BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofit publishers, academic institutions, research libraries, and research funders in the common goal of maximizing access to critical research. Classification, natural history, and evolution of Neorthopleurinae Opitz (Coleoptera, Cleridae). Part III. The Genera Agaphalera Opitz, Allochotes Westwood, Kataspinula Opitz, Lebasiella Spinola, Loedelia Lucas, Patuleius Fairmaire, Rifkindius Opitz, and Romanaeclerus Winkler Author(s): Weston Opitz Source: Pan-Pacific Entomologist, 89(4):244-258. 2013. Published By: Pacific Coast Entomological Society DOI: http://dx.doi.org/10.3956/2013-20.1 URL: http://www.bioone.org/doi/full/10.3956/2013-20.1 BioOne (www.bioone.org ) is a nonprofit, online aggregation of core research in the biological, ecological, and environmental sciences. BioOne provides a sustainable online platform for over 170 journals and books published by nonprofit societies, associations, museums, institutions, and presses. Your use of this PDF, the BioOne Web site, and all posted and associated content indicates your acceptance of BioOne’s Terms of Use, available at www.bioone.org/page/ terms_of_use . Usage of BioOne content is strictly limited to personal, educational, and non-commercial use. Commercial inquiries or rights and permissions requests should be directed to the individual publisher as copyright holder.
Transcript
BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofitpublishers, academic institutions, research libraries, and research funders in the common goal of maximizing access tocritical research.
Classification, natural history, and evolution ofNeorthopleurinae Opitz (Coleoptera, Cleridae). Part III. TheGenera Agaphalera Opitz, Allochotes Westwood, KataspinulaOpitz, Lebasiella Spinola, Loedelia Lucas, Patuleius Fairmaire,Rifkindius Opitz, and Romanaeclerus WinklerAuthor(s): Weston OpitzSource: Pan-Pacific Entomologist, 89(4):244-258. 2013.Published By: Pacific Coast Entomological SocietyDOI: http://dx.doi.org/10.3956/2013-20.1URL: http://www.bioone.org/doi/full/10.3956/2013-20.1
BioOne (www.bioone.org) is a nonprofit, online aggregation of core research in thebiological, ecological, and environmental sciences. BioOne provides a sustainable onlineplatform for over 170 journals and books published by nonprofit societies, associations,museums, institutions, and presses.
Your use of this PDF, the BioOne Web site, and all posted and associated contentindicates your acceptance of BioOne’s Terms of Use, available at www.bioone.org/page/terms_of_use.
Usage of BioOne content is strictly limited to personal, educational, and non-commercialuse. Commercial inquiries or rights and permissions requests should be directed to theindividual publisher as copyright holder.
One intent of this publication is to elucidate the phylogenetic relationships among
the eight genera listed in the title. The phylogeny of the other 14 genera of
Neorthopleurinae is presented in part II of this series of publications (Opitz in
preparation) whose purpose is to bring to light Neorthopleurinae systematics. The
genera included in this work were recently described and copiously illustrated
elsewhere (Opitz 2009b). In this treatise I make known generic level phylogenetics,
increase the species inventory of Neorthopleurinae by three, and present for the firsttime keys of the known species of Agaphalera and Loedelia. A generic description of
Agaphalera and Loedelia is provided to assure a continuance of understanding of
those generic-level characteristics relevant to the new species descriptions. Rifkindius
and Kataspinula remain monotypic. It needs to be clarified that Opitz (2009b: 185,
Character 15 Intraspicular plate: (0) present; (1) absent
Character 16 Dorsal plate of phallobase: (0) not elongated; (1) elongated
Character 17 Funicular antennomere 5: (0) not larger than six; (1) larger than six
Character 18 Tegmen: (0) lobed distally; (1) not lobed distallyCharacter 19 Antennomeres 9 and 10: (0) not acuminate anterodistally; (1) acuminate
anterodistally
Character 20 Basal denticle: (0) small; (1) large
Character 21 Unguis: (0) not bifid; (1) bifid
246 THE PAN-PACIFIC ENTOMOLOGIST Vol. 89(4)
2010: 50) erroneously noted that the tarsal pulvillar formula for Romanaeclerus asbeing 0-0-0, when in fact it is 2-2-2 (Opitz 2009a). Taxonomically, what remains to be
done with this monophyletic branch of Neorthopleurinae is to comprehensively
revise the Asiatic genus Allochotes and the Madagascan genus Patuleius.
Agaphalera Opitz (Opitz 2009b:142)
(Figs. 2, 3, 7–10, 13–15, 19, 21, 22)
Type Species. Lebasiella janthina LeConte 1866:99.
