Pain, 41 (1990) 71-80 Elsevier 71 PAIN 01564 Central effect of the non-steroid anti-inflammatory agents, indometacin, ibuprofen, and diclofenac, determined in C fibre-evoked activity in single neurones of the rat thalamus Ilmar Jurna a and Kay Bnme b a Institut ftir Pharmakologie und Toxikologie, Universittit des Saarlandes, D-6650 Homburg/ Saar (F. R. G.), and b Institut ftir Pharmakologie und Toxikologie, Universitiit Niirnberg-Erlangen, D-8520 Erlangen (F.R. G.) (Received 31 October 1989, accepted 10 November 1989) SummarY This study aimed to investigate if the non-steroid anti-inflammatory agents, indometacin, ibuprofen, and diclofenac, are capable of depressing sensory responses of the nociceptive system by a central action. For this purpose, experiments were carried out on rats under urethane anaesthesia in which activity was elicited by electrical stimulation of afferent C fibres in the sural nerve. Recordings were made ipsi- or contralaterally from single neurones in the dorsomedial part of the ventral nucleus (VDM) of the thalamus. The 3 drugs produced a dose-dependent depression of the evoked activity which amounted to about 60% of the controls at the highest doses employed and lasted longer than 60 min. Their potency ranking, according to the EDs, values (in brackets), is: indometacin (5 mg/kg) > diclofenac (10.9 mg/kg) > ibuprofen (15.6 mg/kg). The results suggest that a central action might contribute to the analgesia produced by these non-steroid anti-inflammatory agents. Key words: Thalamus; Nociception; Analgesia: Non-steroid anti-inflammatory agents; Indometacin; Ibuprofen; Diclofenac; Central action Introduction Analgesic agents are frequently divided into 2 groups of drugs which can be classified as acting centrally or peripherally. This classification de- rives from the results obtained by Lim and co- workers [28,42] in an animal model of visceral pain elicited by bradykinin. This showed that morphine abolished the nociceptive responses by acting on the central nervous system while non- opioid analgesic agents such as acetylsalicylic acid, Correspondence to: Prof. Dr. Ilmar Juma, Institut ftir Pharmakologie und Toxikologie, Universitgt des Saarlandes, D-6650 Homburg/f&r, F.R.G. some pyrazolone derivatives and the precursor of paracetamol, phenacetin, acted in the periphery. The concept of a peripheral action of analgesic agents possessing anti-inflammatory and anti- pyretic properties gained strong support from the observation that some prostaglandins potentiate nociceptive responses to various algogenic sub- stances [10,15,18,29,30,36], and that non-opioid analgesic or non-steroid anti-inflammatory agents inhibit the formation [17,19,54] and the release of prostaglandins induced by algogenic substances in different tissues [16,32,46,58]. Moreover, it was demonstrated that acetylsalicyclic acid reduced the activation of nociceptive responses elicited by al- gogenic substances [26,45]. However, there are some arguments against an 0304-3959/90/$03.50 0 1990 Elsevier Science Publishers B.V. (Biomedical Division)
Transcript
Pain, 41 (1990) 71-80
Elsevier
71
PAIN 01564
Central effect of the non-steroid anti-inflammatory agents, indometacin, ibuprofen, and diclofenac, determined in C
fibre-evoked activity in single neurones of the rat thalamus
Ilmar Jurna a and Kay Bnme b
a Institut ftir Pharmakologie und Toxikologie, Universittit des Saarlandes, D-6650 Homburg/ Saar (F. R. G.),
and b Institut ftir Pharmakologie und Toxikologie, Universitiit Niirnberg-Erlangen, D-8520 Erlangen (F.R. G.)
(Received 31 October 1989, accepted 10 November 1989)
SummarY This study aimed to investigate if the non-steroid anti-inflammatory agents, indometacin, ibuprofen, and diclofenac,
are capable of depressing sensory responses of the nociceptive system by a central action. For this purpose, experiments were carried
out on rats under urethane anaesthesia in which activity was elicited by electrical stimulation of afferent C fibres in the sural nerve.
