Elisabetta Vivarelli, Luigi Frati, Giulio Cossu and Alessandro GiacomelloFiordaliso, Monica Salio, Massimo Battaglia, Michael V.G. Latronico, Marcello Coletta,
Elisa Messina, Luciana De Angelis, Giacomo Frati, Stefania Morrone, Stefano Chimenti, FabioIsolation and Expansion of Adult Cardiac Stem Cells From Human and Murine Heart
is online at: Circulation Research Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer about this process is available in the
located, click Request Permissions in the middle column of the Web page under Services. Further informationEditorial Office. Once the online version of the published article for which permission is being requested is
can be obtained via RightsLink, a service of the Copyright Clearance Center, not theCirculation Researchin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on May 2, 2014http://circres.ahajournals.org/Downloaded from by guest on May 2, 2014http://circres.ahajournals.org/Downloaded from by guest on May 2, 2014http://circres.ahajournals.org/Downloaded from by guest on May 2, 2014http://circres.ahajournals.org/Downloaded from by guest on May 2, 2014http://circres.ahajournals.org/Downloaded from by guest on May 2, 2014http://circres.ahajournals.org/Downloaded from by guest on May 2, 2014http://circres.ahajournals.org/Downloaded from by guest on May 2, 2014http://circres.ahajournals.org/Downloaded from by guest on May 2, 2014http://circres.ahajournals.org/Downloaded from by guest on May 2, 2014http://circres.ahajournals.org/Downloaded from by guest on May 2, 2014http://circres.ahajournals.org/Downloaded from by guest on May 2, 2014http://circres.ahajournals.org/Downloaded from by guest on May 2, 2014http://circres.ahajournals.org/Downloaded from by guest on May 2, 2014http://circres.ahajournals.org/Downloaded from by guest on May 2, 2014http://circres.ahajournals.org/Downloaded from by guest on May 2, 2014http://circres.ahajournals.org/Downloaded from by guest on May 2, 2014http://circres.ahajournals.org/Downloaded from by guest on May 2, 2014http://circres.ahajournals.org/Downloaded from by guest on May 2, 2014http://circres.ahajournals.org/Downloaded from by guest on May 2, 2014http://circres.ahajournals.org/Downloaded from by guest on May 2, 2014http://circres.ahajournals.org/Downloaded from by guest on May 2, 2014http://circres.ahajournals.org/Downloaded from by guest on May 2, 2014http://circres.ahajournals.org/Downloaded from by guest on May 2, 2014http://circres.ahajournals.org/Downloaded from
Elisabetta Vivarelli, Luigi Frati, Giulio Cossu, Alessandro Giacomello
Abstract—Cardiac myocytes have been traditionally regarded as terminally differentiated cells that adapt to increased workand compensate for disease exclusively through hypertrophy. However, in the past few years, compelling evidence hasaccumulated suggesting that the heart has regenerative potential. Recent studies have even surmised the existence ofresident cardiac stem cells, endothelial cells generating cardiomyocytes by cell contact or extracardiac progenitors forcardiomyocytes, but these findings are still controversial. We describe the isolation of undifferentiated cells that growas self-adherent clusters (that we have termed “cardiospheres”) from subcultures of postnatal atrial or ventricular humanbiopsy specimens and from murine hearts. These cells are clonogenic, express stem and endothelial progenitor cellantigens/markers, and appear to have the properties of adult cardiac stem cells. They are capable of long-termself-renewal and can differentiate in vitro and after ectopic (dorsal subcutaneous connective tissue) or orthotopic(myocardial infarction) transplantation in SCID beige mouse to yield the major specialized cell types of the heart:myocytes (ie, cells demonstrating contractile activity and/or showing cardiomyocyte markers) and vascular cells (ie,cells with endothelial or smooth muscle markers). (Circ Res. 2004;95:911-921.)
Key Words: adult stem cell � myocardial regeneration and angiogenesis
Cardiac myocytes have been traditionally regarded asterminally differentiated cells that adapt to increased
work and compensate for disease exclusively through hyper-trophy.1 In the past few years, compelling evidence hasaccumulated suggesting that the heart has regenerative poten-tial.2–5 The origin and significance of the subpopulation ofreplicating myocytes are unknown; these issues could berelevant to understand the for mechanisms coaxing endoge-nous cardiomyocytes to reenter the cell cycle and to thesearch for strategies to transplant cardiac progenitor cells.6 Infact, although embryonic stem cells have an exceptionalcapacity for proliferation and differentiation, potential immu-nogenic, arrhythmogenic, and, particularly, ethical consider-ations limit their current use. Moreover, autologous trans-plantation of skeletal myoblasts has been considered becauseof their high proliferative potential, their commitment to awell-differentiated myogenic lineage, their resistance to is-chemia, and their origin, which overcomes ethical, immuno-logical, and availability problems. However, even if phase IIclinical trials with autologous skeletal myoblasts are ongoing,several problems related to potentially life-threatening ar-
rhythmia (perhaps reflecting cellular uncoupling with hostcardiomyocytes7) must be taken into account when thisapproach is considered. Furthermore, although cardiomyo-cytes can be formed, at least ex vivo, from different adultstem cells, the ability of these cells to cross lineage bound-aries is currently causing heated debate in the scientificcommunity,8 with the majority of reports indicating neoan-giogenesis as the predominant in vivo effect of bone marrowor endothelial progenitor cells.9,10
This report describes the identification and preliminary char-acterization of cells from the adult human and murine heart,which have the properties of cardiac stem cells. Because thesecells also have been isolated and expanded from human heartbiopsy specimens, they could have a significant impact on futureclinical strategies to treat patients with heart disease.
Materials and MethodsTissue SamplesHuman tissue was derived from atrial or ventricular biopsy speci-mens belonging to patients (1 month to 80 years of age) undergoingheart surgery, in conformation with the guidelines of the Italian
Original received December 5, 2003; first resubmission received June 16, 2004; second resubmission received August 24, 2004; revised resubmissionreceived September 28, 2004; accepted September 29, 2004.
From the Department of Experimental Medicine and Pathology (E.M., S.M., M.V.G.L., L.F., A.G.), University La Sapienza, Rome; IRCCS “Neuromed (E.M.,S.M., M.V.G.L., L.F., A.G.),” Pozzilli (IS); Department of Histology and Medical Embriology (L.D.A., M.C., E.V., G.C.), University La Sapienza, Rome;Campus-Biomedico University (G.F.), Rome; Institute of Neurobiology and Molecular Medicine (M.B.), Consiglio Nazionale delle Ricerche, Rome; Stem CellResearch Institute (G.S.), Ospedale San Raffaele, Milan; Istituto di Ricerche Farmacologiche “Mario Negri” (S.C., F.F., M.S.), Milan, Italy.
Correspondence to Alessandro Giacomello, Department of Experimental Medicine and Pathology, University La Sapienza, 00161 Rome, Italy; [email protected]; and to Giulio Cossu, Department of Histology and Medical Embriology, University La Sapienza, 00161 Rome, Italy.E-mail [email protected]
Department of Health. Murine tissue was derived from the hearts ofpreviously characterized homozygous MLC1/3F-nlacZ11 and cTnI-nlacZ12 transgenic mice expressing a nuclear lacZ transgene underthe transcriptional control of the striated muscle myosin light chainor cTnI promoters, respectively, homozygous B5-eGFP mice,13
homozygous GFP-cKit14 mice, MLC3F-nlacZ/B5-eGFP, MLC3F-nlac-Z/GFP-cKit, and cTnI-nlacZ/B5-eGFP cTnI-nlac-Z/GFP-cKitcrossed mice, SCID mice, and SCID beige mice (Charles RiverItalia, Lecco, Italy).
