Chronic Treatment of Aged Mice With L-Deprenyl Produces Marked Striatal MAO-B
Inhibition But No Beneficial Effects on Survival, Motor Performance, or Nigral
Lipofuscin Accumulation
D O N A L D K. I N G R A M , *l H A R V E Y L. W l E N E R , t M A R K E. C H A C H I C H , * J E F F R E Y M. L O N G , * J O H N H E N G E M I H L E * A N D M A D I GUPTA~:
*Molecular Physiology and Genetics Section, Nathan W. Shock Laboratories, Gerontology Research Center, National Institute on Aging, National Institutes of Health, Bayview Research Campus,
4940 Eastern Avenue, Baltimore, MD 21224 :'College of Pharmacy and Allied Health Professions, Department of Pharmaceutical Sciences,
St. John's University, Jamaica, NY 11439 ~:Department of Anatomical Sciences and Neurobiology, Health Sciences Center,
University of Louisville, Louisville, KY 40292
Rece ived 11 D e c e m b e r 1992; Accep ted 26 Apri l 1993
INGRAM, D. K., H. L. WIENER, M. E. CHACHICH, J. M. LONG, J. HENGEMIHLE AND M. GUPTA. Chronic treatment of aged mice with L-deprenyl produces marked striatal MAO-B inhibition but no beneficial effects on survival, motor performance, or nigral lipofuscin accumulation. NEUROBIOL AGING 14(5) 431-440, 1993.--Male C57BL/6J mice were provided l-deprenyl (at 0, 0.5 mg/kg or 1.0 mg/kg per day) in their drinking water beginning at 18 months of age. A battery of motor tests, including open-field, tightrope, rotorod, inclined screen, runwheel, and rotodrum tests, was administered before treatment and then 6 months later at 24 months of age. A subsample of mice was retested again at 27 months of age. An untreated group of 9-month-old mice served as young controls. Deprenyl treatment reduced striatal MAO-B activity by up to 60% after 6 months on treatment but had no significant effects on striatal catecholamine levels. No significant effects of deprenyl treatment were observed on body weight, fluid intake, or survival of the mice. Chronic deprenyl treatment also did not affect motor performance in any test, except rotodrum performance at 27 months of age, which was significantly better in the 1.0 mg/kg group treated group compared to controls. No age or deprenyl effects were observed with respect to cell counts in the substantia nigra. However, nigral cells containing lipofuscin increased with age, but this neurohistochemical parameter was also unaffected by deprenyl treatment.
Aging Lifespan Catecholamines Dopamine Behavior Neuroanatomy Substantia nigra Parkinson's disease
L-DEPRENYL (phenylisopropyl-N-methylpropynylamine, also known as selegiline) was introduced by Knoll et al. (36) as a potent, irreversible blocker of monoamine oxidase Type B (MAO- B) activity used primarily as an antidepressant. Subsequent studies have indicated its potential as a safe therapeutic agent for a variety of neurodegenerative diseases and possibly for aging. According to numerous studies summarized by Knoll (31-35), the compound has several marked effects that facilitate the activity of dopami- nergic neurons in the nigrostriatal system selectively, including increasing the firing rate of dopaminergic neurons as well as in-
1 To whom requests for reprints should be addressed.
creasing the release and turnover of dopamine while inhibiting its reuptake.
Interest in the potential use of deprenyl to protect against neu- rodegenerative disorders was stimulated by reports that the com- pound could block nigrostriatal toxicity induced by 6-hydroxy- dopamine in rats (29) and by MPTP (1-methyl-4-phenyl-l,2,5,6- tetrahydropyridine) in mice (20) and in monkeys (11). Although this action likely pertained primarily to MAO inhibition, these findings generated another hypothesis that age-related decline in nigrostriatal function through cell loss, receptor loss, membrane
431
432 INGRAM ET AL.
alterations, and lipofuscin accumulation might result from the gen- eration of oxy-radicals through the normal action of MAO-B me- tabolism of dopamine ( 10,30,31). Specifically, MAO degradation of dopamine produces hydrogen peroxide that can induce lipid peroxidation or oxidization of sulfhydryl groups of proteins or generate superoxide or hydroxyl radicals (10), all of which can impair function through a variety of mechanisms that can lead to neuronal death. It is also generally agreed that brain MAO activity increases with age (46,47,53); therefore, these detrimental activ- ities could accelerate with advancing age.
Several studies have indicated the possible involvement of ox- ygen radicals in the pathogenesis of Parkinson's disease related to evidence of increased lipid peroxidation in substantia nigra and reduced levels of catalase and glutathione peroxidase to detoxify hydrogen peroxide and oxy-radicals, respectively (see ref. 1 for review). Multi-site studies treating Parkinson's patients with de- prenyl in conjunction with standard pharmacological treatments (L-dopa) or antioxidants (vitamin E) indicated the therapeutic ben- efits of deprenyl for retarding the progression of the disease (5,6, 17,44,51) and for possibly increasing survival of treated patients (7). However, these reports have not always been positive with respect to long-term therapeutic benefits of deprenyl treatment or to the avoidance of side effects at higher doses (14,15,16).
Based on the observations of increased brain MAO activity among Alzheimer's patients (2,42) as well as disturbances to monoamine neurotransmission (37), deprenyl has also been used in the treatment of memory impairment observed in this disease. Modest but statistically significant improvement in performance on cognitive tests has been observed following deprenyl treatment; however, the effects primarily involved performance dependent on attention and sustained effort (43,50).
Perhaps the most dramatic claims for the therapeutic benefits of long-term deprenyl treatment have emerged from Knoll's reports (32,33) of extended lifespan and restored sexual function among aged male rats. Recent confirmation of the positive effect of de- prenyl on lifespan in aged male F-344 rats has been reported (28,39); however, the effect was far less dramatic than reported by Knoll (32,33). The observed effects on sexual function could be linked logically to effects that deprenyl might have on dopaminer- gic function in relation to "drive-motivated behavior" (33). How deprenyl might enhance survival is less clear. Knoll (34) has hy- pothesized that the compound might reduce neuronal death in- duced by specific endogenous neurotoxins generated by the MAO catalysis of dopamine. Moreover, he has reported that treatment of young rats with deprenyl increases the striatal activity of super- oxide dismutase (SOD), which is an endogenous scavenger of the superoxide radical (33). These results have been confirmed in recent reports indicating that deprenyl treatment increased the ac- tivity of SOD and catalase but not glutathione peroxidase in the rat striatum (8,9). Cohen and Spina (12) have indicated that deprenyl treatment can reduce the level of glutathione disulfide induced by haloperidol treatment in mice as an index against oxidative stress. These findings support the view of a generalized effect of deprenyl on reducing impairment due to oxy-radicals in support of the free radical theory of aging (19).
