ANTARCTIC ECOSYSTEM Marfa Vernet and Raymond C. Smith
ABSTRACT
Ultraviolet radiation (UVR) affects biotic and abiot ic factors in marine ecosystems. Effects on organisms are mostly deleterious due co dam
age to DNA and cellular proteins that are involved in biochemical processes and which ultimately affect growth and reproduction. Differential sensitivity among microalgal species to UVR has been shown to shift community composition. As a result of this shift. the total primary production for the com munity may be maintained at pre-UVR levels . Similar impacts and mechanisms are expected in Antarctic waters. The overall effect of UVR on the ecosystem needs ro include relevant feedback mechanisms which can diminish, and sometimes reverse, deleterious effeers on population growth. For example. it has been speculated that UVR can increase iron-limited phytoplankton populations by photoinduced reduction of Fe l - to Fe' -. a more soluble form of iron and readily avai lable for algal and bacterial uptake. An equally positive feedback can be attributed to diminished g razi ng by zooplankton . Thus. energy flow among [he troph ic levels can decrease as a result of damage to a certain trophic level. but overall biomass and ecosystem production might remain relatively unchanged .
Similar positive and negative feedbacks associated wit h UVR are related to the dissolved organic matter (DOM) pool, known to be recycled by baererial activity. Although it cou ld be expected t hat bacterial production in Antarctic surface waters would decrease when exposed to UVR, this effect can be counteracted by increased substrate nutrient availabi lity. Photolysis of high-molecular weight molecules by UVR produces
248 Tile E((ecls of Ozone Opp/etion on AqUcl tic ECOS~/5 tellls
higher availab ili ty of low-molec ul ar weight mo lec ul es readi ly ta ken up by bacrer ia . This step might be of greate r impOfWnCt
in high latit ude (:'(osys(:'ms where low bacterial producrion has bten attributed to low su bstrate ava ilability.
Similarly, inc reased nutrients for bacte ri a l act ivity originate fro m phocolys is of h igh-molecular weight moitcul es which are known to release NH.j • and am ino ac ids under UVR. T he DaM pool might also inc rease th rough phytoplankton excrerio n of organic maner, a process known to occur under a lgal stress. On rhe ot her hand, a decrease in DaM by diffus io n from zooplankton fecal pell ers is ex peered in surface waters due [() decreased g razing.
In summary, we argue t hat rhe understand ing of the effecr of U VR on Anta rcric ecosystems is more t han the sum of the effect of radiation on ind ividua l spec ies, g iven t hat alteration of inte rspec ific interactions can exace rbate , dim ini sh and somet imes reverse known physiological damage . This, p illS complex and nonlinea r feedback mechan isms associated wit h UV R effects m ake p rediction at the ecosystem leve l uncertai n .
INTRODUCTION A rt~cent characte ristic phenomenon of
the Antarctic ecosystem is the well-k now n springti me decrease in stratospheri c ozone, known as the ozone hole. It is confined to the po la r vo rtex over the Antarc ti c continent , from September to December of each yea r. H owever, once the wintf: r/spring vortex breaks dow n, its effc:ns reach mid lat itudes, mostly during the mont h of Decem ber, I alt hough it has a lso been detected in s ub- an rarctic e nvi ronme n rs duri ng the sp ring . .! There has been sig nifi can t ann ual and intera nnua l var iab ility in Antarcti c ozone, and , conseq uen rl y, in changes in ozone-re la ted incident u lt rav ioler radiation (UVR). During the last two decades major internat ional efforts have focused o n the physics and chemi stry of the Eart h's atmosphere w ith emphasis on understanding processes that control the ozone layer, while
studies on the effects of UV on th t b iosphe re, in parti cular a t the co m munit}1 and ecosyst tm level, ha ve been re la ti ve ly lim ited. '
ln te res t in U V effects on aquatic ecosystem s is in creasing because ozone deplet ion is not restr icted to th e area over Anta rctica and signi fi can t reductions have been reported in the Nort hern H emisphere : I
. ';
Hemi spherical trends a rt superimposed on high in terannual variab ility, as poi nted our by Michaels Ct a i,' w here low ozone during 1992 can be assoc iated with a drop in sunspots, a strong EI Nino event and the erupti on of Mount Pinatubo, a ll of which can potentia ll y decrease ozone in the Stratosphe re . Othe r popul a ted areas, such ns Sout h Ameri ca, Australia, New Zea land and Sout h Africa a re affected, in panicular at the t ime of [he vortex disappearance, probably as an effter of dilution.' ··'
It has been est im ated tha t aquati c ecosystems fix bttween 30 and 50 Gt of carbon pe r yea r, whi ch is roug hl y half the rota l g lo ba l fixation of ca rbon. H•
ltl
Consequently, the threat of increased UV R on surface layers of the ocean on mnrine product iv ity is of considerable conce rn . Estimates fo r t he Sourhern Ocea n range frolll 1-5 Gt C y- '." For the Southern O cean, ice a lgae are esti mateu to contribute lip to
30 % of t he total pr im ary produCtion . I ..?
Traditionally, pred ic tion of U V efftcts on ecosystems have assumed a linear addition of U V effecrs on different level s of tht food chai n whe re t he f in a l effec r on h ig i1<:r troph ic Itve l p redators, s lIch as penguins. wha les and sea ls, have been in fe rred from t he cumul ative effeCt o n p ri ma ry produce rs and grazers. I ., I n other words, tht- roral effect of UV at a givf'n troph ic levc:l has bee n assumed co be the combinat ion of U V effeCts on the p revious trophi c level i1(.1ded to t he di rect effeCt of U V on t he leve l icself. For examp le, in itia l studies on UV effeCts o n marine nlga l communities rt'po rted decreased [Ota l pr imary producriv-ity and shi fts between species rownrds It-ss UV -B -sensitive spec ies as well as l:I drop in total species diversity, assum ing consmnr
Effects of Ultraviolet Radiation on the Pelagic Antarctic Ecosystem
grazing.14"17 In contrast, recent trophic-level assessments suggest that differentialUV sensitivity between algae and herbivores may contribute to an increase in algae by exerting a stronger UV influenceon the grazers. 18-ly An analogous influenceon zooplankton, thus reducing zooplankton grazing, could counteract UV photo-inhibition on phytoplankton growth. Inaddition to biological factors, UVR affectsabiotic processes which affect directly orindirectly the food web. These factors areeither chemical (e.g. nutrients) or relatedto the dissolved organic matter (DOM)pool which is intrinsically related to themicrobial loop.20 Such an alteration of theecosystem functioning would result in a decrease of transfer of energy through thefood web.21
In this chapter we summarize what isknown of the UVR effects on different levels of the Antarctic food web, with emphasis on the relationships between trophicspecies, and what is known of the UV effects on abiotic processes affecting the foodweb. Several recent reviews on UVR effectson aquatic and Antarctic ecosystem13-22have given excellent summary of the UVphotobiology and that information will notbe rephrased here. We present evidence tosuggest that research required for understanding UV effects on Antarctic ecosystems will necessitate ecosystem studies inaddition to detailed determination of UVRon specific processes related to any giventrophic level.
UV RADIATION IN THESOUTHERN OCEAN
Estimation of quantitative effects ofultraviolet radiation (UVR) on biologicalsystems requires knowledge of the incidentspectral irradiance and a biological weighting function (BWF), which provides thewavelength-dependency of biological action. Because BWFs are heavily weightedin the UV-B region of the spectrum, highspectral resolution is required for accurateestimation of effective biological doses.Smith et al23 have developed a high spec
tral resolution (1 nm) air and in-waterspectroradiometer and Booth et al24 havedeveloped the U.S. National ScienceFoundation UV Network which provideshigh resolution data at three locations inthe Antarctic continent. Alternatively, narrow band instruments (e.g. Bio-SphericalInstrument PUV series) can, in conjunction with an adequate full spectral model,be used to estimate incident spectral irradiance with adequate resolution. BWFs,specific to the target unit, have been developed. For Antarctica, stepwise functionsfor the BWF for photosynthesis have beendeveloped by Helbling et al,25 Lubin etal,26 Smith et al23 and Boucher et al27which have yielded results similar to themore detailed determination of Cullen etal.28 Other BWFs have been developed intemperate areas for plant chloroplasts29 andDNA.3(1 There is a paucity of BWFs forother processes, for other levels of the foodchain, not only for Antarctica but everywhere. This is a serious constraint formodeling and predictive purposes.
Actinometry (e.g. refs. 31, 32) has notbeen used extensively in Antarctic studies.On the other hand, a biological dosimeter,based on the response of an organism toUVR, has been used. This method providesa relative unit to assess potential effects ofUV exposure on a specific organism or target molecule. Once the response of theorganism to UV is evaluated under standard conditions, i.e. by exposure to natural UV radiation, we can say the organismhas been calibrated. A relative estimate ofpotential UV damage can then be estimated. The potential benefit of the biological dosimeter resides in being a relativelymore easy and inexpensive method, onceit has been carefully evaluated. The maindisadvantage is the exacting dosimetry required for quantitative calibration. It canalso be used to compare biological effectson very diverse environments with or without very different UV climatology. Although a biological dosimeter was carefullyevaluated for an Antarctic coastal site it hasnot been used extensively use in the
250 The £{(ecls or Ozone Depletion on Aquatic Ecosystems
regio n. H Both rh e acr inom erry and rhe biological dosimeter give broad band estimates of UV R unl ess rhe incident radiacion is differentia ll y screened, usually with filrers. ,H
CLIMATOLOGY OF UV RADIATION U leraviolet radi ation (U VR) levels are
mostly controlled by atmospheri c ozone, cloud cover, and solar zenith angle with ozone concentration being relatively specific to the UV-B reg ion. )·' Natural variability in these env ironmental variables g ive ri se to a very high natural variabi lity in UVR , with ozone pri marily affcccing rhe relative ratios of UV-B to UVR, photosynthetic available radi at ion (PAR ), or rota I irrad ianee. The dynamic nature of rhe polar vortex containing rhe ozone hole has given rise to large changes in these UV-B ratios on rim e sc al es of severa l days or less (Fig . 15. 1). The polar vortex, and corresponding ly , the ozone hole, is often elongated in shape, gi ving rise to an uneven di stribution of UV-B at locations within the Antarctic continent.35 The natural-short te rm variability (hours to days) due to
changes in cloud cover and solar zenith ang le compounds the diffi cu lty in assessing the innuence of increased UV-B levels on natu ral sys tems.B,Y; The resultant effect is that natural variability (cloudi ness)
Fig. 15, J. Daily maximum UV·A irradiances (3 60· 400 nm) from J 5 Decem· ber / 989 10 7 Februa ry 199.3 ~1 r McMurdo Sta tion (77.5/ 'S, /66.40 ' £) shown (1S a fun ction of da}'s before and after solstice. Redrawn from Booth et ai, 1994.
