Characterization of Listeria monocytogenes isolated from Ganges water, human clinical and milk samples at Varanasi, India Dharmendra K. Soni a , Rakesh K. Singh b , Durg V. Singh b , Suresh K. Dubey a,⇑ a Environmental Microbiology Laboratory, Department of Botany, Banaras Hindu University, Varanasi, India b Infectious Disease Biology, Institute of Life Sciences, Bhubaneswar, India article info Article history: Received 21 May 2012 Received in revised form 22 September 2012 Accepted 24 September 2012 Available online 28 November 2012 Keywords: Listeria monocytogenes Serotype identification ERIC- and REP-PCR Virulence genes abstract Listeria monocytogenes isolated from Ganges water, human clinical and milk samples were characterized by antibiotic susceptibility, serotype identification, detection of virulence genes and ERIC- and REP-PCR fingerprint analyses. All isolates were uniformly resistant to ampicillin, except two isolates, and showed variable resistance to gentamicin, cotrimoxazole, ofloxacin, rifampicin and tetracycline. Of the 20 isolates found positive for pathogens, seven (four human and three water isolates) belong to serogroups 4b, 4d and 4e; six (one human and five water isolates) belong to serogroups 1/2c and 3c; four milk isolates belong to serogroups 1/2b and 3b; and three milk isolates belong to serogroups 1/2a and 3a. Two water isolates, all human isolates, except one (Pb1) lacking inlJ gene, and three milk isolates possess inlA, inlC, plcA, prfA, actA, hlyA and iap genes. The remaining water and milk isolates showed variable presence of inlJ, plcA, prfA, and iap genes. ERIC- and REP-PCR based analyses collectively indicated that isolates of human clinical samples belong to identical or similar clone and isolates of water and milk samples belong to different clones. Overall study demonstrates the prevalence of pathogenic L. monocytogenes species in the environmental and clinical samples. Most of the isolates were resistant to commonly used antibiotics. Ó 2012 Elsevier B.V. All rights reserved. 1. Introduction Human listeriosis, a public health problem, has been reported in the industrialized nations (Poulsen et al., 2011). Pregnant women, neonates, elderly, or immunocompromised people are particularly susceptible to Listeria which manifests as abortion, stillbirth, septi- cemia, meningitis and meningoencephalitis (WHO, 2004). The genus Listeria, a Gram-positive bacteria, comprised of eight species Listeria monocytogenes, Listeria seeligeri, Listeria ivanovii, Listeria innocua, Listeria welshimeri, Listeria grayi, Listeria marthii and Liste- ria rocourtiae (Graves et al., 2010; Leclercq et al., 2010) of which L. monocytogenes and L. ivanovii, are considered virulent (Guillet et al., 2010). L. ivanovii is rarely associated with human infection (Cummins et al., 1994). L. monocytogenes are usually susceptible to wide range of anti- biotics, the resistance to several antibiotics has been reported by several workers (Carpentier and Courvalin, 1999; Nwachukwu et al., 2010). Drug-sensitive strains of L. monocytogenes were iso- lated from clinical and food samples by Dhanashree et al. (2003). Sharma et al. (2012) reported multidrug-resistance strains from milk samples. A number of virulence factors such as internalins (encoded by inlA, inlC, inlJ), listeriolysin O (LLO encoded by hlyA), actin (actA), phosphatidyl-inositol-phospholipase C (PI-PLC en- coded by plcA), iap (invasion associated protein encoded by iap) and virulence regulator (encoded by prfA) have been reported for their important role in the virulence and pathogenicity of L. mono- cytogenes (Vazquez-Boland et al., 2001; Liu et al., 2007). Serotyping has been used to characterize L. monocytogenes that differs in their virulence and pathogenicity (Doumith et al., 2004; Liu, 2006). Although 13 serotypes of L. monocytogenes have been reported, only three serotypes (1/2a, 1/2b and 4b) are frequently isolated from the clinical cases of which 1/2a is mostly isolated from food and 4b from human epidemics (Liu, 2006). All strains isolated from human clinical, food and environment belong to a small number of serotypes (Farber and Peterkin, 1991; Chen et al., 2010). Therefore, highly discriminatory typing methods that correlate with serotyping are necessary. Among various ap- proaches for molecular typing of L. monocytogenes, PFGE (pulsed field gel electrophoresis) has been considered to be the ‘‘gold stan- dard’’ due to its high reproducibility and discriminatory ability (Swaminathan et al., 2001; Gerner-Smidt et al., 2006). However, it is labor intensive and time consuming method. REP (repetitive element sequence) and ERIC (enterobacterial repetitive intergenic consensus) – PCR based approaches have been found to be rela- tively simple and cost-effective for the genus Listeria and have shown to generate DNA fingerprints comparable to PFGE that allow discrimination within a single bacterial species (Jersek et al., 1999; 1567-1348/$ - see front matter Ó 2012 Elsevier B.V. All rights reserved. http://dx.doi.org/10.1016/j.meegid.2012.09.019 ⇑ Corresponding author. Tel.: +91 542 2307147; fax: +91 542 2368174. E-mail address: [email protected](S.K. Dubey). Infection, Genetics and Evolution 14 (2013) 83–91 Contents lists available at SciVerse ScienceDirect Infection, Genetics and Evolution journal homepage: www.elsevier.com/locate/meegid
Transcript
Infection, Genetics and Evolution 14 (2013) 83–91
Contents lists available at SciVerse ScienceDirect
Characterization of Listeria monocytogenes isolated from Ganges water, humanclinical and milk samples at Varanasi, India
Dharmendra K. Soni a, Rakesh K. Singh b, Durg V. Singh b, Suresh K. Dubey a,⇑a Environmental Microbiology Laboratory, Department of Botany, Banaras Hindu University, Varanasi, Indiab Infectious Disease Biology, Institute of Life Sciences, Bhubaneswar, India
a r t i c l e i n f o a b s t r a c t
Article history:Received 21 May 2012Received in revised form 22 September2012Accepted 24 September 2012Available online 28 November 2012
Keywords:Listeria monocytogenesSerotype identificationERIC- and REP-PCRVirulence genes
1567-1348/$ - see front matter � 2012 Elsevier B.V. Ahttp://dx.doi.org/10.1016/j.meegid.2012.09.019
Listeria monocytogenes isolated from Ganges water, human clinical and milk samples were characterizedby antibiotic susceptibility, serotype identification, detection of virulence genes and ERIC- and REP-PCRfingerprint analyses. All isolates were uniformly resistant to ampicillin, except two isolates, and showedvariable resistance to gentamicin, cotrimoxazole, ofloxacin, rifampicin and tetracycline. Of the 20 isolatesfound positive for pathogens, seven (four human and three water isolates) belong to serogroups 4b, 4dand 4e; six (one human and five water isolates) belong to serogroups 1/2c and 3c; four milk isolatesbelong to serogroups 1/2b and 3b; and three milk isolates belong to serogroups 1/2a and 3a. Two waterisolates, all human isolates, except one (Pb1) lacking inlJ gene, and three milk isolates possess inlA, inlC,plcA, prfA, actA, hlyA and iap genes. The remaining water and milk isolates showed variable presence ofinlJ, plcA, prfA, and iap genes. ERIC- and REP-PCR based analyses collectively indicated that isolates ofhuman clinical samples belong to identical or similar clone and isolates of water and milk samples belongto different clones. Overall study demonstrates the prevalence of pathogenic L. monocytogenes species inthe environmental and clinical samples. Most of the isolates were resistant to commonly used antibiotics.
� 2012 Elsevier B.V. All rights reserved.
