Page 1
1
Cleavage of the Xylosyl Serine Linkage between a Core Peptide and a
Glycosaminoglycan Chain by Cellulases
Keiichi Takagaki, Mito Iwafune, Ikuko Kakizaki,
Keinosuke Ishido, Yoji Kato*, and Masahiko Endo
Department of Biochemistry, Hirosaki University School of Medicine,
5 Zaifu-cho, Hirosaki 036-8562 and
*Laboratory of Food Science, Faculty of Education, Hirosaki University,
1 Bunkyo-cho, Hirosaki 036-8560,
Japan
Running title: Release of Glycosaminoglycan Chains by Cellulases
Address for correspondence: Prof. Masahiko Endo, M.D.
Department of Biochemistry,
Hirosaki University School of Medicine,
5 Zaifu-cho, Hirosaki 036-8562,
Japan
Tel: +81-172-39-5015
Fax: +81-172-39-5016
Copyright 2002 by The American Society for Biochemistry and Molecular Biology, Inc.
JBC Papers in Press. Published on February 27, 2002 as Manuscript M111985200 by guest on June 20, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Page 2
2
SUMMARY
We previously found that endo-β-xylosidase from Patinopecten is an endo-type
glycosidase that cleaves the xylosyl serine linkage between a glycosaminoglycan chain and
its core protein (Takagaki, K., Kon, A., Kawasaki, H., Nakamura, T., Tamura, S., and Endo,
M. (1990) J. Biol. Chem. 265, 854-860). Screening for endo-β-xylosidase activity in several
cellulases detected this activity in the enzymes from Aspergillus niger, Penicillium
funiculosum, Trichoderma reesei, Trichoderma viride and Irpex lacteus. The cellulase derived
from Aspergillus niger was purified and its molecular weight was determined to be 26,000 by
SDS-PAGE. Examination of the specificity of the cellulase revealed that 1) the enzyme acts
on the linkage region (xylosyl serine) between a core peptide and a glycosaminoglycan chain;
2) enzymatic activity is greater with shorter glycosaminoglycan chains; 3) the enzyme readily
hydrolyzes the linkage in glycosaminoglycan-peptides, but intact proteoglycan is cleaved
only slowly; 4) the activity is unaffected by the glycosaminoglycan component (chondroitin
sulfate, dermatan sulfate and heparan sulfate). Judging from these enzymatic characteristics,
this cellulase is different from the endo-β-xylosidase of Patinopecten. We believe that this
cellulase will become a useful tool in the further development of glycotechnology, because,
like the endo-β-xylosidase of Patinopecten, it enables the release of intact
glycosaminoglycans from glycosaminoglycan-peptides.
by guest on June 20, 2018http://w
ww
.jbc.org/D
ownloaded from
Page 3
3
INTRODUCTION
Proteoglycans (PGs) are widely distributed in connective tissue and on the cell surface of
mammalian tissues, and are functional materials influencing cell growth, differentiation and
morphogenesis (1, 2). It is known that PGs consist of a core protein linked to
glycosaminoglycan (GAG) chains, and that the GAG chains interact with a number of growth
factors (3) and with important functional proteins such as antithrombin III (4-6) and heparin
cofactor II (7). Although GAG chains are broadly divided into chondroitin sulfate (ChS),
dermatan sulfate (DS), heparan sulfate (HS), heparin, hyaluronic acid and keratan sulfate by
their particular combination of repeating disaccharide units, ChS, DS, HS and heparin are all
attached to a serine residue of a core protein by the linkage region GlcAβ1-3Galβ1-3Galβ1-
4Xylβ1-O-Ser (8). Recently, it has been shown that the linkage region is modified by
phosphate and sulfate groups (9-11). Such diversity of linkage regions is notable, because it
suggests that the modification is a signal involved in the control of GAG biosynthesis (11).
The metabolism of PGs in vivo has not yet been clarified. Matsue and Endo (12)
demonstrated that urinary GAG chains bear in part glucuronic acid, galactose or xylose
residues at their reducing terminals. These observations demonstrate that nonterminal internal
glucuronide, galactoside and xyloside linkages of GAG chains are cleaved in tissues. Indeed,
Takagaki et al. (13, 14) found endo-type glycosidase activities acting on the linkage regions
in rabbit liver, in the form of ChS-degrading endo-β-glucuronidase, endo-β-galactosidase and
endo-β-xylosidase activities. These enzymes appear to act in the initial catabolism of PGs in
animal tissues. One of them, endo-β-xylosidase, was purified from the mid-gut gland of the
mollusk Patinopecten (15). It specifically hydrolyzes the internal xylosyl serine (Xyl-Ser)
linkage between a core protein and a GAG chain (ChS, DS and HS) and is therefore a useful
tool for isolating native GAG chains, including the linkage region, from GAG-peptides.
Cellulases endolytically hydrolyze 1,4-β-glucoside linkages, and are widely distributed
in plants, microorganisms and in particular, fungi. Furthermore, the presence of endogenous
by guest on June 20, 2018http://w
ww
.jbc.org/D
ownloaded from
Page 4
4
cellulases in animals has been suggested recently (16-21). There is evidence that exo-type
enzymes recognize both xyloside and glucoside linkages. An exo-xylosidase acting on p-
nitrophenyl β-xylose and p-nitrophenyl β-glucose has been purified from Charonia lampas
(22, 23). Additionally, it was suggested that β-glucosidase purified from Aspergillus sojae has
β-xylosidase activity (24), and a single protein in pig kidney has both β-xylosidase activity
and β-glucosidase activity (25). Xylose and glucose have very similar configurations, with
the C-5 of xylose having a hydrogen atom rather than the hydroxymethyl group of glucose.
Therefore, it is likely that enzymes that recognize glucose will also recognize xylose.
Additionally, it is of interest to determine whether the endo-β-xylosidase derived from
Patinopecten was originally a cellulase, or whether it is a cellulase with endo-β-xylosidase
activity, or whether the two activities represent independent enzymes.
In this paper, we demonstrate that a fungal cellulase is able to hydrolyze the Xyl-Ser
linkage between the core peptide and the GAG chain of GAG-peptides. Although this
cellulase has endo-β-xylosidase activity, it is different from the endo-β-xylosidase of
Patinopecten.
by guest on June 20, 2018http://w
ww
.jbc.org/D
ownloaded from
Page 5
5
EXPERIMENTAL PROCEDURES
Materials — Cellulase preparations (EC 3.2.1.4) derived from Aspergillus niger (1.6
units/mg, 1 unit defined as the amount of enzyme that liberates 1 µmol of reducing sugar as
glucose in 1 h at pH 4.5 at 37°C), Penicillium funiculosum (5.1 units/mg, 1 unit defined as the
amount of enzyme that liberates 1 µmol of reducing sugar as glucose in 1 h at pH 5.0 at
37°C), Trichoderma reesei (6.3 units/mg, 1 unit defined as the amount of enzyme that
liberates 1 µmol of reducing sugar as glucose in 1 h at pH 5.0 at 37°C), and Trichoderma
viride (4.0 units/mg, 1 unit defined as the amount of enzyme that liberates 1 µmol of reducing
sugar as glucose in 1 h at pH 5.0 at 37°C) were purchased from Sigma Chem. Co., St. Louis,
MO. A cellulase preparation derived from Eupenicillium sp. (1.0 units/mg, 1 unit defined as
the amount of enzyme that liberates 1 µmol of 4-methylumbelliferone from 4-
methylumbelliferyl-β-lactoside in 1 h at 37°C) was a gift from Dr. Yasushi Mitsuishi,
National Institute of Advanced Industrial Science and Technology, Japan. Cellulase from
Irpex lacteus (1.07 units/mg, 1,000 units defined as the amount of enzyme that completely
destroys 2 sheets of quantitative filter paper 1 cm x 1 cm in 1 min at pH 5.0 at 37°C) was a
gift from Kyowa Hakko Kogyo Co., Tokyo, Japan. Cellulases from Pisum sativum L. and
Populus nigra (2 units/ml and 1 unit/ml, respectively, 1 unit defined as the amount of enzyme
that causes a 1% decrease in viscosity of carboxymethyl-cellulose in 2 h at pH 6.2 at 35°C)
were gifts from Dr. Takahisa Hayashi, Wood Research Institute of Kyoto University, Japan.