Diagnosis. Specimens of Agaphalera resemble superficially those of Loedelia. But,
in Agaphalera specimens the denticle of the unguis is large and somewhat truncate
(Fig. 19) not acuminate as in specimens of Loedelia (Fig. 20).
Figure 1. Habitus of Loedelia maculicollis (LeConte).
2013 OPITZ: TAXONOMY OF NEORTHOPLEURINAE (COLEOPTERA: CLERIDAE) 247
Description. Size: Length 3.2–5.5 mm; width 1.4–2.4 mm. Form (Figs. 21, 22):
Oblong short, robust, deep bodied, about 2 times longer than broad. Vestiture: Disc
of cranium and pronotum densely vested with stout setae, elytra vested only with 1usetae. Head (Fig. 13): Cranium subspheroid, frons very wide, indented with shallow
setiferous punctations, latter not widely separated; gula (Fig. 15) very small, process
Diagnosis. Specimens of this species resemble superficially the members of
Agaphalera cymatilis, sp. nov., but in specimens of A. corallina the epipleural margins
are subconvex, whereas in cymatilis specimens the epipleural margins are subparallel.
Description. Size: Length 5.0 mm; width 2.0 mm. Form (Fig. 21): Integumental
Color: Antennae and terminal palpomeres brown, forebody and legs yellow-brown,tarsi infuscated, elytra, pterothoracic venter, and abdomen dark blue. Integumental
Structure: Cranium and pronotum minutely punctated; elytral surface punctations
shallow, punctations aseriate; antenna capitate, capitulum short and lax, funicular
distally, lobes fimbriate; phallic apex digitiform; phallobasic rod large and cordate;
phallobasic apodeme broadly dilated distally.
Variation. The elytral punctations are slightly more pronounced in one specimen
from Acatlan.
Etymology. The trivial name corallinus (5 coral-red) is a Latin adjective. I refer to
the coloration of the forebody of this beetle.
Agaphalera cymatilis, sp. nov.
(Figs. 2, 9, 10, 22)
Holotype U. Mexico, Colima, Lago La Maria, 41009, 22 July 1995, beating Acacia,
J. Rifkind, A. Reifschneider, colls. (LACM).
Paratypes. Seven specimens. Mexico: Colima: Lago La Maria, 22-VII-1995, beatingAcacia, 1250 m, J. Rifkind, A. Reifschneider (WOPC, 1): Jalisco: 12 km S Autlan, 16-
VII-1990, E. Giesbert (WOPC, 1): Nayarit: 15 km N of Chapalilla, 24-VII-1993, beating
2013 OPITZ: TAXONOMY OF NEORTHOPLEURINAE (COLEOPTERA: CLERIDAE) 253
Loedelia Lucas 1920:380
(Figs. 1, 4–6, 11, 12, 16–18, 20, 23, 24)
Type species. Necrobioides mexicana Gahan 1910:76; Wolcott 1947:88; Corporaal
1950:312; Opitz 2002:280, 2009b:161.
Necrobioides Gahan 1910:76.
Diagnosis. Within Neorthopleurinae, only in members of Loedelia is the denticle of
the ungues acuminate (Fig. 20).
Description. Size: Length 5.0–6.0 mm; width 2.0–3.0 mm. Form (Fig. 1): Oblong
short, robust, deep body, about 2 times longer than broad. Vestiture: Disc of
cranium and pronotum densely vested with stout setae, elytra vested only with 1usetae. Head (Fig. 16): Cranium subspheroid, frons very wide, indented with very
small setiferous punctations, latter widely separated; gula (Fig. 18) very small,
sutures oblique, gular process short, narrow, and minutely forked; labrum short,
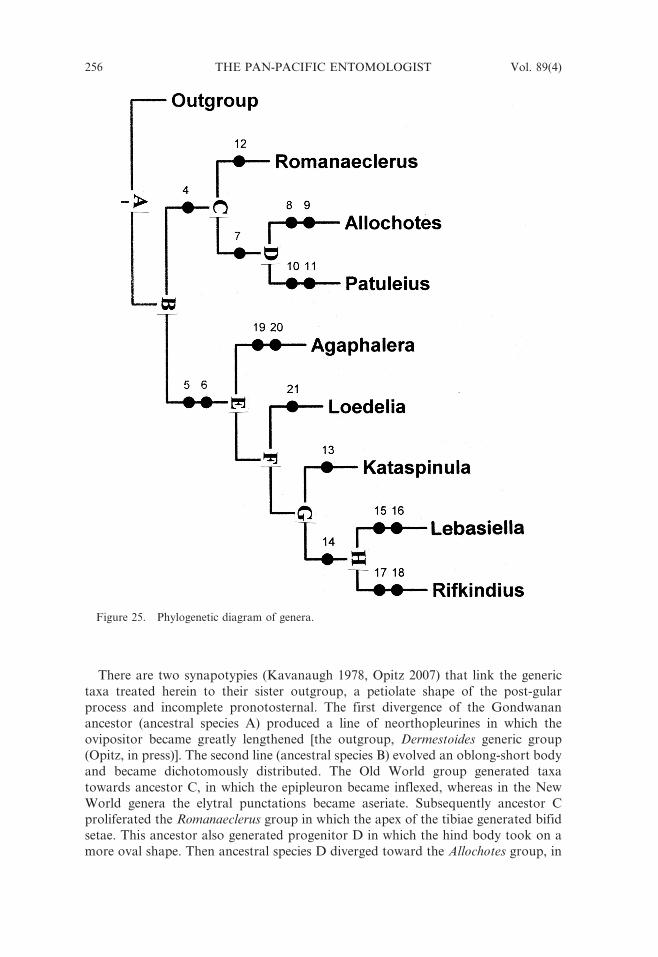
Perhaps the most outstanding information intrinsic in the phylogenetic hypothesis
of the taxa in this treatise (Fig. 25) is their clear separation into two geographical
components; the Old World genera and the New World genera. Such distributional
and evolutionary relationship evidence suggest that the taxa under consideration