1. Introduction
Human listeriosis, a public health problem, has been reported inthe industrialized nations (Poulsen et al., 2011). Pregnant women,neonates, elderly, or immunocompromised people are particularlysusceptible to Listeria which manifests as abortion, stillbirth, septi-cemia, meningitis and meningoencephalitis (WHO, 2004). Thegenus Listeria, a Gram-positive bacteria, comprised of eight speciesListeria monocytogenes, Listeria seeligeri, Listeria ivanovii, Listeriainnocua, Listeria welshimeri, Listeria grayi, Listeria marthii and Liste-ria rocourtiae (Graves et al., 2010; Leclercq et al., 2010) of which L.monocytogenes and L. ivanovii, are considered virulent (Guilletet al., 2010). L. ivanovii is rarely associated with human infection(Cummins et al., 1994).
L. monocytogenes are usually susceptible to wide range of anti-biotics, the resistance to several antibiotics has been reported byseveral workers (Carpentier and Courvalin, 1999; Nwachukwuet al., 2010). Drug-sensitive strains of L. monocytogenes were iso-lated from clinical and food samples by Dhanashree et al. (2003).Sharma et al. (2012) reported multidrug-resistance strains frommilk samples. A number of virulence factors such as internalins(encoded by inlA, inlC, inlJ), listeriolysin O (LLO encoded by hlyA),
ll rights reserved.
: +91 542 2368174.ey).
actin (actA), phosphatidyl-inositol-phospholipase C (PI-PLC en-coded by plcA), iap (invasion associated protein encoded by iap)and virulence regulator (encoded by prfA) have been reported fortheir important role in the virulence and pathogenicity of L. mono-cytogenes (Vazquez-Boland et al., 2001; Liu et al., 2007).
Serotyping has been used to characterize L. monocytogenes thatdiffers in their virulence and pathogenicity (Doumith et al., 2004;Liu, 2006). Although 13 serotypes of L. monocytogenes have beenreported, only three serotypes (1/2a, 1/2b and 4b) are frequentlyisolated from the clinical cases of which 1/2a is mostly isolatedfrom food and 4b from human epidemics (Liu, 2006). All strainsisolated from human clinical, food and environment belong to asmall number of serotypes (Farber and Peterkin, 1991; Chenet al., 2010). Therefore, highly discriminatory typing methods thatcorrelate with serotyping are necessary. Among various ap-proaches for molecular typing of L. monocytogenes, PFGE (pulsedfield gel electrophoresis) has been considered to be the ‘‘gold stan-dard’’ due to its high reproducibility and discriminatory ability(Swaminathan et al., 2001; Gerner-Smidt et al., 2006). However,it is labor intensive and time consuming method. REP (repetitiveelement sequence) and ERIC (enterobacterial repetitive intergenicconsensus) – PCR based approaches have been found to be rela-tively simple and cost-effective for the genus Listeria and haveshown to generate DNA fingerprints comparable to PFGE that allowdiscrimination within a single bacterial species (Jersek et al., 1999;
84 D.K. Soni et al. / Infection, Genetics and Evolution 14 (2013) 83–91
Chen et al., 2010). REP and ERIC sequences act as primer bindingsites to amplify the genome of a variety of bacteria by PCR (Woodset al., 1993).
L. monocytogenes is ubiquitous and widely distributed in theenvironment. It has been isolated from a variety of sources, includ-ing soil, vegetation, food of animal origin, such as meat and dairyproducts, silage, fecal material, sewage and water (Orndorff et al.,2006; Gandhi and Chikindas, 2007). The incidence of listeriosismay varies between 0.1 and 11.3 per million in different countries(Anon., 2004). However, no such type of evidence is available in In-dia and the epidemiological data available till date are not ade-quate for assessing the extent of infection in human beings(Barbuddhe et al., 2012). Only a few reports on the incidence ofL. monocytogenes have been documented for food (Aurora et al.,2008; Kalorey et al., 2008) and clinical samples (Dhanashreeet al., 2003; Kaur et al., 2007) from India. However, to the best ofour knowledge no published information on the prevalence of L.monocytogenes in fresh water bodies like lakes, rivers etc., of Indiaare available. The river Ganges, the most sacred river of India, isone of the ten large rivers of the World. In Varanasi it travels some7 km distance with a half-moon curvature between its two tribu-taries, Assi in the south and Varuna in the north. Around 60,000people take holy dip in the river daily and almost all of them sipthe water. Also, out of the total potable water demand of 220 mil-lion liters per day (MLD), river Ganges serves as the source fornearly 110 MLD of raw water drawn from its Bhadaini ghat waterintake (Mishra, 2005). Therefore, studies on river Ganges with ref-erence to a pathogen like L. monocytogenes is imperative as a majorpart of Indian population depends on river Ganges for bathing,drinking and sustenance.
Considering the above facts, in this study, we characterized L.monocytogenes isolated from the river Ganges water, human clini-cal and milk samples for the presence of virulence genes and theirresponse to antibiotic susceptibility. We also used ERIC- and REP-PCR techniques to generate DNA fingerprints of L. monocytogenesisolated from various sources and to see clonal relationshipsamong them.
2. Materials and methods
2.1. Study site and sample collection
A total of 712 samples were collected from Varanasi, UttarPradesh, India, during June 2009–July 2010 and tested, of which100 were water samples from the river Ganges, 300 samples werefrom humans and 312 samples were from milk and milk products.The Ganges water samples were collected ten meters away fromthe bank of the river and one meter deep at around 9.00 am fromfour sites along ghats (Ravidas ghat, Assi ghat, Bhadaini ghat andDr. Rajendra Prasad ghat) of Varanasi (25�200N and 83�E), India(Fig. 1). The ghat is a flight of stones steps originating from citylanes and reaching up to the river water front. Samples were alsocollected 100 m upstream from the reference point. Taking the out-fall of Assi drain as the reference point, Ravidas ghat, Assi ghat,Bhadaini ghat and Dr. Rajendra Prasad ghat (Dr. R. P. ghat) arearound 1 m, 650 m, 1.3 km and 2.6 km downstreams. All humanclinical samples were collected from private and government hos-pitals, pregnant women with bad obstetric history like repeatedabortion, still births and pre-term labor, of which 75 each werefrom vaginal swabs, cervical swabs, placental tissues and blood,respectively. Out of the 168 milk samples, 120 were from cow milkand 48 from pasteurized milk and out of the 144 milk products, 48each were from cheese, butter and ice-cream, respectively. All thesamples were collected aseptically, quickly transported to the lab-oratory under chilled condition and were processed within 24 h ofcollection.
2.2. River water quality measurements
The parameters related to water quality of river Ganges such aspH, temperature, turbidity, dissolved oxygen (DO), biological oxygendemand (BOD) and fecal coliform count (FCC) are regularly deter-mined by Swatcha Ganga Research Laboratory (SGRL), Varanasi,India, as per the standard protocol of American Public HealthAssociation (APHA, 1992). The data obtained by SGRL during June2009–July 2010 have been used for ascertaining the pollution levelin the river water at Varanasi which is primarily due to dischargeof domestic sewage.
2.3. Isolation and identification of L. monocytogenes
L. monocytogenes were isolated from the samples following thestandard double enrichment method as described by ISO 11290:1with slight modifications (Anon., 1997). Briefly, 25 ml of water,15 ml of milk, 5 g or ml of milk product, placental bit and blood,and cervical or vaginal swab were separately inoculated into 225,135, 45 and 10 ml of half-Fraser broth (Difco, USA), respectivelyand incubated for 24 h at 30 �C. Second enrichment was done byadding 0.1 ml from the overnight grown culture into 10 ml of fullconcentration of selective agents (Fraser broth, Difco, USA) andincubated for 48 h at 37 �C with subsequent spreading on PALCAMagar (Difco, USA) and incubated again for 48 h at 37 �C. Gray-greenish colonies with black sunken center and black halo werepicked up and confirmed by Gram staining, biochemical tests suchas catalase test, methyl red–Voges–Proskauer (MR–VP) reaction,nitrate reduction and motility at 20–25 �C, acid production fromrhamnose, xylose, mannitol, a-methyl-D-mannopyranoside andCAMP test with Staphylococcus aureus and Rhodococcus equi (See-liger and Jones, 1986). L. monocytogenes strain MTCC1143, S. aureusstrain MTCC1144 and R. equi strain MTCC1135 were used as con-trols. All the L. monocytogenes isolates and control strains were pre-served in tryptic soy agar slants at room temperature for use asworking stock in the laboratory.
2.4. Antibiotic susceptibility test
All L. monocytogenes isolates were tested for antimicrobial sus-ceptibility for 10 commonly used antibiotics in veterinary and hu-man therapy (Lyon et al., 2008; Nwachukwu et al., 2010), using thedisc diffusion method of Bauer et al. (1966). Antibiotics discs (Ox-oid, UK) with the following concentrations were used: ampicillin(A, 10 lg), chloramphenicol (C, 30 lg), ciprofloxacin (Cf, 5 lg),cefoxitin (Fox, 30 lg), co-trimoxazole (SXT, 25 lg), gentamicin(G, 10 lg), oflaxacin (Of, 5 lg), rifampicin (R, 5 lg), streptomycin(S, 10 lg) and tetracycline (T, 30 lg). The diameter of the zone ofclearance was recorded and interpreted following the guidelineof the Clinical and Laboratory Standards Institute (CLSI, 2006) forGram-positive bacteria.
2.5. DNA isolation
Chromosomal DNA was extracted from L. monocytogenes iso-lates grown overnight at 37 �C with shaking at 200 oscillationsper min in brain heart infusion broth (BHIB, Difco, USA) using themethod of Moon et al. (2007) with modification. Bacterial cellsfrom 10 ml of culture were pelleted by centrifugation at8000 rpm at 4 �C for 15 min, washed three times in normal salinesolution (0.9%), suspended in 500 ll of lysozyme solution[10 mM Tris–HCl (pH 8), 2.5 mg lysozyme (Sigma) per ml] and5 ll of lysostaphin (100 lg/ll; Sigma) and incubated for 2 h at37 �C. Thereafter 250 ll of Tris–EDTA buffer [50 mM Tris–HCl(pH 8) and 100 mM EDTA (Sigma)], 100 ll of SDS [1% (w/v)],50 ll of proteinase K solution (1 mg/ml; Sigma) were added and
Fig. 1. Location of water sampling sites along the river Ganges at Varanasi, India.
D.K. Soni et al. / Infection, Genetics and Evolution 14 (2013) 83–91 85
the mixture again was incubated for 2 h at 50 �C. Cell lysates werethen extracted once with phenol: chloroform (1:1) mixture andtwice with chloroform: isoamyl alcohol (12:1) mixture after incu-bating the first extract with RNase (10 mg/ml; Sigma) for 2 h at37 �C. The precipitation of DNA was carried out with NaCl (5 M)and 2 vol of chilled ethanol (100%) with incubation at �20 �C forovernight. The DNA was dissolved in sterile MQ water and the con-centration and purity of the DNA was determined in Eppendorfspectrophotometer by using automatic calculation of the ratio ofoptical densities at 260 and 280 nm.
2.6. Species and serogroup identification
The presence of the internalin gene (inlA) was determined byPCR to identify L. monocytogenes as described by Liu et al. (2007)and serogroup (1/2a, 1/2b, 1/2c, and 4b) in L. monocytogenes wasdetermined by multiplex PCR described by Doumith et al. (2004).Amplification was performed in a 0.2 ll tube containing 10� PCRbuffer (100 mM Tris (pH 9.0), 500 mM KCl, 15 mM MgCl2, 0.1% gel-atin) (Bangalore Genei), 2 mM MgCl2, 0.3 mM dNTPs, forward andreverse primers of lmo0737, ORF2819, and ORF2110 (1 lM);lmo1118 (1.5 lM); and prs (0.2 lM), 2 U of Taq DNA polymerase(Bangalore Genei) and 3 ll of DNA template (50 ng/ll). PCR wasperformed with an initial denaturation at 94 �C for 5 min, 35 cyclesin sequence 94 �C for 30 s, 54 �C for 75 s and 72 �C for 75 s, and fi-nal extension at 72 �C for 10 min in a thermal cycler (MyCyler™Thermal Cycler, Bio-Rad Laboratories, Inc, Australia). The PCR prod-ucts were analyzed by 1.5% agarose gel electrophoresis, stained
with ethidium bromide, visualized by UV transilluminator andphotographed.
2.7. Detection of virulence genes
The presence of internalin genes inlA, inlC and inlJ was deter-mined by multiplex PCR described by Liu et al. (2007). PCR wasperformed in a reaction volume of 25 ll containing 10� PCR buffer(100 mM Tris (pH 9.0), 500 mM KCl, 15 mM MgCl2, 0.1% gelatin)(Bangalore Genei), 2 mM MgCl2, 0.2 mM dNTPs, 40 pmol of inlA,30 pmol of inlC and 20 pmol of inlJ, 1 U of Taq DNA polymerase(Bangalore Genei) and 3 ll of DNA template (50 ng/ll) under thefollowing cyclic conditions: initial denaturation at 94 �C for2 min, 30 cycles in sequence 94 �C for 20 s, 55 �C for 20 s and72 �C for 50 s, and final extension at 72 �C for 2 min. The multiplexPCR technique described by Notermans et al. (1991) and subse-quently modified by Kaur et al. (2007) was also used to detect plcA,hlyA, actA, iap and prfA genes. PCR was performed in 25 ll reactionmixture consisting of 10� PCR buffer (Bangalore Genei), 2 mMMgCl2, 0.2 mM dNTPs, 0.1 lM of forward and reverse primers ofeach genes, 1.5 U Taq DNA polymerase (Bangalore Genei) and3 ll of DNA template (50 ng/ll) under the following cyclic condi-tions: initial denaturation at 95 �C for 2 min, 35 cycles in sequence95 �C for 15 s, 60 �C for 30 s and 72 �C for 90 s, and final extensionat 72 �C for 10 min. The PCR products were analyzed by 1.5%agarose gel electrophoresis, stained with ethidium bromide, visu-alized by UV transilluminator and photographed. The details of
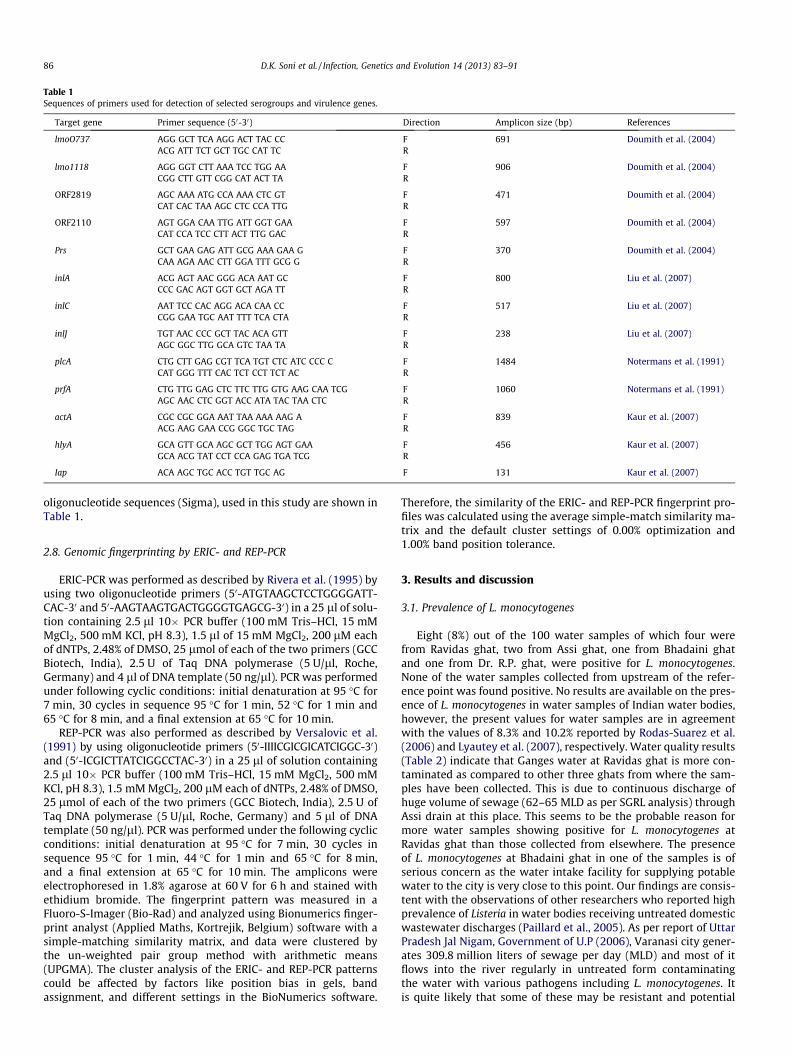
Table 1Sequences of primers used for detection of selected serogroups and virulence genes.
Target gene Primer sequence (50-30) Direction Amplicon size (bp) References
lmoO737 AGG GCT TCA AGG ACT TAC CC F 691 Doumith et al. (2004)ACG ATT TCT GCT TGC CAT TC R
lmo1118 AGG GGT CTT AAA TCC TGG AA F 906 Doumith et al. (2004)CGG CTT GTT CGG CAT ACT TA R
ORF2819 AGC AAA ATG CCA AAA CTC GT F 471 Doumith et al. (2004)CAT CAC TAA AGC CTC CCA TTG R
ORF2110 AGT GGA CAA TTG ATT GGT GAA F 597 Doumith et al. (2004)CAT CCA TCC CTT ACT TTG GAC R
Prs GCT GAA GAG ATT GCG AAA GAA G F 370 Doumith et al. (2004)CAA AGA AAC CTT GGA TTT GCG G R
inlA ACG AGT AAC GGG ACA AAT GC F 800 Liu et al. (2007)CCC GAC AGT GGT GCT AGA TT R
inlC AAT TCC CAC AGG ACA CAA CC F 517 Liu et al. (2007)CGG GAA TGC AAT TTT TCA CTA R
inlJ TGT AAC CCC GCT TAC ACA GTT F 238 Liu et al. (2007)AGC GGC TTG GCA GTC TAA TA R
plcA CTG CTT GAG CGT TCA TGT CTC ATC CCC C F 1484 Notermans et al. (1991)CAT GGG TTT CAC TCT CCT TCT AC R
prfA CTG TTG GAG CTC TTC TTG GTG AAG CAA TCG F 1060 Notermans et al. (1991)AGC AAC CTC GGT ACC ATA TAC TAA CTC R
actA CGC CGC GGA AAT TAA AAA AAG A F 839 Kaur et al. (2007)ACG AAG GAA CCG GGC TGC TAG R
hlyA GCA GTT GCA AGC GCT TGG AGT GAA F 456 Kaur et al. (2007)GCA ACG TAT CCT CCA GAG TGA TCG R
Iap ACA AGC TGC ACC TGT TGC AG F 131 Kaur et al. (2007)
86 D.K. Soni et al. / Infection, Genetics and Evolution 14 (2013) 83–91
oligonucleotide sequences (Sigma), used in this study are shown inTable 1.
2.8. Genomic fingerprinting by ERIC- and REP-PCR
ERIC-PCR was performed as described by Rivera et al. (1995) byusing two oligonucleotide primers (50-ATGTAAGCTCCTGGGGATT-CAC-30 and 50-AAGTAAGTGACTGGGGTGAGCG-30) in a 25 ll of solu-tion containing 2.5 ll 10� PCR buffer (100 mM Tris–HCl, 15 mMMgCl2, 500 mM KCl, pH 8.3), 1.5 ll of 15 mM MgCl2, 200 lM eachof dNTPs, 2.48% of DMSO, 25 lmol of each of the two primers (GCCBiotech, India), 2.5 U of Taq DNA polymerase (5 U/ll, Roche,Germany) and 4 ll of DNA template (50 ng/ll). PCR was performedunder following cyclic conditions: initial denaturation at 95 �C for7 min, 30 cycles in sequence 95 �C for 1 min, 52 �C for 1 min and65 �C for 8 min, and a final extension at 65 �C for 10 min.
REP-PCR was also performed as described by Versalovic et al.(1991) by using oligonucleotide primers (50-IIIICGICGICATCIGGC-30)and (50-ICGICTTATCIGGCCTAC-30) in a 25 ll of solution containing2.5 ll 10� PCR buffer (100 mM Tris–HCl, 15 mM MgCl2, 500 mMKCl, pH 8.3), 1.5 mM MgCl2, 200 lM each of dNTPs, 2.48% of DMSO,25 lmol of each of the two primers (GCC Biotech, India), 2.5 U ofTaq DNA polymerase (5 U/ll, Roche, Germany) and 5 ll of DNAtemplate (50 ng/ll). PCR was performed under the following cyclicconditions: initial denaturation at 95 �C for 7 min, 30 cycles insequence 95 �C for 1 min, 44 �C for 1 min and 65 �C for 8 min,and a final extension at 65 �C for 10 min. The amplicons wereelectrophoresed in 1.8% agarose at 60 V for 6 h and stained withethidium bromide. The fingerprint pattern was measured in aFluoro-S-Imager (Bio-Rad) and analyzed using Bionumerics finger-print analyst (Applied Maths, Kortrejik, Belgium) software with asimple-matching similarity matrix, and data were clustered bythe un-weighted pair group method with arithmetic means(UPGMA). The cluster analysis of the ERIC- and REP-PCR patternscould be affected by factors like position bias in gels, bandassignment, and different settings in the BioNumerics software.
Therefore, the similarity of the ERIC- and REP-PCR fingerprint pro-files was calculated using the average simple-match similarity ma-trix and the default cluster settings of 0.00% optimization and1.00% band position tolerance.
3. Results and discussion
3.1. Prevalence of L. monocytogenes
Eight (8%) out of the 100 water samples of which four werefrom Ravidas ghat, two from Assi ghat, one from Bhadaini ghatand one from Dr. R.P. ghat, were positive for L. monocytogenes.None of the water samples collected from upstream of the refer-ence point was found positive. No results are available on the pres-ence of L. monocytogenes in water samples of Indian water bodies,however, the present values for water samples are in agreementwith the values of 8.3% and 10.2% reported by Rodas-Suarez et al.(2006) and Lyautey et al. (2007), respectively. Water quality results(Table 2) indicate that Ganges water at Ravidas ghat is more con-taminated as compared to other three ghats from where the sam-ples have been collected. This is due to continuous discharge ofhuge volume of sewage (62–65 MLD as per SGRL analysis) throughAssi drain at this place. This seems to be the probable reason formore water samples showing positive for L. monocytogenes atRavidas ghat than those collected from elsewhere. The presenceof L. monocytogenes at Bhadaini ghat in one of the samples is ofserious concern as the water intake facility for supplying potablewater to the city is very close to this point. Our findings are consis-tent with the observations of other researchers who reported highprevalence of Listeria in water bodies receiving untreated domesticwastewater discharges (Paillard et al., 2005). As per report of UttarPradesh Jal Nigam, Government of U.P (2006), Varanasi city gener-ates 309.8 million liters of sewage per day (MLD) and most of itflows into the river regularly in untreated form contaminatingthe water with various pathogens including L. monocytogenes. Itis quite likely that some of these may be resistant and potential
Table 2Range of values of some gross water quality parameters for Ganges water at Varanasi. Assi drain upstream: upstream of reference point (control). Source: SGRL (Swatcha GangaResearch Laboratory), Varanasi, India.
Parameters Sampling points (ghats)
Ravidas Assi Bhadaini Dr. Rajendra Prasad Assi drain upstream
D.K. Soni et al. / Infection, Genetics and Evolution 14 (2013) 83–91 87
carriers of virulent genes. This aspect requires further detailedstudy.
Five (1.7%) out of the 300 clinical samples out of which four(5.3%) were from placental bit (75), one (1.3%) from vaginal swab(75) but none from either cervical swab or blood were positivefor L. monocytogenes. Dhanashree et al. (2003) reported 3.1% posi-tive infection of L. monocytogenes in placental bits and Kaur et al.(2007) showed that 3.2% (placental bits) and 1.6% (vaginal swab)samples were infected with L. monocytogenes.
Seven (5.8%) out of 120 cow milk samples were positive for L.monocytogenes. None of pasteurized milk and milk products(cheese, butter and ice-cream) was found positive for L. monocytog-enes. Earlier studies have demonstrated the presence of L. monocyt-ogenes in milk samples; however, our result is in accordance withthe values reported by Kalorey et al. (2008), 5.1% milk sampleswere positive for L. monocytogenes. Dhanashree et al. (2003) alsofound that none of the pasteurized milk and milk products (cheeseand ice-cream) was tested positive for L. monocytogenes.
3.2. Antibiotics susceptibility
Isolates of L. monocytogenes recovered from the water, humanclinical and milk samples were tested for their antibiotic suscepti-bility. Majority of isolates, irrespective of source of isolation,showed complete resistance to ampicillin but variable resistanceto cefoxitin, co-trimoxazole, gentamicin, oflaxacin, rifampin, andtetracycline (Table 3). All the isolates were susceptible to otherantibiotics tested. The selected antibiotics in our study are cur-rently used in the clinical treatment of listeriosis. For example,
Table 3Source of isolation, serogroup, antibiogram, ERIC- and REP-fingerprints and virulence profi
Isolates Source of isolation Date of isolation Serogroup Antibiogram
W1 Water 15.11.2009 4b, 4d, 4e A, Fox, TW2 Water 15.12.2009 4b, 4d, 4e A, R, TW3 Water 15.01.2010 4b, 4d, 4e A, R, SXT, TW4 Water 15.02.2010 1/2c, 3c A, R, SXT, TW5 Water 15.12.2009 1/2c, 3c A, Of, R, SXT, TW6 Water 15.01.2010 1/2c, 3c A, R, SXT, TW7 Water 15.01.2010 1/2c, 3c A, R, SXT, TW8 Water 15.01.2010 1/2c, 3c A, G, R, SXT, TPb1 Human 03.06.2009 1/2c, 3c A, R, G, Of, SXT, TPb2 Human 08.08.2009 4b, 4d, 4e A, R, G, Of, SXT, TPb3 Human 14.11.2009 4b, 4d, 4e A, R, G, Of, SXT, TPb4 Human 17.02.2010 4b, 4d, 4e A, R, G, Of, SXT, TVS1 Human 05.05.2010 4b, 4d, 4e A, R, G, Of, SXT, TM2 Milk 05.06.2009 1/2a, 3a A, R, G, Of, SXT, TM3 Milk 10.07.2009 1/2a, 3a A, R, SXTM14 Milk 12.09.2009 1/2b, 3b, 7 A, R, SXT, TM21 Milk 17.12.2009 1/2b, 3b, 7 A, R, SXTM25 Milk 09.02.2010 1/2b, 3b, 7 R, SXTM28 Milk 12.04.2010 1/2a, 3a A, R, SXTM38 Milk 15.07.2010 1/2b, 3b, 7 R, SXT
W, water from the river Ganges; Pb, human placental bit; VS, human vaginal swab; M,A, ampicillin; Fox, cefoxitin; R, rifampicin; G, gentamicin; Of, ofloxacin; SXT, co-triomox
antibiotics used in treatment of many human disease (ampicillin,gentamicin, streptomycin, and trimethoprim/sulfamethoxazole),and in veterinary medicine (ampicillin, ciprofloxacin, gentamicin,penicillin, streptomycin, tetracycline, and trimethoprirn/sulfa-methoxazole) (Lyon et al., 2008; Nwachukwu et al., 2010).
L. monocytogenes is generally susceptible to wide range of anti-biotics (Hof et al., 1997). However, multidrug-resistant strainshave been isolated from food, environment and sporadic cases ofhuman listeriosis (Franco-Abuin et al., 1994; Carpentier et al.,1995), after the isolation of one multi drug-resistant strain inFrance in 1988 (Poyart-Salmeron et al., 1990). L. monocytogenesresistance to antibiotics like ampicillin, erythromycin, gentamicin,kanamycin, penicillin, streptomycin, sulphonamide, trimethoprim,tetracycline, and rifampicin have also been reported (Carpentierand Courvalin, 1999; Nwachukwu et al., 2010; Yan et al., 2010).Since the first detection of tetracycline resistant strain in 1988 inFrance (Poyart-Salmeron et al., 1990), other workers have fre-quently reported tetracycline resistance among L. monocytogenesisolated from different sources (Carpentier and Courvalin, 1999;Prazak et al., 2002).
In India, Dhanashree et al. (2003) reported that all the isolatesof L. monocytogenes were sensitive to ampicillin, ciprofloxacin,cotrimoxazole, erythromycin, penicillin and chloramphenicol,while Sharma et al. (2012) reported that most of L. monocytogenesisolates were multidrug resistant; several isolates were resistant tonalidixic acid, amoxycillin + sulbactam, cloxacillin, erythromycin,kanamycin and vancomycin and many isolates also exhibitedintermediate resistance to erythromycin, vancomycin, streptomy-cin and cloxacillin. In the present study, all isolates, irrespective
les of L. monocytogenes used in this study.
ERIC type REP type Presence of following genes determined by PCR
88 D.K. Soni et al. / Infection, Genetics and Evolution 14 (2013) 83–91
of the source, showed resistance to multiple antibiotics and themajority to tetracycline. These observations support the previousstudies which indicate that the indiscriminate use of antibioticsincluding tetracycline on human, discharge of contaminated efflu-ent/waste in the environment and agriculture (e.g., as growth pro-moting and prophylactic agents in animals) could lead to theemergence of antibiotic resistance (Khachatourarians, 1998). Themultidrug-resistant organisms that have the ability to cause hu-man infection can create considerable medical and public healthcomplications (Rodas-Suarez et al., 2006; Nwachukwu et al.,2010). Moreover, application of feces or dung slurries of infected(or carrier) animals onto agricultural land as manure can also serveas source of the organism (Chukwu et al., 2006). This antibiotic-resistant bacteria arising from such agricultural practices can causeinfection in humans via the ingestion of uncooked food (cropsgrown in soil irrigated with contaminated water and/or fertilizedwith Listeria-contaminated manure or biosolids) can thus facilitatetrans-border resistance (Nightingale et al., 2004).
For the last 30 years, a combination of trimethoprim (TMP) andsulfamethoxazole (SMX) has been used as human and veterinarymedicine (Poros-Gluchowska and Makiewicz, 2003). Standardtherapy for listeriosis remains the administration of ampicillin orpenicillin G combined with an aminoglycoside gentamicin. Thesecond obvious choice is combination of trimethoprim (TMP) andsulfamethoxazole (SMX) (Carpentier and Courvalin, 1999). Theantibiotics resistance can also be due to the selective pressure ofantibiotics in clinic and environment (Khachatourarians, 1998;Hanchun et al., 2004) or acquisition of antibiotic resistant genesby integrons and other insertion elements (Dider et al., 2000). Inaddition, occurrences of temporal and spatial changes in antibioticresistance pattern are well known (Lyon et al., 2008; Yan et al.,2010). The majority of isolates of L. monocytogenes, irrespectiveof the source, showed resistance to first line antibiotics includinggentamicin (for human isolates) and second line antibiotics, TMPand SMX, raised alarm about listeriosis treatment in this region.Therefore, the result of the present study indicates the prevalenceof multi-drug resistant isolates of L. monocytogenes in water, hu-man clinical and milk samples, and underline the need for activeand continuous surveillance of their antibiotic resistance.
3.3. Species and serovar identification
All the 20 isolates gave positive results for gene encoding forinternalin A (inlA) indicating that all strains belong to L. monocytog-enes. In serotype specific multiplex PCR, three milk isolates gavepositive results for lmo0737 gene, four milk isolates for ORF2819gene, five water and one placental bit isolates for lmo1118 andlmo0737 genes and three each of human placental bit and waterand one vaginal swab for ORF2110 gene indicating that these iso-lates belong to 1/2a, 3a; 1/2b, 3b, 7; 1/2c, 3c; and 4b, 4d, 4e sero-groups, respectively (Table 3). The result of serogrouping alsorevealed that serogroup 4b was isolated from human and water,and serogroups 1/2a and1/2b were isolated from milk. Theseobservations thus corroborate the findings of other workers whoreported isolation of serogroups 1/2a, 1/2b and 4b from clinicalcases and association of 4b serotype in causing human listeriosis(Farber and Peterkin, 1991; Doumith et al., 2004; Liu, 2006). Incontrast, isolation of serogroups 1/2c and 3c from human andwater may indicate the role of this serogroup in causing infection.However, a detail study including large number of isolates isneeded to establish this fact.
3.4. Virulence associated genes
Twenty isolates of L. monocytogenes were examined for thepresence of virulence genes. The majority of the isolates, irrespec-
tive of the source, showed positive results for inlA, inlC and inlJ. Allisolates were positive for actA and hlyA genes. On the other hand,all the isolates from human clinical and majority isolates from milkgave positive results for plcA, prfA, actA and iap genes. The remain-ing isolates showed presence of variable virulence genes. The dis-tribution of virulence in these isolates is shown in Table 3.
Pathogenicity tests are mandatory to differentiate betweenpathogenic and non-pathogenic strains of L. monocytogenes exist-ing in nature. Several protocols have been developed for assess-ment of L. monocytogenes virulence including in vivo bioassaysand in vitro cell assays. The in vivo method has limitations becauseof its high cost and use of animals. On the other hand, in vitro assayis hampered by its lack of desired reproducibility and the time re-quired completing the analysis. PCR based assays for key virulence-associated genes show rapid and reproducible results. L. monocyt-ogenes is known to possess virulence genes comprising prfA, plcA,hlyA, mpl, actA and plcB located on a virulence pathogenicity island,prfA virulence gene cluster (pVGC) (Vazquez-Boland et al., 2001).These virulence gene determinants are important for the intracel-lular life cycle including vacuole escape and cell to cell spread. ThehlyA encoding listeriolysin O (LLO) helps L. monocytogenes to es-cape from the host cell vacuole, and plcA and plcB play a role indestabilization of cell vacuole membranes (Galliard et al., 1991;Smith et al., 1995). The actA that encodes the ActA protein isresponsible for the accumulation of host actin and for mobility.The role of other virulence genes such as inlA and inlB has been re-ported in human listeriosis (Chakraborty et al., 1992; Liu et al.,2007). The presence of either or both inlC and inlJ genes involvesin L. monocytogenes strain indicate its potential virulence and abil-ity to cause mouse mortality via the intraperitoneal route. It, how-ever, does not necessarily indicate the certainty of the strainscontaining these genes to produce disease in humans via the con-ventional oral ingestion. As human listeriosis is usually a resultfrom ingestion of L. monocytogenes contaminated food, it requiresinvolvement of many other known genes (plcA, prfA, hly, actA,and iap) (Liu et al., 2007). The present results suggest that usingmultiplex PCR of inlA, inlC and inlJ genes species- and virulence-specific properties of L. monocytogenes can be detected together.Results of this study also indicate that isolates of L. monocytogenes,irrespective of the source of isolation, possess internalin inlJ genesindicating their ability for internalization of pathogen into the cell.However, all isolates from human clinical, two from water and fourfrom milk samples possess virulence genes encoding for inlA, inlC,inlJ, plcA, prfA, actA, hlyA and iap indicate that these isolates possessall properties of virulent strain. They had ability to enter via intra-peritoneal route or through the use of contaminated food andcause infection. These findings are similar to those of other work-ers who reported isolation of virulent L. monocytogenes from pla-cental bit and vaginal swab (Dhanashree et al., 2003; Kaur et al.,2007). Moreover, isolates from water and milk samples showedvariation in virulence genes profile that could be because of the ab-sence of virulence genes or occurrence of some mutation in thesegenes (Rodas-Suarez et al., 2006; Kalorey et al., 2008).
3.5. ERIC- and REP-PCR fingerprint analysis
ERIC-PCR of genomic DNA from L. monocytogenes isolates fromGanges water, human clinical and milk yielded twelve fingerprintprofiles (profile 1 through XII) consisting of three to nineteenbands of sizes ranging 140–5200 bases (Fig. 2). Five fingerprintprofiles were shown by eight isolates from the Ganges water.Although four of the five human isolates yielded similar fingerprintprofiles, the fifth one had a distinct profile similar to that of themilk isolate. Interestingly, six milk isolates yielded five distinct fin-gerprint profiles, but are not comparable to the water and humanisolates.
Fig. 2. DNA fingerprints generated by ERIC-PCR amplification from Ganges water, human clinical and milk isolates of L. monocytogenes. Lanes M, 1-kb molecular weightladder; (a) lanes 1–8, water isolates W1, W2, W3, W4, W5, W6, W7 and W8; lanes 9–13, human isolates Pb1, Pb2, Pb3, Pb4 and VS1; (b) lanes 14 to 20, milk isolates M2, M3,M14, M21, M25, M28 and M38, respectively. (c) Digitized ERIC-PCR profile obtained from genomic DNA of water, human and milk isolates of L. monocytogenes. Thedendrogram was generated by using the Bionumerics Fingerprint Analyst Software (Applied Maths), and data were clustered by the unweighted pair group method witharithmetic means. Similarity of the ERIC-PCR fingerprint profiles was calculated using the average simple-match similarity matrix and the default cluster settings of 0.00%optimization and 1.00% band position tolerance were used.
D.K. Soni et al. / Infection, Genetics and Evolution 14 (2013) 83–91 89
REP-PCR of genomic DNA from L. monocytogenes from varioussources resulted in amplification of multiple DNA fragments(126–6800 bases long). Like the ERIC-profile, five fingerprint pro-files were shown by eight isolates from Ganges water (Fig. 3).Although four human isolates yielded similar fingerprint profiles,the fifth one was distinct. Interestingly, seven milk isolates yieldedfive distinct fingerprint profiles, quite different from those of wateror human isolates.
ERIC-sequences are 124 to 127 bases long element consisting ofhighly conserved central inverted repeat, and located in extragenicregions of the bacterial genome. REP elements (38-bp long) containsix degenerate positions and a 5-bp variable loop between eachside of the conserved palindrome (Versalovic et al., 1991; Riveraet al., 1995; Jersek et al., 1999; Fugett et al., 2007). ERIC- andREP-PCR were used to measure the relatedness of L. monocytogenesfrom Ganges water, human clinical and milk and its serotypes. Hu-man clinical isolates showed similar ERIC and REP fingerprints butquite distinct from the water and milk isolates. Overall, there wasstrong correlation among the PCR methods applied, and pathogenisolated from human clinical yielded fingerprints identical or simi-
lar but distinct from the water or milk isolates. The observed differ-ence in the banding pattern among the isolates from differentsources suggests the divergence in genomic organization thatmight have arisen from genetic reassortment applicable to differentecological niches over time. There was no correlation between PCRfingerprint profiles and the serogroups of L. monocytogenes. Fur-thermore, there was no correlation between fingerprint profilesand the source of isolation. These findings are very similar to thoseof others wherein food isolates showed clearly different finger-prints relative to human and animal isolates (Jersek et al., 1999;Fugett et al., 2007). Isolates belonging to different or individualserogroup could be differentiated into different molecular sub-types; the approach considered to be more accurate than the sero-typing. Jersek et al. (1999) reached a similar conclusion that not allL. monocytogenes bacteria in food, inflict disease because of inter-strain differences in their virulence potential. Result of this studyadds further evidence to the potential of REP-PCR as the efficientand sensitive molecular typing tool for L. monocytogenes of differentorigin. These observations thus suggest that REP- and ERIC-PCR canbe used to trace the origin of isolates for epidemiological purposes.
