Page 1
Protein Crystallization:
A Method to Study Phenylalanine
Mutation Effects on Protein Efficiency
Presented By: Christine Meyer
Mentor: Dr. Steven Berry
Department of Chemistry & Biochemistry
Spring 2013 Senior Symposium
Page 2
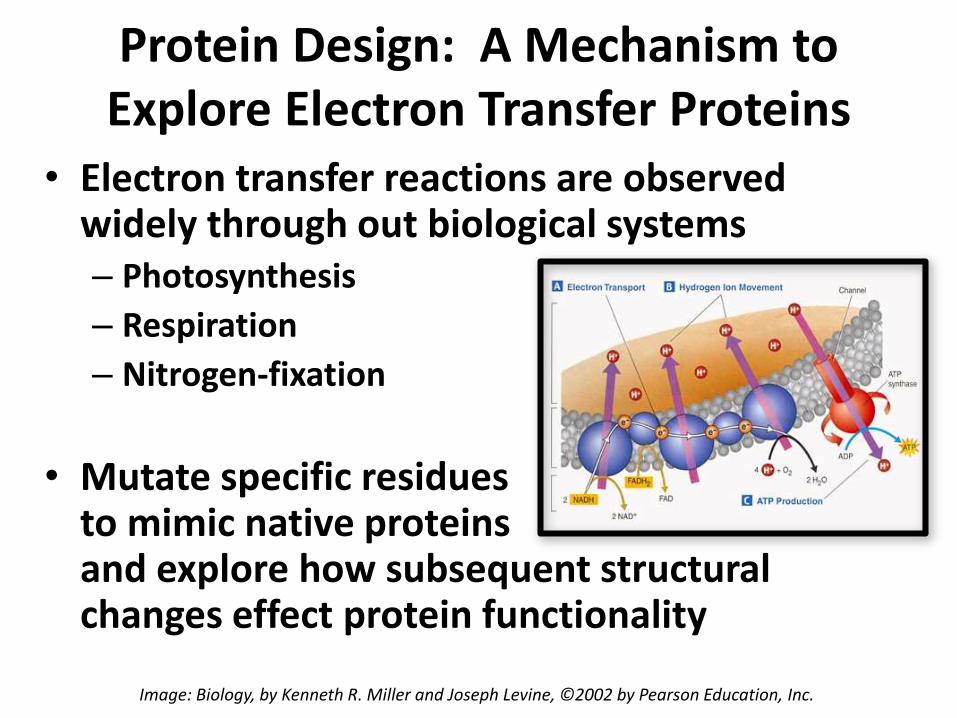
Protein Design: A Mechanism to Explore Electron Transfer Proteins
• Electron transfer reactions are observed widely through out biological systems– Photosynthesis
– Respiration
– Nitrogen-fixation
• Mutate specific residues to mimic native proteins and explore how subsequent structural changes effect protein functionality
Image: Biology, by Kenneth R. Miller and Joseph Levine, ©2002 by Pearson Education, Inc.
Page 3
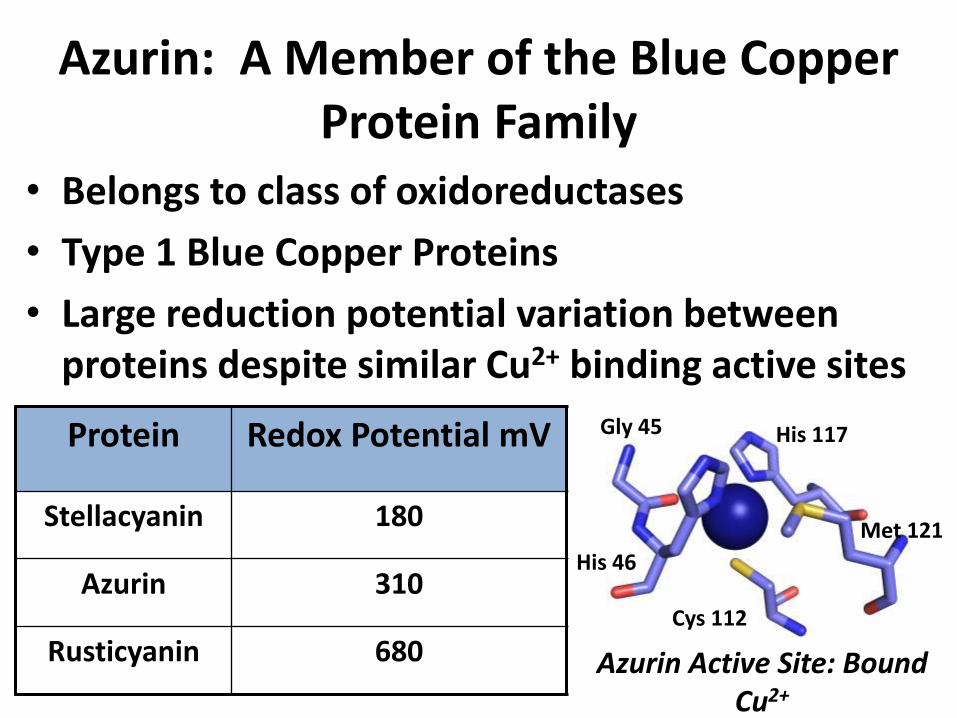
Azurin: A Member of the Blue Copper Protein Family
• Belongs to class of oxidoreductases
• Type 1 Blue Copper Proteins
• Large reduction potential variation between proteins despite similar Cu2+ binding active sites
Protein Redox Potential mV
Stellacyanin 180
Azurin 310
Rusticyanin 680 Azurin Active Site: Bound Cu2+
His 46
Cys 112
Gly 45 His 117
Met 121
Page 4
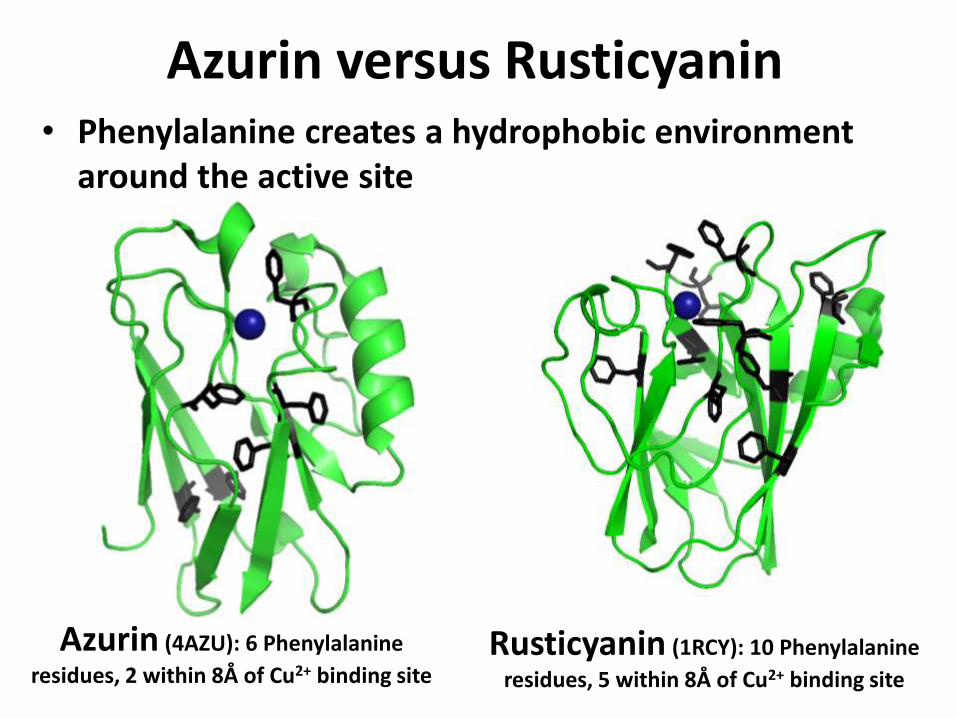
