UNIVERSITE PARIS-NORD UFR LETTRES, SCIENCES DE L’HOMME ET DES SOCIETES DIPLOME D’ETUDES APPROFONDIES DE BIOLOGIE DU COMPORTEMENT (Responsable : Professeur C. BAUDOIN) COMPORTEMENT PREDATEUR DES MERLES (TURDUS MERULA) : CONTRIBUTION A LA SELECTION FREQUENCE DEPENDANTE. Par Nicolas BUSQUET Behavioural and Evolutionary Ecology Research group (BEER) University of Southampton (UK) Responsable : Dr J. A. ALLEN Septembre 1999
Transcript
UNIVERSITE PARIS-NORD UFR LETTRES, SCIENCES DE L’HOMME ET DES SOCIETES
DIPLOME D’ETUDES APPROFONDIES DE BIOLOGIE DU COMPORTEMENT
(Responsable : Professeur C. BAUDOIN)
COMPORTEMENT PREDATEUR DES MERLES (TURDUS MERULA) : CONTRIBUTION A LA SELECTION FREQUENCE DEPENDANTE.
Par Nicolas BUSQUET Behavioural and Evolutionary Ecology Research group (BEER) University of Southampton (UK) Responsable : Dr J. A. ALLEN
Septembre 1999
Table des matières
TABLE DES MATIERES ........................................................................................................................... 1
3.1 EXPERIENCES SANS OBSERVATION ................................................................................................ 12 3.2 EXPERIENCES AVEC OBSERVATION................................................................................................ 13
3.2.1 La population prédatrice .......................................................................................................... 13 3.2.2 Analyse du comportement des merles (Turdus merula)............................................................ 14
3.2.2.1 Site 1 ................................................................................................................................................ 14 3.2.2.2 Site 2 ................................................................................................................................................ 17 3.2.2.3 Site 5 ................................................................................................................................................ 19
4.1 LA POPULATION PREDATRICE ........................................................................................................ 22 4.2 LE COMPORTEMENT PREDATEUR DES MERLES............................................................................... 22
Chaque visite est consacrée à un seul type de proie. La sommation de ces
actions différentes aboutit à une surprédation du type rare.
Sélection au niveau populationnelSélection au niveau individuel
pr
élev
é
nom
bre
pr
élev
é
nom
bre
Les proies rares sont prélevés de façon excessive à chaque visite. Tous les
individus effectuent une sélection anti-apostatique.
Tableau 1 a. et b. : présentation des deux hypothèses théoriques extrêmes de réalisation d'une sélection fréquence dépendante anti-apostatique.
Ces deux hypothèses de départ représentent les deux modalités extrêmes
ableau 1). Le comportement des oiseaux peut se situer n'importe où sur le continuum
ntre ces deux hypothèses. La sélection est testée en comparant les visites observées avec
es visites fictives correspondant à une prédation aléatoire. L'explication proximale de ce
u ces comportements est envisagé en utilisant une démarche modélisatrice.
3
2 Matériel et méthodes 2.1 Matériel biologique
Les oiseaux étudiés sont les passereaux que l’on peut trouver dans les parcs et
jardins d'Europe. Les espèces observées sont le merle noir (Turdus merula), la grive
musicienne (Turdus Philomenos), la mésange bleue (Parus caeruleus), la mésange
charbonnière (Parus major), le rouge-gorge (Erythacus rubecula), le verdier (Carduelis
chloris), et l’étourneau sansonnet (Sturnus vulgaris). Cette guilde de passeraux constitue
la population prédatrice susceptible d’effectuer une sélection sur une population proie
composée de vers. Ces oiseaux sont étudiés par observation, sans aucune manipulation.
2.2 Dispositif expérimental
2.2.1 Les proies
Les proies utilisées sont de même taille, soit rouges, soit jaunes. Ces appâts sont
fabriqués en mélangeant 750 g de farine et 250 g de saindoux. A cette pâte est ajouté un
colorant alimentaire selon les proportions suivantes : 20 ml de colorant « strawberry red »
(code K6020) pour les proies rouges et 16 ml de colorant « egg yellow » (code K6013)
pour les proies jaunes. La pâte ainsi colorée est ensuite transformée en cylindres de 6 mm
de diamètre par le biais d’un appareil electro-ménager modifié. Les cylindres sont alors
alignés et découpés pour donner des proies de 7 mm de long.
2.2.2 Conditions expérimentales
Dans toutes les expériences, 200 proies sont présentées: 180 du type commun, et
20 du type rare, de facon à obtenir un ratio 9:1 en début de l’expérience. Les deux
couleurs sont utilisées alternativement en tant que proies communes ou rares. Le support
sur lequel sont disposées les proies est une plaque de bois de 45 cm de côté. Les proies
sont disposées aléatoirement à l’interieur d’un cercle central de 20 cm de diamètre. La
densité de proies ainsi obtenue est de 7440 m2.
2.3 Protocoles
Chaque expérience se subdivise en douze séances. Pour chaque site, six séances
sont réalisées en condition “rouge rare, jaune commun” et six en conditions “jaune
4
commun, rouge rare”. L’ordre de ces deux conditions est determiné aléatoirement et
indépendamment pour chaque site, de facon à limiter l’effet d’ordre. Les séances
commencent chaque matin entre 6h00 et 8h00, quand le temps le permet.
Pour la première série d’expériences, quatre sites sont étudiés (réalisation avril-
mai). Ces quatres sites sont le jardin botanique de l'université de Southampton (site 1),
l'entrée du bâtiment de médecine et de biologie (site 2), l'arrière de ce bâtiment (site 3) et
un site près du parking de la faculté de médecine et de biologie (site 4). Les expériences
sont réalisées de façon simultanée, de manière à standardiser au maximum ces
expériences, et ainsi être sûr de mesurer une différence entre sites et non pas une
différence entre heures ou entre conditions climatiques.
Les sites 2 et 3 abritent des expériences sans observation et les deux autres font
l’objet d’expériences avec observation, l’un directement (site 1), l’autre par le biais d’un
caméra vidéo (site 4). Ces deux types d’expériences sont décrits en détail plus bas. Les
quatre sites sont très distincts les uns des autres: le jardin botanique (site 1) se trouve à
environ un kilomètre de la faculté de médecine et de biologie, et les trois autres sites,
distants entre eux d'au moins 200 mètres, se situent de part et d'autre de ce bâtiment.
Afin d’obtenir plus de données sur le comportement des oiseaux à chaque visite,
deux autres expériences sont conduites (réalisation début juillet). Les deux sites choisis
pour ces expériences sont l’entrée du bâtiment de la faculté de médecine et de biologie
(site 2), ayant déjà fait l’objet d’une expérience sans observation, et le jardin de John
Allen, situé à environ cinq kilomètres de l’Université (site 5). Les deux sites sont étudiés
simultanément, le premier (site 2) par observation directe, le deuxième (site 5) par le biais
d’une caméra video. Ces deux expériences sont destinées à vérifier la reproductibilité des
expériences réalisées au mois de mai.
2.3.1 Expériences sans observation
Le premier objectif de ces expériences est de vérifier qu’il existe bien une
sélection fréquence dépendante de type anti-apostatique lorsque les oiseaux sont
confrontés à une population de proies dimorphiques.
Le second objectif est d’appréhender, de manière indirecte, la variabilité
comportementale inter-individuelle dans l’établissement de ce type de prédation. En effet,
5
si, sur un même site, la force ou la direction de la sélection fréquence dépendante varie
d’une séance à l’autre, ou bien si ces paramètres varient d’une condition à l’autre (rouge
rare ou jaune rare), cela signifie que le comportement des oiseaux n’est pas stereotypé : il
existe des différences comportementales entre les individus se nourrissant sur la
population proie. Une comparaison inter-site est également effectuée afin de savoir si les
populations prédatrices présentent une action sélective différente sur la population proie.
La population proie est laissée sur le site jusqu'à ce que les prédateurs aient consommé
entre 10 et 50% des proies. Une séance est comptabilisée dans les résultats si la prédation
reste dans ces limites, et si il reste au moins une proie du type rare disponible en fin de
séance , ceci afin de pouvoir effectuer l'analyse statistique (cf plus bas).
2.3.2 Expériences avec observation
Ces expériences sont destinées à effectuer une analyse fine du comportement des oiseaux
lorsqu’ils sont confrontés à la population proie. Les objectifs sont :
- Déterminer la composition spécifique de la population prédatrice.
- Déterminer quelles espèces effectuent une sélection fréquence dépendante anti-
apostatique.
- Estimer la contribution de chaque espèce observée à la sélection fréquence
dépendante globale.
- Appréhender les modalités sous-jacentes à cette sélection au sein d’une espèce : y-a-
t’il une sélection anti-apostatique à chaque visite ou est-ce la sommation de toutes les
visites qui aboutit à une telle action?
Dans ces expériences, la population proie est observée en continu : l'appartenance
spécifique de chaque oiseau est connu, ainsi que l'heure à laquelle il est venu s'alimenter.
En étant posté à une dizaine de mètres, il est possible, grâce à une paire de jumelles,
d’observer le comportement des oiseaux sans les déranger. Le nombre de proies
prélevées de chaque type est comptabilisé pour chaque visite. Par convention, on
considère qu’un proie est consommée dès qu’un oiseau l’a prise dans son bec. Or il arrive
très fréquemment que le prédateur lâche l'appât après l’avoir prélevé, celui ci retombant
dans la population proie. Il est alors difficile de savoir si la proie suivante de même type
prise par l’oiseau est une nouvelle ou bien celle déja comptabilisée. Dans ce cas, il est
6
nécessaire de recompter les proies pour être absolument certain du nombre de proies de
chaque type consommées. De manière générale, un recomptage en cours d’expérience est
réalisé à chaque fois qu’il y a doute sur l’action prédatrice de l’oiseau. Dans ces
conditions, les données concernant la séquence exacte de prise de nourriture sont rares.
Les expériences avec observation se sont déroulées en parallèle avec les expériences sans
observation, ce qui implique que les séances d’observations ont été arbitrairement
limitées à une heure et demie, de façon à optimiser la prise d’information.
Les données sont collectées sur une fiche de saisie sur laquelle sont inscrites la date,
l’heure de mise à disponibilité des proies et les effectifs en début d’expérience.
2.4 Méthodes statistiques d’analyse des résultats
les outils statistiques utilisées doivent permettre de mesurer la force la direction
de la sélection fréquence dépendante ou de la sélection fréquence indépendante.
2.4.1 Expériences sans observation
Pour quantifier la sélection d'un type de proie, Manly (1974) a mis au point un
indice, β, qui se calcule de la façon suivante:
+
=
rR
logcC
log
rR
log
rareβ
où C et R sont les nombres de proies au début de l'expérience respectivement communes
et rares, et c et r sont les nombres de proies restants à la fin de l'expérience,
respectivement communes et rares. β varie entre 0 et 1. Pour chaque séance, le β est
calculé pour la forme rare, le β pour la forme commune étant l’opposé (βcommun= 1 - βrare).
Le βrare est la statistique de test. Si β est égal à 0,5 , il n’y a pas de préférence pour l’une
ou l’autre des deux formes .Si βrare est proche de 1, cela signifie que les proies du type
rare sont proportionnellement plus prélevées que les proies communes : il y a donc une
sélection anti-apostatique. Si βrare est proche de 1 dans les deux conditions, on peut
conclure à une sélection fréquence dépendante, au moins en partie.
7
2.4.2 Expériences avec observation
Les données récoltées lors des séances d’observations sont les suivantes: le
nombre de proies du type commun disponibles, le nombre de proies de type rare
disponibles et le nombre de proies de chaque type prélevées par l’individu observé. A
partir de ces données, il est possible de vérifier qu'une espèce effectue un sélection
fréquence dépendante en utilisant le facteur β de la façon suivante: les nombres de proies
rares et communes avant et après chaque visite d'un individu de l'espèce étudiée sont
additionnés et le β est calculé à partir de ces sommes. Le β reflète donc la direction et la
force de la sélection par une espèce dans une condition et sur un site.
2.4.2.1 Difficultés
Les oiseaux se nourrissant des appâts ont à tout moment à leur disposition une
quantité de proies comprise entre 200 (au début de chaque séance) et 100. Le nombre de
proies qu’un individu prélève n’est pas contraint. De plus, si l’on excepte la première
visite de chaque séance, chaque oiseau se trouve face à une situation légèrement
différente. Bien qu'un type de proie soit toujours clairement beaucoup plus abondant que
l’autre, le ratio change. La densité des proies reste très élevée (supérieure à 3720 m2)
mais diminue au fur et à mesure de la prédation. Quoique la population à laquelle est
confronté un oiseau soit dépendante de l’action des précédents prédateurs, aucune
propriété fondamentale (rareté, densité) n'est grandement modifiée.
Dans ces conditions, il est impossible de comparer les prises alimentaires
simplement en terme de nombre de proies de chaque type prélevées. Le nombre total de
proies prises par l'animal est exprimé en pourcentage du nombre total de proies
disponibles avant chaque visite. De la même façon, le nombre de proies du type rare
prises est exprimé en pourcentage par rapport au nombre de proies rares disponibles.
Mais cette transformation n’enlève rien au caractère fondamentalement discret de ces
données. En effet, par convention, un oiseau prélève toujours un nombre entier de proies.
Chaque visite est donc définie par une taille (nombre de proies prélevées), un
nombre de proies rares prélevées, des conditions initiales. L'ensemble des visites
observées sur un site n'est de ce fait pas directement comparable d'une condition à l'autre
(rouge commun ou jaune conmun) : il est impossible d’effectuer un traitement statistique
8
direct de ces données qui puisse permettre de tester une différence dans l'action sélective.
De plus, le nombre de visites observées sur un site n'étant pas contraint, une comparaison
de la même condition d'un site à l'autre n'est pas non plus possible.
2.4.2.2 Approche probabilistique
Afin de pouvoir appréhender le comportement prédateur des individus d'une
espèce, il est impératif de disposer d'une série de données avec lesquelles comparer les
visites observées. Ces données doivent être des visites de même taille effectuées dans les
mêmes conditions. Pour obtenir ces données, des simulations par ordinateurs sont réalisés
en utilisant une approche probabilistique. Pour tester l'hypothèse nulle "les oiseaux
choisissent leurs proies au hasard", un premier ensemble de visites fictives est généré. Un
modèle probabilistique est ensuite utilisé pour générer une deuxième série de données, en
modifiant différents facteurs influant sur la probabilité de prélever un proie rare. Ces
données correspondent toujours à des visites de même taille réalisées dans les mêmes
conditions. L'objectif du modèle est de voir si le comportement prédateur est prédictible,
donc s'il est possible d'obtenir des visites fictives de même type que les visites réelles.
Les visites observées sont comparées avec les visites générées par la simulation de
prédation aléatoire puis avec les visites générées par le modèle probabilistique. La
variable choisie pour effectuer cette comparaison est la probabilité associée à chaque
visite, calculée selon l'hypothèse nulle.
2.4.2.2.1 Simulation de prédation aléatoire
Cette simulation est réalisée en utilisant un programme développé en language
Microsoft® Qbasic. Il s'agit de simuler des visites effectuées par des oiseaux choisissant
leurs proies au hasard, c'est-à-dire indépendamment de leur couleur. Le programme
fournit une série de données fictives correspondant à des visites de même taille que celles
observées. De plus, la simulation prend en compte les conditions initiales avant chaque
visite. Par exemple, si un oiseau observé a pris 4 proies parmi 162 jaunes et 18 rouges, la
simulation va générer une visite fictive de 4 proies parmi cette population de proies
disponibles. La règle de décision utilisée par le programme est basée sur un calcul
probabilistique. La probabilité de prélever une proie rare est égale à la fréquence des
proies rares dans la population de proies totale au moment de chaque choix. L'ordinateur
9
effectue les choix pour aboutir à la visite de façon successive. A chaque itération, le
choix précédent est pris en compte pour calculer la probabilité du choix suivant. Cette
procédure est illustrée dans la figure 1. Les visites simulées correspondant à une
prédation aléatoire sont donc effectuées choix par choix, comme les visites réelles.
rarep R
R C=
+
rarep R
R C=
−− +
11( )
rarep R
R C=
+ −( )1
proie rare
proie commune
1er choix 2eme choix
rarep R
R C=
+
rarep R
R C=
−− +
11( )
rarep R
R C=
+ −( )1
proie rare
proie commune
1er choix 2eme choix
proie rare
proie commune
proie rare
proie commune
Figure 1 : Procédure de choix utilisée par la simulation de prédation aléatoire: probabilité Prare de prélever une proie rare lors de chaque itération. R= nombre de proies rares disponibles, C= nombre de proies communes disponibles.
2.4.2.2.2 Modèle probabilistique
Le modèle probabilistique permet de simuler une attirance ou une répulsion pour
la proie rare. Ceci est réalisé en modifiant la formule probabilistique de choix de la façon
suivante:
CR
RRy
xP
rare +
=
Deux facteurs de nature différente sont intégrés dans l'équation probabilistique: x est un
facteur de préférence pour la couleur: il est constant lors de chaque simulation. Le facteur
y/R permet de simuler une action de la raréfaction: quand y/R augmente, la probabilité de
prendre une proie rare augmente. y peut prendre une valeur constante ou bien être égal à
R (dans ce cas, il n'y a pas d'action de la raréfaction). L'effectif des proies rares en début
de séance étant de 20, c'est cette valeur qui est attribuée à y pour simuler un effet de la
raréfaction.
10
Grâce à ce modèle probabilistique, l'ordinateur détermine de nouveau une série de
visites de façon similaire à la simulation aléatoire, c'est-à-dire de façon itérative. La série
de données obtenue en utilisant ces probabilités modifiées correspond à des visites par
des oiseaux possédant une préférence ou une répulsion pour la couleur rare, et répondant
ou non à la rareté de celle-ci.
2.4.2.2.3 Calcul des probabilités associées à chaque visite
La probabilité associée à chaque visite observée ou simulée est calculée selon
l'hypothèse nulle "les oiseaux choisissent leur proies au hasard". A chaque choix fait par
l'animal, la probabilité de prendre une proie rare est égale à la fréquence des proies rares
parmi la population totale. Les proies n'étant pas renouvellées, cette fréquence varie.
Selon le premier choix, la probabilité du deuxième choix se trouve modifié (figure 1) : il
y a une proie en moins de l'une des deux couleurs donc une probabilité moindre de
choisir une proie de ce type.
Chaque visite observée est la résultante des choix successifs effectués par
l'individu. La probabilité d'un protocole de visite particulier est calculé en multipliant les
probabilités de chaque choix composant cette visite. Ce protocole de visite particulier
correspond à des choix successifs dans un ordre précis. La probabilité d'un protocole
quelque soit la séquence est égale à la somme des probabilités de tous les protocoles
particuliers aboutissant à la prise du même nombre de proies de chaque type.
Un autre programme informatique en language Microsoft ® Qbasic permet de
calculer la probabilité associée à chaque visite (probabilité associée à un protocole
quelque soit la séquence) conformément à l'hypothèse nulle. Ce programme associe donc
aux visites observées et simulées une série de probabilités. La comparaison des
probabilités obtenues pour les données réelles et pour les données issues de la simulation
de prédation aléatoire permet de tester la différence entre la sélection observée et une
prédation aléatoire. Le "spectre" de probabilités obtenue en appliquant le modèle
probabilistique est lui aussi directement comparable avec le spectre de probabilités
associées aux visites observées. C'est cette comparaison qui permet de valider ou non le
modèle.
11
3 Résultats 3.1 Expériences sans observation
Ces expériences permettent d'appréhender l'homogénéité du comportement
aboutissant à la sélection fréquence dépendante par le biais d'une étude sur différents
sites. Les sites 1, 2 et 3 sont étudiés. Pour les 6 séances de chaque condition le β est
calculé pour la forme rare. Les résultats sont présentés dans la figure 2. Le site 1, qui a
aussi fait l'objet d'observation, est le seul à montrer une sélection des proies rares
significative dans les deux conditions (Test T de student par rapport à la valeur théorique
de 0,5 (pas de sélection) : p=0,002 quand les proies rouges sont les proies rares, p<0,001
quand ce sont les proies jaunes qui sont rares). Les deux autres sites montrent une
sélection significative seulement dans une des deux conditions. Pour le site 3, cette action
s'observe au détriment des proies rouges quand elles sont rares (p<0,001), mais l'action
anti-apostatique n'est pas statistiquement significative envers les proies jaunes quand
elles sont rares (p=0,427). L'étude du site 2 révèle que si l'action anti-apostatique envers
les proies jaunes est très marquée (p<0,001), la sélection des proies rouges quand elles
FigurCompsignif
0.80
0.89
0.56
0.71
0.60
0.75
0.0
0.5
1.0
site 1 site 2 site 3
Bet
a
180 rouges, 20 jaunes 180 jaunes, 20 rouges
*****
***
***N.S.
N.S.
e 2 : β moyen calculé pour la forme rare dans les expériences sans observation. araison avec la fréquence indépendance (β=0,5): ***: p<0,001, **: p<0,01, N.S.: non-icatif. Sur chaque site, N=6 pour chaque condition.
12
sont rares n'est pas significative (p=0,193). Ce premier résultat montre que la sélection
anti-apostatique n'est pas un phénomène constant, puisqu'il existe de grandes variations
entre les sites. Il y a donc des différences dans l'action globale d'une population prédatrice
confronté à une population proie hétérogène à haute densité.
Une comparaison des données obtenues dans les deux conditions au sein de
chaque site permet de montrer, là encore, une grande hétérogeneïté dans l'action
prédatrice. La différence observée dans l'action prédatrice sur le site 1 est significative
(simulation Monte-Carlo à 99% d'intervalle de confiance , 100.000 itérations, p= 0,032):
il y a donc une action anti-apostatique plus prononcée envers les proies jaunes. Le même
résultat se retrouve pour le site 2, avec là aussi une préférence pour les proies rares jaunes
par rapport aux proies rares rouges (p= 0,015). En revanche, l'étude du site 3 ne révèle
pas de différence significatives dans la surprédation des proies rares (p= 0,079). Ceci
s'explique par la très grande variabilité du βrare calculé en situation "180 rouges, 20
jaunes". On retrouve d'ailleurs une variabilité similaire en situation "180 jaunes, 20
rouges" sur le site 2.
Globalement, ces expériences sans observation montrent qu'il existe bien une
sélection anti-apostatique quand les oiseaux sont confrontés à une population hétérogène
à forte densité. Cette sélection est fréquence dépendante mais la variabilité de l'action
prédatrice implique l'intervention d'une composante fréquence indépendante de
préférence pour l'une ou l'autre des deux couleurs.
3.2 Expériences avec observation
3.2.1 La population prédatrice
Une prédation n'a été observée que sur trois des quatres sites étudiés : aucune
visite n'a été effectuée sur le site 4.
Sur les trois sites où des visites ont eu lieu, 7 espèces de passereaux ont été
observées en train de se nourrir sur la population d'appâts. Ces espèces sont, par ordre
décroissant de nombre de visites, le merle noir, le rouge-gorge, la mésange bleue,
l’étourneau sansonnet, la grive musicienne, le verdier et la mésange charbonnière. Ces
oiseaux ont effectué 410 visites et prélevé 1554 proies. Le tableau 1 présente la
composition spécifique de la population prédatrice observée pour chacun de ces trois
13
sites, d’abord en nombre de visites puis en nombre de proies prélevées. Ces résultats sont
présentés quelques soient les conditions (rouge rare ou jaune rare).
Il apparaît clairement que sur les trois sites les merles sont les prédateurs les plus
importants, tant en nombre de visites (82%) qu'en nombre de proies prélevées (93%). Les
données concernant les autres espèces observées sont trop peu nombreuses pour pouvoir
effectuer une analyse comportementale. L'étude se concentre donc sur le comportement
des merles.
N visites N proies N visites N proies N visites N proies N visites N proiesT. merula 61 454 25 174 250 811 336 1439E. rubec. 9 9 20 20 16 18 45 47P. caer. 12 12 - - - - 12 12St. Vulg. - - - - 11 45 11 45
Tableau 2 : Composition spécifique de la population prédatrice sur les trois sites d'observation, exprimée en nombre de visites (N visites) et en nombre de proies prélevées (N proies). Les espèces sont : T.merula : merle noir ; E.rubec. : rouge-gorge ; P. caer. :mésange bleue ; St vulg. : étourneau sansonnet ; T. philom. : grive musicienne ; C. chloris: verdier ; P. major : mésange charbonnière.
3.2.2 Analyse du comportement des merles (Turdus merula)
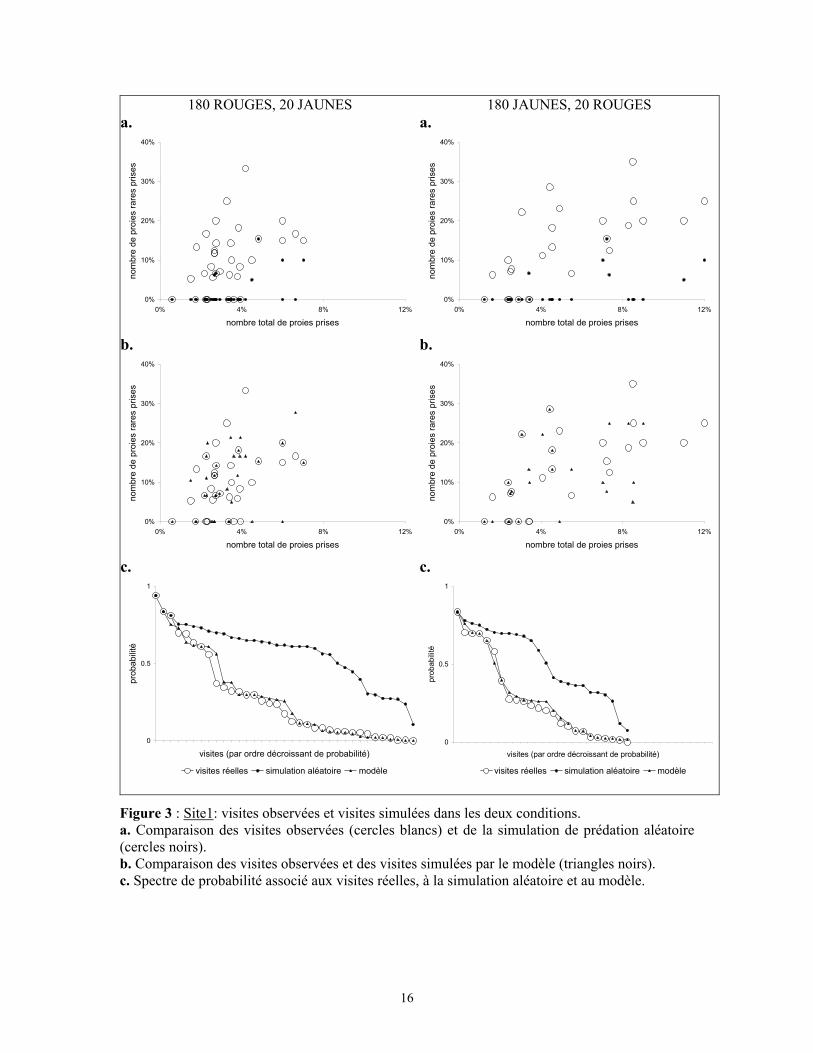
3.2.2.1 Site 1
Sur ce site, 61 visites de merles ont été observées pour un total de 454 proies. 214
proies ont été prises en 35 visites en condition "rouge commun, jaune rare", et 246 proies
ont été prélevées en 26 visites en condition "jaune commun, rouge rare". La figure 3a
expose les choix observés lors de chaque visite dans les deux conditions.
Les merles ont effectué une sélection anti-apostatique dans les deux conditions.
Le β calculé est de 0,803 pour les proies rares jaunes, et de 0,762 pour les proies rouges
rares. Les visites effectuées dans les deux conditions sont de tailles statistiquement
différentes (simulation Monte-Carlo de la méthode exacte, 100.000 itérations, N=61,
p=0,008) : le nombre total moyen de proies prises quand le jaune est commun est de 7,9 ±
6,7, et ce nombre total moyen quand le rouge est commun est de 6,1 ± 2,9 (moyenne ±
écart-type). Les merles ont prélevé des proies des deux types lors de la plupart des visites.
14
Ces visites "mixtes" sont au nombre de 27 sur 35 en condition de départ "180 rouges, 20
jaunes", et de 20 sur 26 en condition inverse.
La figure 3a présente également les visites générées par la simulation de prédation
aléatoire. La majorité de ces visites correspondent à des prises de proies de type commun,
comme le montre le regroupement de ces visites sur l'axe des abscisses. L'analyse du
spectre de probabilités, c'est à dire des probabilités calculées selon l'hypothèse nulle
(figure 3c), montre que les probabilités associées aux visites réelles sont globalement plus
faibles que celles associés à la simulation de prédation aléatoire. Dans les deux
conditions, ces deux spectres de probabilités sont statistiquement différents (simulation
Monte-Carlo de la méthode exacte : 100.000 itérations : p<0,001). La comparaison des
probabilités de la simulation aléatoire et des visites réelles, associée à la représentation
graphique de la figure 3a, montre que la majorité des oiseaux effectuent une sélection
fréquence dépendante anti-apostatique.
Etablissement du modèle.
En se basant sur les données réelles, le modèle probabilistique est testé sur le
principe suivant : la probabilité de choisir une proie rare est la résultante d'une préférence
fréquence-indépendante pour ce type de proie (x) et d'un effet de la raréfaction (y/R) sa
fréquence dans la population proie lors de chaque visite. La meilleure adéquation est
obtenue avec les paramètres suivants:
jaune rare, rouge commun rouge rare, jaune commun
CR
RR20
2P
jaune +
=CR
RR20
1.8P
rouge +
=
où R est le nombre de proies rares disponible avant chaque prise de proie par l'oiseau, et
C le nombre de proies communes disponibles au même moment. Le facteur de préférence
invariant x est de 2 pour les proies rares jaunes et de 1.8 pour les proies rares rouges. y
est égal au nombre de proies disponibles au début de chaque séance, c'est-à-dire 20. Le
facteur y/R augmente dès que des proies rares sont consommées.
Les visites simulées par ces modèles sont représentées dans la figure 3, d'abord en
comparaison avec les visites réelles en terme de nombre de proies prises (figure 3b) puis
sous la forme du spectre de probabilité associés à ces visites (figure 3c). Ces spectres
15
180 ROUGES, 20 JAUNESa.
0%
10%
20%
30%
40%
0% 4% 8% 12%
nombre total de proies prises
nom
bre
de p
roie
s ra
res
pris
es
b.
0%
10%
20%
30%
40%
0% 4% 8% 12%
nombre total de proies prises
nom
bre
de p
roie
s ra
res
pris
es
c.
0
0.5
1
visites (par ordre décroissant de probabilité)
prob
abilit
é
visites réelles simulation aléatoire modèle
180 JAUNES, 20 ROUGESa.
0%
10%
20%
30%
40%
0% 4% 8% 12%
nombre total de proies prises
nom
bre
de p
roie
s ra
res
pris
es
b.
0%
10%
20%
30%
40%
0% 4% 8% 12%
nombre total de proies prises
nom
bre
de p
roie
s ra
res
pris
es
c.
0
0.5
1
visites (par ordre décroissant de probabilité)
prob
abili
té
visites réelles simulation aléatoire modèle
Figure 3 : Site1: visites observées et visites simulées dans les deux conditions. a. Comparaison des visites observées (cercles blancs) et de la simulation de prédation aléatoire (cercles noirs). b. Comparaison des visites observées et des visites simulées par le modèle (triangles noirs). c. Spectre de probabilité associé aux visites réelles, à la simulation aléatoire et au modèle.
16
de probabilités ne sont pas statistiquement différents de ceux des visites observées
(p=0.785 quand le rouge est la couleur rare, p=0.775 quand le jaune est la couleur rare).
La modélisation en utilisant deux coefficients x et y est donc valide: elle fournit une
rationalisation simple du comportement des merles face à une population proie telle que
celle présentée. Les merles montrent une préférence pour les proies rares qui serait le
produit d'une attirance spécifique pour une couleur quand elle est rare (facteur 2 pour les
proies jaunes rares, facteur 1.8 pour les proies rouges rares) et qui augmente avec la
raréfaction de cette proie (facteur 20/R). De plus, la différence de 10% dans le facteur x
utilisé pour le modèle correspond à la différence des β calculés dans les deux conditions.