Endo-β-xylosidase was purified from Patinopecten as described previously (15). 4-
Methylumbelliferyl GAG (GAG-MU), which is an artificial substrate for endo-β-xylosidase,
was prepared from cultured medium of human skin fibroblasts by the method previously
reported (26). After digestion of GAG-MU with hyaluronidase from Streptomyces
hyalurolyticus (Seikagaku Kogyo Co., Tokyo, Japan), Galβ1-4Xylβ1-MU and Galβ1-
3Galβ1-4Xylβ1-MU were purified by Bio-Gel P-4 (Bio-Rad, Richmond, CA) column
chromatography as described previously (27, 28). To obtain ∆GlcAβ1-3GalNAcβ1-3GlcAβ1-
by guest on June 20, 2018http://w
ww
.jbc.org/D
ownloaded from
Page 6
6
3Galβ1-3Galβ1-4Xylβ1-MU and ∆GlcAβ1-3Galβ1-3Galβ1-4Xylβ1-MU, GAG-MU was
digested with chondroitinase ABC and chondroitinase ACII (Seikagaku Kogyo Co.) as
described by Saito et al. (29), and then purified by Sephadex G-50 (Amersham Pharmacia
Biotec AB, Uppsala, Sweden) column chromatography. 4-Methylumbelliferyl-β-cellobioside
(cellobiose-MU) and 4-methylumbelliferyl-β-xylose (Xyl-MU) were purchased from Sigma
Chem. Co.
Chondroitin sulfate proteoglycan (ChS-PG), dermatan sulfate proteoglycan (DS-PG) and
heparan sulfate proteoglycan (HS-PG) were extracted from salmon cartilage, pig skin and
bovine lung, and purified by standard procedures as described by Heinegård and Hascall (30).
These PGs were digested with actinase E (Kaken Pharmaceutical Co., Tokyo, Japan) in 0.1 M
Tris-HCl buffer, pH 8.0, containing 10 mM CaCl2 at 50°C for 24 h. After digestion, the GAG-
peptides (ChS-peptides, DS-peptides and HS-peptides) were separated by DEAE-cellulose
(Whatman Chemical Separation Ltd., Maidstone, United Kingdom) column chromatography
and then purified by Sephacryl S-200 HR (Amersham Pharmacia Biotec AB) column
chromatography. The molecular weights of the peptide moieties of the ChS-peptides, DS-
peptides and HS-peptides were all about 1,000 as estimated from gel-filtration
chromatography based on the method of Ishido et al. (31). The linkage regions of the GAG-
peptides were analyzed for sulfate and phosphate contents based on the method of Takagaki
et al. (28). Sulfate or phosphate were not detected; therefore, it was considered that the GAG-
peptides used in this study did not contain phosphorylated xylose or sulfated galactose. All
other chemicals were obtained from commercial sources.
Fluorescence labeling with 2-aminopyridine — Fluorescence labeling of the reducing
terminal of the oligosaccharides with 2-aminopyridine (PA; Wako Pure Chemical Ind. Co.,
Tokyo, Japan) was carried out as described previously (32).
Assay of endo-β-xylosidase activity and cellulase activity — Endo-β-xylosidase activity was
by guest on June 20, 2018http://w
ww
.jbc.org/D
ownloaded from
Page 7
7
assayed as described previously (15). A complete incubation mixture (200 µl), which
contained 2 µM GAG-MU as a substrate, 0.1 M glucono-1,5-lactone (Wako Pure Chem. Co.)
as an inhibitor of exo-glucosidase, 0.1 M sodium acetate buffer, pH 5.0, and enzyme solution,
was incubated at 37°C for 1 h. The reaction was stopped and fluorescence developed by
adding 1 ml of 0.5 M glycine-NaOH buffer, pH 10.4. The fluorescence was measured on a
spectrofluorometer (Hitachi F-4500; Hitachi, Tokyo, Japan) at excitation and emission
wavelengths of 350 and 450 nm.
Cellulase activity was assayed with 2 µM cellobiose-MU as substrate based on the
method of Chernoglazov et al. (33).
One unit was defined as the amount of enzyme that liberated 1 µmol/min of MU from
GAG-MU or cellobiose-MU.
Another assay for cellulase activity using CM-cellulose as a substrate was performed as
described by Hurst et al. (34).
Assay of hydrolysis of natural substrates — Hydrolysis of natural substrates, GAG-peptides
(10 nmol) or intact ChS-PG (10 nmol; the molar amount of ChS-PG was calculated as the
molar amount of the GAG sugar chains), was assayed as described previously (35). Each
GAG-peptide was incubated with the purified cellulase from A. niger at 37°C for 12 h, and
the reducing terminals of the liberated GAG chains were labeled with PA. Then, each GAG-
PA chain was hydrolyzed in 2 M HCl at 100°C for 2 h. PA-xylose was detected by HPLC
with an Ultrasphere ODS column (4.6 mm x 250 mm; Beckman Coulter Inc., Fullerton, CA).
The molar amounts of the GAG-peptides were determined as the molar amount of PA-xylose
(Takara Shuzo, Kyoto, Japan).
Purification of cellulase— The following procedures were conducted at 4°C. First,
commercial cellulase (30 mg of protein) from A. niger was applied to a column of Sephacryl
S-100 HR (1.5 cm x 102 cm; Amersham Pharmacia Biotec AB) equilibrated previously with
by guest on June 20, 2018http://w
ww
.jbc.org/D
ownloaded from
Page 8
8
20 mM Tris-HCl buffer, pH 7.0, at a flow rate of 15 ml/h, and the fractions with cellulase
activity and endo-β-xylosidase activity were pooled. Next, the active fractions were applied
to a column of POROS PI (4.6 mm x 100 mm; Applied Biosystems Japan, Tokyo, Japan)
equilibrated previously with 20 mM Tris-HCl buffer, pH 7.0. The column was eluted with a
linear gradient of NaCl from 0 to 1 M containing the above buffer at a flow rate of 10 ml/min.
The fractions with both activities were pooled and, lastly, the pool was subjected to
isoelectric focusing using a Rotofor IEF Cell (Bio-Rad) with ampholyte (Bio-Rad) ranging
from pH 3.0 to 10.0 for 4 h.
High-performance liquid chromatography (HPLC) — A high-performance liquid
chromatograph (Hitachi L-6200) connected to a fluorescence detector (Hitachi F-1050) was
used. MU-derivatives and MU were analyzed on an Ultrasphere ODS column (4.6 mm x 250
mm; Beckman Coulter Inc.) with a linear gradient of acetonitrile from 0% to 30% for 50 min
at a flow rate of 1.0 ml/min at 30°C. For detection, eluates were monitored at excitation and
emission wavelengths of 325 and 380 nm. PA monosaccharides were analyzed on an
Ultrasphere ODS column (4.6 mm x 250 mm; Beckman Coulter Inc.) with 0.25 M sodium
citrate and 1% acetonitrile at a flow rate of 0.5 ml/min at 30°C, as described previously (26).
