ž .rosci. 9 1986 486–489.
w x23 V.D. Ramirez, F. Marcus, Dopamine-releasing protein and antibody,Patent no. 5, 149,786, 1992.
w x24 J. Rudge, R. Alderson, E. Pasnikowski, J. McClain, N. Ip, R.Lindsay, Expression of ciliary neurotrophic factor and the neu-rotrophins — nerve growth factor, brain-derived neurotrophic factor,and neurotrophin 3 — in cultured rat hippocampal astrocytes, Eur. J.
Ž .Neurosci. 4 1992 459–471.w x25 W. Sandrock, J. Goodearl, W. Yin, D. Chang, D. Fischbach, ARIA
is concentrated in nerve terminals at neuromuscular junctions and atŽ .other synapses, J. Neurosci. 15 1995 6124–6136.
w x26 D. Schaar, B. Sieber, C. Dreyfus, I. Black, Regional and cell-specificŽ .expression of GDNF in rat brain, Exp. Neurol. 124 1993 368–371.
w x27 D. Schaar, B. Sieber, A. Sherwood, D. Dean, G. Mendoza, L.Ramakrishnan, S. Dreyfus, I. Black, Multiple astrocyte transcriptsencode nigral trophic factors in rat and human, Exp. Neurol. 130Ž .1994 387–393.
w x29 C. Suter-Crazzolara, K. Unsicker, GDNF mRNA levels are inducedŽ .by FGF-2 in rat C6 glioblastoma cells, Mol. Brain Res. 41 1994
175–182.w x30 A. Verity, T. Wyatt, B. Hajos, R. Eglen, P. Baecker, R. Johnson,
Regulation of glial cell line-derived neurotrophic factor release fromŽ .rat C6 glioblastoma cells, J. Neurochem. 70 1998 531–539.
w x31 R. Westermann, M. Hardung, D. Meyer, P. Ehrhard, U. Otten, K.Unsicker, Neuronotrophic factors released by C6 glioma cells, J.
Ž .Neurochem. 50 1988 1747–1758.w x32 F. Zafra, D. Lindhoim, E. Castren, J. Hartikka, H. Thoenen, Regula-
tion of brain-derived neurotrophic factor and nerve growth factormRNA in primary cultures of hippocampal neurons and astrocytes,