The objective of the present study was to extend the findings related to the effects of deprenyl on aging in the nigrostriatal dopaminergic system in another species. We selected the C57BL/ 6J (B6) mouse to provide an actuarial, behavioral, neurochemical, and morphological analysis of chronic deprenyl treatment. This inbred mouse strain is known to exhibit age-related declines in motor performance (23,25), striatal dopamine receptor concentra- tion (21), selected markers of dopamine metabolism in the nigro- striatal system (13) as well as nigral accumulation of lipofuscin (40). Compared to other strains, B6 mice have relatively high
levels of striatal MAO-B activity (55), which makes this strain particularly sensitive to MPTP toxicity (48). Wiener et al. (56) demonstrated in young BALB/cBy mice that 4-week treatment with deprenyl (10 mg/kg twice weekly) could reduce MAO-B activity by over 85% and increase binding of [3H] mazindol as a measure of the dopamine uptake carrier. In the present study, we began treatment at 18 months of age and carried the mice through to either 24 or 27 months of age. Because the mean lifespan of B6 mice in our laboratory has been estimated to be about 26-27 mo (18), this intervention was initiated much earlier than reported in previous rat studies of chronic deprenyl treatment on parameters of aging (32,33,39). If deprenyl retarded brain aging generally, we expected to observe a significant increase in survival, improved motor performance as assessed in a battery of age-sensitive tests, and reduced accumulation of nigral cells containing lipofuscin pigment.
METHOD
Subjects
Male C57BL/6J were obtained around 3 (n = 30), 17 (n = 90), or 24 months of age (n = 24) from the colony maintained at the Gerontology Research Center (GRC: Baltimore, MD). The mice were housed in groups of 5 in plastic cages with wood shav- ings for bedding. Fresh cages and bedding were provided weekly. The cages were located in a vivarium used exclusively for the study and maintained automatically at 28°C with a relative humid- ity of 70% and a photocycle set at 12L: 12D cycle with lights on at 06:00. The mice were fed a NIH-07 diet ad lib from hoppers located in their cages. Distilled water was provided ad lib to each cage from a glass bottle with a stainless steel sipper tube equipped with ball bearings to reduce spillage. A sentinel animal system was used to monitor pathogen conditions in the vivarium. Periodic sacrifices of the sentinel animals during the study indicated that the vivarium was free of common infectious pathogens as described previously (49).
Motor Battery: Test 1
Beginning at 3 or 18 months of age, the mice were tested in a battery of motor tests according to the procedures and schedule described below and summarized in Table 1. All testing occurred during the light portion of the photocycle. During all tests the mice were first brought in their home cage to the test room and allowed to acclimate for at least 30 min.
Open-Field (Day I). This test assesses the amount of locomo- tor activity during brief exposure to a novel environment. Previous studies using other apparatus have demonstrated an age-related decline in this behavioral response in B6 mice (23) and in rats using the same apparatus (38).
The apparatus consists of a translucent plastic box (49 cm square x 12 cm high with a lid containing ventilation holes) placed inside an aluminum frame containing a 15 x 15 matrix of infrared photosensors attached to a microcomputer (Optovarimex, Columbus Instruments, Columbus, OH). The plastic box and frame were housed inside a larger opaque black plastic box equipped with a translucent white lid to permit indirect illumina- tion from room lights and containing a ventilation fan in the center of the lid.
To begin the 10-min session, the experimenter removed the mouse from its home cage, placed it onto the middle of the floor of the apparatus, and quickly closed the tops of both the inner and outer boxes. About 10 s after placement of the mouse, a computer program was activated which could record the distance traveled based on the pattern of beam interruptions. Following removal of
CHRONIC DEPRENYL TREATMENT IN AGED MICE 433
TABLE l PSYCHOMOTOR TEST BATTERY
Day Test Performance Assessed
1 Open-Field
2 Tight Rope
3 Rotorod
4 Inclined Screen
5-8 Runwheel
9-10 Rotodrum
Locomotor activity during l0 min in a novel, illuminated environment
Time able to remain suspended by front paws from a taut string averaged over 3 trials
Falls during 3-rain placement on a plastic rod rotating at 3 rpm
Locomotor activity during l0 min on a wire mesh grid inclined 60 °
Locomotor activity during 72 h in a runwheel cage
Ability to remain on a rotating drum with different speeds presented during 1-min trials (0--60 cm/s)
the mouse at the end of each session, the experimenter removed all boli with a vacuum cleaner, sprayed the floor of the box with a 95% ethanol solution, and wiped the floor dry with paper towels. The performance variable for this test was the total distance moved during the 10-min session.
Tightrope (Day 2). This test measured motor strength and co- ordination as the ability to remain suspended by front paws from a taut string. The test has proven to be highly age sensitive in B6 mice (23) and in rats when using a wire or narrow rod (38). The apparatus consisted of cotton string tied tautly between two posts of a stainless box that contained wood shavings to soften the fall of the mouse from the string. The mouse was lifted gently by its tail from its home cage and suspended above the tightrope and then allowed to grasp the string securely with both front paws. The experimenter then moved the mouse's body below the tightrope and released it. If the mouse released its grip, it fell about l0 cm into the wood shavings and was given about 30 s to rest before retesting. Three such trials were given, with the performance vari- able estimated as the mean time before falling averaged across trials.
Rotorod (Day 3). Motor coordination was measured in this test by the mouse's ability to prevent falling from a rotating plastic rod. This test has also produced age-related performance declines in B6 mice (23) using the same apparatus and in rats using a similar apparatus (38). The apparatus consisted of a scored plastic rod (4.5 cm in diameter) that is rotated at 3 rpm through a pulley system connected to a 6 rpm motor. To begin a trial, the experi- menter picked up the mouse by its tail and placed it onto the rotating rod such that its body was parallel to the axis of the rod. A timer was then activated. To remain on the rotorod, the mouse was required to turn 90 ° and begin to locomote. If the mouse lost its balance, it fell about 25 cm onto a foam rubber cushion, and the experimenter deactivated the timer. After about 5-10 s, the mouse was returned to the rotorod and the clock reactivated. The perfor- mance measure was the number of falls during 3-min exposure to the rotorod. After testing each mouse, the experimenter cleaned the apparatus with an ethanol-soaked cloth and wiped it dry with paper towels.
Inclined screen (Day 4). This was a new apparatus designed to measure locomotor activity on a wire mesh that was tilted 60 ° to the horizontal to the plane of the table. Although not tested pre- viously in B6 mice, age-related deficits in performance in a similar apparatus had been observed in rats (54). The apparatus used was identical to that described for the open-field test with the following
exceptions. First, instead of a plastic floor, the floor was con- structed of a wire mesh (40 cm x 40 cm) attached to the plastic side (10 cm high). This box was still surrounded by the infrared matrix that permitted recording of locomotor activity. Second, instead of being housed inside the larger box, the apparatus was placed onto a wooden frame that tilted it 60 ° from the horizontal. Third, to prevent the mouse from crawling onto the side of the box closest to the top, a piece of clear plastic was tilted toward the inside of the box. The performance variable was the same as that measured in the open-field--distance traveled, but presumably this task required additional skills in motor coordination.
