Cholangiocyte cilia express TRPV4 and detect changesin luminal tonicity inducing bicarbonate secretionSergio A. Gradilone*, Anatoliy I. Masyuk*, Patrick L. Splinter*, Jesus M. Banales*†, Bing Q. Huang*, Pamela S. Tietz*,Tatyana V. Masyuk*, and Nicholas F. LaRusso*‡
*Department of Internal Medicine, Division of Gastroenterology and Hepatology, Mayo Clinic College of Medicine, Rochester, MN 55905; and †Centro deInvestigacion Medica Aplicada (CIMA) and Centro de Investigacion Biomedica en red Enfermedades Hepaticas y Digestivas (CIBEREHD), E-31008,Pamplona, Spain
Edited by David E. Clapham, Harvard Medical School, Boston, MA, and approved October 10, 2007 (received for review June 26, 2007)
Cholangiocytes, epithelial cells lining the biliary tree, have primarycilia extending from their apical membrane into the ductal lumen.Although important in disease, cilia also play a vital role in normalcellular functions. We reported that cholangiocyte cilia are sensoryorganelles responding to mechanical stimuli (i.e., luminal fluid flow)by alterations in intracellular Ca2� and cAMP. Because cholangiocytecilia are also ideally positioned to detect changes in composition andtonicity of bile, we hypothesized that cilia also function as osmosen-sors. TRPV4, a Ca2�-permeable ion channel, has been implicated insignal transduction of osmotic stimuli. Using purified rat cholangio-cytes and perfused intrahepatic bile duct units (IBDUs), we found thatTRPV4 is expressed on cholangiocyte cilia, and that hypotonicityinduces an increase in intracellular Ca2� in a TRPV4-, ciliary-, andextracellular calcium-dependent manner. The osmosensation of lu-minal tonicity by ciliary TRPV4 induces bicarbonate secretion, themain determinant of ductal bile formation, by a mechanism involvingapical ATP release. Furthermore, the activation of TRPV4 in vivo, by itsspecific agonist, 4�PDD, induces an increase in bile flow as well as ATPrelease and bicarbonate secretion. Our results suggest that cholan-giocyte primary cilia play an important role in ductal bile formation byacting as osmosensors.
bile secretion � osmosensor � calcium � ATP release � purinergic receptor
Bile initially secreted by hepatocytes is modified by cholangio-cytes (epithelial cells lining the bile ducts) via absorptive and
secretory processes (1). Although cholangiocytes account for only3–5% of the liver cell population, they generate up to 40% of totalbile volume (1, 2). Hepatocytes secrete osmotically active solutes,primarily bile salts, and glutathione, establishing a gradient thatgenerates water movement via aquaporin water channels (1–4).Primary bile then flows through the lumen of intrahepatic bile ductswhere cholangiocytes modify it by absorption of solutes, mainly bilesalts and glucose, and secretion of ions like Cl� and HCO3
�.Ultimately, these processes lead to water secretion (i.e., ductal bileproduction) by cholangiocytes, also involving aquaporin waterchannels (2, 5).
Cholangiocytes, but not hepatocytes, express primary cilia, whichare microtubule-based organelles (6), extending from their apicalplasma membrane into the ductal lumen. Thus, cholangiocyte ciliaare ideally positioned to detect changes in the flow, composition,and tonicity of luminal bile (7). Until recently, primary cilia wereconsidered vestigial organelles with no physiologically relevantfunctions (8). Triggered by observations that mutations in ciliary-associated proteins cause disease (e.g., situs inversus, hydrocepha-lus, obesity, and polycystic kidney and liver disease), primary ciliahave become the subject of intense investigation (9–11).
Primary cilia are also important to normal cell function. Inmammals, primary cilia detect different stimuli, including light (byphotoreceptors) and urine flow (by primary cilia on renal tubularepithelia) (9, 12). We reported that primary cilia on cholangiocytesact as mechanosensors, responding to luminal fluid flow by alter-ations in intracellular Ca2� and cAMP. The ciliary proteins in-volved in this transduction of mechanical stimuli included polycys-
tin-1 (PC1), a cell surface receptor; polycystin-2 (PC2), a Ca2�
channel; and adenylyl cyclase isoform 6 (7). Given that cholangio-cyte cilia are exposed to various osmotic stresses induced by themovement of osmotically active molecules into and out of bileflowing through the lumen of intrahepatic bile ducts, we hypoth-esized that these organelles might also function as osmosensors.
TRPV4 is a member of the transient receptor potential (TRP)superfamily of cation channels that function as integrators ofphysical and chemical stimuli. This Ca2�-permeable channel func-tions as an osmosensor, being activated by anisosmotic conditions(13–20). In Trpv4-transfected CHO cells, TRPV4 participates inregulatory volume decrease (RVD), sensing a hypotonic stimulusand mediating influx of extracellular calcium that activates signalingcascades resulting in secretion of K� and Cl�, followed by loss ofintracellular water (13, 21). The worm, Caenorhabditis elegans,expresses Osm-9, a gene of the TRP family and an ortholog ofTrpv4, the protein product of which is found in its ciliated sensoryneurons. Worms with a mutation in Osm-9 have defects in theirresponse to osmotic, mechanical, and odorant stimuli, defects thatare rescued by neuron-directed transgene expression of TRPV4(22). In mammals, TRPV4 is expressed in multiple organs (e.g.,kidney, trachea, lung, and brain) in cells exposed to changes intonicity of fluid (14, 23, 24).
In the work described here using rat and mouse cholangiocytes,perfused intrahepatic bile duct units (IBDUs), and in vivo ratmodels, we show that TRPV4 is expressed on cholangiocyte cilia,and that hypotonicity induces an increase in intracellular Ca2� in aTRPV4-, ciliary-, and extracellular calcium- dependent manner.The osmosensation of luminal tonicity detected by ciliary TRPV4induces bicarbonate secretion, the main determinant of ductal bileformation, through a mechanism involving ATP release into theductal lumen. Our results provide evidence of TRPV4 expressionin primary cilia and suggest that cholangiocyte primary cilia play animportant role in the regulation of ductal bile formation by actingas osmosensors.
ResultsCholangiocytes Express TRPV4 Message and Protein. We performedRT-PCR on total RNA from isolated mouse and rat cholangiocytes(25) and cultured mouse (NMC) and rat (NRC) cholangiocytes (26)with cDNA from kidney as positive control. As shown in Fig. 1A,both mouse and rat cholangiocytes express Trpv4 mRNA. Wesequenced the PCR products and verified the identities of the
Author contributions: S.A.G., A.I.M., P.L.S., and N.F.L. designed research; S.A.G., A.I.M.,P.L.S., J.M.B., and B.Q.H. performed research; J.M.B., P.S.T., and T.V.M. contributed newreagents/analytic tools; S.A.G., A.I.M., P.L.S., J.M.B., and T.V.M. analyzed data; and S.A.G.and N.F.L. wrote the paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.
‡To whom correspondence should be addressed. E-mail: [email protected].
This article contains supporting information online at www.pnas.org/cgi/content/full/0705964104/DC1.
amplicons by database homology searches (BLAST; National Cen-ter for Biotechnology Information, National Institutes of Health).
As shown in Fig. 1B, immunoblotting for TRPV4 detected adouble band between 75 and 100 kDa in rat kidney and IBDUs, apattern similar to that reported in mouse kidney (27); only the highmolecular-weight isoform was seen in NRCs. Bands were absentwhen TRPV4 antibody was preabsorbed with a molar excess ofimmunizing peptide. To further evaluate the identity of the TRPV4band, we obtained total protein from NMCs transfected to over-express TRPV4 (NMC-Trpv4) and NMC-Trpv4 cotransfected witha shRNA against TRPV4 (NMC-Trpv4-shRNA) cell lines. Westernblots showed a similar double band pattern in all samples (Fig. 1C);as expected, protein expression was higher in the overexpressingNMC-Trpv4 cell line than in NMC and NMC-Trpv4-shRNA. Afterincubation with N-glycosidase, the highest molecular-mass bandshifted to the 75-kDa band (Fig. 1C), indicating the presence ofN-glycosylation in the TRPV4 molecule, as reported (28), andfurther confirming antibody specificity.
TRPV4 Protein Is Expressed on Cholangiocyte Primary Cilia. Weperformed immunofluorescent confocal microscopy on NMC andNMC-Trpv4 cultured for 7 days after confluence, a manipulationthat results in the development of mature cilia on cholangiocytes inculture (29). Using antibodies to TRPV4 and to acetylated �-tu-bulin, a known component of the ciliary axoneme, we found thatTRPV4 is expressed in cholangiocyte cilia (Fig. 2A). Immunoflu-orescent confocal microscopy on rat liver tissue sections confirmedthe ciliary localization of TRPV4 in vivo (Fig. 2B). The presence ofTRPV4 in cilia was also confirmed by immunogold electron mi-croscopy on IBDUs (Fig. 2C). Finally, a highly enriched fraction ofcilia isolated from NRCs (29) stained positively for TRPV4(Fig. 2D).
Extracellular Hypotonicity Induces a Ca2� Response in Cholangiocytesin a TRPV4- and Extracellular Ca2�-Dependent Manner. It was previ-ously reported that Mz-Cha-1 cells, derived from a human cholan-giocarcinoma, respond to hypotonicity by an increase in [Ca2�]i (30,31). To test the possibility that TRPV4 is involved in this response,we exposed NMC, NMC-TrpV4, and NMC-Trpv4-shRNA cells insuspension and preloaded with fura-2 to a hypotonic buffer (200mOsm/liter) and measured [Ca2�]i. NMCs maintain a stable [Ca2�]iwhen in isotonic buffer but show a 2-fold increase in [Ca2�]i whenin hypotonic buffer [supporting information (SI) Fig. 8A, P � 0.05,n � 3]. TRPV4-overexpressing cells show a 6-fold increase in[Ca2�]i when exposed to hypotonicity, a response that was impairedwhen NMC-Trpv4-shRNA cells were used (SI Fig. 8A, P � 0.05, n �
Fig. 1. TRPV4 message and protein are expressed in cholangiocytes. (A) RT-PCRfor Trpv4 on normal mouse cholangiocytes cell line (NMC), freshly isolated mousecholangiocytes (FIMC), isolated rat bile ducts, normal rat cholangiocytes in pri-mary culture (NRC), and NMC-overexpressing TRPV4 (NMC-Trpv4). Kidney RNAwasusedaspositivecontrol. (B)Thirtymicrogramsoftotalhomogenateofratbileducts and NRCs was loaded onto 7.5% SDS–polyacrylamide gels. Kidney and CHOcells were used as positive and negative controls, respectively. (C) Ten microgramsof NMC, NMC-Trpv4, and NMC-Trpv4-shRNA total homogenates was immuno-blotted with anti-TRPV4. Anti-actin immunoblots were used as loading controls.Anti-TRPV4 immunoblot of NMC-Trpv4 homogenates before (�) or after (�)digestion with peptide/N-glycosidase (PNGaseF).