have had an extensive evolutionary history that might have extend back to an era
when the continents were amassed into the great ancient Southern Hemisphere
land mass known as Gondwanaland. The evolution and diversification of these
neorthopleurines are probably linked to ancient angiosperm proliferations and to the
rise of ancient forests of gymnosperms. It is widely known that as a group the
Cleridae is mostly xylophilous (Balduf 1935) and that angiosperms flourished and
radiated throughout Gondwanaland (White 1988). Other discussions that link
ancestral Cleridae to a probable Gondwanan origin is found in Opitz (2003).
2013 OPITZ: TAXONOMY OF NEORTHOPLEURINAE (COLEOPTERA: CLERIDAE) 255
There are two synapotypies (Kavanaugh 1978, Opitz 2007) that link the generic
taxa treated herein to their sister outgroup, a petiolate shape of the post-gular
process and incomplete pronotosternal. The first divergence of the Gondwanan
ancestor (ancestral species A) produced a line of neorthopleurines in which the
ovipositor became greatly lengthened [the outgroup, Dermestoides generic group
(Opitz, in press)]. The second line (ancestral species B) evolved an oblong-short body
and became dichotomously distributed. The Old World group generated taxa
towards ancestor C, in which the epipleuron became inflexed, whereas in the New
World genera the elytral punctations became aseriate. Subsequently ancestor C
proliferated the Romanaeclerus group in which the apex of the tibiae generated bifid
setae. This ancestor also generated progenitor D in which the hind body took on a
more oval shape. Then ancestral species D diverged toward the Allochotes group, in
Figure 25. Phylogenetic diagram of genera.
256 THE PAN-PACIFIC ENTOMOLOGIST Vol. 89(4)
which the humerus became bulgy and the phallic plate spinous, and the Madagascan
Patuleius group, in which the epipleuron became extensively ridged.
The ancestral stock of the New World taxa (ancestral species E) proliferated the
Mexican Agaphalera group, characterized by a large basal denticle and the anterodistal
angle of antennomere 9 and 10 became subacuminate. Ancestor E also generated
ancestor F, which eventually evolved the progenitors of the Loedelia group, in which
the unguis became bifid, and the progenitors of ancestor G. The latter evolved theMexican Kataspinula group, in which the phallic membrane evolved two rows of
spines, and progenitor H. Ancestral species H generated the Lebasiella group. Among
these species, the intraspicular plate was lost and the dorsal plate of the phallobase
became elongated. The monotypic Rifkindius group forms the complementary stock, in
which the tegminal lobes were lost and funicular antennomere 5 enlarged.
ACKNOWLEDGMENTS
My gratitude to John Dorshorst for his review of the manuscript and to Gregory
Zolnerowich (Kansas State University) for providing SEM time.
LITERATURE CITED
Balduf, W. V. 1935. The Bionomics of Entomophagous Coleoptera. John S. Swift, New York, 220 pp.
Barr, W. F. 1950. Contributions towards a knowledge of the insect fauna of Lower California.No. 12. Coleoptera: Cleridae. Proceedings of the California Academy of Sciences, FourthSeries 24(12):485–519.
Corporaal, J. B. 1950. Cleridae. In: W. D. Hincks (Ed.). Coleopterorum Catalogus Supplementa,Pars 23 (editio secunda). W. Junk, Gravenhage, 373 pp.
Ekis, G. 1977. Classification, phylogeny, and zoogeography of the checkered beetle genus Perilypus(Coleoptera: Cleridae: Clerinae). Smithsonian Contributions to Zoology 227:1–38.
