CHANGES IN THE ACTIVITY AND SUBCELLULAR DISTRIBUTION OF PPO IN ‘DELICIOUS’ APPLES DURING CONTROLLED ATMOSPHERE STORAGE DIANE M. BARRETT’ Department of Food Science and Technology Oregon State University Corvallis, OR 97331 C. Y. LEE Department of Food Science and Technology New York State Agricultural Experiment Station Cornell University Geneva, NY 14456 and FRANK W. LIU Department of Pomology Cornell University Ithaca, NY 14850 Received for Publication September 4, 1990 Accepted for Publication April 8, 1990 ABSTRACT Polyphenol oxidase (PPO) activify in 4,000 X g, 100,000 X g, 200 x g and soluble fractions of ‘Delicious’ apples was monitored during 28 weeks of controlled atmosphere storage. PPO activity in 4,000 X g and 100,000 X g fractions decreased, while that in soluble and 200 X g fractions increased. These shifts in subcellular location occurred sooner in apples which were stored under high CO, conditions (2.54% O,, 8-12% CO,) than those under normal CA conditions (2% O,, 3% CO,). Polyacrylamide gel electrophoresis of 4,000 x g fractions isolated from ‘Delicious’ apples at harvest showed 2 bands with ‘Address correspondence to: Dr. Diane M. Barrett, Dept. of Food Science & Technology, Wiegand Hall, Oregon State University, Corvallis, OR 97331. Journal of Food Biochemistry 15 (1991) 185-199. All Rights Reserved. 0 Copyright 1991 by Food & Nutrition Press, Inc., Trumbull, Connecticut. 185
Transcript
CHANGES IN THE ACTIVITY AND SUBCELLULAR DISTRIBUTION OF PPO IN ‘DELICIOUS’ APPLES
DURING CONTROLLED ATMOSPHERE STORAGE
DIANE M. BARRETT’
Department of Food Science and Technology Oregon State University
Corvallis, OR 97331
C. Y. LEE
Department of Food Science and Technology New York State Agricultural Experiment Station
Cornell University Geneva, N Y 14456
and
FRANK W. LIU
Department of Pomology Cornell University Ithaca, N Y 14850
Received for Publication September 4, 1990 Accepted for Publication April 8, 1990
ABSTRACT
Polyphenol oxidase (PPO) activify in 4,000 X g, 100,000 X g, 200 x g and soluble fractions of ‘Delicious’ apples was monitored during 28 weeks of controlled atmosphere storage. PPO activity in 4,000 X g and 100,000 X g fractions decreased, while that in soluble and 200 X g fractions increased. These shifts in subcellular location occurred sooner in apples which were stored under high CO, conditions (2 .54% O,, 8-12% CO,) than those under normal CA conditions (2% O,, 3% CO,). Polyacrylamide gel electrophoresis of 4,000 x g fractions isolated from ‘Delicious’ apples at harvest showed 2 bands with
‘Address correspondence to: Dr. Diane M. Barrett, Dept. of Food Science & Technology, Wiegand Hall, Oregon State University, Corvallis, OR 97331.
Journal of Food Biochemistry 15 (1991) 185-199. All Rights Reserved. 0 Copyright 1991 by Food & Nutrition Press, Inc., Trumbull, Connecticut. 185
186 D. M . BARREIT, C. Y. LEE and F. W. LIU
PPO activio, while stored samples all had 3 bands. Isoenzyme patterns were found to vary in digerent subceilular fractions.
INTRODUCTION
Apples are frequently stored in controlled atmosphere (CA) conditions which utilize low 0, (-1-3%) and high CO, (-1-5%) concentrations to slow respi- ration and maintain high quality fruit for longer periods of time. High CO, concentrations (>7%) have also been used as prestorage treatments when cooling facilities are limited, and some investigators (Couey and Olson 1977; Pomtt and Meheriuk 1977; Mattus 1982) report improvements in firmness. Deviations from optimum 0, or CO, concentration often result in undesirable changes in fruit quality, and brown discoloration of external or internal tissues is one of the most serious symptoms of improper storage conditions.
Enzymatic browning of fruit tissues is catalyzed primarily through the oxi- dation of polyphenolic substrates, located in the vacuole or cytoplasm, by the enzyme polyphenol oxidase (PPO), which is located primarily in the plastids. In the intact tissue, compartmentalization prevents the enzymatic browning re- action in the presence of oxygen. However, membrane weakening under con- ditions of senescence, mechanical injury or stress, such as chilling injury, drought, or exposure to extremes in 0, or CO,, may result in decompartmen- talization and facilitation of enzymatic browning.