Azurin versus Rusticyanin
Azurin (4AZU): 6 Phenylalanine
residues, 2 within 8Å of Cu2+ binding siteRusticyanin (1RCY): 10 Phenylalanine
residues, 5 within 8Å of Cu2+ binding site
• Phenylalanine creates a hydrophobic environment around the active site
Page 5
Azurin Phenylalanine Mutant Series
• Single Mutants– Leu33Phe
– Met44Phe
– Leu86Phe
• Double Mutant:– Leu33Phe & Met44Phe
– Met44Phe & Leu86Phe
• Triple Mutant:– Leu33Phe, Met44Phe,
Leu86PheS.M. Berry et al. Journal of Inorganic Biochemistry 104 (2010) 1071-1078
Residue Substitution ΔE, mV (±5)
ΔLeu33Phe
WTL33F 12
M44FL33F/M44F 10.5
M44F/L86FL33F/M44F/L86F 16.5
ΔMet44Phe
WTM44F 32.5
L33FL33F/M44F 31
L33F/L86FL33F/M44F/L86F 39
ΔLeu86Phe
WTL86F 27.5
L33FL33F/L86F 27
L33F/M44F L33F/M44F/L86F 35
• Key residues in secondary coordination sphere were mutated to Phe to resemble native Rusticyanin secondary coordination sphere
Page 6
Experimental Procedure
• Grow crystals to identify structural reason for reproducible redox potential change upon introduction of Phe residue
• Begin by growing Phe Azurin crystals using the hanging drop method
Page 7
Growing Phe Azurin Crystals• Crystal Box: The Hanging Drop method
Page 8
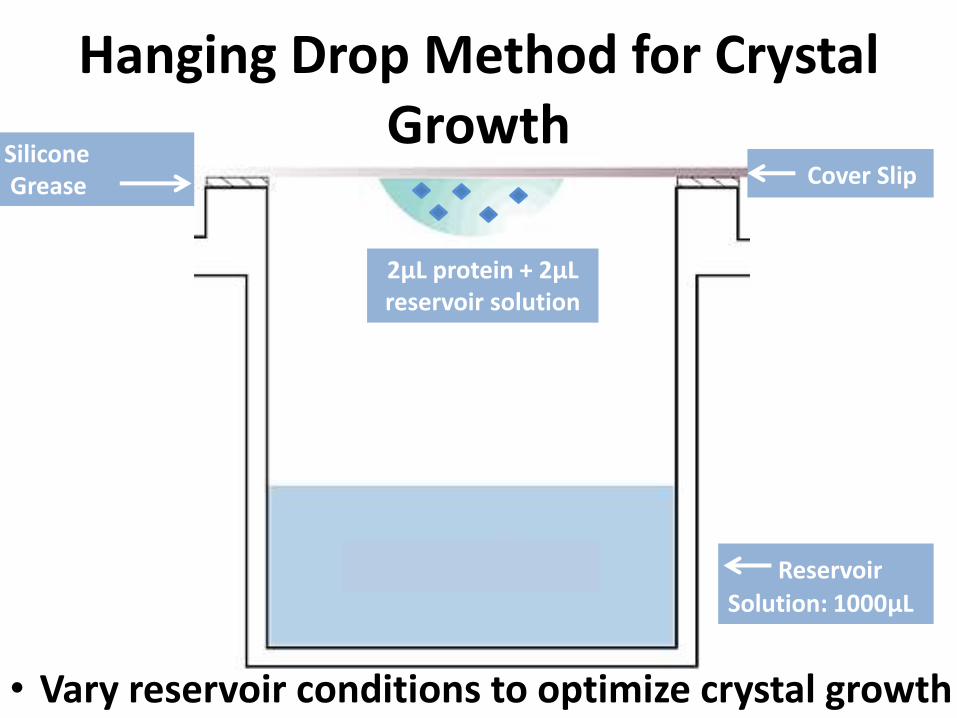
Hanging Drop Method for Crystal Growth
Cover Slip
2μL protein + 2μL reservoir solution
SiliconeGrease
Reservoir
Solution: 1000μL
• Vary reservoir conditions to optimize crystal growth
Page 9
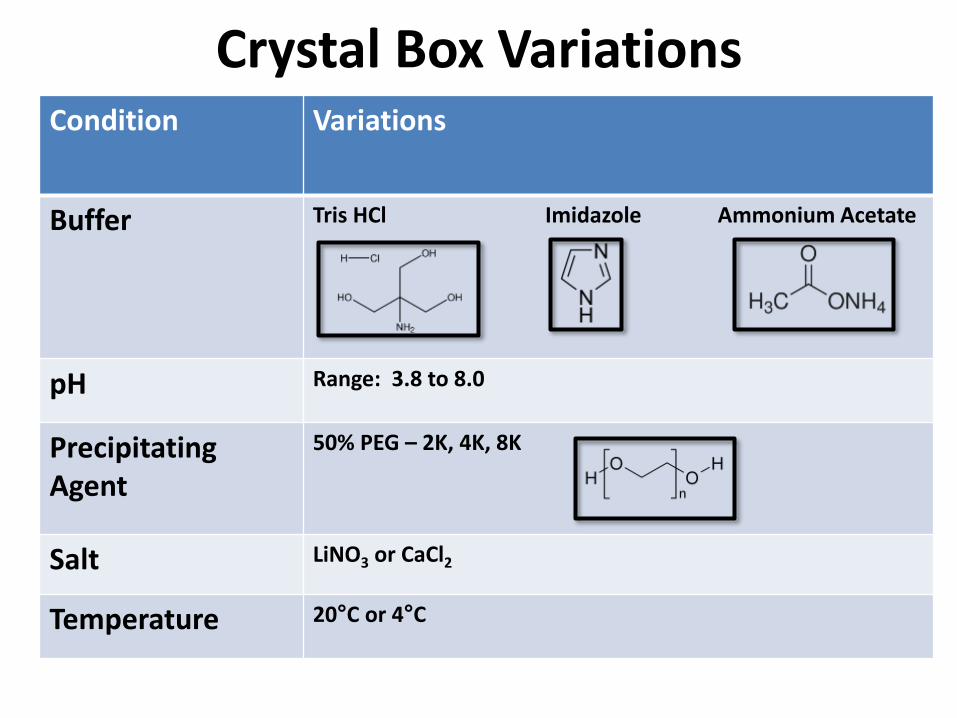
Crystal Box VariationsCondition Variations
Buffer Tris HCl Imidazole Ammonium Acetate
pH Range: 3.8 to 8.0
Precipitating Agent
50% PEG – 2K, 4K, 8K
Salt LiNO3 or CaCl2
Temperature 20°C or 4°C
Page 10
Crystal Box Reservoir Variations
Decreasing [50%-PEG]
Increasing [Salt]
30% 26% 22% 18% 14% 10%
0.03 M
0.09 M
0.15 M
0.21 M
• 100 mM Tris-HCl, 5 mM CuSO4, plus varied PEG and CaCl2
Page 11
Crystal Pictures
Increasing [CaCl2]
Decreasing [50%-PEG]
• Less PEG resulted in slower crystal growth• Varying CaCl2 resulted in different crystal sizes
Page 12
Example of Ordered Crystals
Page 13
Dehydration: Improved Crystal Order• A technique used to decrease mosaicity
– Mosaicity: skewed arrangement of molecules
– Before Dehydration: Range 1.0-1.5°
– After Dehydration: Range 0.3-0.5°
Before Dehydration, high mosaicity
After Dehydration,low mosaicity
H2O
H2O
H2O
H2O
+ Glycerol
& PEG
Page 14
Crystal Pinning
• Pin crystal on loop
• Flash freeze in liquid nitrogen
• Store in liquid nitrogen
Page 15
Crystal Diffraction: Rigaku Rapid II©
Nitrogen Stream
Pinned CrystalCollimator
Cu Source
Page 16
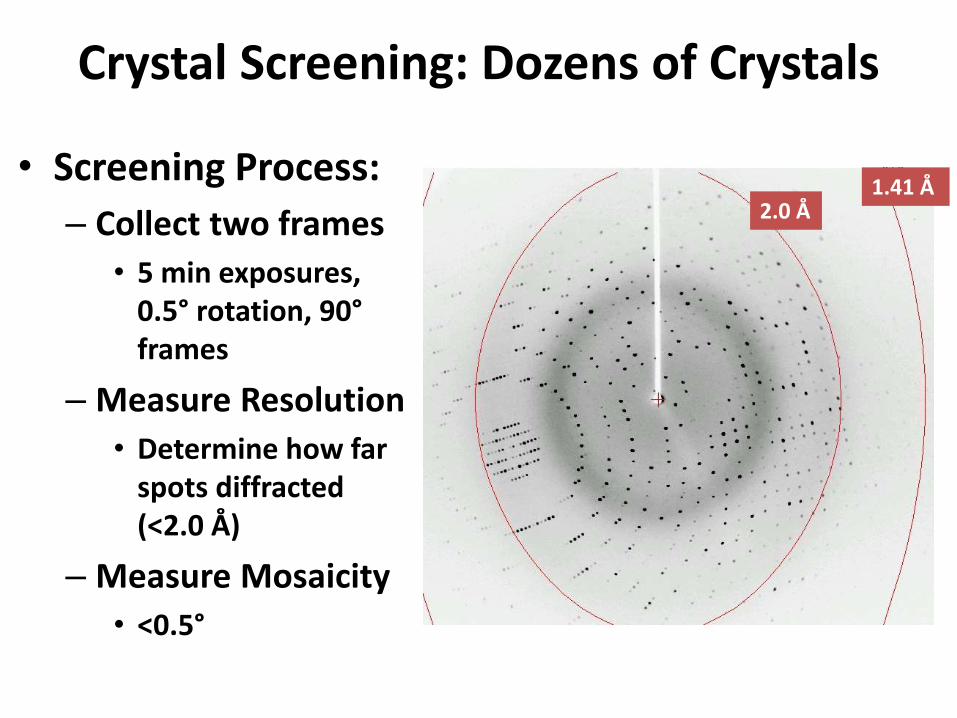
Crystal Screening: Dozens of Crystals
• Screening Process:
– Collect two frames
• 5 min exposures, 0.5° rotation, 90°frames
– Measure Resolution
• Determine how far spots diffracted (<2.0 Å)
– Measure Mosaicity
• <0.5°
2.0 Å1.41 Å
Page 17
Collecting Data on the Best Crystal
• Collect 400 to 500 frames
– Expose 20 minutes, 0.5° rotation
– Rotate 0.5°
– Repeat 400-500 times
Image Plate
Nitrogen Stream
Collimator Pinned Crystal
Goniometer
Page 18
Solving the Crystal Structure
• CrystalClear© Software for integration
• CCP4© for molecular replacement and refinement
• Coot© for manipulating to fit electron density
CrystalClear Software CCP4 Coot
Page 19
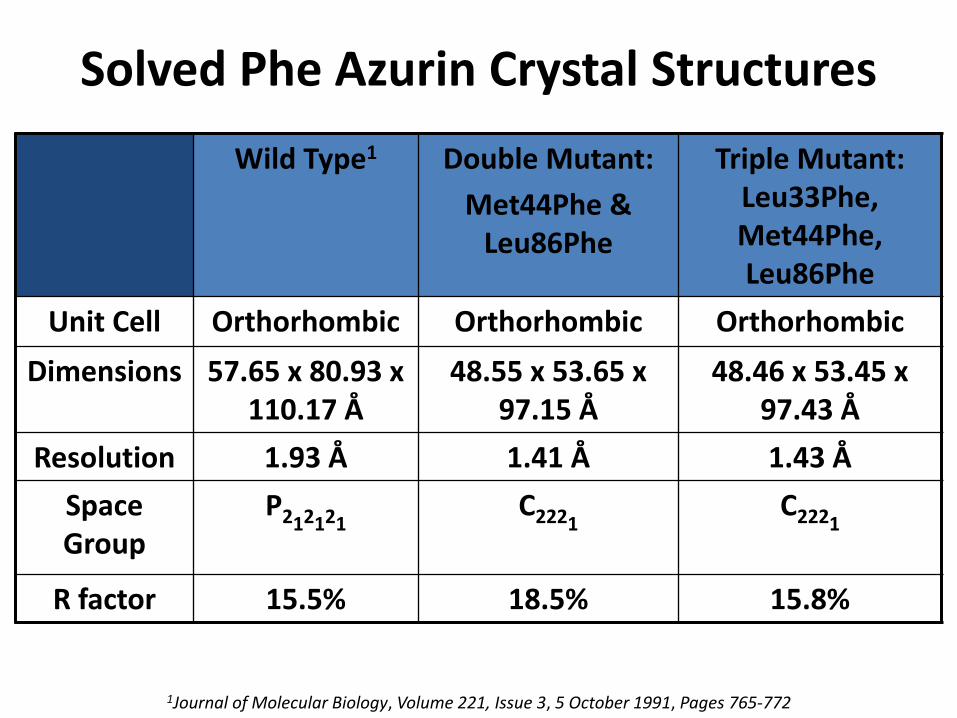
Solved Phe Azurin Crystal Structures
Wild Type1 Double Mutant:
Met44Phe & Leu86Phe
Triple Mutant: Leu33Phe, Met44Phe, Leu86Phe