Fig. 2. TRPV4 is expressed on cholangiocyte cilia. (A) Confocal microscopy onnormal and Trpv4-transfected NMCs. Cells were stained with anti-acetylated�-tubulin (�-tub) in red and anti-TRPV4 in green. (B) Normal rat liver tissueshowing a longitudinal cut of an intrahepatic bile duct and the expression ofTRPV4 on cholangiocyte cilia (yellow). (C) Immunogold electron microscopyindicates the presence of TRPV4 on cholangiocyte cilia in IBDUs isolated from ratliver (arrows). (D) Confocal immunofluorescence showing rat cholangiocytes‘‘Peel off’’ isolated cilia costained with anti-acetylated �-tubulin (red) and anti-TRPV4 (green).
Gradilone et al. PNAS � November 27, 2007 � vol. 104 � no. 48 � 19139
3). The hypotonicity-induced increases in [Ca2�]i depended on thepresence of Ca2� in the extracellular media (SI Fig. 8B). As apositive control, we measured the effect of the TRPV4 agonist,4�PDD, on intracellular Ca2�, and we found an increase in [Ca2�]ithat was dose-dependent (SI Fig. 8C). These data suggest thatcholangiocyte TRPV4 is involved in a hypotonicity-induced Ca2�
influx from the extracellular media.
Luminal Hypotonicity Induces TRPV4- and Ciliary-Dependent Changesin [Ca2�]i Levels in Cholangiocytes of Microperfused IBDUs. To test thehypothesis that ciliary TRPV4 could sense luminal changes inosmolarity, we used a previously developed physiologically relevantexperimental model: microperfused IBDUs prepared from rat liverby microdissection (7, 32). To avoid a potential confoundingmechanical response by cholangiocyte cilia, luminal perfusion waskept at a rate that would not stimulate a mechanical ciliary response[�20 nl/min (7)]. Because we had previously observed that osmot-ically induced water movement into or out of the luminal space ofIBDUs occurs very rapidly depending on the osmotic gradientbetween the lumen and the bathing buffer (32), we inducedintraluminal hypotonicity by bathing the exterior of IBDUs inhypotonic KRB (100 mOsm/liter). This maneuver resulted in arapid decrease in fluorescence of a nonabsorbable fluorescent
marker in the luminal perfusate, consistent with a drop in luminalosmolarity (Fig. 3A) and validating this experimental approach tomodify the luminal tonicity of IBDUs. When cholangiocytes ofIBDUs were preloaded with a Ca2�-sensitive fluorescent dye(fluo-4), we found that a decrease in intraluminal tonicity caused anincrease in dye fluorescence (i.e., an increase in [Ca2�]i) (Fig. 3 Aand B). The hypotonicity-induced increase in [Ca2�]i levels was45 � 9.9% compared with basal isotonic conditions. To addresswhether cholangiocyte cilia were involved in this hypotonicity-induced [Ca2�]i signaling response, we deciliated IBDUs withchloral hydrate (ClHy), a technique described and validated by usand others (7, 9, 33). Functional studies showed that in IBDUstreated with ClHy, the hypotonicity-induced rise in [Ca2�]i wasreduced by 93.8 � 10.6% (Fig. 3D), suggesting that cholangiocytecilia are involved in the transmission of hypotonic stimuli into an[Ca2�]i response.
To explore whether TRPV4 is involved in the hypotonicity-induced [Ca2�]i response, we inhibited endogenous Trpv4 in IBDUsby a specific siRNA. As shown in Fig. 3C, both Trpv4 mRNA andprotein decreased by �70% with this reagent. The hypotonicity-induced increase in [Ca2�]i of IBDUs treated with a scrambledsiRNA was not different from that in normal IBDUs. In IBDUstreated with Trpv4-siRNA, the [Ca2�]i response was reduced by90.1 � 4.7% (Fig. 3D), supporting TRPV4 involvement in thehypotonicity-induced increase in [Ca2�]i level.
Hypotonicity Induces TRPV4- and Ciliary-Dependent Luminal Alkalin-ization and ATP Release. To determine a possible physiological roleof luminal tonicity sensation by ciliary TRPV4, we measuredchanges in luminal pH as a reflection of bicarbonate secretion usingthe cell-impermeable pH-sensitive dye, BCECF-dextran (32, 34,35). As shown in Fig. 4, hypotonicity, as expected, induced adecrease in the luminal isosbestic fluorescence intensity because ofwater movement into the ductal lumen. As a consequence of thisstimulus, control IBDUs showed an increase in luminal pH of0.23 � 0.04 unit/min (Fig. 4 B and C). Treatment of IBDUs with theTrpv4-siRNA or with ClHy decreased the hypotonicity-inducedluminal pH increase by 71 � 17% and 89 � 3%, respectively. Theseexperiments demonstrate that luminal hypotonicity sensed by cil-iary TRPV4 contributes to regulation of bicarbonate secretion bycholangiocytes.
To address mechanisms linking the hypotonicity-induced activa-tion of ciliary TRPV4 with bicarbonate secretion, we tested thehypothesis that ATP release mediates this phenomenon. As shownin Fig. 5A, hypotonicity induced a 5-fold increase in ATP release inNMCs compared with isotonic conditions. This result is consistentwith data previously reported on Mz-ChA-1 cells (36). Cellsoverexpressing TRPV4 showed a 7-fold increase in ATP releasewhen exposed to hypotonicity, an effect that was reduced by 70%when cells were cotransfected with a Trpv4-shRNA (Fig. 5A).TRPV4 activation by 4�PDD induced a 3-fold increase in extra-cellular ATP (Fig. 5B). Furthermore, the hypotonicity-inducedincrease in luminal pH was prevented when IBDUs were perfusedwith suramin, a purinergic receptor antagonist (Fig. 5C). Both the
Fig. 3. Luminal hypotonicity induces an [Ca2�]i increase in a TRPV4- andcilia-dependent manner. (A) Representative traces of IBDUs microperfused withKRB containing 1 mmol/liter of FS (gray) or IBDUs loaded with fluo-4 (black). Thetonicity of the bath perfusate was changed and fluorescence intensity followedover the time. (B) Fluorescence tracing of fluo-4 loaded IBDUs perfused withisotonicmannitol-KRBbuffer (gray)orhypotonicKRBbuffer (black)over time. (C)Trpv4 qRT-PCR normalized to 18s RNA and Western blot analysis on IBDUsincubated overnight with scrambled or Trpv4-siRNA. (D) Fluo-4 fluorescencechanges inducedbyhypotonicityoncontrol,Trpv4-siRNAandClHytreatedIBDUs.(*, P � 0.01, n � 6.)
Fig. 4. Luminal tonicity changes in IBDUsinduce cholangiocyte bicarbonate secretionin a TRPV4- and cilia-dependent manner.Changes in ductular pH (�pH) induced byhypotonicity were examined by using lumi-nal perfusion with BCECF-dextran. (A) WhenIBDUs were exposed to hypotonicity, watermoved into the ductal lumen inducing a de-crease in the isosbestic fluorescence of thedye. (B) Control IBDUs (F) showed a net in-crease in luminal pH over time that was pre-vented in the siRNA- (■ ) or ClHy- (�) treated IBDUs. (C) Slopes of the first portion of the curves after the addition of hypotonic buffer were calculated and expressedas �pH units/min. (*, P � 0.05; #, P � 0.01 compared with control group, n � 4.)
19140 � www.pnas.org�cgi�doi�10.1073�pnas.0705964104 Gradilone et al.
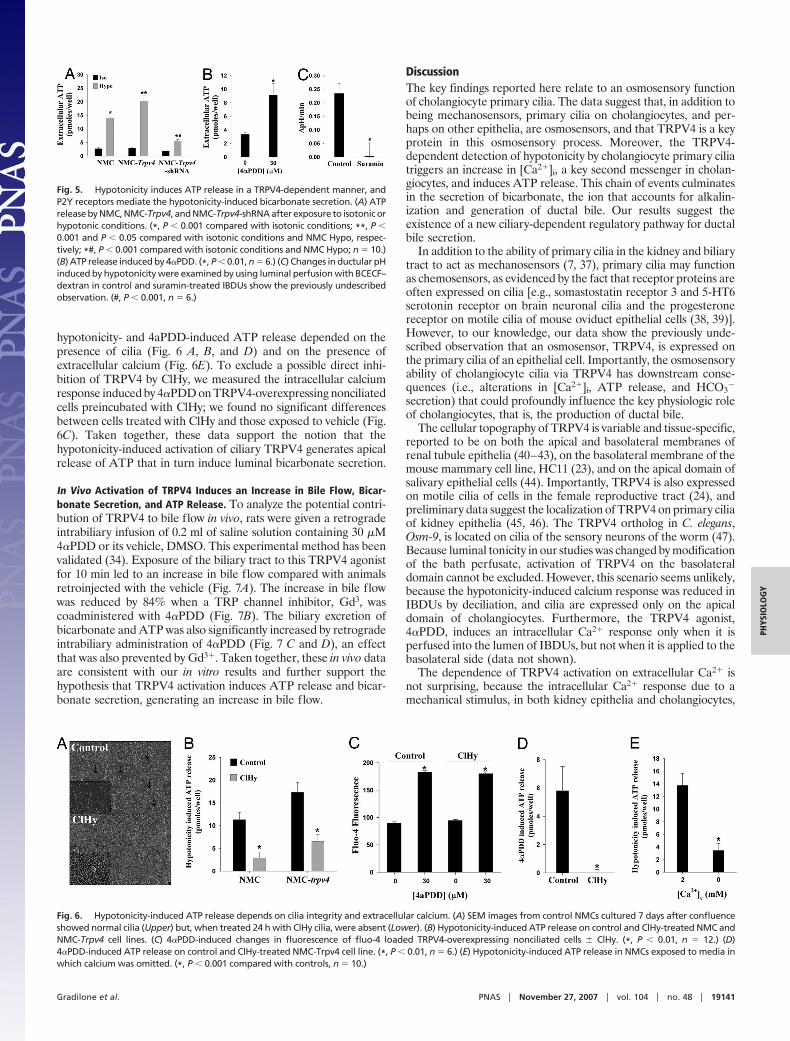
hypotonicity- and 4aPDD-induced ATP release depended on thepresence of cilia (Fig. 6 A, B, and D) and on the presence ofextracellular calcium (Fig. 6E). To exclude a possible direct inhi-bition of TRPV4 by ClHy, we measured the intracellular calciumresponse induced by 4�PDD on TRPV4-overexpressing nonciliatedcells preincubated with ClHy; we found no significant differencesbetween cells treated with ClHy and those exposed to vehicle (Fig.6C). Taken together, these data support the notion that thehypotonicity-induced activation of ciliary TRPV4 generates apicalrelease of ATP that in turn induce luminal bicarbonate secretion.