For detection, eluates were monitored at excitation and emission wavelengths of 320 and 400
nm. PA-glucose, PA-galactose and PA-xylose (Takara Shuzo) were used as standards.
Electrophoresis — Native polyacrylamide gel electrophoresis (PAGE) was done in 7.5%
polyacrylamide gel at 4°C. Protein was stained with Coomassie Brilliant Blue R-250, and
duplicate gels were cut into 5-mm segments. Each segment was extracted with 10 mM Tris-
HCl buffer, pH 7.0, at 4°C for 24 h, and the activity was assayed. SDS-PAGE was carried out
in 10% polyacrylamide gel according to the methods of Weber and Osborn (36). Protein was
stained with Coomassie Brilliant Blue R-250.
by guest on June 20, 2018http://w
ww
.jbc.org/D
ownloaded from
Page 9
9
Ion-spray mass spectrometry — Mass spectra were obtained on an API-100 triple-quadruple
mass spectrometer (PE SCIEX, Ontario, Canada) equipped with an atmospheric pressure
ionization source, as described previously (27, 37). The samples were dissolved in 50%
methanol and injected at 2 µl/min with a micro-HPLC syringe pump. In positive-ion mode,
scanning was done from m/z 30 to 250 during the 1-min scan (ten cycles).
Protein assay – Protein was determined by the method of Bradford (38), with bovine serum
albumin as a standard.
by guest on June 20, 2018http://w
ww
.jbc.org/D
ownloaded from
Page 10
10
RESULTS
Screening of endo-β-xylosidase activity in cellulases — Endo-β-xylosidase activity, which
hydrolyzed the Xyl-MU linkage of GAG-MU as a substrate, and cellulase activity, which
hydrolyzed cellobiose-MU as a substrate, were assayed in eight cellulase preparations from
five kinds of ascomycetes, one basidiomycetes and two plants, and in endo-β-xylosidase from
Patinopecten mid-gut gland (Fig. 1). Endo-β-xylosidase activity was found in the five
cellulase preparations from Aspergillus niger, Penicillium funiculosum, Trichoderma reesei,
Trichoderma viride and Irpex lacteus. Particularly high endo-β-xylosidase activities were
detected in the cellulases from A. niger and P. funiculosum. These results indicated that some
cellulases have endo-β-xylosidase activity in addition to cellulase activity. Fig. 1-I shows that
the endo-β-xylosidase from Patinopecten appeared to be different from the cellulases, having
much higher endo-β-xylosidase activity and very little cellulase activity.
To study the purity of each cellulase preparation, electrophoresis in 7.5% polyacrylamide
gel was performed and then the gels were stained with Coomassie Brilliant Blue. As a result,
several protein bands were observed in the cellulase preparations, except for that from
Eupenicillium sp. Duplicate gels were cut into 5-mm segments, extracted individually for 24
h with 10 mM Tris-HCl buffer, pH 7.0, at 4°C, and endo-β-xylosidase activity and cellulase
activity were measured in each fraction. These results with native PAGE are summarized in
Fig. 2. In the cellulase preparation from A. niger, endo-β-xylosidase activity was detected in
the same fractions in which cellulase activity was detected (Fig. 2-A). Similar results were
obtained with the preparations from T. reesei, T. viride, and I. lacteus (Fig. 2-C, D and F). In
contrast, the cellulase preparation from Eupenicillium sp. had only cellulase activity but not
endo-β-xylosidase activity (Fig. 2-E). In P. funiculosum, both activities were detected in
segment numbers 4 - 6 and only cellulase activity was detected in segment numbers 7 - 9 (Fig.
2-B). These results suggested that there are two types of cellulase, one having both endo-β-
xylosidase and cellulase activities and one having only cellulase activity.
by guest on June 20, 2018http://w
ww
.jbc.org/D
ownloaded from
Page 11
11
Purification of the cellulase from A. niger — In order to verify that a single enzyme protein
has both cellulase and endo-β-xylosidase activities, we performed the following experiment.
Although the β-xylosidase activity in P. funiculosum was higher than that in A. niger (Fig. 1),
the cellulase from A. niger is more widely available commercially. Thus, from the
commercial cellulase preparation the enzyme was purified 5-fold at 11% yield in endo-β-
xylosidase activity and cellulase activity by the sequential use of Sephacryl S-100 HR
column chromatography, POROS PI column chromatography and isoelectric focusing
electrophoresis (Table I). The cellulase exhibited endo-β-xylosidase activity during the course
of purification, and the ratio of each enzyme activity was almost constant. Fig. 3 shows that
native PAGE of this purified enzyme yielded a single band, with an estimated molecular
weight of 26,000 by SDS-PAGE. These results showed that the single protein band has the
activities of endo-β-xylosidase and cellulase. Additionally, the optimal pH for the enzyme
toward CM-cellulose was 3.8 (data not shown). The molecular weight of this enzyme and its
optimal pH toward CM-cellulose corresponded with the properties of cellulase from A. niger,
which has been purified previously (34, 39).
Action of the purified cellulase from A. niger on artificial substrates — To examine the action
of the purified cellulase from A. niger on the artificial substrates GAG-MU and cellobiose-
MU, the enzymatic products were subjected to reverse-phase HPLC (Fig. 4). After incubation
with the purified cellulase, the peaks of GAG-MU and cellobiose-MU had disappeared and
new fluorogenic components were found to be eluted at the position of authentic MU (Fig. 4).
The new fluorogenic components were collected and ion-spray mass spectrometry analysis
was carried out. The spectra of both new fluorogenic components showed two peaks of m/z
[M+H]+ ion and [M+Na]+ ion at m/z 177 [M+H]+ and 199 [M+Na]+, respectively (Fig. 5). The
combined results of HPLC analysis and ion-spray mass spectrometry indicated that this
fluorogenic component was MU, presumably released from the corresponding reducing ends
by guest on June 20, 2018http://w
ww
.jbc.org/D
ownloaded from
Page 12
12
of GAG-MU and cellobiose-MU.
To identify the newly exposed reducing terminal sugars of the other incubation products,
they were fluorescence-labeled with PA and hydrolyzed with 2 M HCl at 100°C for 2 h. After
N-acetylation with acetic anhydride, PA-sugars were subjected to reverse-phase HPLC on an
Ultrasphere ODS column as described previously (35). An elution profile is shown in Fig. 6.
The PA-sugar derived from GAG-MU gave a single peak at the position of standard PA-Xyl
(Fig. 6-A). On the other hand, that from cellobiose-MU gave a single peak at the position of
standard PA-glucose (Fig. 6-B). These data proved that xylose and glucose were exposed at
the reducing terminal ends of the sugar chains after digestion of GAG-MU and cellobiose-
MU with the cellulase. Thus, these results showed that the purified cellulase from A. niger
was able to hydrolyze endolytically the 4-methylumbelliferyl-β-xyloside linkage and the 4-
methylumbelliferyl-β-glucoside linkage of GAG-MU and cellobiose-MU, respectively.
Comparison of endo-β-xylosidase activity and cellulase activity of purified enzyme from A.
niger – The optimal pH values, time courses, and effects of metal ions were compared for
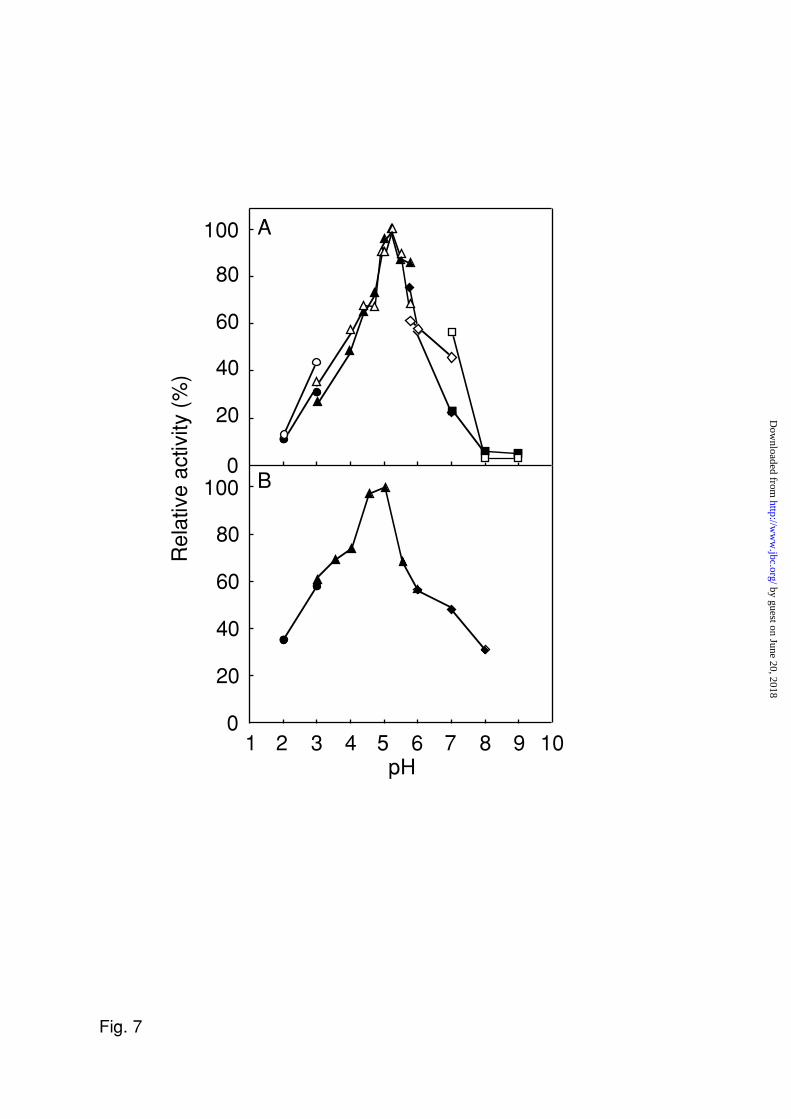
both hydrolysis reactions. For both activities, optimal pH was around 5.0 and the activity
curves were similar (Fig. 7-A). In the time course studies, hydrolysis by endo-β-xylosidase
was slower than that by cellulase (data not shown). Neither activity was affected by
monovalent cations or EDTA, with little difference discernible between endo-β-xylosidase
activity and cellulase activity. However endo-β-xylosidase activity was slightly decreased by
bivalent cations compared with cellulase activity (Table II).
Effect of GAG size on the endo-β-xylosidase activity from A. niger — To investigate the effect
of GAG size on the purified cellulase, several MU derivatives were incubated with the
purified cellulase. After incubation, liberated MU was measured by HPLC. Hydrolysis
activity was greater with shorter GAG chains, but Xyl-MU was not hydrolyzed (Table III).
Hydrolysis by the endo-β-xylosidase from Patinopecten, obtained from a previous report (15),
by guest on June 20, 2018http://w
ww
.jbc.org/D
ownloaded from
Page 13
13
is also shown. Endo-β-xylosidase from Patinopecten hydrolyzed GAG-MU efficiently, but
oligosaccharides were unable to serve as substrates. We concluded that substrate specificity
for GAG chain length is different between the purified cellulase from A. niger and the endo-
β-xylosidase from Patinopecten.
Activity of the endo-β-xylosidase from A. niger toward natural substrates — The properties of
the endo-β-xylosidase activity from A. niger toward natural substrates were examined.
First, in order to investigate whether the enzyme can act on intact PG, the time course of
hydrolysis of ChS-PG was compared with that of ChS-peptide (Fig. 8). The results showed
that the hydrolysis rate of ChS-PG was small compared with that of ChS-peptide. Therefore,
it is difficult for the enzyme to cleave the Xyl-Ser linkage of PG.
Secondly, the effects of pH and metal ions on the activity toward ChS-peptide were
examined to determine optimal assay conditions. With ChS-peptide as substrate, the optimal
pH was around 5.0 (Fig. 7-B), which corresponded with the optimal pH values toward
cellobiose-MU and GAG-MU (Fig. 7-A). In addition, the effect of metal ions on endo-β-
xylosidase activity toward ChS-peptide was similar to that on activity toward GAG-MU
(Table II).
Thirdly, the effect of GAG components on hydrolysis activity was examined at pH 5.0
without any metal ions. The purified cellulase hydrolyzed the Xylβ1-O-Ser linkage of ChS-
peptide, DS-peptide and HS-peptide. Hydrolysis rates of the GAG-peptides are shown in
Table IV. The Km value for ChS-peptide was estimated to be 0.84 mM by Lineweaver-Burk
plot (data not shown).
by guest on June 20, 2018http://w
ww
.jbc.org/D
ownloaded from
Page 14
14
DISCUSSION
PGs are composed of a core protein and a GAG chain whose molecular weight is several
tens of thousands (1, 2). The GAG chains are attached to serine residues of the core protein
via a linkage region, GlcAβ1-3Galβ1-3Galβ1-4Xylβ1-O-Ser, which is common to ChS, DS,
HS and heparin (8). In this paper, we showed that the cellulase purified from A. niger has
endo-β-xylosidase activity, which cleaves the Xyl-Ser linkage between the core protein and
the GAG chain of GAG-peptides, in addition to containing endo-β-glucosidase. An endo-β-
xylosidase with similar catalytic activity has been purified from Patinopecten and
characterized (15). Examination of the specificity of the cellulase from A. niger revealed that
1) the enzyme acts on the linkage region (Xyl-Ser) between a core peptide and GAG chains;
2) enzymatic activity is greater with shorter GAG chains; 3) the enzyme readily hydrolyzes
the linkage in GAG-peptides, but intact PG is cleaved only slowly; 4) the activity is
unaffected by the GAG component (ChS, DS and HS). This enzyme was similar to endo-β-
xylosidase from Patinopecten in 1), 3) and 4). However, Patinopecten endo-β-xylosidase
catalyzes hydrolysis of long GAG chains, but short GAG chains are hardly cleaved.
Additionally, the amino acid sequence of endo-β-xylosidase from Patinopecten showed no
homology to that of the cellulases (unpublished data). Taken together with the fact that the
endo-β-xylosidase from Patinopecten had little cellulase activity, these observations show
that the Patinopecten enzyme is different from the cellulase purified in this study.
It is known that some glycosidases recognize both xyloside linkages and glucoside
linkages as a result of the similarity of the configurations of these molecules. An exo-type
glycosidase acting on both p-nitrophenyl β-glucoside and p-nitrophenyl β-xyloside has been
purified from Charonia lampas (22, 23). Moreover, it has been suggested that β-glucosidase
purified from Aspergillus sojae has β-xylosidase activity (24), and a single protein in pig
kidney has both β-xylosidase activity and β-glucosidase activity (25). Also, it has been
reported that addition of Xyl-MU to cultured human skin fibroblasts or Chinese hamster
by guest on June 20, 2018http://w
ww
.jbc.org/D
ownloaded from
Page 15
15
ovary cells induces SAα2-3Galβ1-4Xylβ1-MU, which is initiated by β-xyloside as a primer,
rather than the expected SAα2-3Galβ1-4Glcβ1-MU, which is initiated by β-glucose as a
primer (40, 41), suggesting that the galactosyltransferases involved in biosynthesis cannot
distinguish between xylose and glucose. Similarly, cellulase, which is a glucosidase, seems
also to show endo-type β-xylosidase activity.