Numerous investigators have proposed that changes in cell membrane penne- ability and release of either stored substrates from the vacuole or bound enzymes from organelles may be involved in enzymatic browning. Hare1 et al. (1964) were the first to report a soluble PPO which increased in activity during apple ripening, and chloroplast and mitochondria1 fractions which decreased in activity. Since that time, others (Walker and Wilson 1975; Stelzig et al. 1972; Vamos- Vigyazo et al. 1985b) have noted the existence of soluble and membrane-bound forms of the enzyme. Today there is general agreement that the soluble form originates from the particulate form, and that this solubilization takes place to a greater degree as ripening and senescence progress.
The primary objective of this study was to examine the effects of a low O,, high CO, atmosphere on the activity and subcellular distribution of apple PPO during a typical storage season. Comparison was made to an atmosphere with a relatively higher CO, (8-12%) content, in order to determine whether advan- tages existed over normal CA conditions in terms of improved apple quality. PPO activity and subcellular location were also examined after apples were removed from CA storage and held in air at 18°C for 7 days, simulating a marketing situation. As another means of following changes in PPO activity and distribution during storage, subcellular fractions of all samples were analyzed using polyacrylamide gel electrophoresis.
SUBCELLULAR ACTIVITY PPO I87
EXPERIMENTAL
Raw Materials and Storage Conditions
‘Delicious’ apples were harvested from Cornell University Orchards and a flow-through simulated CA storage system, as described by Liu and Samelson (1986) was utilized. Five gallon (19 L) glass jars, each containing 45 apples from the same tree, were stored at 0” * 1°C. Gas mixtures containing the appropriate amounts of O,, CO,, and N, were mixed and humidified as described previously (Liu and Samelson 1986) and flowed at a rate of 200 mL/min.
Apples were stored under normal CA and varying CO, conditions during both 1987/88 and 1988/89. The same trends were observed during both years; there- fore, only the representative data from the 1988/89 season will be presented. The conditions for the varying CO, storage environment were modeled after Liu and Pan (1989). Normal CA gas mixtures were composed of 2% 0,, 3% CO,, 95% N,. The varying CO, condition changed with time in storage as described in Table 1.
Apples were removed from storage after 7 , 14, 21 and 28 weeks in the first year study and 7, 11.5, 14, 21 and 28 weeks in the second year study. A portion of the apples was analyzed immediately, while the other portion was analyzed after being kept in regular air storage at 18°C for 7 days. The latter treatment was representative of a simulated marketing condition.
Polyphenol Oxidase Extraction and Subcellular Fractionation
Polyphenol oxidase was separated into ‘plastid’ (4,000 X g), ‘mitochondrial’ (100,000 X g), ‘cell wall’ (200 X g) and soluble fractions using a modified method of Mayer er al. (1964). One hundred grams of apple flesh were ho- mogenized with 100 mL of cold extraction buffer (0.2 M Na,HPO,-NaH,PO, buffer with 0.4 M sucrose, pH 7.3) which contained 0.01 M sodium ascorbate,
188 D. M. BARRETT, C. Y. LEE and F. W. LIU
for 1 min. The homogenate was filtered through a 10 pm milk filter (Agway, Inc., Syracuse, New York), the pH of the filtrate was adjusted to 7.3 with 1 .O N NaOH and then centrifuged (Sorvall RC-5B Refrigerated Superspeed Centri- fuge) at 200 X g for 5 min. The supernatant was decanted and saved, and the pellet was stirred gently in an additional 100 mL of extraction buffer, and centrifuged again at 200 X g for 10 min. The pellet containing the 200 X g fraction was resuspended in 25 mL of extraction buffer, and the supernatant was combined with that obtained earlier.
The fraction containing the ‘plastids’ was obtained by centrifuging the su- pernatant for 15 min at 4,000 X g. The precipitate was washed with an additional 100 mL extraction buffer and centrifuged again at 4,000 X g for 15 min. The supernatant was decanted and combined with that obtained earlier, and the pellet was resuspended in 100 mL extraction buffer. The supernatant was finally cen- trifuged for 1 h at 100,000 X g (Beckman L2-65B Ultracentrifuge). The pellet obtained, which contained the ‘mitochondrial’ fraction, was resuspended in 10 mL extraction buffer. The resultant supernatant, referred to here as the soluble fraction, was dialyzed overnight against 0.05 M Na,HPO,-NaH,PO, buffer and analyzed the following day.