In Vivo Activation of TRPV4 Induces an Increase in Bile Flow, Bicar-bonate Secretion, and ATP Release. To analyze the potential contri-bution of TRPV4 to bile flow in vivo, rats were given a retrogradeintrabiliary infusion of 0.2 ml of saline solution containing 30 �M4�PDD or its vehicle, DMSO. This experimental method has beenvalidated (34). Exposure of the biliary tract to this TRPV4 agonistfor 10 min led to an increase in bile flow compared with animalsretroinjected with the vehicle (Fig. 7A). The increase in bile flowwas reduced by 84% when a TRP channel inhibitor, Gd3, wascoadministered with 4�PDD (Fig. 7B). The biliary excretion ofbicarbonate and ATP was also significantly increased by retrogradeintrabiliary administration of 4�PDD (Fig. 7 C and D), an effectthat was also prevented by Gd3�. Taken together, these in vivo dataare consistent with our in vitro results and further support thehypothesis that TRPV4 activation induces ATP release and bicar-bonate secretion, generating an increase in bile flow.
DiscussionThe key findings reported here relate to an osmosensory functionof cholangiocyte primary cilia. The data suggest that, in addition tobeing mechanosensors, primary cilia on cholangiocytes, and per-haps on other epithelia, are osmosensors, and that TRPV4 is a keyprotein in this osmosensory process. Moreover, the TRPV4-dependent detection of hypotonicity by cholangiocyte primary ciliatriggers an increase in [Ca2�]i, a key second messenger in cholan-giocytes, and induces ATP release. This chain of events culminatesin the secretion of bicarbonate, the ion that accounts for alkalin-ization and generation of ductal bile. Our results suggest theexistence of a new ciliary-dependent regulatory pathway for ductalbile secretion.
In addition to the ability of primary cilia in the kidney and biliarytract to act as mechanosensors (7, 37), primary cilia may functionas chemosensors, as evidenced by the fact that receptor proteins areoften expressed on cilia [e.g., somastostatin receptor 3 and 5-HT6serotonin receptor on brain neuronal cilia and the progesteronereceptor on motile cilia of mouse oviduct epithelial cells (38, 39)].However, to our knowledge, our data show the previously unde-scribed observation that an osmosensor, TRPV4, is expressed onthe primary cilia of an epithelial cell. Importantly, the osmosensoryability of cholangiocyte cilia via TRPV4 has downstream conse-quences (i.e., alterations in [Ca2�]i, ATP release, and HCO3
�
secretion) that could profoundly influence the key physiologic roleof cholangiocytes, that is, the production of ductal bile.
The cellular topography of TRPV4 is variable and tissue-specific,reported to be on both the apical and basolateral membranes ofrenal tubule epithelia (40–43), on the basolateral membrane of themouse mammary cell line, HC11 (23), and on the apical domain ofsalivary epithelial cells (44). Importantly, TRPV4 is also expressedon motile cilia of cells in the female reproductive tract (24), andpreliminary data suggest the localization of TRPV4 on primary ciliaof kidney epithelia (45, 46). The TRPV4 ortholog in C. elegans,Osm-9, is located on cilia of the sensory neurons of the worm (47).Because luminal tonicity in our studies was changed by modificationof the bath perfusate, activation of TRPV4 on the basolateraldomain cannot be excluded. However, this scenario seems unlikely,because the hypotonicity-induced calcium response was reduced inIBDUs by deciliation, and cilia are expressed only on the apicaldomain of cholangiocytes. Furthermore, the TRPV4 agonist,4�PDD, induces an intracellular Ca2� response only when it isperfused into the lumen of IBDUs, but not when it is applied to thebasolateral side (data not shown).