Endo-β-xylosidase activity was compared with endo-β-glucosidase activity in the
purified cellulase from A. niger. The optimal pH was around 5.0 and the activity curves were
similar for both activities. Hydrolysis by endo-β-xylosidase was slower than that by endo-β-
glucosidase (data not shown). The inhibitory effects of metal ions were similar for endo-β-
xylosidase activity and endo-β-glucosidase activity across the range of 11 reagents examined.
Endo-β-xylosidase activity was slightly inhibited by bivalent cations compared with cellulase
activity. GAG-MU was used as a substrate for assay of endo-β-xylosidase activity. GAG
contains a large number of negative charges such as sulfate groups and carboxyl groups (42).
It is known that bivalent cations, for example Ca2+, can form chelates between the negative
charges of GAG (43). Thus, the configuration of the entire GAG may be altered by binding of
cation, which may have affected enzymatic activity.
The properties of endo-β-xylosidase activity toward natural substrates were investigated.
The optimal pH for, and the effect of metal ions on, hydrolysis of ChS-peptide were similar to
those of hydrolysis of GAG-MU. The enzyme hydrolyzed the Xyl-Ser linkages of ChS-
peptide, DS-peptide and HS-peptide, but not ChS-PG. In addition, the hydrolysis of HS-
peptide was faster than that of ChS-peptide and DS-peptide.
Although GAG chains are attached to serine residues of the core protein via the common
linkage region (8), it has been shown recently that the linkage region is modified by
phosphate and sulfate groups (9-11). Additionally, it has been suggested that the linkage
region can act as a recognition signal for sorting in the biosynthesis of different GAG chains
(11). The linkage regions of the GAG-peptides used in this study did not contain
phosphorylated xylose or sulfated galactose. Therefore, it is unclear whether the enzyme can
by guest on June 20, 2018http://w
ww
.jbc.org/D
ownloaded from
Page 16
16
act on substrates containing phosphorylated xylose and/or sulfated galactose, and this
requires further investigation.
Endo-β-xylosidase activity was found in cellulase preparations from A. niger, P.
funiculosum, T. reesei, T. viride and I. lacteus, although not in the preparations from
Eupenicillium sp., P. nigra and P. sativum. Consequently, this activity is not specific for the
cellulase from A. niger. Although the presence or absence of endo-β-xylosidase activity may
be related to the different action patterns of cellulases, for example endolytic or exolytic
cleavage, this is not clear at the present time.
The structural diversity of GAG chains, in terms of sugar chain length, sugar chain
composition and position, and degree of sulfation, is well established. However, all GAG
chains are attached to a xylose residue, which forms an O-glycoside linkage (9). A chemical
method, β-elimination (44), is known to release GAG chains, but may cause their
decomposition by the peeling reaction. On the other hand, endo-β-xylosidase is useful for
analysis of GAG chains, since it specifically releases intact GAG chains from GAG-peptides.
Using this enzyme, it was possible to analyze GAG chains from 50 mg wet weight of animal
tissues with the combined use of PA-labeling at the reducing terminal of the released sugar
chains and HPLC (45). Furthermore, this enzyme is useful in the molar quantification of
GAG chains (45), in giving directivity to GAG chains (46), and in preparing artificial
substrates for novel endo-type enzymes (35). Therefore, we believe that cellulases with endo-
β-xylosidase activity will become useful tools for structural and functional analysis.
by guest on June 20, 2018http://w
ww
.jbc.org/D
ownloaded from
Page 17
17
REFERENCES
1. Rodén, L. (1980) in The Biochemistry of Glycoproteins and Proteoglycans (Lennarz, W.
J., ed.), pp. 267-371, Plenum Publishing Corp., New York
2. Hascall, V. C. (1986) in Functions of the Proteoglycans (Evered, D., and Whelan, J.,
eds.), pp. 1-8, John Wiley & Sons, New York
3. Raab, G., and Klagsbrun, M. (1997) Biochim. Biophys. Acta 1333, 179-199
4. Casu, B., Oreste, P., Torri, G., Zoppetti, G., Choay, J., Lormeau, J. C., Petitou, M., and
Sinay, P. (1981) Biochem. J. 197, 599-609
5. Thunberg, L., Bäckström, G., and Lindahl, U. (1982) Carbohydr. Res. 100, 393-410
6. Grootenhuis, P. D. J., and van Boeckel, C. A. A. (1991) J. Am. Chem. Soc. 113, 2743-
2747
7. Maimone, M.M., and Tollefsen, D.M. (1990) J. Biol. Chem. 265, 18263-18271
8. Lindahl, U., and Rodén, L. (1972) Glycoproteins (Gottschalk, A., ed.), pp. 491-517,
Elsevier Scientific Publishing Co., Amsterdam
9. Oegema, T. R., Jr., Kraft, E. L., Jourdian, G. W., and Van Valen, T. R. (1984) J. Biol.
Chem. 259, 1720-1726
10. Fransson, L. Å., Silverberg, I., and Carlstedt, I. (1985) J. Biol. Chem. 260, 14722-14726
11. Sugahara, K., Yamashina, I., De Waard, P., Van Halbeek, H., and Vliegenthart, J. F. G.
(1988) J. Biol. Chem. 263, 10168-10174
12. Matsue, H., and Endo, M. (1987) Biochim. Biophys. Acta 923, 470-477
13. Takagaki, K., Nakamura, T., Majima, M., and Endo, M. (1985) FEBS Lett. 181, 271-274
14. Takagaki, K., Nakamura, T., and Endo, M. (1988) Biochim. Biophys. Acta 966, 94-98
15. Takagaki, K., Kon, A., Kawasaki, H., Nakamura, T., Tamura, S., and Endo, M. (1990) J.
Biol. Chem. 265, 854-860
16. Smant, G., Stokkermans, J. P. W. G., Yan, Y., De Boer, J. M., Baum, T. J., Wang, X.,
Hussey, R. S., Gommers, F. J., Henrissat, B., Davis, E. L., Helder, J., Schots, A., and