~
'1' E
== ~
0 1::. CI> u c: co 'is CO ~
.!:::
3
2
can cou nteract UVR increases. Further, rccent work (Gautier et ai, Un iversity or Cal iforni a Sanra Barbara, U .S., personal com muni ca ti on) suggests that t he co mbined influence of cloud cover and surface reflectance influences these UV -8 ratios. As not much is known w ith respect to the effen of th is variability on organi sms and processes, it is roo soon to predict t he effect of thi s variabil ity either to enhance or decrease U V effects o n Antarcti c ecosystems.
TRANSMISSION OF UV IN SURFACE WATERS AND ICE
Transmi ss ion of UVR wi thin the water col umn is a key element in assess ing UV effeCts in marine sys tems. Light transmission is affected by wa ter itse lf, as well as particulate and dissolved organic matter (POM and DOM, respect ively) within the water column , Water is known to be a relatively strong UV absorber37-39 and spectral attenuati on coefficients have been published for cleat natu ral wate rs. 3M H owever, in natura l waters, particulate and di ssolved o rgani c matte r strong ly absorb U VR and t hese in-water cons t ituents are hig hl y variable. In blue, more transparent oligotrophic waters, biolog icall y sign ifica nt UV doses (a n penetrate several tens of met ers. J n con trast, mort productive coastal waters,
o 125 Days before or after solstice
Effects of Ultraviolet Radiation on the Pelagic Antarctic Ecosystem
with higher particle concentration (e.g.>3 mg chlorophyll a m-3) can have attenuation coefficients nearly an order of magnitude higher, limiting significant penetration depths to the order of meters.40 DOMshows an even stronger attenuation in theUVR'10,41 and can effectively limit significant penetration depths to a meter or less.For example, Kramer42 estimated that thecombination of high POM and DOM inDutch coastal waters would limit UVRtransmission in the water column to suchan extent that no UV effects or planktonicorganisms were expected. High POM absorption in Antarctic waters43 and probablyin ice-edge blooms,44 would limit UVtransmission in late spring and summerdue to high production, but not duringearly spring (e.g. October) where chlorophyll (chl) a levels are usually lower than0.5 mg m-3.45 The paucity of absorprionestimates for POM, and in particular forDOM, make it difficult to speculate ontheir effect in Antarctic waters, althoughsimilar levels of DOM as in other parts ofthe world would support the hypothesis ofimportant UVR absorption by DOM(Fig. 15.2).46 Estimated UV effects atdepths of about 20 m in the vicinity ofPalmer might be due in part to thecontribution of DOM absorption.'3-4^
The role of DOC in light attenuationis intimately related to other environmental changes. For example, in boreal lakes,the decreased amount of DOC, caused byan increase in average temperature and
acidification in the last 20 years, was related to increased UVR in the water column.48 In the case of Antarctic waters, acomplex mix of competing feedback mechanisms make estimating changes in UVR,due to environmental change, speculative.
There are relatively few direct observations on the optical properties of Antarctic ice and snow. These observationssuggest that UV transmission in the ice ismaximum in October due to relativelyhigh transparency in spring. Based on theseobservations, it is expected that ice algae,associated with bottom communities in iceflows, potentially can be exposed to relatively high levels of UV-B. These UV-Blevels have increased by as much as an order of magnitude under the ozone hole.49
THE FOOD WEB
Phytoplankton
PhotosynthesisDeleterious effect of UV-B on photo
synthesis has been studied both in culturesand in the field, in particular for Antarctic phytoplankton. The reader is referredto reviews done in the last few years thatcover this subject extensively (e.g. refs. 22,36, 50, 51 and references therein). Overall, UV-B inhibits primary production by30-509? of shielded samples^2 with a strongdepth gradient from surface to about20-50 m.23-33-33 All these experiments arebased on 6-24 h incubations, either in situ
, - , -'-
Of in incuharors t'xposl·d ro sllnli,;.:hr. On rhe ;I\'l"r~l,!.!t' for rhe wafer column. primary producriCln <.I,.-creilses by (i- I 2';; ~'. ~! durin,!! spri nt!rime IlZUIlt" deplt'r ioll O\'f: r Anrarnir wafer resulring in ;t 2r;; fed unioll in rht' rt"arl y pri mary protlu([ ion {'Sf i nlil(:"S for rill:
l1lilrt.:inal ict' z() n l·. ~'H t' lbling e( ai " has(:d on dint-fene assumprions and mt'rhodol()~~y.
l'srimilrt' rhe: decrease in primary produc{ion ro he n,I,) ';; for elu' en ri re iu:-frt't' wafers sourh of rht' Polilr From. A UV inhibition fUllcr ion for pho{()synrhesis has been described by Cu lltn and Nealt. 'i'i Tht biol ogica l weighring funcrion for Anran.:ric phytoplankton. I1tct'ssaQI to sca le UV R ro biological efTc:niv(: irratiianct', has been d(:,,[t'rm int'd for Il aru ral popu larions by Lubin e( ai, '" Helbling « ai," Smi(h or al,2 ; Buucher tr .. 12- ;Ind Nl'ale ef alY'
Nutrient uptake Very lin ll' is known or th t' c:ffen of
UVR on nutrient upwkt' in Anrarnic phyroplankron. Srudit's on tempe ra te species sugt!:t's r (hat nirrogenast' , t he t' ll zy ml' rt'lart'd co nirrogtn assimi lation in phyropiankwil. is acrivared by PAR '- anJ i l1-acrivnfed by UV -B rac1iarion. '~ In cont raSt, ammonium IIp[;d~t' seems less afft'<.:[(.·d. ~".f'O
Overall. amino acid (Ollct'lH rar iol1 in rhe ce ll dtt'reastd undtr UV-B. (01 Tht effect is also Idr on t'l1zymc:s rdartd (0 amino acid metaholism . UVR diminishes synrht'sis ilnd il1rra( .. ~ lI lIhir iln:umuiarion of alanint· ilnd va li nt"'! whi lt synrlll·sis ilnd acculllulat ion
of t!luramie ac id incr('· 'ISl' dul' {(I inhibition or g lu rama((:' synr hast ' .... or ,l.duram'Hl' dehyd rogen;lst: , t,l! Tht'se rt:slIl tS are si m i lar ro
metabolic changes obsl'rvt'd in phrroplankron under nirroge:n streSS. suggesring rhar UV-B suppresses nirrogt'n assimilation inro ce:lls, t>; Decre;lsl,d N H I' uprakt' by PadM'a
spp, under UV-B and high intensity UV-A was interprered as rt'dllced supply of ATP and NA DPH from direre offeres of UV-B on rht: phorosynrheri <.: apparacus and pigl11t'nt b le;tching, 1>4I Simi lar dTe(ts of UVR nn Anrarctic spec it's will havt ro bt assllmtd until t'xptrimtnrs are ca rried Ollt ror Anrarc..' ric, or at leasr, polar p hyroplankron,
Exudation Tht' amount of excrilcc: l lular c lrhu n
produced by phyroplankron has lx'ell ;1 COIi
tro\'ersial subjeL't for St:\,eral decades ,""
Excrerion of carbon hy pilOtosy ntht'ril' 01"
ganisms is a widespread process ;,lSSOl ial l,d with photosyn thtsi s,h~ On the: iI\'l' ra,l.!l·. phytoplankton txcrl'te:s 5-15'A' of rhl' C;II'
bon in corporilted in parri c ulatt" maner. bot h in monospecific n ilrure:s and in nafU
rid populatiol1s(,'j ,t>:-! and tht' amOlllH e x
crett'd is a conStant proportion oi' phtllosynthetic: ratts, Several st lldi t's have poimnl OLit t hin a large: proportion of p llOwsyn chetic ca rbon got'S throug h a DOC phas("'" for at It'<lst shurr pt'riods of t ime:. - II Undl'!' chest' conditions. bt'tween 20-()OIfi oi' plwrosynthiltt' must go inru rht' DOC pool l t l
explain the DOC ( ha n gc:s obser\' ed . I'
mainly dur ing spring bloom t'\'l'IltS in (('Ill
pt'riltl' waters, Additional organi( ('ar hon t'xCfetion in phytoplankton seem s associated with physiological imbalance due to ('V("ll['"
such as nitrogen limitation : I ,-; in particular lIndtr high-light condit ions,-' III rill'
ritld. rhe transfer of ct'lls to higher irradi · anct" might produ<.:e excess phorosynthatt'. ('-'<'.'i Nutrient limitation is ohsl'fn·d
during !att' grow rh sragt's in batr h nLlturc:s · I or at tht' e:nd or rht spring hlotlill .
High DOC c() llLenrrations havt' al so IWl' 1l
obse rvt'd after a Pb{((:IIIJ.flis sp. b loom, - ~, "
This t'xce:ss carhon t'xcre:red mi,ldn 11l' ;1 ... •
sociatni wirh increast'd in trace:llular Llrhll h)'<.iratt. as in diatoms -1.-- bur nor ohsl'f\'l'I1
in dinotlagellat(:s .--' Vt'r)' lit tle is known of ("xud,lrioll hy
Antarnic phyrophinkwn illld [he: l Oll'ol'· tillenr implicilrion for (il(: DOC pool. \{ l'rent resulrs in the Arni c sllg,cesr ,I I. :r,~l '
,111101l1l( (If exrrace llular carb on Oh,'ol'r\'nl see mt'J to bt: rt' latc:d to phywplallkr tll l
composition (i,e. (ells w hi( h produ <.' l' 11111 -
c: ilal:e ror colonial forn"larion) .lntl (0 ;J It':-:-..t' r e:xr;IH {(l in sitU n itra te lim i(;lrion. · ... In Arnic Wa tt'r. Cbdt'/fIl't' I'IIS .fll(itl/i.l' a ll oc;nnl 4 0 '/f of total ca rbon incorporatt'tI ;IS ("st[.l ctllll iar under (ondit ions of low si lic ic acid (<<),2 PNf) and measurable nirril(t' concentrations (0.5 -2.5 JlNO, Similar exrr;u.:l'lIuI:1r
carbon produccion was found in iI mixru rlo
Eifects of Ultrilvio/et Radiation on t/u" Pel.lgic Antarctic Ecos}lsrem 1.03
of C. l{J(ia/il and P. p'JlldJdii ar rh t Po la r Front and rhe ma rgina l ice zo nt wir h h ig her IlUrri e lH conct nrrarion s (5-1 0 ~tM n it rate ).