Processing, Isolation, and Cryopreservation ofSphere-Forming CellsIsolated myocardial tissue was cut into 1- to 2-mm3 pieces, washedwith Ca2�-Mg2�–free phosphate-buffered solution (PBS) (Invitro-gen), and digested three times for 5 minutes at 37°C with 0.2%trypsin (Invitrogen) and 0.1% collagenase IV (Sigma, Milan, Italy).The obtained cells were discarded, and the remaining tissue frag-ments washed with complete explant medium (CEM) (Iscove’sModified Dulbecco’s Medium [IMDM] supplemented with 10%fetal calf serum, 100 U/mL penicillin G, 100 �g/mL streptomycin,2 mmol/L L-glutamine, and 0.1 mmol/L 2-mercaptoethanol) werecultured as explants in CEM at 37°C and 5% CO2. After a periodranging from 1 (embryo) to 3 (adult) weeks, a layer of fibroblast-likecells was generated from adherent explants over which small,phase-bright cells migrated. These phase-bright cells were collectedby pooling two washes with Ca2�-Mg2�–free PBS, one wash with0.53 mmol/L EDTA (Versene, Invitrogen) (1 to 2 minutes), and onewash with 0.5 g/L trypsin and 0.53 mmol/L EDTA (Invitrogen) (2 to3 minutes) at room temperature under visual control. The cellsobtained (from 104 to 4�105 cells/explant) were seeded at �0.5 to2�105 cells/mL in poly-D-lysine-coated multiwell plates (BD Bio-scences, Milan, Italy) in cardiosphere-growing medium (CGM)(35% complete IMDM/65% DMEM–Ham F-12 mix containing 2%B27, 0.1 mmol/L 2-mercaptoethanol, 10 ng/mL epidermal growthfactor [EGF], 20 ng/mL basic fibroblast growth factor [bFGF],40 nmol/L cardiotrophin-1, 40 nmol/L thrombin, antibiotics, andL-Glu, as in CEM). Isolation of the cardiosphere-forming cells couldbe performed at least 4 times at 6- to 10-day intervals from the sameexplant. Cardiospheres (CSs) were passaged every 2 to 3 days bypartial changing of the medium and mechanical trituration of thelarger clusters. Movies of cultured CSs, available in the online datasupplement at http://circres.ahajournals.org, were recorded using aNikon-COOLPIX-4500 digital camera connected to a Leitz invertedmicroscope. For cryopreservation, we used CEM/DMEM–Ham F12at 50:50, 5% B27, and 10% DMSO as the freezing medium.
Extensive descriptions of BrdUrd labeling, clonal analysis, differ-entiation on substrate-coated surface, coculture experiment, immu-nocytochemistry, flow cytometric analysis, in vivo analysis, andheterotopic and orthotopic transplantation are provided in the onlinedata supplement.
ResultsIsolation, Expansion, and Features in Culture ofSphere-Generating CellsSphere-generating cells were obtained by mild enzymaticdigestion of explanted human atrial or ventricular biopsyspecimens and embryo, fetal, and postnatal mouse hearts.Soon after the generation of a layer of fibroblast-like cellsfrom well-adherent explants, some small, round, phase-brightcells began to migrate over this coat. These cells could beharvested periodically by treatment with EDTA and mildtrypsinization and were allowed to grow on poly-D-lysine–coated culture surfaces in a low-serum (3.5% fetal calf serum)medium supplemented with a serum substitute (B27), growthfactors (EGF and bFGF), cardiothrophin-1 (CT-1),15 andthrombin.16 During the first week of culture, the last factor ledto a 7-fold increase in the number of spheres with respect to
