104
Control of Animal Development by Steroid Hormones Craig T. Woodard Mount Holyoke College
| Date post: | 21-Dec-2015 |
| Category: |
Documents |
| View: | 215 times |
| Download: | 0 times |

Control of Animal Development by Steroid Hormones
Craig T. Woodard
Mount Holyoke College






BACKGROUND
• The life cycle of Drosophila melanogaster has a duration of ten to twelve days, during which the embryo develops into a larvae to a stationary pupa and finally ecloses into the adult fly. This transition from larvae to adult is known as metamorphosis and is controlled by the steroid hormone, ecdysone.
•
The Life Cycle of Drosophila melanogaster

20-hydroxyecdysone

Drosophila Life Cycle
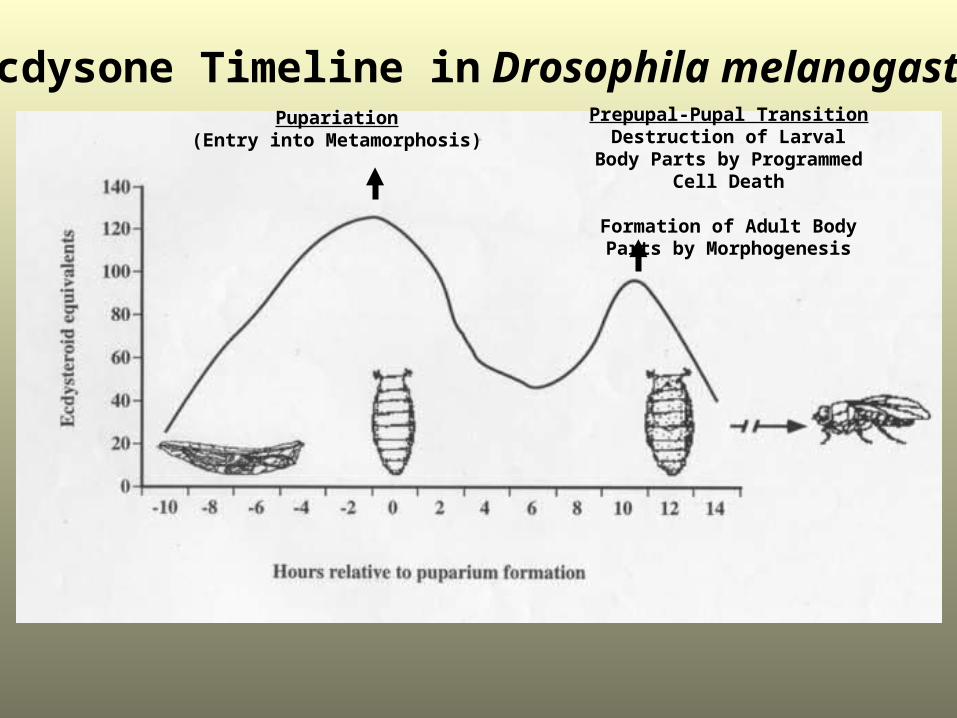
Ecdysone Timeline in Drosophila melanogasterPupariation
(Entry into Metamorphosis)Prepupal-Pupal Transition
Destruction of Larval Body Parts by Programmed Cell Death
Formation of Adult Body Parts by Morphogenesis

How can a single steroid hormone How can a single steroid hormone elicit different responses at elicit different responses at
different times in development?different times in development?

Ecdysone Timeline in Drosophila melanogasterPupariation
(Entry into Metamorphosis)Prepupal-Pupal Transition
Destruction of Larval Body Parts by Programmed Cell Death
Formation of Adult Body Parts by Morphogenesis

How can a single steroid hormone How can a single steroid hormone elicit different responses in elicit different responses in
different parts of the body during different parts of the body during development?development?

Ecdysone Timeline in Drosophila melanogasterPupariation
(Entry into Metamorphosis)Prepupal-Pupal Transition
Destruction of Larval Body Parts by Programmed Cell Death
Formation of Adult Body Parts by Morphogenesis

Morphogenesisof Adult Body Parts
Destruction of Larval body Parts by Programmed Cell Death
Pupariation (Entry into
Metamorphosis)

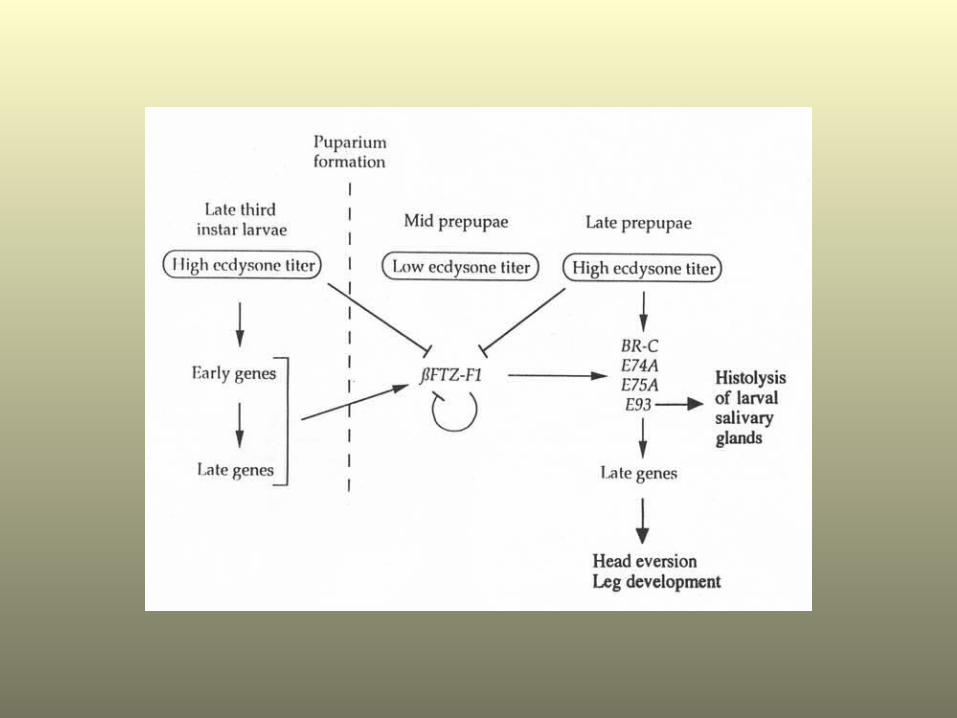
HypothesisA. ßFTZ-F1 provides the early genes, the BR-C, E74A and
E75A with the competence* to be reinduced by the prepupal ecdysone pulse.
1) These early genes then direct morphogenesis of adult body parts.
B. ßFTZ-F1 provides the prepupal stage-specific E93 early gene with the competence* to be induced by ecdysone.
ßFTZ-F1 thus directs the stage-specificity of the E93 response to ecdysone
1) E93 then directs programmed cell death in larval body parts.
*Competence the ability to respond to an inductive signal