Fairmaire, L. 1902. Materiaux pour la faune coleopterique de la region malgache. Annales de laSociete Entomologique de France 71:530–568.
Gahan, C. J. 1910. Notes on the Cleridae and descriptions of some new genera and species of thisfamily of Coleoptera. Annals and Magazine Natural History 5(8):55–76.
Goloboff, P. A. 2003. NONA version 2.0. A tree searching program. Privately distributed by theauthor.
Hennig, W. 1966. Phylogenetic Systematics. University Press, Urbana, Illinois, 263 pp.Kavanaugh, D. H. 1978. Hennigian phylogenetics in contemporary systematic: principles, methods,
and uses, pp. 139–150. In: Biosystematics in Agriculture. Beltsville Symposia in AgriculturalResearch, 2, Allenheld, Osmun and Company, Halstead Press, New York, New York.
LeConte, J. L. 1866. New species of North American Coleoptera. Part I. Smithsonian MiscellaneousCollection 6(167):1–177.
LeConte, J. L. 1874. Descriptions of new Coleoptera chiefly from the Pacific slope of NorthAmerica. Transactions of the American Entomological Society 5:43–72.
LeConte, J. L. 1881. Descriptions of new species. In: J. L. LeConte & G. H. Horn, Description ofnew species of North American Coleoptera. Transactions of the Kansas Academy of Science7:74–77.
Lucas, R. 1920. Catalogus alphabeticus generum et subgenerum Coleopterorum orbis terrariumTotius (famil., trib., subtr., sect. incl.) Pars. 1. Archiv fuer Naturgeschite 84 (Abt. A):289–448.
Mayr, E. 1963. Animal Species and Evolution. Harvard University Press, Cambridge, 797 pp.
Nixon, K. C. 2002. Winclada version 1.00.08. Privatetly published by the author.Nixon, C. N. & J. A. Carpenter. 1993. On outgroups. Cladistics 9:413–426.
Opitz, W. 2002. Family 73. Cleridae Latrielle 1804, pp. 267–280. In: R. H. Arnett Jr, M. C. Thomas,P. E. Skelley & J. H. Frank (Eds.). American Beetles, Volume 2. Boca Raton. CRC Press,Gainesville, xiv + 861 pp.
Opitz, W. 2003. Spermatophores and spermatophore producing internal organs of Cleridae(Coleoptera: Clerinae): Their biological and phylogenetic implications. The ColeopteristsBulletin 57(2):167–190.
2013 OPITZ: TAXONOMY OF NEORTHOPLEURINAE (COLEOPTERA: CLERIDAE) 257
Opitz, W. 2007. Classification, natural history, and evolution of Epiphloeinae (Coleoptera,Cleridae). Part IV. The genera Pyticeroides Kuwert, 1894 and Diapromeces Opitz, 1997.Entomologica Basilensia et Collectionis Frey 29:77–166.
Opitz, W. 2009a. Revision of the African checkered beetle genus Romanaeclerus Winkler, 1960(Coleoptera: Cleridae: Korynetinae). Journal of Afrotropical Zoology 5:53–70.
Opitz, W. 2009b. Classification, natural history, and evolution of Neorthopleurinae subfa.nov.(Coleoptera, Cleridae) Part I. Generic composition of the subfamily and key to genera.Entomologica Basiliensia et Collectionis Frey 31:135–207.
Opitz, W. 2010. Classification, natural history, phylogeny, and subfamily composition of theCleridae and generic content of the subfamilies (Coleoptera: Cleroidea). EntomologicaBasiliensia et Collectionis Frey 32:31–128.
Opitz, W. (in press). Classification, natural history, and evolution of Neorthopleurinae Opitz(Coleoptera, Cleridae) Part II. The Dermestoides generic group. Giornale Italiano diEntomologia.
Pic, M. 1939. Mutations et nouveautes diverses. Melanges exotico-Entomologiques 71:1–36.Pic, M. 1940. Diagnoses de Coleopteres exotiques. L’Echange, Revue Linneenne 56(482):10–12.Standfuss, M. 1896. Handbuch der palaarktischen Gross-Schmetterlinge fur Forscher und Sammler.
Gustav Fischer, Jena, 392 pp.Tuomikosky, R. 1976. Notes on some principles of phylogenetic systematics. Annales Entomologici
Fennici 33(3):137–147.Westwood, J. O. 1875. Description of a new genus of clerideous Coleoptera from the Malayan
Archipelago. Transactions of the Entomological Society of London Part II, 1852:241–242.White, M. E. 1988. The Greening of Gondwana. Reed Books, New South Wales, 256 pp.Wolcott, A. B. 1947. Catalogue of North American beetles of the family Cleridae. Fieldiana: