17
Page 106 Chapter 5 Analysis of cysteine residues and inhibitor studies

Page 106
Chapter 5 Analysis of cysteine residues and inhibitor studies

Chapter 5
Page 107
SACOL0085 peptidase is found to be grouped in M20D family of peptidases
according to the sequence similarity. This was observed when a homology search
for this polypeptide sequence was done with National Center for Biotechnology
Information TBLASTN, against the EMBL, DDBJ, GenBank, and PDB databases.
The other members of this class for which the three-dimensional structure is known
include-YxeP from Bacillus subtillis (PDB id: 1YSJ), an auxin conjugate
aminohydrolase ILL2, from Arabidopsis thaliana (PDB id: 1XMB) (Bitto et al., 2009)
and HmrA, an antibiotic resistance factor from Staphylococcus aureus subsp. aureus
N315 (PDB id: 3RAM) (Botelho et al., 2011).
The enzymes in M20D family are previously reported to show catalytic activity in the
presence of two metal ions generally preferring Mn2+ or Cu2+ as cofactors (LeClere et
al., 2002; Bitto et al., 2009). Even though, the catalytic machinery for M20D class of
metalloenzymes has been established previously with example of ILL2 from
Arabidopsis thaliana (Bitto et al., 2009), more emphasis on the structural
characteristics which modulate function stay less explored.
ILL2 from Arabidopsis thaliana (PDB id: 1XMB) which belong to M20D family is an
amidohydrolase that release the active plant hormone indole-3-acetic acid (IAA) from
its conjugated storage forms (Bitto et al., 2009). This protein shows 32 % sequence
homology with SACOL0085 protein and shows monomer architecture having similar
amino acids conserved (1Cys, 1Glut and 3His) as the active site residues as in case
of SACOL0085 when a multiple sequence alignment is made (Figure 5.1).
YxeP protein from Bacillus subtilus was another M20D family protein which showed
34 % sequence homology with SACOL0085 peptidase having a dimer quaternary
architecture. However, it remains less characterized till date to comment on it
functional significance. This protein also have similar amino acids conserved as the
active site residues as in case of SACOL0085 and 1XMB (Figure 5.1).
Another member of M20D family is HmrA from Staphylococcus aureus subsp.
aureus N315 having 25 % sequence similarity with SACOL0085 and is a tetramer
having 1Cys, 2 Glu and 3 His as active site residues. An additional Glu residue
(Glu128) other than the conserved Glu129 is found playing a role in catalysis and is
an exception in HmrA when compared to 1XMB and YxeP. The functional

Chapter 5
Page 108
characterization showed that this protein is an antibiotic resistance factor of
methicillin-resistant Staphylococcus aureus (MRSA) (Botelho et al., 2011). It is an
unspecific double-zinc endopeptidase which can hydrolyze peptide bonds in
extended peptides, unlike SACOL0085 peptidase, which acted primarily as a
dipeptidase.
The active site residues were found conserved in 1XMB, YxeP and HmrA proteins all
representing M20D family of metallopeptidases on carrying out a multiple sequence
alignment (Figure 5.1) using T-COFFEE online server Version-8.06 at the EBI
(Notredame et al., 2000). The same residues were found conserved in SACOL0085
as well. The metal-coordinating residues in the active site of 1XMB, YxeP and HmrA
include a conserved cysteine that clearly distinguishes this protein from previously
structurally characterized members of the M20 peptidase family. Importantly, Cys137
of ILL2, Cys98 of YxeP and Cys93 HmrA are all analogous to Cys103 of
SACOL0085 (Table 5.1). These Cys residues were found coordinating with both
metals in the active sites of 1XMB, YxeP and HmrA. The coordination of metals by
this bridging cysteine residue seems to be a unique feature of members of the
peptidase M20D family that clearly distinguishes these enzymes from other
structurally characterized enzymes from the M20 family.
The active site of 1XMB harbors an absolutely conserved glutamate (Glu173), which
is well positioned to act as a general acid base residue. Such a glutamate residue is
also found conserved among SACOL0085, HmrA and YxeP protein (Table 5.1) of
M20D family. Thus, it suggests that these enzymes likely uses a catalytic
mechanism that follows the paradigm established for the enzymes of the M20
peptidase family.
Cys103, Glu139, His105, His164 and His362 are the catalytic site residues expected
in SACOL0085 which are found to be conserved as active site residues in 1XMB,
YxeP and HmrA proteins, all belonging to M20D family having a bridging cysteine
residue coordinating with two metal ions at the catalytic site (Table 5.1).