Recordings were made ipsi- or contralaterally from single neurones in the dorsomedial part of the ventral nucleus (VDM) of the
thalamus. The 3 drugs produced a dose-dependent depression of the evoked activity which amounted to about 60% of the controls at
the highest doses employed and lasted longer than 60 min. Their potency ranking, according to the EDs, values (in brackets), is:
indometacin (5 mg/kg) > diclofenac (10.9 mg/kg) > ibuprofen (15.6 mg/kg). The results suggest that a central action might
contribute to the analgesia produced by these non-steroid anti-inflammatory agents.
Analgesic agents are frequently divided into 2 groups of drugs which can be classified as acting centrally or peripherally. This classification de- rives from the results obtained by Lim and co- workers [28,42] in an animal model of visceral
pain elicited by bradykinin. This showed that morphine abolished the nociceptive responses by
acting on the central nervous system while non- opioid analgesic agents such as acetylsalicylic acid,
Correspondence to: Prof. Dr. Ilmar Juma, Institut ftir
Pharmakologie und Toxikologie, Universitgt des Saarlandes,
D-6650 Homburg/f&r, F.R.G.
some pyrazolone derivatives and the precursor of paracetamol, phenacetin, acted in the periphery. The concept of a peripheral action of analgesic agents possessing anti-inflammatory and anti- pyretic properties gained strong support from the
observation that some prostaglandins potentiate nociceptive responses to various algogenic sub-
stances [10,15,18,29,30,36], and that non-opioid analgesic or non-steroid anti-inflammatory agents inhibit the formation [17,19,54] and the release of
prostaglandins induced by algogenic substances in different tissues [16,32,46,58]. Moreover, it was demonstrated that acetylsalicyclic acid reduced the activation of nociceptive responses elicited by al- gogenic substances [26,45].
exclusively peripheral action of the non-opioid analgesic agents in depressing pain sensation. Thus, e.g., acetylsalicylic acid and some pyrazo-
lone derivatives causing analgesia are much weaker
inhibitors of prostaglandin synthesis than the more modern non-steroid anti-inflammatory agents
[6,19,21,31], and paracetamol does not inhibit the formation of prostaglandins in the periphery [7.22]
but reduces cyclooxygenase activity in brain tissue [22,23]. Still, the non-opioid analgesic agents are fairly effective and reliable in relieving weak to
moderate pain.
Some observations indicate an additional central action of these drugs. Sodium salicylate
increased the nociceptive threshold in rats to stimulation of the lateral hypothalamus [12], and
acetylsalicylic acid administered by microinjection into the preoptic region of the hypothalamus de-
pressed nociceptive responses in monkeys in a way that depended on intact functioning of mono-
aminergic pathways [55]. Indometacin and di- clofenac administered by intracerebroventricular
injection inhibited nociceptive responses in arthri- tic rats [47]. In human volunteers, acetylsalicylic acid reduced the late pain-related response in the
electroencephalogram evoked by electrical stimu- lation of the tooth [ll], and ketoprofen elevated
the threshold of nociceptive reflex activity elicited in the femoral biceps muscle by electrical stimula-
tion of the sural nerve [60].
Since electrical nerve stimulation caused release
of prostaglandins from the frog spinal cord [51] and the cat cerebral cortex [SO], it has been pro- posed that prostaglandins may facilitate nocicep- tive signal transmission in the central nervous
system, and that non-opioid analgesic or non- steroid anti-inflammatory agents abolish this faci-
litatory action [16,62]. Our own experiments have shown that activity
in single neurones in the rat thalamus, elicited by electrical stimulation of afferent C fibres in the
sural nerve, is depressed by paracetamol, lysine acetylsalicylate and the pyrazolone derivative, metamizol [8,9]. Since this model allows one to establish central drug effects, the action of the non-steroid anti-inflammatory agents, indometa- tin, ibuprofen, and diclofenac, was studied in sim- ilar experiments.