These resu lrs s tlt:t::esr ch.u species composicion and rheir p hys iolo,!-:ica l sta te may largtl y control extracel lular (;.\rbon producti on in t he field. -'J Alt houg h low n itratt' is known to inlTt'ast' exudation. - ' thi s t'ffl'n
is not l'x pel'Cl'd in Ancarnic open watl'rs; however. t h is tffecr mig h t bt observed during o r after massive co;\sral b looms. 1'1.).;(1
In spite of the obviolls imporranct (If ph),top lank wn ex udacion o n tht carbon C},c!t and as subst rate for the microbial loop, no s tud it'S have bet'n ca rrit'd out on tht efftn of UV-B on txud;\ t ion, fo r tithtr tt'mpt'ratt' or pola r phytop lan k ton. In gtlleral, ex ud at ion illlTeast's wht n a lgat a rt' stressed and it can be specul attd chat UV -B stress wou ld act in a s imi lar way.
Respiration Changes in OHC in rCO.,! observed in
the Bt lli nghaust n Sea in the s pring of 19YO combined wit h changes in ce ll abulldan ce in th t' colon ial prymnes iop h y te Pbm!O(}'llis sp. sugges t t hat under im:reased U V-B radia tion , as mtasu red undt' r dtcrt aseJ ozo nt (onctnt rat ion , t htrt is an increast in t ht' ratio of [0[;11 communi ty n:sp irat ion (0 p hotosynthes is.xl H eterot rophic respiration increases were ;.ltuibuted co incrtased bantria l substrate du t' to (til lysis.
Growth Tht' effect of U V- B on marint ph y
wpla nk w n grow th has btt n shown to be spt'c its-sptci fi c. For stveral cu lwres of tel11-ptratt sptci es, spec ific grow th rate was affecced negativ(·ly by U V_B. ~.!-N I In the dia
com P b(((,(" /(/(/),/IIIII Iri(II/·IJIIIIIIII. no d tuease in UV R se ns it iv it y was observed wit h t illl t.~ .! Simi lar results were observed on .) 0
txper imtnrs on Anra rcric ph ytop la nkton dominated by C',rt'lbrrlJl (ri lll,b)/I1'" whe re .;..:rowrh rates d tcreasl'J by IOO'J on (t· ll s exposed to UV-A + UV- B + PAR and by SO'/( whe n ex posnl to UV- A + P,\R. as co mpared to conrrol s t'xpost'd to P ,\ R
onl y .'''~ O n rht other hanJ. act ivt grow rh of ( oasra l sptc ies was obst rvtJ for 12 days a t Pa lm e r Station whert d iarom ( u lrurt's wtre kept at in sit u sol'ar radi.niol1. sh No ditfere nc<:' was found also bttwee n t reat
ments (UV R + PAR "s. PAR on ly) Ii" rh" colon i.li prym nes iophy rt P/;at IJIJ.fliJ sp .. althoutih tht'st' cu lwn:s did nor g row. T h is lack of ellel'( was observed in spitt' of tht' we ll -doc umented inhibirion of pho(Osy nr h ts i s .!\·.!('-.!~ ti.)f Anrarcric ph}lroplankron in
ex pt' ri me nrs frol11 1-14 h and po inrs to
wards di lh:rtnr conrro ls of p horosynr htsis
and growth and bttwetll sho rt - vs. lon,grefm t:lltns of U V-B . Ir has been nott'J fi.lf some timt rh at cau tion must be used when inft rfing longe r tt' rm eco logical COI1-
St'lluencts from short-term obstrva rions.N-Mixing of (t lls in th t uppt r watt'r col
umn . in partic ular within rhe mixt d laye r, affens rht' avtragt irraJiance in whi ch a (t.1I is txposecl d uri ng rh t day. \"i.NS,""·) Sev
t' ral s rud its havt spt'nlia[td aboU( rhe poss ib lt role of a lleviat ion fro m U VR in Anta rctic wate rs if ce ll s are mi xed detptr in rhe wate r colllmn . "i (j ·~tI ") 1 Expt rimt'nts where
U VR inre nsity was ma nipul ated to rtstm ble m ix ing in the u pper water co lu mn showed increased product ion in cloud y days while rhe effect was opposirt on sunn y Jays. ~ 1 Phyroplankron dominattJ by t ht d iarom T ba/fl11;Olir(1 grarititl showed less phoroin h ibit ion when tx poseJ to va riable radiari on:''! supportin,!-: t ht' hypothtsis char mixing mi g ht provide UV-B protection. ,I,
Cell size Coastal wattrs have. on th t' avt' ragt_ a
hi ,!-: h t r proportion of htrge r ct' ll s chan ope n warers.''' For example. more than :-lO 'A of the nearshort p hyr(lp lan kcon biomass was assoc iated w ith ct ll s > 10 pm in T er re Ade li t' durin~ sum mer wh ilt 70 km offshort'. ct' li s > I () 11 m rep rt'st llted o n ly .)WJ of t ht, wra l b iomass and '51) r;1 of cht, (e ll s were be rwt'tn 1 _ 1()~lm . ' JI \'(Iithin coas ra l
wate rs. hit:h Chi (/ acc u m ul ar io n s ( i .e . b looms ) a rl- do min.Hed b~' lar~e n -lls (e.!! . > ~o pm ) while low Chi (f ( oncencrar iolls afe dominared by sm all er u : II S .. -l11.'1\ A differ
obst" rvnl for diatom cultll rt"s.'Jh show hi~ h t' r
d;'lma~e on smalltr ct' lI s, and Wt m i!-dH spt'udatt' that oceani c ph ytoplankton rnay havt a hightr st' ns itiv ity to l1V-B . In addition , U VR int:re<1sts ce ll s ize;.! assm:ia ttd with a concomltanr reducrio ll in specific growth ra tts.
Species composition In itia l t'xptrimtnts with ttmperatt
phytoplankton, showing difft' renri .1i se ns it ivi ty to UV-B by d iffc.: rt'nr spt:cies ,l - sugges t a change in spec ies composition in long-ttrm U V-B exposure with more UVtoleram spec ies ultimatel y dominating. I ('
As mentioned above. there is a wide range of interspeci fi c UV- B stns itivity on growth and surv iva l, with smaller ce ll s being mo re se nsitivt'. due to a hig htr surface to volu me ratio as a resul t of ce ll sizt' and ce ll shapt' Y6 In addi t ion to sizt, an increased UV-8 stns itivity in tlabe ll an:s, as compared with diatoms, was observtd in natural populat ions of AntarCtic phyroplankron . 'i •. 1)
Thi s di fft'rence ( an be att ri bmed in parr to si ze (fla g ell a tes are on the average smaller than Anrarct ic diatom s) and to increased UV-absorbing propercit's of diatomsl)1 rt'lat eJ to the presenct' o f mycospo rint-like amino acids whi ch are bt' li evt'd to reduce dt' lt'te ri om effects by UV-B on g row th :'" The predicted sh ift from less to more resistant spec its (e.g. from fbgellarts ro diawms) was obsNvt'd in a 2-wt"t"k experimtnt of nar ura l Anrarctic populations t"xpost"d ro amnienr U VR, although si milar ChI tI and parricuhue carbon acc umularion were ohserved undtr UVR and UVR + PAR ." Unde r lIV R the amount of U V ahsorhing compound s (t.t!. m y<:ospori ne-l i ke a 111 i no .. It'ids) increased as well. As a result of rhi s shift in speci es composition. a decreased sensiriv iry of photosynthe s is wa s obse rv ed in rhe ph yroplankton t"x posed to U VR . The hi g her res isranct" by d iarnms, as cornpared wit h fiage ll a rts ( in parricu lar the colonia l prymnc:siophyte. P /)rft'{JI),JliJ jJOIlc/Jf!lii, ref. R I ), see ms to be rehHed [() a lower efft"([ on phorosynt hes is as wtl l as nitrat t' uprake. $·)
Few sw d its a re a\'a ilablt' on d"(tc£ s Ill' UV R at lon,!-!t"r rime sca les. rvldvl inn t't ill' " docllmt"med no changts in diatom spl' r il'., co mpos ition in laminared sed im enrs ill Anrarctic anox ic fio rds for t ht las( 20 year,. coincidin,g w it h t he dtcre;lst' of OZOIll' . Howeve r, as nored by Bothwell and coworkers ls the limi ted data provided hy McMinn et <1 1'/1' do no r subsrami <1rt" rill' ir implied lack or a UV-B eHtcr.