Runwheel (Days 5--8). Intended as a measure of general loco- motor activity, this apparatus has also been described previously and has yielded evidence of age-related performance declines in B6 mice (23) and in rats in a similar apparatus (38). The mouse is housed individually inside an activity-wheel cage (Wahmann, Bal- timore, MD) constructed from stainless steel and consisting of a stationary compartment (15 × 9 × 6 cm) and a run-wheel (15 cm in diameter), which is entered through a guillotine door from the stationary cage. Food and distilled water were provided in the stationary cage. This activity-wheel cage was housed inside a sound-attenuated wooden box equipped with a ventilation fan and automatically controlled lighting maintaining the same photope- riod experienced in the vivarium. The mouse was placed into the activity-cage about 2 h before the offset of lights on Day 5 and removed about the same time on Day 8. The performance variable for this test was the mean daily activity as measured by wheel revolutions.
Rotodrum (Days 9-10). This test was developed for B6 mice as a measure of maximum running speed requiring motor coordina- tion (23). A previous study in rats had documented age-related decline in this parameter of motor performance (22). The appara- tus is a commercially available automated drum exerciser designed for rodents (DREX, Columbus Instruments, Columbus, OH). It consisted of a drum (30 cm in diameter and 9.5 cm wide) con- raining a corrugated, rubber mat along the outside surface. Con- trolled by a microprocessor, a motor can be programmed to turn the drum at various speeds for various time intervals. The roto- drum is enclosed inside a clear plastic box that contains a stainless steel grid shelf (15 cm × 9 cm) positioned at a height about one-sixth below the diameter of the drum. A series of infrared sensors is located about 2 cm above the grid floor and is used to detect the presence of the mouse (and its tail) to activate a pulsed foot shock (0.5 mA at 0.5 s) and to record the number of visits to as well as the time on the shelf.
On Day 9 each mouse was provided a training session. The experimenter placed the mouse onto the stationary rotodrum for 2 min. If the mouse moved onto the shelf, it received the foot shock until it returned to the rotodrum. The apparatus was then pro- grammed to turn the rotodrum at a speed of 1 crn/s for a 1-min trial followed by a 1-min rest period when the rotodrum was stationary. This procedure was repeated by incrementing the speed of the rotodrum by 5 crn/s for each trial up to a maximum of 35 crn/s. If the mouse was exposed to the electrified shelf for 30 s, it was removed from the apparatus and the session was terminated. In this fashion, virtually all mice learned to run on top of the roto- drum with little possibility of being shocked at the lower speeds. On Day 10, the test session was provided by following the same general procedure with the following changes. The first trial began as 10 crn/s, and was incremented by 2 crn/s following each rest period thereafter to a maximum of 60 cm/s. Criterion for removal was 2 consecutive trials in which the mouse was detected on the shelf for at least 30 s. Thus, the performance measure was the maximum running speed achieved before being removed from the apparatus.
434 INGRAM ET AL.
Drug Treatment
About 1 week following the last behavioral test, the mice were assigned to treatment groups. Assignment to groups was quasi- random based on groups of 5 cage-mates as the experimental units and with adjustments made to insure equivalent baseline body weights across the 18-month-old groups. One 3-month and one 18-month-old mouse were dropped from the experiment because of poor performance in the tests.
Deprenyl (L-deprenyl; Research Biochemicals) was provided in the drinking water (distilled water). Specifically, body weight of each mouse and water intake of each cage were monitored semi-weekly by weighing the mice and water bottles, and per mouse consumption and body weight were then estimated for each cage. Based on these estimates, the concentration of deprenyl was adjusted to provide dosages of 0.5 mg/kg/day (LO-DEP) or 1.0 mg/kg/day (HI-DEP) to the experimental groups. Control groups of young (Y-CON) and aged (A-CON) mice received distilled water only. Cages were examined daily to check for any leakage of water tubes and to remove any dead mice.
Motor Battery: Test 2
After 6 months on their respective treatments, all mice were retested in the motor battery when the young group was about 9-10 month old and the aged group was about 24--25 months old. At this time, a group of 24-month-old mice was obtained from the GRC colony, and placed on the control, L0-DEP, and HI-DEP regimens (n = 8) for a period of 2-3 weeks. These mice were to be used for the analysis of MAO-B activity related to acute treat- ment.
Neurochemical Analysis
About 1 week after completing the motor battery, randomly selected members of each group were sacrificed by cervical dis- location, brains were quickly extracted, the striata were dissected on a cooled surface, and these along with the remaining whole brain were frozen on dry-ice, and then were stored at - 70°C until time of assay.
MAO-B assays. Striatal MAO-B activity was determined by the spectrofluorometric method of Naoi and Nagatsu (41). Briefly, one striatum was homogenized in 200 ixl of 0.25 M sucrose using an ultrasonic cell disrupter, and 50 ~,1 aliquots were assayed for MAO-B activity using kynuramine as substrate.
Catecholamine assays. Striatal samples for catecholamine analysis were prepared and assayed by reversed-phase high- performance liquid chromatography with electrochemical detec- tion by the method of Sershen et al. (45).
Neurohistochemistry. Frozen 16 Izm thick sections were cut using a Reichert cryostat and processed for histofluorescence using the sucrose-potassium phosphate-glyoxylic acid (SPG) method of de la Torre (52). Every fourth 16 Izm section was mounted on glass slides, fixed in 4% paraformaldehyde, rinsed in water and stained with cresyl violet. Using coded slides from all different groups of mice, the number of yellow fluorescent profiles depict- ing lipofuscin were quantitated from every eighth 16 ~m section through the substantia nigra using a Nikon microscope. Using cresyl violet stained sections, the numbers of neurons and glia were quantitated in the substantia nigra pars compacta (SNC) and substantia nigra pars reticulata (SNR) separately.
Motor Battery: Test 3
After the second test, a subsample of mice (n = 26) were maintained on their respective treatments and then retested in the
motor battery about 3 months later when the young group was 12-13 months of age and the aged groups were 27-28 months of age.
Statistical Analysis
Data from assays for striatal MAO-B activity were analyzed using a one-way analysis of variance (ANOVA) followed by a two-tailed t test comparing each group to the aged controls. Data on body weight and fluid intake were submitted to a 4 x 8 (treat- ment group x treatment interval) ANOVA. Survival data were analyzed by a nonparametric survival function analysis (BMDP-- Survivor Function P1L). Histological data were analyzed by one- way ANOVA followed by two-tailed t test comparisons. Signifi- cant differences were accepted at p < 0.05.