Morphogenesisof Adult Body Parts
Destruction of Larval body Parts by Programmed Cell Death
Pupariation (Entry into
Metamorphosis)

HypothesisA. ßFTZ-F1 provides the early genes, the BR-C, E74A and
E75A with the competence* to be reinduced by the prepupal ecdysone pulse.
1) These early genes then direct morphogenesis of adult body parts.
B. ßFTZ-F1 provides the prepupal stage-specific E93 early gene with the competence* to be induced by ecdysone.
ßFTZ-F1 thus directs the stage-specificity of the E93 response to ecdysone
1) E93 then directs programmed cell death in larval body parts.
*Competence the ability to respond to an inductive signal
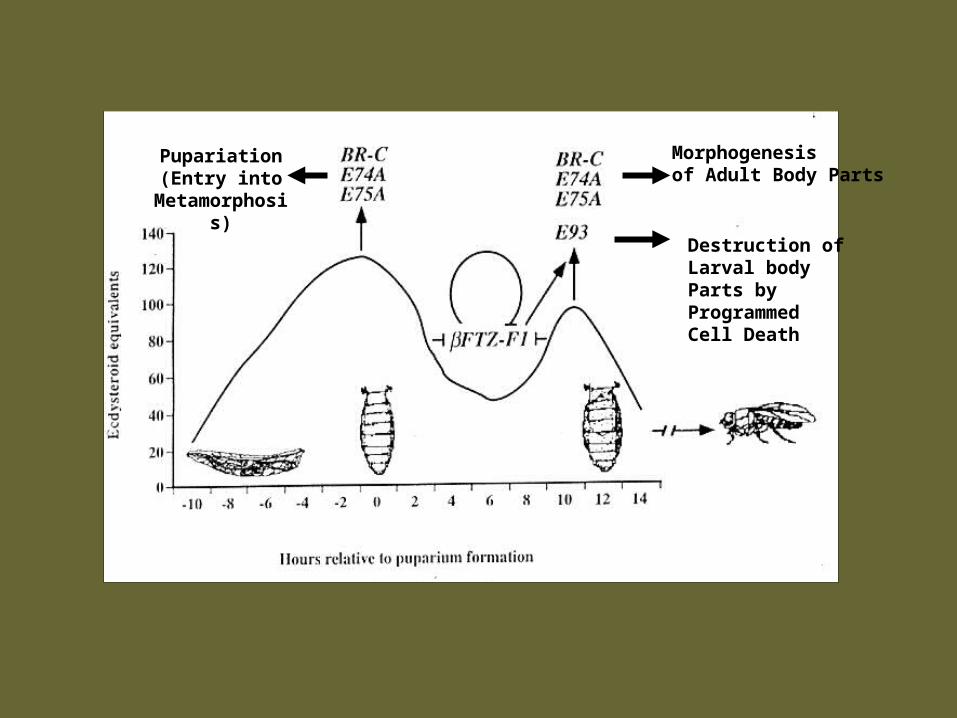
Morphogenesisof Adult Body Parts
Destruction of Larval body Parts by Programmed Cell Death
Pupariation (Entry into
Metamorphosis)

Evidence in Support of our Hypothesis
• ßFTZ-F1 protein binds to the E93, E74A, E75A, and BR-C genes.
• Ectopic (over) expression of ßFTZ-F1 in transgenic larvae provides E93 with the competence to respond (prematurely) to the late larval ecdysone pulse.
• Induction of BR-C, E74A and E75A transcripts by ecdysone is enhanced significantly by ectopic expression of ßFTZ-F1.
• A Loss-of-function mutation in ßFTZ-F1 results in dramatic reductions in E93, E74A, E75A, and BR-C activation at the end of the prepupal stage.
• A loss-of-function mutation in ßFTZ-F1 results in pupal lethality with defects in larval salivary gland programmed cell death, head eversion, and leg elongation.

Levels of Early Gene Transcripts are Reduced in ßFTZ-F1 Mutant
Prepupae

ßFTZ-F1 Mutants Exhibit Pupal Lethality and Defects in
Morphogenesis
• head eversion
• leg elongation
• wing extension

Mutations in ßFTZ-F1 Disrupt Leg Morphogenesis
Control ßFTZ-F1 Mutant
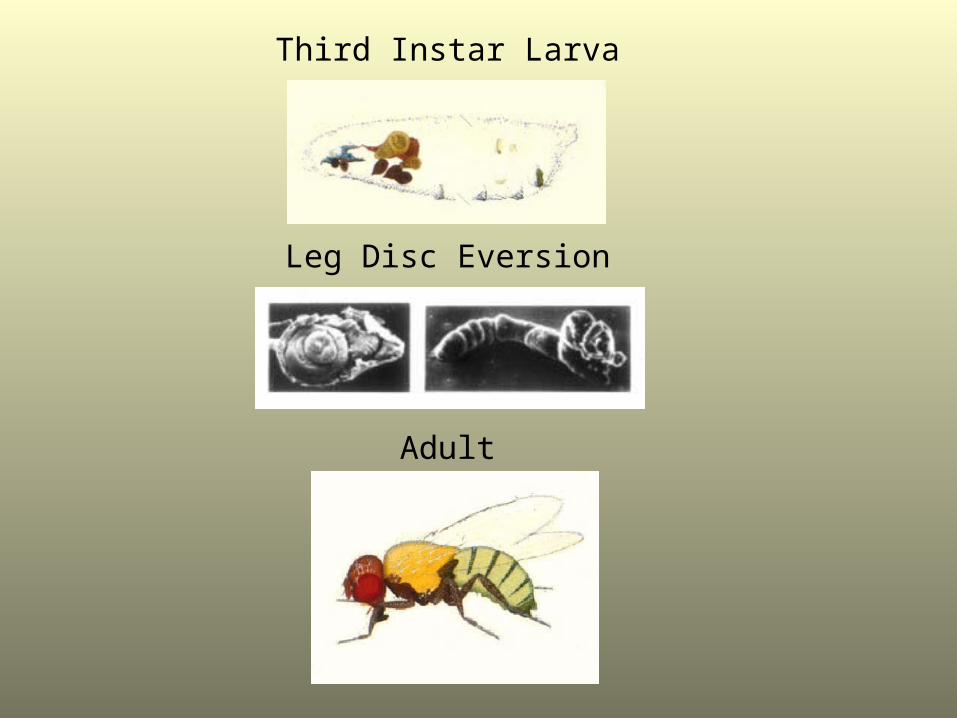
Third Instar Larva
Leg Disc Eversion
Adult

Cell Shape Changes During Leg Disc Elongation
Courtesy of Condic et al. 1991. Development 111:23-33
a b



Comparative Leg Development
Control
ßFTZ-F1 Mutant

Possible Causes of Short Legs
1) Contraction of the muscles is too weak in ßFTZ-F1 mutants.
2) There is something wrong with the leg imaginal discs in ßFTZ-F1 mutants, which prevents them from extending.


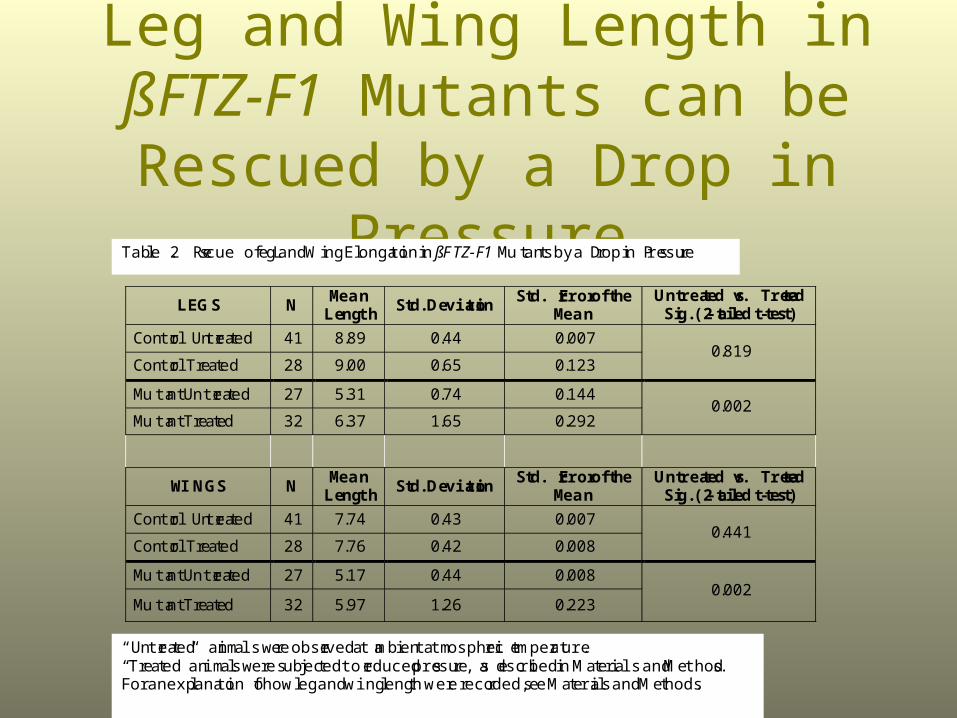
Leg and Wing Length in ßFTZ-F1 Mutants can be Rescued by a Drop
in Pressure
LEGS NMean
LengthStd. Deviation
Std. Error of theMean
Untreated vs. TreatedSig. (2-tailed t-test)
Control Untreated 41 8.89 0.44 0.007
Control Treated 28 9.00 0.65 0.1230.819
Mutant Untreated 27 5.31 0.74 0.144
Mutant Treated 32 6.37 1.65 0.2920.002
WINGS NMean
LengthStd. Deviation
Std. Error of theMean
Untreated vs. TreatedSig. (2-tailed t-test)
Control Untreated 41 7.74 0.43 0.007
Control Treated 28 7.76 0.42 0.0080.441
Mutant Untreated 27 5.17 0.44 0.008
Mutant Treated 32 5.97 1.26 0.2230.002
Table 2. Rescue of Leg and Wing Elongation in ßFTZ-F1 Mutants by a Drop in Pressure
“Untreated” animals were observed at ambient atmospheric temperature.“Treated” animals were subjected to reduced pressure, as described in Materials and Methods.For an explanation of how leg and wing length were recorded, see Materials and Methods.
Significant Difference
Significant Difference

Leg and Wing Length in ßFTZ-F1 Mutants can be Rescued by a Drop in
PressureControl Untreated vs. Treated
0255075100
10.5109.598.587.576.565.55Denticle Belt Reached by Legs
Percent of Animals
UntreatedTreated
Control Untreated vs. Treated
0255075100
10.5109.598.587.576.565.55Denticle Belt Reached by Wings
Percent of Animals
UntreatedTreated
Mutant Untreated vs. Treated
0255075100
10.5109.598.587.576.565.55Denticle Belt Reached by Legs
Percent of Animals
UntreatedTreated
Mutant Untreated vs. Treated
0255075100
10.5109.598.587.576.565.55Denticle Belt Reached by Wings
Percent of Animals
UntreatedTreated

Possible Causes of Short Legs
1) Contraction of the muscles is too weak in ßFTZ-F1 mutants.
---------------------------------------------------------------2) There is something wrong with the leg imaginal
discs in ßFTZ-F1 mutants, which prevents them from extending.
RULED OUT

Possible Causes of Short Legs
1) Contraction of the muscles is too weak in ßFTZ-F1 mutants.
This is supported by our careful observations of control and ßFTZ-F1 mutant animals going through the Prepupal-Pupal Transition.
The ßFTZ-F1 mutants exhibit severe defectsin muscle contractions.

Conclusions: Morphogenesis
ßFTZ-F1 directs the muscle movements that generate internal pressure (at the appropriate time), which drives extention of legs and wings, and eversion of the heads.
We are attempting to determine which ßFTZ-F1 target genes are involved in these processes.

Morphogenesisof Adult Body Parts
Destruction of Larval body Parts by Programmed Cell Death
Pupariation (Entry into
Metamorphosis) Target Genes?


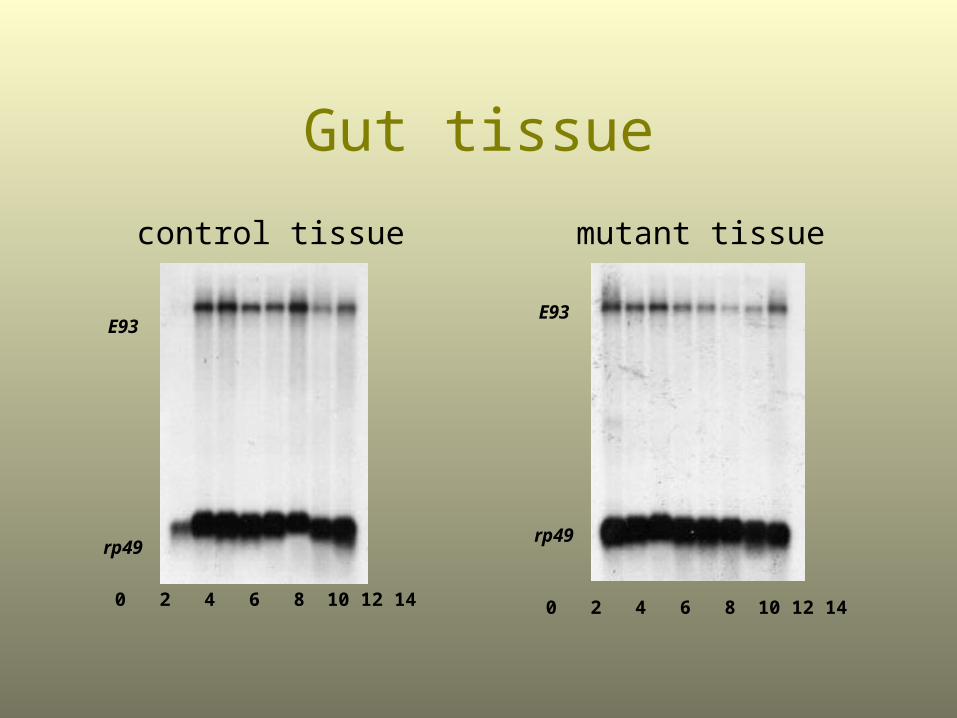
ßFTZ-F1 Mutants Fail to Activate E93 in the Larval Salivary glands
control ßFTZ-F1 mutant
E93
rp49
E93
rp49
0 2 4 6 8 10 12 14 0 2 4 6 8 10 12 14

ßFTZ-F1 Mutants Fail to Destroy Larval Salivary Glands
• Normal salivary gland histolysis ßFTZ-F1 Mutant
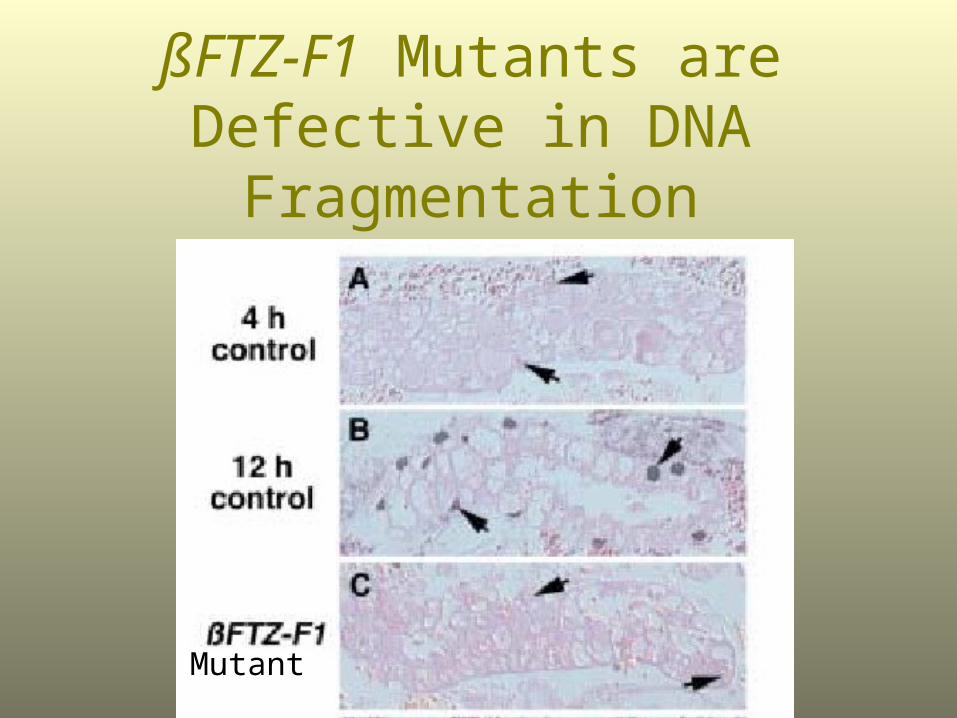
ßFTZ-F1 Mutants are Defective in DNA Fragmentation
Mutant


w
w;P[F-F1]
Ectopic Expression of ßFTZ-F1 Induces Premature Activation of E93

Ectopic Expression of ßFTZ-F1 Induces Premature Cell
Death

Ectopic Expression of ßFTZ-F1 Activates Cell Death Genes
(a Caspase)
(an Apaf-1 homolog)
LOADING CONTROL

Induction of Cell Death by ßFTZ-F1 Requires E93

Activation of Cell Death Genes by ßFTZ-F1 Requires E93

Conclusions: Programmed Cell Death
ßFTZ-F1 enables ecdysone to activate E93 in cells (such as those in the larval salivary gland) that are to be destroyed by programmed cell death.
E93 then activates other genes that direct programmed cell death.
Thus, the right cells are destroyed at the right time.

