Complete Oxidation of Toluene under Strictly AnoxicConditions by a New Sulfate-Reducing Bacterium
RALF RABUS,1 RALPH NORDHAUS,2 WOLFGANG LUDWIG,3 AND FRIEDRICH WIDDELl*Max-Planck-Institut fir marine Mikrobiologie, Fahrenheitstrasse 1, W-2800 Bremen 33,1 Fraunhofer Institut
fir Atmosphanrische Umweltforschung, W-8100 Gannisch-Partenkirchen,2 and Institut fiirMikrobiologie, Technische Universitat, W-8000 Munich 2, 3 Germany
Received 14 January 1993/Accepted 3 March 1993
A toluene-degrading sulfate-reducing bacterium, strain Tol2, was isolated from marine sediment understrictly anoxic conditions. Toluene was toxic if applied directly to the medium at concentrations higher than 0.5mM. To provide toluene continuously at a nontoxic concentration, it was supplied in an inert hydrophobiccarrier phase. The isolate had oval, sometimes motile cells (1.2 to 1.4 by 1.2 to 2.0 ,um). The doubling time was27 h. Toluene was completely oxidized to C02, as demonstrated by measurement of the degradation balance.The presence of carbon monoxide dehydrogenase and formate dehydrogenase indicated a terminal oxidation ofacetyl coenzyme A via the CO dehydrogenase pathway. The use of hypothetical intermediates of toluenedegradation was tested in growth experiments and adaptation studies with dense cell suspensions. Results donot support a degradation of toluene via one of the cresols or methylbenzoates, benzyl alcohol, or phenylacetateas free intermediate. Benzyl alcohol did not serve as growth substrate; moreover, it was a strong, specificinhibitor of toluene degradation, whereas benzoate utilization was not affected by benzyl alcohol. Sequencingof 16S rRNA revealed a relationship to the metabolically dissimilar genus Desulfobacter and on a deeper levelto the genus Desulfobacterium. The new genus and species Desulfobacula toluolica is proposed.
Interest in the biodegradation of aromatic hydrocarbonsarises from their environmental impact as well as fromquestions concerning biochemical reactions. Anthropogenicsources such as petroleum spillage or refuse disposal sitesaccount for contamination of soil and water by benzene,toluene, ethylbenzene, and xylene (31). Interestingly, tolu-ene can also be biologically produced in anoxic hypolimnia(21). The presence of aromatic hydrocarbons in anoxichabitats evokes an interest in organisms that may degradesuch compounds without molecular oxygen. Anaerobic deg-radation of aromatic hydrocarbons has long been disputedbecause no activation reactions other than a direct attackwith molecular oxygen (17) seemed conceivable. Neverthe-less, studies with anaerobic enrichment cultures have dem-onstrated that aromatic hydrocarbons were degraded with-out 02 (13, 19, 22) and that oxygen in phenolic compoundsformed as intermediates was derived from H2O (32). Inrecent years, pure cultures of toluene-degrading and denitri-fying (2, 12, 15, 28) or ferric iron-reducing (23) bacteria havebeen described. The degradation of toluene with sulfate asan electron acceptor has been observed in enrichment cul-tures with river sediment (6) but not to date in pure cultures.A crucial step in the anaerobic degradation of toluene is
the initial activation of the aromatic hydrocarbon. Formationof intermediary products by denitrifying Pseudomonas sp.strain K172 suggested that anaerobic toluene oxidation oc-curs via benzyl alcohol and benzaldehyde (2, 3). Becausemetabolic by-products of anaerobic toluene degradationhave been identified in a denitrifying pure culture (14) and asulfate-reducing enrichment culture (7), an oxidative con-densation of toluene with acetyl coenzyme A (acetyl-CoA)to phenylpropionyl-CoA has been proposed as an alternativestarting reaction. Introduction of an hydroxyl group into thenucleus or a carboxylation reaction has been discussed as an
* Corresponding author.
additional activation reaction (2, 23, 28), but experimentalresults do not support such a mechanism.
Several species of sulfate-reducing bacteria are known toutilize aromatic compounds as sole electron donors andcarbon sources. However, all aromatic substrates used bypure cultures described to date either carry a functionalgroup (e.g., benzoate, phenol, or aniline) (5, 27, 34) orpossess a heteroatom in the ring (e.g., indole or quinoline)(4, 34). Recently, a sulfate-reducing bacterium has beenisolated that completely oxidizes long-chain saturated hy-drocarbons such as hexadecane to CO2 under strictly anoxicconditions (1). Here we report the isolation of a pure cultureof a novel marine sulfate-reducing bacterium that anaerobi-cally degrades an aromatic hydrocarbon, toluene. The con-sumption of hypothetical intermediates was studied to deter-mine possible starting reactions of toluene degradation bythe new isolate.
MATERIALS AND METHODS
Sources of bacteria. Toluene-degrading sulfate reducerswere enriched from anoxic, sulfide-rich marine sedimentsamples from Eel Pond, a seawater pond in Woods Hole,Mass. Desulfobacterium anilini, Desulfobacterium phenoli-cum, and Desulfococcus multivorans (DSM 4660, 3384, and2059, respectively) were from the Deutsche Sammlung vonMikroorganismen und Zellkulturen, Braunschweig, Ger-many.Media and conditions of cultivation. The enrichment, iso-
lation, and cultivation of toluene degraders were carried outin a defined, bicarbonate-buffered, sulfide-reduced saltwatermedium generally used for marine sulfate reducers; themedium contained high concentrations of Na2SO4 (4 g/liter),NaCl (20 g/liter), and MgCl2. 6H20 (3 g/liter) and a traceelement mixture chelated with sodium EDTA (34). Forroutine cultivation, sodium sulfide was added at a concen-tration of 1 mM; for experiments with resuspended cells, a
1444
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 21
Jan
uary
202
2 by
124
.219
.211
.107
.
TOLUENE OXIDATION BY A SULFATE-REDUCING BACTERIUM 1445
concentration of only 0.2 mM was applied. Sodium dithion-ite (15 to 20 mg/liter) was added as additional reductantbefore inoculation as described previously (34).
