Chromosomal directionality of DNA mismatch repair in Escherichia coli A. M. Mahedi Hasan and David R. F. Leach 1 Institute of Cell Biology, School of Biological Sciences, University of Edinburgh, Edinburgh EH9 3BF United Kingdom Edited by Peggy Hsieh, National Institute of Diabetes and Digestive and Kidney Diseases, National Institutes of Health, Bethesda, MD, and accepted by the Editorial Board June 16, 2015 (received for review March 17, 2015) Defects in DNA mismatch repair (MMR) result in elevated muta- genesis and in cancer predisposition. This disease burden arises because MMR is required to correct errors made in the copying of DNA. MMR is bidirectional at the level of DNA strand polarity as it operates equally well in the 5′ to 3′ and the 3′ to 5′ directions. However, the directionality of MMR with respect to the chromo- some, which comprises parental DNA strands of opposite polarity, has been unknown. Here, we show that MMR in Escherichia coli is unidirectional with respect to the chromosome. Our data demon- strate that, following the recognition of a 3-bp insertion-deletion loop mismatch, the MMR machinery searches for the first hemime- thylated GATC site located on its origin-distal side, toward the rep- lication fork, and that resection then proceeds back toward the mismatch and away from the replication fork. This study provides support for a tight coupling between MMR and DNA replication. mismatch repair | recombination | E. coli D NA can be mutated following damage caused by exposure to chemical or physical mutagens or by errors in DNA metabolism, during replication, recombination, or repair (1). Although replicative DNA polymerases have proofreading activities, they cannot avoid a low frequency of incorporation of noncomplementary deoxynucleoside triphosphates (dNTPs). It has been estimated that there are 10 −5 –10 −6 misincorporation events per replicated base pair (2). Uncorrected errors of this kind (known as mismatches) will be converted into mutations, which have the potential to perturb biological processes in the next round of DNA replication. The DNA mismatch repair (MMR) system is a key DNA guardian that ensures the removal of the misincorporated nucleotides and thereby maintains ge- nomic integrity. The MMR system has a defined substrate range, with different efficiencies of correction of single base mismatches and a 3–4 base size limit for the correction of insertion/deletion loops (IDLs) (3–6). This system is conserved in almost all or- ganisms, with the exception of most Actinobacteria and Molli- cutes, and parts of the archaea (7). When the replication machinery inserts a noncomplementary dNTP in the nascent strand of the E. coli chromosome, the MutS protein binds to the mismatch and recruits the MutL protein (8). These two proteins activate the endonuclease MutH, which nicks the unmethylated strand of a hemimethylated GATC site to ini- tiate removal of the nascent strand containing the mismatch (9, 10). Following the passage of the replisome, GATC motifs re- main transiently hemimethylated before methylation of the na- scent strand by the Dam methyltransferase enzyme and can therefore be used to distinguish between parental and nascent strands (11, 12). In vitro, MutH can distinguish between these strands by using hemimethylated GATC motifs located within a 2-kb distance of the mismatch, either on the 3′ or the 5′ side (13, 14). UvrD helicase uses the incision made by MutH at the hemimethylated GATC motif as an entry point to unwind the nascent strand. One or more of the four exonucleases (ExoI, ExoVII, RecJ, and ExoX), depending on the required polarity of degradation, resects the unwound nascent strand (15, 16). The single-stranded parental DNA is immediately bound by the single-strand DNA-binding protein (SSB) (17). The DNA poly- merase III holoenzyme then correctly resynthesizes the nascent strand and DNA ligase seals the remaining nick (16, 18). Most of the previous studies of MMR have been based on in vitro experiments using linear or closed-circular heteroduplex DNA substrates in defined systems (19–21). These in vitro studies have shown that an incision of the nascent strand can occur either on the 3′ or the 5′ side of a mismatch, depending on the position of the hemimethylated GATC motif recognized by MutH (22, 23). Using electron microscopy and end labeling, Grilley et al. have shown that the single-stranded DNA region created after an ex- cision reaction lies between the mismatch and the closest GATC motif (15). A hemimethylated GATC motif recognized on the 3′ side of the mismatch requires a 3′ to 5′ exonuclease (e.g., ExoI or ExoX), and ExoVII or RecJ cleaves the single-stranded DNA when a hemimethylated GATC motif is recognized on the 5′ side of the mismatch (5′ to 3′ cleavage).Therefore, in vitro, the cleav- age reaction of the MMR system is bidirectional (15, 16). How- ever, in vivo, an MMR system that is bidirectional with respect to DNA polarities could nevertheless be unidirectional with respect to the chromosome. In fact, a unique directionality of MMR with respect to DNA replication would explain the evolution of bidir- ectionality at the level of strand polarities as mismatches on both the leading and lagging strands need to be repaired. Fig. 1A dis- tinguishes directionality at the level of strand polarity from di- rectionality at the level of the chromosome and illustrates how directionality relative to the replication fork can lead to bidi- rectionality of resection polarities. Blackwood and collaborators found that MMR at the site of an unstable trinucleotide repeat (TNR) array caused a stimulation of recombination at a nearby 275-bp tandem repeat. This stimulation occurred only when the tandem repeat was placed on the origin-proximal side the TNR that had generated a high frequency substrate for MMR (24). This Significance DNA mismatch repair (MMR) is critical to avoid mutations that can lead to genetic disease, cancer, and death. The MMR sys- tem is evolutionarily conserved from bacteria to humans and is an example of a remarkable molecular machine. Here we show that this machine operates unidirectionally with respect to the chromosome of Escherichia coli. The most likely explanation for this directionality is that the MMR machinery is associated with the complex responsible for DNA replication. We suggest that this association facilitates the mechanism of strand discrimi- nation in MMR. Author contributions: A.M.M.H. and D.R.F.L. designed research; A.M.M.H. performed re- search; A.M.M.H. and D.R.F.L. analyzed data; and A.M.M.H. and D.R.F.L. wrote the paper. The authors declare no conflict of interest. This article is a PNAS Direct Submission. P.H. is a guest editor invited by the Editorial Board. Freely available online through the PNAS open access option. 1 To whom correspondence should be addressed. Email: [email protected]. This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10. 1073/pnas.1505370112/-/DCSupplemental. 9388–9393 | PNAS | July 28, 2015 | vol. 112 | no. 30 www.pnas.org/cgi/doi/10.1073/pnas.1505370112
Transcript
Chromosomal directionality of DNA mismatch repair inEscherichia coliA. M. Mahedi Hasan and David R. F. Leach1
Institute of Cell Biology, School of Biological Sciences, University of Edinburgh, Edinburgh EH9 3BF United Kingdom
Edited by Peggy Hsieh, National Institute of Diabetes and Digestive and Kidney Diseases, National Institutes of Health, Bethesda, MD, and accepted by theEditorial Board June 16, 2015 (received for review March 17, 2015)
Defects in DNA mismatch repair (MMR) result in elevated muta-genesis and in cancer predisposition. This disease burden arisesbecause MMR is required to correct errors made in the copying ofDNA. MMR is bidirectional at the level of DNA strand polarity as itoperates equally well in the 5′ to 3′ and the 3′ to 5′ directions.However, the directionality of MMR with respect to the chromo-some, which comprises parental DNA strands of opposite polarity,has been unknown. Here, we show that MMR in Escherichia coli isunidirectional with respect to the chromosome. Our data demon-strate that, following the recognition of a 3-bp insertion-deletionloop mismatch, the MMR machinery searches for the first hemime-thylated GATC site located on its origin-distal side, toward the rep-lication fork, and that resection then proceeds back toward themismatch and away from the replication fork. This study providessupport for a tight coupling between MMR and DNA replication.