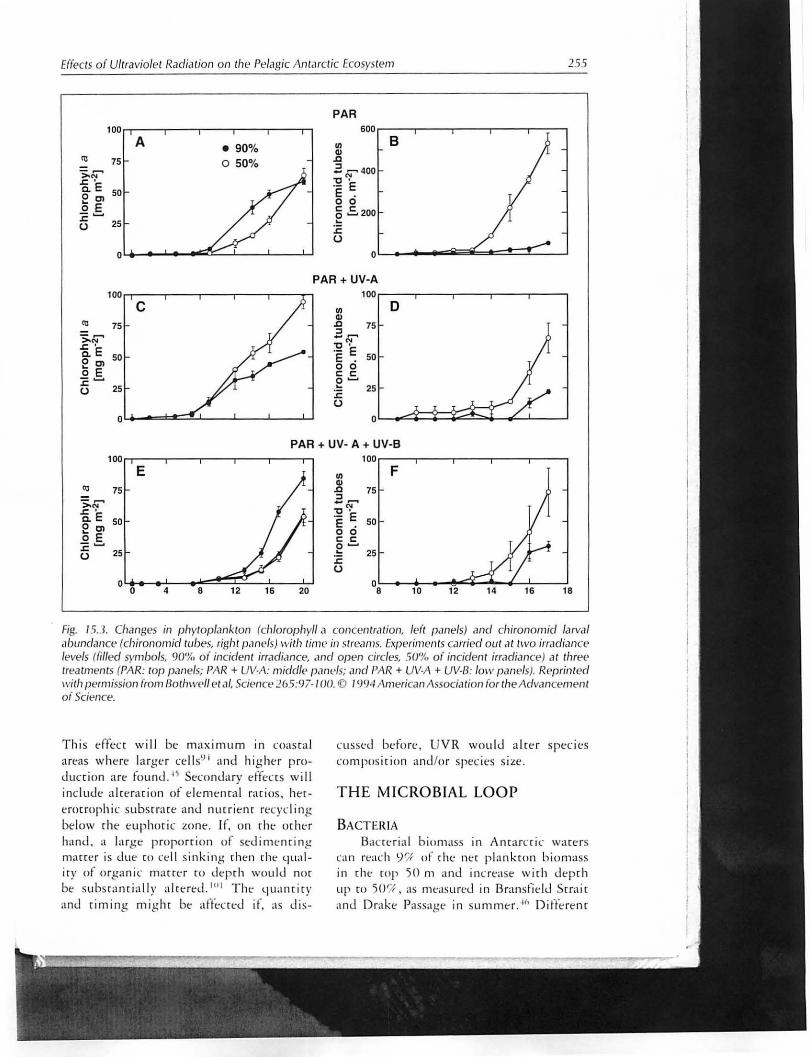
ZOOPLANKTON UV t'ffec£s on zooplankton , under nor
mal and decreased ozone conditions in rempt'rare waters, afftct zooplankton su rvival. reproduct ion and grazing .'}'} It is nor cif.'ar fro m these res ults if d t"c reastd g ra zi n,t: would resu lt in a reversa l of UV ent-crs 011
phyrop lankton, as observed for a chrnnomid/diilwm interaction in te mpl'ract.' freshwatt'r strt"am beds (Fi g, 15, .1 ), \Y/ t' call expecc that a 50 '/{ mortality of a g razer would decrease g razi ng p ress ure and fa vor phyroplankron growth. The poss ibi lity of grazing revtrs ing delttt"riulis effeCts of UV on phyroplankron and rhe relarivt' imporulIlce of grazing in controll ing phytoplank ton popu lat ion growth in any ,g ivt"n comf11uniry is currtntly a mar rer of specula rioll . U ndt"r current U V irradinllce. ovt'fall tll'creast' in primary production by U V in rill' Antarcti c t"up hotic zone is t'st ir1"wted .11
6-2Y/f of marg inal ice zone production .:" :' Th e ovtra ll rt"su lt would depend on rill' dft'Ct of UVR on Anrarct ic graze rs, an'raged for rhe t uphor ic ZOIlt". and on rilllt' .sca les represen tative of phywplankro n ;I t
clll'nulatioll at ambienr tempt'rctrurl' (d.IY:[(l wtl'ks. if Wt' assume i t spt'cifi c ~ro\\,lh ra rt' of 0.1-0, ,) d- I), ll
SEDIMENTATION Pottnria l changes in g razing prt'ssufl'
wi ll affect st"dimelltatiol1 of particu l:lIl' matter. In areas whert:· organic matter snli· men rar ion OU t of rhe euphoric zone is d Ul '
to g ra zt r ( i,e, k rill ) ftcal pe l lt'r s . II ~ ' Wl'
m ig- In ex pen a shifr ro cdl st d imt'ntation. assuming no change in p rima ry p roduction . Thus, rhe pulse of orbani c matte r after a bloom could consist mainl y of intilCt cell s.
Effects of Ultraviolet Rae/;alion on tile Pelagic Antarctic Ecosys tem 255
PAR 100 600
A • 90% .. B .. '" 75 0 50% .c =~ .a N" 400 > N .c ' "0 ' a.E .- E o '"
50 E . (; E o 0
:;:-25
e oS 200 U :c
U
0 0
PAR + UV-A 100 100
C .. 0 .. '" 75 .c 75
" >'N" ; ~ .c ' a.E .- E 50 0", 50 E .
(; E 00 c: c:
:;:-25 e- 25 U :c
U
0 0
PAR + UV- A + UV-B 100 100
E .. F ttl
.. 75 .c 75
~N" " .c ' ; ~ a.E 50 .- E 50 o '"
E . (; E o 0
c: c: :;:-25 e- 25 U :c
U
0 0 0 • • 12 16 20 • 10 12 ,. 16 ,.
Fig. 15.3. Changes in pi1ytopl.lnktoll (chlorophyll a conce/Hr.ltiofl, left pclnels) and chirollolllid larval abundance (cilironomid wbps, right panels) with tim e> in streams. Expprimenls carried Qut at two irradiance levels (filled s}'mbols, 90'1./ of inciden t irradi.lIlce, .1fld open circles, 5()% of incident irradiatlce} at three treatments (PAR: top panels: PAR + UV·A: middle parll'/s; and PAR + UV·A + UV·13: low pclnels). Reprinted with permission from Bothwell et aI, Science 26.5:97·/ 00. [) 1994 American Association [or the Ad va ncem ellt o f Science.
T his tffecr w ill be maxil11um in coasral art'as where larger ce ll s') I and hight f product ion art fO Ll nu. ''j Secondary effects wil l include alrt'r<lriol1 of t' lel11t'nral rar ios . hertrorroph ic subsr rare and IllHrienr recycling below rhe euphoric zone. If, on rhe m htr hand. a large p roportion of stdim (.' nrin .c manef is due [0 cell si nking rhen rhe lJual iry of orga ni c ma[[e r co deprh wou ld nor bt" subs ranriall y alrered. 1I 1! The q uan r iry and riming mighr bt afte:ctt"d if. as Jis-
cussed before , UV R wou ld a lre r speCies composirion and /or specit"s s ize.
THE M ICROBIAL LOO P
B ACTERIA
Bant'fi.d biomass in Anrarn ic warers can reach 9 '/f of rhl' nee plan kcon biomass in rhe rop 50 m and increase wirh deprh lip co 507r . as Illeilsurt"d in Bransfield Srrai r and Drakt Passage in summer. 1(' DitYt" rtlH
from othtr parts o( rhe oceall, th l' n: is no correlarion hl"twl'L'n phyropl.inkron and bacterial biomass in Anran.:t ic warers'l l,li l
,;
and rhe reasoll (or this differt'nct' I S
unclear. 1(1;
UV R reduces hacrni;ll acriv:ry III remperare (oastal waters in rhe rop 5 m of the water co lumn, with no indirarion of higher resisran(e in surfan' popu lations as opposed ro chose from depth .1II1 Inhibit ion was observed at an irradi<1 nce equal to
0. 7 W 111 -'; . UV-B was also found to photochem ically degrade bacterial extrace llular t'nzymes . I!H The com bination of decreased bacterial activity and the degradarion of ex tracellular enzymes redu ct'S t he flow or energy through rhl" microbial loop. Thi s ('ffeer is COLI l1[eracred , or at leasr diminished , by rhe increase in bacterial subsrrate due to phorodegradat ion of DOM. Increased bacteria l activity at low UV -8 irradian ce wirh respect to da rk up t akt· (F ig. 15.4) was attributed to this process.
PHOTO-OXIDATION OF DOM UV-B inreranion wit h DOM is known
to produce oxygen radica ls and hydrogen peroxide (H,;O,! ) which call be considered oxidarive agell[s of biologica l membranes and have a negat ive impaer on plankton ic comI11L1nities.l!l ~ In addirion , mulriple studies have documenred rhe photo-ox idat ion of nOM responsib le for degrading highmolt'nilar weighr DOM inro low-l11olecu-
140
lar wei ,~.dH DOM (e.g. Fig. 15 . ) ) 111 '1 101.
which is readily ilvailab le for bacrni ;d (()nsumprion. ICI~.IIlS
The imporrance of rhe s izt dass 011
bantrial produnivity is srill a ma rr<: r Ill' debart. as Amon and Benner lll'l found rhat although bacter ial growth effi(it'l1(il's WL"rl'
hightr ,t[ low-mo lecular we ight DOM , rll~
tal baneri;.11 growth and respirarion wa ~ h igher at high-molt'cu lar wt'ight DOJ\I ( > I UOO da lrons). n:,sulring in a hi gher Clr~ bon based rilre of ut il ization. It is ((){) <:arlr to assess the degree to which UV pllOr() ~
oxidat ion of DOM would be or impOrrall ll" in Anrarnic surface waters. Given rhe dl' ~
bart' on whether bacterial aniviry is de ~ pressed at low temperature, I 111. 11 I and rill' potential ro le of substrate on po lar h.lnerial metabo lism, ll l rhe role of phy[()plallk ~
ron as providers of labi Ie DOC and p lw('ooxidation of DOM by UVR ',Ht' hoth critica l to An t:.1n: tic ecosystems.
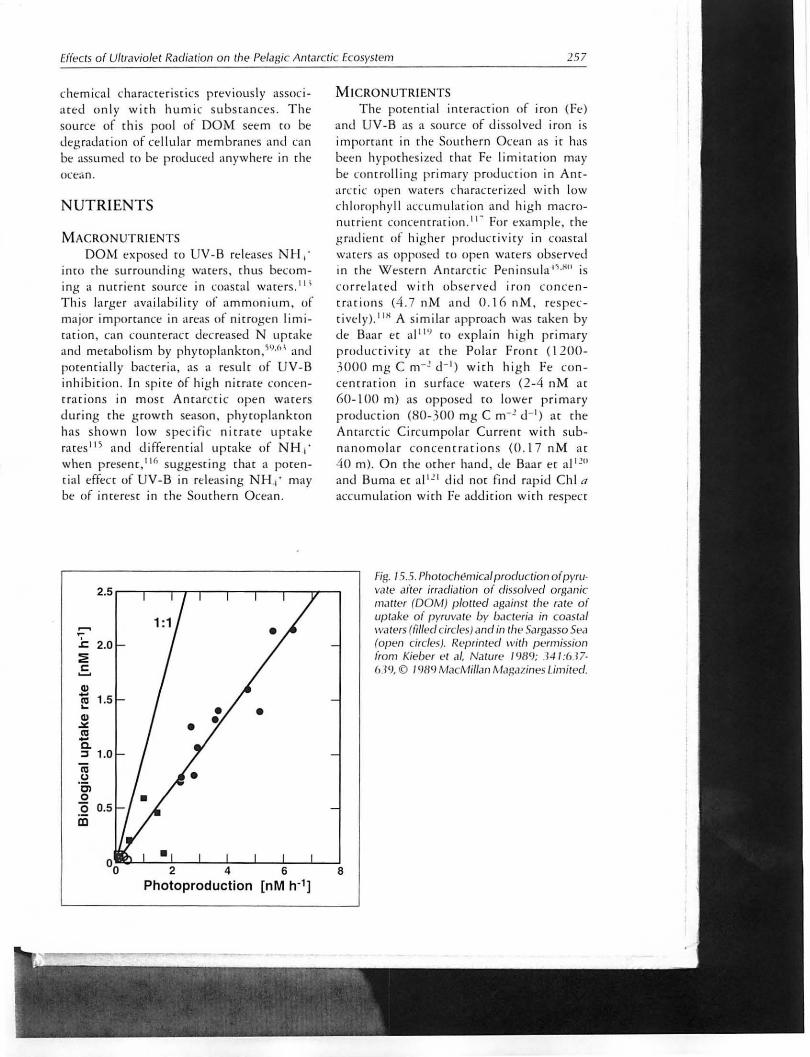
Phor()chemi ca l p roduction of d isso lvl'll am ino ilc ids from hum ic substances hav t· been shown co increase bacterial prmllJ r ~ tion in temperare coasta l warers. 11 \ UV-B was found to be t he most anive porrion or the solar spe([rum for this process whi(h cou ld be due both (0 high t r energy <Ind hig her absorption by the targt'r moIL-cull'. A lthough no or low humic acids are expected in Anrarctica, Lara and Thol1l;IS III have ident ified recalcirrant DOM produl..·· rion by marine p h ytoplankcon w irh
y = 52.7 - 40.7· log(x)
Fig . IS . ..J. /3aCfl'ri.1/ s('condclr~ ' produc· tion fB SJJ ) ,J.s a (unction o( UV·/3 (,leliatioll. Notl-' hight·,. produclion ,J( low u\I·n wilh rpspl-'c/ 10 rldr}.. U/)/tlkl'. Rl" drdwn (rolll /-h-' mell p i .1/. t"I,IIl1fl' H, /:7 f 7·7 1'). COfJ rrigIJI, A/d c/\/ifl.ln
EfI"ects of Ultraviolet Radiation on the Pe/<lgic Antarctic Ecosys tem 257
chem ical characttr ist ics p rtv iously assoc iated on ly w ith humi c subs tan ces. Th e source of thi s pool of DOM see m to be degradation of ctllul ar mt'mbranes and can be assumt'd ro bt' prouuced anywhe re in tht' occ-;m .