by guest on June 20, 2018http://w
ww
.jbc.org/D
ownloaded from
Page 18
18
Bakker, J. (1998) Proc. Natl. Acad. Sci. U. S. A. 95, 4906-4911
17. Girand, C., and Jouanin, L. (1999) Insect Biochem. Mol. Biol. 29, 1129-1142
18. Watanabe, H., Noda, H., Tokuda, G., and Lo, N. (1998) Nature 394, 330-331
19. Tokuda, G., Lo, N., Watanabe, H., Slaytor, M., Matsumoto, T., and Noda, H. (1999)
Biochim. Biophys. Acta 1447, 146-159
20. Lo, N., Tokuda, G., Watanabe, H., Rose, H., Slaytor, M., Maekawa, K., Bandi, C., and
Noda, H. (2000) Curr. Biol. 10, 801-804
21. Byrne, K. A., Lehnert, S. A., Johnson, S. E., and Moore, S. S. (1999) Gene 239, 317-324
22. Fukuda, M., Muramatsu, T., and Egami, F. (1969) J. Biochem. (Tokyo) 65, 191-199
23. Fukuda, M., and Egami, F. (1969) J. Biochem. (Tokyo) 66, 157-164
24. Kimura, I., Yoshioka, N., and Tajima, S. (1999) J. Biosci. Bioeng. 87, 538-541
25. Robinson, D., and Abrahams, H. E. (1967) Biochim. Biophys. Acta 132, 212-214
26. Takagaki, K., Kon, A., Kawasaki, H., Nakamura, T., and Endo, M. (1989) J. Biochem.
Biophys. Methods 19, 207-214
27. Takagaki, K., Kojima, K., Majima, M., Nakamura, T., Kato, I., and Endo, M. (1992)
Glycoconjugate J. 9, 174-179
28. Takagaki, K., Nakamura, T., Kon, A., Tamura, S., and Endo, M. (1991) J. Biochem.
(Tokyo) 109, 514-519
29. Saito, H., Yamagata, T., and Suzuki, S. (1968) J. Biol. Chem. 243, 1536-1542
30. Heinegård, D., and Hascall, V. C. (1974) J. Biol. Chem. 249, 4250-4256
31. Ishido, K., Takagaki, K., Iwafune, M., Yoshihara, S., Sasaki, M. and Endo, M. (2002) J.
Biol. Chem. in press
32. Kon, A., Takagaki, K., Kawasaki, H., Nakamura, T., and Endo, M. (1991) J. Biochem.
(Tokyo) 110, 132-135
33. Chernoglazov, V. M., Jafarova, A. N., and Klyosov, A. A. (1989) Anal. Biochem. 179,
186-189
34. Hurst, P. L., Nielsen, J., Sullivan, A., and Shepherd, M. G. (1977) Biochem. J. 165, 33-41
by guest on June 20, 2018http://w
ww
.jbc.org/D
ownloaded from
Page 19
19
35. Takagaki, K., Nakamura, T., Kawasaki, H., Kon, A., Ohishi, S., and Endo, M. (1990) J.
Biochem. Biophys. Methods 21, 209-215
36. Weber, K., and Osborn, M. (1969) J. Biol. Chem. 244, 4406-4412
37. Takagaki, K., Munakata, H., Nakamura, W., Matsuya, H., Majima, M., and Endo, M.
(1998) Glycobiology 8, 719-724
38. Bradford, M. M. (1976) Anal. Biochem. 72, 248-254
39. Okada, G (1985) Agric. Biol. Chem. 49, 1257-1265
40. Shibata, S., Takagaki, K., Nakamura, T., Izumi, J., Kojima, K., Kato, I., and Endo, M.
(1995) J. Biol. Chem. 270, 13794-13798
41. Freeze, H. H., Sampath, D., and Varki, A. (1993) J. Biol. Chem. 268, 1618-1627
42. Roux, L., Holojda, S., Sundblad, G., Freeze, H.H., and Varki, A., (1988) J. Biol. Chem.
263, 8879-8889
43. Hunter, G. K., Wong, K. S., and Kim, J. J. (1988) Arch. Biochem. Biophys 260, 161-167
44. Heinegård, D. (1972) Biochim. Biophys. Acta 285, 193-207
45. Takagaki, K., Takeda, Y., Nakamura, T., Daidouji, K., Narita, H., and Endo, M. (1994) J.
Biochem. Biophys. Methods 28, 313-320
46. Takagaki, K., Nakamura, T., Izumi, J., Saitoh, H., Endo, M., Kojima, K., Kato, I., and
Majima, M. (1994) Biochemistry 33, 6503-6507
by guest on June 20, 2018http://w
ww
.jbc.org/D
ownloaded from
Page 20
20
Footnotes:
This work was supported by Grants-in-Aid (nos. 09358013, 11121203, 11470029,
12680603 and 12793010) for Scientific Research from the Ministry of Education, Culture,
Sports, Science, and Technology of Japan, and grants from the Aomori Support Center for
Industrial Promotion.
The abbreviations used are: PG, proteoglycan; GAG, glycosaminoglycan; ChS,
chondroitin sulfate; DS, dermatan sulfate; HS, heparan sulfate; Ser, serine; MU, 4-
methylumbelliferone; PA, 2-aminopyridine; HPLC, high-performance liquid
chromatography; PAGE, polyacrylamide gel electrophoresis; GlcA, glucuronic acid; Gal,
galactose; Xyl, xylose; ∆GlcA, D-gluco-4-enepyranosyluronic acid; GalNAc, N-
acetylgalactosamine; SA, sialic acid; IdoA, iduronic acid. All sugars mentioned in this paper
are of D configuration except for iduronic acid.
by guest on June 20, 2018http://w
ww
.jbc.org/D
ownloaded from
Page 21
21
FIGURE LEGENDS
Fig. 1. Comparison between endo-β-xylosidase activity and cellulase activity in
cellulase preparations from various origins and in endo-β-xylosidase. Each activity was
measured using 1 unit of each enzyme as defined in Materials under “EXPERIMENTAL
PROCEDURES”. Endo-β-xylosidase activity was measured with 2 µM GAG-MU as
substrate (solid bars), and cellulase activity was measured with 2 µM cellobiose-MU as a
substrate (open bars). The cellulase preparations were derived from A, A. niger; B, P.
funiculosum; C, T. reesei; D, T. viride; E, Eupenicillium sp.; F, I. lacteus; G, P. sativum L.; H,
P. nigra; and I, endo-β-xylosidase from Patinopecten.
Fig. 2. Native PAGE of various cellulase preparations. Each cellulase preparation (A, A.
niger; B, P. funiculosum; C, T. reesei; D, T. viride; E, Eupenicillium sp.; F, I. lacteus) was
separated in 7.5% polyacrylamide gel and stained with Coomassie Brilliant Blue. Duplicate
gels were cut into 5-mm segments, and extracted individually for 24 h with 10 mM Tris-HCl
buffer, pH 7.0, at 4°C, and endo-β-xylosidase activity (solid bars) and cellulase activity (open
bars) were measured in each fraction.
Fig. 3. Native PAGE (A) and SDS-PAGE (B) of the purified cellulase from A. niger.
PAGE and SDS-PAGE were performed in 7.5% and 10% polyacrylamide gel, respectively, at
a constant 20 mA. Proteins were stained with Coomassie Brilliant Blue.
Fig. 4. Analysis by HPLC of GAG-MU (A and B) and cellobiose-MU (C and D) before
and after digestion with the purified cellulase from A. niger. MU derivatives were
analyzed by HPLC before digestion (A and C) and after digestion (B and D) with the purified
cellulase from A. niger. An Ultrasphere ODS column (4.6 mm x 250 mm) was used with a
linear gradient of acetonitrile from 0% to 30% for 50 min at a flow rate of 1.0 ml/min at 30°C.
by guest on June 20, 2018http://w
ww
.jbc.org/D
ownloaded from
Page 22
22
For detection, eluates were monitored at excitation and emission wavelengths of 325 and 380
nm. The arrow indicates the elution position of authentic MU.
Fig. 5. Ion spray mass spectra of new fluorogenic products obtained from GAG-MU
and cellobiose-MU by cellulase digestion. GAG-MU (A) and cellobiose-MU (B) were
digested with the purified cellulase from A. niger, and positive-mode mass spectra of the
hydrolysates were analyzed. The spectra were acquired after injection of the samples
dissolved in a mobile phase of 50% methanol.
Fig. 6. HPLC chromatograms of the reducing terminal sugars in the hydrolysates
obtained from GAG-MU and cellobiose-MU by cellulase digestion. GAG-MU (A) and
cellobiose-MU (B) were digested with the purified cellulase from A. niger. The hydrolysates
were fluorescence-labeled with PA and hydrolyzed with 2 M HCl at 100°C for 2 h. After N-
acetylation with acetic anhydride, PA-sugars were subjected to reverse-phase HPLC. An
Ultrasphere ODS column (4.6 mm x✖250 mm) was used with 0.25 M sodium citrate and 1%
acetonitrile at a flow rate of 0.5 ml/min at 30°C. For detection, eluates were monitored at
excitation and emission wavelengths of 320 and 400 nm. The arrows indicate the elution
positions of the PA-monosaccharide standards: 1, PA-glucose; 2, PA-galactose; 3, PA-xylose.