Unit Cell Orthorhombic Orthorhombic Orthorhombic
Dimensions 57.65 x 80.93 x 110.17 Å
48.55 x 53.65 x 97.15 Å
48.46 x 53.45 x 97.43 Å
Resolution 1.93 Å 1.41 Å 1.43 Å
Space Group
P212121C2221
C2221
R factor 15.5% 18.5% 15.8%
1Journal of Molecular Biology, Volume 221, Issue 3, 5 October 1991, Pages 765-772
Page 20
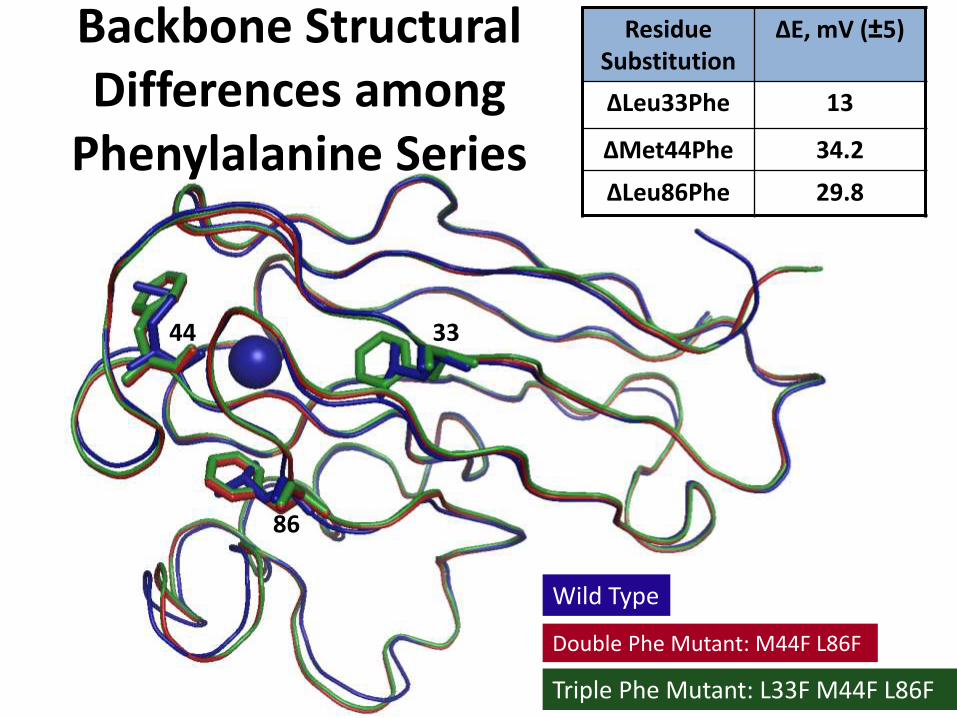
Backbone Structural Differences among
Phenylalanine Series
Wild Type
Double Phe Mutant: M44F L86F
Triple Phe Mutant: L33F M44F L86F
33
86
44
Residue Substitution
ΔE, mV (±5)
ΔLeu33Phe 13
ΔMet44Phe 34.2
ΔLeu86Phe 29.8
Page 21
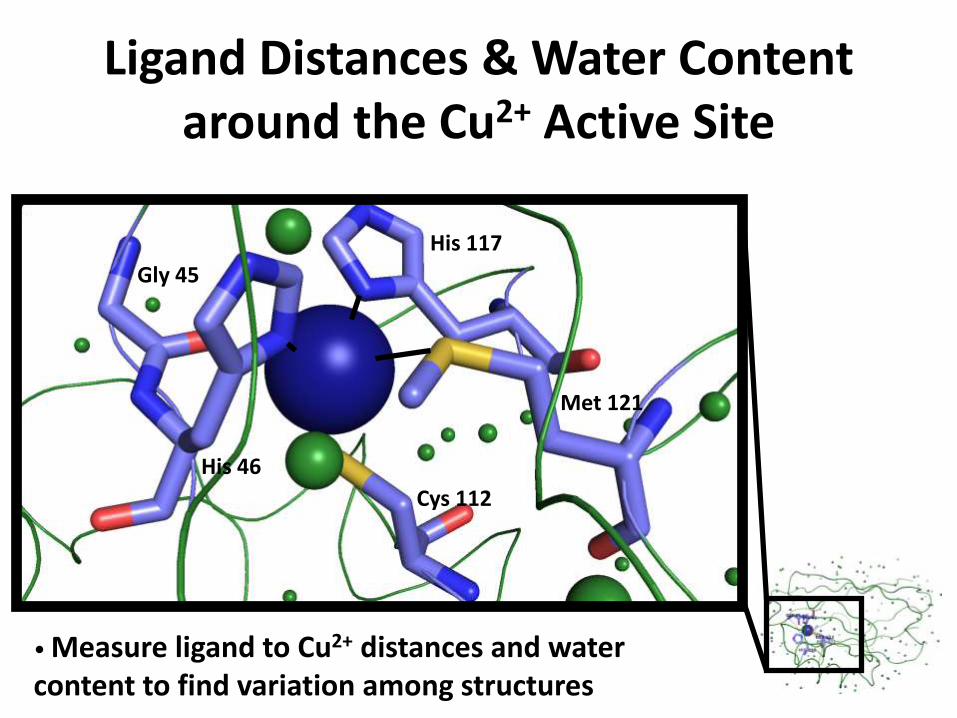
Ligand Distances & Water Content around the Cu2+ Active Site
Gly 45
His 117
His 46
Cys 112
Met 121
• Measure ligand to Cu2+ distances and water content to find variation among structures
Page 22
Structural differences between MutantsWild Type (PDB ID –
4AZU)
Double Mutant:
Met44Phe & Leu86Phe
Triple Mutant: Leu33Phe, Met44Phe, Leu86Phe
Water Content: within 8 Å
2 Molecules 4 Molecules 3 Molecules
His 46: nitrogen 2.0 Å 2.0 Å 2.0 Å
His 117: nitrogen 2.1 Å 2.0 Å 2.0 Å
Cys 112: sulfur 2.3 Å 2.2 Å 2.2 Å
Met 121: sulfur 3.2 Å 3.4 Å 3.3 Å
Gly 45: oxygen (carbonyl group)
2.8 Å 3.0 Å 3.1 Å
Gly 45: carbonyl group to Cu2+ angle
133.9° 129.9 ° 133.2 °
• Copper active sites are the same for the three proteins
Page 23
Conclusions• Purified and grew crystals of the double and
triple Phe azurin mutants
• Screened crystals using Rigaku Rapid II©
diffractometer
• Solved crystal structures of triple and double Phe mutant
• Compared and contrasted differences between wild-type Azurin and Phe mutants
– Backbone disruption of H-bonding network
– Water content near Cu2+
– Carbonyl ligand bonding angle
Page 24
Future Directions
• Crystallize and determine structure of WT
– Purify and crystallize WT azurin
• Determine structure of single Phe azurin mutants
– Currently screening crystals of Leu86Phe
– Purify and crystallize Met44Phe azurin
• Computational analysis of dipole distribution
Page 25
Acknowledgements• Dr. Berry
• Melanie Ladd
• Sarah Pedersen
• Dr. Nemykin
• Dr. Carter
• University of Minnesota Undergraduate Research Opportunities Program
• Department of Chemistry and Biochemistry
Page 27
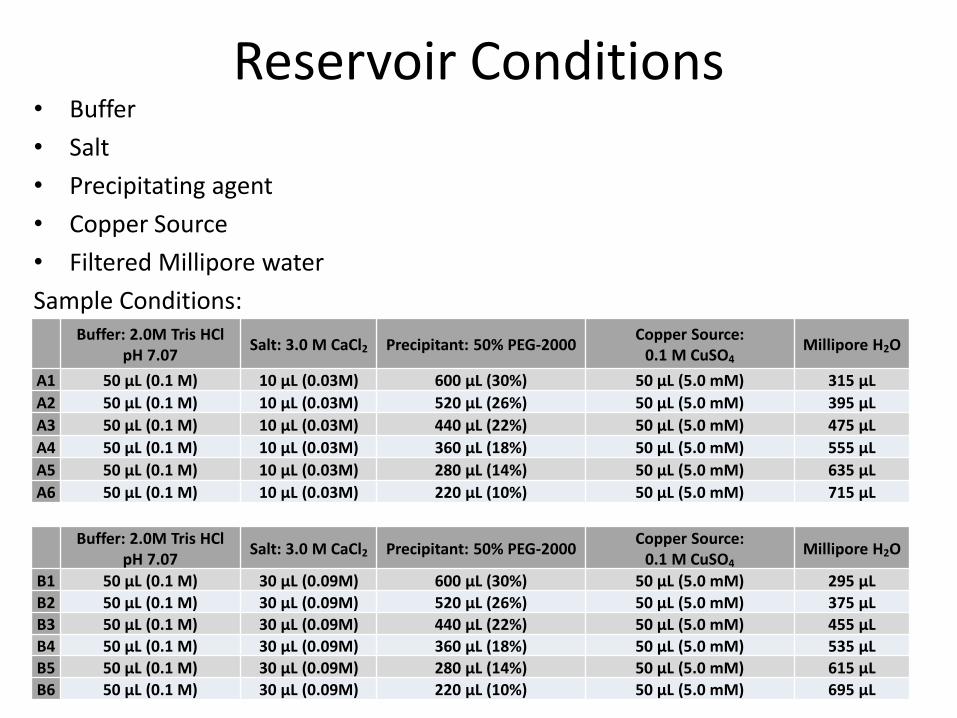
Reservoir Conditions• Buffer
• Salt
• Precipitating agent
• Copper Source
• Filtered Millipore water
Buffer: 2.0M Tris HClpH 7.07
Salt: 3.0 M CaCl2 Precipitant: 50% PEG-2000Copper Source:
0.1 M CuSO4Millipore H2O
A1 50 µL (0.1 M) 10 µL (0.03M) 600 µL (30%) 50 µL (5.0 mM) 315 µL
A2 50 µL (0.1 M) 10 µL (0.03M) 520 µL (26%) 50 µL (5.0 mM) 395 µL
A3 50 µL (0.1 M) 10 µL (0.03M) 440 µL (22%) 50 µL (5.0 mM) 475 µL
A4 50 µL (0.1 M) 10 µL (0.03M) 360 µL (18%) 50 µL (5.0 mM) 555 µL
A5 50 µL (0.1 M) 10 µL (0.03M) 280 µL (14%) 50 µL (5.0 mM) 635 µL
A6 50 µL (0.1 M) 10 µL (0.03M) 220 µL (10%) 50 µL (5.0 mM) 715 µL
Sample Conditions:
Buffer: 2.0M Tris HCl pH 7.07
Salt: 3.0 M CaCl2 Precipitant: 50% PEG-2000Copper Source:
0.1 M CuSO4Millipore H2O
B1 50 µL (0.1 M) 30 µL (0.09M) 600 µL (30%) 50 µL (5.0 mM) 295 µL
B2 50 µL (0.1 M) 30 µL (0.09M) 520 µL (26%) 50 µL (5.0 mM) 375 µL
B3 50 µL (0.1 M) 30 µL (0.09M) 440 µL (22%) 50 µL (5.0 mM) 455 µL
B4 50 µL (0.1 M) 30 µL (0.09M) 360 µL (18%) 50 µL (5.0 mM) 535 µL
B5 50 µL (0.1 M) 30 µL (0.09M) 280 µL (14%) 50 µL (5.0 mM) 615 µL
B6 50 µL (0.1 M) 30 µL (0.09M) 220 µL (10%) 50 µL (5.0 mM) 695 µL
Page 28
Protein Purification
• Grow E. coli containing a plasmid that codes for Phe mutants
• Use osmotic shock to separate cells from protein
• Purify the protein through SP-sepharous (cation-exchange column) and Q-column (anion-exchange column)
• Titrate with copper
• Purify through gel size exclusion column
Page 29
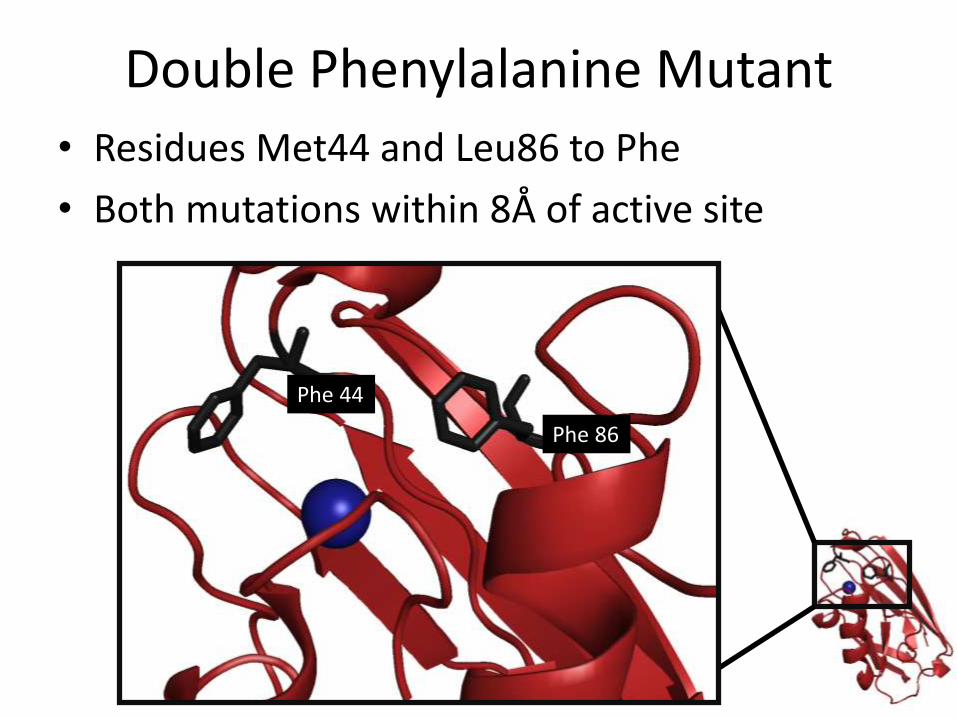
Double Phenylalanine Mutant
• Residues Met44 and Leu86 to Phe
• Both mutations within 8Å of active site
Phe 86
Phe 44





















![[My] Experiences building games in Visual Basic & Flash Focus on 'cannonball' Jeanine Meyer Math Senior Seminar.](https://static.documents.pub/doc/80x56/5697bfd21a28abf838cab7b9/my-experiences-building-games-in-visual-basic-flash-focus-on-cannonball.jpg)

![PECTIN METHYLESTERASE48 Is Involved in - Plant … METHYLESTERASE48 Is Involved in Arabidopsis Pollen Grain Germination1[OPEN] Christelle Leroux, Sophie Bouton, Marie-Christine Kiefer-Meyer,](https://static.documents.pub/doc/80x56/5b2f3aa57f8b9a94168c813d/pectin-methylesterase48-is-involved-in-plant-methylesterase48-is-involved-in-arabidopsis.jpg)