Fig. 3. REP-PCR profile obtained by using DNA from Ganges water, human clinical and milk isolates of L. monocytogenes. Lanes M, 1-kb molecular weight ladder; (a) lanes 1–8,water isolates W1, W2, W3, W4, W5, W6, W7 and W8; lanes 9–13, human isolates Pb1, Pb2, Pb3, Pb4 and VS1; (b) lanes 14–20, milk isolates M2, M3, M14, M21, M25, M28and M38, respectively. (c) Digitized fingerprints of water, human and milk isolates of L. monocytogenes generated by REP-PCR amplification. The dendrogram was constructedby using the Bionumerics Fingerprint Analyst Software (Applied Maths) as described in the legend of Fig. 2.
90 D.K. Soni et al. / Infection, Genetics and Evolution 14 (2013) 83–91
4. Conclusions
Isolates of L. monocytogenes recovered from clinical and envi-ronmental samples belonging to 4b, 4d, 4e or 1/2c, 3c or 1/2a, 3aor 1/2b, 3b, 7 serogroups exhibited multiple antibiotic resistanceand showed the presence of variable virulence genes. REP- andERIC-PCR results showed that human clinical isolates yielded sim-ilar PCR fingerprint profiles indicating close relationships amongthem. The pathogenicity and multi-resistance attributes againstcommonly used antimicrobial drugs may pose serious threats tohuman health. Thus, there is an urgent need to carry out extensiveresearch on L. monocytogenes with special reference to its habitatsuch as water, food and clinical samples in order to understandits further transmission route in the environment and resistanceto antibiotics.
Acknowledgments
This study was supported by the Indian Council of Medical Re-search, New Delhi Grant No. 5/3/3/10/2007-ECD-I to SKD, and fundcontributed by the Department of Biotechnology, New Delhi to
Institute of Life Sciences. The authors are grateful to ProfessorS.N. Upadhyay, Department of Chemical Engineering and Technol-ogy, I.I.T. (BHU), Varanasi for his critical suggestions on manuscriptand Swatcha Ganga Research Laboratory (SGRL), Varanasi, India,for providing the river Ganges water quality data. One of theauthors (RKS) is grateful to UGC, New Delhi, for financial assistance(JRF).
References
American Public Health Association, 1992. Standard Methods for the Examinationof Water and Wastewater, 18th ed. American Public Health Association,Washington, DC.
Anon., 1997. EN ISO 11290-1 microbiology of food and animal feeding stuffs –horizontal method for the detection and enumeration of Listeria monocytogenes– Part 1: Detection. International Organization for Standardisation, Geneva.
Aurora, R., Prakash, A., Prakash, S., Rawool, D.B., Barbuddhe, S.B., 2008. Comparisonof PI-PLC based assays and PCR along with in vivo pathogenicity tests for rapiddetection of pathogenic Listeria monocytogenes. Food Control 19, 641–647.
Barbuddhe, S.B., Malik, S.V.S., Kumar, J.A., Kalorey, D.R., Chakraborty, T., 2012.Epidemiology and risk management of listeriosis in India. Int. J. Food Microbiol.154, 113–118.
D.K. Soni et al. / Infection, Genetics and Evolution 14 (2013) 83–91 91
Bauer, A.W., Kirby, W.M., Sherris, J.C., Turck, M., 1966. Antibiotic susceptibilitytesting by a standardized single disk method. Am. J. Clin. Pathol. 45, 493–496.
Carpentier, E., Courvalin, P., 1999. Antibiotic resistance in Listeria spp.. Antimicrob.Agents Chemother. 43, 2103–2108.
Carpentier, E., Gerbaud, G., Jacquet, C., Rocourt, J., Courvalin, P., 1995. Incidence ofantibiotic resistance in Listeria species. J. Infect. Dis. 172, 277–281.
Chakraborty, T., Leimeister-Washter, M., Domann, E., Hartl, M., Goebel, W.,Nichterlein, T., Notermans, S., 1992. Coordinate regulation of virulence genesin Listeria monocytogenes requires the product of the prfA gene. J. Bacteriol. 174,568–574.
Chen, B., Pyla, R., Kim, T., Silva, J.L., Jung, Y., 2010. Prevalence and contaminationpatterns of Listeria monocytogenes in catfish processing environment and freshfillets. Food Microbiol. 27, 645–652.
Chukwu, O.O.C., Ogbonna, C.I.C., Olabode, O.A., Chukwu, D.I., Owuliri, F.C., Nwankiti,O.O., 2006. Listeria monocytogenes in Nigerian processed meats and ready to eatdairy products. Niger. J. Microbiol. 20, 900–904.
Clinical and Laboratory Standards Institute (previously National Committee onClinical Laboratory Standards), 2006. Performance Standards for AntimicrobialDisk Susceptibility Tests; Approved Standard-Ninth Edition (M2-A9). Clinicaland Laboratory Standards Institute, Wayne, PA.
Cummins, A.J., Fielding, A.K., McLauchlin, J., 1994. Listeria ivanovii infection in apatient with AIDS. J. Infect. 28, 89–91.
Dhanashree, B., Otta, S.K., Karunasagar, I., Goebel, W., Karunasagar, I., 2003.Incidence of Listeria spp. in clinical and food samples in Mangalore, India.Food Microbiol. 20, 447–453.
Dider, M., Broderich, D., Vera, A.W., Julian, D., 2000. Antibiotic resistance in theECOR collection: integrons and identification of a novel genes. Antimicrob.Agents Chemother. 15, 1568–1574.
Doumith, M., Buchriester, C., Glaser, P., Jacquet, C., Martin, P., 2004. Differentiationof the major Listeria monocytogenes serovars by multiplex PCR. J. Clin. Microbiol.42, 3819–3822.
Franco-Abuin, C.M., Quinto-Fernandez, E.J., Fente-Sampayo, C., Rodriguez-Otero, J.L.,Dominguez-Rodriguez, L., Sepeda-Saez, A., 1994. Susceptibilities of Listeriaspecies isolated from food to nine antimicrobial agents. Antimicrob. AgentsChemother. 38, 1655–1657.
Fugett, E.B., Schoonmaker-Bopp, D., Dumas, N.B., Corby, J., Wiedmann, M., 2007.Pulsed field gel electrophoresis (PFGE) analysis of temporally matched Listeriamonocytogenes isolated from human, clinical cases, food, ruminant farms, andurban and natural environments reveals source associated as well as widelydistributed PFGE types. J. Clin. Microbiol. 45, 865–873.
Galliard, J.L., Berche, P., Frehel, C., Gouin, E., Cossart, P., 1991. Entry of L.monocytogenes into cells is mediated by internalin, a repeat proteinreminiscent of surface antigen from Gram-positive cocci. Cell 65, 1127–1141.
Gandhi, M., Chikindas, M.L., 2007. Listeria: a foodborne pathogen that knows how tosurvive. Int. J. Food Microbiol. 113, 1–15.
Gerner-Smidt, P., Hise, K., Kincaid, J., Hunter, S., Rolando, S., Hyytia-Trees, E., Ribot,E.M., Swaminathan, B., 2006. PulseNet USA: a five-year update. FoodbornePathog. Dis. 3, 9–19.
Guillet, C., Join-Lambert, O., Monnier, A.L., Leclercq, A., Mechai, F., Mamzer-Bruneel,M.F., Bielecka, M.K., Scortti, M., Disson, O., Berche, P., Vazquez-Boland, J.,Lortholary, O., Lecuit, M., 2010. Human Listeriosis caused by Listeria ivanovii.Emerg. Infect. Dis. 16, 136–138.
Hanchun, Y., Sheng, C., David, G.W., Shaohua, Z., Patric, M.D., Robert, W.,Jianghong, M., 2004. Characterization of multiple antimicrobial-resistantEscherichia coli isolates from chicken and swine in China. J. Clin. Microbiol.42, 3484–3489.
Hof, H., Nichterlien, T., Krestschmar, M., 1997. Management of listeriosis. Clin.Microbiol. Rev. 10, 345–357.
Jersek, B., Gilot, P., Gubina, M., Klun, N., Mehle, J., Teherneva, E., Rijpens, N., Herman,L., 1999. Typing of Listeria monocytogenes strains by repetitive elementsequence-based PCR. J. Clin. Microbiol. 37, 103–109.
Kalorey, D.R., Warke, S.R., Kurkure, N.V., Rawool, D.B., Barbuddhe, S.B., 2008. Listeriaspecies in bovine raw milk: a large survey of Central India. Food Control 19,109–112.
Kaur, S., Malik, S.V.S., Vaidya, V.M., Barbuddhe, S.B., 2007. Listeria monocytogenes inspontaneous abortions in humans and its detection by multiplex PCR. J. Appl.Microbiol. 103, 1889–1896.
Khachatourarians, G.G., 1998. Agricultural use of antibiotics and the evolutionand transfer of antibiotic-resistant bacteria. Can. Med. Assoc. J. 159, 1129–1136.
Leclercq, A., Clermont, D., Bizet, C., Grimont, P.A., Le Flèche-Matéos, A., Roche, S.M.,Buchrieser, C., Cadet-Daniel, V., Le-Monnier, A., Lecuit, M., Allerberger, F., 2010.Listeria rocourtiae sp. nov.. Int. J. Syst. Evol. Microbiol. 60, 2210–2214.
Liu, D., 2006. Identification, subtyping and virulence determination of Listeriamonocytogenes, an important foodborne pathogen. J. Med. Microbiol. 55, 645–659.
Liu, D., Lawrence, M.L., Austin, F.W., Ainsworth, A.J., 2007. A multiplex PCR forspecies- and virulence-specific determination of Listeria monocytogenes. J.Microbiol. Methods 71, 133–140.
Lyautey, E., Lapen, D.R., Wilkes, G., McCleary, K., Pagotto, F., Tyler, K., Hartmann, A.,Piveteau, P., Rieu, A., Robertson, W.J., Medeiros, D.T., Edge, T.A., Gannon, V.,Topp, E., 2007. Distribution and characteristics of Listeria monocytogenes isolatesfrom surface waters of the south nation river watershed, Ontario, Canada. Appl.Environ. Microbiol. 73, 5401–5410.
Lyon, S.A., Berrang, M.E., Fedorka-Cray, P.J., Fletcher, D.L., Meinersmann, R.J., 2008.Antimicrobial resistance of Listeria monocytogenes isolated from a poultryfurther processing plant. Foodborne Pathog. Dis. 5, 253–259.
Mishra, V.B., 2005. The Ganga at Varanasi and a travail to stop her abuse. Curr. Sci.89, 755–763.
Moon, J.S., Lee, A.R., Kang, H.M., Lee, E.S., Kim, M.N., Paik, Y.H., Park, Y.H., Joo, Y.S.,Koo, H.C., 2007. Phenotypic and genetic antibiogram of methicillin-resistantstaphylococci isolated from bovine mastitis in Korea. J. Dairy Sci. 90, 1176–1185.
Nightingale, K.K., Schukken, Y.H., Nightingale, C.R., Fortes, E.D., Ho, A.J., Her, Z.,Grohn, Y.T., McDonough, P.L., Wiedmann, M., 2004. Ecology and transmission ofListeria monocytogenes infecting ruminants and in farm environment. Appl.Environ. Microbiol. 70, 4458–4467.
Notermans, S.H., Dufrenne, J., Leimeister-Wachter, M., Domann, E., Chakraborty, T.,1991. Phosphatidylinositol-specific phospholipase C activity as a marker todistinguish between pathogenic and non-pathogenic Listeria species. Appl.Environ. Microbiol. 57, 2666–2670.
Nwachukwu, N.C., Orji, F.A., Iheukwumere, I., Ekeleme, U.G., 2010. Antibioticresistant environmental isolates of Listeria monocytogenes from anthropogeniclakes in Lokpa-Ukwu, Abia State of Nigeria. Aust. J. Basic Appl. Sci. 4, 1571–1576.
Orndorff, E.O., Hamrick, T.S., Smoak, I.W., Havell, E.A., 2006. Host and bacterialfactors in listeriosis pathogenesis. Vet. Microbiol. 114, 10–15.
Paillard, D., Dubois, V., Thiebaut, R., Nathier, F., Hoogland, E., Caumette, P., Quentin,C., 2005. Occurrence of Listeria spp. in effluents of French urban wastewatertreatment plants. Appl. Environ. Microbiol. 71, 7562–7566.
Prazak, M.A., Murano, E.A., Mercado, I., Acuff, G.R., 2002. Antimicrobial resistance ofListeria monocytogenes isolated from various cabbage farms and packing shedsin Texas. J. Food Prot. 65, 1796–1799.
Rivera, I.G., Chowdhury, M.A.R., Huq, A., Jacobs, D., Martins, M.T., Colwell, R.R., 1995.Enterobacterial repetitive intergenic consensus sequences and the PCR togenerate fingerprints of genomic DNA from Vibrio cholerae O1, O139 and non-O1 strains. Appl. Environ. Microbiol. 61, 2898–2904.
Rodas-Suarez, O.R., Flores-Pedroche, J.F., Betancourt-Rule, J.M., Quinones-Ramirez,E.I., Vazquez-Salinas, C., 2006. Occurrence and antibiotic sensitivity of Listeriamonocytogenes strains isolated from oysters, fish and estuarine water. Appl.Environ. Microbiol. 72, 7410–7412.
Seeliger, H.P.R., Jones, D., 1986. Listeria. In: Sneath, P.H.A., Maine, N.S., Sharpe, M.E.,Holt, J.G. (Eds.), Bergey’s Manual of Systematic Bacteriology, 9th ed. Williamsand Wilkins, Baltimore, Maryland, pp. 1235–1245.
Sharma, D., Sharma, P.K., Saharan, B.S., Malik, A., 2012. Isolation, identification andantibiotic susceptibility profiling of antimicrobial resistant Listeriamonocytogenes from dairy milk. Int. J. Microb. Resour. Technol. 1, 1–4.
Smith, G.A., Klarsfeld, A., Msadek, T., Cossart, P., 1995. The two distinctphospholipases C of Listeria monocytogenes have overlapping roles in escapefrom a vacuole and cell to cell spread. Infect. Immun. 63, 4231–4237.
Swaminathan, B., Barrett, T.J., Hunter, S.B., Tauxe, R.V., 2001. PulseNet: the molecularsubtyping network for foodborne bacterial disease surveillance, United States.Emerg. Infect. Dis. 7, 382–389.
Uttar Pradesh Jal Nigam, 2006. Ganga action plan (phase II) at Varanasi. <http://www.upjn.org/plan-II.aspx>.
Versalovic, J.T., Koeuth, T., Lupski, J.R., 1991. Distribution of repetitive DNAsequences in Eubacteria and application to fingerprinting of bacterialgenomes. Nucleic Acid Res. 19, 6823–6831.
WHO, 2004. World Health Organization and Food and Agriculture Organization.Risk assessment of Listeria monocytogenes in ready-to-eat foods, MRA Series No.4–5. Rome, Italy.