Fig. 7. Effect of pH on the purified cellulase from A. niger. GAG-MU (solid symbols)
and cellobiose-MU (open symbols) were incubated with the purified enzyme (A). ChS-
peptide was incubated with the enzyme (B). The assay method for enzyme activity followed
that described in “EXPERIMENTAL PROCEDURES” except for the composition of the
buffer. The following buffers were used: circles, 0.1 M glycine-HCl buffer; triangles, 0.1 M
sodium acetate buffer; diamonds, 0.1 M sodium phosphate buffer; squares, 0.1 M Tris-HCl
buffer.
by guest on June 20, 2018http://w
ww
.jbc.org/D
ownloaded from
Page 23
23
Fig. 8. Time courses of the hydrolysis of natural substrates by the purified cellulase
from A. niger. ChS-PG (open circles) and ChS-peptide (solid circles) were incubated with the
purified enzyme as described under “EXPERIMENTAL PROCEDURES”.
by guest on June 20, 2018http://w
ww
.jbc.org/D
ownloaded from
Page 24
6
.
7
0
1
.
0
2
.
0
3
.
0
A B C D E F G H I
0
.
5
1
.
0
1
.
5
0
A c
t
i
v
i
t
y
o
f
e
n
d
o
-β
-
x
y
l
o
s
i
d
a
s
e
(
u
n
i
t
,
)
A c
t
i
v
i
t
y
o
f
c
e
l
l
u
l
a
s
e
(
u
n
i
t
,
)
F
i
g
.
1
by guest on June 20, 2018http://w
ww
.jbc.org/D
ownloaded from
Page 25
A
S e
g
m
e
n
t
n
u
m
b
e
r
B
S e
g
m
e
n
t
n
u
m
b
e
r
C
S e
g
m
e
n
t
n
u
m
b
e
r
0 0 .
2 0
.
4 0
.
6 0
.
8 1
1 2
1 1
1 0987654321
A
c
t
i
v
i
t
y
(
u
n
i
t
s
/
m
l
)
0 0 .
2 0
.
4 0
.
6 0
.
8 1
1 2
1 1
1 0987654321
A
c
t
i
v
i
t
y
(
u
n
i
t
s
/
m
l
)
0 0 .
2 0
.
4 0
.
6 0
.
8 1
1 2
1 1
1 0987654321
A
c
t
i
v
i
t
y
(
u
n
i
t
s
/
m
l
)
D
S e
g
m
e
n
t
n
u
m
b
e
r
0 0 .
2 0
.
4 0
.
6 0
.
8 1
1 2
1 1
1 0987654321
A
c
t
i
v
i
t
y
(
u
n
i
t
s
/
m
l
)E
S e
g
m
e
n
t
n
u
m
b
e
r
F
S e
g
m
e
n
t
n
u
m
b
e
r
0 0 .
2 0
.
4 0
.
6 0
.
8 1
1 2
1 1
1 0987654321
A
c
t
i
v
i
t
y
(
u
n
i
t
s
/
m
l
)
0 0 .
2 0
.
4 0
.
6 0
.
8 1
1 2
1 1
1 0987654321
A
c
t
i
v
i
t
y
o
f
c
e
l
l
u
l
a
s
e
(
u
n
i
t
s
/
m
l
)
F
i
g
.
2
by guest on June 20, 2018http://w
ww
.jbc.org/D
ownloaded from
Page 26
+
-
(
)
(
)
A B
1
7
5
k
8
3
k
6
2
k
4
7
.
5
k
3
2.
5
k
2
5
k
F
i
g
.
3
by guest on June 20, 2018http://w
ww
.jbc.org/D
ownloaded from
Page 27
R
e
t
e
n
t
i
o
n
t
i
m
e
(
m
i
n
)
F l
u
o
r
e
s
c
e
n
c
e
i
n
t
e
n
s
i
t
y
0 1 0 2
0 3
0 4
0 5
0
A
B
D
C
F
i
g
.
4
by guest on June 20, 2018http://w
ww
.jbc.org/D
ownloaded from
Page 28
m
/
z1
5
0 2
0
0
R e
l
a
t
i
v
e
i
n
t
e
n
s
i
t
y
A
B
1
7
7
1
7
7
1
9
9
1
9
9[ M
+
H
]+
1
[ M
+
N
a
]+
1
[ M
+
H
]+
1
[ M
+
N
a
]+
1
0
F
i
g
.
5
by guest on June 20, 2018http://w
ww
.jbc.org/D
ownloaded from
Page 29
R
e
t
e
n
t
i
o
n
t
i
m
e
(
m
i
n
)0 1
0 2
0 3
0 4
0
F l
u
o
r
e
s
c
e
n
c
e
i
n
t
e
n
s
i
t
y
1 2 3
B
A
F
i
g
.
6
by guest on June 20, 2018http://w
ww
.jbc.org/D
ownloaded from
Page 30
0
2
0
4
0
6
0
8
0
1
0
0
0
2
0
4
0
6
0
8
0
1 2 3 4 5 6 7 8 9 1
0
R e
l
a
t
i
v
e
a
c
t
i
v
i
t
y
(
%
)
p
H
A
B1
0
0
F
i
g
.
7
by guest on June 20, 2018http://w
ww
.jbc.org/D
ownloaded from
Page 31
0
2 0
4 0
6 0
8 0
1 0
0
0 5 1 0 1
5 2
0 2
5
T
i
m
e
(
h
)
R e
l
a
t
i
v
e
a
c
t
i
v
i
t
y
(
%
)
F
i
g
.
8
by guest on June 20, 2018http://w
ww
.jbc.org/D
ownloaded from
Page 32
A
,
e
n
d
o
-β-
x
y
l
o
s
i
d
a
s
e
a
c
t
i
v
i
t
y
;
B
,
c
e
l
l
u
l
a
s
e
a
c
t
i
v
i
t
y
.
A B A B
Commercial enzyme 3 0 . 0 2 9 . 7 5 5 . 9 1 . 0 1 . 9 1 0 0 1 0 0 1 . 0 1 . 0Sephacryl S-100 HR 7 . 6 9 . 8 2 3 . 1 1 . 3 3 . 0 3 3 4 1 1 . 3 1 . 6POROS PI 1 . 2 3 . 5 1 0 . 6 3 . 0 8 . 8 1 2 1 9 3 . 0 4 . 6Rotofor 0 . 6 3 . 3 6 . 0 5 . 5 1 0 . 0 1 1 1 1 5 . 5 5 . 3
Totalactivity
TABLE IPurification of cellulase from A. niger
Purification
Bunits
RecoveryTotal
m g units/mg
proteinA
Specif icactivity
fo ld%A B
-
by guest on June 20, 2018http://www.jbc.org/Downloaded from
Page 33
T
A
B
L
E
I
I
E
f
f
e
c
t
s
o
f
v
a
r
i
o
u
s
a
g
e
n
t
s
o
n
c
e
l
l
u
l
a
s
e
f
r
o
m
A
.
n
i
g
e
r
T
h
e
e
n
z
y
m
e
w
a
s
a
s
s
a
y
e
d
u
n
d
e
r
n
o
r
m
a
l
c
o
n
d
i
t
i
o
n
s
d
e
s
c
r
i
b
e
d
i
n"
E
X
P
E
R
I
M
E
N
T
A
L
P
R
O
C
E
D
U
R
E
S
"
e
x
c
e
p
t
f
o
r
t
h
e
p
r
e
s
e
n
c
e
o
f
v
a
r
i
o
u
sa
g
e
n
t
s
.