that obtained using the medium supplemented with the otherfactors, either alone or in combination. Time-course obser-vations of cells derived from human and murine explantsshowed that early after their seeding (30 minutes), some ofthese cells began to divide while still in suspension. Mostcells became loosely adherent, whereas others remained insuspension, and some contaminating fibroblast-like cellsattached firmly to the poly-D-lysine coat. Cellular divisionsalso were evident from the loosely adherent cell populationand produced clusters of small, round, phase-bright cells (thatwe termed CSs) after 10 to 12 hours (Figure 1a). Within 24 to36 hours of their appearance, CSs increased in size and someof them detached from the culture surface; after 48 to 72hours, most CSs were between 20 and 150 �m in size, and,when not subjected to mechanical dissociation, the largestcontained dark zones within the inner mass (Figure 1a).
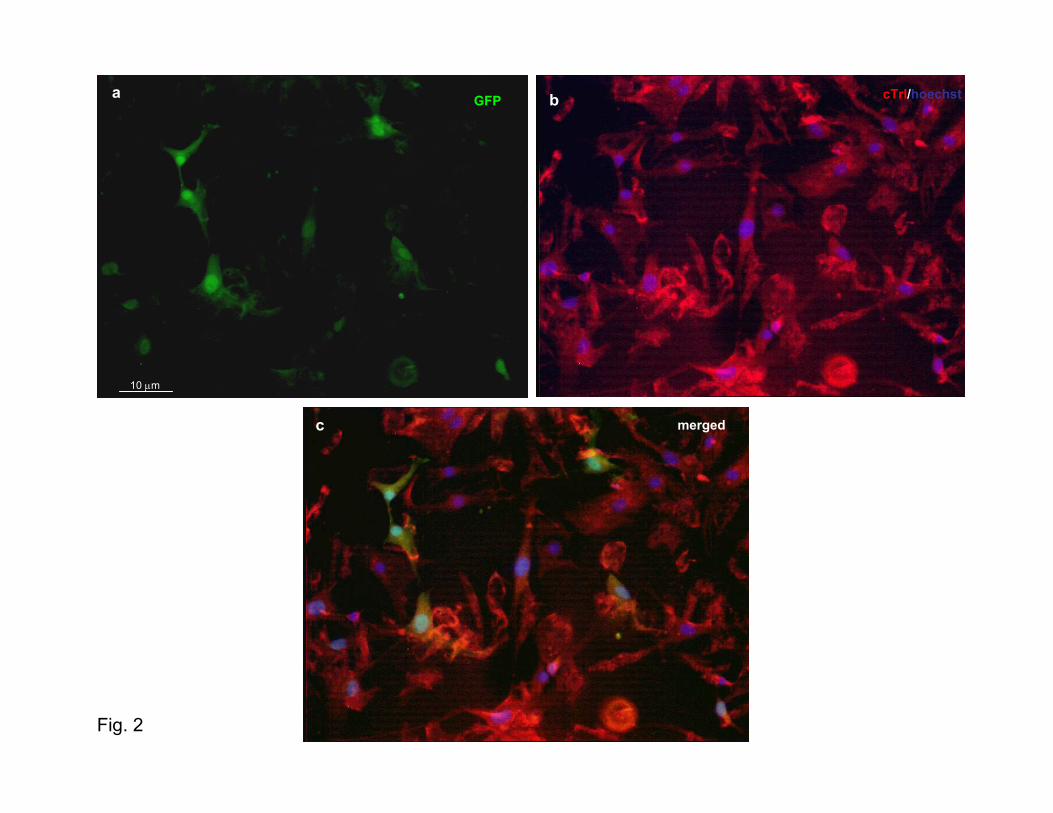
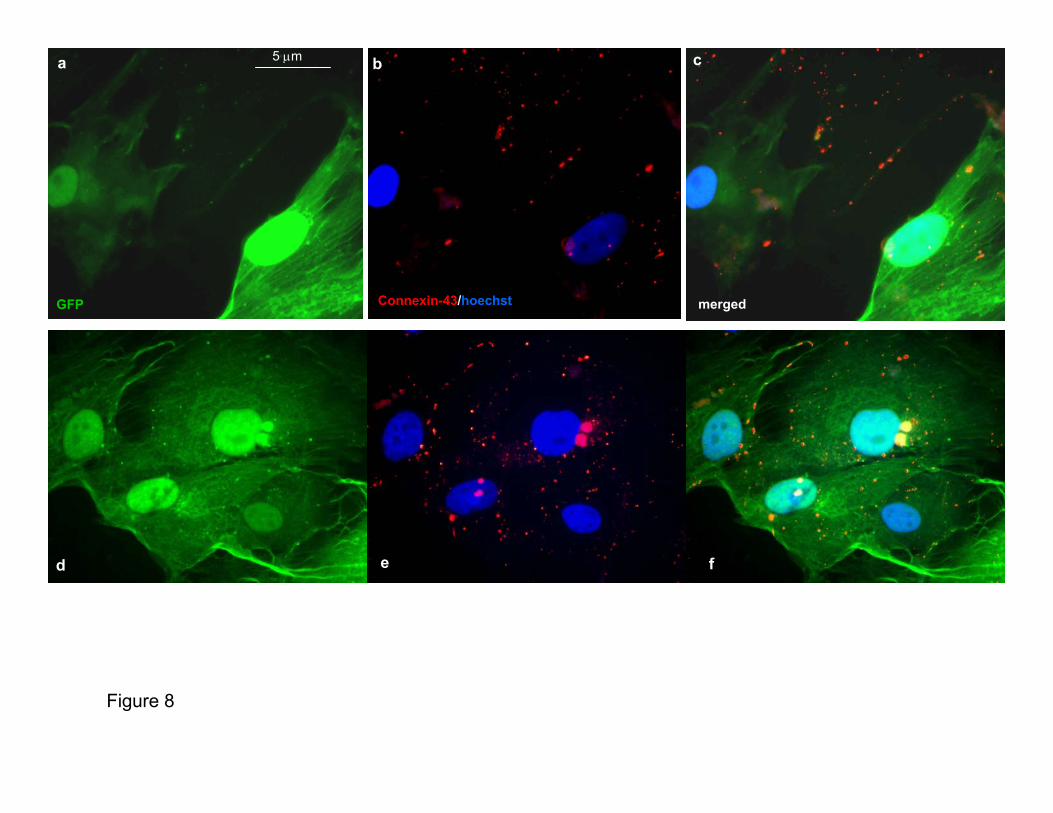
Murine CSs started beating spontaneously soon after theirgeneration (Supplementary Movie: mouse CSs movie 1a) andmaintained this function during their life span (Supplemen-tary Movie: mouse CSs movie 1b), whereas human CSs didso only when cocultured with rat cardiomyocytes (Supple-mentary Movie: human CSs movie 1a and 1b). To be sure thatcontraction was a new trait acquired by the CSs cells,GFP-labeled human CSs (partially or totally dissociated)were cocultured with cardiomyocytes prestained (Supple-mentary Human CSs Movie 2b through 2d) or not prestained(Supplementary Human CSs Movie 3a through 3d) with Dil.Contracting GFP-labeled cells were observed after 48 hoursof coculture; furthermore, Cx-43 immunostaining performedon the cocultures of human GFP-transduced CSs with unla-beled neonatal rat cardiomyocytes showed the typical punc-tuate fluorescence pattern of the main gap junction protein ofthe heart along the cytoplasmatic membrane of the humancells (Figure 2d and Supplementary Figure VIII), suggestingthat a functional connection is created between the twocellular populations.
ClonogenesisCSs were found to be composed of clonally derived cells anddid not simply represent cellular aggregates. In fact, whenhuman GFP-transduced CSs or murine CSs (derived fromeGFP/MLC3F or eGFP/cTrI mice) were dissociated andplated as single cells on mitomycin-treated STO fibroblast-coated 96-well plates (or clonally diluted on 10-cm Petridishes), fluorescent spheres were generated with a 1% to 10%efficiency (Figure 2a). These spheres could be subcloned onpoly-D-lysine-coated surfaces, showing the same functionaland phenotypic behavior in culture as the nonclone-derivedCSs. In fact, 3 days after their appearance, some of theMLC3F-nlacZ/B5-eGFP or cTnI-nlacZ/B5-eGFP miceclone–derived CSs started to beat (supplementary clonemovie), and, after 48 hours of culture with CEM, the majority(6 of 7) of these showed expression of the lac-Z transgenewithin the nuclei after specific histochemical staining (Figure2b1 and 2b2 and Supplementary Figure I). Moreover, humanclones derived from a single GFP-labeled cell started asynchronous beating and expressed cTnI after 48 hours ofcoculture with rat cardiomyocytes (Supplementary Moviehuman CSs 2a and 2a1 and Supplementary Figure II).
912 Circulation Research October 29, 2004
by guest on May 2, 2014http://circres.ahajournals.org/Downloaded from
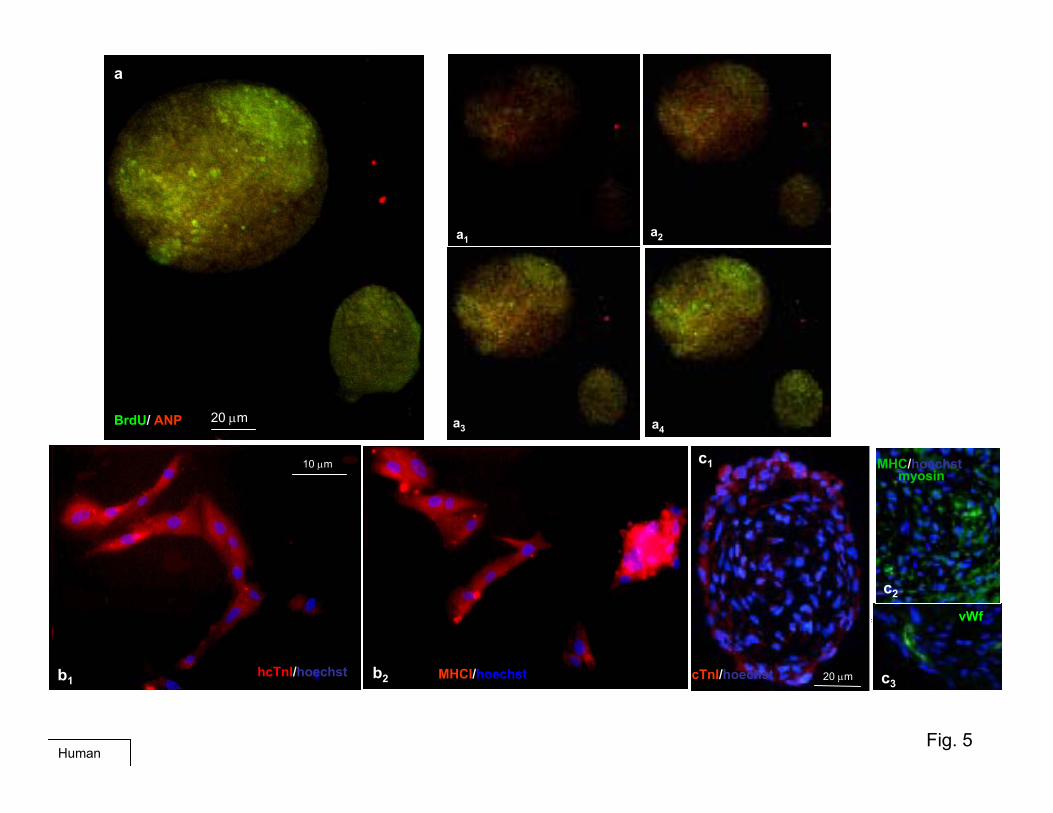
Furthermore, when BrdUrd was added to the culturemedium, virtually all cells in the small CSs and those of theinner part of the largest CSs were labeled (Figure 3a),indicating that these cells were newly generated (Supplemen-tary Figures III through Va).
ProliferationHuman CS-generating cells were capable of self-renewal.With periodical dissociation, together with partial substitutionof CGM every 2 to 3 days, a log-phase expansion of sphereswas obtained (Figure 1b). Mouse CS growth was slower(probably because of the more differentiated features as-sumed in culture, such as beating) and serum-dependent asfor the human CSs (Figure 1b).
As shown in Figure 3a and Supplementary Figure V,confocal immunofluorescence analysis of BrdUrd-labeled
human CSs with anti-BrdUrd (green) and cardiac-troponin I(cTnI) or atrial natriuretic peptide (ANP) (red) revealedBrdUrd-positive cells, particularly in the inner of the spheres,whereas cTnI-positive or ANP-positive cells were mainlylocalized in the external layers. Similar features are shown inSupplementary Figures III and IV. BrdUrd-labeled cells (red)mostly localized in the center of a CS and colocalize with theHoechst-labeled nuclei, whereas cardiac myosin heavy chain(MHC)-expressing cells (green) were preferentially located inthe boundary layers. Furthermore, several CS cells expressedcardiac differentiation markers (cTnI, ANP) while still divid-ing, as indicated by BrdUrd incorporation (Figure 3a andSupplementary Figure Va), suggesting that early cardiac differ-entiation already occurred during the proliferation phase of theirgrowth. Usually within 10 days, some spheres became adherent,
Figure 1. CS proliferation. a, Phase micrograph of floating CSs (cultured from �24 hours to �48 hours) derived from primary culture of ahuman atrial biopsy sample. b, Proliferation curves of human and mouse CSs (derived from 8 different subjects [left] and from prenatal andpostnatal hearts [middle and right], respectively) in the presence (middle) and absence (right) of 3.5% serum. Number of spheres refers to themean number per well from which 90% of the spheres were withdrawn at each time point for further analysis. Note the different pattern of prolifera-tion between the human and mouse CSs and the rapid rise of the curves, followed by an irreversible decline in the serum-free conditions.
Messina et al Mammalian Adult Cardiac Stem Cells 913
by guest on May 2, 2014http://circres.ahajournals.org/Downloaded from
showing a more flattened morphology. Some small cells even-tually migrated out from these “sun-like” spheres in the form ofadherent (differentiated) or small, round cells that could generatenew spheres. After thawing from cryopreservation, CSs prolif-erated again, maintaining their ability to beat (SupplementaryMovie: human CSs movie).
Immunophenotype Characterization andFluorescence-Activated Cell Sorting AnalysisPhenotypic analysis of newly developing human and mouseCSs revealed expression of endothelial (KDR (human)/flk-1[mouse], CD-31) and stem cell (CD-34, c-kit, sca-1) markers.As shown in Figure 3b, CSs at the 2- to 10-cell stage strongly
Figure 2. Clonogenesis and coculturefeatures. a, Fluorescence analysis of asingle cell (upper right) (obtained from adissociated GFP-expressing CS) whenplated by limiting dilution on mitomycin-treated STO fibroblast-coated 96-wellplates in CGM over the course of thegeneration of the GFP-labeled clone. Thisclone could be passaged and expandedon poly-D-lysine coat (lower left). b, X-Galstaining of a eGFP/MLC3F clone(obtained in the same way as werehuman clones) after 48 hours of exposureto growth factor-free medium. In theseconditions, clone cells become more flat-tened, with many nuclei appearing blue,demonstrating that a differentiation pro-cess occurred (see also SupplementaryFigure I and Supplementary clone mov-ies). c, Fluorescence analysis of partiallydissociated eGFP-labeled human CSs at96 hours of coculture with rat cardiomyo-cytes. The same green cells that showeda synchronous contraction with cardio-cytes (see supplementary human CSsmovies) express cTnI. d, Fluorescentanalysis of connexin-43 expression (red)in eGFP-labeled human CSs coculturedwith rat cardiomyocytes, as in (c). Apunctuate red fluorescence is present inthe cell membrane of human cells (seeSupplementary Figure VIII).
914 Circulation Research October 29, 2004
by guest on May 2, 2014http://circres.ahajournals.org/Downloaded from
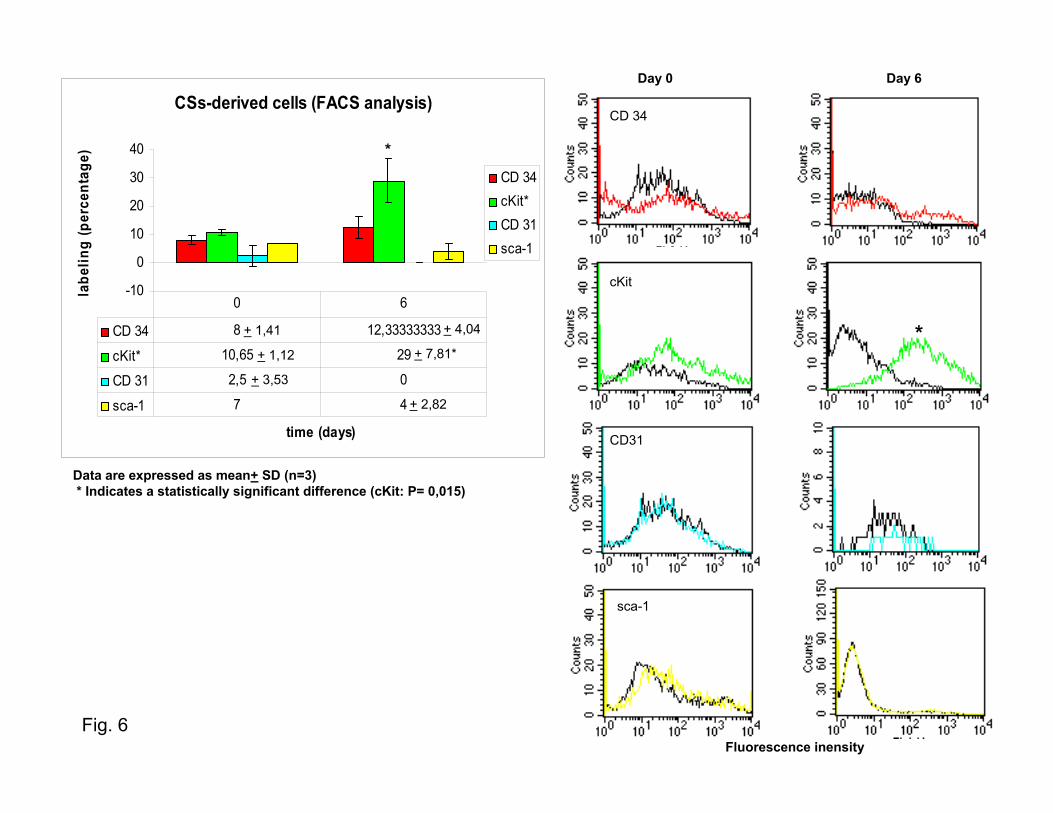
Figure 3. CSs BrdUrd incorporation and CSs characterization. a, Fluorescence confocal analysis of BrdUrd-labeled human CSs for car-diac differentiation markers: 6-�m scans (from the periphery to the center of the sphere) and final pictures (small and large images,respectively) of BrdUrd (green) and cTnI (red) (see Supplementary Figures III through V). b, Confocal analysis of human CSs after 12hours of culture: CD-34, CD-31, KDR, and c-Kit labeling of CS-generating cells at the beginning of sphere formation. c, fluorescence-activated cell sorting analysis of postnatal mouse CSs-derived cells. A time course at 0 and 6 days was used, and the phenotype pro-file for CD34, cKit, Cd31, and sca-1 expression was analyzed and shown as a percentage of positive events. Data are presented asmean�SD (n�3). *Statistically significant difference from 0 days. See the graphics in the Table and in Figure 6.