RESULTS
Striatal MAO-B Activity
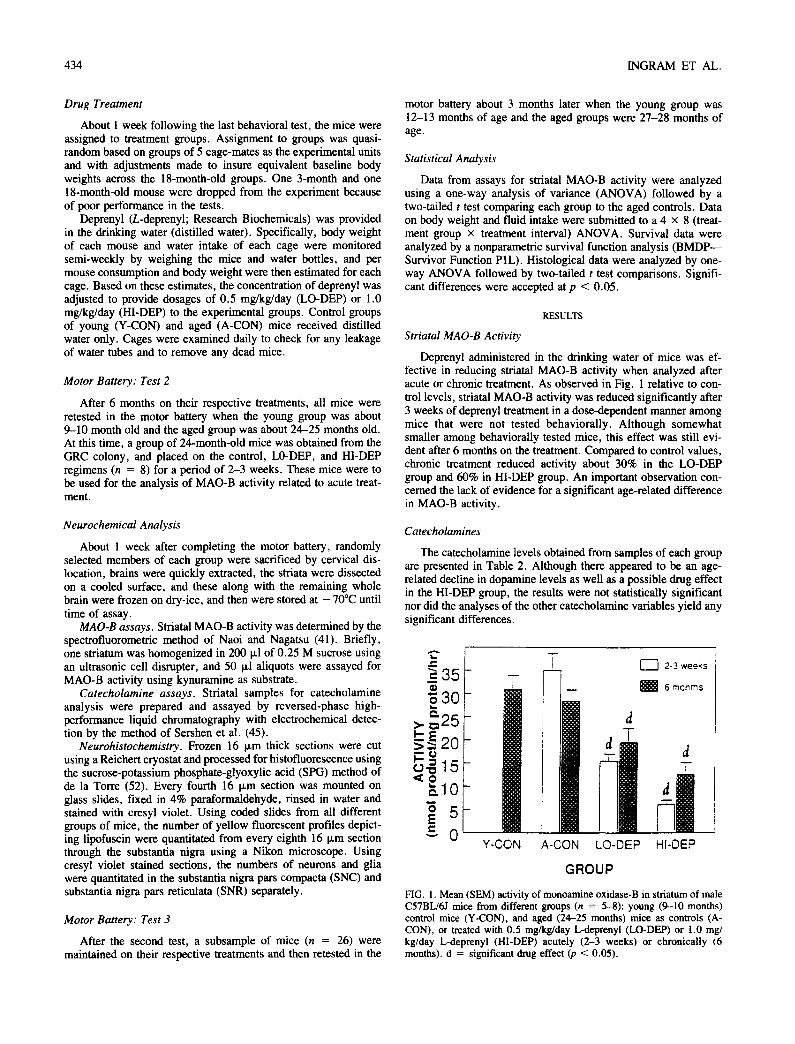
Deprenyl administered in the drinking water of mice was ef- fective in reducing striatal MAO-B activity when analyzed after acute or chronic treatment. As observed in Fig. 1 relative to con- trol levels, striatal MAO-B activity was reduced significantly after 3 weeks of deprenyl treatment in a dose-dependent manner among mice that were not tested behaviorally. Although somewhat smaller among behaviorally tested mice, this effect was still evi- dent after 6 months on the treatment. Compared to control values, chronic treatment reduced activity about 30% in the LO-DEP group and 60% in HI-DEP group. An important observation con- cerned the lack of evidence for a significant age-related difference in MAO-B activity.
Catecholamines
The catecholamine levels obtained from samples of each group are presented in Table 2. Although there appeared to be an age- related decline in dopamine levels as well as a possible drug effect in the HI-DEP group, the results were not statistically significant nor did the analyses of the other catecholamine variables yield any significant differences.
e,- .= 35
~25
"o '<C 15
o 5 E £ 0
7 i ~ - l 2.3 weeks
Y-CON A-CON LO-DEP HI-DEP
GROUP
FIG. 1. Mean (SEM) activity of monoamine oxidase-B in striatum of male C57BL/6J mice from different groups (n = 5-8): young (9-10 months) control mice (Y-CON), and aged (24-25 months) mice as controls (A- CON), or treated with 0.5 mg/kg/day L-deprenyl (LO-DEP) or 1.0 mg/ kg/day L-deprenyl (HI-DEP) acutely (2-3 weeks) or chronically (6 months), d = significant drug effect (p < 0.05).
* Means (SEM) in pmoles/mg protein for dopamine (DA); dihydroxy- phenylacetic acid (DOPAC homovanillic acid (HVA); 3-methoxytryamine (3-MT); 5-hydroxytryptamine (5-HT); and 5-hydroxyindoleacetic acid (5- HIAA).
n = 5 - 7 .
Body Weight and Fluid Intake
Chronic deprenyl treatment did not significantly affect body weight or fluid intake in the aged mice. Figure 2 shows that body weight remained constant throughout the 6-month treatment period for all mice except for Y-CON mice which increased in body weight for about 16 weeks to reach the plateau observed in older mice. The ANOVA for body weight data yielded no significant main effect of treatment time but did reveal the significant group x time interaction attributed to the growth of Y-CON mice. The ANOVA of fluid intake revealed no significant main effects of group or treatment time or their interaction.
Survival
As observed in Fig. 3, deprenyl had no significant effect on survival. Among all aged groups, survival after 6 months of treat- ment was about 70% and after 9 months, about 55%. However,
FIG. 2. Mean (SEM) body weight and fluid intake measures of male C57BI../6J mice from different groups (n = 15-30 for body weight and 6 for fluid intake which represents the intake per mouse in a cage): young (3 months at start of treatment) control mice (Y-CON), and aged (18 months at start of treatment) mice as controls (A-CON), or treated with 0.5 mg/ kg/day L-deprenyl (LO-DEP) or 1.0 mg/kg/day L-deprenyl (HI-DEP) acutely (2-3 weeks) or chronically (6 months).
(5 Z _> > cr
Go I-- z LLI 0 n" LU O.
100
90
8O
7oi
6O
5O 0
- : Y -CON {3 "~ A-CON i - - • LO .DE= I ! - - - - i i N I -OE~
\ ' . , 1 - - - a ~ . : . . . . . C . . m '
I I 1 f { ! i
4 8 12 16 20 24 28
WEEK
FIG. 3. Percent distribution of survival in male C57BL/6J mice from different groups (n = 29-30 at start of experiment): young (3 months) control mice (Y-CON); aged (18 months) control mice (A-CON); treated with 0.5 mg/kg/day L-deprenyl (LO-DEP); and 1.0 mg/kg/day L-deprenyl (HI-DEP) for 6 months.
statistical analysis did not reveal any significant differences among the aged groups at either treatment interval.