NUTRIENTS
MACRONUTRIENTS
DOM exposed to U V-B re leases NH .. in to dle su rround ing watNS, thus becoming a nllcritnr sOlln.:t' in coastal wate rs. ll' Thi s larger availabi lity of ammonium, of major im porranct in areas of nitrogt ll li m ica rion , can counteract dtcreast'd N upcake and mecabol ism by ph ytoplan kton, W.6' and potenria lly bacrtria , as a rtsu lr of U V-B inhibition . In spite of h igh nitrate conct ntrarion s in mos t Anta rnic open wattrs d uring the growth season, phytop lank ton has shown low spec ific nitrate lIptakt rates l
! ') and differentia l uptake of NH j '
wht n p resent, ll(' suggesting that a portnrial efferr of UV-B in releas ing N H ., · may be of interts t in tht Southern Ocea n.
M ICRON UTRIENTS The potentia l inreract ion of iron ( Fe)
and UV-B as a source of dissolved iron is imporcanr in t he Southern Ocean as it has betn hypothesized that Fe limitation may be con troll ing primary product io n in Antareric open waters characte ri zed with low chl orophyll acc umul at ion and hig h macronutr ient conct nrrat ion . ll - For txample . t he g radient of hightr productivity in coasta l wa(t'rs as opposed to optn W;l(t rS observed in th t' \'(/ts tern Antarctic Pen insu la 1').1"11 is co rre laetd wit h observe d iron concen tration s (4.7 nM a nd 0. 1 () nM, res pectively). lIS A sim ila r approach was taken by de Baar et a l II,) to explain h igh primary productivity at tht Po lar f ront ( 120() .1 000 mg C 01-' d- ') wit h hi g h Fe co ncentratIOn in surface waee rs (2-4 nM at 60- 100 m) as opposed to lowe r primary production (80- .,00 mg C m-' d- ') a t the Antarctic Ci rcumpolar Curn: nt with su bnan o molar co ncentra tion s (0. 17 nM a t 40 m ). On the other hand , de Baa r e t all.!o and Burna tt al 12 1 did not fin d rapi d C hi fl
accumu lation with Fe addi tion wit h respect
2.5 '--'--'-"--'--"----'--"-IT--'
Fig. / .5.5. Photochemical production ofpyru· va te after irradidtion of di$solved organic matter (DOM ) plotted agaimt the ra te of uptake of fJ}' fl lV.1U· by b,lCteria in coastal waters (tilled circles) and in the Sarg.15so Spa (op en circles). ReprinfE>d with permiSSion from Kie/)C'r t'l aI, Na tllre 19119; 341 :637· 639,!D /989 Mac/!"Iiflan M.lgazines Limited.
.2! .. 1.5 ~
~ E. :::l 1.0 .. " .0, o o 0.5 iii
• °0~~--2~~-~4~-L-~6~-L-~8
Photoproduction [nM h-1]
I ,
258 The E[fects of OlOnt~ De:'pletion on Aqucl t ic Ecos~/s t(:,lll s
1.0,--,--,--,--,----;,----;,----;,----;,----;n
i' 0.8 .;;.
~ 0.6
"C ~ 0.4 ::J
"C
~ 0.2
•
30 60 Time [min)
to controls in t he Wedde ll/Scotia Seas (both ueatments g rew at simi lar leve ls). The authors concluded t hat incubation effects overrode meta l, and in part icula r, Fe add it ion d ut' in pare to rhe excl usion ur large grazers from the t'xperi mental vessels. Iron add itions shifts phyroplankton composition from flage llates to diatoms, both in Antarct ic l1 1 and in eq uatoria l Pac ific waters. I 11 The ir rt'sults were nor as dramatic as t hose obse rved by Helbli ng et al 1'! \ who found increased pr imary product ivity and m icrozooplankton population in surface pe lag ic waters afte r addit ion of Fe. No effect was obse rved in deep pe lagic waters or coas tal wate rs off Seal Island . A sh ift to la rger cells is si milar to orher experi mentS of phytop lankton exposed to UVR I IJ.91'l whic h were attributed to differential cell su rvival and DNA damage.
In marine ox ic waters, Ft: " is the more stable form while Fe ':!' is mort' solub le and readily ava ilable to phytoplankton and bacter ial uprake .l'?-I T he concent ration of Ft: (111 ), (the sum of dissolved inorganic spec it'S) is t he relevant fac£O r ro cons ider with respect to the uptakt' of inorganic iron. I.? '
Its conce ntration va ri t's from lO-H to t 0-')
M. Rect'nt data indicates that 99.9% of t he dissolved iron in surface warers is bound with in organic co mplexes, resulting in subpicomolar concent ration of disso lved Fe(l II ). It is be lieved that the ligands for iron may orig inate from phytoplankton_ L!~
90
Fig. 15.6. Photoreduction of Fe(lII) in sealvaler (p/-I B.O·a.1 1 in the presence o( the diatom Phaeodactylum tricornu tunl under UVR. Fe(lI/) concerHr<l· lion of 5 JIM; diatom cOIlC(~n · Ira tion of I O~ cells mI' J. Re:" dra~·vn (rom Klima et ai, Marin(' Chemistry J 7: 15-27. Com/right 1992, with kind permission (rom Elsevier NL.
Sun light inc reases rates of oxidarion and reduction of iron , enhancing labile Ft· concentrations and p hytop lankton uptake. A lrhoug h UV-B photoreduces Fe(l ll ) to Fe(lI ) associated to inorgan ic ligand COI11-
p lexes, a larger reduction power is expecrnl from organic chromophores. 115 ReduCtion of organic l igands may occu r by rhe p horoproduced superoxide radical (0'-). In addit ion, ox idation of Pe(lI) can occur with p hotoproduced H 10 .! .
Photo-reducr ion of Fe( I1I ) to Fe(ll) is also attr ibuted to the act ion of marine p hytop lankton (F ig. 15.6). H igh conce ntrations of Pe( II ) were observed during p hytoplan kton spri ng blooms in J apal1(~se coastal waters. l Ui Experimt'nts wit h filrrare from a d iatom cult ure resu lted in photored uction of Fe(l l ) after addition of 5 pM Ft'(J lI ). T hi s proct'ss was attr ibured to rhe re least' of hydroca rboxylic acids by phytoplankto n, know n co reduce Fe(l ll ) ro Fe( Il ) in t he prt'st'nce of sun light 1.!·! and is more pronounced at lowt'r temperawres (5 0
vs. 20°C), important for Antarctic warers (su rface water tem peraturt' va ries from - 1.Bo to +2.5°C).
CONCLUSIONS Two important conc lus ions ca n be
d rawn from this discussion . First , evicit'nce has accumulatt'd to indicate that an aSSt"SS mt'nt of UV effects on Antarctic ecos}'sterns or marine ecosystems in genemi, will
Effects of Ultraviolet Radiation on the Pelagic Antarctic Ecosystem 259
gr.1lng h~zlng \ 1 Fig. 151 Scheme showing biotic (fu lf line) and abiotic (dashed line) relationships in the upper water column in the ocean, based on interactions discussed in the text. The arrow shows the direction of energy flow.
. • I~kag. "'"
MACROZOOPLANKTON • MICROZOOPLANKTON grazing
require experimenmtion on the ecosystem as a whole, or at least , isolate pares of it which include several interactions (i.e. the microbial loop). The predictive capability of adding effects on individual pools in the system is limited and experiments in temperate areas suggest that this can even be erroneous. Each level or species is not acting in a vacuum and biocic and abiot ic interactions will modify its genotypic response to UVR. Second, it is not possible to est imate UV effects on ecosystems withOut concurrent effort toward understanding environmental and biological forces which drive the system. Thus, UV effects are an added stress upon the system and need to be considered in conjunction with other potential limiting factors, such as nutrients, and other driving forces, such as mixing and ice cover.
In general, we speculate that a more profound and permanent effect of UVR might be the alteration of interaction between singular elements in the ecosystem than the direct effect of UV in inhibition of that same element (Fig. 15.7) . For example , changes in species composition might overshadow decrease in total primary producrion;16.19 increased substrate for heterotrophic activity might balance UV inhibition of bacterial growth; IO~ changes in iron availabili ty l 25 could counteraCt photosynthetic photoinhibition. The conse-
quences are far reaching in that the overall carbon balance might change due to
different proportions of carbon burial related to potential changes in cell size, grazing and subsequent sedimentation altering the CO2 interaction between atmosphere and oceans.
REFERENCES 1. Atkinson Rj, Matthews WA, Newman PA
et al. Evidence of rhe mid-latitude impact of Antarctic ozone depletion. Nature 1989; 340:290-294.
2. Diaz SB, Frederick JE, Lucas T et a1. Solar ultraviolet irradiance at Tierra del Fuego: comparisons of measurements and calculations over a full annual cycle. J Geophys
Lett 1996; 23:355-358. 3. Frederick JE, Lubin D. Solar ultraviolet ir
radiance at Palmer Station, Antarctica. In: Weiler (S, Penhale PA, eds. Ultravioler Radiation in Antarctica: Measurements and Biological Effects. Vol 62. Washington, D.C.: American Geophysical Union: Antarctic Research Series, 1994:43-5 2.