Morphogenesisof Adult Body Parts
Destruction of Larval body Parts by Programmed Cell Death
Pupariation (Entry into
Metamorphosis) Target Genes?
Cell Death Genes

Acknowledgments• Mount Holyoke College• Leg Morphogenesis• Tina M. Fortier**• Priya Vasa• Paejonette Jacobs
• E93 and Programmed Cell Death
• Tina M. Fortier**• Samara Brown**• Zareen Gauhar • Michael Chapman• Biology 340 Classes
• Mutagenesis of ßFTZ-F1• Jennifer R. McCabe• Lynn L’Archeveque• Margaret Lobo• Emily McNutt
• ßFTZ-F1 Gene Structure• Dana Cruz• Tetyanya Obukhanych• Petra Scamborova
• University of Utah• Carl Thummel
• Julie Broadus
• Bart Endrizzi
• University of Maryland
• Eric Baehrecke
• Cheng Yu Lee
• Special Thanks for Technical Assistance
• George Cobb
• Rachel Fink
• Janice Gifford
• Tamara Hjermstad
• Diane Kelly
• This research was funded by the National Science Foundation
•Mechanism of ßFTZ-F1 Action•Diyya Mathur
•Genome-Wide Functions of ßFTZ-F1•Katie McMenimen•Vidya Anegundi•Rhiana Menen
•Other•Cindy Chang•Jacque Miller

Acknowledgements
• Mount Holyoke College
• Tina M. Fortier**
• Priya Vasa
• Samara N. Brown**
• **put this presentation together
• Thanks to these folks from the University of Utah for help in making the movies.
• Carl S. Thummel
• Pamela Reid


Induction of cell death by ßFTZ-F1 requires E93

Levels of early gene transcripts are reduced in ßFTZ-F1 mutant
prepupae

Salivary glands
control tissue mutant tissue
E93
rp49
E93
rp49
0 2 4 6 8 10 12 14 0 2 4 6 8 10 12 14

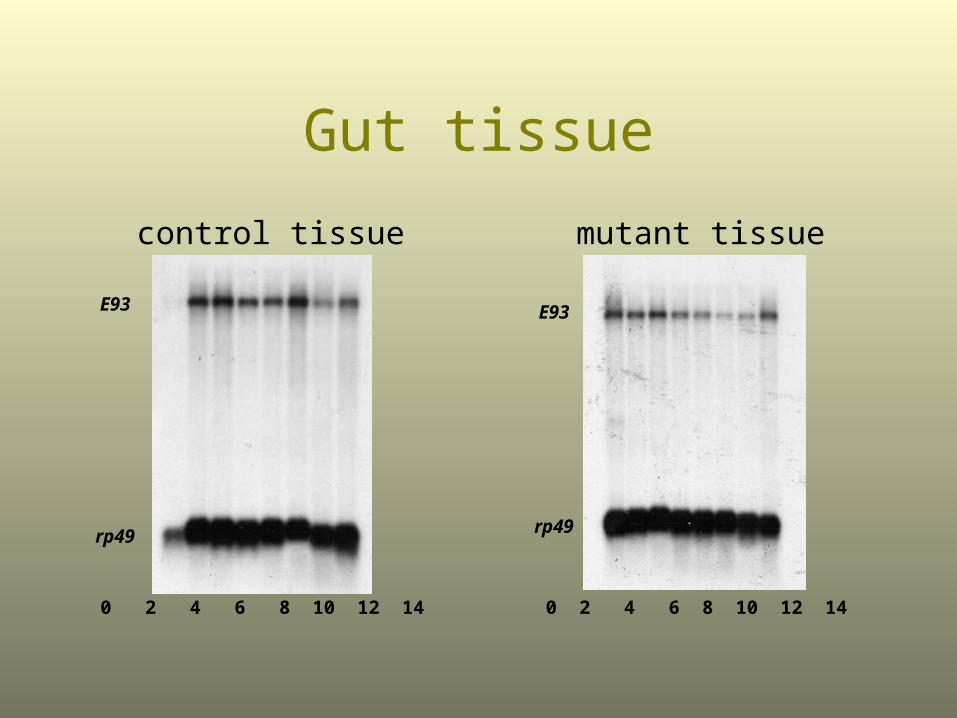
Gut tissue
mutant tissuecontrol tissue
E93
rp49
0 2 4 6 8 10 12 14
E93
rp49
0 2 4 6 8 10 12 14

SG gut fat CNS SG
hsßF
TZ
-F
1C
ontrol
hsßF
TZ
-F
1C
ontrol
hsßF
TZ
-F
1C
ontrol
hsßF
TZ
-F
1C
ontrol
hsßF
TZ
-F
1C
ontrol

Acknowledgements
• Mount Holyoke College
• Tina M. Fortier**
• Samara N. Brown**
• Michael Chapman
• Priya Vasa
• Dana Cruz
• Zareen Gauhar
• Thanks to these folks from the University of Utah for help in making the movies.
• Carl S. Thummel
• Pamela Reid

Normal Leg Development



Acknowledgements
• Mount Holyoke College
• Tina M. Fortier**
• Samara N. Brown**
• Michael Chapman
• Jennifer R. McCabe
• Priya Vasa
• Dana Cruz
• Zareen Gauhar
• Lynn L’Archeveque
• Margaret Lobo
• Emily McNutt
• Tetyanya Obukhanych
• Petra Scamborova
• University of Utah
• Carl S. Thummel
• Eric H. Baehrecke
• Julie Broadus
• Bart Endrizzi

HypothesisA. ßFTZ-F1 provides the prepupal stage-specific
E93 early gene with the competence* to be induced by ecdysone
1) ßFTZ-F1 thus directs the stage-specificity of the E93 response to ecdysone.
B. ßFTZ-F1 provides the early genes, the BR-C, E74A and E75A with the competence* to be reinduced by the prepupal ecdysone pulse.
*Competence the ability to respond to an inductive signal
Third Instar Larva
Leg Disc Eversion
Adult
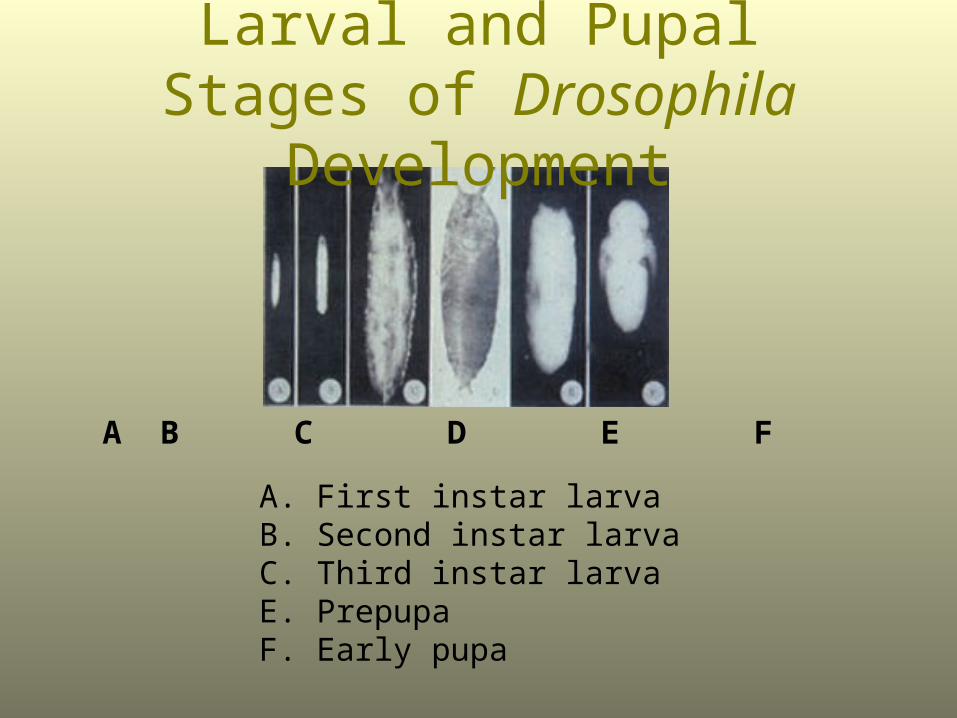
Larval and Pupal Stages of Drosophila Development
A B C D E F
A. First instar larvaB. Second instar larvaC. Third instar larvaE. PrepupaF. Early pupa
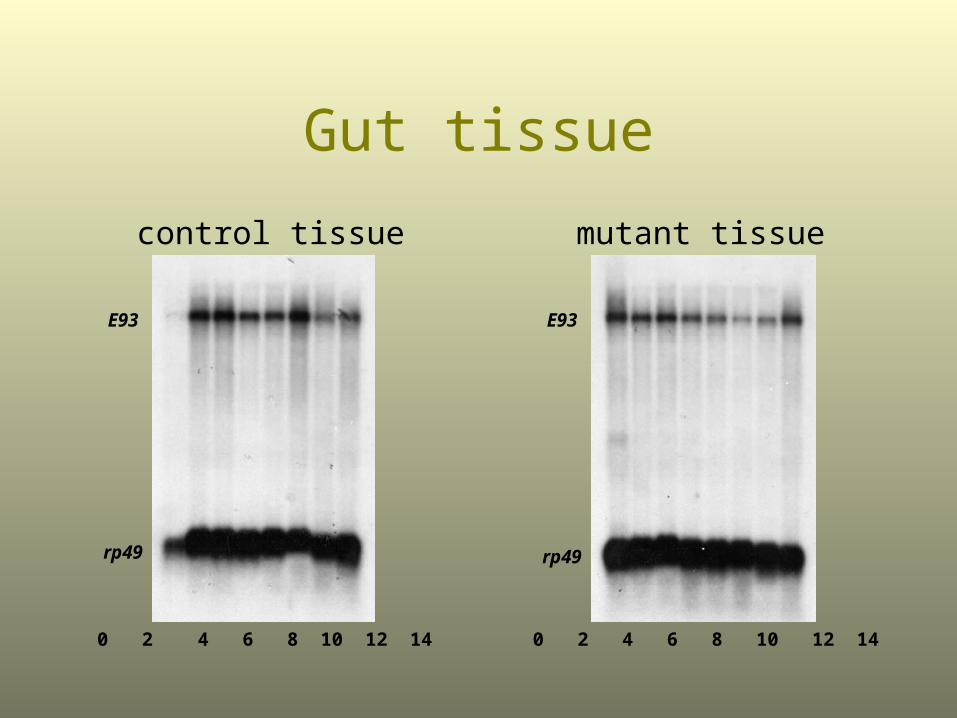
Gut tissue
mutant tissuecontrol tissue
E93
rp49
0 2 4 6 8 10 12 14
E93
rp49
0 2 4 6 8 10 12 14

Gut tissue
mutant tissuecontrol tissue
E93
rp49
0 2 4 6 8 10 12 14
E93
rp49
0 2 4 6 8 10 12 14

Gut tissue
mutant tissuecontrol tissue


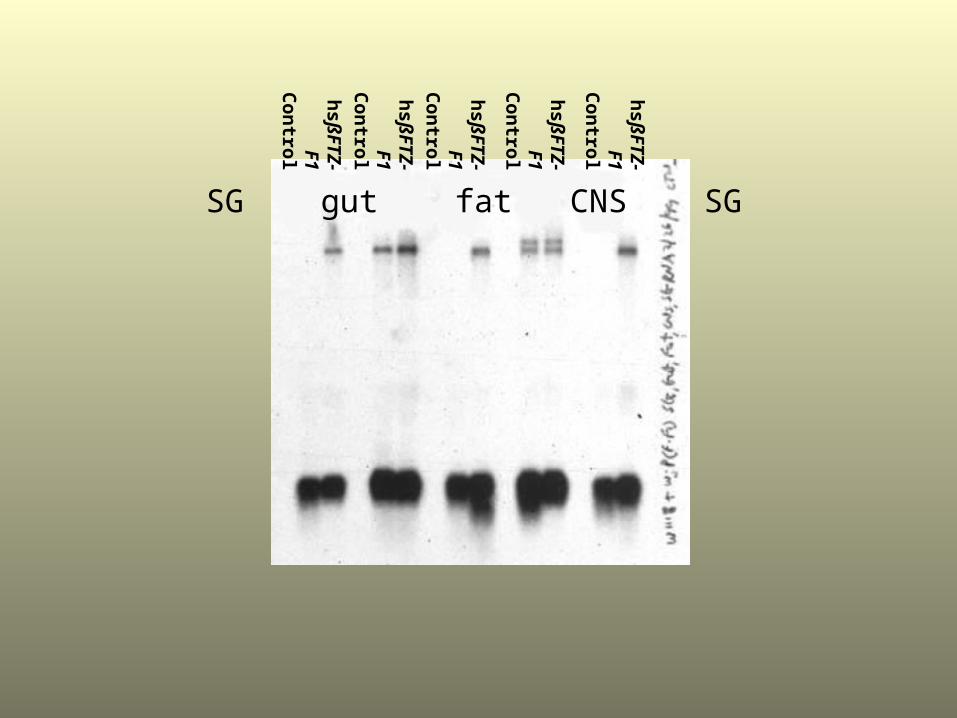
SG gut fat CNS SG
hsßF
TZ
-F
1C
ontrol
hsßF
TZ
-F
1C
ontrol
hsßF
TZ
-F
1C
ontrol
hsßF
TZ
-F
1C
ontrol
hsßF
TZ
-F
1C
ontrol


Gut tissue
mutant tissuecontrol tissue
E93
rp49
0 2 4 6 8 10 12 14
E93
rp49
0 2 4 6 8 10 12 14


Third Instar Larva
Leg Disc Eversion
Adult

Larval and Pupal Stages of Drosophila Development
A B C D E F
A. First instar larvaB. Second instar larvaC. Third instar larvaE. PrepupaF. Early pupa

Gut tissue
mutant tissuecontrol tissue
E93
rp49
0 2 4 6 8 10 12 14
E93
rp49
0 2 4 6 8 10 12 14
Gut tissue
mutant tissuecontrol tissue
E93
rp49
0 2 4 6 8 10 12 14
E93
rp49
0 2 4 6 8 10 12 14

Gut tissue
mutant tissuecontrol tissue


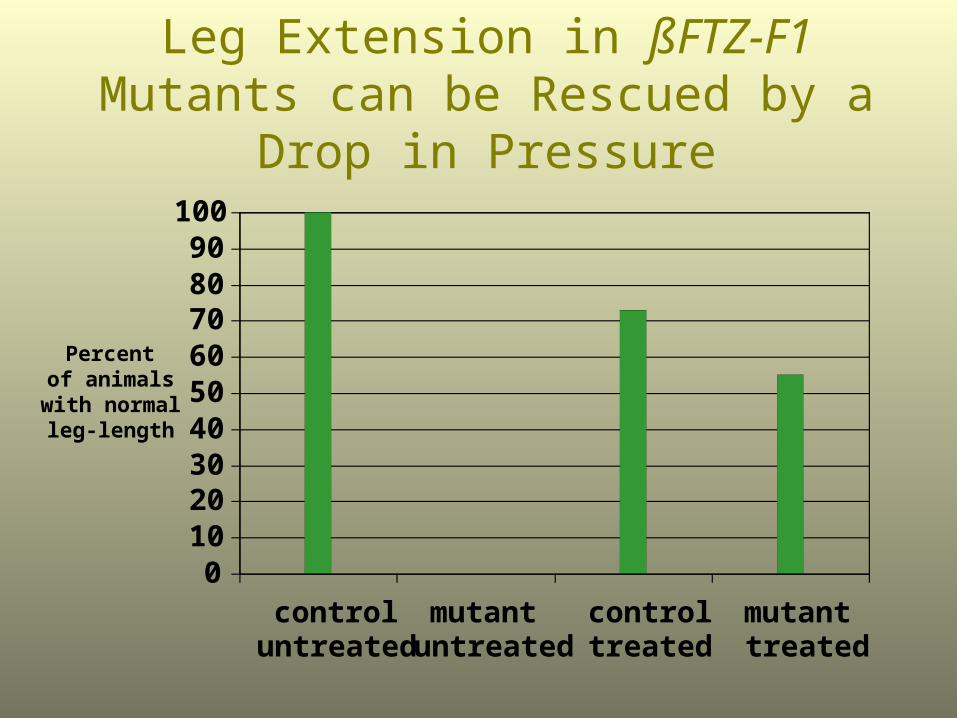
0102030405060708090
100
controluntreated
mutantuntreated
controltreated
mutanttreated
Leg Extension in ßFTZ-F1 Mutants can be Rescued by a Drop in Pressure
Percent of animals
with normal leg-length

BACKGROUND
• The life cycle of Drosophila melanogaster has a duration of ten to twelve days, during which the embryo develops into a larvae to a stationary pupa and finally ecloses into the adult fly. This transition from larvae to adult is known as metamorphosis and is controlled by the steroid hormone, ecdysone.
•
The Life Cycle of Drosophila melanogaster

Fig C. ECR Expression in Tissues

THE CHEMICAL STRUCTURE OF ECDYSONE

Ecdysone Timeline in Drosophila melanogaster

IN WHICH OTHER TISSUES DOES THE EXPRESSION OF ßFTZ-F1
AFFECT THE ECDYSONE INDUCTION OF BR-C, E74A, E75A AND E93
TRANSCRIPTION?

What is the molecular mechanism by which ßFTZ-F1 exerts its function to regulate early gene expression?
•Does ßFTZ-F1 induce expression of the ecdysone-receptor complex to facilitate the induction of the early genes?
•To test this hypothesis, in vitro experiments and Northern blot hybridization analysis was used to see if there is any ECR induction in the mid-third instar larval tissues.

EXPERIMENTAL DESIGN
• Transformant Flies called P[F-F1] were used that express a high level of ßFTZ-F1 mRNA upon heat shock.
• Control w1118 and transformant w;P[F-F1] mid-third instar larvae were heat shocked for 30 min and the tissues were immediately dissected in oxygenated Robb’s saline.
• The organs were then cultured in the presence of oxygen at 25 C for 2 hr with or without ecdysone.
• Total RNA was extracted from the tissues and analyzed for E93 mRNA by Northern blot hybridization. The Northern blot was also probed with rp49 (gene encoding ribosomal protein) as a control for loading and transfer.

How can a single steroid hormone How can a single steroid hormone elicit different responses at elicit different responses at
different times in development?different times in development?


Induction of cell death by ßFTZ-F1 requires E93

Control Untreated vs. Treated
0255075100
10.5109.598.587.576.565.55Denticle Belt Reached by Legs
Percent of Animals
UntreatedTreated
Control Untreated vs. Treated
0255075100
10.5109.598.587.576.565.55Denticle Belt Reached by Wings
Percent of Animals
UntreatedTreated
Mutant Untreated vs. Treated
0255075100
10.5109.598.587.576.565.55Denticle Belt Reached by Legs
Percent of Animals
UntreatedTreated
Mutant Untreated vs. Treated
0255075100
10.5109.598.587.576.565.55Denticle Belt Reached by Wings
Percent of Animals
UntreatedTreated

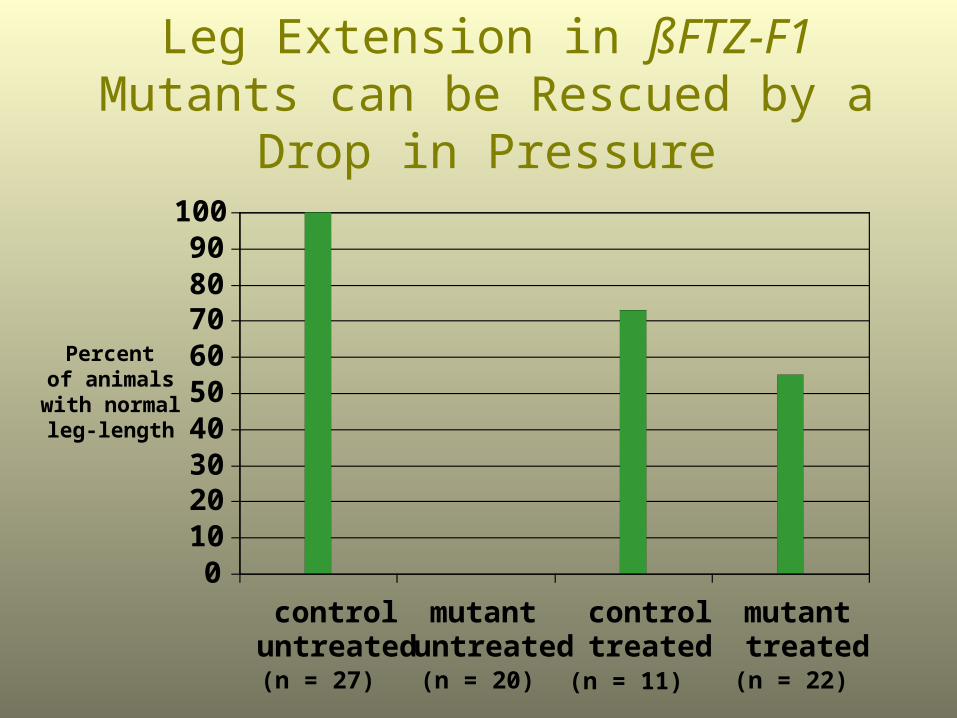
0102030405060708090
100
controluntreated
mutantuntreated
controltreated
mutanttreated
Leg Extension in ßFTZ-F1 Mutants can be Rescued by a Drop in Pressure
Percent of animals
with normal leg-length
(n = 27) (n = 20) (n = 11) (n = 22)
Leg and Wing Length in ßFTZ-F1 Mutants can be Rescued by a Drop
in Pressure
LEGS NMean
LengthStd. Deviation
Std. Error of theMean
Untreated vs. TreatedSig. (2-tailed t-test)
Control Untreated 41 8.89 0.44 0.007
Control Treated 28 9.00 0.65 0.1230.819
Mutant Untreated 27 5.31 0.74 0.144
Mutant Treated 32 6.37 1.65 0.2920.002
WINGS NMean
LengthStd. Deviation
Std. Error of theMean
Untreated vs. TreatedSig. (2-tailed t-test)
Control Untreated 41 7.74 0.43 0.007
Control Treated 28 7.76 0.42 0.0080.441
Mutant Untreated 27 5.17 0.44 0.008
Mutant Treated 32 5.97 1.26 0.2230.002
Table 2. Rescue of Leg and Wing Elongation in ßFTZ-F1 Mutants by a Drop in Pressure
“Untreated” animals were observed at ambient atmospheric temperature.“Treated” animals were subjected to reduced pressure, as described in Materials and Methods.For an explanation of how leg and wing length were recorded, see Materials and Methods.


Evidence in Support of our Hypothesis
• Staining with anti-ßFTZ-F1 antibodies shows ßFTZ-F1 protein bound to the 2B5, 74EF, 75B and 93F puff loci in prepupal salivary gland polytene chromosomes. [Lavorgna, et al. (1993) PNAS 90: 3004- 3008]
• Ectopic expression of ßFTZ-F1 provides E93 with the competence to respond to the late larval ecdysone pulse. [Woodard et al. (1994) Cell 79: 607-615]
• ßFTZ-F1 protein binds E93 genomic sequences. [E. Baehrecke, unpublished].
• Induction of BR-C, E74A and E75A transcripts by ecdysone is enhanced significantly by ectopic ßFTZ-F1. [Woodard et al. (1994) Cell 79: 607-615]
• A Loss-of-function mutation in ßFTZ-F1 results in dramatic reductions in E93, E74A, E75A, and BR-C transcripts at the end of the prepupal stage. [Broadus et al. (1999) Molecular Cell 3: 143-149]
• A loss-of-function mutation in ßFTZ-F1 results in pupal lethality with defects in larval salivary gland programmed cell death, head eversion, and leg elongation. [Broadus et al. (1999) Molecular Cell 3: 143-149]

ßFTZ-F1 mutants exhibit pupal lethality and defects in
morphogenesis

Induction of Cell Death by ßFTZ-F1 Requires E93

