Methods
The experiments were carried out on rats of either sex (Sprague-Dawley/SIV; 250-300 g body weight). The animals were housed in macrolon cages (6 animals to a cage) and fed on standard
diet (Altromin) and tap water ad libitum. They received an intraperitoneal injection of urethane
(1.2 g/kg) to induce and maintain anaesthesia for surgery and the experiment proper. At the end of surgery, an additional subcutaneous injection of
urethane (120 mg/kg) was given. This treatment warranted a sleeping time of more than 6 h [8],
during which no righting reflexes were present; while preparing the animals, searching for neu-
rones in the thalamus and recording of activity before and after drug administration lasted no
longer than 3 h. The animals breathed sponta- neously. Body temperature was monitored in the rectum and kept between 37.5 and 38’ C by radiant heat.
The procedure to prepare the animals for the experiment and to elicit and record nociceptive
activity from thalamic neurones has already been described in detail [9]. A brief summary is pre- sented: a cannula was inserted into a tail-vein for i.v. injection. The left sural nerve was prepared for electrical stimulation with a pair of platinum wire
electrodes and cut distal to the electrodes to ex-
clude drug actions in the nociceptor region. The nerve was stimulated employing single rectangular
impulses delivered from a Grass stimulator (Model S44) with stimulus isolation unit at a frequency of 0.1 Hz. an impulse duration of 0.5 msec and a strength that was 2-2.5 times higher than that producing maximum responses and was supra-
maximal for afferent C fibres in the sural nerve (48-68 V). These stimulation parameters have been employed in a previous investigation [37] in which activity in single axons of the rat spinal cord was elicited by electrical stimulation of the sural nerve. Recording of activity from single neurones in the dorsomedial part of the ventral nucleus (VDM) of the thalamus on the left or right side was per- formed with tungsten microelectrodes (tip diame- ter 1 pm; resistance 10 MO) at the coordinates AP: 1.9-2.7 mm, L: 2.5 mm, and V: 5.0-7.0 mm [20]. Neurones responding to stimulation of noci-
73
ceptive afferents in the sural nerve were recorded ipsi- or ~ntralater~ly, i.e., on the left or right side. At the end of the experiments, the position of the microelectrode tip was marked by passing a current of approximately 4 mA for 30 set (Grass lesion maker). The animals were killed with an overdose of pentobarbital and the brain was per- fused by an intraarterial injection of a 10% for- maldehyde solution. The brain was removed, fixed in Bouin’s solution and embedded in paraplast. Serial sections (10 pm) were stained with gallo-
b control
.
Indometacin 8 mg/kg t.v.
cyanin-chromalum [14] and counterstained with phloxin.
The activity of singIe neurones or axons was amplified (WPI preamplifier Model DAM-SA) and displayed on a cathode ray oscilloscope (Fig. lb). Signals recorded from thalamic neurones were passed through a window discriminator (WPI Model 121) and evaluated with a personal com- puter (Tandon XT 10) and MBATE software. The number of addresses used was 256 and the dura- tion of each address 8 msec. Peristimulus histo-
lndometacin 8 mg/kg i.v.
I. counts
50 minrlg
Fig 1. Depression by indometacin of activity in single neurones of the thalamus VDM evoked by sural nerve stimulation. a: site of recording the activity presented in b and c. b: activation produced by supramaximal electrical stimulation (52 V) of the ipsilateral sural nerve before and after intravenous injection of indometacin. Stimulus artifacts are indicated by dots under recordings. c: peristimulus histograms (10 trials each) of the activity. Stimulation is indicated by dots under the recordings. Horizontal scales: total time of recording. Vertical scale: number of impulse discharges counted in each computer address (total number of addresses 256).
Rat under urethane anaesthesia.
74
grams consisting of 10 consecutive responses of thalamic neurones to sural nerve stimulation were
summed each time and electronically integrated. Spontaneous and evoked activities were treated separately. The interval between each measure-
ment consisting of 10 consecutive responses was 5 min or longer. The integrations of activity were
pooled for statistical evaluation. Four to 6 de- terminations were made before drug administra-
tion and these served as controls when they were stable. When 6-8 determinations showed changes
of more than 10% of the mean evoked activity, the neurone was abandoned and another one was
searched for. Only one injection was given to each animal. Therefore, the number of thalamic neu-
tones or ascending axons from which recordings
were made, the number of rats employed and the number of experiments carried out are identical.
Significant differences were established by apply- ing Student’s t test for paired or unpaired sam-
ples.
The drugs used were diclofenac (sodium salt: Sigma), gallamine triethiodide (Flaxedilm, Abbott). ibuprofen (Sigma), indometacin (sodium salt. ~~onfortid~; Dumex) and urethane (Riedel-De
HaEn). Diclofenac and indometacin were injected intravenously; ibuprofen was administered by in-
traperitoneal injection. The doses of diclofenac and indometacin indicated in the text refer to the
salts.
Results
In total. 70 neurones in the VDM nucleus of the rat thalamus responding to suprama~mal elec- trical stimulation of the ipsi- or contralateral sural nerve were studied. Forty-five neurones were
activated from the ipsilateral sural nerve and 25 neurones from the contralateral nerve. Peristimu- lus histograms, as shown in Fig. la, were obtained by sampling 10 consecutive responses. No ‘wind-
up’ of spontaneous activity occurred when supramaximal stimulation of the sural nerve was applied at the rate (0.1 Hz) chosen for these experiments. When the strength of stimulation was reduced below that causing activation of ascending axons in the spinal cord from nocicep-
tive afferents in the sural nerve [37], i.e., below 20 V, none of the neurones tested responded to stimulation. Moreover, all neurones responding to sural nerve stimulation could also be activated by
squeezing more than one paw and pinching the skin of various parts of the body. However, none
of these neurones responded to touching or gentle stroking of the skin, nor to air puffs applied to it. The interval between the stimulus and the maxi- mum of evoked activity in the peristimulus histo-
grams of the controls varied between 100 and 300 msec [see also ref. 91. In Fig. lc, the interval was 290 msec and the distance between the proximal
stimulation electrode (cathode} and the dorsal root entry zone containing afferents from the sural nerve was 82 mm. This yields an apparent conduc-
tion velocity of 0.28 m/set which would be in accordance with activation by afferent C fibres.
In experiments carried out on 7 thalamic neu-
rones. saline injected intravenously in the same volume (1 ml/kg) as the i.v. administered drugs (indometacin and diclofenac) caused no signifi-
cant change in the evoked activity. An intravenous injection of urethane (120 mg/kg) made in 4 additional experiments was not followed by a change in the activity evoked in thalamic neu- rones. Since all neurones were influenced in the
same way by the drugs tested. evaluation of drug effects was performed without distinguishing be- tween ipsi- and contralateral activation.
lndometacin injected intravenously at a dose of 8 mg/kg markedly reduced the activity evoked in
thalamic neurones (Fig. la and b). The depression amounted to about 60% of the control activity between 5 and 20 mm after the injection and was si~ificant from 5 min until the end of the experi-
ment at 60 min (Fig. 2af. .A lower dose of in- dometacin (4 mg/kg) was less effective than the high dose, but it significantly reduced the evoked activity from 5 to 30 mm after the injection. The lowest dose of indometacin tested (2 mg/kg) pro- duced no significant effect (Fig. 2af. From the dose-response relationship of the effects de- termined 10 min after the administration, an ED,,,
of 5 mg/kg was calculated (Fig. 2b). Ibuprofen administered by intraperitoneal in-
jection at a dose of 20 mg/kg caused a significant and long-lasting depression of activity evoked in
the thalamus that amounted to about 60% of the control activity (Fig. 3a). A lower dose of ibupro- fen (10 rn~~kg) was Sess effective but ~~~~f~~t~~ reduced the activity by 30% of the controls. The lowest dose of ibuprofen (5 mgfkg) failed to affect the evoked activity (Fig. 3a). From the
120
L@4
A A I
%
100 /
f 60 If
Fig. 2. Depression by indometacin of activity evoked in thalamic neuranes of the rat. a: time course of the depression
caused by intravenous injection of 1 different doses of in-
dometacin. Ordinate: changes in activity in per cent of the
mean value of the controls. Abscissa: time in mm before and
after injeztion. Points in the curves are mean values of de-
ter~a~o~~ %&xx nmnber is indicated in brackets for each
dose of indometacin. Open symbols: values differ si@nficautiy
from controls (P < 0.05 or less). b: dose-response line of the
depressant effect of indometacin established from the values
obtained 10 min after injectiou. Ordinate: indomet&n-in-
duced change in activity in per cent of mean values of the
eontrois. Abscissa: dose of indometacin on ~uga~~~~ s&e.
a
120
w
100
2a “PV ” 20 40 60 mln
,, r
KWROFEN tp.