The dependence of TRPV4 activation on extracellular Ca2� isnot surprising, because the intracellular Ca2� response due to amechanical stimulus, in both kidney epithelia and cholangiocytes,
Fig. 5. Hypotonicity induces ATP release in a TRPV4-dependent manner, andP2Y receptors mediate the hypotonicity-induced bicarbonate secretion. (A) ATPrelease by NMC, NMC-Trpv4, and NMC-Trpv4-shRNA after exposure to isotonic orhypotonic conditions. (*, P � 0.001 compared with isotonic conditions; **, P �0.001 and P � 0.05 compared with isotonic conditions and NMC Hypo, respec-tively; *#, P � 0.001 compared with isotonic conditions and NMC Hypo; n � 10.)(B) ATP release induced by 4�PDD. (*, P � 0.01, n � 6.) (C) Changes in ductular pHinduced by hypotonicity were examined by using luminal perfusion with BCECF–dextran in control and suramin-treated IBDUs show the previously undescribedobservation. (#, P � 0.001, n � 6.)
Fig. 6. Hypotonicity-induced ATP release depends on cilia integrity and extracellular calcium. (A) SEM images from control NMCs cultured 7 days after confluenceshowed normal cilia (Upper) but, when treated 24 h with ClHy cilia, were absent (Lower). (B) Hypotonicity-induced ATP release on control and ClHy-treated NMC andNMC-Trpv4 cell lines. (C) 4�PDD-induced changes in fluorescence of fluo-4 loaded TRPV4-overexpressing nonciliated cells � ClHy. (*, P � 0.01, n � 12.) (D)4�PDD-induced ATP release on control and ClHy-treated NMC-Trpv4 cell line. (*, P � 0.01, n � 6.) (E) Hypotonicity-induced ATP release in NMCs exposed to media inwhich calcium was omitted. (*, P � 0.001 compared with controls, n � 10.)
Gradilone et al. PNAS � November 27, 2007 � vol. 104 � no. 48 � 19141
PHYS
IOLO
GY
and the odorant receptors in olfactory epithelial, all depend onextracellular Ca2� (7, 48, 49).
Studies carried out on Trpv4-null mice show that TRPV4 isnecessary for the maintenance of osmotic equilibrium, and it isconceivable that TRPV4 acts as an osmotic sensor in the centralnervous system (16). TRPV4 also plays a role in maintenance ofcellular osmotic homeostasis (50). Here we show the function ofTRPV4 as osmosensor in a different context, i.e., not in theregulation of global body osmolarity but in the specific sensation ofbile tonicity, leading to the secretion of ions such as bicarbonate inresponse to a decrease in the tonicity of bile. Although activationof TRPV4 has been reported to induce an increase in paracellularpermeability for small solutes in an epithelial cell line, this effectoccurs over hours and, thus, is not likely relevant to our findings,which occur much more rapidly (23). The mechanism that links thehypotonicity-induced TRPV4-mediated [Ca2�]i increase with thestimulation of bicarbonate secretion appears to involve ciliary- andTRPV4-dependent hypotonicity-induced ATP release. ATP ispresent in bile in physiological concentrations (51). Moreover,cholangiocytes express a number of P2Y and P2X nucleotidereceptors on their apical domain, and their stimulation by ATPinduces apical Cl� secretion and provokes alkalinization of bile viaCl�/HCO3
� exchange activity mediated by AE2 (35, 36, 52–54).Interestingly, an intracellular P2X receptor required for osmoreg-ulation in Dictyostelium discoideum was recently reported (55).Based on this information, we speculated that ATP released by thehypotonic activation of ciliary TRPV4 would activate purinergicreceptors on the apical domain of cholangiocytes, which might theninfluence cholangiocyte transport processes and ultimately ductalbile secretion. Our data support this chain of events.
The physiological implications of an osmosensory function ofcholangiocyte cilia are unclear. Bile is considered isotonic, but notechnology permits measurements of variations in bile tonicityduring its long passage through the ductal lumen, from the hepa-tocyte canaliculus to the common bile duct; indeed, measurementsof bile tonicity can be made only on bile collected at the most distalend of the biliary tree (i.e., the common bile duct). It is known thatcholangiocytes can absorb glucose and bile salts from primary bilegenerated by hepatocytes (1, 32). Thus, it seems plausible that theseabsorptive processes can generate transient decreases in ductal biletonicity that could be sensed by cholangiocyte cilia. As shown by ourin vivo and in vitro results, the activation of ciliary TRPV4, in turn,induces a bicarbonate secretory response, mediated by the apicalrelease of ATP and purinergic receptor activation. This secretionwould restore the tonicity of ductal bile and also generate watersecretion as indicated by the increased bile flow.
In summary, our study shows that primary cilia on cholangiocytesmay function as osmosensors, and that ciliary osmosensation,mediated by a mechanism that involves an increase in [Ca2�]i andapical ATP release, induces luminal bicarbonate secretion. Thephysiological, pathophysiological, and therapeutic implications ofthis phenomenon remain to be fully elucidated.
Materials and MethodsAnimals. Male Sprague–Dawley rats (225–250 g) were maintainedon a standard diet. In vivo experiments using a validated retrogradeintrabiliary infusion model were performed as described (34). Allexperimental procedures were approved by the Animal Use andCare Committee of the Mayo Foundation.
Solutions. The composition of Ringer-HCO3� buffer (KRB) was as
follows: 120.0 mmol/liter of NaCl, 5.9 mmol/liter of KCl, 1.2mmol/liter of Na2HPO4, 1.0 mmol/liter of MgSO4, 25.0 mmol/literof NaHCO3, 1.25 mmol/liter of CaCl2, and 5.0 mmol/liter ofD-glucose, pH 7.4. For tonicity-change studies, isotonic KRB wasprepared with 40 mmol/liter of NaCl and completed to isotonicitywith mannitol. For hypotonic KRB, addition of mannitol wasomitted. In calcium-free KRB, CaCl2 was removed.
Cell Culture. NMCs were grown in Eagle’s modified essentialmedium supplemented with 10% (vol/vol) FBS and penicillin(100 units/ml)/streptomycin (100 �g/ml). NRCs were grown asdescribed (56).
Isolated IBDUs. IBDUs were isolated from normal rat liver asdescribed (32).
RT-PCR. Specific oligonucleotides were synthesized based on thepublished sequence for rat Trpv4 5�-TACCACGGTGGACTAC-CTGAG-3� (forward) and 5�-CATGATGCTGTAGGTC-CCTGT-3� (reverse).
Trpv4 Cloning, shRNA Constructs, and Stable Transfection. Trpv4 wascloned from NMC total RNA. RT-PCR amplification was achievedwith Trpv4 specific primers (5�-CCAAGCTTGCCACCATGGCA-GATCCTGGTGATGGT-3� and 5�-CCGGATCCAGTGGGGC-ATCGTCCGT-3�), and cloned in frame into the pcDNA6/myc-Hismammalian expression vector. After selection, cells were cotrans-fected with the Trpv4-shRNA expressing vector. Top- and bottom-strand oligonucleotides corresponding to the nucleotide sequenceof the target site (AACATGAAGGTCTGTGACGAG) werecloned into pSilencer3.1-H1 puro shRNA vector.
Trpv4 Silencing by siRNA and Chemical Deciliation on IBDUs. IsolatedIBDUs were cultured for 24 h with 20 nmol/liter of functional orscrambled Trpv4-siRNA (target sequence, 5�-AAGAACTCAG-GCACAGATGAA-3�) or with 4 mmol/liter of ClHy (57).
Measurement of [Ca2�]i Level in Suspended Cholangiocytes. Cells(1.0 106) were washed and loaded with 3 �M Fura-2/AM for 30min at 37°C. After washing, cells were resuspended in isotonic orhypotonic KRB and fluorescence-quantified. Excitation was per-formed at both 340 and 380 nm, with emission determined at 520nm. [Ca2�]i was calculated from the ratios of these values as
Fig. 7. In vivo activation of TRPV4 induced an increased bile flow and bicarbonate and ATP secretion. (A) Effect of intrabiliary administration of 30 �M 4�PDD or itsvehicle DMSO on bile flow. (B) Effect of 100 �M Gd3� on the 4�PDD-induced increase in bile flow. (C and D) Biliary bicarbonate and ATP excretion on control vs.4�PDD-treated rats. (*, P � 0.01, n � 4.)
19142 � www.pnas.org�cgi�doi�10.1073�pnas.0705964104 Gradilone et al.
described (58). Alternatively, cells were seeded on 96-well plates,loaded with fluo-4, and, after washing, stimulated with KRBcontaining 4�PDD or its vehicle DMSO. In related experiments,cells were preincubated with ClHy before Fluo-4 loading.
Microperfusion of IBDUs. Individual isolated IBDUs were mountedin a custom-designed temperature-controlled chamber on the stageof an inverted fluorescence microscope, as described (32). Thelumen was perfused at very low rate (20 nl/min). The externalsurface of the perfused IBDU was simultaneously bathed withisotonic or hypotonic KRB.
Measurement of Water Movement into the Lumen of IBDUs. Thelumen of IBDUs was perfused with KRB containing 1 mmol/literof fluorescein-5-(6)-sulfonic acid, trisodium salt (FS), a fluorescentvolume marker. To decrease luminal tonicity, water movement intothe lumen was induced by changing the tonicity of the bath solutionand measured as described (32).
Measurement of [Ca2�]i Level in Cholangiocytes of MicroperfusedIBDUs. Changes in [Ca2�]i levels of cholangiocytes of KRB-perfusedIBDUs were detected by using the cell-permeant Ca2�-sensitivefluorescent probe Fluo-4 AM, as we described in detail (35).
Measurement of pH in the IBDU Lumen. The luminal pH of perfusedIBDUs was measured by using the cell-impermeant pH-sensitivedye, BCECF-dextran, as described (32). IBDUs were perfused withHCO3
�-free buffer containing BCECF-dextran (pH 7.2) and simul-taneously bathed with isotonic or hypotonic KRB. After excitationat 495 nm, luminal pH was measured as the ratio of emissionintensities at 530 nm (pH- and concentration-sensitive) and 440 nm(only concentration-sensitive), respectively. In situ calibrationcurves were generated at the end of each experiment to convertfluorescence-excitation ratios (F495/F440) to pH values (32, 35).
ATP Release. NMC, NMC-trpv4, and NMC-Trpv4-shRNA werecultured 7 days after confluence in 24-well plates. After washingthree times with KRB, cells were incubated for 2 min in 200 �l ofisotonic KRB. Two hundred microliters of isotonic KRB or waterwas added and incubated for 4 min. The viability of the cells was notaffected. Alternatively, cells were incubated with KRB containing4�PDD or its vehicle DMSO. Media were then collected, centri-fuged for 10 min 500 g, and passed to clean tubes. ATPconcentration was measured with a luciferin–luciferase biolumi-nescence assay kit.
Immunoblot Analysis of TRPV4. Protein fractions were subjected to7.5% SDS–polyacrylamide gel and transferred to nitrocellulosemembranes. After blocking, blots were incubated overnight at 4°Cwith rabbit affinity-purified polyclonal TRPV4 antibody (1:5,000,Alomone Labs), washed, and incubated 1 h at room temperaturewith HRP-conjugated goat anti-rabbit secondary antibody (1:5,000dilution). For protein detection, ECL system was used. Selectedsamples were digested with 10 �g/ml peptide/N-glycosidase F.
Immunofluorescence Confocal Microscopy. Samples were incubatedwith antibodies against acetylated �-tubulin (1:500, Sigma–Aldrich)and TRPV4 (1:100) overnight at 4°C followed by incubation for 1 hwith fluorescent secondary antibodies (1:100). Nuclei were stainedwith DAPI.
Immunogold and Scanning Electron Microscopy. Immunogold elec-tron microscopy of IBDUs and SEM of NMCs were performed asdescribed (29, 59).
Cilia Isolation. Cilia were isolated from cultured cells by using thepeeloff technique as described (29).
This work was supported by the PKD Foundation (Grant 132a2f, to S.A.G.),the National Institutes of Health (Grant DK24031-29, to N.F.L.), theRamon Areces Foundation (J.M.B.), and the Mayo Foundation.
Mol Reprod Dev 71:444–452.25. Roberts SK, Yano M, Ueno Y, Pham L, Alpini G, Agre P, LaRusso NF (1994) Proc Natl Acad
Sci USA 91:13009–13013.26. Vroman B, LaRusso NF (1996) Lab Invest 74:303–313.27. Liedtke W, Friedman JM (2003) Proc Natl Acad Sci USA 100:13698–13703.28. Xu H, Fu Y, Tian W, Cohen DM (2006) Am J Physiol 290:F1103–F1109.29. Huang BQ, Masyuk TV, Muff MA, Tietz PS, Masyuk AI, Larusso NF (2006) Am J Physiol
291:G500–G509.30. Roman RM, Wang Y, Fitz JG (1996) Am J Physiol 271:G239–G248.31. Feranchak AP, Doctor RB, Troetsch M, Brookman K, Johnson SM, Fitz JG (2004) Gastroen-
281:G1059–G10567.36. Roman RM, Feranchak AP, Salter KD, Wang Y, Fitz JG (1999) Am J Physiol 276:G1391–G1400.37. Praetorius HA, Frokiaer J, Nielsen S, Spring KR (2003) J Membr Biol 191:193–200.38. Michaud EJ, Yoder BK (2006) Cancer Res 66:6463–6467.39. Teilmann SC, Clement CA, Thorup J, Byskov AG, Christensen ST (2006) J Endocrinol
Plomann M, Heller S (2006) J Biol Chem 281:18753–18762.41. Taniguchi J, Tsuruoka S, Mizuno A, Sato J, Fujimura A, Suzuki M (2007) Am J Physiol
292:F667–F673.42. Tian W, Salanova M, Xu H, Lindsley JN, Oyama TT, Anderson S, Bachmann S, Cohen DM
(2004) Am J Physiol 287:F17–F24.43. Wu L, Gao X, Brown RC, Heller S, O’Neil RG (2007) Am J Physiol, 293:F1699-F1713.44. Liu X, Bandyopadhyay B, Nakamoto T, Singh B, Liedtke W, Melvin JE, Ambudkar I (2006)
J Biol Chem 281:15485–15495.45. Giamarchi A, Padilla F, Coste B, Raoux M, Crest M, Honore E, Delmas P (2006) EMBO Rep
7:787–793.46. Kottgen M (2007) Biochim Biophys Acta 1772:836–850.47. Qin H, Burnette DT, Bae YK, Forscher P, Barr MM, Rosenbaum JL (2005) Curr Biol 15:1695–1699.48. Nauli SM, Alenghat FJ, Luo Y, Williams E, Vassilev P, Li X, Elia AE, Lu W, Brown EM, Quinn
SJ, et al. (2003) Nat Genet 33:129–137.49. Liu G, Badeau RM, Tanimura A, Talamo BR (2006) J Neurochem 96:1591–1605.50. Liedtke W (2006) J Endocrinol 191:515–523.51. Chari RS, Schutz SM, Haebig JE, Shimokura GH, Cotton PB, Fitz JG, Meyers WC (1996)
Am J Physiol 270:G246–G252.52. Zsembery A, Spirli C, Granato A, LaRusso NF, Okolicsanyi L, Crepaldi G, Strazzabosco
M (1998) Hepatology 28:914–920.53. Schlenker T, Romac JM, Sharara AI, Roman RM, Kim SJ, LaRusso N, Liddle RA, Fitz JG
(1997) Am J Physiol 273:G1108–G11017.54. Banales JM, Prieto J, Medina JF (2006) World J Gastroenterol 12:3496–3511.55. Fountain SJ, Parkinson K, Young MT, Cao L, Thompson CR, North RA (2007) Nature
448:200–203.56. Muff MA, Masyuk TV, Stroope AJ, Huang BQ, Splinter PL, Lee SO, Larusso NF (2006)