35
Chromosome Organization and Molecular Structure
| Date post: | 26-Dec-2015 |
| Category: |
Documents |
| Upload: | daniel-harrington |
| View: | 223 times |
| Download: | 0 times |

Chromosome Organization and Molecular Structure

Chromosomes & Genomes
Chromosomes complexes of DNA and proteins – chromatin Viral – linear, circular; DNA or RNA Bacteria – single, circular Eukaryotes – multiple, linear
Genome The genetic material that an organism possesses Nuclear genome Mitochondrial & chloroplasts genome

Genome is Infectious particles containing nucleic acid surrounded by a protein capsid
Rely on host cell for replication, transcription, translation
Exhibit a limited host range Genomes vary from a few thousand to a
hundred thousand nucleotides
Viruses

Some Virus StructuresPhage
Capsid protein
TMV


In a region called the nucleoid
DNA in direct contact with cytoplasm
Bacterial Chromosomes

Bacterial Chromosomes
Chromosomal DNA is compacted ~ 1000 fold to fit within cell

Size Escherichia coli
~ 4.6 million bp Haemophilus influenzae
~ 1.8 million bp Composition
E coli ~6000 genes Genes encoding proteins
for related functions arranged in operons
Intergenic regions nontranscribed DNA
Single origin of replication (Ori)
Bacterial Chromosomes

Prokaryotic Gene (Operon) Structure Stop Codon
TAA, TAG, TGARegulatoryElements
Cistron 1
Coding Sequence= ORF
+1 ATGStop Codon
TAA, TAG, TGA ATG
Coding Sequence= ORF
Cistron 2
Promoter& Operator
DNA
Terminatorsequence
Regulatory and Coding Sequence Unit = Operon
Protein A Protein B
Structural or Coding SequencesRegulatory Sequences

Eukaryotic species contain one or more sets of chromosomes (ploidy level)
Each set is composed of several linear chromosomes
DNA amount in eukaryotic species is greater than that in bacteria
Chromosomes in eukaryotes are located in the nucleus To fit in there, they must be highly compacted
This is accomplished by the binding of many proteins The DNA-protein complex is termed chromatin
Copyright ©The McGraw-Hill Companies, Inc. Permission required for reproduction or display
Eukaryotic Chromosomes
10-21

vary substantially in size variation not related to complexity of the
species i.e - a two fold difference in genome size
between two salamander species Size difference due to accumulation of
repetitive DNA sequences
Eukaryotic genomes

Figure 10.10
Variations in DNA Content

Eukaryotic chromosomes are long, linear DNA molecule
Three types of DNA sequences are required for chromosome replication and segregation Origins of replication (multiple) Centromeres (1) Telomeres (2)
Eukaryotic Chromosome Organization

Centromere
Kinetochore proteins
Origin of replication
Origin of replication
Origin of replication
Origin of replication
Telomere
Telomere
GenesRepetitive sequences

Chromosome Organization Genes located between centromere & telomeres
hundreds to thousands of genes lower eukaryotes (i.e. yeast)
Genes are relatively small Very few introns
higher eukaryotes (i.e. mammals) Genes are long Have many introns
Non-gene sequences Repetitive DNA
Telomere Centromere Satellite

Eukaryotic Gene Structure
Promoter/Enhancer
Cis-Regulatory Elements
Start Codon ATG
Exon1 Exon2 Exon3
Stop CodonTAA, TAG, TGA

Sequence complexity refers to the number of times a particular base sequence appears in the genome
2 main types of sequences Moderately repetitive Highly repetitive (low complexity)
Repetitive Sequences

Unique or non-repetitive sequences Found once or a few times in the genome Includes structural genes as well as intergenic areas
Moderately repetitive Found a few hundred to a few thousand times Includes
Genes for rRNA and histones Origins of replication Transposable elements
Repetitive Sequences
10-28

Highly repetitive Found tens of thousands to millions of times Each copy is relatively short (a few nucleotides to several
hundred in length)
Some sequences are interspersed throughout the genome Example: Alu family in humans
Other sequences are clustered together in tandem arrays Example: centromeric satellite & telomeric regions
Repetitive Sequences

Stretched end to end, a single set of human chromosomes will be over 1 meter long nucleus is only 2 to 4 m in diameter
The compaction of linear DNA in eukaryotic chromosomes involves interactions between DNA and various proteins Proteins bound to DNA are subject to change during the
life of the cell These changes affect the degree of chromatin compaction
Eukaryotic Chromatin Compaction

Histone proteins basic (+ charged lysine & arginine)amino acids that bind DNA backbone
Four core histones in nucleosome Two of each of H2A, H2B, H3 & H4
Fifth histone, H1 is the linker histone
Figure 10.14
Nucleosomes

Figure 10.14
Nucleosomes

Beads on a String
Overall structure of connected nucleosomes resembles “beads on a string” Shortens DNA length ~ seven-fold

Nucleosomes associate to form more compact structure - the 30 nm fiber
Histone H1 plays a role in this compaction
Nucleosomes Join to Form 30 nm Fiber
10-54

Copyright ©The McGraw-Hill Companies, Inc. Permission required for reproduction or display
The two events we have discussed so far have shortened the DNA about 50-fold
A third level of compaction involves interaction between the 30 nm fiber and the nuclear matrix
Further Compaction of the Chromosome
10-57
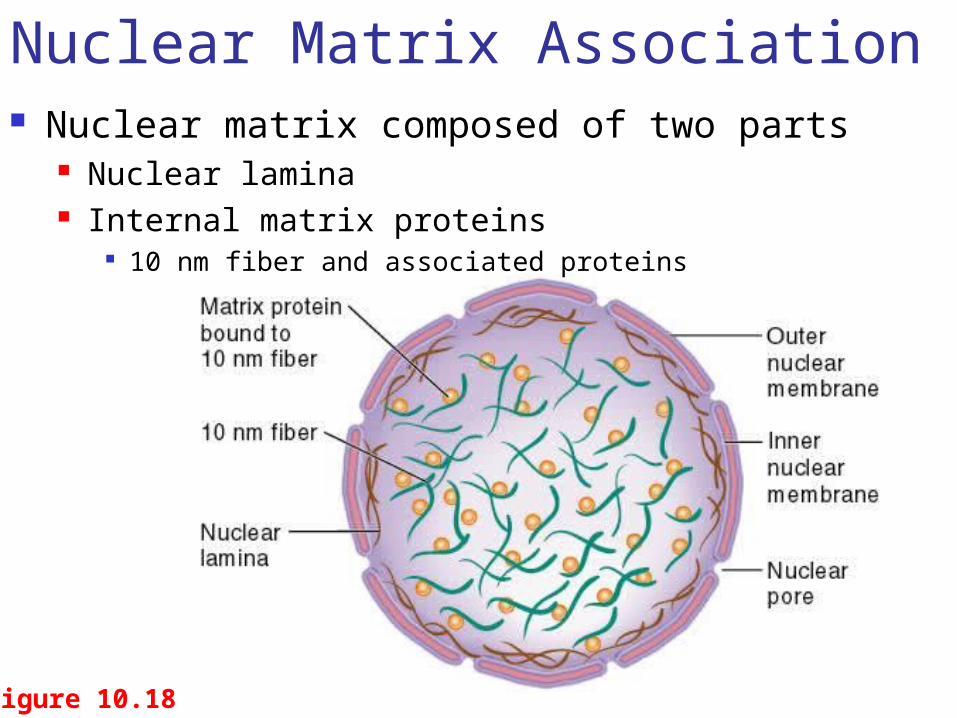
Figure 10.18
Nuclear Matrix Association Nuclear matrix composed of two parts
Nuclear lamina Internal matrix proteins
10 nm fiber and associated proteins
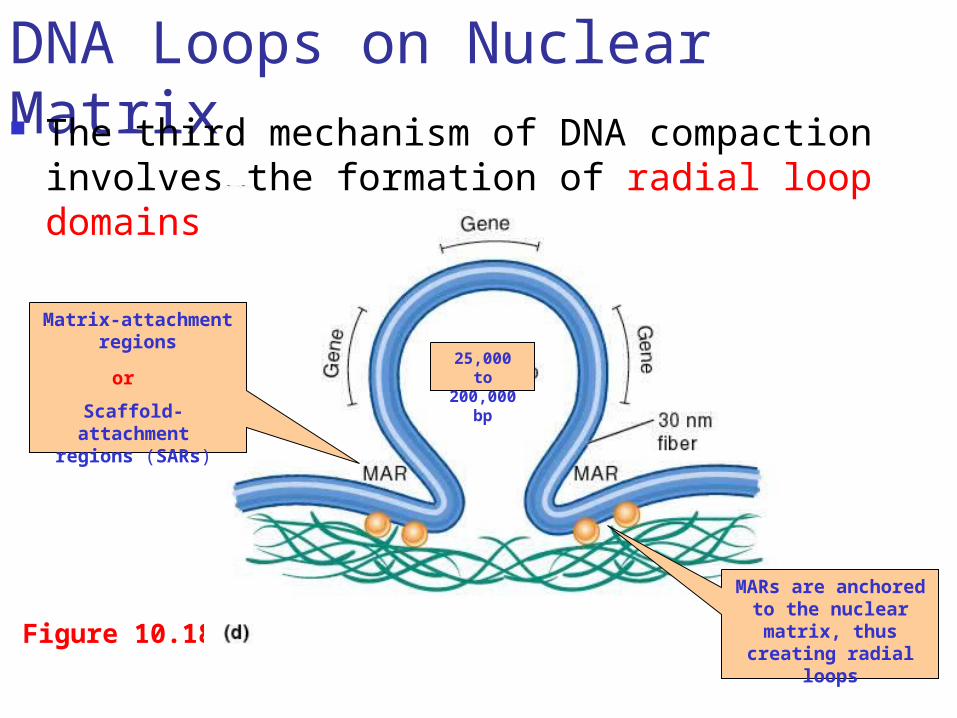
Figure 10.18
Matrix-attachment regions
Scaffold-attachment regions (SARs)
or
MARs are anchored to the nuclear matrix, thus creating radial
loops
25,000 to 200,000 bp
DNA Loops on Nuclear Matrix The third mechanism of DNA compaction involves the
formation of radial loop domains

Copyright ©The McGraw-Hill Companies, Inc. Permission required for reproduction or display
The attachment of radial loops to the nuclear matrix is important in two ways 1. It plays a role in gene regulation
Discussed in Chapter 15
2. It serves to organize the chromosomes within the nucleus
Each chromosome in the nucleus is located in a discrete and nonoverlapping chromosome territory
Refer to Figure 10.19
Further Compaction of the Chromosome
10-60

Compaction level of interphase chromosomes is not uniform
Euchromatin Less condensed regions of chromosomes Transcriptionally active Regions where 30 nm fiber forms radial loop domains
Heterochromatin Tightly compacted regions of chromosomes Transcriptionally inactive (in general) Radial loop domains compacted even further
Heterochromatin vs Euchromatin

Types of Heterochromatin
Figure 10.20
Constitutive heterochromatin Always heterochromatic Permanently inactive with regard to transcription
Facultative heterochromatin Regions that can interconvert between euchromatin
and heterochromatin Example: Barr body

Figure 10.21

10-64Figure 10.21
Compaction level in euchromatin
Compaction level in heterochromatin
During interphase most chromosomal
regions are euchromatic

Condensed chromosomes are referred to as metaphase chromosomes
During prophase, the compaction level increases
By the end of prophase, sister chromatids are entirely heterochromatic
These highly condensed metaphase chromosomes undergo little gene transcription
In metaphase chromosomes, the radial loops are compacted and anchored to the nuclear matrix scaffold
Metaphase Chromosomes

The condensation of a metaphase chromosome by condensinFigure 10.23
The number of loops has not changedHowever, the diameter of each loop is smaller
Condensin travels into the nucleus
Condensin binds to chromosomes and compacts the
radial loops
During interphase, condensin is in the cytoplasm
Chromosome Condensation

The alignment of sister chromatids via cohesinFigure 10.24
Cohesins along chromosome arms are
released
Cohesin remains at centromere
Cohesin at centromer is degraded
Chromosomes During Mitosis

