*o 6aglkg it+8)
*a f# mgfkg fN.77
AD 20 mglkg (N*5)
b OOI- .*
91
60
EOaa * 15.6 me/kg
0 r 80.66
p =o.aaaa
5 10 2a v/kg Fig. 3. Depression by ibuprafen of activity evoked in thalamic ueurones of the rat. Ibuprofen was administered by in-
trap&oneaZ injection. Other details as in legend to Fig. 2.
dose-response re’lationship of the depressant cf- feet produced by ibuprofen 10 min after the injec- tion, an EDso of 15.6 mg/kg was calculated (Fig. 3b).
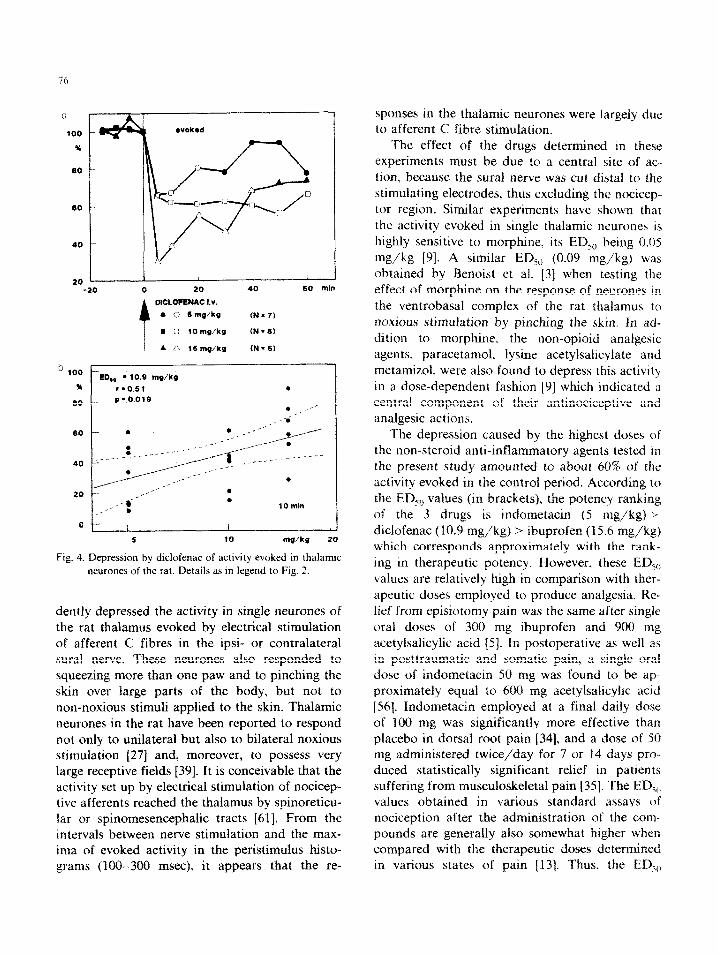
~~~~ufena~ injected ~n~av~ous~~ at doses of 15 mg/kg, If3 mg/kg> and 5 mg/kg dose-depen- dently and significantly reduced the activity evoked in thalamic neurones (Fig. 4a). The EDsv of diclofenac established from the dose-response retationship 10 mm after the inje&on is 10.9 mgJkg (Fig_ 4b).
Discmssion
The 3 non-steroid anti-inff ammatory agents, in- dometa~~n~ ibnprofen, and dicfofenac, dose-depen-
20 1 -20
I / I 0 20 40 60 mln
DICLOFENAC 1.~.
l 0 Smg/kg fN=7)
M n lOmp/kg <N=8)
A P. 15mg/kQ 1N=6)
b 100
*
60
40
20
ED,. - 10.9 mg/kg rg0.51
0 / / I 5 10 mu/k9 20
Fig. 4. Depression by diclofenac of activity evoked in thalamic
neurones of the rat. Details as in legend to Fig. 2.
dently depressed the activity in single neurones of the rat thaiamus evoked by electrical stimulation of afferent C fibres in the ipsi- or contralateral sural nerve. These neurones also responded to squeezing more than one paw and to pinching the skin over large parts of the body, but not to
non-noxious stimuli applied to the skin. Thalamic neurones in the rat have been reported to respond not only to unilateral but also to bilateral noxious stimulation [27] and, moreover, to possess very large receptive fields [39]. It is conceivable that the activity set up by electrical stimulation of nocicep- tive afferents reached the thalamus by spinoreticu- lar or spinomesencephalic tracts [61]. From the intervals between nerve stimulation and the max- ima of evoked activity in the peristimulus histo- grams (100-300 msec), it appears that the re-
sponses in the thalamic neurones were largely due to afferent C fibre stimulation.
The effect of the drugs determined in these experiments must be due to a central site of ac- tion, because the sural nerve was cut distal to the
stimulating electrodes. thus excluding the nocicep- tor region. Similar experiments have shown that the activity evoked in single thalamic neurones is
highly sensitive to morphine, its ED,,, being 0.05 mg/kg [9]. A similar EDS,, (0.09 mg/kg) was obtained by Benoist et al. [3] when testing the
effect of morphine on the response of neurones in the ventrobasal complex of the rat thalamus to noxious stimulation by pinc~ng the skin. In ad-
dition to morphine. the non-opioid anatgesic agents, paracetamol, Iysine acetylsalicylate and
metamizol, were also found to depress this activity in a dose-dependent fashion [9] which indicated a central component of their antinociceptive and analgesic actions.
The depression caused by the highest doses of the non-steroid anti-inflammatory agents tested in the present study amounted to about 60% of the activity evoked in the control period. According to
the EDsCt values (in brackets), the potency ranking of the 3 drugs is indometacin (5 mg/kg) >
diclofenac (10.9 mg/kg) > ibuprofen (I 5.6 mg/kg) which corresponds approximately with the rank-
ing in therapeutic potency. However, these EL&,) values are relatively high in comparison with ther-
apeutic doses employed to produce analgesia. Re- lief from episiotomy pain was the same after single oral doses of 300 mg ibuprofen and 900 mg acetylsalicylic acid [5]. In postoperative as well as in posttraumatic and somatic pain, a single oral
dose of indometacin 50 mg was found to be ap- proximately equal to 600 mg acetylsalicylic acid [56]. Indometacin employed at a final daily dose of 100 mg was significantly more effective than
placebo in dorsal root pain [34], and a dose of 50 mg administered twice/day for 7 or 14 days pro- duced statistically significant relief in patients suffering from musculoskeletal pain [35]. The ED,,, values obtained in various standard assays of nociception after the administration of the COIII-
pounds are generally also somewhat higher when compared with the therapeutic doses determined in various states of pain f13]. Thus. the ED,,,
values of indometacin ranging between 0.5 and 30 mg/kg correspond to a single oral pain relieving dose of 30 mg (i.e., about 0.4 mg/kg), and the ED,, values of ibuprofen ranging between 0.9 and 25 mg/kg correspond to a single therapeutic dose of 400 mg (i.e., about 5 mg/kg). This obviously means that a central and a peripheral action con- tribute to the relief of pain caused by these drugs. It is interesting to note that in rats with adjuvant arthritis, indometacin reduced the activity evoked in thalamic neurones by transcutaneous electrical stimulation or electrical stimulation of the tibialis nerve, the respective ED,, values being markedly lower than in the present experiments, i.e., 0.32 mg/kg and 0.66 mg/kg [48]. This depression was explained in terms of an inhibition of prostaglan- din synthesis in the central nervous system and the periphery, and it would require that the in- flammatory process in the periphery lowered the threshold of thalamic neurones to electrical nerve stimulation. It has also been reported that di- clofenac caused a depression of about 50% of the activity in rat thalamus neurones elicited by apply- ing pressure to an inflamed joint [l]. This effect was produced by a dose of 6 mg/kg, which is about half the ED,, of diclofenac determined in the present experiments, and it was attributed to a peripheral action of the drug. Alternatively, it must be taken into account that chronic in- flammatory processes may increase the respon- siveness of the nociceptive system and that in this condition analgesic agents may be more effective. It has been demonstrated in experiments carried out on arthritic rats that dorsal horn neurones normally responding only to noxious stimuli ex- hibited extremely high spontaneous activity and could also be activated by non-noxious stimuli [44], or that these neurones had unusually large receptive fields [33]. Moreover, neurones in the thalamus responding to noxious stimulation may be activated in arthritic rats by otherwise innocu- ous stimuli [25]. These changes in the behaviour of nociceptive neurones are attributed to central rather than peripheral events resulting from the inflammation. It is in accordance with this view that not only the antinociceptive effect of acetyl- salicylic acid 1491 but also that of morphine [38] were found to be enhanced in arthritic rats, It is
conceivable that the EDso values of the central effects of the drugs deter~ned in this study would be lower in arthritic rats.
Various mechanisms could account for a central action of the non-steroid anti-inflammatory agents. In the first instance, these drugs might interfere with the formation of prostaglandins involved in nociceptive transmission (see Introduction). An- other possibility is that they act by interfering with transmitters or modulators in the nociceptive system. A central antinociceptive effect of acetyl- salicylic acid apparently depended on an intact catechola~ner~c function 1551. It has been sug- gested that diclofenac-induced analgesia is media- ted in part by endogenous opioid peptides [43,59], or that it blocks the release of serotonin induced by central administration of bradykinin [4].
Furthermore, a correlation has recently been reported to exist between the depression of noci- ceptive activity evoked in thalamic neurones of rats and the levels and turnover of serotonin in the brain of the animals caused by lysine acetylsalicy- late [24]. This correlation might be explained in terms of a release of tryptophan from its albumin binding sites by the salicylic acid derivative [57]. Such a mechanism could also explain why the 3 drugs tested in the present study were almost equally effective despite the considerable dif- ferences in their clinical potency.
Finally, it is known that salicylic acid and some of its derivatives markedly change the electrophys- iological properties of neuronal membranes [2,40,41,52] and block impulse conduction in nerve fibres [53], but similar observations have not yet been made with other non-steroid anti-infla~a- tory agents. Although their particular mode of action needs to be elucidated, the results presented provide further evidence that non-steroid anti-in- flammatory agents can cause analgesia by a central action.
Acknowledgements
This study was supported by the Schwerpunkt ‘Nociception und Schmerz’ of the Deutsche For- schungsgemeinschaft (Ju 27/8-l). Indometacin for i.v. injection was kindly provided by Dumex, Cope~agen/Denmark.
The authors are grateful to Mrs. Birgit Spohrer and Mrs. Karin Wolske for skillful technical assis-
tance.
References
1
2
3
4
5
6
7
x
9
10
II
12
Attal, N., Kayser, V., Eschalier. A.. Benoist, J.M. and
Guilbaud. G., Behavioural and electrophysiological evi-
dence for an analgesic effect of non-steroidal anti-in-
57 Tagliamonte, A., Tagliamonte, P., Perez-Cruet, J.. Stem, S.
and Gessa, G.L., Effect of psychotropic drugs on
tryptophan concentrations in the rat brain, J. Pharmacol. Exp. Ther., 177 (1971) 475-480.
58 Vane, J.R., Inhibition of prostaglandin synthesis as a mech-
anism of action for aspirin-like drugs, Nature (New Biol.),
231 (1971) 232-235.
59 Vescovi, P., Passeri, M.. Gerra, G. and Grossi, E., Nal-
oxone inhibits the early phase of diclofenac analgesia in man, Pain Clin., 1 (1987) 151-155.
60 Willer, J.C. et Harrewyn, J.M., Effet inhibiteur central du
ketoprofene intraveneux sur le r&exe nociceptif de flexion 62 Yaksh, T.L., Central and peripheral mechanisms for the chez I’homme, Presse Med., 16 (1987) 63-67. antialgesic action of acetylsalicylic acid. In: H.J.M. Bamett.
61 Willis, W.D., The Pam System, The Neural Basis of Noci- J. Hirsh and J.F. Mustard (Eds.), Acetyls~icyIi~ Acid: New
ceptive Transmission in the Mammalian Nervous System, Aspects for an Old Drug, Raven Press, New York, 1982.