4. Frederick)E, Soulen PF, Diaz SB et al. Solar ultraviolet irradiance observed from Southern Argentina: September 1990 (0 March 1991.) Geophys Res 1993; 98:8891-8897.
5. Kerr )B, McElroy CT. Evidence for large upward trends of ultraviolet-B radiation linked to ozone depletion. Science 1993; 262:1032- 1034.
260 The Effects of Ozone Depletion on Aquatic Ecosystems
6. Kerr JB, McElroy CT. Analyzins ultraviolet-B radiation : Is there a trend? Science
Productivity of rhe Ocean: Present and Past. John Wiley and Sons, 1989:85-97.
9. Falkowski PG, Woodhead AD, eds. Primary Producciviry and Biogeochemical Cycles in the Sea. New York: Plenum Press, 1992: 213-237.
10. Antoine D. Andre J, More l A. Oceanic
primary product ion : II. Estimation at glo
bal scale from satellite (Coastal Zone Color Scanner) chlorophyll. Glob Biogeochem Cyc 1996; 10:57-69.
II. Smith RC, Baker KS, Byers ML ec a!. Primary productivity of rhe Palmer Long-Term Ecological Research Area and rhe Souchern Ocean. J Mar Sysc 1996b; (in press).
12. Rivkin RB, Putt M, Alexander SP et al. Biomass and produccion in polar piankronic and sea ice microbial communities: a comparative 5[ucly . Mar BioI 1989; 101 : 273-283.
13. Hader D-P, WorreS( RC, Kumar HD et al. Effects of increased solar ultraviolet radiation on aquatic ecosystems. Ambia 1995; 24:174-180.
15 . WorreS( RC, Van Dyke H, Thomson BE. Impacr of enhanced simulated solar ultraviolet radiation upon a marine community. Phocochem Phocobiol 1978; 27:471 -478.
16. Worrest RC, Wolniakowski KU, SCOtt JD et a!. Sensitivity of marine phytoplanktOn to UV-B radiation : impact upon a model ecosystem. Phorochcm PhotObiol 1981; 33:223-227.
17 . Calkins j. Thordard~ttir T. The ecological significance of solar UV radiation on aquatic organisms. Nacure 1980; 283:563-566.
19. Bothwell ML, Sherbot D, Roberge AC et al. Influence of natural ultraviolet radiation on lotic peri phytic diacom community
growth, biomass accrual, and species composition: shore-term versus long-rerm effects. J Phycol 1993; 29:24-35.
20. Azam F, Fenchel T, Field JG et a!. The ecological role of water-column microbes in the sea. Mar Ecol Pros Ser 1983; 10: 257-263.
2 1. Voytek MA . Addressing the biological effecrs of decreased ozone on the Anrarccic environment. Ambio 1990; 19:52-6 1.
22. Weiler CS, Penhale PA , eds. Ultraviolet Radiation in Amarceica: Measuremems and Biologi ca l Effects. Vol 62. WashingtOn, D.C. : American Geophysical Union, 1994.
23. Smith RC, Preze1in BB, Baker KS et a!. Ozone depletion: ultraviolet radiation and phycoplankron biology in Anrarctic waters. Science 1992; 255:952-959.
24. Booth CR, Lucas TB, Morrow JH. Ultraviolet radiation in Ancarctica: Measurements and biologica l effects. In: Weiler CS, Penhale PA, eds.Ultraviolet Radiation in Antarctica: Measuremencs and Biological Effeces. Vol 62. Washington, D.C.: American Geophysical Union, Antarcti c Resea rch Series 1994:17-37.
25. Helblins EW, Villafane V, Ferrario M et 011. Impact of naeural ultraviolet radiation on rates of photosynthesis and on specific marine phytoplankton species. Mar Ecol Prog Ser 1992; 80:89-1 00.
26. Lubin D, Micchell BG, Frederick JE ec a!. contribution toward understanding rhe biospherical significance of Ancarctic ozone depletion . J Geophys Res 1992; 97(D8): 7817-7828.
27. Boucher N, Prczcl in BB, Evens T C[ 0.1.
lcecolors '93: biological weighting funcrion for rhe ultraviolet inhibirion of carbon fixation in a natural Antarceic phytoplankton community. Ant J US 1994; XXIX: 272-275.
28. Cullen JJ, Neale PJ , Lesser MP. Biological weigh ring function for the inhibition of phytoplankron photosynthesis by ultraviolet radiation. Science 1992; 258:646-650.
29. Jones LW, Kok B. Phocoinhibition ofchloroplast reaCtions. Il . Multiple effects. Plant Physiol 1966; 41:1044-1049.
30. Serlow RB. The wavelengrhs in sunlight effective in producing skin cancer: A [hea-
Effects of Ultraviolet Radiation on the Pelagic Antarctic Ecosystem 26 1
rerical analysis. Proc Nar Acad Sci USA
19 74 ; 7 1 :.3.36.)-3 .)66.
5 1. \'{Iood \'(IF. Phomadaprive responses of the
tropical red alga 1:.1((/)1.','1111(1 .1'/ 1';(1 1101/ Schmirz
(Gigarrinales) ({) ultra-violet radiation .
Aquatic Bown)' 1989: 53:.4 I-5 1.
32. Morales RGE. Jam GP, Cabrera $. Solar
ultraviolet radi ·;\tio!l meaSllrl'menrs by
v- ni trobenzaldehydc.· aCt i nonH,: ( ry . Li III nol
Oceanow 19>13: ,, 11 :7 0.) -705.
:n. Karentz D. Luczl' 1.1-1 . Evaluation or bio
log ically harmful ultf;lvioll'r radiation in
Antarctica with a biological dosimeter de
signed for aquat ic envi ronmt:nrs. Limllol
Oceanogr 1990; ,);;:54>1-56 1.
,,4. Roy CR. Gies HP, Tomlinson D\XI et al.
EtTecrs of Owne Depletion on the U ltravio
let Radiation Environml'nr at thl' Austra
li an Stat ions in Antarcrica. In : \Xleiler CS,
Penhale PA. eds. U ltravio let Radiation in
Antarctica: Me,lsun:menrs and Biological
Effects. Vol 62. Washingron. D.C.: Aml'ri
can Geophys ical Union, 11)94: 1-1 5,
35. Smith RC. Ownl' , middle ultravio let radia
tion and the aquaric environment. Phoro
chern Photobiol 1989; 50:459-468. 36. Smith RC, Culle n JJ. Ellens of UV radia
t ion on phytoplankton. Rev Geophys 19>15;
Supplement: 12 1 1-1 22., .
57 . Sm ith Re, Baker KS. Relllote sensing of
chlorophyll. In: Godby EA. Ouerlllan J. eds.
COSPAR , The Cont ri bur ion of Space Ob
se rvations co G loba l rood Informarion Sys
rems. Oxford, New York : Pergalllon Press,
1978:161-1 72. 38. Smith RC, Bnker KS. Optical prop('rries of
rhe clearest narural ware rs COO-SOO 11111).
Appl Optics 1981 ; 20:1 77- 184. 39. Kirk JTO . H,lrg re<lves SR. Morri s DIl (' r
al. Measurements of U V -S radiarion in twO
freshwa ter lakes: an insrrument inrt' rcom
pa ri son. Arch Hydrobiol Still Ergl' bn
Lin"lOol 1994 ; .1.1:7 1-99.
40. Baker KS, Smirh RC. Middle ulravioler ir
radiance at the ocean surface: measure menrs
and models. In: Calkins J. ed. TIll' Rol l' of
Ulrravio let Radiarion in lvla rin c.· Ecosystems.
Nnv York: Plt-num Publishin,L: Co ..
1>182:71)-9 1.
4 1. Bri cilud A. Mort! 1\, Pril'llf L. Absorprion
by dissolved o rganic matter o( tile sea (yel-
low subsrance) in the UV and visib l(:' do
mains. Limnol Oc{;'ano~r 19R 1; 26:4 .)-,)3.
-12. Krame r K. Effects of increased solar uv-b
radiarion on coastal marine ecosystems: An
overview. In: Bt'uktma JJ, \\! oil' \\!). Brouns
J. eds. EX]lened EHi:ns of Climatic Changl'
on Marine Coasta l EcoSYStl·n1S. Bos ron:
Kluwer Acadl'lllice, 191)() : 1 1)5-21 (1.
"n . Mi{chell BG, Holm- Hansen O. Obstrv:l
tions and modtl int: or rhe Ancarnic phy
toplankton crop in rda rion to mixing d l' (1th .
Deep-Sea Res 11)91; .1H:9H 1-1 007.
44. Smith \'\10. Ntlson DM . Phytoplankton
bloom produced by a receding ic(' t'd,L;e in
the Ross S(:'a: spari;iI CUill'rl'nCe with tht'
d (:' nsity field. Science 1985: 210:1r,5-1 r,().
4 5. Sm ith RC, Baker KS, Verner M. Season.d
and interannual variabiliry of phyrplankron
biomass west of the Anrarrr ic Pen insula. J
Mar Sysr 1996n; (in prl'ss) .
46. Mullins B\'{I. Priddle J . Relarionsh ips be
(ween bacteria and phywplankron in the
BransfIeld Strait and Somhern Drake Pas
sage . Brirish Anrarni c S urvey I >I~7;
76 :51-64. 47. Holm-Hansen O. Mitchell BG. Vernet M,
5 1. Cullen JJ, Neale PJ. U ltravio ler radiarion.
()zoni,: deplerion. and marine photosynrht'
s is. Photos Res 1994; 59 :,,0 5- .1 20.
52. Villat;1I1e VE, i-leib lin.L! [\'\1. l-!olm - H:lIls('n
a et al. Acclimarization of Antarct ic natu
ral phytoplan kton assl'll1bia~es wlll'1l ex
posed w so lar uirraviol l't radiario1l . .J PI Rt's
1995; 17:2295-2306.
262 The Efrects or Ozone Depletion on Aquatic Ecosystems
53. Gieskes W, Kraay G \X' . Transmission of ulrraviolet lighr in the Weddell Sea: reporr of rhe first measurt ments made in the AntarctIc. BIOMASS Newsle([er 1990; 12: 12-1 4.
54. Helbling EW, Villafaiie V, Holm-H. nsen O. EffeCts of ultraviolet radiation on Antarctic marine phytoplanktOn photOsynthesis with particular attention to the inOuence of mixing. In : Wei ler CS, Penhale P, eds. Ultraviolet Radiation in Antarctica: Measurements and Biological Effects. Vol 62. \X'ashingtOn, D.C.: Antarnic Research Series, 1994:207 -227.