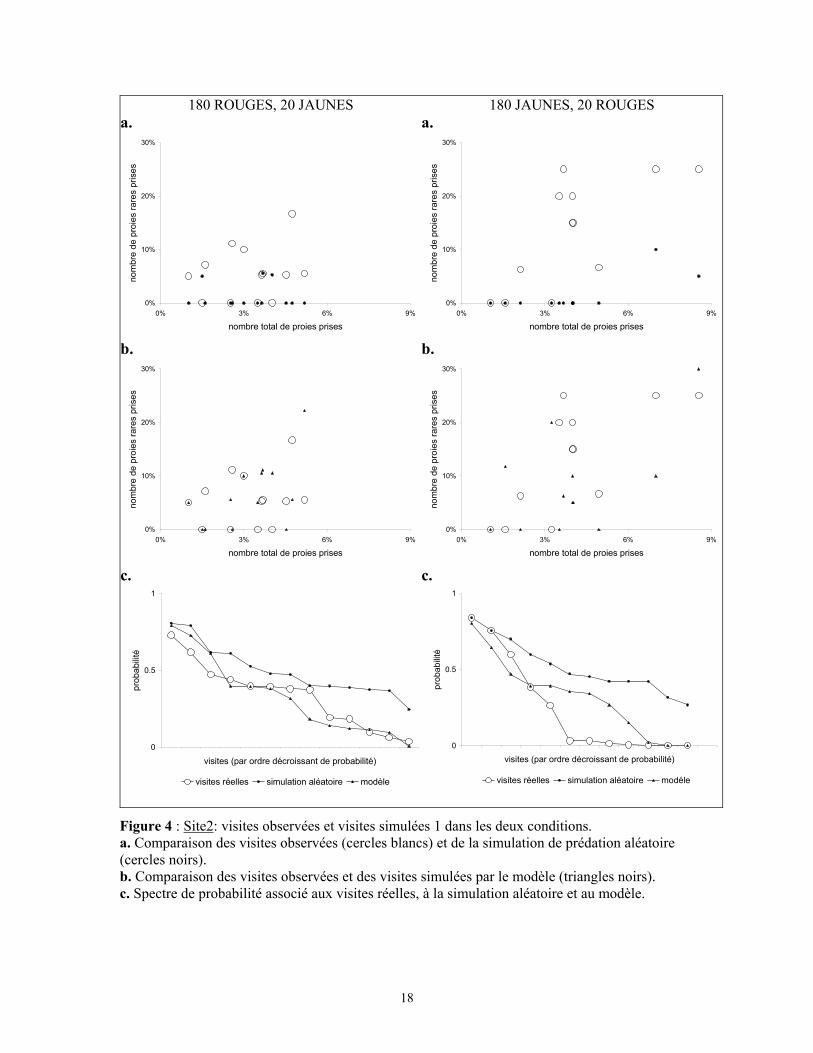
3.2.2.2 Site 2
Les visites observées sur ce site sont du même type que sur le site 1. Le nombre
de visites sur ce site est cependant inférieur : 13 en condition "180 rouges, 20jaunes", et
12 en condition inverse. Les merles effectuent une sélection fréquence dépendante de
type anti-apostatique (β=0,649 pour les proies rares jaunes, β=0,832 pour les proies rares
rouges). Les visites observées sont représentées dans la figure 4a. Les prises alimentaires
sont de taille semblable dans les deux conditions (nombre total moyen = 7 ± 3,5). Cette
taille n'est pas significativement de la taille moyenne observée sur le site 1. La figure 4a
représente également les visites générées par la simulation de prédation aléatoire.
Le modèle probabilistique est testé avec les paramètres établis pour le site 1 (x= 2
pour la probabilité de choisir une proie jaune rare, x=1,8 pour la probabilité de choisir
une proie rare rouge ; y=20). La répartition des visites fictives générées en utilisant le
modèle est représentée dans la figure 4b. La comparaison du spectre de probabilité
associé à ces visites avec celui associé aux visites réelles montre une différence non
significative dans les deux cas: p=0,624 quand le jaune est la couleur commune, et
p=0,891 quand le jaune est la couleur rare (figure 4c) ; le modèle semble là encore
adéquat pour prédire le comportement des merles.
Le spectre de probabilités obtenu pour la simulation de prédation aléatoire est
significativement différent du spectre obtenu pour les visites réelles quand le rouge est la
couleur rare (p=0,013). La différence n'est pas significative pour la situation inverse
(p=0,062).
17
180 ROUGES, 20 JAUNESa.
0%
10%
20%
30%
0% 3% 6% 9%
nombre total de proies prises
nom
bre
de p
roie
s ra
res
pris
es
b.
0%
10%
20%
30%
0% 3% 6% 9%
nombre total de proies prises
nom
bre
de p
roie
s ra
res
pris
es
c.
0
0.5
1
visites (par ordre décroissant de probabilité)
prob
abilit
é
visites réelles simulation aléatoire modèle
180 JAUNES, 20 ROUGESa.
0%
10%
20%
30%
0% 3% 6% 9%
nombre total de proies prises
nom
bre
de p
roie
s ra
res
pris
es
b.
0%
10%
20%
30%
0% 3% 6% 9%
nombre total de proies prises
nom
bre
de p
roie
s ra
res
pris
es
c.
0
0.5
1
visites (par ordre décroissant de probabilité)
prob
abili
té
visites réelles simulation aléatoire modéle
Figure 4 : Site2: visites observées et visites simulées 1 dans les deux conditions. a. Comparaison des visites observées (cercles blancs) et de la simulation de prédation aléatoire (cercles noirs). b. Comparaison des visites observées et des visites simulées par le modèle (triangles noirs). c. Spectre de probabilité associé aux visites réelles, à la simulation aléatoire et au modèle.
18
3.2.2.3 Site 5
Les visites effectuées par les merles sur ce site sont en contraste complet avec les
visites observées sur les sites 1 et 2. Ce site présente une activité prédatrice de la part des
merles très intense. 250 visites y ont été observées, et ces visites correspondent à 811
proies prélevées. Ces visites sont de plus petite taille que celles observées sur le site 1 et
le site 2 (p<0,001) : les merles ont pris en moyenne 3.2 proies (±2,4). Il n'y a pas de
différence significative dans le nombre de proies prélevées par visite entre les deux
conditions ( Simulation Monte-Carlo de la méthode exacte, 100.000 itérations, N=250 ;
p=0,137). De plus, les visites observées sont en général très courtes (moins de 5
secondes). De nombreux merles ont présenté des comportements agonistiques (26 visites
interrompues par un congénère, 2 combats).
Les merles effectuent une sélection fréquence dépendante dans les deux
conditions (β=0,85 quand le rouge est rare ; β=0,77 quand le jaune est le couleur rare).
Les visites observées sont représentées dans la figure 5a. Le comportement des merles
apparaît extrêmement différent du comportement observé sur les deux premiers sites : les
oiseaux ont une forte tendance à ne prélever qu'un type de proie, le plus souvent le type
commun, comme le montre le regroupement des visites sur l'axe des abscisses dans la
figure 5a. La figure 5b représente les visites fictives obtenues grâce à la simulation de
prédation aléatoire. Le modèle développé est inadéquat pour prédire le comportement des
merles sur ce site.
La sélection anti-apostatique est le fait de quelques visites seulement, comme le
montre la comparaison des visites observées et des visites générées par la simulation de
prédation aléatoire (figure 5). Ces visites sont celles où le prédateur prélève uniquement
des proies rares ou bien les deux types de proies à la fois. La figure 6 présente les
spectres de probabilités associés aux visites observées dans les deux conditions en
comparaison avec les spectres de probabilités des visites générées par la simulation
aléatoire. Contrairement aux deux sites précédents, où les spectres de probabilités étaient
de forme radicalement différente, ceux établis pour les visites du site 5 apparaissent
simplement décalés : Comme le montre le figure 6, ce décalage est dû aux visites où seul
le type rare est prélevé.
19
180 ROUGES, 20 JAUNESa.
-10%
0%
10%
20%
30%
40%
50%
60%
70%
80%
0% 3% 6% 9%
nombre total de proies prises
nom
bre
de p
roie
s ra
res
pris
es
b.
-10%
0%
10%
20%
30%
40%
50%
60%
70%
80%
0% 3% 6% 9%
nombre total de proies prises
nom
bre
de p
roie
s ra
res
pris
es180 JAUNES, 20 ROUGES
a.
-10%
0%
10%
20%
30%
40%
50%
60%
70%
80%
0% 3% 6% 9%
nombre total de proies prises
nom
bre
de p
roie
s ra
res
pris
es
b.
-10%
0%
10%
20%
30%
40%
50%
60%
70%
80%
0% 3% 6% 9%
nombre total de proies prises
nom
bre
de p
roie
s ra
res
pris
es
Figure 5 : Site 5: visites observées et visites simulées 1 dans les deux conditions. La taille des cercles est proportionnelle au nombre de visites. a. Visites observées. b. Visites générées par la simulation de prédation aléatoire.
Les deux spectres de probabilités sont significativement distincts des spectres de
prédation aléatoire: p=0,017 quand le jaune est la couleur rare, p<0,001 quand le rouge
est rare (simulation Monte-Carlo de la méthode exacte, 100000 itérations, N=126 en
conditions "180 rouges, 20 jaunes", N=124 en condition inverse). Cette différence de
significativité et le décalage différent par rapport au spectre de probabilités établi pour la
simulation de prédation aléatoire confirme le résultat du β calculé pour la forme rare : sur
ce site, les merles manifestent globalement une préférence pour les appâts rouges quand
ils sont rares.
20
Frlcuab
a.
180 rouges, 20 jaunes
0
0.5
1
visites (par ordre décroissant de probabilité)
prob
abili
té
proies communes proies communes et rares proies rares simulation aléatoire
b.
180 jaunes, 20 rouges
0
0.5
1
visites (par ordre décroissant de probabilité)
prob
abilit
é
proies communes proies communes et raresproies rares simulation aléatoire
igure 6 : Site5: Spectre de probabilités des visites observées et simulées. Les petits cercles noirs eprésentent les visites générées par la simulation de prédation aléatoire. Les grands cercles sont es visites observées. Les couleurs indiquent les visites où l'oiseau n'a prélevé que des proies ommunes (cercles blancs), des proies des deux types (cercles gris clair) ou des proies rares niquement (cercles gris fonçé). . Situation de départ: 180 rouges, 20jaunes. . Situation de départ: 180 jaunes, 20 rouges.
21
4 Discussion
4.1 La population prédatrice
Sur l'ensemble des expériences sans observation, il existe une tendance forte vers
la sélection fréquence dépendante anti-apostatique : à forte densité, les proies rares
subissent une prédation plus forte que les proies communes. Ceci est en accord avec
l'étude d'Allen & Anderson (1984). Cependant, il existe une grande variabilité dans la
force de cette sélection, que ce soit entre sites ou entre conditions sur un même site.
Différentes populations prédatrices ont donc des actions variées sur la population proie.
Cette variabilité ne peut être comprise qu'en effectuant une série d'études sur de
nombreux sites en connaissant la composition spécifique et le taux de prédation, afin
d'appréhender l'effet de ces deux facteurs sur la sélection fréquence dépendante.
4.2 Le comportement prédateur des merles
L'approche probabilistique développée lors de cette étude est un nouvel élément
méthodologique dans l'étude des comportements conduisant à une sélection fréquence
dépendante. En effet, le fait de pouvoir comparer statistiquement un ensemble de visites
observées avec des visites générées par une simulation de prédation aléatoire permet de
s'affranchir de la standardisation jusque là inhérente aux études sur la sélection fréquence
dépendante (situation de départ constante, ratio prédeterminé entre les deux proies). Le
modèle probabilistique développé permet de mimer le comportement des merles sur les
sites 1 et 2. Il fournit une rationalisation simple du processus de choix des proies par les
merles : la sélection des proies rares peut être comprise par l'interaction d'une préférence
pour une couleur quand elle est rare et d'un effet de la raréfaction. Comme la simulation
de prédation aléatoire, ce modèle est adaptable et testable pour des situations naturelles
non contraintes. Il est alors envisageable d'étudier le comportement d'animaux se
nourrissant sur des proies naturelles polymorphiques, par exemple, comme suggéré par
Allen (1988), des merles prélevant des baies (exemple de proies disponibles à très haute
densité).
Par ailleurs, l'étude de merles marqués permettraient d'appréhender la variabilité
interindividuelle de la préférence pour la couleur rare (facteur x). Le facteur (y/R) est
22
toujours égal à (20/R) dans le modèle utilisé, 20 étant le nombre de proies rares
disponibles en début de séance: des expériences réalisées avec un nombre de départ
différent permettraient de connaître la variabilité de y. En effet, l'application du modèle
lors de cette étude ne permet pas de savoir si les oiseaux réagissent à une proportion
(pourcentage de proies rares parmi la population totale) ou à un nombre absolu de proies
restantes. De plus, malgré son adéquation avec les visites observées sur les sites 1 et 2, le
modèle ne permet pas toujours de mimer le comportement des merles.
La présente étude a en effet mis en évidence de grandes variations dans le
comportement prédateur des merles lorsqu'ils sont confrontés à une population de proies
dimorphiques. Les différences observées entre les sites 1 et 2 et le site 5 sont
vraisemblablement plus dépendantes du contexte que d'une variabilité interindividuelle.
En effet, sur chacun des trois sites, il existe une grande homogénéité du comportement
prédateur. Le nombre estimé de merles observés sur les trois sites est respectivement de
4, 2 et 8 (estimations basées sur des différences de sexe, d'âge ou de caractéristiques
physiques). Il serait cependant nécessaire de disposer d'oiseaux marqués pour pouvoir
l'affirmer. Les 6 individus des sites 1 et 2 montrent un comportement en accord avec le
modèle, c'est-à-dire qu'ils ont tendance à prélever les deux types de proies lors de chaque
visite. En revanche, au maximum un individu semble manifester ce type de
comportement sur le site 5. Si cette estimation est vraie, la probabilité que cette
différence entre sites soit due à la seule variabilité interindividuelle est de 0,031.
La compétition est plus forte sur le site 5 que sur les sites 1 et 2, comme le montre
la taille plus faible des prises alimentaires, le nombre de visites très important et la
fréquence des comportements agonistiques. Cette forte compétition semble affecter le
comportement des prédateurs: les oiseaux ont tendance à prélever un seul type de proie à
chaque visite. Plusieurs hypothèses peuvent rendre compte de ce comportement :
Greenwood (1984) suggère qu'un prédateur pour lequel le coût de l'échantillonnage est
très élevé va avoir tendance à "essayer" le type de proie le plus abondant. Il se forme
ainsi lors de sa première visite une préférence pour la proie commune. Quand cette proie
devient rare, elle est prélevée du fait de cette préférence acquise, d'où une action anti-
apostatique. Cependant, cette hypothèse implique que tous les prédateurs n'essaient
jamais qu'une seule proie. Or, si elles sont minoritaires, les visites où les deux types sont
23
prélevés n'en existent pas moins. Il est alors possible de développer une hypothèse
alternative : le choix du type de proie à prélever est dépendant d'une visite où les deux
types sont échantillonnés. En fonction du succès rencontré avec les deux types lors de
cette visite, l'oiseau choisit un type de proie et s'y tient pour quelques jours, jusqu'à ce
qu'il réévalue la valeur de chaque type dans une autre visite "test". Cette hypothèse est en
meilleur accord avec les observations réalisées lors de la présente étude et est
concordante avec les résultats obtenus par Allen et Anderson (1984) sur trois merles en
captivité se nourrissant sur une population de proies vertes et brunes à haute densité et
présentant un ratio 9:1. La captivité implique en effet qu'il y ait une compétition pour la
seule ressource alimentaire disponible, la population d'appâts. Si la compétition affecte le
comportement prédateur, il paraît justifié d'étudier le rôle joué par l'état motivationnel des
oiseaux dans l'établissement d'un des deux types de comportements observés. L'effet de
la période de l'année (période d'élevage de jeunes, reproduction…) pourrait être étudiée
par l'observations d'oiseaux bagués en nature. L'étude d'oiseaux en captivité permettrait
d'étudier l'effet de la disponibilité de la ressource alimentaire.
La sélection fréquence dépendante anti-apostatique par les merles peut être
réalisée à chaque visite (sélection au niveau individuel, sites 1 et 2) ou lors d'une minorité
de visites fortement anti-apostatiques (sélection au niveau populationnel, site 5). Le
déterminisme de ces deux scénarios semble être contextuel (lié au taux de compétition).
La sélection au niveau individuel est dépendante de la fréquence des proies rares et
modulée par un facteur de préférence pour le type de proie. Grâce aux nouveaux outils
méthodologiques développés lors de la présente étude, il est désormais possible
d'analyser le comportement conduisant à une sélection fréquence dépendante anti-
apostatique d'animaux en situation naturelle. Une étude de la variabilité interspécifique et
interindividuelle de ce comportement apparaît donc plus que jamais nécessaire. Remerciements:
Je tiens à remercier le Dr Toby Carter pour son aide à la programmation, Frédéric
Sohm pour ces commentaires sur le rapport et tout particulièrement le Dr John Allen pour
ses conseils et son soutien tout au long de ce stage.
24
25
5 Bibliographie
Allen, J. A. 1972. Evidence for stabilizing and apostatic selection by wild passerines. - Nature 237: 348-349.
Allen, J. A. 1988. Frequency-dependent selection by predators. - Phil. Trans. Roy. Soc. Lond. B 319: 485-503.
Allen, J.A. & Anderson, K. P. 1984 Selection by passerine birds is anti-apostatic at high prey density.- Biol J. Linn. Soc. 23, 237-246.
Allen, J. A., Anderson, K. P. & Tucker, G. M. 1987. More than meets the eye - a simulation of natural selection. - J. Biol. Educ. 21: 301-306.
Allen, J. A., Raison, H. E. & Weale, M. E. 1998. The influence of density on frequency-dependent selection by wild birds feeding on artificial prey. – Proc. R. Soc. Lond. B 265: 1031-1035.
Ayala, F. J. & Campbell, C. A. 1974 Frequency dependent selection. A. Rev. Ecol. Syst. 5, 115-138. Church, S. C., Jowers, M. & Allen, J. A. 1997. Does prey dispersion affect frequency-dependent
predation by wild birds? Oecologia 111: 292-296 Clarke, B. C. 1962 a. Balanced polymorphism and the diversity of sympatric species. In Taxonomy
and geography (ed. D. Nichols), pp. 47-70. (Systematics Association publ. No. 4.) Oxford : Systematics Association.
Greenwood, J. J. D. 1984. The functional basis of frequency-dependent food selection. - Biol. J. Linn. Soc. 23: 177-199.
Greenwood, J. J. D., Blow, N. C. & Thomas , G. E., 1984 b. More mice prefer rare food. Biol. J. Linn. Soc. 23: 211-219.
Greenwood, J. J. D., Cotton, P. A. & Wilson, D. M., 1989. Frequency dependent selection on aposematic prey: some experiments. Biol. J. linn. Soc. 36: 213-226.
Greenwood, J. J. D., Johnston, J. P. & Thomas, G. E., 1984 a. Mice prefer rare food. Biol. J. Linn. Soc. 23, 201-210.
Horsley, D. T., Lynch, B. M., Greenwood, J. J. D., Hardman, B. & Moseley, S. 1979. frequency-dependent selection by birds when the density of prey is high. - J. Anim. Ecol. 48: 483-490.
Landeau, L. & Terborgh, J. 1986. Oddity and the "confusion effect" in predation. Anim Behav. 34, 1372-1380.
Lawrence, E. S. 1985 a. Evidence for search image in blackbirds Turdus merula L. : short-term learning. Anim. Behav. 33: 929-937.
Lawrence, E. S., 1985 b. Evidence for search image in blackbirds Turdus merula L. : long-term learning. Anim. Behav. 33: 1301-1309.
Manly, B. J. F., 1974. A model for certain types of selection experiment. – Biometrics 30: 281-294. Mueller, H. C., 1968. Prey selection: oddity or conspicuousness? Nature 217, 92. Mueller, H. C., 1971. Oddity and specific search image more important than conspicuousness in prey
selection. Nature 233, 345-346. Oghuchi, O., 1978. Experiments on the selection against colour oddity of water fleas by three-spined
sticklebacks. Z. Tierpsychol. 47, 154-267. Pielowski, Z., 1959. Study on the relationship: predator (goshawk) - prey (pigeon). Bull. Acad. pol.
sci. Cl. II Sér. Sci. biol. 7, 401-403. Raymond, D. L., 1984. Wild birds prefer the familiar of striped and unstriped artificial prey. – Biol.
J. Linn. Soc. 23: 229-235. Raymond, D. L. & Allen, J. A., 1990. Wild birds prefer the familiar colour pattern when feeding on
similar artificial morphs. - Oikos 57: 175-179. Sherratt, T. N. & MacDougall, A. D., 1995. Some population consequences of variation in
preference among individual predators. Biol. J. Linn. Soc. 55: 93-107. Wilson S. E., Allen J. A. & Anderson K. P. 1991. Fast movement of densely aggregated prey
increases the strenght of anti-apostatic selection by wild birds. Biol. J. Linn. Soc. 41: 375-380.



































![NLTE simulations with CRASH code and related issues Fall 2011 Review Igor Sokolov [with Michel Busquet and Marcel Klapisch (ARTEP)]](https://static.documents.pub/doc/80x56/56649eac5503460f94bb25e2/nlte-simulations-with-crash-code-and-related-issues-fall-2011-review-igor-sokolov.jpg)