N
o
n
e 1
0
0 1
0
0 1
0
0C
d
C
l2 7
4 7
1 6
9F
e
S
O 4 3 4 1Z
n
C
l2 9
9 9
0 9
4C
u
S
O4 9
4 7
5 8
1
B
a
C
l 2 9
5 7
4 8
5M
g
S
O4 9
7 8
5 8
9
M
n
C
l2 9
5 8
6 9
5
C
a
C
l2 1
0
4 9
2 9
5N
a
C
l 9
7 9
8 1
0
6K
C
l 9
9 9
9 9
1E
D
T
A 1
0
2 9
5 1
0
6
1 0
m
M %
C
o
m
p
o
u
n
dC
h
S
-
p
e
p
t
i
d
e
R
e
l
a
t
i
v
e
a
c
t
i
v
i
t
y
c e
l
l
o
b
i
o
s
e
-
M
U G
A
G
-
M
U
by guest on June 20, 2018http://w
ww
.jbc.org/D
ownloaded from
Page 34
T
A
B
L
E
I
I
IC
o
m
p
a
r
a
t
i
v
e
a
c
t
i
v
i
t
y
o
f
c
e
l
l
u
l
a
s
e
p
u
r
i
f
i
e
d
f
r
o
m
A
.
n
i
g
e
r
A
f
t
e
r
1
µ
M
s
u
b
s
t
r
a
t
e
s
w
e
r
e
i
n
c
u
b
a
t
e
d
w
i
t
h
p
u
r
i
f
i
e
d
c
e
l
l
u
l
a
s
e
f
r
o
m
A
.
n
i
g
e
r,
t
h
e
l
i
b
e
r
a
t
e
d
M
U
w
a
s
a
s
s
a
y
e
d
b
y
H
P
L
C
.
T
h
e
p
r
e
p
a
r
a
t
i
o
n
o
f
s
u
b
s
t
r
a
t
e
s
i
s
d
e
s
c
r
i
b
e
d
i
n
"
E
X
P
E
R
I
M
E
N
T
A
LP
R
O
C
E
D
U
R
E
S
"
.
A
n
U
l
t
r
a
s
p
h
e
r
e
O
D
S
c
o
l
u
m
n
(
4
.
6
m
m
x
2
5
0
m
m
)
w
a
s
u
s
e
d
w
i
t
h
a
l
i
n
e
a
r
g
r
a
d
i
e
n
t
o
fa
c
e
t
o
n
i
t
r
i
l
e
f
r
o
m
0
%
t
o
3
0
%
f
o
r
5
0
m
i
n
a
t
a
f
l
o
w
r
a
t
e
o
f
1
.
0
m
l
/
m
i
n
a
t
3
0
°
C
.
F
o
r
d
e
t
e
c
t
i
o
n
,
e
l
u
a
t
e
sw
e
r
e
m
o
n
i
t
o
r
e
d
a
t
e
x
c
i
t
a
t
i
o
n
a
n
d
e
m
i
s
s
i
o
n
w
a
v
e
l
e
n
g
t
h
s
o
f
3
5
0
a
n
d
4
5
0
n
m
.
T
h
e
a
c
t
i
v
i
t
i
e
s
w
e
r
ed
e
t
e
r
m
i
n
e
d
u
s
i
n
g
t
h
e
p
e
a
k
a
r
e
a
o
f
a
u
t
h
e
n
t
i
c
M
U
.
M
U
d
e
r
i
v
a
t
i
v
e
G
a
l
-
X
y
l
-
M
U
G
a
l
-
G
a
l
-
X
y
l
-
M
U
∆G
l
c
A
-
G
a
l
-
G
a
l
-
X
y
l
-
M
U
∆G
l
c
A
-
G
a
l
N
A
c
-
G
l
c
A
-
G
a
l
-
G
a
l
-
X
y
l
-
M
U
- (
G
l
c
A
/
I
d
o
A
-
G
a
l
N
A
c
(
S
O3 )
)n-
G
l
c
A
-
G
a
l
-
G
a
l
-
X
y
l
-
M
U
X
y
l
-
M
U
2 6
4
2
1 3
4
1 0
0
P
u
r
i
f
i
e
d
c
e
l
l
u
l
a
s
ef
r
o
m
A
.
n
i
g
e
r
1 7
3
1 4
8
1 2
7
3 0
1 0
0
3 0
E
n
d
o
-β-
x
y
l
o
s
i
d
a
s
ef
r
o
m
P
a
t
i
n
o
p
e
c
t
e
n
b
<
1
R
e
l
at
i
v
e
a
c
t
i
v
i
t
y
a
a T
h
e
a
c
t
i
v
i
t
y
s
h
o
w
s
t
h
e
r
e
l
a
t
i
v
e
r
a
t
e
w
h
e
n
t
h
e
a
c
t
i
v
i
t
y
o
f
h
y
d
r
o
l
y
s
i
s
u
s
i
n
g
G
A
G
-
M
U
a
s
a
s
u
b
s
t
r
a
t
e
i
s
1
0
0
.b T
h
e
d
a
t
a
i
s
o
b
t
a
i
n
e
d
f
r
o
m
T
a
k
a
g
a
k
i
e
t
a
l
.
(
1
5
)
.
by guest on June 20, 2018 http://www.jbc.org/ Downloaded from
Page 35
T
A
B
L
E
I
V
E
f
f
e
c
t
o
f
h
y
d
r
o
l
y
s
i
s
a
c
t
i
v
i
t
y
t
o
w
a
r
d
n
a
t
u
r
a
l
s
u
b
s
t
r
a
t
e
s
S
u
b
s
t
r
a
t
e
C
h
S
-
p
e
p
t
i
d
e 1 0
0
D
S
-
p
e
p
t
i
d
e 9 7
H
S
-
p
e
p
t
i
d
e 1 2
4
R
e
l
a
t
i
v
e
a
c
t
i
v
i
t
y a
T
h
e
a
c
t
i
v
i
t
y
s
h
o
w
s
t
h
e
r
e
l
a
t
i
v
e
a
c
t
i
v
i
t
y
o
fh
y
d
r
o
l
y
s
i
s
u
s
i
n
g
C
h
S
-
p
e
p
t
i
d
e
a
s
a
s
u
b
s
t
r
a
t
ei
s
1
0
0
.
a
E
a
c
h
s
u
b
s
t
r
a
t
e
w
a
s
i
n
c
u
b
a
t
e
d
w
i
t
h
t
h
e
p
u
r
i
f
i
e
de
n
z
y
m
e
u
n
d
e
r
t
h
e
t
y
p
i
c
a
l
c
o
n
d
i
t
i
o
n
s
d
e
t
a
i
l
e
d
i
n"
E
X
P
E
R
I
M
E
N
T
A
L
P
R
O
C
E
D
U
R
E
S
"
.
by guest on June 20, 2018http://w
ww
.jbc.org/D
ownloaded from
Page 36
Masahiko EndoKeiichi Takagaki, Mito Iwafune, Ikuko Kakizaki, Keinosuke Ishido, Yoji Kato and
glycosaminoglycan chain by cellulasesThe cleavage of the xylosyl serine linkage between a core peptide and a
published online February 27, 2002J. Biol. Chem.
10.1074/jbc.M111985200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on June 20, 2018http://w
ww
.jbc.org/D
ownloaded from




















































![Systematic Mutagenesis of Serine Hydroxymethyltransferase · Systematic Mutagenesis of Serine Hydroxymethyltransferase Reveals an Essential Role in Nematode Resistance1[OPEN] Pramod](https://static.documents.pub/doc/80x56/5d512bba88c993ee1f8b8729/systematic-mutagenesis-of-serine-hydroxy-systematic-mutagenesis-of-serine-hydroxymethyltransferase.jpg)