Messina et al Mammalian Adult Cardiac Stem Cells 915
by guest on May 2, 2014http://circres.ahajournals.org/Downloaded from
reacted with antibodies against these antigens. In largerspheres, the expression pattern of some of these markers(particularly cKit) was similar to that of the BrdUrd-labeling(positive staining in the center and in some peripheral zones,generating satellite spheres; data not shown).
A time course (0 and 6 days) of the quantitative charac-terization of CS cells with these stem and endothelial markerswas performed by fluorescence-activated cell sorting analysis(Figure 3c and Supplementary Figure VI). As shown at thebeginning of their formation (0 days), the phenotype of thesecells seems to reflect the epifluorescent microscopy analysiswith �10% of positive staining for all four phenotypes.However, at 6 days, cKit appears to be the only conservedmarker, suggesting that the cKit� cells could be the main onescontributing to the maintenance of proliferation. The initialcell-labeling may reflect an early activation state, as has beensuggested for CD-34 in several systems.17 Fluorescencemicroscopy analysis performed on cryosectioned human CSsrevealed expression of cardiac differentiation markers (cTnI,MHC) and endothelial markers (von Willebrand factor)(Supplementary Figure Vc1 through Vc3). When totally orpartially dissociated into single cells and cultured oncollagen-coated dishes in the same medium as the explants,mouse and human CS-derived cells assumed a typical car-diomyocyte morphology, phenotype (Supplementary FiguresVb1 through Vb2 and VIIc and VIId), and function docu-mented (in the mouse only) by spontaneous contraction(Supplementary Movie: mouse CSs movie 2a and 2b).
Human CSs did not beat spontaneously; however, thesebegan to beat within 24 hours when cocultured with postnatalrat cardiomyocytes, losing their spherical shape and assuminga “sun-like” appearance. Markers of cardiac differentiationwere coexpressed within GFP in labeled human CSs cells(Figure 2c).
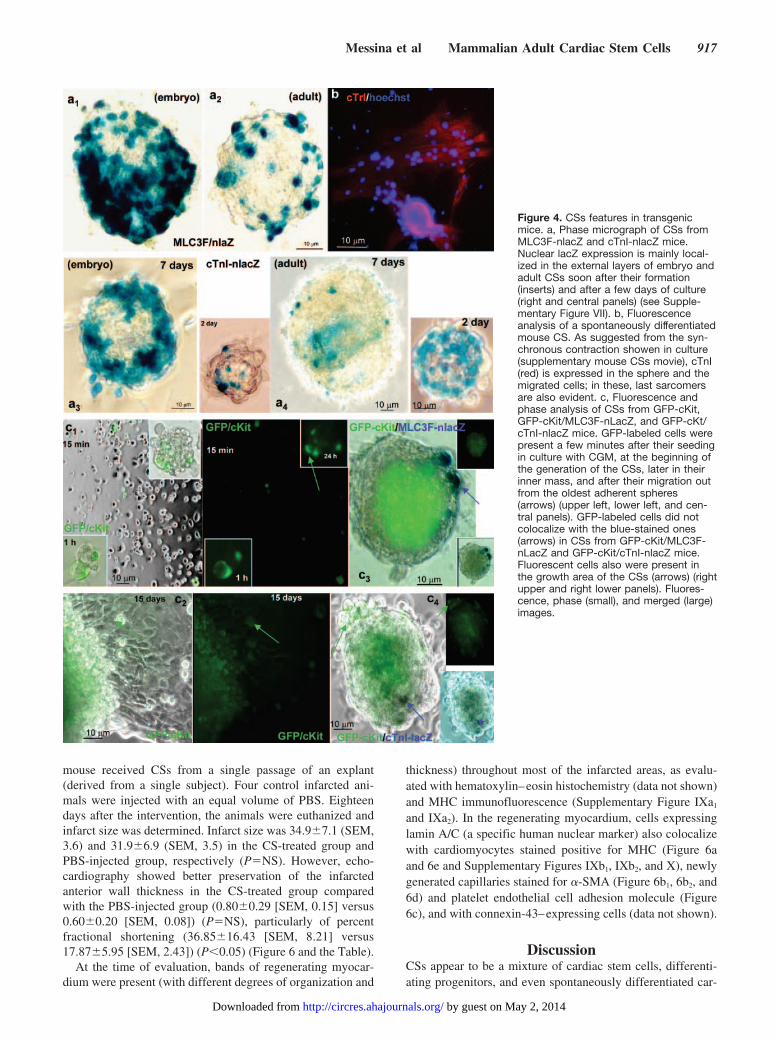
Transgenic MiceTo follow the differentiation process of CSs during theprenatal and postnatal age, MLC3F-nlacZ and cTnI-nlacZmice were used.11–12 These mice express a form of lacZtransgene that localizes within the nucleus under the skeletaland cardiac muscle myosin light chain or cardiac troponin Ipromoter, respectively. CSs obtained from embryonic day 9to 12, fetal day 17 to 18, and from neonatal and adult miceshowed spontaneous expression of the reporter gene in variablepercentages (10% to 60%) of spheres in the different cultureconditions used (Figure 4a1 through 4a4 and SupplementaryFigure VIIa1, VIIa2, VIIb1, and VIIb2). Moreover, regarding thehuman ones, CS-generating cells from mice expressed stem(CD-34, sca-1, cKit) and endothelial cell markers (flk-1, CD-31)(data not shown).
On this basis, we used transgenic mice expressing GFPunder the control of the c-kit promoter14 to further clarify thecellular origin of these spheres and to follow the pattern oftheir growth process. As shown in Figure 4c1, GFP-positivecells were present from the beginning of the formation of theCSs and, albeit with reduced fluorescence intensity, also laterwithin the mass of cells of the CSs and in cells migrating fromold adherent “sun-like” CSs (Figure 4c2). Moreover, assuggested by the growth pattern of human CSs, when satellite
secondary CSs appeared to detach from the primary ones,GFP-positive cells localized on the margins of the latter andin the inner part of the former.
We studied this process in double-heterozygous miceobtained from GFP-cKit/MLC3F-nlacZ or GFP-cKit/cTnI-nLacZ crossings. As shown in Figure 4c3 and 4c4, �-Galpositivity did not colocalize with GFP in cells present withinthe growing areas.
In Vivo Survival and Morpho-functional Potentialof the CardiospheresTo investigate the survival and morpho-functional potentialof the CSs in vivo, two sets of experiments were performed.In the first, CS cells were injected in the dorsal subcutaneousregion of SCID mice. In the second, they were injected intothe hearts of SCID beige mice, acutely after myocardialinfarction. The objective of ectopic transplantation experi-ments was to study the pattern and the behavior of growth ofCSs in a neutral milieu (ie, without specific cardiac induction)to verify their unique potential of generation of the maincardiac cell types and to exclude the potential of neoplastictransformation. For these experiments, �60 pooled spheres/inoculum/mouse from prenatal and postnatal MLC3F-nlacZ/B5-eGFP mice, TnI-nlacZ/B5-eGFP mice, MLC3F-nlacZ/CD-1 mice, and cTnI-nlacZ/CD-1 mice were used. Duringthe first 10 days, beating was appreciable through the skinover the injection site, distant from large blood vessels. Onday 17, animals were euthanized and the inoculum recog-nized as a translucent formation, grain-like in size, wrappedin ramified vessel-like structures. Observation of unfixedcryosections by fluorescence microscopy (Figure 5a1 through5a4) revealed the presence of open spheres from which cellsappeared to have migrated. Clusters of “black holes,” partic-ularly in the periphery of the structure, were evident. Thetissue contained tubular formations, surrounded by nuclei(Hoechst-positive), identified as cardiac sarcomeres by cTnIand sarcomeric myosin immunostaining (Figure 5b3 through5b6). �-Smooth muscle actin (�-SMA)-positive structures(known to be transiently expressed during cardiomyogen-esis)2,18 were present in the remainder of the spheres andassociated with the vasculature (the clusters of “black holes”)(Figure 5a3 through 5a5). This exhibited well-differentiatedstructures with a thin endothelium expressing vascularendothelial–cadherin (Figure 5b1) and a relative large lumencontaining erythrocytes (Figure 5a3), indicating the establish-ment of successful perfusion by the host. Light microscopicobservation of the inoculum, after X-gal staining, showedstrong nuclear expression of striated muscle-specific lacZ inthe remainder of the spheres and in some cells close to them(Figure 5b2). No multidifferentiated structures suggesting thepresence of tumor formation were observed.
To test the acquisition of functional competence and thecardiac regenerative potential of the CSs when challengedinto an infarcted myocardium, orthotopic transplantationexperiments with human CSs were performed. To performthese, thawed (cryopreserved) adult human CSs from threeatrial (one male and two female) and one ventricular (onefemale) biopsy specimens were injected into the viablemyocardium bordering a freshly produced infarct. Each
916 Circulation Research October 29, 2004
by guest on May 2, 2014http://circres.ahajournals.org/Downloaded from
mouse received CSs from a single passage of an explant(derived from a single subject). Four control infarcted ani-mals were injected with an equal volume of PBS. Eighteendays after the intervention, the animals were euthanized andinfarct size was determined. Infarct size was 34.9�7.1 (SEM,3.6) and 31.9�6.9 (SEM, 3.5) in the CS-treated group andPBS-injected group, respectively (P�NS). However, echo-cardiography showed better preservation of the infarctedanterior wall thickness in the CS-treated group comparedwith the PBS-injected group (0.80�0.29 [SEM, 0.15] versus0.60�0.20 [SEM, 0.08]) (P�NS), particularly of percentfractional shortening (36.85�16.43 [SEM, 8.21] versus17.87�5.95 [SEM, 2.43]) (P�0.05) (Figure 6 and the Table).
At the time of evaluation, bands of regenerating myocar-dium were present (with different degrees of organization and
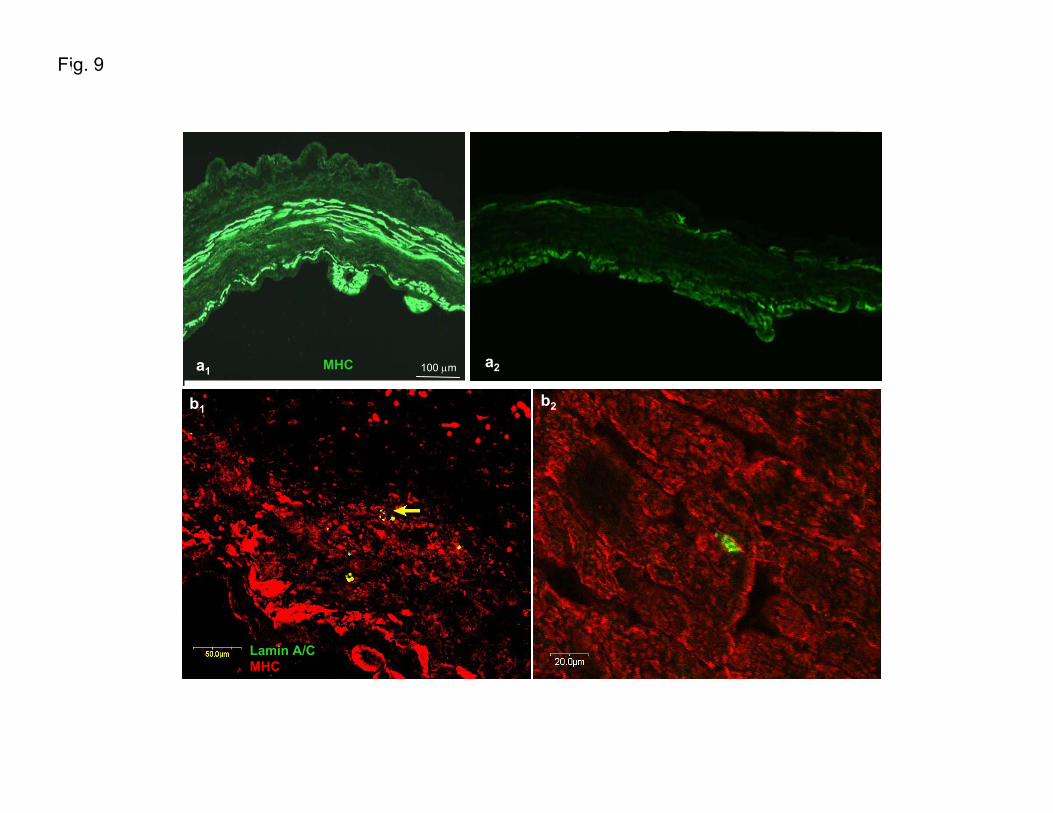
thickness) throughout most of the infarcted areas, as evalu-ated with hematoxylin–eosin histochemistry (data not shown)and MHC immunofluorescence (Supplementary Figure IXa1
and IXa2). In the regenerating myocardium, cells expressinglamin A/C (a specific human nuclear marker) also colocalizewith cardiomyocytes stained positive for MHC (Figure 6aand 6e and Supplementary Figures IXb1, IXb2, and X), newlygenerated capillaries stained for �-SMA (Figure 6b1, 6b2, and6d) and platelet endothelial cell adhesion molecule (Figure6c), and with connexin-43–expressing cells (data not shown).
DiscussionCSs appear to be a mixture of cardiac stem cells, differenti-ating progenitors, and even spontaneously differentiated car-
Figure 4. CSs features in transgenicmice. a, Phase micrograph of CSs fromMLC3F-nlacZ and cTnI-nlacZ mice.Nuclear lacZ expression is mainly local-ized in the external layers of embryo andadult CSs soon after their formation(inserts) and after a few days of culture(right and central panels) (see Supple-mentary Figure VII). b, Fluorescenceanalysis of a spontaneously differentiatedmouse CS. As suggested from the syn-chronous contraction showen in culture(supplementary mouse CSs movie), cTnI(red) is expressed in the sphere and themigrated cells; in these, last sarcomersare also evident. c, Fluorescence andphase analysis of CSs from GFP-cKit,GFP-cKit/MLC3F-nLacZ, and GFP-cKt/cTnI-nlacZ mice. GFP-labeled cells werepresent a few minutes after their seedingin culture with CGM, at the beginning ofthe generation of the CSs, later in theirinner mass, and after their migration outfrom the oldest adherent spheres(arrows) (upper left, lower left, and cen-tral panels). GFP-labeled cells did notcolocalize with the blue-stained ones(arrows) in CSs from GFP-cKit/MLC3F-nLacZ and GFP-cKit/cTnI-nlacZ mice.Fluorescent cells also were present inthe growth area of the CSs (arrows) (rightupper and right lower panels). Fluores-cence, phase (small), and merged (large)images.
Messina et al Mammalian Adult Cardiac Stem Cells 917
by guest on May 2, 2014http://circres.ahajournals.org/Downloaded from
Figure 5. In vivo analysis (ectopic CSs inoculum). a1 to a5, Ectopic transplantation of CSs from MLC3F-nlacZ/B5-eGFP mouse to SCIDmouse (upper left panels). Fluorescence analysis of unfixed cryosections (a1, a2, and a4) from the subcutaneous dorsal inoculum (day17). GFP cells seemed to have migrated from the spheres, whereas clusters of vessel-like structures (a2) could be observed mainly inthe external area. Staining for SMA of one of these cryosections showed positive immunoreaction of the sphere and some cells withinthe inoculum (a5). b-1 to b6, Fluorescence (b3 to b4) and phase analysis (b5 to b6) of fixed and immunostained cryosections from dorsalinoculum of CSs from MLC3F-nlacZ/CD-1 and cTnI-lacZ/CD-1 mice. Tubular structures were stained for sarcomeric myosin (b3 to b5)and cTnI (b4 through b6). X-Gal staining labeled the cells within and those migrating from CS (b2). Endothelial markers (SMA and vascu-lar endothelial–cadherin) stained the vasculature (“black holes”) (a3 and b1).
918 Circulation Research October 29, 2004
by guest on May 2, 2014http://circres.ahajournals.org/Downloaded from
Figure 6. In vivo analysis (orthotopic transplantation of humanCSs). Orthotopic transplantation performed in a SCID-beigemouse. Cryopreserved human CSs were transplanted into theviable myocardium bordering a freshly produced infarct. Con-focal analysis of cryosectioned left ventricular heart 18 daysafter the coronary ligature shows that (a) cardiomyocytesexpressing MHC (red) in the regenerating myocardium (partic-ularly those indicated by the two central arrows) also stainpositive for lamin A/C (green) (a specific human nuclearmarker). In these cells, MHC expression is evident mainly inthe perinuclear area (see Supplementary Figure X). Lamin A/C-labeled cells (red) are present in newly generated capillariesstaining for �-SMA (b1 through d), and platelet endothelial celladhesion molecule (c). d, Confocal analysis of colocalization oflamin A/C-labeled cells (red) with the newly generated capillar-ies staining for �-smooth muscle actin. e, Low-magnificationimage shows viable lamin A/C-expressing cells (green) inregenerating myocardium expressing MHC (red).
Messina et al Mammalian Adult Cardiac Stem Cells 919
by guest on May 2, 2014http://circres.ahajournals.org/Downloaded from
diomyocytes. Vascular cells were also present, depending onthe size of the sphere and time in culture. It is possible that,as for neurospheres,19 differentiating/differentiated cells stopdividing and/or die, whereas stem cells continue to proliferatein an apparently asymmetric way, giving rise to manysecondary spheres and to exponential growth in vitro. Me-chanical dissociation favors this process. Death, differentia-tion, and responsiveness to growth factors of the differentcells within the CSs could depend on the three-dimensionalarchitecture and on localization within the CSs.20 The spon-taneous formation of spheres is a known prerogative of neuralstem cells, some tumor cell lines (LIM),21 endothelial cells,22
and fetal chicken cardiomyocytes.23 All these models (oursincluded) that mimic the true three-dimensional architectureof tissues consist of spheroids of aggregated cells thatdevelop a two-compartment system composed of a surfacelayer of differentiated cells and a core of unorganized cellsthat first proliferate and then disappear over time (perhapsthrough apoptotic cell death). As well-documented in fetalchick cardiomyocytes and endothelial cell spheroid culture,three-dimensional structure affects the sensitivity of cells tosurvival and growth factors.21,22 In particular, central spheroidcells do not differentiate and are dependent on survivalfactors to prevent apoptosis, whereas the cells of the surfacelayer seem to differentiate beyond the degree that can beobtained in two-dimensional culture and become independentof the activity of survival factors.23 Furthermore, cell–cellcontact and membrane-associated factors, known to be im-portant for the division of neural precursor cells,24 could beinvolved in our system. This is in accordance with the notionthat stem cells (or cells with stem cell function) will onlyretain their pluripotency within an appropriate environment,as suggested by the “niche” hypothesis.25
Thus CSs can be considered clones of adult stem cells,maintaining their functional properties in vitro and in vivoafter cryopreservation.
While the experiments performed for this article wereongoing, two articles were published concerning the isolationof cardiac stem cells or progenitor cells from adult mamma-lian hearts.26,27 Isolation of these cells was based exclusivelyon the expression of a stem cell-related surface antigen: c-kitin the first article and Sca-1 in the second one. In the firststudy,26 freshly isolated c-kit� Lin� cells from rat hearts werefound to be self-renewing, clonogenic, and multi-potent,exhibiting biochemical differentiation into the myogenic cell,smooth muscle cell, or endothelial cell lineage but failing tocontract spontaneously. When injected into an ischemic heart,these cells regenerated functional myocardium. In the secondstudy,27 Sca-1� cKit� cells from mice hearts were induced invitro to differentiate toward the cardiac myogenic lineage inresponse to 5-azacytidine. When given intravenously afterischemia/reperfusion, these cells targeted injured myocardi-um and differentiated into cardiomyocytes, with and withoutfusion with the host cells. Our data obtained on GFP-cKittransgenic mice also suggest that the adult cardiac stem cell iscKit�. It is possible that CSs enclose a mixed population ofcells that, as in the niche, could promote the viability of cKitprogenitors and contribute to their proliferation. The dataobtained in the present article confirm the existence of adultcardiac stem cells/progenitor cells. More importantly, theydemonstrate for the first time to our knowledge that it ispossible to isolate cells from very small fragments of humanmyocardium and expand these cells in vitro many-fold(reaching numbers that would be appropriate for in vivotransplantation in patients) without losing their differentiationpotential. Previously unforeseen opportunities for myocardialrepair could now be identified.
AcknowledgmentsThis study was supported by Italian Ministery of the University andof the Scientific and Technological Research (MURST) and byspecific funds of the University of Rome “La Sapienza” from stem
Myocardial Repair (Echocardiography)
CAL�CSs(N�4)
CAL(N�6)
Sham(N�3)
LVIDd, mm 4.10 3.57 2.13*†
SD�0.85 SD�0.33 SD�0.06
SEM�0.42 SEM�0.13 SEM�0.03
AWThd, mm 0.80 0.60 1.03†
SD�0.29 SD�0.20 SD�0.01
SEM�0.15 SEM�0.08 SEM�0.01
FS, % 36.85 17.87* 59.13*†
SD�16.43 SD�5.95 SD�4.56
SEM�8.21 SEM�2.43 SEM�2.63
Regional wall motion abnormality, % 33.82 30.88
SD�10.05 SD�14.71
SEM�6.53 SEM�7.35
*P�0.05 vs CAL�CSs.†P�0.05 vs CAL.Effect of human CSs orthotopic transplantation on echocardiographic index of myocardial
performance. Data are presented as mean�SD.LVIDd indicates left ventricular internal dimension at end diastole; AWThd, anterior wall thickness;
FS, fractional shortening.
