Europ.j.Proristol, 23,111-121 (1987) European Journal of
PROTISTOLOGY
Cortical Morphogenesis and Conjugation Process inCaenomorpha medusula (Ciliophora, Heterotrichida)
Ana Martin-Gonzalez, Susana Serrano and Dimas FernandezGalianoDepartamento de Microbiologia, Facultad de Biologia, Universidad Complutense.Madrid, Espana
SUMMARY
The new oral infraciliature comes from the posterior longitudinal proliferation of numeroussomatic kineties of the parental perizonal zone during the bipartition of Caenomorpha medusula. The new perizonal zone of the opisthe originates from proliferation of the anterior extremes of all the somatic kineties that make up the parental perizonal zone. Theproter retainsboth, the oral infraciliature and the perizonal zone of the parental cell.
Conjugation in C. medusula presents three maturation divisions and threepostzygotic divisions. During conjugation the oral infraciliature of each conjugant degenerates to be replacedby a new one.
Introduction
Caenomorphidae, which includes the generaCaenomorpha, Cirranter and Ludic, is a single family inthe suborder Armophorina (order Heterotrichida) [7,21] .The genus Caenomorpba has several fresh water species[19] with polysaprobic habitats and Caenomorphamedusula is one of the most frequent to be found in zoneswith a high concentration of H2S.
An ultrastructural work [30] and several morphologicalstudies [16, 19, 40] have been carried out on this species.But as far as we know, no study has yet been made ofstomatogenesis and conjugation in members of the suborder Armophorina.
In this present paper the morphogenetical events andnuclear phenomena occurring the asexual and sexualphases of the life cyclein C. medusula are described for thefirst time.
Material and Methods
Samples with C. medusula were collected from the Aulenciariver (Madrid, Spain) and cultures were made using a soil extractmedium previously inoculated with Enterobacter aerogenes andmaintained at a constant temperature of 20 ± 1"C. Subcultureswere made periodically to obtain a high population growth
necessary to studybinary fission. Some cultures were transferredto test tubes with Dryl's solution [9] to induce conjugation. Aliquotsof these cultures were stained every Y2 hour to threee hoursonce the greater part of the pairs had separated.
The staining proceding used was the pyridinated silver carbonatemethod [13]. Theorcein acetochlorhidric stain[20] wasalsoused to record thedifferent nuclear stages during theasexual andsexual phases of the life cycle.
Results
General Morphology
Caenomorpba medusula is a bell-likeshaped heterotrichwith a posterior spine. The nuclear apparatus is constituted by two or three macronuclear nodes (although up tofive nodes were exceptionally observed), and an oval micronucleus placed near the macronuclear nodes (Figs. 1, 2).
The somatic infraciliature consists only of the followingstructures: one perizonal zone, two kineties of the bell,and two very short kinetics situated at the base of thespine. The perizonal zone (PZ) begins in the right ventraledge of the bell, continues along the dorsal edge and thencurves inwards on the left side towards the left ventral partwere it ends near the spine. This zone is composed of morethan 100 oblique kineties, each averaging 20-24 pairs ofkinetosomes, except for those on the left ventral zone
0932-4739/88/0023-0111 $0.00/0
2
.~
..-. ./
1K
6Fig. 1. Caenomorpha medusula. Ventral view of a vegetative cell showing the infraciliature of the two kineties of the bell (BK),of theperizonal zone (PZ) and also the two kineties of the spine (SK) (x 720). - Fig. 2. Specimen of C. medusula with three spine kineties(SK). AZM-adoral zone of membranelles, PM-paroral membrane. Note that there are two macronuclear nodes (MA) and onemicronucleus (MI) (x 720). - Fig. 3. Detail of the left ventral zone. In this part of the cell each adoral membranelle has four rows ofkinetosomes, one of which is very short having only two or three kinetosomes (~). The paroral membrane (PM) and the two kinetiesof the spine can also be observed (SK) (x 1400). - Fig. 4. Morphogenetic stage of division in C. medusula. The cytoplasmic prolongation of the spine disappears although the kineties of this structure (SK) do not degenerate. Arrow points the antiapical proliferation ofthe most of the kineties of the perizonal zone (x 550). - Fig. 5. Detail of the left ventral zone of the same specimen of Fig. 4. Note thefibrillar connections between the submembranellar and subparoral fibres (~) (x 1320). - Fig. 6. Photomicrograph showing the oralprimordia in a dividing cell. The submembranellar and subparoral fibres have degenerated and there is a partial desdifferentiation ofthe parental buccal infraciliature (X 1550).