mismatch repair | recombination | E. coli
DNA can be mutated following damage caused by exposureto chemical or physical mutagens or by errors in DNA
metabolism, during replication, recombination, or repair (1).Although replicative DNA polymerases have proofreadingactivities, they cannot avoid a low frequency of incorporation ofnoncomplementary deoxynucleoside triphosphates (dNTPs). Ithas been estimated that there are 10−5–10−6 misincorporationevents per replicated base pair (2). Uncorrected errors of thiskind (known as mismatches) will be converted into mutations,which have the potential to perturb biological processes in thenext round of DNA replication. The DNA mismatch repair(MMR) system is a key DNA guardian that ensures the removalof the misincorporated nucleotides and thereby maintains ge-nomic integrity. The MMR system has a defined substrate range,with different efficiencies of correction of single base mismatchesand a 3–4 base size limit for the correction of insertion/deletionloops (IDLs) (3–6). This system is conserved in almost all or-ganisms, with the exception of most Actinobacteria and Molli-cutes, and parts of the archaea (7).When the replication machinery inserts a noncomplementary
dNTP in the nascent strand of the E. coli chromosome, the MutSprotein binds to the mismatch and recruits the MutL protein (8).These two proteins activate the endonuclease MutH, which nicksthe unmethylated strand of a hemimethylated GATC site to ini-tiate removal of the nascent strand containing the mismatch (9,10). Following the passage of the replisome, GATC motifs re-main transiently hemimethylated before methylation of the na-scent strand by the Dam methyltransferase enzyme and cantherefore be used to distinguish between parental and nascentstrands (11, 12). In vitro, MutH can distinguish between thesestrands by using hemimethylated GATC motifs located within a2-kb distance of the mismatch, either on the 3′ or the 5′ side (13,14). UvrD helicase uses the incision made by MutH at thehemimethylated GATC motif as an entry point to unwind thenascent strand. One or more of the four exonucleases (ExoI,ExoVII, RecJ, and ExoX), depending on the required polarity ofdegradation, resects the unwound nascent strand (15, 16). Thesingle-stranded parental DNA is immediately bound by the
single-strand DNA-binding protein (SSB) (17). The DNA poly-merase III holoenzyme then correctly resynthesizes the nascentstrand and DNA ligase seals the remaining nick (16, 18).Most of the previous studies of MMR have been based on
in vitro experiments using linear or closed-circular heteroduplexDNA substrates in defined systems (19–21). These in vitro studieshave shown that an incision of the nascent strand can occur eitheron the 3′ or the 5′ side of a mismatch, depending on the positionof the hemimethylated GATC motif recognized by MutH (22, 23).Using electron microscopy and end labeling, Grilley et al. haveshown that the single-stranded DNA region created after an ex-cision reaction lies between the mismatch and the closest GATCmotif (15). A hemimethylated GATC motif recognized on the 3′side of the mismatch requires a 3′ to 5′ exonuclease (e.g., ExoI orExoX), and ExoVII or RecJ cleaves the single-stranded DNAwhen a hemimethylated GATC motif is recognized on the 5′ sideof the mismatch (5′ to 3′ cleavage).Therefore, in vitro, the cleav-age reaction of the MMR system is bidirectional (15, 16). How-ever, in vivo, an MMR system that is bidirectional with respect toDNA polarities could nevertheless be unidirectional with respectto the chromosome. In fact, a unique directionality of MMR withrespect to DNA replication would explain the evolution of bidir-ectionality at the level of strand polarities as mismatches on boththe leading and lagging strands need to be repaired. Fig. 1A dis-tinguishes directionality at the level of strand polarity from di-rectionality at the level of the chromosome and illustrates howdirectionality relative to the replication fork can lead to bidi-rectionality of resection polarities. Blackwood and collaboratorsfound that MMR at the site of an unstable trinucleotide repeat(TNR) array caused a stimulation of recombination at a nearby275-bp tandem repeat. This stimulation occurred only when thetandem repeat was placed on the origin-proximal side the TNRthat had generated a high frequency substrate for MMR (24). This
Significance
DNA mismatch repair (MMR) is critical to avoid mutations thatcan lead to genetic disease, cancer, and death. The MMR sys-tem is evolutionarily conserved from bacteria to humans and isan example of a remarkable molecular machine. Here we showthat this machine operates unidirectionally with respect to thechromosome of Escherichia coli. The most likely explanation forthis directionality is that the MMR machinery is associated withthe complex responsible for DNA replication. We suggest thatthis association facilitates the mechanism of strand discrimi-nation in MMR.
Author contributions: A.M.M.H. and D.R.F.L. designed research; A.M.M.H. performed re-search; A.M.M.H. and D.R.F.L. analyzed data; and A.M.M.H. and D.R.F.L. wrote the paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission. P.H. is a guest editor invited by the EditorialBoard.
Freely available online through the PNAS open access option.1To whom correspondence should be addressed. Email: [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1505370112/-/DCSupplemental.
result suggested that MMRmight be directional in a chromosomalcontext. We have now tested this hypothesis and shown that theMMR system of E. coli has a unique chromosomal directionality.
ResultsMMR Is Unidirectional in the E. Coli Chromosome. Length instabilityat the level of a single trinucleotide unit in a CTG·CAG repeatarray that is inserted in the chromosome of E. coli is elevated inMMR deficient cells (24). This elevation of mutation occurs to anequal extent in mutS, mutL, and mutH mutants indicating thatcorrection of the 3-base IDL, precursor to a single trinucleotiderepeat unit change, occurs by a normal MMR reaction involvingrecognition by MutS and strand cleavage by MutH (24). This is asexpected for a 3-base IDL, which has been shown to be correctedwith an efficiency close to that of a G·T mismatch (the best rec-ognized single base mismatch) (6) and contrasts with changes oftwo or more CTG·CAG trinucleotide units, which are not ele-vated in MMR deficient cells (24), as predicted by the inability ofthe MMR system to recognize an IDL of 5 bases (6). Accordingly,a CTG·CAG repeat array can be used as a source of frequent3-base IDL mismatches at a defined chromosomal locus for thestudy of MMR in vivo. In this work, we have used a repeated arrayof 98 copies of CTG·CAG (294 bp) where the CTG repeats arelocated on the leading-strand template in the lacZ gene of theE. coli chromosome (Fig. 1B). The variation of length of the repeatarray at the level of a single trinucleotide unit has been defined as“single-unit instability,” which is used as a quantitative phenotypereflecting the efficiency of the MMR system. We have focused onthe directionality of MMR as defined by the use of GATC sites onboth sides of the TNR. This was carried out by generating site-specific mutations that modified the GATC sites but retained theircoding sequences via synonymous substitutions.No significant change of single-unit instability was observed
following the modification of all of the GATC motifs within a2.3-kb region on the origin-proximal side of the TNR. In thissituation, the next available GATC site was located at a distanceof 2,339 bp from the TNR (P2339) and the frequency of single-unit instability remained equivalent to that observed for wildtype cells [D472(wt)] (Fig. 2). We conclude that GATC motifswithin the first 2.3 kb on the origin-proximal side of the TNR arenot required for the excision reaction during DNA mismatchrepair while origin-distal GATC motifs are present.On the other hand, the frequency of single-unit instability of
the TNR started to increase when GATC motifs beyond 1 kbwere modified on the origin-distal side of the TNR (D1356 on-ward in Fig. 2), while keeping the origin-proximal GATC motifsintact. The level of single-unit instability increased 2.9-fold overthat observed for [D472(wt)] when all of the available GATCmotifs within a 2-kb region on the original-distal side of the TNRwere modified (D2369). However, the level of single-unit in-stability observed for D2369 did not reach the level of that in amutS background. The difference of instability frequencies be-tween the mutS background and D2369 is statistically significantat a 95% confidence level (p value = 0.0317), indicating that anin vivo separation of 2 kb between the mismatch and the closestremaining GATC site did not totally abolish MMR. This con-trasts with the situation observed in vitro where a separation of2 kb abrogates MMR (15, 25, 26). Nevertheless, our data clearlyshow that the DNA mismatch repair system has a preference forrecognition of GATC sites on the origin-distal side of the TNR.Our next step was to extend the distance between the nearest
available GATC motif and a mismatch further beyond the 2 kbin vitro limit. As there were many closely spaced GATC motifsjust beyond the 2-kb boundary on the origin-distal side of theTNR in the E. coli genome, we extended the GATC-free regionby inserting a non E. coli DNA sequence in the strain D2369. A2.3-kb-long GATC motif-free DNA sequence was isolated fromthe fission yeast Schizosaccharomyces pombe and inserted at asite 200 bp from the TNR on the origin-distal side. In that strain[D4600(Sp)], the total length of GATC motif-free sequence onthe origin-distal side of the TNR reached 4.6 kb. The frequency
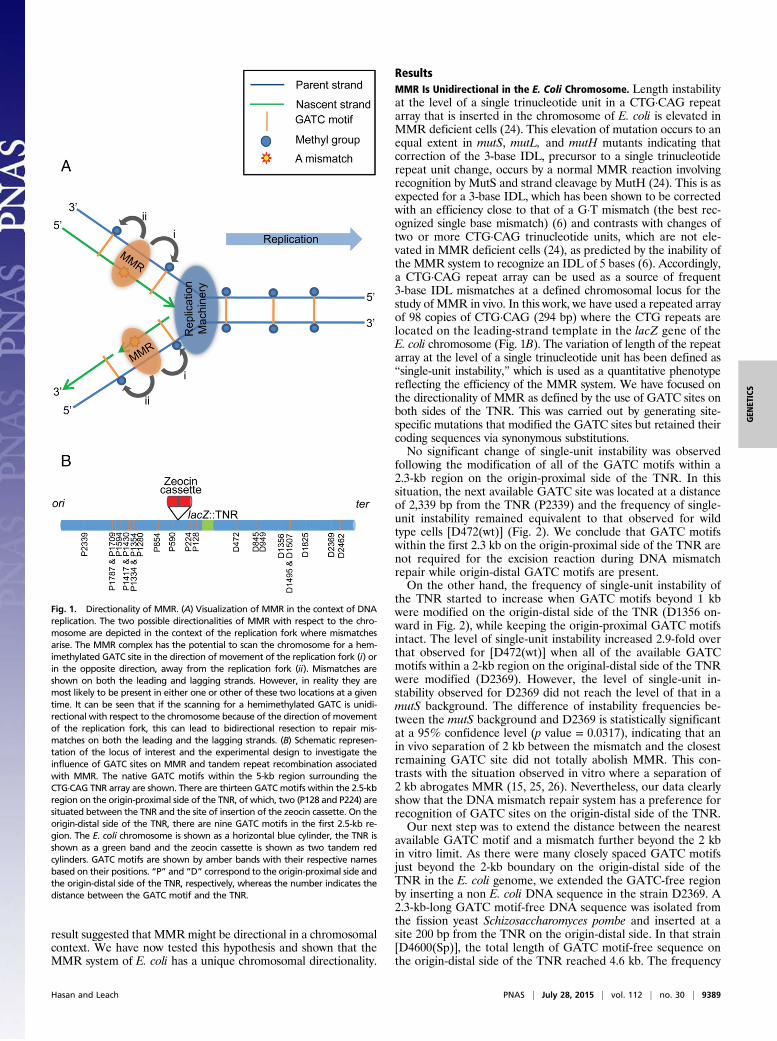
Fig. 1. Directionality of MMR. (A) Visualization of MMR in the context of DNAreplication. The two possible directionalities of MMR with respect to the chro-mosome are depicted in the context of the replication fork where mismatchesarise. The MMR complex has the potential to scan the chromosome for a hem-imethylated GATC site in the direction of movement of the replication fork (i) orin the opposite direction, away from the replication fork (ii). Mismatches areshown on both the leading and lagging strands. However, in reality they aremost likely to be present in either one or other of these two locations at a giventime. It can be seen that if the scanning for a hemimethylated GATC is unidi-rectional with respect to the chromosome because of the direction of movementof the replication fork, this can lead to bidirectional resection to repair mis-matches on both the leading and the lagging strands. (B) Schematic represen-tation of the locus of interest and the experimental design to investigate theinfluence of GATC sites on MMR and tandem repeat recombination associatedwith MMR. The native GATC motifs within the 5-kb region surrounding theCTG·CAG TNR array are shown. There are thirteen GATC motifs within the 2.5-kbregion on the origin-proximal side of the TNR, of which, two (P128 and P224) aresituated between the TNR and the site of insertion of the zeocin cassette. On theorigin-distal side of the TNR, there are nine GATC motifs in the first 2.5-kb re-gion. The E. coli chromosome is shown as a horizontal blue cylinder, the TNR isshown as a green band and the zeocin cassette is shown as two tandem redcylinders. GATC motifs are shown by amber bands with their respective namesbased on their positions. “P” and “D” correspond to the origin-proximal side andthe origin-distal side of the TNR, respectively, whereas the number indicates thedistance between the GATC motif and the TNR.
Hasan and Leach PNAS | July 28, 2015 | vol. 112 | no. 30 | 9389
GEN
ETICS
of single-unit instability in this strain rose close to that observedfor a DNAmismatch repair deficient background (mutS; p value =0.9451 at 95% confidence level), indicating little or no MMR(Fig. 2). This implies that the GATC sites on the origin proximalside of the TNR (which remained present in this situation) werenot used to direct repair.Our data have allowed us to calculate the relative efficiency of
MMR as a function of the distance between a mismatch and itsnearest GATC motif on the origin-distal side of the TNR (Fig.S1). The efficiency of the MMR system in a wild type cell hasbeen defined as 1 and that in a mutS mutant has been set to 0.Sequential modifications of GATC motifs on the origin-distalside result in the loss of efficiency of MMR as depicted in Fig.S1. Fitting an exponential trend line to the data reveals that theMMR system is 50% less efficient when the distance between theTNR and the next available GATC motif is 2.8 kb.To determine whether a single origin-distal GATC site located
close to the TNR was able to restore MMR, we inserted an ec-topic GATC site (via synonymous substitution) at 567 bp on theorigin-distal side of the TNR in the strain D2369 (D567-1G; Fig.S2). In this strain, the frequency of single-unit instability of theTNR returned to a similar level to that observed in wild type cells[D472(wt)]. This experiment suggested that a single origin-distalGATC site at 567 bp from the TNR was efficiently recognized. Toconfirm that this site was indeed recognized at close to 100%efficiency, we modified the same region (567 bp origin distal tothe TNR) to include three tandemly repeated GATC sites (D567-3G). Because that did not further increase the efficiency of MMRwe concluded that a single origin-distal GATC site at this distancefrom the TNR was already recognized with maximal efficiency.
The Efficiency of MMR Depends on the Length of the Excision Tract.To investigate whether the observed decrease in efficiency ofMMR with respect to the separation between the mismatch andthe closest origin-distal GATC site was determined by recognition
of the GATC site implicated in the reaction or the length of theexcision tract, we investigated a second reaction implicatingMMR. It has previously been shown that recombination at a splitzeocin resistance recombination reporter cassette (zeocin cassette)on the origin-proximal side of a TNR (Fig. S3) is stimulated byMMR (24). The rate of recombination was found to be a functionof the distance between the TNR and the zeocin cassette andcould be detected as far as 6.3 kb away on the origin-proximal sideof the TNR. Here, we have studied the recombination level at thezeocin cassette as a function of the location of the first availableGATC site to initiate the MMR reaction. In this experiment, weplaced the zeocin cassette at 500 bp on the origin-proximal side ofthe TNR, as indicated in Fig. 1B. In the absence of the TNR, therewas a basal rate of recombination [D472(wt) TNR–; Fig. 3]. Thislevel increased sevenfold in the presence of the TNR [D472(wt)TNR+]. This increase was dependent on MMR as only the basallevel of recombination was detected in a mutS mutant containingthe TNR (D472 mutS TNR+). This reaction has previously beenshown to be dependent on MutS, MutL, and MutH, with the ar-gument that (like MMR) it is dependent on an excision tractinitiated by cleavage of a GATC site by MutH endonuclease (24).In MMR proficient cells, mutation of both GATC motifs, P128and P224, located between the TNR and the zeocin cassette didnot affect the level of recombination (P590 TNR+). Therefore,this MMR excision reaction is not influenced by the presence ofthe GATCmotifs between the TNR and the zeocin cassette on theorigin-proximal side of the mismatch. A gradual decrease in therate of recombination at the zeocin cassette was observed whensequential modifications of GATC motifs extended beyond a 1-kbdistance on the origin-distal side of the TNR (D1356 TNR+ andbeyond). A 15% and a 19% decrease of the rate of recombinationwere observed, compared with the strain with intact GATC motifs[D472(wt) TNR+], in the strains with the first available GATCmotif at 1,356 bp (D1356 TNR+) and 1,825 bp (D1825 TNR+)away on the origin-distal side of the TNR. However, the rate ofrecombination did not decrease to the level observed in cells de-void of the TNR (TNR–) or mutated in the MMR system (D472mutS TNR+), even after modification of all GATC motifs within2 kb (D2369 TNR+) or 4.6 kb [D4600(Sp) TNR+] on the origin-distal side of the TNR (Fig. 3). Nevertheless, these experiments
Fig. 2. MMR as a function of GATC site availability. Frequencies of single-unitinstability of the TNR in strains with different distances to the nearest origin-proximal or origin-distal GATC sites. The distance to the first available GATC sitefrom the TNR is indicated on the x axis and the height of the correspondingcolumns shows the frequency of single-unit instability (y axis). Error bars rep-resent SEs of means. Strains are numbered according to the distance to the firstGATC motif present, with “P” and “D” representing the origin-proximal sideand the origin-distal side of the TNR respectively. The strain containing the wild-type GATC motif is named D472(wt) and D4600(Sp) represents the strain in-cluding a 2.3-kb GATC-free section of S. pombe DNA with a 4.6-kb GATC-freeregion on the origin-distal side of the TNR. The mutS strain refers to the strainD472 mutS TNR+ in the Table S1, which contains all of the native GATC sitesfrom the chromosome. All of the strains used in this assay contain the TNR.
Fig. 3. Tandem repeat recombination as a function of GATC site availabil-ity. Rates of recombination at a zeocin cassette in strains with differentdistances to the nearest origin-proximal or origin-distal GATC sites. The ratesof recombination at the zeocin cassette are shown along the y axis for strainswith different distances to the first available GATC site from the TNR (in-dicated along the x axis by the number following “P” for origin-proximalside and “D” for origin-distal side of the TNR). Error bars represent upperand lower limits of 95% confidence intervals.
9390 | www.pnas.org/cgi/doi/10.1073/pnas.1505370112 Hasan and Leach
revealed that, as for MMR, the rate of recombination was sensi-tive to the separation of the TNR and the zeocin cassette from thefirst available origin-distal GATC site.We have quantified the effect of the distance between the
mismatch and the first available GATC site on the origin-distalside of the TNR by plotting the relative recombination efficiencyas a function of this distance (see Supporting Information for thecalculation of relative recombination efficiencies). Here, we havedefined the strain with wild type GATC sites [D472(wt) TNR+] tohave a relative recombination efficiency of 1 and themutSmutant(D472mutS TNR+) to have a relative recombination efficiency of0. As can be seen in Fig. S4, the recombination efficiencies de-creased as the distance between the first available GATC site andthe TNR (or the zeocin cassette) increased. However, the slope ofthe exponential fitted curve is approximately half that for MMR(compare Figs. S4 and S1). The data predict that a 50% re-combination efficiency will be reached when 6 kb separates thefirst available GATC site and the TNR.The observations that the efficiencies of MMR and of zeocin
cassette recombination are affected by the separation between theTNR and first available origin-distal GATC site argue that origin-distal GATC sites are implicated in both reactions. However, thedifference in the efficiencies of the two reactions was surprisinggiven that both excision to mediate MMR and excision to stimu-late zeocin cassette recombination were expected to pass throughthe TNR and to enable repair. We considered that sensitivity tothe separation between the TNR (or zeocin cassette) and the firstavailable origin-distal GATC site might either reflect a diminishedprobability of recognizing a GATC site with distance from themismatch or a diminished probability of an excision tract coveringthe distance between the GATC site recognized and the TNR (orzeocin cassette). Given that there is no reason to expect two dif-ferent modes of recognition of the same origin-distal GATC site,we argue that two lengths of excision tracts must exist. We proposethat MMR is primarily mediated via shorter tracts that do notneed to extend far beyond the mismatch, whereas longer tracts areresponsible for zeocin cassette recombination at a greater distancebeyond the initiating mismatch. Because both reactions areexpected to involve excision tracts that remove the mismatch inthe TNR, the shorter tracts must be more frequent than the longertracts to explain the observation of two different efficiencies as afunction of separation of the mismatch and the first availableorigin-distal GATC site. The fact that two reactions, which areinitiated at the same GATC sites, are differentially sensitive to thedistance of the first origin-distal GATC site from the TNR pro-vides strong evidence that the lengths of the two different classesof excision tracts are the primary in vivo determinants of theability of a distant GATC to stimulate MMR or recombination.
RecQ Helicase Is Not Implicated in the Longer Excision Tracts. We havetested whether the RecQ helicase might be implicated in the longerexcision tracts, by extending excision tracts initiated by UvrD. Thiswas an attractive hypothesis as RecQ is a non-MMR helicase witha 3′ to 5′ directionality that can bind ssDNA—dsDNA junctionsand stimulate recombination. However, the rates of recombinationat the zeocin cassette were similar independently of the presence ofRecQ in strains where 4.6 kb of DNA separated the TNR and thefirst origin-distal GATC site [D4600(Sp) TNR+] (Fig. S5).
DiscussionMMR Is Unidirectional with Respect to the E. coli Chromosome. Wehave shown that to correct IDL mismatches the MMR system ofE. coli uses the GATC sequences present on the origin-distal sideof a TNR located in the chromosomal lacZ gene. Given that IDLsare created during DNA replication and that hemimethylatedGATC sites, which direct the strand specificity of MMR, aretransiently generated during DNA replication, we propose that itis the first GATC site between the mismatch and the replication
fork that is normally recognized during MMR. Unidirectionalityof the MMR system with respect to chromosomal DNA replica-tion explains bidirectionality at the level of the strand excision, asMMR is required to occur on both the leading and lagging strandof the replication fork (15).Our assay relies on the observation that the frequency of single-
unit TNR changes (+1 and −1 TNR) in a CTG·CAG repeat array isdependent on MMR catalyzed by MutS, MutL, and MutH (24).This is consistent with a conventional MMR reaction to correct a3-nucleotide IDL, generated in the TNR during DNA replicationand known to be a good substrate for MMR (6). For this reason, wehave made the simplifying assumption that the TNR is providing anelevated frequency of 3-nucleotide IDLs for conventional MMRdetected in our single-unit instability assay. However, we cannotformally exclude the possibility that some other features of the re-peat array are contributing to the frequency of single-unit changes.It is well known that trinucleotide repeats can form pseudohairpinsin vitro (27–32) that have been implicated in large-scale TNRchanges in many experimental systems including the E. coli chro-mosome (33, 34). We suspect that these large pseudohairpinstructures are not implicated in our single-unit TNR changes, but ifthey are, the repair reaction must involve the three components ofconventional MMR (MutS, MutL, and MutH) and must demon-strate the chromosomal directionality that we have detected.Further experiments are required to determine the molecular
basis of this directionality. However, codirectionality of hemi-methylated GATC recognition and DNA replication provides amechanism for origin-distal GATC motif scanning via a knowninteraction between the MutS protein and the β-clamp (35, 36).Following recognition of the mismatch by an ADP-bound formof MutS, this protein is converted into an ATP-bound slidingclamp (2). The interaction between MutS-ATP and the β-clampof the replisome would provide the possibility of a rapid anddirected scanning for a hemimethylated GATC site on the ori-gin-distal side of the mismatch, as shown in Fig. 4. Upon iden-tification of a hemimethylated GATC site, the incision of thenascent strand would be carried out by MutH in conjunction withthe matchmaker protein MutL.
The Maximal Distance, Measured Between a GATC Site and a Mismatch,for Productive MMR Is Longer in vivo than in vitro. Our data suggestthat the efficiency of MMR is reduced to 50% in the presence of2.8 kb of DNA between the mismatch and the GATC site used toinitiate resection. This distance is greater than the limit of 2 kbobserved in vitro (15, 25, 26). This difference could be explained ifscanning for a hemimethylated GATC site is unidirectional in vivobecause of the interaction between MutS and the β-clamp (asdescribed in Fig. 4). If so, scanning for a hemimethylated GATCsite might in fact not be the factor determining the maximal in vivodistance separating a mismatch and a GATC site for productiveMMR, while it might be critical in vitro. We have determined thatonly seven regions of the E. coli chromosome have GATC motifsseparated by distances greater than 2.8 kb. These regions are ei-ther derived from prophages or present in rhs loci, which have anunusual base composition and are subject to complex inheritancepatterns (37–40).
Different Lengths of MMR Excision Tracts Are Formed. MMR me-diates the repair of mismatches and stimulates recombination ata zeocin cassette located up to 6.3 kb away from a TNR (24).Both reactions involve the recognition of a GATC site on theorigin-distal side of the TNR. However, the zeocin cassette re-combination reaction implies the existence of longer resectiontracts extending beyond that observed in vitro (15). Here, weshow that MMR and zeocin cassette recombination are differ-entially affected by the distance between the first origin-distalGATC site and the TNR. This differential effect argues that twodifferent lengths of excision tracts are generated during MMR.
Hasan and Leach PNAS | July 28, 2015 | vol. 112 | no. 30 | 9391
The shorter tracts must be more frequent as they are primarilyresponsible for MMR and only need to reach the mismatch forrepair to occur. We have no evidence for where they terminate.However, they may indeed terminate within 100 nucleotidesbeyond the mismatch, as demonstrated in vitro (15). The longerexcision tracts that lead to zeocin cassette recombination must beless frequent as they do not determine the efficiency of MMRbut extend significantly beyond the mismatch. They are lesssensitive to the distance between the mismatch and the firstGATC site, suggesting that they are intrinsically longer (moreprocessive resection). We have eliminated the possibility that thelonger excision tracts are caused by the action of the RecQhelicase. Further work is required to determine the molecularbasis of the differences between these lengths of excision tracts.For example, it is known that several alternative nucleases cansubstitute for each other in excision tract processing (16, 41) andthis nuclease choice might determine excision tract length.
The Success of MMR as a Function of the Distance between theMismatch and the First Origin-Distal GATC Site Is Determined by theLength of the Excision Tract. We have considered two possibilitiesto explain the effect of the separation of the mismatch from thefirst origin-distal GATC site on the efficiency of MMR and ofzeocin cassette recombination. First, the recognition of a hemi-methylated GATC site is sensitive to the distance from the mis-match. Second, the processivity of resection determines the lengthof the excision tract. Given that there is no reason to propose twodifferent modes of GATC recognition for excision tracts of dif-ferent lengths, the fact that these events are differentially affectedby the distance between the first origin-distal GATC and the TNRargues that it is the length of the excision tract that determines thesuccess of MMR. This fits with the model presented in Fig. 4,where MutS is carried forward with the replisome facilitating theunidirectional scanning of DNA for a hemimethylated GATC site.We can estimate the length of the excision tracts that mediateMMR assuming that GATC sites are recognized equally well, arerecognized independently of their distance from the TNR in therange of distances that we have investigated and that resectionfrom the first origin-distal site (at 472 bp from the TNR) alwaysreaches the mismatch. Given these assumptions, the observationthat the efficiency of MMR is reduced to 50% with a separation of2.8 kb between the mismatch and the first available GATC siteargues that 50% of these excision tracts measure this distance. By asimilar argument, our data argue that 50% of the excision tractsthat mediate zeocin cassette recombination measure 6 kb plus thedistance required to uncover at least one copy of the repeatedsection of the zeocin cassette. Because the zeocin cassette is lo-cated 500 bp on the origin-proximal side of the TNR, the zeocincassette includes a repeated sequence of 275 bp and the TNRmeasures 294 bp a successful excision tract must reach ∼1.1 kbfarther than the distance between the recognized GATC site andthe TNR. The 50% length of these longer excision tracts is there-fore estimated to be ∼7 kb.
Implications for Eukaryotic MMR Systems. In eukaryotic cells, MMR isprimarily initiated by the MutSα (MSH2/MSH6) or MutSβ (MSH2/MSH3) complex, which is then followed by the MutLα (MLH1/PMS) protein. There is no separate MutH endonuclease homolog.However, MutLα has been shown to have endonuclease activity(42). Strand discrimination is carried out via the recognition of nicksin the nascent strand and it has recently been shown that these nickscan be generated via the removal of ribonucleotides from this DNAstrand (43, 44). The roles of helicases and exonucleases in eukary-otic MMR are only partially understood. No helicase has beenshown to be required for the resection reaction but this does notexclude the possible involvement of helicases (45). The exonucleaseactivity of Exo1 can provide 5′ to 3′ resection in the absence of ahelicase (46). Furthermore the endonuclease activity of MutLα cannick a strand 5′ of a mismatch that was originally nicked on the 3′side, allowing access to a 5′ to 3′ nuclease such as Exo1 (42, 47).However, the 3′ to 5′ exonuclease activities of polymerases δ ande have also been implicated in MMR (48). As in E. coli, theeukaryotic MutS complexes interact with the sliding clamp (49, 50).Therefore, whatever the transduction pathway to resection and thefinal polarity of that resection, we expect that MMR in eukaryoticcells will also show chromosomal directionality.
Materials and MethodsBacterial Strains. The construction and genotypes of the strains used areprovided in the Supporting Information.
MMR Analysis. Fragment length analysis of the TNR is described in theSupporting Information.
Recombination Analysis. Fluctuation analysis of recombination frequenciesat the zeocin cassette is described in the Supporting Information.
Fig. 4. Model for the chromosomal directionality of MMR. (A) Recognition ofa mismatch by MutS. MutS is closely associated to the replisome through itsinteraction with the β-clamp. Upon recognition of a mismatch the MutS dimeradopts its ATP-bound form and becomes a sliding clamp. (B) Recognition of anorigin-distal hemimethylated GATC site by the MutSLH complex. Throughcontinued association with the β-clamp, MutS slides with the replisome andbecomes associated with MutL and MutH. This triple complex recognizes thefirst hemimethylated GATC site encountered in the direction of replisomemovement. In these figures, mismatches are shown on both the leading andlagging strands. However, in reality replisome complexes are likely to encountera mismatch on either the leading or the lagging strand at a particular time.
9392 | www.pnas.org/cgi/doi/10.1073/pnas.1505370112 Hasan and Leach
3. Gradia S, Acharya S, Fishel R (2000) The role of mismatched nucleotides in activatingthe hMSH2-hMSH6 molecular switch. J Biol Chem 275(6):3922–3930.
4. Schofield MJ, et al. (2001) The Phe-X-Glu DNA binding motif of MutS. The role ofhydrogen bonding in mismatch recognition. J Biol Chem 276(49):45505–45508.
5. Jiricny J, Su SS, Wood SG, Modrich P (1988) Mismatch-containing oligonucleotideduplexes bound by the E. coli mutS-encoded protein. Nucleic Acids Res 16(16):7843–7853.
6. Parker BO, Marinus MG (1992) Repair of DNA heteroduplexes containing small het-erologous sequences in Escherichia coli. Proc Natl Acad Sci USA 89(5):1730–1734.
7. Sachadyn P (2010) Conservation and diversity of MutS proteins. Mutat Res 694(1-2):20–30.
8. Su SS, Modrich P (1986) Escherichia coli mutS-encoded protein binds to mismatchedDNA base pairs. Proc Natl Acad Sci USA 83(14):5057–5061.
9. Wang H, Hays JB (2003) Mismatch repair in human nuclear extracts: Effects of internalDNA-hairpin structures between mismatches and excision-initiation nicks on mis-match correction and mismatch-provoked excision. J Biol Chem 278(31):28686–28693.
10. Wang H, Hays JB (2004) Signaling from DNA mispairs to mismatch-repair excision sitesdespite intervening blockades. EMBO J 23(10):2126–2133.
11. Urig S, et al. (2002) The Escherichia coli dam DNA methyltransferase modifies DNA ina highly processive reaction. J Mol Biol 319(5):1085–1096.
12. Barras F, Marinus MG (1989) The great GATC: DNA methylation in E. coli. TrendsGenet 5(5):139–143.
13. Lahue RS, Su SS, Modrich P (1987) Requirement for d(GATC) sequences in Escherichiacoli mutHLS mismatch correction. Proc Natl Acad Sci USA 84(6):1482–1486.
14. Bruni R, Martin D, Jiricny J (1988) d(GATC) sequences influence Escherichia coli mis-match repair in a distance-dependent manner from positions both upstream anddownstream of the mismatch. Nucleic Acids Res 16(11):4875–4890.
15. Grilley M, Griffith J, Modrich P (1993) Bidirectional excision in methyl-directed mis-match repair. J Biol Chem 268(16):11830–11837.
16. Burdett V, Baitinger C, Viswanathan M, Lovett ST, Modrich P (2001) In vivo re-quirement for RecJ, ExoVII, ExoI, and ExoX in methyl-directed mismatch repair. ProcNatl Acad Sci USA 98(12):6765–6770.
17. Ramilo C, et al. (2002) Partial reconstitution of human DNA mismatch repair in vitro:Characterization of the role of human replication protein A. Mol Cell Biol 22(7):2037–2046.
18. Lahue RS, Au KG, Modrich P (1989) DNA mismatch correction in a defined system.Science 245(4914):160–164.
19. Thomas DC, Roberts JD, Kunkel TA (1991) Heteroduplex repair in extracts of humanHeLa cells. J Biol Chem 266(6):3744–3751.
20. Blackwell L J, Bjornson KP, Modrich P (1998) DNA-dependent activation of thehMutSalpha ATPase. J Biol Chem 273(48):32049–32054.
21. Dzantiev L, et al. (2004) A defined human system that supports bidirectional mis-match-provoked excision. Mol Cell 15(1):31–41.
22. Modrich P (1989) Methyl-directed DNA mismatch correction. J Biol Chem 264(12):6597–6600.
23. Au KG, Welsh K, Modrich P (1992) Initiation of methyl-directed mismatch repair. J BiolChem 267(17):12142–12148.
24. Blackwood JK, Okely EA, Zahra R, Eykelenboom JK, Leach DRF (2010) DNA tandemrepeat instability in the Escherichia coli chromosome is stimulated by mismatch repairat an adjacent CAG·CTG trinucleotide repeat. Proc Natl Acad Sci USA 107(52):22582–22586.
25. Pukkila PJ, Peterson J, Herman G, Modrich P, Meselson M (1983) Effects of high levelsof DNA adenine methylation on methyl-directed mismatch repair in Escherichia coli.Genetics 104(4):571–582.
26. Modrich P (1991) Mechanisms and biological effects of mismatch repair. Annu RevGenet 25:229–253.
27. Gacy AM, Goellner G, Jurani�c N, Macura S, McMurray CT (1995) Trinucleotide repeatsthat expand in human disease form hairpin structures in vitro. Cell 81(4):533–540.
28. Mitas M, Yu A, Dill J, Haworth IS (1995) The trinucleotide repeat sequence d(CGG)15forms a heat-stable hairpin containing Gsyn. Ganti base pairs. Biochemistry 34(39):12803–12811.
29. Smith GK, Jie J, Fox GE, Gao X (1995) DNA CTG triplet repeats involved in dynamicmutations of neurologically related gene sequences form stable duplexes. NucleicAcids Res 23(21):4303–4311.
30. Yu A, Dill J, Mitas M (1995) The purine-rich trinucleotide repeat sequences d(CAG)15and d(GAC)15 form hairpins. Nucleic Acids Res 23(20):4055–4057.
31. Yu A, et al. (1995) The trinucleotide repeat sequence d(GTC)15 adopts a hairpinconformation. Nucleic Acids Res 23(14):2706–2714.
32. Petruska J, Hartenstine MJ, Goodman MF (1998) Analysis of strand slippage in DNApolymerase expansions of CAG/CTG triplet repeats associated with neurodegenera-tive disease. J Biol Chem 273(9):5204–5210.
33. Zahra R, Blackwood JK, Sales J, Leach DRF (2007) Proofreading and secondary struc-ture processing determine the orientation dependence of CAG x CTG trinucleotiderepeat instability in Escherichia coli. Genetics 176(1):27–41.
34. Andreoni F, Darmon E, Poon WCK, Leach DRF (2010) Overexpression of the single-stranded DNA-binding protein (SSB) stabilises CAG*CTG triplet repeats in an orien-tation dependent manner. FEBS Lett 584(1):153–158.
35. López de Saro FJ, Marinus MG, Modrich P, O’Donnell M (2006) The beta sliding clampbinds to multiple sites within MutL and MutS. J Biol Chem 281(20):14340–14349.
37. Feulner G, et al. (1990) Structure of the rhsA locus from Escherichia coli K-12 andcomparison of rhsA with other members of the rhs multigene family. J Bacteriol172(1):446–456.
39. Minet AD, Rubin BP, Tucker RP, Baumgartner S, Chiquet-Ehrismann R (1999) Ten-eurin-1, a vertebrate homologue of the Drosophila pair-rule gene ten-m, is a neu-ronal protein with a novel type of heparin-binding domain. J Cell Sci 112 (Pt 1):2019–2032.
40. Aggarwal K, Lee KH (2011) Overexpression of cloned RhsA sequences perturbs thecellular translational machinery in Escherichia coli. J Bacteriol 193(18):4869–4880.
41. Kunkel TA, Erie DA (2005) DNA mismatch repair. Annu Rev Biochem 74:681–710.42. Kadyrov FA, Dzantiev L, Constantin N, Modrich P (2006) Endonucleolytic function of
MutLalpha in human mismatch repair. Cell 126(2):297–308.43. Ghodgaonkar MM, et al. (2013) Ribonucleotides misincorporated into DNA act as
strand-discrimination signals in eukaryotic mismatch repair. Mol Cell 50(3):323–332.44. Lujan SA, Williams JS, Clausen AR, Clark AB, Kunkel TA (2013) Ribonucleotides are
signals for mismatch repair of leading-strand replication errors. Mol Cell 50(3):437–443.
45. Song L, Yuan F, Zhang Y (2010) Does a helicase activity help mismatch repair in eu-karyotes? IUBMB Life 62(7):548–553.
46. Genschel J, Bazemore LR, Modrich P (2002) Human exonuclease I is required for 5′ and3′ mismatch repair. J Biol Chem 277(15):13302–13311.
47. Kadyrov FA, et al. (2007) Saccharomyces cerevisiae MutLalpha is a mismatch repairendonuclease. J Biol Chem 282(51):37181–37190.
48. Tran HT, Gordenin DA, Resnick MA (1999) The 3′–>5′ exonucleases of DNA poly-merases delta and epsilon and the 5′–>3′ exonuclease Exo1 have major roles inpostreplication mutation avoidance in Saccharomyces cerevisiae. Mol Cell Biol 19(3):2000–2007.
49. Flores-Rozas H, Clark D, Kolodner RD (2000) Proliferating cell nuclear antigen andMsh2p-Msh6p interact to form an active mispair recognition complex. Nat Genet26(3):375–378.
50. Kleczkowska HE, Marra G, Lettieri T, Jiricny J (2001) hMSH3 and hMSH6 interact withPCNA and colocalize with it to replication foci. Genes Dev 15(6):724–736.
51. Merlin C, McAteer S, Masters M (2002) Tools for characterization of Escherichia coligenes of unknown function. J Bacteriol 184(16):4573–4581.
52. Spell RM, Jinks-Robertson S (2004) Determination of mitotic recombination rates byfluctuation analysis in Saccharomyces cerevisiae. Methods Mol Biol 262:3–12.
Hasan and Leach PNAS | July 28, 2015 | vol. 112 | no. 30 | 9393