55. Cullen ]], Ne.le Pl . Qu.nrifyi ng rhe effects of ultrav iolet radiat ion on aquatic photOsynthesis. In : YamamotO H , Smith CM, eds. Phocosynthct ic Responses to the Environment. Washing tOn, D.C.: American Sociery of PI . nr Physiologisrs , 1993:45 -60.
56. Neale P] , Lesser MP, Cullen]). Effects of ultraviolet radiation on ehe photOsynthesis of phytopl ankton in the v ic init y of McMurdo Sracion, AntarCtica. In: Weiler CS, Penhale PA , eds. Ultraviolet Radiat ion in Anrarnica: Measurements and Biolog ical EffeCts. Vol 62. Washington, D.C.: Antarctic Research Series, 1994: 125- 142.
57. Collos Y, Slawyk G. Nitrogen uptake and assimilation by marine phyroplankton . In : Falkowsk i PG , ed . Primary Productivity in the Sea. 3 1. New York : Plenum Pr<.-ss, 1980: 195 -2 11.
58. Dohler G. lmpacr of UV-B radiation on [1 ~N]ammonia and [n N]nicrate uptake of
Oil)'/II'" brighlll/e/lii. Photobiochem Phowbiophys 1986; I 1: II 5-1 21.
59. D6hl t r G . Impact of UV-B radiation on uptake of I ~N -ammonia and n N-nitrate by phywplankron of the Wadden Sea. Mar Bioi 1992 ; 112:485 -489.
60. Dohler G, Buchmann T. Effects of UV-A and UV -8 irrad iance on pigments and n N _
ammonium 'lss imilation of rhe Haptophycean Pm,fom. ] PI Phys 1995; 146:29-34.
61. Goes ]1, H. nda N , Taguchi S er . 1. Impact of UV radia tion on the production panerns and composition of dissolved free and combined amino acids in marine phytOplankron.] Plankron Res 1995 ; 17:1 337- 1362.
62. Sinha RP, Kumar HD , Kumar A Ct al. Effeccs of UV-B irradiaeion on g rowt h, survival, pigmemarion and nierogen mctaboli sm en zymes in cyanoba cteria . Ana Prorozool 1995; 34:187- 192.
63. Goes ]l , Handa N , T.g uchi S er.1. Changes in ehe panerns of biosynthesis and composition of amino acids in a marine ph)l
tOplankte r exposed co ultraviolet-B radiation : nieroge n lim itaeion impli cated. Phorochem Phorobiol 1995; 62:7 03-7 10.
64. Sharp JH . Excretion of organic maHer by marine phytoplankton: do healthy cells do ir ' Limnol Oce.nogr 1977; 22:381 -389.
65. Fogg GE, N.lewajko C, W.rt WD. Exrracellular produces of phyroplankcon phorosynrhesis. Proc R Soc Lond Ser B 1965; 162:5 17-534.
66. Bj~rnsen PK. Phyroplankton exudarion of organic matter: why do healthy cells do it ? Limnol O ccanogr 1988; 33: 151-1 54.
67. Wood AM, Rai H, Garnier J et al. Praceical approaches co algal excreeion. Mar M icrob Food Webs 1992; 6:2 1-38.
68. Mag ue TH, Friberg E, Hughes D] er ,I. Exeracellular release of carbon by marine phytoplankton ; a physiological approach. Limnol O ceanogr 1980; 25 :262-279.
69. Wi lliams PJ Ie B. The importance of losses during microbial growth: commemary on the phys iology, measuremem and ecolosy of the release of dissolved organic marerial. Mar Microb Food Webs 1990; 4 : 175-206.
70. Kirchman DL, Suzuki Y, Garside C er ill. 1-1 igh curnover rates of dissolved organic carbon during a spring phytoplankeon bloom . Narure 1991; 352:612-614.
7 1. Myklesrad S, Haug A. Production of carbo· hydra tl"s by the mar ine diarom C b(lel O(frDS
(Ijfi,,;! vOIr /l lillei (Gran) Husted. 1. Effect of (he concentration of nurriems in rhe cul {Ure medium. J Exp Mar Bioi Ecol 1972;
9: 125 -1 36. 72. Sakshaug E, Myklesrad S, Krogh T er al.
Product ion of protein and carbohydrate in the Dinoflagellate Amphidi1lill1lJ ((Ir/L.,.i. Some prdiminary resul ts . Norw J Bor 1975; 20:2 11 -2 18.
73. Hellebusc JA . Excreeion of some org.mic co mpounds by marine phytop lan kton . Limnol O ceanogr 1965; 10: 192-206.
Effects of Ultraviolet Radiation on the Pelagic Antarctic Ecosystem 263
74. Myklesrad S. Production of carbohydratts by marine planktonic diatoms. I. Comparison of nine different species in culture. J Exp Mar Bioi Ecol 1974; 15:26 1-274.
75. Veldhuis MJW, Admiraal W . Transfer of phorosymhetic products in gelatinous colonies of PhaeoryJtiJ pouchetii (Haptophyceae> and its effect on the measure. Mar Ecol Prog Ser 1985; 26:301-304.
76. Davidson AT, Marcham H] . Prot ist abundance and carbon concentration dur ing a PhaeoryJlis-clominated bloom at an Antarcric sire. Polar Bioi 1992; 12:387-395.
77. Richardson TL, Cullen JJ . Changes in bouyancy and chemical composition during growth of a coastal marine diatom: ecological and biogeological consequences. Mar Ecol Prog Ser 1995; PP 77-90 and VI 28, NI- 3
78. Vernet M, Matrai PA. Symhesis of part iculate and extracellular carbon by phytOplanktOn in the Barents Sea. J Geophys ResOceans 1996; in press.
79. Smich DC, Steward GF, Long RA ec a1. Bacterial mediat ion of carbon fluxes during a diatom bloom in a mesocosrn. Deep-Sea Res 11 1995; 42:75 -97.
80. Holm-Hansen 0 , Micchell BG. Spacial and temporal d istribution of phytOplanktOn and primary production in the western Bransfield Strai t region. Deep-Sea Res 199 1; 38,:96 1-980.
8 1. Karentz D, Spero HJ. Response of a natural PhaeoryJlis population to ambient fluctuations of UVB radiation caused by Antarctic ozone deple t ion. ] Plankton Res 1995; 17: 177 1-1 789.
82. Behrenfeld MJ, Hardy J T, Lee HI. Chronic effects of ultrav iolet-B rad iation on growth and cell volume of PhaeodactyluTlI (Bac illariophyceae). J Phycol 1992; 28:757 -760.
83. Hargraves PE , Zhang J , Wang R ec a1. G rowth cha racte ri stics of the di ato m PJeJldollitzJchia pUlIgem and P. fraudulen ttl exposed to ultraviolet rad iation. Hydrobiologia 1993; 269/270:207-2 12.
84. Lesser MP. Accl imation of phycoplankton to UV -B rad iation: oxidative stress and phmoinhibition of photosynthes is are not prevented by UV-absorbing compounds in rhe dinoflagellate Prorocentrllm micam. Mar Ecol Prog Ser 1996; 132:287-297.
85 . Vernet M. UV radiation in Antarct ic waters: response of phytOplanktOn pig mencs. In: Mi <chell BG, Holm-Hansen 0 , Sobolev I, cds. Response of marine phytoplank ton to natural variations in UV-B fl ux. Washing ton, D.C: Chemical Manufacturers Assoc iat ion, Proceed ings of a W orkshop , Scripps Institut ion of Oceanography, La Jolla, CA, April 5., 1990.
86. Karentz D. Ultraviolet colerance mechanisms in Ancarctic marine organisms. In: Weiler CS, Penhale PA , cds. Ultraviolet radiation in Antarctica: Measurements and Biological Effects. Vol 62. Washington, D.C: American Geophysical Union: Ancarc tic Research Series, 1994:93- 110.
87. Smith RC, Baker KS. Stratospheric ozone, middle ultraviolet radiation and carbon-1 4 measurements of marine productivity. Science 1980; 208:592-593.
88. Smith RC Baker KS. Assessment of the influence of enhanced UV-B on marine primary productivity. In : Calkins ] , ed . The Role of Solar Ultraviolet Radiation in Marine Ecosystems. New York: Plenum Publishing Co., 1982:509-537.
89. Kullenberg G. Note on the role of vertical mixing in relation to effec ts of UV radiation on the marine environmenc. In : Calkins ] , cd. The Role of Solar Ultrav iolet Radiation in Marine Ecosys tems. New York : Plenum Press, 1982:283-292.
90. Bidigare RR . POtencial effects of UV-B radiation on marine organisms of the southern ocean: dist ri butions of phytoplankton and krill during austral spri ng. Photochem Phocobiol 1989; 50:469-477.
91. Karentz D. Ecological considerat ions of Antarctic ozone depletion. Antarctic Science 199 1; 3:3- 11.
92. Ferreyra GA, Schloss fR , Demers S et al. PhytOplankton responses to natural ultraviolet irradiance during ea rly spring in the Weddell-Scotia confluence: an ex peri menral approach. An< J US 1994; XXI X: 268-270.
93. Malone TC Size-fractionated primary productivity of marine phytoplankton. In : Falkowski PG, ed. Primary Productiv ity in the Sea. New York, London: Plenum Press, 1980:30 1-319.
264 The Effects of Ozone Depletion on Aquatic Ecosystems
94 . Fiala M, Delille D. Variability and inceractions of phytoplankton and bacrerioplankton in the Antarctic neritic area. Mar Eeol Prog Ser 1992; 89:135-146.
95 . Bidigare RR, Iriane )L, Kang S-H et a1 . Phytoplankton: Quancirarive and Qualitative Assessmencs . In: Ross RM, Hofmann EE, Quetin LB, eds. Foundations for Ecological Research West of the Ancarctic Peninsula. Washington, D.C.: American Ge0-physical Union, 1991; 70:173-198.
96. Karencl D, Cleaver )E, Mitchell DL. Cell survival characteristics and molecular responses of Antarctic phycoplankron to ultraviolet-b radiation . J Phycol 1991; 27:326-341.
97. Vernet M, Brody EA, Holm-Hansen 0 et al. The response of Antarctic phytoplankton to ultraviolet light: absorption, photosynthesis, and taxonomic composition. In: Weiler CS, Penhale PA, eds. Ultra Violet Radiation in Antarctica: Measurements and Biological Effects. Vol 62 . Washington, D.C.: American Geophysical Union, 1994: 143-158.
98. McMinn AD, Heijnis H, Hodgson D. Minimal effects of UVB radiation Amarctic diatoms over the past 20 years. Nature 1994; 370:547-549.