10
Cortical Morphogenesis and Conjugation in Caenomorpha medusula . 113
7
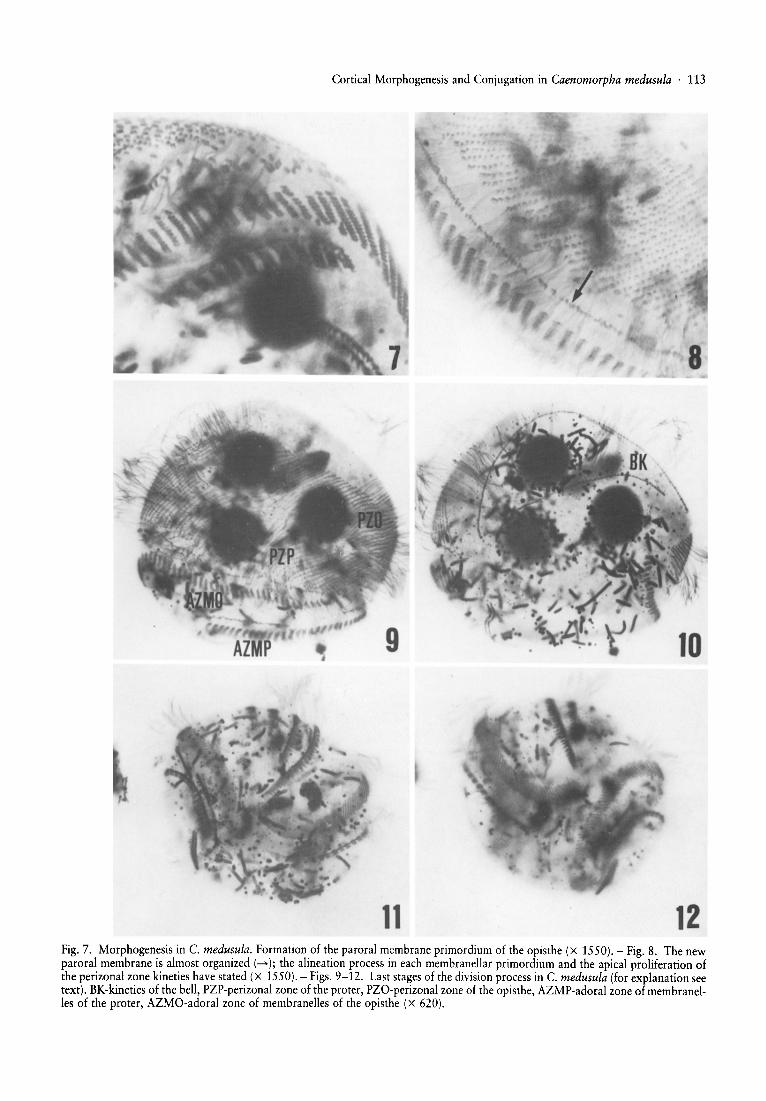
11 12Fig. 7. Morphogenesis in C. medusula. Formation of the paroral membrane primordium of the opisthe (x 1550). - Fig. 8. The newparoral membrane is almost organ ized (--+) ; the alineation process in each membranellar primordium and the apical proliferation ofthe perizonal zone kineties have stated (X 1550). - Figs. 9-12. Last stages of the division process in C. medusula (for explanation seetext). BK-kineties of the bell, PZP-perizonal zone of the proter, PZO-perizonal zone of the opisthe, AZMP-adoral zone of membranelles of the proter, AZMO-adoral zone of membranelles of the opisthe (x 620).
114 . A. Martin-Gonzales, S. Serrano and D. Fernandez-Galiano
where the number of kinetosomes per kinety decreasesprogressively (Figs. 1,2). The two kineties of the bell aredifferent in length; the shortest one is in the middle ventralpart and has a longitudinal trajectory whilst the secondone extends anteriorly towards the dorsal side (Fig. 1).Each of these kineties has two rows of kinetosomes. Thekineties of the spine are made up of two rows of kinetosomes and sometimes three kineties can be observed in thiszone (Figs. 1, 2).
The oral infraciliature consists of one adoral zone ofmembranelles (AZM) and one paroral membrane. TheAZM is parallel to the perizonal zone and has 37-45membranelles, each with three or four rows of kinetosomes. Not all the membranelles are of the same length,the most posterior ones being longer than the others. Theparoral membrane is located between the adoral zone ofmembranelles and the perizonal zone and it is a diplostichomonade which extends from the base of the spine on
Fig. 13. Scheme showing the morphogenetic events during division of C. medusula.
Cortical Morphogenesis and Conjugation in Caenomorpha medusula . 115
the ventral side to the dorsal part where it ends beforereaching it. Located beneath the AZM and the paroralmembrane are two fibres: the submembranellar fibre andthe subparoral fibre with some fibrillar connectionsbetween them (Figs. 2, 3, 5).
Binary Fission (Fig. 13)
During the asexual division of the vegetative cell thebody adopts an ellipsoidal shape and the cytoplasmic prolongation of the spine dissappears although the infraciliature of this zone remains intact. The cellular movementdecreases greatly during morphogenesis even though theciliate never loses its cilia with the result that these cellssink down to the bottom of the culture container.
We have divided morphogenesis into eight stages:Stage 1. - The kinetosomes of the somatic kineties in the
perizonal zone lose their fibrillar derivates. Then thesesomatic kineties undergo a longitudinal and antiapicalproliferation following a left to right gradient except forthe most right kineties where no sign of proliferation canbe observed (Figs. 13a, 4, 5).
Stage 2. - The kineties in the perizonal zone continueproliferating and the fibrillar derivates between the AZMand the paroral membrane disappear. Simultaneously apartial desdifferentiation of the parental paroral membrane occurs (Fig. 13b and 6).
Stage 3. - The posterior extremes of the kineties of theperizonal zone that have been proliferating graduallybreak and imigrate to an intermediate zone closer to theoral structures. Each of these kinetal segments is made upof two rows of kinetosomes which decrease in length asthey follow a right (Figs. Be, 6).
Stage 4. - The anterior extremes of these kinetal segments or membranellar primordia break off and then alengthened and anarchic field of pairs of kinetosomes isformed between the membranellar primordia and theparental perizonal zone (Figs. 13d, 7).
Stage 5. - The somatic kineties of the perizonal zoneincrease in length due to an apical proliferation (Fig. Be).
Stage 6. - The kinetosomal field placed between theperizonal zone and the membranellar primordia isarranged giving rise to a row of pairs of kinetosomeswhich represents the new paroral membrane of theopisthe. A rearrangement of the kinetosomes in the oldparoral membrane was also observed (Fig. 13g).
Stage 7. - When the new paroral membrane is completely formed a delineation of one or two rows of kinetosomes takes place, so that each membranelle exhibits thesame number of rows as those in the vegetative cell (Figs.13g,8).
At the same time, the kineties of the spine and those ofthe bell, which had lost their fibrillar derivates, increase inlength.
Stage 8. - When all the infraciliar structures are formedin the proter and in the opisthe, a complex succession ofmorphogenetic movements takes place in the cell.
The perizonal zone breaks into two parts and later anunciliated area appears between them, the anterior part ofwhich constitutes the perizonal zone of the opisthe and theposterior part the perizonal zone of the proter. Both partsare ciliated but their kinetosomes do not bear fibrillar derivates (Figs. 9, 10).
Initially, the cell elongates transversally so that its widthis greater than its length. As a result, the two kineties ofthe spine break into two fragments, one of which is in theright posterior half of the body. Later, the posterior peri-
.>. /.~
.. .. r. 1\x x /\ x x ./\ x x
l_-=-o-=::::=:::::=---==::====~./~. /()~.