The PPO assay procedure used was a modification by Lee and Smith (1979) of the colorimetric method of Ponting and Joslyn (1948). Two hundred pL of enzyme extract were added to 2.8 mL of a 0.05 M catechol (Sigma Chemical Co.) solution in 0.2 M Na,HPO,-0.1 M H,C,H,O, buffer, pH 6.5, at 25°C and the change in absorbance at 420 nm was recorded. One unit of enzyme activity was defined as the quantity of enzyme responsible for a change in absorbance of 0.00 Umin.
Polyacrylamide Gel Electrophoresis
Polyacrylamide gel electrophoresis (PAGE) was carried out under discontin- uous nondenaturing conditions as described by Laemmli (1970) and Davis (1964). The separating gel and stacking gel were composed of 10.0% and 4.0% acrylamide, respectively. ‘Plastid’ (4,000 x g), ‘mitochondrial’ (100,OOO x g), and ‘cell wall’ (200 X g) samples, extracted as described above, were prepared by centrifugation at the appropriate centrifugal force and resuspension of the pellets in 500 pL of sample buffer (0.0625 M Tris-HCI, pH 6.8, 10% glycerol, 1.25 x bromophenol blue, 2% CHAPS, Bio-Rad). The resuspended pellet was stirred, sonicated, and the suspensions were centrifuged at 1900 X g for 20 min in a Centricon microconcentrator. Soluble storage samples were also concentrated using the microconcentrator, and 10% glycerol was added prior to electrophoresis. A commercial mushroom PPO standard (EC 1.14.18.1, 2,000- 4,000 unitshg solid) and a combination of 6 molecular weight standards (Sigma Chemical Co.) were used for comparative purposes. The following standards
SUBCELLULAR ACTIVITY PPO 189
were loaded: lysozyme 14.3 kD, carbonic anhydrase 29.0 kD, P-lactoglobulin 36.8 kD, bovine albumin 66.0 kD, alcohol dehydrogenase 150.0 kD, and P-amy- lase 200.0 kD. One hundred pL aliquots of all samples were used for electro- phoresis. The gels were stained for protein using 0.1% Coornassie Blue R-250 (Bio-Rad) and for PPO activity with L-P-3, dihydroxyphenylalanine (DOPA) as described by Wissemann and Montgomery ( 1 985).
RESULTS AND DISCUSSION
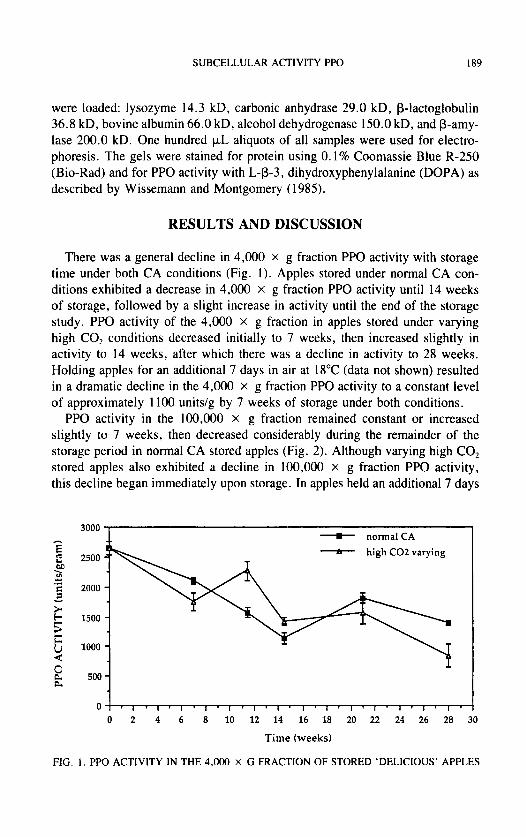
There was a general decline in 4,000 x g fraction PPO activity with storage time under both CA conditions (Fig. I ) . Apples stored under normal CA con- ditions exhibited a decrease in 4,000 X g fraction PPO activity until 14 weeks of storage, followed by a slight increase in activity until the end of the storage study. PPO activity of the 4,000 X g fraction in apples stored under varying high CO, conditions decreased initially to 7 weeks, then increased slightly in activity to 14 weeks, after which there was a decline in activity to 28 weeks. Holding apples for an additional 7 days in air at 18°C (data not shown) resulted in a dramatic decline in the 4,000 X g fraction PPO activity to a constant level of approximately 1100 units/g by 7 weeks of storage under both conditions.
PPO activity in the l00,OOO X g fraction remained constant or increased slightly to 7 weeks, then decreased considerably during the remainder of the storage period in normal CA stored apples (Fig. 2). Although varying high CO, stored apples also exhibited a decline in 100,000 x g fraction PPO activity, this decline began immediately upon storage. In apples held an additional 7 days
FIG. 2. PPO ACTIVITY IN THE 100,OOO X G FRACTION OF STORED ‘DELICIOUS’ APPLES
at 18°C (data not shown), there was a drastic decline in 100,000 X g fraction PPO activity, to approximately 600 units/g by 7 weeks of storage under both conditions.
There was a general increase in PPO activity in the soluble fraction after 14 weeks of either normal CA or high CO, storage (Fig. 3). After this time, soluble PPO activity in both immediately analyzed and air stored apples was slightly greater in apples stored under normal CA conditions than in those under high CO, conditions.
PPO activity in the 200 X g fraction remained fairly constant until 11.5 weeks of storage under both normal CA and varying high CO, conditions (Fig. 4). At 14 weeks in varying high CO, stored apples and at 21 weeks in normal CA stored apples, there was a slight increase in PPO activity, followed by a decline. A similar trend of increase, followed by decline in activity was demonstrated following an additional 7 days at 18°C (data not shown). In apples stored under varying high CO, conditions, changes in 200 X g fraction PPO activity occurred earlier than in apples stored under normal CA conditions. After 28 weeks of storage, apples stored under both conditions, either with or without holding in air, had similar 200 x g fraction PPO activity (300-400 units/g).
During both storage years and under both conditions of the present study, there was a general decrease in PPO activity in the 4,000 x g and 100,000 x g fractions, and an increase in the soluble and 200 X g associated activity. Harel ef al. (1966) found similar results for ‘Grand Alexander’ apples which had been stored for up to 3 months in air at 2°C. In another study, Harel ef al. (1964) found that PPO activity in the mitochondria1 fraction of ‘Grand Alexander’ apples
SUBCELLULAR ACTIVITY PPO 191
1 normalCA * high CO2 varying
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30
Time (weeks)
FIG. 3 . PPO ACTIVITY IN THE SOLUBLE FRACTION OF STORED 'DELICIOUS' APPLES
had decreased by 93% after 9 months of cold storage. Chloroplast fraction activity was also reported to decrease with storage, while soluble PPO activity increased slightly. There have been no previous studies of controlled atmosphere storage effects on PPO activity in various subcellular fractions.
In both years there was a general reduction in total PPO activity with time under both storage conditions (Fig. 5 ) . Total activity was calculated by summing activities in 4,000 x g, 100,OOO x g, soluble and 200 X g fractions. Total PPO activity in normal CA stored apples appeared to decrease to 14 weeks, then
f & 800- . .- 5 600-
E & 800- . .- 5 600-
e c 2 0 0 1 I normalCA
* high CO2 varying
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30
Time (weeks)
FIG. 4. PPO ACTIVITY IN THE 200 X G FRACTION OF STORED 'DELICIOUS' APPLES
192 D. M . BARRE’IT, C. Y . LEE and F. W. LIU
FIG. 5 . TOTAL PFQ ACTIVITY IN STORED ‘DELICIOUS’ APPLES
increase slightly to 28 weeks. Total activity in varying high CO, stored apples decreased initially, and remained relatively constant until 11.5 weeks, after which there was a decline in activity until the end of the storage period. Apples held an additional 7 days at 18°C (data not shown) had exhibited a sharp reduction in total activity by 7 weeks of storage, and activity remained constant at ap- proximately 2300 units/g for the remainder of the storage period.
Most investigators have noted a decline in total PPO activity in stored apples (Harel et al. 1966; Vamos-Vigyazo et al. 1976, 1985a, 1985b; Prabha and Patwardhan 1986; Coseteng and Lee 1987). This decline may be explained in terms of reduced PPO synthesis with tissue age (Smock and Neubert 1950; Hyodo and Uritani 1966); a relatively greater rate of degradation than of synthesis (Mayer 1987; Burton 1988); substrate binding may result in conforrnational changes (Frylinck et al. 1987); PPO may be loosened from protective membranes (Tolbert 1973; Lieberei and Biehl 1978; Meyer and Biehl 1981); activators such as fatty acids or organic acids may decline in concentration (Hutcheson and Buchanan 1980); or phenolic substrate synthesis may decrease and result in a seeming reduction in PPO activity (Harel et al. 1966).
The brief increases in 4,000 X g fraction PPO activity seen in both years after 14 weeks of storage under normal CA conditions and after 7 weeks of storage under high CO, conditions may be explained in the light of changes in plastid permeability. Studies involving cell fractionation, histochemistry , im- munology and chemical or genetic modification of the plastid have fairly con- clusively located polyphenol oxidase in the lamellae and membrane-bound inclusions of the chloroplasts, and specifically on the thylakoid membrane. Chlo- rophyll disappearance, and chloroplast breakdown, are among the earliest symp-
SUBCELLULAR ACTIVITY PPO I93
toms of senescence (Butler and Simon 1971). The granal lamellae degenerates, and a simple membrane system forms in its place, with an increase in size and number of osmiophilic globules. It may be that the observed increases in PPO activity took place at a time when the chloroplast was degenerating and reforming into a simpler structure. PPO may have been released from the thylakoid mem- brane, and/or the increased permeability of the chloroplast itself may have al- lowed for greater substrate access to the enzyme. In either case, it would appear as though there were a temporary increase in PPO activity. After formation of the simpler membrane system, PPO activity may decline due to decreased perme- ability, enzyme depletion, inactivation or tighter binding.
Several investigators have reported that the mitochondria are more resistant to breakdown than other organelles and persist long after senescence has begun (Bain and Mercer 1964; Baker 1975). Although PPO activity decreased overall in this fraction, the decline took place at a later time than that observed in the case of the 4,000 X g fraction. The decline also appeared to occur at an earlier time in high CO, stored apples than in normal CA stored apples. If the PPO activity found in this fragment actually represented that bound to broken plastid fragments or in secondary association with mitochondria and other organelles, it might be that activity decreased due to the action of proteases, turnover, release from protective membranes or conformational changes.
Several studies have shown that a soluble PPO does exist in apples (Hare1 et al . 1964, 1965; Stelzig et al. 1972; Walker and Wilson 1975; Ben-Shalom et al . 1977) and investigators assume that this originates from the solubilization of the thylakoid-bound enzyme. In the present investigation, PPO activity in the soluble fraction displayed a generally increasing trend, but the extent of the increase did not equal that of the loss in activity which occurred in the 4,000 x g and 100,000 X g fractions. I t may be that solubilized PPO is more available to participate in enzymatic browning reactions, to be attacked by proteases or to be otherwise degraded. The temporary increase, then decrease in 200 X g PPO activity during storage may indicate association of soluble PPO with the cell wall, followed by loss due to utilization or degradation. The activity of both soluble and 200 x g fractions is relatively low, and the variations observed may be due to the extraction procedure. It is interesting to note that, while PPO activity declined dramatically in particulate fractions of apples stored an addi- tional 7 days at 18"C, the reverse was true for soluble and 200 X g fractions. This is further evidence to substantiate the theory that the membrane-bound PPO becomes solubilized.
In general, apples stored under high CO, conditions had slightly lower PPO activity in the particulate fractions and underwent declines in activity earlier than those stored under normal CA conditions. Carbon dioxide is known to inhibit certain enzymes, including PPO (Murr and Morris 1974; Siriphanich and Kader
194 D. M. BARRETI', C. Y. LEE and F. W. LIU
1985). In addition, Sears and Eisenburg (1961) showed that either increasing or decreasing CO, from its equilibrated tension led to molecular redistribution of lipids at the interface of microbial membranes. It was assumed this altered interfacial tension could result in membrane collapse and change in the activity of membrane-associated enzymes. Just such a phenomenon might occur in fruit stored under high CO, conditions. Although the theory of altered interfacial tension was based on studies with microorganisms, Frenkel and Patterson (1977) noted that a similar phenomenon might be occumng in Bartlett pears which were stored under 20% CO, conditions.