Toluene, ethylbenzene, and xylenes were filter sterilizedthrough solvent-resistant cellulose filters (pore size, 0.2 ,um).To avoid toxic effects, the aromatic hydrocarbons wereadded as dilute solutions (usually 2% [vol/vol]) in an inertlipophilic solvent. For routine cultivation, pharmaceutical-grade, nonfluorescent mineral oil (viscosity, 150 mPa/s;Lamotte, Bremen, Germany) served as the carrier phase; foran exact measurement of toluene consumption, the less-viscous 2,2,4,4,6,8,8-heptamethylnonane (Alfa Products,Karlsruhe, Germany) was used. Per liter of medium, 10 to 30ml of carrier phase was added. Stocks of sterile, deaeratedmineral oil or heptamethylnonane were stored asepticallyunder an N2 atmosphere as described previously (1). Poorlysoluble p-hydroxybenzyl alcohol and p-hydroxybenzalde-hyde were added from filter-sterilized 0.75 M solutions in2-propanol. 2-Propanol was neither utilized by nor inhibitoryto the bacteria at the given concentration. Other organicsubstrates were added from aqueous and autoclaved orfilter-sterilized stock solutions (34).Enrichment and routine cultivation on toluene (Fluka,
Neu-Ulm, Germany) were carried out in tubes (20 ml) or flatbottles (50 or 100 ml) containing a small gas phase (approx-imately one-eighth of the culture volume) of N2-CO2 (90:10[vol/vol]). Tubes and bottles were sealed with butyl or blackrubber stoppers, inverted, and incubated at an angle slightlyinclined from the horizontal on a rotary shaker at low speed(70 rpm); in this way, an optimal supply of the culture withtoluene from the carrier phase was guaranteed while itscontact with the stoppers was prevented (1). Incubationoccurred at 28°C in the dark.Chemicals were of analytical grade.Isolation. Toluene-degrading sulfate reducers were puri-
fied via repeated agar dilution series in culture tubes (160 by16 mm) as described previously (34). The solidified agarcolumn was covered with mineral oil (approximately 1 mlper tube) containing 1 to 2% (vol/vol) toluene; tubes wereanoxically sealed and incubated in an upright position at28°C. Isolated colonies were transferred to liquid medium.For purity control, cultures on toluene and benzoate were
examined routinely by phase-contrast microscopy. Further-more, cultures were transferred to medium containing 0.5 gof yeast extract per liter and 5 mM glucose or pyruvate andexamined microscopically.For maintenance, cells were grown on 4 mM benzoate,
stored at 4°C, and transferred every 4 to 6 weeks.Analysis of rRNA. Preparation of genomic DNA, in vitro
amplification of 16S rRNA genes, and direct sequencing ofamplified DNA were performed as described elsewhere (33).The sequenced rRNA gene fragments are homologous topositions 8 to 1542 of the Escherichia coli 16S rRNA gene(9). The 16S rRNA gene sequences were inserted into analignment of more than 900 homologous sequences frombacteria for comparative analysis. Distance matrices werecalculated from this alignment by using a newly developedprogram (SEQDIS [28a]). Phylogenetic trees were recon-structed by applying distance, maximum-parsimony, andmaximum-likelihood methods by using the programsNEIGHBOR and DNAPARS of the PHYLIP package (16)and fastDNAml (25a), respectively.
Determination of degradation balances and growth param-eters. For the measurement of degradation balances, diffu-sion of toluene from the medium into the stoppers wasminimized to the extent possible. This was achieved by using
special flat vials (1) with a small orifice (inner diameter, 9.5mm) relative to the volume (130 ml). Anoxic, reducedmedium (110 ml), inoculum (5 ml), and anoxic heptamethyl-nonane (3 ml) were added aseptically to the vials that werekept under an anoxic atmosphere. Vials were anoxicallysealed with butyl stoppers and inverted. Defined amounts oftoluene were then added through the stoppers by means ofsterile microliter syringes (1). Released toluene dropletsascended and dissolved in the carrier phase without cominginto contact with the stopper.Samples for chemical analyses were taken by sterile,
N2-flushed microliter syringes (1).The growth yield could not be determined directly by
weighing dried cells because removal of salts from the cellsby washing caused lysis. Therefore, cell dry mass had to bedetermined indirectly. The optical density of cultures grownon defined amounts of toluene was measured at 660 nm. Theratio of the optical density to the cell mass per volume wasdetermined once by centrifugation of a defined culturevolume and subsequent chemical oxidation (see below).
Chemical analyses. Sulfide was quantified by the methy-lene blue formation reaction in microassays as describedpreviously (1). In addition, a sulfide test with Cu2+ ionsyielding CuS was applied (10).
Toluene in the aqueous phase was measured by high-performance liquid chromatography (Sykam, Gilching/Mu-nich, Germany) on a Spherisorb OD S2 reverse-phase col-umn (5 by 250 mm) with 100% acetonitrile as the eluent andmonitored at 205 nm. Toluene in heptamethylnonane fromgrown cultures was determined by diluting aliquots in pureheptamethylnonane and measuring the absorbance at 261nm. The method was calibrated with defined, freshly pre-pared solutions of toluene in the solvent.The guanine-plus-cytosine content of the DNA was deter-
mined by thermal denaturation (24). E. coli K-12 served as a
reference.Cell dry mass was quantified by complete oxidation with
chromium(VI) (dichromic acid) yielding chromium(III) (25).The decrease in chromium(VI) is a direct measure of thereducing equivalents derived from a sample and, because ofthe relatively constant average oxidation state of cell carbon,is also a measure of cell mass. Cells from an original culturevolume of 20 to 100 ml were centrifuged, washed in 300 mMNa2SO4 to remove oxidizable chloride ions, and freeze-dried. The pellet was resuspended in 2 ml of a K2Cr2O7solution (2% [wt/wt]) in concentrated H2SO4 and heated inan aluminum block to 130°C for 30 min. The decrease inchromium(VI) was measured at 440 nm. The method was
calibrated with various amounts of yeast extract (c8 mg perassay).Measurement of catabolic activities in whole cells. All steps
for harvesting cells and preparing suspensions were carriedout inside an anoxic chamber. Centrifuged cells were resus-
pended in anoxic salt medium without organic substrate toobtain an approximately 10-fold cell density. Aliquots of 10ml were distributed to culture tubes (15 ml). Dithionite (15mg/liter) was added as described previously (34). To some ofthe tubes, rifampin in a solution in methanol (20 mg/ml) wasadded to a final concentration of 100 ,ug/ml. To the tubesused for experiments with toluene, 0.5 ml of mineral oil wasadded. After the tubes were sealed with butyl rubber stop-pers, the gas phase from the anoxic chamber was replacedby N2-CO2 (90:10). Organic compounds to be tested were
added via syringes; tubes with toluene were gently shaken as
described for routine cultures. Utilization of the applied
VOL. 59, 1993
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 21
Jan
uary
202
2 by
124
.219
.211
.107
.
APPL. ENvIRON. MICROBIOL.
FIG. 1. Phase-contrast photomicrograph of the newly isolatedsulfate-reducing bacterium, strain Tol2, grown on toluene. Bar, 10,um.
substrate was monitored by measuring the sulfide in aliquotstaken with microliter syringes.Enzyme assays. All steps for harvesting cells and prepara-
tion of cell extracts were performed inside an anoxic cham-ber or under gassing with N2. Cells from a 2-liter culturewere centrifuged, washed with anoxic salt medium, andresuspended in 2 ml of Tris-HCI buffer (100 mM, pH 7.5)containing 5.0 mM MgCl2- 6H20 and 2.5 mM dithioerythri-tol. The suspension was passed through an anoxic, refriger-ated French pressure cell at 1,000 lb/in2. After centrifugationat 30,000 x g for 15 min, the supernatant was stored underN2 at 4°C. Carbon monoxide dehydrogenase, formate dehy-drogenase, and 2-oxoglutarate dehydrogenase were assayedwith methyl viologen as the electron acceptor as describedpreviously (1).
Protein was determined by the method of Bradford (8)with bovine serum albumin as a standard.
Nucleotide sequence accession number. Data are depositedunder EMBL accession number X70953.
RESULTS
Enrichment and isolation. Anoxic saltwater medium (28mM sulfate) dispensed in 100-ml bottles was inoculated withmarine sediment slurry (approximately 5 ml). The mediumwas covered with mineral oil containing 5 or 15% (vol/vol)toluene and incubated at 28°C. Cultures were briefly shakenonce per week. After 8 weeks, 15 mM sulfide had beenformed in the presence of 5% toluene in the carrier phase.Controls with mineral oil alone never yielded more than 2mM sulfide. With 15% toluene, even less sulfide had beenproduced than that in the control. In consideration of theobvious toxicity of toluene, concentrations lower than 5% inthe carrier phase were tested in subcultures. Indeed, theydeveloped faster with less toluene. For further cultivation, acarrier phase with 2% toluene was used.The second subculture of the enrichment served as start-
ing inoculum for an agar dilution series. After 14 days, ochreto yellowish-brown colonies could be recognized. Coloniesnext to the carrier phase were largest in size (diameter, 1mm). After a second dilution series, colonies were trans-ferred to liquid medium and incubated at 28°C. Growthoccurred within 5 to 8 days on toluene as well as benzoate.Of four isolates, one strain (Tol2) was selected for furtherstudies.
Purity controls in media with various organic compoundsalways revealed the same cell type.
Morphological, physiological, and other characteristics.Cells of strain Tol2 (Fig. 1) were oval to nearly coccoid (1.2
to 1.4 ,um by 1.2 to 2.0 ,um) and sometimes motile. The cellsoccasionally adhered to the glass surface of culture vessels.Slime capsules were detected under the light microscopeafter negative staining.
Strain Tol2 was able to oxidize various classes of organicsubstrates. Compounds were tested as electron donors forgrowth and sulfate reduction by Tol2. Each compound wastested twice, at either two identical or two different concen-trations. In addition to toluene (2% in mineral oil), the isolateused other aromatic compounds, some short-chain alcohols,and a number of aliphatic acids (specifically, p-cresol [1mM], benzaldehyde [1 mM], benzoate [2 and 6 mM], phe-nylacetate [2 and 4 mM], p-hydroxybenzaldehyde [1 mM],p-hydroxybenzoate [1 mM], ethanol [10 mM], 1-propanol [5mM], 1-butanol [5 mM], butyrate [5 mM], pyruvate [10mM],succinate [5 mM], fumarate [5 mM], malate [5 mM], andglutarate [2 mM]). The compounds tested that were notutilized by Tol2 for growth or sulfate reduction includedcyclohexane, methylcyclohexane, benzene, o-xylene, m-xy-lene, p-xylene, ethylbenzene (all 1 and 2% in mineral oil);benzyl alcohol (0.5 and 1 mM), phenol (0.5 and 2 mM),o-cresol (0.5 and 1 mM), m-cresol (0.5 and 1 mM), p-hy-droxybenzyl alcohol (0.5 and 1 mM), o-methylbenzoate (0.2and 2 mM), m-methylbenzoate (0.2 and 2 mM), p-methyl-benzoate (0.2 and 2 mM), phenylpropionate (0.2 and 4 mM),H2 (with CO2 and 1 mM acetate), methanol (10 mM),2-propanol (5 mM), formate (10 mM, with 1 mM acetate),acetate (10 mM), propionate (5 and 10 mM), isobutyrate (5mM), valerate (5 mM), 2-methylbutyrate (3 mM), 3-methyl-butyrate (3 mM), caproate (2 mM), caprylate (0.5 mM),palmitate (0.5 mM), and lactate (10 mM). (Growth on bu-tyrate occurred only after 7 weeks. Because of poor solubil-ity in water, p-hydroxybenzyl alcohol was added from a 0.75M solution in 2-propanol, which was neither utilized by norinhibitory to the bacteria.)
If toluene was dissolved directly in the medium in theabsence of a carrier phase, growth was inhibited by concen-trations higher than 0.5 mmol/liter. In mineral oil as thecarrier phase (30 ml/liter of medium), 2% (vol/vol) tolueneyielded an equilibrium concentration in the aqueous phase ofabout 0.25 mmol/liter.The optimal growth temperature was 28°C, and the opti-
mal pH was between 7.0 and 7.1. Growth of strain Tol2depended on high concentrations of NaCl (340 mM) andMgCl2. 6H20 (15 mM) and a supplementation with vita-mins. Initial growth of freshly inoculated cultures was im-proved by applying dithionite as additional reductant. Theshortest doubling time on toluene was 27 h.
Strain Tol2 was highly sensitive to 02-Thermal denaturation of DNA revealed a guanine-plus-
cytosine content of 42 mol%.Natural relationships to other sulfate reducers. Relation-
ships derived from 16S rRNA sequencing are depicted inFig. 2.
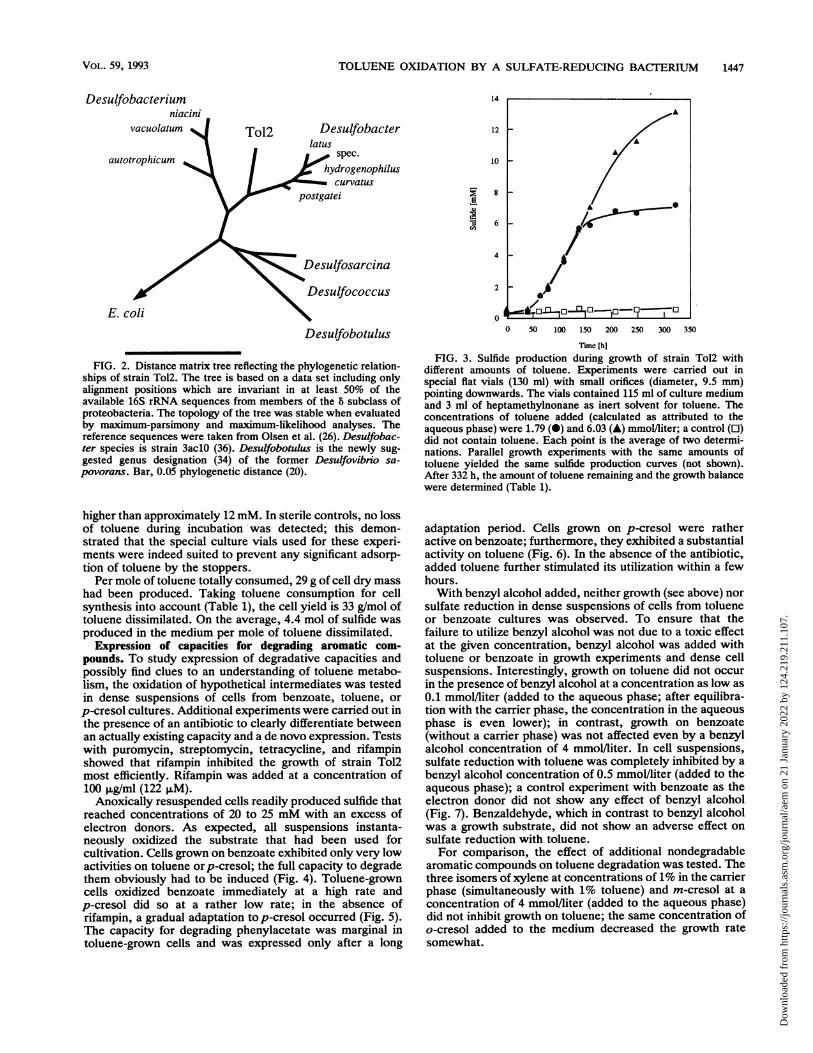
Degradation balance and growth yield. The degradationbalance was determined with low and high concentrations oftoluene (1.79 and 6.03 mmol/liter, respectively, calculated asattributed to the aqueous phase) dissolved in a carrier phase.Production of sulfide was monitored during incubation (Fig.3), and the toluene remaining in the aqueous and hydropho-bic phases was measured after 332 h (Table 1). Tolueneadded at a low concentration had almost completely van-ished. In the experiment with the higher toluene concentra-tion, almost two-thirds had disappeared. A comparison ofsulfide production curves (Fig. 3) suggests that growth andtoluene oxidation are impeded by sulfide at concentrations
1446 RABUS ET AL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 21
Jan
uary
202
2 by
124
.219
.211
.107
.
TOLUENE OXIDATION BY A SULFATE-REDUCING BACTERIUM 1447
Desulfobacteriumniacini
14
12
10
E
c.8
Desulfobotulus
FIG. 2. Distance matrix tree reflecting the phylogenetic relation-ships of strain Tol2. The tree is based on a data set including onlyalignment positions which are invariant in at least 50% of theavailable 16S rRNA sequences from members of the 8 subclass ofproteobacteria. The topology of the tree was stable when evaluatedby maximum-parsimony and maximum-likelihood analyses. Thereference sequences were taken from Olsen et al. (26). Desulfobac-ter species is strain 3aclO (36). Desulfobotulus is the newly sug-gested genus designation (34) of the former Desulfovibrio sa-povorans. Bar, 0.05 phylogenetic distance (20).
higher than approximately 12 mM. In sterile controls, no lossof toluene during incubation was detected; this demon-strated that the special culture vials used for these experi-ments were indeed suited to prevent any significant adsorp-tion of toluene by the stoppers.
Per mole of toluene totally consumed, 29 g of cell dry masshad been produced. Taking toluene consumption for cellsynthesis into account (Table 1), the cell yield is 33 g/mol oftoluene dissimilated. On the average, 4.4 mol of sulfide wasproduced in the medium per mole of toluene dissimilated.
Expression of capacities for degrading aromatic com-pounds. To study expression of degradative capacities andpossibly find clues to an understanding of toluene metabo-lism, the oxidation of hypothetical intermediates was testedin dense suspensions of cells from benzoate, toluene, orp-cresol cultures. Additional experiments were carried out inthe presence of an antibiotic to clearly differentiate betweenan actually existing capacity and a de novo expression. Testswith puromycin, streptomycin, tetracycline, and rifampinshowed that rifampin inhibited the growth of strain Tol2most efficiently. Rifampin was added at a concentration of100 ,ug/ml (122 jLM).Anoxically resuspended cells readily produced sulfide that
reached concentrations of 20 to 25 mM with an excess ofelectron donors. As expected, all suspensions instanta-neously oxidized the substrate that had been used forcultivation. Cells grown on benzoate exhibited only very lowactivities on toluene orp-cresol; the full capacity to degradethem obviously had to be induced (Fig. 4). Toluene-growncells oxidized benzoate immediately at a high rate andp-cresol did so at a rather low rate; in the absence ofrifampin, a gradual adaptation to p-cresol occurred (Fig. 5).The capacity for degrading phenylacetate was marginal intoluene-grown cells and was expressed only after a long
8
6
0 50 100 150 200 250 300 350
Time [h]FIG. 3. Sulfide production during growth of strain Tol2 with
different amounts of toluene. Experiments were carried out inspecial flat vials (130 ml) with small orifices (diameter, 9.5 mm)pointing downwards. The vials contained 115 ml of culture mediumand 3 ml of heptamethylnonane as inert solvent for toluene. Theconcentrations of toluene added (calculated as attributed to theaqueous phase) were 1.79 (0) and 6.03 (A) mmol/liter; a control (0)did not contain toluene. Each point is the average of two determi-nations. Parallel growth experiments with the same amounts oftoluene yielded the same sulfide production curves (not shown).After 332 h, the amount of toluene remaining and the growth balancewere determined (Table 1).
adaptation period. Cells grown on p-cresol were ratheractive on benzoate; furthermore, they exhibited a substantialactivity on toluene (Fig. 6). In the absence of the antibiotic,added toluene further stimulated its utilization within a fewhours.With benzyl alcohol added, neither growth (see above) nor
sulfate reduction in dense suspensions of cells from tolueneor benzoate cultures was observed. To ensure that thefailure to utilize benzyl alcohol was not due to a toxic effectat the given concentration, benzyl alcohol was added withtoluene or benzoate in growth experiments and dense cellsuspensions. Interestingly, growth on toluene did not occurin the presence of benzyl alcohol at a concentration as low as0.1 mmol/liter (added to the aqueous phase; after equilibra-tion with the carrier phase, the concentration in the aqueousphase is even lower); in contrast, growth on benzoate(without a carrier phase) was not affected even by a benzylalcohol concentration of 4 mmolfliter. In cell suspensions,sulfate reduction with toluene was completely inhibited by abenzyl alcohol concentration of 0.5 mmol/liter (added to theaqueous phase); a control experiment with benzoate as theelectron donor did not show any effect of benzyl alcohol(Fig. 7). Benzaldehyde, which in contrast to benzyl alcoholwas a growth substrate, did not show an adverse effect onsulfate reduction with toluene.For comparison, the effect of additional nondegradable
aromatic compounds on toluene degradation was tested. Thethree isomers of xylene at concentrations of 1% in the carrierphase (simultaneously with 1% toluene) and m-cresol at aconcentration of 4 mmol/liter (added to the aqueous phase)did not inhibit growth on toluene; the same concentration ofo-cresol added to the medium decreased the growth ratesomewhat.
VOL. 59, 1993
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 21
Jan
uary
202
2 by
124
.219
.211
.107
.
1448 RABUS ET AL.
TABLE 1. Quantification of toluene consumption and sulfate reduction to sulfide during growth of strain Tol2a
Toluene concn (mmol/liter)
Total added' Present in the Present in the Total utilized Sulfide Molar ratio ofExpt (calculated as aqueous phase carrier ihne (calculated as producedd dissimilation
attributed to ae pas attributed to Dissimilated (mmoliter) (sulfide/toluene)after 332 h of after 332 h of atiue o Dsiiaec (mlltr slietleeaqueous incubation aqueousphase) incubation phase)
Strain Tol2 with small 1.79 0.00 0.02 1.77 1.53 7.1 4.64amt of toluene
Strain Tol2 with large 6.03 0.10 2.26 3.67 3.18 13.1 4.12amt of toluene
Strain Tol2 without 0.00 0.00 0.0toluene (control)
Sterile medium with 1.90 0.10 1.80 0.00 0.0toluene (control)
a The cultures used to monitor sulfide production (Fig. 3) were analyzed at the end of growth. The data shown are average values from two parallel cultures.b Supplied from a carrier phase of heptamethylnonane.c Calculated from produced cell dry mass assuming the following assimilation equation (36): 17C6H5CH3 + 25HCO3 + 25H+ + 33H20 736C4H703; thus,
1 mg of dry mass requires 0.00458 mmol of toluene as the carbon source.d Difference between sulfide concentrations at the beginning (from medium and inoculum) and at the end of growth.
Key enzymes of acetyl-CoA oxidation. In cell extract,carbon monoxide dehydrogenase and formate dehydroge-nase were detected; their specific activities (per mass ofprotein) were 0.72 and 0.37 Fimol min-' mg-', respec-tively. No activity of 2-oxoglutarate dehydrogenase wasfound.
Usability of toluene by other sulfate reducers. The capacityfor toluene oxidation was also tested with D. anilini, D.phenolicum, and D. multivorans, which had been originallyisolated on aromatic compounds (5, 27, 34). After 6 weeks ofincubation, D. phenolicum exhibited poor growth whereasthe other species did not grow at all. Growth of controlscontaining both toluene and benzoate verified that the tolu-ene concentration (2% in mineral oil) was not toxic.
25
20 _
r 15
A 10
la
0
0 10 20 30 40 50
25
0 10 20 30 40 50
Time [h]
FIG. 4. Sulfate reduction with aromatic compounds measured indense cell suspensions (3.7 to 5.6 mg of cell dry mass per ml) ofstrain Tol2 grown on benzoate. (A) Suspensions without antibiotic;(B) suspensions with added rifampin (100 ,ug/ml) to inhibit de novo
expression of degradative capacities. Symbols for compounds tested(concentrations given in parentheses): *, benzoate (2 mmol/liter);0, toluene (10 mmol/liter, calculated as attributed to the aqueous
phase), supplied from carrier phase; A,p-cresol (2.9 mmol/liter); 0,
no substrate added. Each point is the average of two determinations.In a second experiment with cells from a separately grown benzoateculture, the same expression of activities was observed.
DISCUSSION
The newly isolated marine strain Tol2 is the first pureculture of a sulfate-reducing bacterium that has been shownto degrade an aromatic hydrocarbon. Even though toluenecould be readily utilized, it was relatively toxic to strainTol2. The sulfate reducer did not tolerate more than 0.5mmol of toluene per liter, which is less than 7% of asaturated solution in water (around 7.3 mM at 23.5°C) (11).Some nitrate-reducing toluene degraders tolerate approxi-mately 1 mM toluene (2, 28); one strain tolerates a concen-
30 30 B /
eT2020
stai To2gono0oun.()Sspninihuniitc
a t 10Aot0
20~~~v
Tie[h]
FIG. 5. Sulfate reduction with aromatic compounds measured indense cell suspensions (3.7 to 5.6 mg of cell dry mass per ml) ofstrain Tol2 grown on toluene. (A) Suspensions without antibiotic;(B) suspensions with added rifampin (100 p.glml) to inhibit de novoexpression of degradative capacities. Symbols for compounds tested(concentrations given in parentheses): o,toluene (10 mmol/liter,calculated as attributed to the aqueous phase), supplied from carrierphase; U, benzoate (2 mmol/liter); A, p-cresol (2.9 mmol/liter); V,phenylacetate (1 mmollliter); 0, no substrate added. The highestvalues for sulfide produced with toluene (arrow) fell outside the panel.Sulfide produced with phenylacetate was finally measured after 98 h(circled symbols). Each point is the average of two measurements. Ina second experiment with cells from a separately grown tolueneculture, the same expression of activities was observed.
A B20
15-U-.
.E
10 _
I1 IU5/z*\ A~. . .
APPL. ENvIRON. MICROBIOL.
_
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 21
Jan
uary
202
2 by
124
.219
.211
.107
.
TOLUENE OXIDATION BY A SULFATE-REDUCING BACTERIUM 1449
30 30
A B
20 20
:510 j10
000 *~~~~~~~~~~~~~~~~~~0 0
0 10 20 30 40 50 0 10 20 30 40 50
Time [h]
FIG. 6. Sulfate reduction with aromatic compounds measured indense cell suspensions (3.7 to 5.6 mg of cell dry mass per ml) ofstrain Tol2 grown on p-cresol. (A) Suspensions without antibiotic;(B) suspensions with added rifampin (100 ,ug/ml) to inhibit de novo
expression of degradative capacities. Symbols for compounds tested(concentrations given in parentheses): A, p-cresol (2.9 mmol/liter);U, benzoate (2 mmol/liter); *, toluene (10 mmol/liter, calculated as
attributed to the aqueous phase), supplied from carrier phase; 0, no
substrate added. The highest values for sulfide produced withtoluene (arrow) fell outside the panel. Each point is the average oftwo measurements.
tration as high as 3 mM (15). Mineral oil or heptamethyl-nonane applied as inert solvent allowed strain Tol2 to becontinuously supplied with toluene while its activity and,therefore, its concentration in the medium were simulta-neously kept low. Carrier phases of hexadecane and otheralkanes have been applied previously for the cultivation ofmicroorganisms on relatively toxic hydrocarbons (for exam-ples, see references 15 and 18).The measured degradation balance (Table 1) is in good
agreement with the following stoichiometry: C6H5CH3 +4.5So42- + 2H+ + 3H20 -- 7HC03- + 4.5H2S (AG' =
-273 kJ/mol of toluene).The free energy is calculated from standard values (11, 29)
for activities of dissolved compounds of 10-2, which in thecase of toluene corresponds to approximately 7 x 10-5mol/liter in water (1% of a saturated solution). It can beinferred from enzymes measured in cell extracts that strainTol2 oxidizes acetyl-CoA via the carbon monoxide dehy-drogenase pathway. This pathway is common to completelyoxidizing sulfate reducers except for Desulfobacter species(1, 35). Despite its capacity for oxidizing organic substratescompletely to CO2, strain Tol2 did not grow on acetate.Several species of completely oxidizing sulfate reducers thatpossess the carbon monoxide dehydrogenase pathway growpoorly or are even unable to grow on free acetate (1, 34, 35).The activation reaction of toluene in the newly isolated
sulfate-reducing bacterium remains unknown. The presentstudies leave open the possibility that toluene is oxidativelycondensed with acetyl-CoA to yield phenylpropionyl-CoAas suggested elsewhere (7, 14). Since free phenylpropionatedoes not appear on this pathway, the failure of strain Tol2 togrow with this compound does not argue against such a
mechanism. Nevertheless, the expression of degradativecapacities measured with whole cells allows some state-ments to be made as to which of the formerly discussed,hypothetical starting reactions are unlikely to occur. Tolu-
30 _
-
E
E-6
r-
c'n
20 _
° 1 '0 5 10 15 20 25 30 35 0 5 10 15 20 25 30 35
Time [h]
FIG. 7. Effect of benzyl alcohol and benzaldehyde on sulfatereduction to sulfide in dense cell suspensions (3.7 to 5.6 mg of celldry mass per ml) with toluene and benzoate as electron donors.Cells had been harvested from cultures grown on toluene or benzo-ate, respectively. Benzyl alcohol (0.5 mmolJliter) and benzaldehyde(0.5 mmol/liter) were added after 5.5 h (arrow). (A) Sulfate reductionwith toluene (10 mmollliter, calculated as attributed to the aqueous
phase; supplied from the carrier phase) without further additions(@), with addition of benzyl alcohol (V), and with addition ofbenzaldehyde (C1). (B) Sulfate reduction with benzoate (2 mmol/liter) without further additions (-), with addition of benzyl alcohol(V), and with addition of benzaldehyde (O); a control experiment(0) was carried out without organic substrate. Each point reflectsthe average of two measurements.
ene-grown cells were not adapted to phenylacetate; o-, m-,and p-methylbenzoate were not growth substrates at all.Therefore, carboxylation of the methyl group or the ring as
a starting reaction to yield a free carboxylate appears doubt-ful if one assumes that the tested compounds could enter thecell as benzoate does. The same conclusion was drawn fromexperiments with denitrifying strains (2, 28). Even thoughp-cresol was, in principle, a good growth substrate, it wasutilized very slowly by toluene-grown cells of strain Tol2(Fig. 5). This observation as well as the failure of the newisolate to grow on other cresols raises doubts about a
degradation of toluene via the introduction of a hydroxylgroup into the ring. Similarly, results of adaptation studieswith a denitrifier were not consistent with a conversion oftoluene to a cresol (2). As for degradation of toluene viabenzyl alcohol, observations are paradoxical at present. Thetoluene-degrading denitrifying strains GS-15 (23) and K 172(2, 28, 30) are able to grow with benzyl alcohol. With thelatter, the conversion of toluene to benzyl alcohol andbenzaldehyde has been observed (3). In contrast, the deni-trifying strains Ti (15) and T (28) and the newly isolatedsulfate reducer strain Tol2 were unable to grow with benzylalcohol; toluene oxidation by the latter was even specificallyinhibited by the alcohol (Fig. 7). This may support thehypothesis that the methyl group of toluene, rather than thering, is the site of attack in the initial reaction which might beimpeded by benzyl alcohol.Taxonomic affiliation of the newly isolated sulfate reducer
with an existing species or genus would lead to inconsisten-cies. Genealogically, strain Tol2 is most closely related toDesulfobacter species (Fig. 2). Physiologically, however,there are marked differences. Desulfobacter species are
rather specialized acetate oxidizers that make use of the
A30 _
B
20--°
10 U
. --0.'o__oo-o0 PL O.
VOL. 59, 1993
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 21
Jan
uary
202
2 by
124
.219
.211
.107
.
1450 RABUS ET AL.
citric acid cycle. The genealogically less closely relatedDesulfobactenum species Desulfococcus and Desulfo-sarcina (Fig. 2) are metabolically more similar to strain Tol2.From other sulfate reducers isolated on aromatic compoundsand described as D. anilini (27), Desulfobactenum indolicum(4), and D. phenolicum (5), sequences of 16S rRNA are notavailable to date. In view of the phylogenetic distances thatexist between other genera, strain Tol2 must be regarded as
a species of a new genus. The designation Desulfobaculatoluolica is proposed.
Description of Desulfobacula gen. nov. Desulfobacula(De.sul.fo.ba' cu.la. L.n. bacula dem. of baca, berry, olive-like fruit; Desulfobacula, sulfate-reducing small berry). Ovalto sometimes coccoid cells that may be motile are charac-teristic. They are strict anaerobes, using sulfate as theterminal electron acceptor that is reduced to sulfide. Aro-matic compounds and a number of low-molecular-weightaliphatic acids and alcohols may be utilized as electrondonors and carbon sources. Electron donors are completelyoxidized to CO2 via the carbon monoxide dehydrogenasepathway. Desulfobacula belongs to the 8 subclass of theproteobacteria; the closest relatives are Desulfobacter spe-
cies.Description of Desulfobacula toluolica sp. nov. Desulfobac-
ula toluolica (tol.u.o' li.ca. L. adj. toluolica, pertaining totoluene). Cells are 1.2 to 1.4 ,um in diameter and 1.2 to 2.0,um in length. Motility may occur. NaCl, MgCl2, and vita-mins are required for growth. Toluene, benzaldehyde, ben-zoate, phenylacetate, p-cresol, p-hydroxybenzaldehyde,p-hydroxybenzoate, glutarate, pyruvate, succinate, fumar-ate, malate, butyrate, ethanol, 1-propanol, and 1-butanolserve as electron donors and carbon sources. Benzyl alco-hol, o-, m-, and p-methylbenzoate, o- and m-cresol, lactate,and H2 are not utilized. The optimum pH is 7.0 to 7.1; theoptimum temperature is 28°C. The guanine-plus-cytosinecontent of the DNA is 42 mol% (by thermal denaturation).Habitat is anoxic marine sediment rich in organic com-
pounds.The strain is deposited in the Deutsche Sammlung von
Mikroorganismen und Zellkulturen GmbH under number7467.
ACKNOWLEDGMENTS
We thank S. Trimkowski and F. Aeckersberg for experimentalhelp. The initial enrichment studies were made possible during themicrobiology summer course at the Marine Biological Laboratory,Woods Hole, Mass., in 1989.
This work was supported by the Deutsche Forschungsgemein-schaft and the Fonds der Chemischen Industrie.
REFERENCES1. Aeckersberg, F., F. Bak, and F. Widdel. 1991. Anaerobic
oxidation of saturated hydrocarbons to CO2 by a new type ofsulfate-reducing bacterium. Arch. Microbiol. 156:5-14.
2. Altenschmidt, U., and G. Fuchs. 1991. Anaerobic degradation oftoluene in denitrifying Pseudomonas sp.: indication of toluenemethylhydroxylation and benzoyl-CoA as central aromatic in-termediate. Arch. Microbiol. 156:152-158.
3. Altenschmidt, U., and G. Fuchs. 1992. Anaerobic toluene oxi-dation to benzyl alcohol and benzaldehyde in a denitrifyingPseudomonas strain. J. Bacteriol. 174:4860-4862.
4. Bak, F., and F. Widdel. 1986. Anaerobic degradation of indoliccompounds by sulfate-reducing enrichment cultures, and de-
scription of Desulfobactenium indolicum gen. nov., sp. nov.
Arch. Microbiol. 146:170-176.5. Bak, F., and F. Widdel. 1986. Anaerobic degradation of phenol
and phenol derivatives by Desulfobacterium phenolicum sp.
nov. Arch. Microbiol. 146:177-180.
6. Beller, H. R., D. Grbic-Galic, and M. Reinhard. 1992. Microbialdegradation of toluene under sulfate-reducing conditions andthe influence of iron on the process. Appl. Environ. Microbiol.58:786-793.
7. Beller, H. R., M. Reinhard, and D. Grbic-Galic. 1992. Metabolicby-products of anaerobic toluene degradation by sulfate-reduc-ing enrichment cultures. Appi. Environ. Microbiol. 58:3192-3195.
8. Bradford, M. M. 1976. A rapid and sensitive method for thequantitation of microgram quantities of protein utilizing theprinciple of protein-dye binding. Anal. Biochem. 72:248-254.
9. Brosius, J., T. J. Dull, D. D. Sleeter, and H. F. Noller. 1981.Gene organization and primary structure of a ribosomal RNAoperon from Escherichia coli. J. Mol. Biol. 148:107-127.
10. Cord-Ruwisch, R. 1985. A quick method for the determinationof dissolved and precipitated sulfides in cultures of sulfate-reducing bacteria. J. Microbiol. Methods 4:33-36.
11. Dean, J. A. 1992. Lange's handbook of chemistry. McGraw-HillBook Co., New York.
12. Dolfing, J., P. Zeyer, P. Binder-Eicher, and R. P. Schwarzen-bach. 1990. Isolation and characterisation of a bacterium thatmineralizes toluene in the absence of molecular oxygen. Arch.Microbiol. 154:336-341.