Chapter 5
Page 109
Figure 5.1 Sequence features of SACOL0085. Multiple sequence alignment of the peptidase from Staphylococcus aureus COL0085; ILL2, an auxin-conjugate amidohydrolase from Arabidopsis thaliana (PDB id: 1XMB); YxeP, a dinuclear metal binding peptidase from Bacillus subtilis (PDB id: 1YSJ) and HmrA, an antibiotic resistance factor from Staphylococcus aureus subsp. aureus N315 (PDB id: 3RAM). Conserved active site residues in all the four sequences are depicted by boxes over the alignment. This alignment was obtained using the T-COFFEE online server Version-8.06 at the EBI (Notredame et al., 2000).
Chapter 5
Page 110
M20D family
Catalytic site residues
Cysteine Glutamate Histidine Histidine Histidine
SACOL0085 Cys103 Glu139 His105 His164 His362
1XMB Cys137 Glu173 His139 His197 His397
1YSJ Cys98 Glu134 His100 His158 His350
3RAM Cys93 Glu129 His95 His156 His341
Table 5.1 Catalytic site residues involved in metal binding. Active site residues conserved in Staphylococcus aureus COL0085 peptidase; ILL2, an auxin-conjugate amidohydrolase from Arabidopsis thaliana (PDB id: 1XMB); YxeP, a binuclear metal binding peptidase from Bacillus subtilis (PDB id: 1YSJ) and HmrA, an antibiotic resistance factor from Staphylococcus aureus subsp. aureus N315 (PDB id: 3RAM).
A B
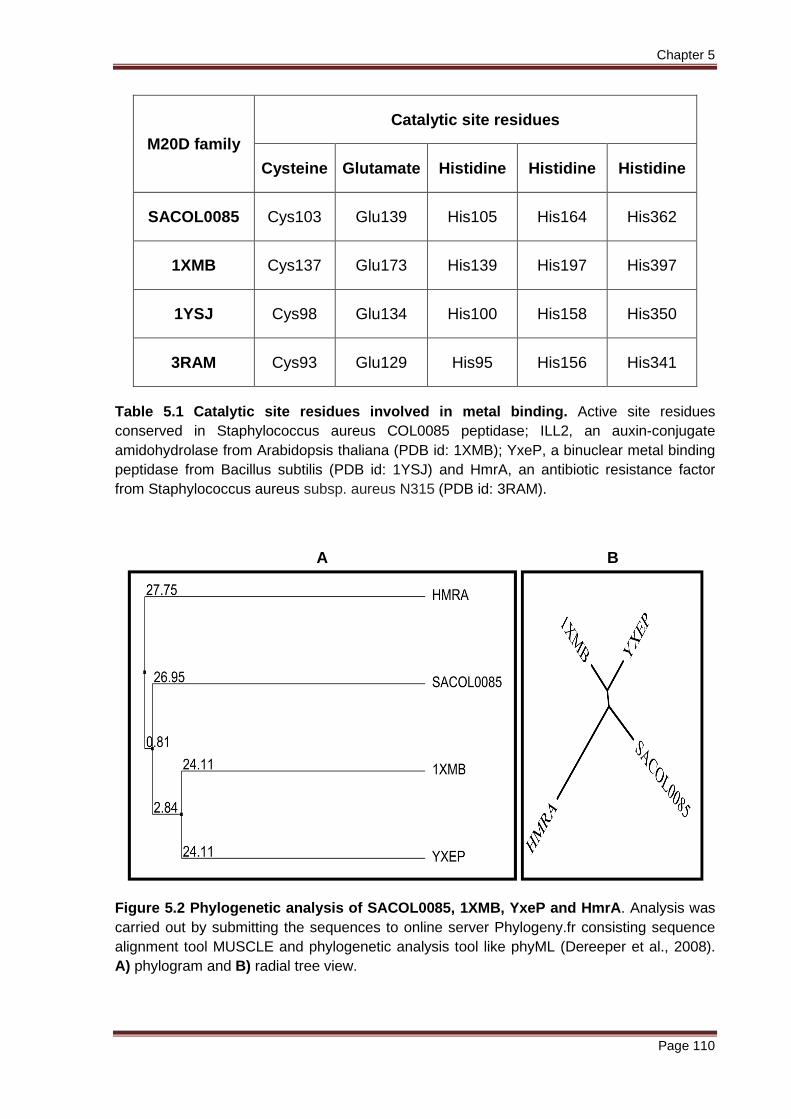
Figure 5.2 Phylogenetic analysis of SACOL0085, 1XMB, YxeP and HmrA. Analysis was carried out by submitting the sequences to online server Phylogeny.fr consisting sequence alignment tool MUSCLE and phylogenetic analysis tool like phyML (Dereeper et al., 2008). A) phylogram and B) radial tree view.

Chapter 5
Page 111
Phylogenetic analysis of the amino acid sequences of SACOL0085, 1XMB, YxeP
and HmrA proteins were found to synchronize with the multiple sequence alignment
data. Analysis were done by submitting the sequences to web server Phylogeny.fr
consisting of sequence alignment tool MUSCLE and phylogenetic analysis tool like
phyML (Dereeper et al., 2008). A tree view was obtained which showed 1XMB and
YxeP nodes (clade 1) having significant sequence conservation, since both were at
equal distance (24.11) from the branch node of SACOL0085 (clade 2). Hence, all
three are highly conserved. This is depicted in the phylogram and radial tree view
(Figure 5.2) and the branch length indicates diversion from the branch node. As the
length of the branch increases, the proteins become distantly related across the
species. However, phylogram and radial tree view data confirmed that sequence
wise these four proteins are conserved across species but they carry out different
functions.
It has been reported that mutations to His397 of ILL2, lead to a complete loss of the
enzymatic activity (Bitto et al., 2009). This observation is consistent with a crucial
role of this histidine in coordinating a metal ion in the active site. Structural similarity
was observed between ILL2 and the enzymes of the M20 peptidase family.
Moreover, the conservation of the key catalytic residue and the similar chemical
nature of substrates (e.g., being amides) of these enzymes suggest that they use a
catalytic mechanism similar to that proposed for the peptidases from M20 family.
Serine- and cysteine peptidase inhibitors exerted no relevant inhibitory activity in
case of HmrA protein. In contrast, it was efficiently inhibited by general zinc-chelating
agents such as EDTA, o-phenanthroline, tetra ethylene pentamine, as well as by
excess zinc (Botelho et al., 2011).
Analysis of cysteine residues and inhibitor studies is the objective of this chapter.
The activity of SACOL0085 peptidase will be analyzed by taking the conserved
active site cysteine residue into consideration. The regulatory mechanism of this
enzyme, if suggested, could potentially be exploited for the development of selective
inhibitors as in case of the protein tyrosine kinases (Schindler et al., 2000) and other
metal dependent peptidases.

Chapter 5
Page 112
5.1 Materials and Methods
5.1.1 Analysis of sulfhydryl groups in the SACOL008 5 protein by DTNB method
5 µM of the apo SACOL0085 protein was denatured by incubation with 7 M
Guanidium hydrochloride (GdnHCl) at room temperature for 2 hours as well as at 37 °C for 2 hours. The free-thiol content was determined by adding 1000-fold molar
excess of 5, 5΄- dithiobis-(2-nitrobenzoic acid) (DTNB or Ellman’s reagent) and
monitoring the absorbance at 412 nm. An extinction coefficient of 13,700 cm-1 M-1 for
the thio nitro benzoate (TNB2-) anion was used to calculate the number of free
sulfhydryl groups under denaturing conditions (Ellman, 1959; Butterworth et al.,
1967). The free-thiol content in the native condition of SACOL0085 protein (in the
absence of GnHCl) was also determined in a similar manner as in case with the
presence of GnHCl described above.
In order to determine the total number of sulfhydryl groups in the reduced
apoenzyme, 5 µM of apo-SACOL0085 protein was incubated with 10 mM of DTT in
ice for 3 hours as well as incubated for 2 hours at 37 °C and then passed through a
desalting column (20×1 cm Sephadex G10 column), prior to the reaction with
Ellman’s reagent. The assay was done both under denaturing condition (in the
presence of GnHCl) and in the native state (in the absence of GnHCl).
5.1.2 Inhibition studies
Carboxymethylation may be carried out without prior reduction to modify only those
cysteine residues that are not involved in disulfide bridges. If cysteine or cystine is
identified in a protein, it requires modification in order to be quantified. Thiol groups
may be blocked by a variety of reagents including mercuric chloride, iodoacetamide,
iodoacetic acid etc. For example, iodoacetate produces the S-carboxymethyl
derivative of cysteine, effectively introducing new negative charges into the protein.
Where such a charge difference is undesirable, iodoacetamide may be used to
derivatize cysteine to S-carboxyamidomethylcysteine (on acid hydrolysis, as for
amino acid analysis, this yields S-carboxymethylcysteine). The charge difference
between these two derivatives has been utilized in a method to quantify the number
of cysteine residues in a protein (Creighton, 1980; Aitken and Learmonth, 1980).

Chapter 5
Page 113
5.1.2.1 Inhibition studies with mercury
Mercuric ion and a number of its compounds are potential inhibitors for a wide
variety of cysteine proteases. This was traditionally considered to be one of the
typical toxicologic properties of heavy metals (Jiang et al., 2007). The enzyme
activity was first tested by performing thin layer chromatography (as described in
Chapter 3, materials and methods, section 3.1.3 ). Phe-Gly (2 mM), the best,
among the substrates tested; was incubated in an assay buffer containing 10 μg
(0.2256 μmol) SACOL0085 enzyme, pre-incubated with 1 mM MnCl2 and two
different conc. of HgCl2 (1 mM and 5 mM) was used.
The inhibitory effect of mercuric ion was also monitored by Cd-ninhydrin method by
setting up reactions in assay buffer containing 10 μg (0.2256 μmol) enzyme, pre-
incubated with 1 mM MnCl2 and different concentrations of HgCl2 varying from 10 μM
to 1000 μM at pH 8.0. Phe-Gly (5 mM) was added and then incubated for 1 hour
followed by the Cd-ninhydrin method.
5.1.2.2 Inhibition studies with iodoacetamide
The effect of cysteine residues on the catalytic properties of the enzyme was also
analyzed by modification with thiol-directed reagent iodoacetamide. The assay was
carried out in assay buffer containing 10 μg (0.2256 μmol) enzyme, pre-incubated
with 1 mM MnCl2 and different concentrations of iodoacetamide varying from 10 μM
to 1000 μM at pH 8.0. Phe-Gly (5 mM) was added and then incubated for 1 hour.
The reaction was monitored by Cd-ninhydrin method.
5.2 Results
5.2.1 Analysis of cysteine residues
There were three cysteine residues in the translated SACOL0085 protein sequence.
As there was no activity observed towards the available substrates in the absence of
divalent metal ions, it could be due to a masking of the catalytic site of the enzyme
as a result of disulfide bond formation between any two of the three Cys residues
present. A recent report also suggested that the activity of similar type of protein was

Chapter 5
Page 114
inhibited by a disulfide bond which stabilized its native form (Girish and Gopal,
2010). Hence, it was necessary to experimentally analyze the number of Cys
residues present in bound or free form.
The Ellman’s experiment using 5, 5’-dithio bis - (2-nitrobenzoic acid) (DTNB) labeling
assay showed no free thiol group under native conditions (in the absence of GnHCl).
Incubation of purified protein at 37 °C for 2 hours prior to the reaction with Ellman’s
reagent (in the absence of GnHCl), showed the presence of 1 free-thiol group.
Conversely, one free thiol group was observed in the presence of GnHCl without
prior incubation of protein as well. However, 3 free thiol groups were observed on
incubating the protein with GnHCl for 2 hours at 37 °C which indicates the influence
of temperature and GnHCl on unfolding the protein exposing all 3 free thiol groups
(Table 5.2).
The experiment with reduced protein (in the presence of 10 mM DTT) showed the
presence of only 1 free-thiol group under denaturing condition (in the presence of
GnHCl) (Table 5.2). However, even DTT cannot reduce buried (solvent-inaccessible)
disulfide bonds, so reduction of disulfide bonds is sometimes carried out under
denaturing conditions (e.g., at high temperatures, or in the presence of a strong
denaturant such as 6 M guanidinium hydrochloride, 8 M urea, or 1 % sodium
dodecylsulfate). When the protein was incubated with DTT at 37 °C for 2 hours,
under denaturing conditions, prior to the treatment with Ellman’s reagent, the
presence of all three free Cys residues in the protein was revealed. Also, under
denaturing condition, only one Cys residue was observed when incubated in ice for 3
hours even in the presence of DTT. These results indicate the combined influence of
temperature and denaturing agent (GnHCl) in unfolding the protein to expose all free
Cys groups in the protein. However, prevalence of a disulfide bond between any of
the three cysteine residues is not supported as results show the presence of three
free thiols both in the absence and presence of DTT. Hence, the apo-protein as such
has three cysteines which are not linked by disulfide bonds.

Chapter 5
Page 115
A. In the absence of DTT
Conditions Incubation condition prior to treatment with Ellman’s reagent
No. of free thiol groups observed
A) Under native condition (in the absence of GnHCl)
a) incubation at room temperature
for 2 hours
b) incubated at 37 °C for 2 hours
0.283 mols
1.060 mols
B) Under denaturing condition (in the presence of GnHCl)
a) incubation at room temperature
for 2 hours 1.046 mols
C) Under denaturing condition (in the presence of GnHCl)
a) incubated at 37 °C for 2 hours 3.030 mols
B. In the presence of DTT (reduced condition)
Conditions Incubation condition prior to
treatment with Ellman’s reagent
No. of free thiol groups observed
A) Under native condition (in the absence of GnHCl)
a) Protein incubated with DTT at 37 °C for 2 hours 1.004 mols
B) Under denaturing condition (in the presence of GnHCl)
a) Protein incubated with DTT in ice for 3 hours b) Protein incubated with DTT at 37 °C for 2 hours
1.007 mols
3.020 mols
Table 5.2 Analysis of cysteine residues by DTNB method. A) In the absence of reducing agent; B) In the presence of reducing agent. 5.2.2 SACOL0085 activity inhibited by cysteine inhi bitors
5.2.2.1 Mercuric ions acting as a potent inhibitor
Thin layer chromatography performed with the reaction mixture samples showed
Hg2+ acting as a potent inhibitor of the SACOL0085 protein and the experimental
results supported that Hg2+ might bind to the conserved active site cysteine residue
of the enzyme thus inhibiting its activity. However, the protein was found getting
precipitated with supplementation of 5 mM of Hg2+. Nevertheless, with 1 mM of Hg2+
no hydrolysis was monitored by thin layer chromatography (Figure 5.3). The extent to

Chapter 5
Page 116
which activity was inhibited was analyzed by carrying out the Cd- ninhydrin method
(Doi et al., 1981).
Figure 5.3 Monitoring the hydrolysis Phe-Gly by a TLC based assay. C corresponds to control. SI corresponds to assay with 10 μg of the enzyme in the presence of 1 mM MnCl2 where hydrolysis of the substrate was monitored as expected. S2 constitute the assay with similar conditions as before but with 1 mM HgCl2 included in the assay buffer and S3 shows the similar assay with 5 mM HgCl2 included in the assay buffer. Both in S2 and S3, activity ceased when Hg2+ ions were present showing their interference in catalytic activity. *Note: PHE corresponds to Phenylalanine (Std); C for control; S1, S2 and S3 are the reaction mixtures; GLY corresponds to Glycine (Std).
Conc. of HgCl 2 (μM)
Specific activity
(mmol/min/mg)
Percentage activity (%)
Control 2.814 100 10 0.7513 26.69 25 0.6488 23.05 50 0.6215 22.08 75 0.601 21.35
100 0.56 19.9 150 0.3825 13.59 250 0.3346 11.89 500 0.1843 6.55
1000 0.1638 5.825 Table 5.3 Peptidase activity of the SACOL0085 enzyme in the presence of Hg2+. The assay was carried out in assay buffer containing 10 μg (0.2256 μmol) enzyme, pre-incubated with 1 mM MnCl2 and different concentrations of HgCl2 varying from 10 μM to 1000 μM. Phe-Gly (5 mM) was added and then incubated for 1 hour. The reaction was monitored by Cd-ninhydrin method. A residual activity of 6 % (approx.) was seen even at 1 mM conc. of Hg2+ ions. However, it confirmed the decrease in activity with increasing concentration of the inhibitor. Relative activity was calculated.

Chapter 5
Page 117
The prevalence of a catalytic cysteine which favors hydrolysis of Phe-Gly dipeptide
by the SACOL0085 enzyme could be predicted since there was 94 % (approx.)
decrease in the activity with 1 mM of Hg2+ in comparison with the activity in its
absence (Table 5.3). Moreover, as the concentration of Hg2+ increased, the activity
decreased. However, a residual activity of about 6 % (approx.) was observed even at
1000 μM of Hg2+ (Table 5.3 and Figure 5.4).
5.2.2.2 Modification by iodoacetamide inhibited SAC OL0085 peptidase activity
Conc. of iodoacetamide
(μM) OD at 508 nm
Control 0.415 10 0.411 25 0.461 50 0.481 75 0.465
100 0.451 150 0.463 250 0.470 500 0.460 1000 0.459
Table 5.4 Assays with iodoacetamide. Assays done with 10 μg of protein pre-incubated with an assay buffer containing different concentration of iodoacetamide varying from 10 μM to 1000 μM. 1 mM MnCl2 is added along with Phe-Gly (5 mM) and incubated for 1 hour followed by Cd-ninhydrin assay.
Iodoacetamide which also alkylates the thiol groups of cysteine residues was tested
to monitor inhibition of enzyme of interest. The activity of SACOL0085 peptidase was
not inhibited even at concentrations of 1 mM of iodoacetamide (Table 5.4). It seems
like the protein is not getting labeled with iodoacetamide. This could be accounted
for the catalytic cysteine not getting exposed to alkylation by iodoacetamide.
Therefore, if accessibility is the reason, the assays could be carried out with
guanidium hydrochloride (GnHCl) since it was found to expose all three cysteine
residues while performing DTNB assay (described in section 5.2.1 of this chapter).
Hence, the assays need to be standardized in the presence of GnHCl as well.

Chapter 5
Page 118
5.2.2.2.1 Assay with guanidium HCl
Conc. of GnHCl (mM) OD at 508 nm
Control 0.421 0.010 0.688 0.050 0.824 0.1 0.882 0.5 0.803 1 0.768
10 0.676 50 0.496
100 0.341 250 0.285 500 0.193 1000 0.170 2000 0.138 3000 0.098 4000 0.068 5000 0.042 6000 0.048
Table 5.5 Assays with guanidium HCl. Assays done with 10 μg of protein pre-incubated with an assay buffer containing different concentration of GnHCl varying from 0.010 mM to 6000 mM. 1 mM MnCl2 is added along with Phe-Gly (5 mM) and incubated followed by Cd-ninhydrin assay. Guanidium HCl at a concentration of 6 M exposed all three free cysteine residues in
the protein as seen in DTNB assay (described in section 5.2.1 of this chapter).
Hence, the assay was performed with different concentrations of GnHCl varying from
0.010 mM to 6000 mM. The results showed maximum activity at a concentration of
0.1 mM in comparison with the activity in its absence (2 fold increase). However,
decrease in activity was observed thereafter with increasing concentration of GnHCl
(Table 5.5) which was possibly due to the denaturation of enzyme.
Consequently the assays were carried out in the presence of 0.1 mM of GnHCl to
enhance the opening up of the SACOL0085 protein exposing the catalytic cysteine
favoring alkylation by iodoacetamide.
However, results showed no inhibition of activity with the modifications employed.
The reason for such behavior was unknown at this point of time and further attempts

Chapter 5
Page 119
were made for confirming whether iodoacetamide is a potent inhibitor of the protein
or not.
In order to confirm the inability of iodoacetamide in alkylating the protein, fresh salt of
iodoacetamide was purchased (Sigma-Aldrich Fine Chemicals, St.Louis, MO, USA)
and used. However, to our surprise the experiment showed inhibitory results
showing a decrease of 74 % activity when compared in its absence (Table 5.6 and
Figure 5.4). Hence, iodoacetamide also proved to be a potent inhibitor of
SACOL0085 protein confirming the presence of a catalytic cysteine favoring
hydrolysis. So the quality of iodoacetamide salt used previously gave us artifacts.
Conc. of
iodoacetamide
(μM)
Specific activity (mmol/min/mg)
Percentage activity (%)
Control 2.818 100
10 2.178 75.23
25 1.960 67.68
50 1.728 59.66
75 1.584 54.71
100 1.434 49.52
150 1.168 40.33
250 0.9493 32.78
500 0.84 29
1000 0.7786 26.88
1500 0.6556 22.64
Table 5.6 Peptidase activity of the enzyme in the presence of iodoacetamide. The assay was carried out in assay buffer containing 10 μg (0.2256 μmol) enzyme, pre-incubated with 1 mM MnCl2 and different concentrations of iodoacetamide varying from 10 μM to 1500 μM. Phe-Gly (5 mM) was added and then incubated for 1 hour. The reaction was monitored by Cd-ninhydrin method. A residual activity of 22 % was seen even at 1.5 mM conc. of iodoacetamide. However, it confirms the activity decreasing with increasing concentration of the inhibitor. Such direct involvement of cysteine residues in the catalytic mechanism has been
reported previously in such group of metallopeptidases. For example, ACY1
inhibition by thiol specific reagents which also belong to M20 family of
metallopeptidases (Loffler et al., 1987, Heese et al., 1989, Wang et al., 1995, Yang
Chapter 5
Page 120
et al., 1996; Bitto et al., 2009). The catalytic site can undergo a conformational
change by substrate-binding (Lindner et al., 2003), metal coordination or by
modification with inhibitors. Such rearrangements can bring changes in the catalytic
site residues in the vicinity of the active site, thus, limiting the hydrolysis of Phe-Gly.
The structural modification induced by the mercuric ions and iodoacetamide may
probably allow the occurrence of this event, resulting in the inactivation of the
enzyme.
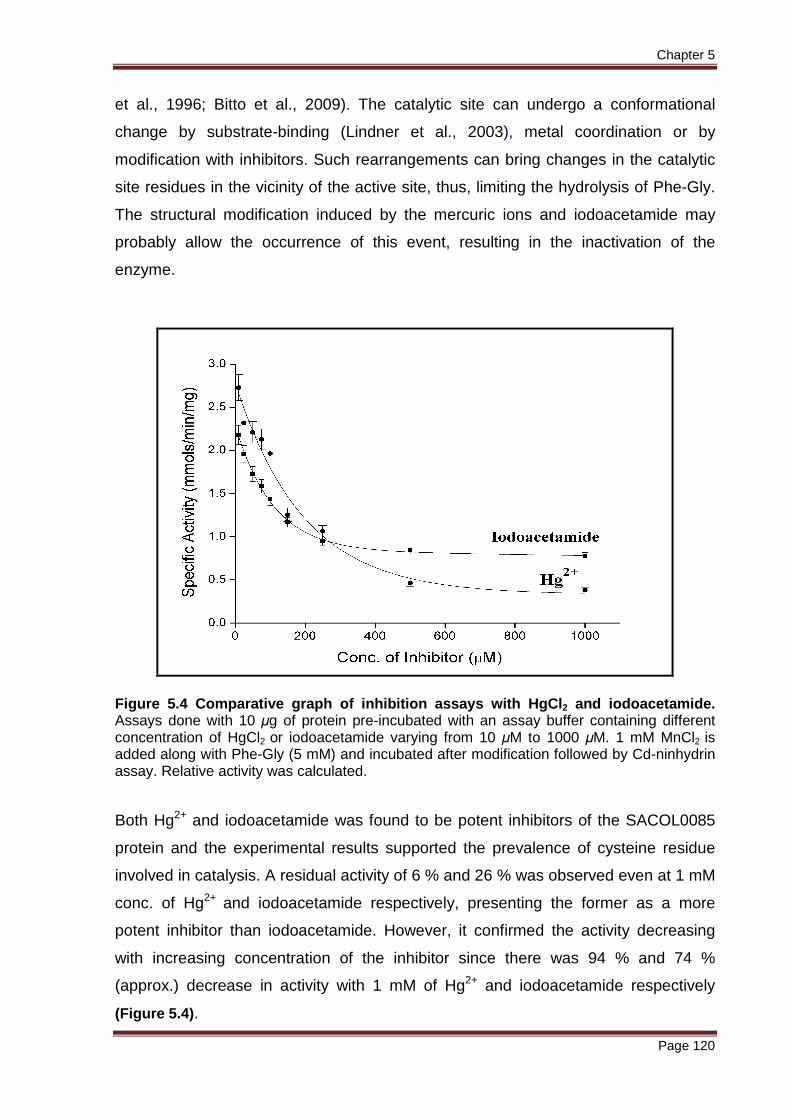
Figure 5.4 Comparative graph of inhibition assays with HgCl2 and iodoacetamide. Assays done with 10 μg of protein pre-incubated with an assay buffer containing different concentration of HgCl2 or iodoacetamide varying from 10 μM to 1000 μM. 1 mM MnCl2 is added along with Phe-Gly (5 mM) and incubated after modification followed by Cd-ninhydrin assay. Relative activity was calculated.
Both Hg2+ and iodoacetamide was found to be potent inhibitors of the SACOL0085
protein and the experimental results supported the prevalence of cysteine residue
involved in catalysis. A residual activity of 6 % and 26 % was observed even at 1 mM
conc. of Hg2+ and iodoacetamide respectively, presenting the former as a more
potent inhibitor than iodoacetamide. However, it confirmed the activity decreasing
with increasing concentration of the inhibitor since there was 94 % and 74 %
(approx.) decrease in activity with 1 mM of Hg2+ and iodoacetamide respectively
(Figure 5.4).

Chapter 5
Page 121
5.3 Discussion
The DTNB experiments showed that the thiol groups of the native SACOL0085
protein were not fully accessible since the titre values increased slowly with time and
temperature such that a clear end point could not be reached. However, the
comparison of experiments with or without reducing agent showed that the
SACOL0085 protein did not have any linked thiol groups and all the three cysteines
were in unbound state. The results confirmed the combined influence of temperature
and denaturing agent (GnHCl) in unfolding the protein to expose all the free Cys
groups in the protein. Hence, the prevalence of a disulfide bond between any of the
three cysteine residues is not supported as results show the presence of three free
thiols both in the absence or presence of DTT. The studies with TCEP also indicated
that the Cys residues were in the reduced form (concluded in chapter 4). Hence, the
apo-protein as such has three cysteines which are not linked by disulfide bonds.
Therefore, Ellman’s procedure confirmed the presence of three Cys residues in
reduced form. The closest sequence homologues of SACOL0085 are- ILL2, an
auxin-conjugate amidohydrolase from Arabidopsis thaliana, an aminopeptidase
YxeP from Bacillus subtilis and an antibiotic resistance factor HmrA from
Staphylococcus aureus subsp. aureus N315 all belonging to M20D family having a
bridging cysteine residue coordinating with the metal ions at the catalytic site.
Although ILL2, YxeP and HmrA show significant sequence homology (34 %, 32 %
and 25 % respectively) with SACOL0085, it is intriguing to note that they however,
do not show same functional aspect as that of SACOL0085. Nevertheless, this
protein could have the same structural fold at the active sites as shown by the
multiple sequence alignment. The amino acids involved in metal binding are
conserved in all the four proteins and the geometric orientations of the ligands and
metal ions in the active site could possibly the same. Hence, such a cysteine residue
could form a key catalytic residue in catalytic mechanism in this protein as well.
Confirmation came when 94 % and 74 % decrease in activity was observed with thiol
directed reagents like Hg2+ and iodoacetamide at 1 mM concentration. Such loss of
activity could be probably because of conformational change in the 3D structure of
the enzyme induced by modification of cysteine at the active site. There are
instances reported before in similar type proteins wherein conformational changes in
the catalytic site are induced by substrate-binding or by metal coordination (Heese et

Chapter 5
Page 122
al., 1989; Jiang et al., 2007). Such rearrangement can bring the catalytic site
residues in the vicinity of the active site, thus, favoring hydrolysis of Phe-Gly. The
structural modification induced by such inhibitors may probably prevent the
occurrence of this event, resulting in the inactivation of the enzyme. These studies
illustrate the contribution of any of the three cysteine residues to the functional
stability of the protein under study or the direct involvement of cysteine residues in
the catalytic machinery of the enzyme. These cysteine residues (probably Cys103
shown as conserved residue in multiple sequence alignment) could be a part of the
active site of the enzyme aiding in the acid base mechanism for hydrolysis of the
specific substrate when bound. The direct involvement of cysteine residues in
catalytic machinery has been a topic of intense debate. This discussion originated
from the observation of ACY1 inhibition by thiol specific reagents which also belong
to M20 family of metallopeptidases (Loffler et al., 1987; Heese et al., 1989; Wang et
al., 1995; Wisena et al., 2004; Herga et al., 2005; Sommaruga et al., 2008; Chang et
al., 2010). Moreover, site-directed mutagenesis experiments could possibly identify
any functionally important cysteine residue being located near one of the metal
coordinating residues, which play a critical role in the enzymatic activity. We intend
to explore more regarding this protein by trying to crystallize it and get a 3-
dimensional structure which could further be utilized for the design of inhibitors that
specifically target the organism’s biosynthesis route and counteract the infectious
state. This will be discussed in the next chapter.

