Motor Performance
Age effects. All tests were expected to yield evidence of cross- sectional age differences when comparing Y-CON to A-CON groups. As depicted in Fig. 4, this prediction of an age-related decline was confLrrned in the tightrope, rotorod, inclined screen, and runwheel tests, but not in the open-field or rotodrum tasks. In addition, a comparison of test scores observed at baseline and at the 6-month retest for the Y-CON and A-CON groups was ex- pected to yield evidence of a longitudinal decline in motor perfor- mance. For the Y-CON group, this prediction was confLrmed in the open-field, tightrope, and inclined screen tests; while for the A-CON group, results of the open-field, inclined screen, and run- wheel tests showed evidence of a longitudinal decline in perfor- mance. Thus, the rotodrum test was the only one showing no age sensitivity in this analysis.
Figure 5 provides a similar comparison but only for those mice that survived the 9 months of treatment. Therefore, because this testing lasted beyond the median age of survival (26 months), this procedure makes this population highly selective. Similar to the analyses just described, significant cross-sectional age differences (Y-CON vs. A-CON at baseline) were confmned for the tightrope, rotorod, and inclined screen task but not for the runwheel task although an age-related decline is apparent. Longitudinal declines in performance for the Y-CON were evident only for the open- field and for inclined screen tasks at the second test only. For A-CON mice, longitudinal declines in performance were observed in the open-field, inclined screen, runwheel, and rotodrum tests; however, the decline was not always consistently linear through to the last test.
Deprenyl effects. When performance in A-CON groups was compared to that of LO-DEP and HI-DEP groups at any age (Figs. 4 and 5), no significant effects of deprenyl treatment were ob- served in any test with only one exception. This occurred in the rotodrum test where the HI-DEP group showed higher maximum running speed compared to the A-CON group after 9 months of treatment.
436 INGRAM ET AL.
OPEN FIELD
~--~50 ~ 6-me Reres, v -- 1 l
so _
2 0 ' I = i i = ~ . 2 o - - -
m 01 Y-CON A-CON LO-OEP HI-DEP Y-CON A-CON LO-DEP HI-DEP
ROTOROD INCLINED SCREEN
TIGHTROPE
[r• Base.tie
6-too Retest
1 14 f-~ Bail,n,, [Z aase,,oe I t 12
~ 8631024 ~ ~ i c ~ i ~ 1 o~,10~15~205 II~ / -- l ._=..cl] i tl ~ i i - -
0 . _ _ 0 Y.CON " ; o ; o ; ; " ; ;
~ ~50 2 I -
0 0 Y-CON A-CON LO-DEP HI-DEP Y-CON A-CON LO-DEP HI-DEP GROUP GROUP FIG. 4. Mean (SEM) values comparing the motor performance of male C57BL/6J mice from different groups (n = 29-30 at start of experiment): young (3 months) control mice (Y-CON), and aged (18 months) mice as controls (A-CON), or treated with 0.5 mg/kg/day L-deprenyl (LO-DEP) or 1.0 mg/kg/day L-deprenyl (HI- DEP) for 6 mo. Data were analyzed as a 4 x 2 (group × test) ANOVA with repeated measures on the last factor. Where significant group effects were noted, these were followed by appropriate F tests for the main effects; and where significant interactions were detected, these were followed by F tests for the simple main effect of test within each group, c denotes significant (p < 0.05) cross-sectional age difference in performance comparing the Y-CON group to the A-CON group. ! denotes a significant (p < 0.05) longitudinal change in performance within a group across tests.
Cell Counts in the Substantia Nigra
Cell counts of neuronal and glial populations in the SNC and SNR were made in all four groups of mice as shown in Fig. 6. Neurons were characterized by a large, relatively uniform size, lightly stained nucleus and Nissl substance in the cytoplasm. Glia were characterized by a small densely stained nucleus and an unstained cytoplasm. It is clear that the number of neurons in the SNC were much higher compared to those in the SNR due to the presence of a significantly large number of dopaminergic neurons
in the SNC. However, no significant effects of age or deprenyl were observed in counts of neurons or glia.
Lipofuscin-Containing Nigral Neurons
The estimated number of lipofuscin profiles is presented in Fig. 7 for each group. A significant age-related increase (--20%) in lipofuscin was observed, but there was no significant difference detected among the aged treatment groups.
FIG. 5. Mean (SEM) values comparing the motor performance of male C57BL/6J mice from different groups (n = 29-30 at start of experiment): young (3 months) control mice (Y-CON), and aged (18 months) mice as controls (A-CON), or treated with 0.5 mg/kg/day L-deprenyl (LO-DEP) or 1.0 mg/kg/day L-deprenyl (HI- DEP) for 9 months; Data were analyzed as a 4 x 3 (group x test) ANOVA with repeated measures on the last factor. Where significant group effects were noted, these were followed by appropriate F tests for the main effects; where significant interactions were detected, these were followed byF tests for the simple main effect of test within each group, c denotes significant (p < 0.05) cross-sectional age difference in performance comparing the Y-CON group to the A-CON group. I denotes a significant (19 < 0.05) longitudinal change in performance within a group across tests, d denotes a significant (p < 0.05) drug effect as compared to respective A-CON group.
DISCUSSION
The present results demonstrate the safety and efficacy of orally administered L-deprenyl as a MAO-B inhibitor for use in chronic studies of mice at the doses studied. When provided 1.0 mg/kg orally for 6 months, L-deprenyl reduced striatal MAO-B activity by 60% compared to control values. We observed no toxic effects on body weight or fluid intake in treated mice. These findings are consistent with the report of no evidence of toxicity
following chronic SC treatment with 0.25 mg/kg deprenyl in rats (39).
Despite these positive findings regarding the pharmacological effects, we did not observe any beneficial effects of chronic L-de- prenyl treatment on survival or motor performance with only one exception--performance on the rotodrum in mice treated for 9 months. Nor did we observe any significant effects of L-deprenyl treatment on catecholamine levels or the accumulation of lipofus- cin pigment in the substantia nigra. The lack of any beneficial
438 INGRAM ET AL.
+1
P 800
E
0
SNC-Neurons SNC-Glla SNR-Neurons SNR-Glia
Groups n Young Control n Aged Control
l AgedLoDcse
q AgedHiDose
FIG. 6. Mean (SEM) counts of neurons and glia in substantia nigra pars compacta (SNC) and pars reticulata (SNR) in male C57BW6J mice from four different groups: young (3 months) control mice (Y-CON), and aged (18 months) mice as controls (A-CON), or treated with 0.5 mglkgiday L-deprenyl (LO-DEP) or 1.0 mglkglday L-deprenyl (HI-DEP) for 6 months. A one-way ANOVA for each variable revealed no significant effects of age or deprenyl treatment @ > 0.05).
effects on age-sensitive parameters, including survival, contrasts with previous findings (4,28,32,33,39). This discrepancy should be addressed with respect to several procedural differences.
Among the major procedural differences from previous pub- lished studies reporting increased survival in L-deprenyl treated rats (33,39) include the species (mouse vs. rats), the age of treat- ment initiation (middle age vs. aged), and the route of adminis- tration (oral vs. SC). We selected the B6 mouse for analysis be- cause of the known sensitivity of this strain for MPTP toxicity (48) and the relatively high level of striatal MAO-B activity (55). We chose to begin treatment at middle age because we considered that earlier intervention might be even more beneficial. And we used the oral route of administration notably because of the practical advantages but also because this is the preferred route in human applications and has been shown to be effective in treatment of Parkinson’s disease (51). Our analysis of striatal MAO-B inhibi- tion indicated that this route of administration was effective; how- ever, the 60% inhibition observed may not have been high enough to obtain the maximal benefits of the treatment. For example, a recent report indicated that oral treatment with 0.5 mgikg begin- ning at 12 months of age in male F-344 rats was successful in increasing median lifespan by about 15% but with a near 90% inhibition of striatal MAO-B activity (4).
The precise dose of deprenyl necessary for optimal effects on different parameters is apparently very dependent on genetic and age factors. Carrillo et al. (9) reported that the striatal increase in SOD activity following 3 weeks of SC treatment with L-deprenyl in rats differed greatly between males and females and across age. For example, these investigators noted that the optimal dose for increasing striatal SOD activity was ten-fold higher in male com- pared to female F-344 rats. Moreover, a dose of 2.0 mgikg L-de- prenyl in young (4-6 months) female rats decreased SOD activi- ties; whereas, there was no effect of this dose in aged (26-28 months) female rats. This reduced effect on SOD activity in older
rats was speculated to be due to increased levels of MAO-B as a substrate for deprenyl; however, MAO-B was not assayed in their study. In our study, we had expected to observe an age-related increase in striatal MAO-B activity, but we did not. In male rat strains for which there is an age-related increase in striatal MAO-B activity, Carrillo et al. (9) predicted that the optimal deprenyl dose for increasing SOD activity would bc lower than in younger rats. Since we did not assay for SOD activity, we do not know if our dosage was optimal in this regard, which might be the most im- portant factor for sparing the integrity of the nigro-striatal pathway.
On the other hand, an important question to consider is why should L-deprenyl increase SOD activity. Some investigators have emphasized that methamphetamine is one of the metabolites of deprenyl which might be toxic (27); thus, the upregulation of SOD might be in response to a toxic insult. Indeed, in brains of persons with Parkinson’s disease, SOD activity has been reported higher in several btain regions, including the substantia nigra, as a possible compensatory response to sustained oxidative stress occurring in mitochondria and neutonal cytosol (1). However, the observation that the deprenyl-induced SOD increase in rat brain was regionally specific, e.g., there was no effect in hippocampus (9), would argue against a generalized response to methamphetamine toxicity.
Regarding the lack of robust effects on motor performance, few other studies have examined this aspect, but a recent chronic study using oral administration of deptenyl (0.5 mg/kg) in male F-344 rats found no significant effects of 6 and 12 months of treatment on performance in several motor tasks (4). Moreover, in a motor learning task, the deprenyl treated animals actually showed some degree of impairment (4). Knoll (33,34) reported that chronic treatment with 0.25 mgikg SC L-deprenyl improved the shuttle avoidance of young male rats that had shown impotence in tests of copulatory abilities. We applied the present battery of motor tests because of its proven age-sensitivity in B6 mice (23-25); however, we had not examined longitudinal change in performance except in one study that was initiated at 24 months of age (26). Inconsistent with past results was the lack of a significant cross-sectional age effect on performance in the open-field and rotodrum tests (22,23). While in the longitudinal analysis of performance in the aged group of mice, significant age effects were observed only in the open-field, tightrope, and inclined screen tests, but none showed a significant effect of deprenyl treatment.
We also found no significant effects of our deprenyl regimen on age-sensitive neurohistological parameters, specifically nigral cells containing lipofuscin. If deprenyl was acting as an antioxi- dant, then we hypothesized that brain lipofuscin accumulation should have been affected as it is with manipulations of other antioxidants, such as Vitamin E (40). Our finding contrasts with a recent report that chronic deprenyl treatment in 19-month-old rats retarded lipofuscin accumulation in hippocampal pyramidal cells as well retarded the age-associated loss in mossy fibers (3). This study employed SC treatment with 0.25 mgfkg and oral treatment at 1.25 and 5 mg/kg for 5 months. Thus, again it is possible that our dose of deptenyl was not high enough to see an effect on lipofuscin accumulation.
Our regimen of deprenyl also had no significant effects on striatal levels of dopamine or its metabolites. This lack of drug treatment effect was consistent with a previous report using de- prenyl injections IP in BALB/cBy mice, which suggested that MAO-B inhibition would have to be greater than 60% to impact on striatal dopamine metabolism (57). Moreover, this same report indicated that chronic deprenyl treatment in a wide range of doses could not restore striatal dopamine following MPTP treatment in young mice.
In summary, our study failed to find any major beneficial ef- fects of chronic oral treatment with L-deprenyl when provided to
CHRONIC DEPRENYL TREATMENT IN AGED MICE 439
1600]
1400
o o 1200
o 1000
o
800' o
600' o
400 E Z
200
(7)
Y-CON A-CON LO-DEP HI-DEP
Group
FIG. 7. Mean (SEM) counts of cells containing lipofuscin fluorescent profiles in substantia nigra of different groups of male C57BL/6J mice: young (3 months) control mice (Y-CON), and aged (18 months) mice as controls (A-CON), or treated with 0.5 mg/kg/day L-deprenyl (LO-DEP) or 1.0 mg/kg/day L-deprenyl (HI-DEP) for 6 months. A one-way ANOVA for each variable followed by individual two-tailed t test comparisons to the A-CON group revealed a significant effect of age (*p < 0.05) but not to deprenyl (p > 0.05).
18-month-old male B6 mice to produce striatal MAO-B inhibition of up to 60%. As the drug has proven safe thusfar, any additional experimentation is certainly justified but will have to consider the issue of dosage, route of administration, and different responses among species to provide support for the generalized effect of chronic L-deprenyl on aging processes.
ACKNOWLEDGEMENTS
We appreciate the assistance of Laura Gordon, Ying Lin, and Michael Anson in tissue preparation. This work was supported in part by USPHS Grant NS24291 to M. G.
This research was fully accredited by the American Association for Accreditation of Laboratory Animal Care.
REFERENCES
1. Adams, J. D., Jr.; Odunze, I. N. Oxygen free radicals and Parkin- son's Disease. Free Radical Biol. Med. 10:161-169; 1991.
2. Adolfsson, R.; Gottfries, C. G.; Oreland, L. Increased activity of brain and platelet monoamine oxidase in dementia of the Alzheimer type. Life Sci. 27:1029-1034; 1980.
3. Amenta, F.; Bronzetti, E.; Felici, L.; Ricci, A.; Cadel, S.; Bongrani, S.; Valsecchi, B. Effect of long term L-deprenyl treatment on the structure of the hippocampus of aged rat. Neurobiol. Aging 13:S 134; 1992.
4. Bickford, P. C.; Heron, C.; Rose, G. M.; Lin, A.; Friedemann, M.; Gerhardt, G. A. Effects of long term deprenyl administration on be- havior and noradrenergic neurotransmission in aged rats (in press).
5. Birkmayer, W.; Riederer, P.; Youdin, M. B. H.; Linauer, W. The potentiation of the antiakinetic effect after L-dopa treatment by an inhibitor of MAO-B, deprenyl. J. Neural Transm. 36:225-228; 1975.
6. Birkmayer, W.; Riederer, P.; Ambrozi, L.; Youdim, M. B. H. Im- plications of combined treatment with madopar and L-deprenyl in Parkinson's disease. Lancet 1:439-444; 1977.
7. Birkmayer, W.; Knoll, J.; Riederer, P.; Hars, V.; Marton, J. Im- provement of life expectancy due to L-deprenyl addition to madopar
treatment in Parkinson's disease: A long-term study. J. Neural Transm. 64:113-127; 1985.
8. Carrillo, M. C.; I Kanai, S.; Nokubo, M.; Kitani, K. (-)Deprenyl induces activities of both superoxide and catalase but not of glutathi- one peroxidase in the striatum of young male rats. Life Sci. 48:517- 521; 1991.
9. Carrillo, M. C.; Kanai, S.; Nokubo, M.; Ivy, G. O.; Sato, Y.; Kitani, K. (-)Deprenyl increases activities of superoxide and catalase in stri- atum but not hippocampus: The sex and age-related differences in the optimal dose in the rat. Exp. Neurol. 116:286-294; 1992.
10. Cohen, G. Monoamine oxidase, hydrogen peroxide and Parkinson's disease. Adv. Neurol. 45:119-125; 1986.
l l . Cohen, G.; Pasik, P.; Cohen, B.; Leist, A.; Mytilineou, C.; Yahr, M. D. Pargyline and deprenyl prevent the neurotoxicity of 1-methyl- 4-phenyl-l,2,3,6-tetrahydropyridine (MPTP) in monkeys. Eur. J. Pharmacol. 106:209-210; 1984.
12. Cohen, G., Spina, M. B. Deprenyl suppresses the oxidant stress as- sociated with increased dopamine turnover. Annals Neurol. 26:689- 690; 1989.
13. Ebel, A.; Strosser, M. T.; Kempf, E. Genotypic differences in central
440 INGRAM ET AL.
neurotransmission responses to aging in mice. Neurobiol. Aging 8: 417-427; 1987.
14. Elizan, T. S.; Yahr, M. D.; Moros, D. A.; Mendoza, M. R.; Pang, S.; Bodian, C. A. L-deprenyl, a MAO-B inhibitor, as an adjunct to conventional L-dopa therapy in Parkinson's disease: Experience in 200 patients. Adv. Neurol. 53:431-435; 1990.
15. Frankel, J. P.; Kempster, P. A.; Stibe, M. H.; Eatough, V. M. H.; Nathanson, M.; Lees, A. J.; Stern, G. M. A double-blind controlled study of high-dose L-deprenyl in treatment of Parkinson's disease. Clin. Neuropharmacol. 12:448--451; 1989.
16. Golbe, L. I.; Lieberman, A. B.; Muenter, M. D.; Ashlskog, J. E.; Gopinathan, G.; Neophytides, A. N.; Foo, S. H.; Duvoisin, R. C. Clin. Neuropharmacol. 11:45-55; 1988.
17. Golbe, L. I. Long-term efficacy and safety of deprenyl (selegiline) in advanced Parkinson's disease. Neurol. 39:1109-1111; 1989.
18. Goodrick, C. L.; Ingram, D. K.; Reynolds, M. A.; Freeman, J. R.; Cider, N. Effects of intermittent feeding upon body weight and lifespan in inbred mice: Interaction of genotype and age. Mech. Age- ing Devel. 55:69-87; 1990.
19. Harman, D. Aging: A theory based on free radical and radiation chemistry. J. Gerontol. 11:289-300; 1956.
20. Heikkila, R. E.; Manzino, L.; Cabbat, F. S.; Duvoisin, R. C. Pro- tection against the dopaminergic neurotoxicity of l-methyl-4-phenyl- 1,2,5,6-tetrahydropyridine by monoamine oxidase inhibitors. Nature 311:467-469; 1984.
21, Henry, J. M.; Filburn, C. R.; Joseph, J. A.; Roth, G. S. Effect of aging on striatal dopamine receptor subtypes in Wistar rats. Neuro- biol. Aging 7:357-361; 1986.
22. Hofecker, G.; Skalicky, M.; Kment, A.; Niedermuller, H. Models of biological age of the rat. I. A factor model of age parameters. Mech. Ageing Devel. 14:345-359; 1980.
23. Ingram, D. K. Toward the behavioral assessment of biological aging in the laboratory mouse: Concepts, terminology, and objectives. Exp. Aging Res. 9:225-238; 1983.
24. Ingram, D. K. Motor performance variability during aging in rodents: An assessment of reliability and validity of individual differences. Ann. NY Acad. Sci. 515:70-96; 1988.
25. Ingram, D. K. Behavioral and neurobiological effects of dietary re- striction in aged rodents. In: Fishbein, L., ed. Biological effects of dietary restriction. Berlin: Springer-Verlag; 1991:157-171.
26. Ingram, D. K.; Reynolds, M. A. Assessing the predictive validity of psychomotor tests as measures of biological age in mice. Exp. Aging Res. 12:155-162; 1986.
27. Karoums, F. L. W.; Chuang, W.; Eisler, T.; Calne, D.; Liebowitz, M. R.; Quotkin, F. M.; Wyatt, R. J. Metabolism of (-)deprenyl to amphetamine and methamphetamine may be responsible for depre- nyl's therapeutic benefit: A biochemical assessment. Neurol. 32:503- 509; 1982.
28. Kitani, K.; Kanai, S.; Sato, Y.; Ivy, G. O.; Carrillo, M. C. Chronic treatment of (-)deprenyl prolongs lifespan of male Fischer 344 rats (in press).
29. Knoll, J. The possible mechanism of action of (-)deprenyl in Parkin- son's disease. J. Neural Transmsn. 43:177-198; 1978.
30. Knoll, J. The pharmacology of selective MAO inhibitors. In: Youdim, M. B. H.; Paykel, E. S., eds. Monoamine oxidase inhibitors. The state of the art. Chichester: John Wiley & Sons; 1981:45-61.
31. Knoll, J. Selective inhibition of B type monoamine oxidase in the brain. A drug strategy to improve the quality of life in senescence. In: Keverling-Buisman, J. A., ed. Strategy of drug research. Amsterdam: Elsevier/North Holland; 1982:107-135.
32. Knoll, J. Long-term (-)deprenyl treatment. Mt. Sinai J. Med. 55:67- 74; 1988.
33. Knoll, J. The striatal dopamine dependence of life span in male rats. Longevity study with (-)deprenyl. Mech. Ageing Devel. 46:237-262; 1988.
34. Knoll, J. The pharmacology of selegiline ((-)deprenyl). New aspects. Acta Neurol. Scand. 126:83-91; 1989.
35. Knoll, J. Nigrostriatal dopaminergic activity, deprenyl treatment, and longevity. Adv. Neurol. 53:425-429; 1990.
36. Knoll, J.; Ecery, Z.; Keleman, K.; Nievel, J.; Knoll, B. Phenyliso- propyl-methylpropinylamine (E-250), a new spectrum psychic ener- gizer. Arch. Int. Pharmacodyn. Ther. 155:154; 1965.
37. Mann, D. M. A.; Lincoln, J.; Yates, P. O.; Stamp, J. E.; Toper, S. Changes in the monoamine containing neurones of the human CNS in senile dementia. Br. J. Pscyhia. 136:533-541; 1980.
38. Markowska, A. J.; Ingram, D. K.; Barnes, C. A.; Spangler, E. L.; Lemken, V. J.; Kametani, H.; Yee, W.; Olton, D. S. Acetyl-1- carnitine 1: Effects on mortality, pathology and sensory-motor per- formance in aging rats. Neurobiol. Aging 11:491-498; 1990.
39. Milgram, N. W.; Racine, R. J.; Nellis, P.; Mendonca, A.; Ivy, G. O. Maintenance of L-deprenyl prolongs life in aged male rats. Life Sci. 47:415-420; 1990.
40. Nandy, K. Lipofuscin as a marker of impaired homeostasis in aging organisms. In: Davis, B. B.; Wood, W. G., eds. Homeostatic func- tion and aging. New York: Raven Press; 1985:13%148.
41. Naoi, M.; Nagatsu, T. Inhibition of monoamine oxidase by 3,4- dihydroxyphenylserine. J. Neurochem. 47:604-607; 1986.
42. Oreland, L.; Gottfries, C. G. Brain and monoamine oxidase in aging and in dementia of Alzheimer's type. Prog. Neuro-Psychopharmacol. Biol. Psychia. 10:533-540; 1986.
43. Piccinin, G. L.; Finali, G.; Piccirilli, M. Neuropsychological effects of L-deprenyl in Alzheimer's type dementia. Clin. Neuropharmacol. 13:147-163; 1990.
44. Przuntek, H.; Kuhn, W. The effect of R-(-)-deprenyl in de novo Parkinson patients on combination therapy with levodopa and decar- boxylase inhibitor. J. Neural Transm. (Suppl). 25:97-104; 1987.
45. Sershen, H. A.; Hashim, A.; Lajtha, A. Behavioral and biochemical effects of nicotine in an MPTP-induced mouse model of Parkinson's disease. Pharmacol. Biochem. Behav. 28:29%303; 1987.
46. Strolin-Benedetti, M. S.; Keane, P. E. Differential changes in mon- oxidase A and B activity in the aging rat brain. J. Neurochem. 35: 1026-1032; 1980.
47. Strolin-Benedetti, M. S.; Dostent, P. Monoamine oxidase, brain age- ing and degenerative diseases. Biochem. Pharmacol. 38:555-561; 1989.
48. Sundstrom, E.; Stromberg, 1.; Tsutsumi, T.; Olson, L.; Jonsson, G. Studies on the effect of I-methyl-4-phenyl-l,2,3,6-tetrahydro- phyridine (MPTP) on central catecholamine neurons in C57BL/6J mice: Comparison with three other strains of mice. Brain Res. 405: 26-38; 1987.
49. Talan, M. I.; Engel, B. T.; Whitaker, J. R. A longitudinal study of tolerance to cold stress among C57BL/6J mice. J. Gerontol. 40:8--14; 1985.
50. Tariot, P. N.; Cohen, R. M.; Sunderland, T.; Newhouse, P. A.; Yount, D.; Mellow, A. M.; Weingartner, H.; Mueller, E. A.; Mur- phy, D. L. L-deprenyl in Alzheimer's disease. Arch. Gen. Psychiat. 44:427-433; 1987.
51. The Parkinson Study Group. Effect of deprenyl on the progression of disability in early Parkinson's disease. N. Eng. J. Med. 321:1364- 1371; 1989.
52. de la Torre, J. C. Standardization of the sucrose-potassium phosphate- glyoxylic acid histofluorescence method for tissue monoamines. Neu- rosci. Lett. 17:33%340; 1980.
53. Venero, J. L.; Machado, A.; Cano, J. Changes in monoamines and their metabolite levels in substantia nigra of aged rats. Mech. Ageing Devel. 49:227-233; 1989.
54. Wallace, J. E.; Krauter, E. E.; Campbell, B. A. Motor and reflexive behavior in the aging rat. J. Gerontol. 35:364-370; 1980.
55. Wiener, H. L.; Hashim, A.; Lajtha, A.; Sershen, H. Age and strain differences in monoamine oxidase B in mouse brain. Res. Commun. Psychol. Psychiat. Behav. 13:306; 1988.
56. Wiener, H. L.; Hashim, A.; Lajtha, A.; Sershen, H. Chronic L-de- prenyl-induced upregulation of the dopamine uptake carrier. Eur. J. Pharmacol. 163:191-194; 1989.
57. Wiener, H. L.; Hashim, A.; Lajtha, A.; Sershen, H. Chronic L-de- prenyl does not alter the restoration of striatal dopamine in MPTP- lesioned mice. J. Neurosci. Res. 23:326-329; 1989.




















![striatal binding of 2-amino-6,7-[3h]dihydroxy-1,2,3,4](https://static.documents.pub/doc/80x56/586bd52b1a28ab84588b456b/striatal-binding-of-2-amino-67-3hdihydroxy-1234.jpg)