13. Edwards, E. A., and D. Grbic.Galic. 1992. Complete mineral-ization of benzene by aquifer microorganisms under strictlyanaerobic conditions. Appl. Environ. Microbiol. 58:2663-2666.
14. Evans, P. J., W. Ling, B. Goldschmidt, E. R. Ritter, and L. Y.Young. 1992. Metabolites formed during anaerobic transforma-tion of toluene and o-xylene and their proposed relationship tothe initial steps of toluene mineralization. Appl. Environ. Mi-crobiol. 58:496-501.
15. Evans, P. J., D. T. Mang, K. S. Kim, and L. Y. Young. 1991.Anaerobic degradation of toluene by a denitrifying bacterium.Appl. Environ. Microbiol. 57:1139-1145.
16. Felsenstein, J. 1982. Numerical methods for inferring phyloge-netic trees. Quart. Rev. Biol. 57:379-404.
17. Gibson, D. T., and V. Subramanian. 1984. Microbial degrada-tion of aromatic hydrocarbons, p. 181-252. In D. T. Gibson(ed.), Microbial degradation of aromatic compounds. MarcelDekker, Inc., New York.
18. Gill, C. O., and C. Ratledge. 1972. Toxicity of n-alkanes,n-alk-1-enes, n-alkan-1-ols and n-alkyl-1-bromides towardsyeasts. J. Gen. Microbiol. 72:165-172.
19. Grbic-Galic, D., and T. M. Vogel. 1987. Transformation oftoluene and benzene by mixed methanogenic cultures. Appl.Environ. Microbiol. 61:491-497.
20. Jukes, T. H., and C. R. Cantor. 1969. Evolution of proteinmolecules, p. 21-132. In N. H. Murano (ed.), Mammalianprotein metabolism. Academic Press, Inc., New York.
21. Juittner, F., and J. J. Henatsch. 1986. Anoxic hypolimnion is asignificant source of biogenic toluene. Nature (London) 323:797-798.
22. Kuhn, E. P., J. Zeyer, P. Eicher, and R. P. Schwarzenbach.1988. Anaerobic degradation of alkylated benzenes in denitrify-ing laboratory columns. Appl. Environ. Microbiol. 54:490-496.
23. Lovley, D. R., and D. J. Lonergan. 1990. Anaerobic oxidation oftoluene, phenol, andp-cresol by the dissimilatory iron-reducingorganism GS-15. Appl. Environ. Microbiol. 56:1858-1864.
24. Marmur, J., and P. Doty. 1962. Determination of the basecomposition of deoxyribonucleic acid from its thermal denatur-ation temperature. J. Mol. Biol. 5:109-118.
25. Marr, L., M. S. Cresser, and L. J. Ottendorfer. 1988. Um-weltanalytik, p. 177-178. Thieme, Stuttgart, Germany.
25a.Olsen, G. J. Personal communication.26. Olsen, G. J., R. Overbeck, N. Larsen, T. L. Marsh, M. J.
McCaughey, M. J. Maciukenas, W. M. Kuan, T. J. Macke, Y.Xing, and C. R. Woese. 1992. The ribosomal database project.Nucleic Acids Res. 20(Suppl.):2199-2200.
27. Schnell, S., F. Bak, and N. Pfennig. 1989. Anaerobic degradationof aniline and dihydroxybenzenes by newly isolated sulfate-reducing bacteria and description of Desulfobactenum anilini.Arch. Microbiol. 152:556-563.
28. Schocher, R. J., B. Seyfried, F. Vazquez, and J. Zeyer. 1991.
APPL. ENVIRON. MICROBIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 21
Jan
uary
202
2 by
124
.219
.211
.107
.
TOLUENE OXIDATION BY A SULFATE-REDUCING BACTERIUM 1451
Anaerobic degradation of toluene by pure cultures of denitrify-ing bacteria. Arch. Microbiol. 157:7-12.
28a.Stuckmann, N., and W. Ludwig. Unpublished data.29. Thauer, R. K., K. Jungermann, and K. Decker. 1977. Energy
conservation in chemotrophic anaerobic bacteria. Bacteriol.Rev. 41:100-180.
30. Tschech, A., and G. Fuchs. 1987. Anaerobic degradation ofphenol by pure cultures of newly isolated denitrifyingpseudomonads. Arch. Microbiol. 148:213-217.
31. U.S. Public Health Service. 1989. Toxicological profile for tolu-ene. Publication ATSDR/TP-89/23. Agency for Toxic Sub-stances and Disease Registry, U.S. Public Health Service,Atlanta.
32. Vogel, T. M., and D. Grbic-Galic. 1986. Incorporation of oxygenfrom water into toluene and benzene during anaerobic fermen-tative transformation. Appl. Environ. Microbiol. 52:200-202.
33. Weizenegger, M., M. Neumann, E. Stackebrandt, and W. Lud-
34. Widdel, F., and F. Bak. 1992. Gram-negative mesophilic sulfate-reducing bacteria, p. 3352-3378. In A. Balows, H. G. Truper,M. Dworkin, W. Harder, and K. H. Schleifer (ed.), Theprokaryotes, 2nd ed., vol. 4. Springer-Verlag, New York.
35. Widdel, F., and T. A. Hansen. 1992. The dissimilatory sulfate-and sulfur-reducing bacteria, p. 583-624. In A. Balows, H. G.Truper, M. Dworkin, W. Harder, and K. H. Schleifer (ed.), Theprokaryotes, 2nd ed., vol. 1. Springer-Verlag, New York.
36. Widdel, F., and N. Pfennig. 1981. Studies on dissimilatorysulfate-reducing bacteria that decompose fatty acids. I. Isola-tion of new sulfate-reducing bacteria enriched with acetate fromsaline environments. Description of Desulfobacter postgateigen. nov., sp. nov. Arch. Microbiol. 129:395-400.


















![Biogeochemical cycling in anoxic sediments · 2007-04-20 · for sulfate reduction and methanogenesis. Hydrogen concentrations respond to T, [X], pH. Laboratory changes correspond](https://static.documents.pub/doc/80x56/5f478b2e8fcaea602546f86a/biogeochemical-cycling-in-anoxic-sediments-2007-04-20-for-sulfate-reduction-and.jpg)