99. Hunter )R, Kaupp SE, Taylor )H. Assessment of effects of UV radiation on marine fisb larvae. In: Calkins), ed. The Role of Solar Ultraviolet Radiation in Marine Ecosystems. New York: Plenum Publishing Co, 1982:459-497.
100. von Bodungen B, Smetacek V, Tiller B er al. Primary production and sedimentation during spring in the Anrarctic Peninsula region. Deep-Sea Res 1986; 33:177-194.
101. Wassmann P, Verner M, Mitchell BG et al. Mass sedimentation of Phaeoryslis poucherii in the Barents Sea. Mar Ecol Prog Ser 1990; 66: 183-195.
102. Bird DF, Karl DM. Bacterial growth, abundance and loss due to protozoan grazing during the 1989 spring bloom. Ant ) US 1990; 25:156-157.
103 . Karl DM. Microbial processes in the Southern Ocean. In: Friedmann EI, ed. Anrarctic Microbiology. New York: John Wiley and Sons, Inc. , 1992.
104. Herndl G), Muller-Niklas G, Frick) . Major role of ultraviolet-B in controlling bacterioplankton growth in the surface layer of the ocean. Nature 1993; 361 :717-719.
105. Mopper K, Zhou X, Kieber R) et al. Photochemical degradation of dissolved organic carbon and its impact on the oceanic catbon cycle. Nature 1991 ; 353:60-62.
106. Kieber D), McDaniel), Mopper K. Phorochemical source of biological substrates in sea water: implications for carbon cycling. Nature 1989; 341 :637-639.
107. Moran MA, Hodson RE . SuppOrt of bacterioplankton production by dissolved humic substances from three marine environments. Mar Ecol Prog Ser 1994; 110:241-247.
108. Morris DP, Zagarese H , Williams CE et al. The attenuation of solar UV radiation in lakes and the role of dissolved organic carbon. Limnol Oceanogr 1995; 40:1381 -1391.
109. Amon RMW, Benner R. Bacterial utilization of different size classes of dissolved organic matter. Limnol Oceanogr 1996; 41 :41-51.
110. Pomeroy LR, Diebel D. Temperature regulation of bacterial activity during the spring bloom in Newfoundland coastal waters. Science 1986; 233:359-361.
111. Thingstad F, Billen G. Microbial degradation of Phaeorysris material in the water column. ) Mar SYSt 1994; 5:55-65.
112. Pomeroy LR, Weibe WG. Energy sources for microbial food web. Mar Microb Food Webs 1993; 7:101-118.
113. Bushaw KL, Zepp RG, Tatr MA et a1 . Photochemical release of biologically available nitrogen from aquatic dissolved organic matter. Nature 1996; 381 :404-407 .
114. Lara R), Thomas DN. Formation of recalcitrant organic matter: humification dynamics of algal derived dissolved organic carbon and its hydrophobic fractions . Mar Chern 1995; 51:193-199.
115. Dugdale RC, Wilkerson FP. Low specific nitrate uptake rate: a common feature of high-nutrient. low-chlorophyll marine ecosystems. Limnal Oceanogr 1991; 36: 1678-1688.
116. Tupas LM, Koike I, Karl DM et al. Nitrogen metabolism by heterotrophic bacterial
Effects of Ultraviolet Radiation on the Pelagic Antarctic Ecosystem 265
l.l ssemblages in Amarcr ic coasra l wmers. Po la, Bioi 1994; 14: 195-204.
11 7. MarrinJH , Gordon RM , Firzwurer SE. The
cas~ for iron . Limnol O ceano,gr 199 1:
56: 1793-1 S02 . 118. Marr in JH , Gordon RM, Fi(zW,lrer SE. Iron
in Aman..' (i c waters. Narure I 99(}; 5-i5:
156- 158. 11 9. De Baa, I-I)W. BlIma AG). Noltin~ Rf et
ill. On iron limirario n in rhe Sourhern
On'an : eXpl'riIllL'mal obsL"rvarions in rhe
\'(/ t:clde ll and Scoria 5e'ls. Mar Ecol Prog 5L" r 1990; 65: lOS- In.
120. De Baa, I-I) W. de ) ong )TM. Baake, DC E er al. im porranc(' of iron for plankton blcxlnlS
and carbon dioxide drawdown in rhe 50urh
ern Oce;ln . N;lrure 1995 : 375:4 12-415 . 121. BlIl11a AG) . De Baa, H) W. Noltig RF et
;I I. Meml enrichmem experimenrs in rhe
\Xleddell-Scoria Seas: Effecrs of iron and
manganese on variolls plankron communi
t ies. Limnol Oceanogr 199 1; 36: 1865 -1 878.
122. Chave, FP. Blick KR . Coale KH et al.
Growth rares. graz ing. sink ing. and iron
limi r;l rion of equaro rial Pacific ph}' (Oplank
ro n. Limnol Oceanogr 199 1: 36: 18 16- 18 .15. 123. Helbl ing EW , Villal,lne V. Holm- Hansen
a Effecr of iron on producr iviry and siZt,
disrr iburion of Anrarcric p hyropl ;l nkron .
Li mnolOceanog' 199 1: 56:1879-I R85. 124. Kuma K. Nakabayashi S, Suzuki Y ('{ a l.
Photo-ftclucrion of Fdlll ) by di ssolved or
g'lnic substances and exis tence of h :(Il ) in seawarer during spring blooms. Mar C hern
1992; 37: 15-27 . 125 . \'{/dls ML. Price N M. Bruland K\'{/ . Iron
<..'htm isrry in seawarer and irs relarionship
co phytop lankton : a workshop reporr. ~'I ar
Chem 1995 ; 48: 157-1 82. 126. Nakabayash i S. Kudo I, Kuma K e t a l.
Exisrence of di ssolved Fe!' in a spring bloom
ar Funka Bay. Jpn Soc Fish Oceanogr 1989:
5.l:649-680.
ENVIRONMENT Ai
INTELLIGENCE UNIT
THE EFFECTS OF OZONE DEPLETION ON AQUATIC ECOSYSTEMS
Donat-P. Hader Friedrich -Alexander -U niversirar
Insritut ftir Botanik und Pharmazeutische Biologie Erlangen, Germany
Academic Press R.G. LANDES COMPANY
AUSTIN
r;:====== CONTENTS ========il
1. Stratospheric Ozone Depletion and Increase in Ultraviolet Radiation .............. ......... .... ............................... .... 1 Donat-P. Hader How Is the Ozone Layer Affected? ........................................................ I UV Increase Was Partially Masked by Air Pollution ........... ........ ........... 2 Negative Effects of Increased Solar UV Radiation on Life ..... ..... ........... 4
2. Making Links: From Causes to Effects to Action ........................ 5 Canice Nolan
3. Consequences of the Effects of Increased Solar Ultraviolet Radiation on Aquatic Ecosystems .... ... ........... .. ... .... ... .... ...... .... . 11 Donat-P. Hader and Robert C. Worrest Importance of Aquatic Ecosystems ...................................••................. II Effects of Solar UV-B on Aquatic Ecosystems .. ... .. .. .. ........ ............ .... .. II Global Distribution of Phytoplankton .... ..... ............ ... ... ....... .............. 12 Penetration of Solar Radiation into the Water Column ..... ..... ..... .. ... ... 12 Macroalgae and Seagrasses ................................ .. ....................... .......... 18 Cyanobacteria ..... ....... ........... .. .... .... ............... ...... ........... ...... .. ............ 19 Screening Pigments ............ ...... ... ....... ... ......... .......... .. ........ ... .............. 21 Carbon Dioxide Uptake and Its Role in Global Warming ................... 2 1 Consumers ........ .... .. ... ... .... .......... ................... .................... ....... .. .. ... .. . 21 Substitutes of CFCs and Their Degradation Products .... .. ......... .... .. .... 25 Consequences .. .. ..... ... .... ..... ....... ..... .. ....... .... ............ .. ..... .. .............. ... . 25
7. Biological UV Dosimetry ....... ..... .. .......... ..... .... ... ... .. ...... ... ...... 119 Gerda Horneck The Biological Effectiveness of Solar UV Radiation ..... .. ...... .... ..... ... . 119 Quantification of the Biological Effectiveness of Solar Radiation .... .. 120 Characteristics of Biological Dosimeters ...................... ......... .. ...... .... . 125 Scope of Application of Biological Dosimeters .................................. 134
8. Role of Uhraviolet Radiation on Bacterioplankron Activity .... 143 Gerhard! Herndl Introduction ........................................... ........... ............. .................. 143 Role of Ultraviolet Radiation on Bacterioplankron Cells ................... 144 Role of Ultraviolet Radiation on Dissolved Organic Matter ..... .. ...... . 148 Interaction Between Bacterioplankron Activiry and DOM
in the Upper Mixed Water Column .. ......................... ... ...... ... .. ... .. 150 Future Research Directions ..... .... ....................... .. .......... ... ................ 150
'I
ill
9. Optical Properties and Phytoplankton Composition in a Freshwater Ecosystem (Main-Donau-Canal} .......... ................ . 155 Maria A. Hiider and Donat-P. Hiider Introduction ....................... .. ....... ....................... ......... .. .. .......... ...... . 155 Materials and Methods ...... ...................... .. ................... ............ ........ 156 Results ............................................................................... ..... .. ........ 157 Discussion ... .......................... ... ..... .................... ..... ..... ...... ................ 168
10. The Effects ofUltraviolet-B Radiation on Amphibians in Natural Ecosystems .. .. ...... ......................... 175 Andrew R. BIa"stein and j oseph M. Kiesecker Initial Laboratory Studies ............................ ..... ...... ....... .. ........ .......... 176 Field Studies ...................... .... .... ................................................. ...... 176 Field Experiments at Relatively High Elevation Sites in Oregon ...... . 177 Field Experiments at Low Elevation Sites in Oregon ..... .................. .. 178 Synergism ofUV-B Radiation with Other Agents .. ........ ................... 180 Discussion ........ .... .. ............ .. ......... .............. ..... ................ .. ..... .. ........ 182 Conclusions ........ ... .......... ......................... ....... .. ......... .. ....... ............. 185
II. Impacts of UV-B Irradiation on Rice-Field Cyanobacteria .... . 189 Rajeshwar P. Sinha and Donat-P. Hiider Introduction .. ....................... ........... ................ ..... ................... ...... ... 189 Conclusion .......... .. .............. ..... .. .......................... .... ...... ... ....... .. ....... 195
12. Studies of Effects of UV-B Radiation on Aquatic Model Ecosystems ................................................ 199 Sterz-Ake Wiingberg arid j ohmme-Sophie Selmer Introduction ......... ............................................... ....... ... ............ ....... 199 Model Ecosystems Used for Assessing Effects ofUV-B Radiation