920 Circulation Research October 29, 2004
by guest on May 2, 2014http://circres.ahajournals.org/Downloaded from
cell research. We thank Gianluigi Condorelli for continuous encour-agement and valuable discussion. We thank Roberto Latini and LidiaStaszewsky in particular for their precious contribution in conductingand analyzing in vivo experiments. We thank Sergio Ottolenghi(Dipartimento Biotecnologie e Bioscienze, Universita Milano-Bicocca) for kindly providing transgenic mice. We are also gratefulto Francesco Musumeci and Fabio Miraldi for kindly supplying someof the heart biopsy specimens. Marta Alessandrini, Dario Sirabella,Lucia Ricci Vitiani, and Ilaria Falciatori (PhD students), Universityla Sapienza of Rome, Italy, are also gratefully acknowledged fortheir availability during the course of this work.
References1. Soonpaa MH, Field LJ. Survey of studies examining mammalian cardio-
myocyte DNA synthesis. Circ Res. 1998;83:15–26 .2. Nadal-Ginard B, Kajstura J, Leri A, Anversa P. Myocyte death, growth,
and regeneration in cardiac hypertrophy and failure. Circ Res. 2003;92:139–150.
3. Condorelli G, Borello U, De Angelis L, Latronico M, Sirabella D, ColettaM, Galli R, Balconi G, Follenzi A, Frati G, Cusella De Angelis MG,Gioglio L, Amuchastegui S, Adorini L, Naldini L, Vescovi A, Dejana E,Cossu G. Cardiomyocytes induce endothelial cells to trans-differentiateinto cardiac muscle: implications for myocardium regeneration. Proc NatlAcad Sci U S A. 2001;98:10733–10738.
4. Laflamme MA, Myerson D, Saffitz JE, Murry CE. Evidence for cardio-myocyte repopulation by extracardiac progenitors in transplanted humanhearts. Circ Res. 2002;90:634–640.
5. Glaser R, Lu MM, Narula N, Epstein JA. Smooth muscle cells, but notmyocytes, of host origin in transplanted human hearts. Circulation. 2002;106:17–19.
6. Hassink RJ, Dowell JD, Brutel de la Riviere A, Doevendans PA, Field LJ.Stem cell therapy for ischemic heart disease. Trends Mol Med. 2003;9:436–441.
7. Menasche P, Hagege AA, Vilquin JT, Desnos M, Abergel E, Pouzet B,Bel A, Sarateanu S, Scorsin M, Schwartz K, Bruneval P, Benbunan M,Marolleau JP, Duboc D. Autologous skeletal myoblast transplantation forsevere postinfarction left ventricular dysfunction. J Am Coll Cardiol.2003;41:1078–1083.
9. Strauer BE, Brehm M, Zeus T, Kostering M, Hernandez A, Sorg RV,Kogler G, Wernet P. Repair of infarcted myocardium by autologousintracoronary mononuclear bone marrow cell transplantation in humans.Circulation. 2002;106:1913–1918.
10. Assmus B, Schachinger V, Teupe C, Britten M, Lehmann R, Dobert N,Grunwald F, Aicher A, Urbich C, Martin H, Hoelzer D, Dimmeler S,Zeiher AM. Transplantation of Progenitor Cells and RegenerationEnhancement in Acute Myocardial Infarction. Transplantation of Pro-genitor Cells and Regeneration Enhancement in Acute MyocardialInfarction (TOPCARE-AMI). Circulation. 2002;106:3009–3017.
11. Kelly R, Alonso S, Tajbakhsh S, Cossu G, Buckingham M. Myosin lightchain 3F regulatory sequences confer regionalized cardiac and skeletalmuscle expression in transgenic mice. J Cell Biol. 1995;129:383–396.
12. Di Lisi R, Millino C, Calabria E, Altruda F, Schiaffino S, Ausoni S.Combinatorial cis-acting elements control tissue-specific activation of thecardiac troponin I gene in vitro and in vivo. J Biol Chem. 1998;273:25371–25380.
13. Hadjantonakis AK, Gertsenstein M, Ikawa M, Okabe M, Nagy A. Gen-erating green fluorescent mice by germline transmission of green fluo-rescent ES cells. Mech Dev. 1998;76:79–90.
14. Cairns LA, Moroni E, Levantini E, Giorgetti A, Klinger FG, Ronzoni S,Tatangelo L, Tiveron C, De Felici M, Dolci S, Magli MC, Giglioni B,Ottolenghi S. Kit regulatory elements required for expression indeveloping hematopoietic and germ cell lineages. Blood. 2003;102:3954–3962.
15. Pennica D, King KL, Shaw KJ, Luis E, Rullamas J, Luoh SM, DarbonneWC, Knutzon DS, Yen R, Chien KR, et al. Expression cloning ofcardiotrophin 1, a cytokine that induces cardiac myocyte hypertrophy.Proc Natl Acad Sci U S A. 1995;92:1142–1146.
16. Sabri A, Guo J, Elouardighi H, Darrow AL, Andrade-Gordon P, SteinbergSF. Mechanisms of protease-activated receptor-4 actions in cardiomyo-cytes. Role of Src tyrosine kinase. J Biol Chem. 2003;278:11714–11720.
17. Zammit PS, Beauchamp JR. The skeletal muscle satellite cell.: stem cellor son o stem cell? Differentiation. 2001;68:193–204.
18. Kruithof BP, Van Den Hoff MJ, Tesink-Taekema S, Moorman AF.Recruitment of intra- and extracardiac cells into the myocardial lineageduring mouse development. Anat Rec. 2003;271A: 303–314.
20. Layer PG, Robitzki A, Rothermel A, Willbold E. Of layers and spheres:the reaggregate approach in tissue engineering. Trends Neurosci. 2002;25:131–134.
22. Korff T, Augustin HG. Integration of endothelial cells in multicellularspheroids prevents apoptosis and induces differentiation. J Cell Biol.1998;143:1341–1352.
23. Armstrong MT, Lee DY, Armstrong PB. Regulation of proliferation ofthe fetal myocardium. Dev Dyn. 2000;219:226–236.
24. Svendsen CN, ter Borg MG, Armstrong RJ, Rosser AE, Chandran S,Ostenfeld T, Caldwell MA. A new method for the rapid and long termgrowth of human neural precursor cells. J Neurosci Methods. 1998;85:141–152.
25. Nilsson SK, Simmons PJ. Transplantable stem cells: home to specificniches. Curr Opin Hematol. 2004;11:102–106.
26. Beltrami AP, Barlucchi L, Torella D, Baker M, Limana F, Chimenti S,Kasahara H, Rota M, Musso E, Urbanek K, Leri A, Kajstura J, Nadal-Ginard B, Anversa P. Adult cardiac stem cells are multipotent and supportmyocardial regeneration. Cell. 2003;114:763–776.
27. Oh H, Bradfute SB, Gallardo TD, Nakamura T, Gaussin V, Mishina Y,Pocius J, Michael LH, Behringer RR, Garry DJ, Entman ML, SchneiderMD. Cardiac progenitor cells from adult myocardium: homing, differen-tiation, and fusion after infarction. Proc Natl Acad Sci U S A. 2003;100:12313–12318.
Messina et al Mammalian Adult Cardiac Stem Cells 921
by guest on May 2, 2014http://circres.ahajournals.org/Downloaded from