Electrophoresis of Apple Polyphenol Oxidase
Results indicated that isoenzyme patterns were similar in samples stored the same length of time, regardless of the storage condition. There were differences observed in samples, however, as storage progressed. Three PPO bands were observed in all apple 4,000 X g fractions of the stored samples, a dark band at R, 0.02, a medium to dark intensity band at R, 0.09, and a faint to medium intensity band at R, 0.23 (Fig. 6). The unstored apple 4,000 X g fraction was unusual in that only the first two PPO bands were observed. In addition, the R, 0.09 band observed in the stored samples decreased in intensity with storage time, while the lower molecular weight band at R, 0.23 became more intense. The standard mushroom PPO, like the 4,000 X g samples, had PPO bands at R, 0.02, 0.09 and 0.23, and, in addition, medium intensity bands were visible at R, 0.60 and 0.91.
When stained for protein, both unstored and stored 4,000 X g samples showed medium to dark intensity bands at R, 0.02, 0.09 and 0.23, which would cor- respond to the PPO bands mentioned above, dark intensity protein bands at R, 0.06 and 0.08, and faint or medium intensity bands at R, 0.30, 0.36 and 0.42. The only bands which were visible in the mushroom PPO standard after both PPO and protein stains were those at R, 0.02 and 0.23. The PPO bands at R, 0.09,0.60 and 0.91 were not visible after protein staining, which would indicate that the protein concentrations were quite low, while the specific activity was high. There were also 9 bands observed in the mushroom PPO standard protein stain, that were not visible after PPO staining.
Comparison of the R, values for the 3 PPO bands observed in the 4,000 x g samples to the molecular weight standard indicates that molecular weights of these three isoenzyme forms might be estimated at 208 kD, 165 kD and 11 1 kD. Very few reports of the electrophoretic pattern of apple PPO exist in the literature. Hare1 and Mayer (1968) used column chromatography to estimate the molecular weight of 3 fractions isolated from apple chloroplasts at 120-130 kD, 60-70 kD and 30-40 kD. The approximate molecular weights obtained in the present study are larger than these, and it may be that the PPO enzyme applied
SUBCELLULAR ACTIVITY PPO 195
.60
.91
1 2 3 4 5 6 7 (B)
.02
.06
.08
.G9
.23
.30
.36
.42
- - 150kD --I---- 66 kD -
1 2 3 4 5 6 7 0
FIG. 6. ELECTROPHORETIC PATTERN OF 'DELICIOUS'
(A) Gel stained with DOPA for PPO activity. (B) Gel stained with Coomassie Blue for protein. Samples: I . Unstored; 2. 7 weeks; 3. 11.5 weeks;
to the electrophoresis gel existed in very aggregated forms. Solubilization of the enzyme from lipid membrane was difficult and, while the zwitterionic de- tergent CHAPS was most successful, it may not have resulted in complete disaggregation.
Isoenzyme patterns for PPO isolated from different subcellular locations also differed. Although the same 3 PPO bands noted above were visible in the 100,OOO X g and 4,000 X g samples, there was a slightly different pattern observed in soluble and 200 X g samples. Soluble samples had 3 bands at Rf 0.02, 0.09 and 0.37 while 200 X g samples had 4 bands at R, 0.02, 0.09, 0.23 and 0.37. Comparison to the molecular weight standards would indicate that the new band
196 D. M. BARRETT, C. Y. LEE and F. W. LIU
at R, 0.37 corresponded to an approximate molecular weight of 47 kD. It may be that this lower molecular weight isoenzyme, which is only found in the soluble and 200 x g fractions, arises due to disaggregation of subunits. This finding may support the theory of PPO solubilization from the plastid and release to the cytoplasm, where the enzyme remains either soluble or associates with the cell wall. Protein bands observed after Coomassie Blue staining were faint; however, the isoenzyme pattern which was visible in all fractions appeared to be similar to that observed earlier in the 4,000 X g fraction.
In conclusion, changes in the subcellular location of apple PPO took place after 7-14 weeks of CA storage, and may be indicative of membrane breakdown and cell decompartmentalization. Plastid degradation may liberate the thylakoid- bound PPO and free it to react with phenolic substrates. Results indicate that PPO activity decreased in membrane (‘plastid’ and ‘mitochondrial’) fractions and increased in soluble and ‘cell wall’ fractions of stored apples. In apples stored under high CO, conditions these changes in subcellular location occurred earlier. Additional storage of apples in air at 18°C for 7 days following removal from CA storage, which simulated a typical marketing period, resulted in drastic losses in PPO activity. If this loss is indeed a measure of enzyme utilization or depletion and internal browning, these results may argue for the maintenance of CA conditions up to the point of consumption.
REFERENCES
BAIN, J . M. and MERCER, F. V. 1964. Organization resistance and the res- piration climacteric. Aust. J . Biol. Sci. 17, 78-85.
BAKER, J. E. 1975. Morphological changes during maturation and senescence. In Postharvest Physiology, Handling and Utilization of Tropical and Sub- tropical Fruits and Vegetables (E. B. Pantastico, ed.), Van Nostrand Reinhold/ AVI, New York.
BEN-SHALOM, N., KAHN, V., HAREL, E. andMAYER, A. M. 1977. Olive catechol oxidase4hanges during fruit development. J . Sci. Food Agric. 28,
BURTON, K. S. 1988. The effects of pre- and post-harvest development on mushroom tyrosinase. J. Hort. Sci. 63(2), 255-260.
BUTLER, R. D. and SIMON, E. W. 1971. Ultrastructural aspects of senescence in plants. In Advances in Gerontological Research (B. L. Strehler ed.), 3,
COSETENG, M. Y. and LEE, C. Y. 1987. Changes in apple polyphenol oxidase and polyphenol concentrations in relation to degree of browning. J . Food Sci.
COUEY, M. and OLSON, K. 1977. Commercihl use of a prestorage carbon
545-550.
73-129.
52(4), 985-989.
SUBCELLULAR ACTIVITY PPO 197
dioxide treatment to retain quality in Golden Delicious apples. In Proceedings of 2nd National CA Research Conference, Michigan State University Horti- cultural Report No. 28, 165-169.
DAVIS, B. J. 1964. Disc electrophoresis 11: Method and application to human serum proteins. Ann. N.Y. Acad. Sci. 121, 404-427.
FRENKEL, C. and PATTERSON, M. E. 1977. Metabolic effects of CO, in Bartlett pears. In Proceedings of the 2nd National CA Research Conference, Michigan State University Horticultural Report No. 28, 108-1 15.
FRYLINCK, L., DUBERY, I. A. and SCHABORT, J. C. 1987. Biochemical changes involved in stress response and ripening behaviour of gamma-irra- diated mango fruit. Phytochemistry 26, 68 1-686.
HAREL, E. and MAYER, A. M. 1968. Interconversion of sub-units of catechol oxidase from apple chloroplasts. Phytochemistry 7, 199-204.
HAREL, E., MAYER, A. M. and SHAIN, Y. 1964. Catechol oxidases from apples, their properties, subcellular location and inhibition. Physiol. Plant.
HAREL, E., MAYER, A. M. and SHAIN, Y. 1965. Purification and multiplicity of catechol oxidase from apple chloroplasts. Phytochemistry 4, 783-790.
HAREL, E., MAYER, A. M. and SHAIN, Y. 1966. Catechol oxidases, en- dogenous substrates and browning in developing apples. Sci. Food Agric. 17,
HUTCHESON, S. W. and BUCHANAN, B. B. 1980. Polyphenol oxidation by Vicia faba chloroplast membranes. Plant Physiol. 66, 1150-1 154.
HYODO, H. and URITANI, I . 1966. A study on increase in o-diphenol oxidase activity during incubation of sliced sweet potato tissue. Plant Cell Physiol. 7,
LAEMMLI, U. K. 1970. Cleavage of structural proteins during the assembly of the head of Bacteriophage T4. Nature (London) 227, 680-685.
LEE, C. Y. and SMITH, N. L. 1979. Blanching effect on polyphenol oxidase activity in table beets. J. Food Sci. 44, 82-83, 86.
LIEBEREI, R. and BIEHL, B. 1978. Activation of latent phenolase from spinach chloroplasts by ageing and by frost. Phytochemistry 17, 1427-1429.
LIU, F. W. and PAN, H. W. 1989. Storing ‘Delicious’ apples in high carbon dioxide atmospheres at above optimum temperatures. Presented at CA Con- ference, Wenatchee, WA, July 1989.
LIU, F. W. and SAMELSON, D. 1986. Rates of change in firmness, acidity, and ethylene production of ‘Mclntosh’ apples in simulated low-ethylene CA storage. J . Amer. SOC. Hort. Sci. 111(3), 404-408.
MATTUS, G. E. 1982. CO, treatment of Golden Delicious apples in Virginia as a supplement to CA storage. In CA Atmospheres for Storage and Transport of Perishable Agricultural Commodities (D. G. Richardson and M. Meheriuk eds.), Oregon State University, Timber Press, Portland.
17, 921-930.
389-392.
137- 144.
198 D. M. BARRETT. C. Y. LEE and F. W. LIU
MAYER, A. M. 1987. Polyphenol oxidases in plants-recent progress. Phy- tochemistry 26, 11-20.
MAYER, A. M., HAREL, E. and SHAIN, Y. 1964. 2,3-Naphthalenediol, a specific competitive inhibitor of phenolase. Phytochemistry 3, 44745 1.
MEYER, H. U. and BIEHL, B. 1981. Activation of latent phenolase during spinach leaf senescence. Phytochemistry 20, 955-959.
MURR, D. P., and MORRIS, L. L. 1974. Influence of 0, and CO, on o- diphenol oxidase activity in mushrooms. J. Amer. SOC. Hort. Sci. 99(2), 155- 158.
PONTING, J. D. and JOSLYN, M. A. 1948. Ascorbic acid oxidation and browning in apple tissue extracts. Arch. Biochem. 19, 47-63.
PORRITT, S. W. and MEHERIUK, M. M. 1977. Effect of CO, treatment on storage behavior of apples and pears. In Proceedings of 2nd National CA Research Conference, Michigan State University Horticultural Report No. 28, 170-1 74.
PRABHA, T. N. and PATWARDHAN, M. V. 1986. Polyphenol oxidase (PPO) and peroxidase (POD) enzyme activities and their isoenzyme patterns in rip- ening fruits. Acta Aliment. 15, 199-207.
SEARS, D. F. and EISENBERG, R. M. 1961. A model representing a phys- iological role of CO, at the cell membrane. J. Gen. Physiol. 44, 869-887.
SIRIPHANICH, J. and KADER, A. A. 1985. Effects of CO, on total phenolics, phenylalanine ammonia lyase, and polyphenol oxidase in lettuce tissue. J. Amer. SOC. Hort. Sci. 110(2), 249-253.
SMOCK, R. M. and NEUBERT, A. M. 1950. Apples and Apple Products, Interscience Publishers, New York.
STELZIG, D. A., AKHTAR, S. and RIBEIRO, S. 1972. Catechol oxidase of Red Delicious apple peel. Phytochemistry 11, 535-539.
TOLBERT, N. E. 1973. Activation of polyphenol oxidase of chloroplasts. Plant Physiol. 51, 234-244.
VAMOS-VIGYAZO, L., GAJZAGO, I., NADUDVARI-MARKUS, V., and MIHALYI, K. 1976. Studies into the enzymic browning and the polyphenol- polyphenol oxidase complex of apple cultivars. Confructa 21( 1/2), 24-35.
KUS, V., HAMORI-SZABO, J. and SASS, P. 1985a. Changes in the poly- phenol-polypheno1 oxidase complex of apples during ripening and storage. Part I: Variations related to cultivar, year and date of picking. Chem. Mik- robiol. Technol. Lebensm. 9, 37-47.
KUS, V., HAMORI-SZABO, J. and SASS, P. 1985b. Changes in the poly- phenol-polyphenol oxidase complex of apples during ripening and storage. Part 11: Effects of storage conditions. Chem Mikrobiol. Technol. Lebensm.
VAMOS-VIGYAZO, L., SCHUSTER-GAJZAGO, I., NADUDVARI-MAR-
VAMOS-VIGYAZO, L., SCHUSTER-GAJZAGO, I., NADUDVARI-MAR-
9. 97-104.
SUBCELLULAR ACTIVITY PPO 199
WALKER, J . R . L. and WILSON, E. L. 1975. Studies on the enzymic browning of apples. Inhibition of apple o-diphenol oxidase by phenolic acids. J . Sci. Food Agric. 26, 1825-1831.
WISSEMANN, K . W. and MONTGOMERY, M. W. 1985. Purification of d’Anjou pear (Pyrus cornrnunis L.) polyphenol oxidase. Plant Physiol. 78, 256-262.






























![The composition function k(x) = (f o g)(x) = f (g(x)) g f g f R->[0,+oo)->[-1, 1]](https://static.documents.pub/doc/80x56/56649f1f5503460f94c36d59/the-composition-function-kx-f-o-gx-f-gx-g-f-g-f-r-0oo-1.jpg)