1\ 1\ /\ /\, /\/\/\ /\/\/\ '
14Fig. 14. Scheme of nuclear changes during the conjugation process in C. medusula.
116 . A. Martin-Gonzales, S. Serrano and D. Fernandez-Galiano
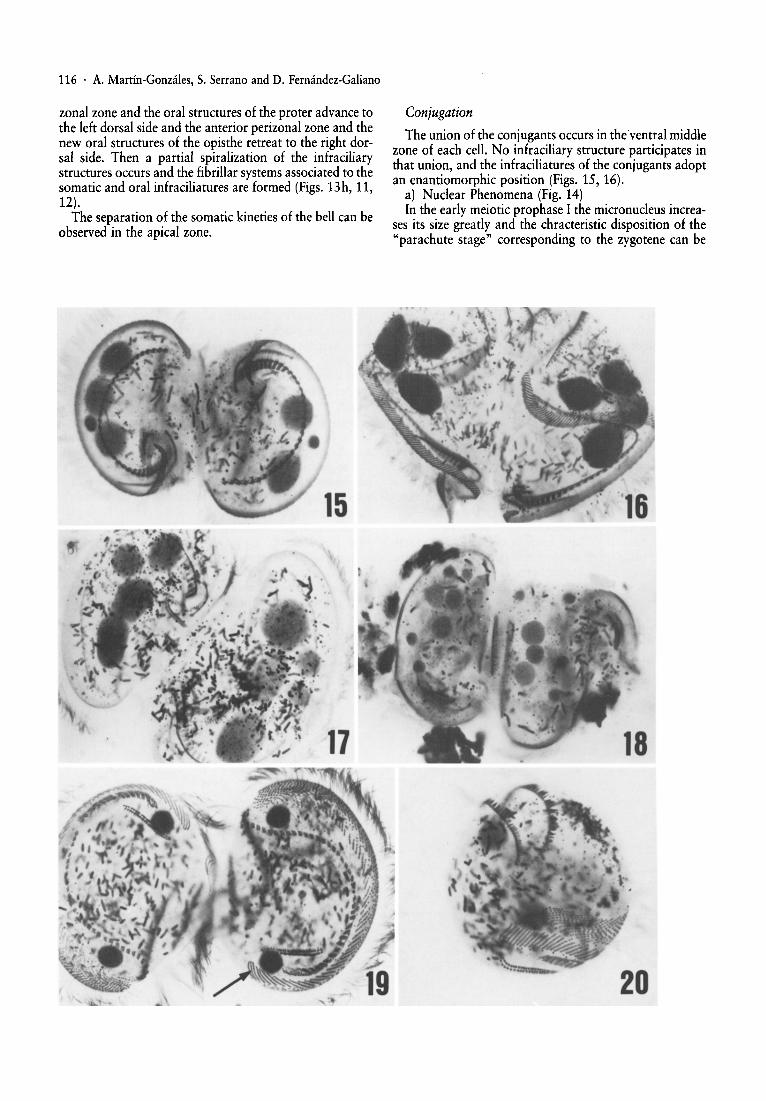
zonal zone and the oral structures of the proter advance tothe left dorsal side and the anterior perizonal zone and thenew oral structures of the opisthe retreat to the right dorsal side. Then a partial spiralization of the infraciliarystructures occurs and the fibrillar systems associated to thesomatic and oral infraciliatures are formed (Figs. 13h, 11,12). . . .
The separation of the somanc kineties of the bell can beobserved in the apical zone.
15
Conjugation
The union of the conjugants occurs in theventral middlezone of each cell. No infraciliary structure participates inthat union, and the infraciliatures of the conjugants adoptan enantiomorphic position (Figs. 15, 16).
a) Nuclear Phenomena (Fig. 14). .In the early meiotic prophase I the micronucleus mcrea
ses its size greatly and the chracteristic disposition of the"parachute stage" corresponding to the zygotene can be
'16
...
17
./
19
18
20
Cortical Morphogenesis and Conjugation in Caenomorpha medusula . 117
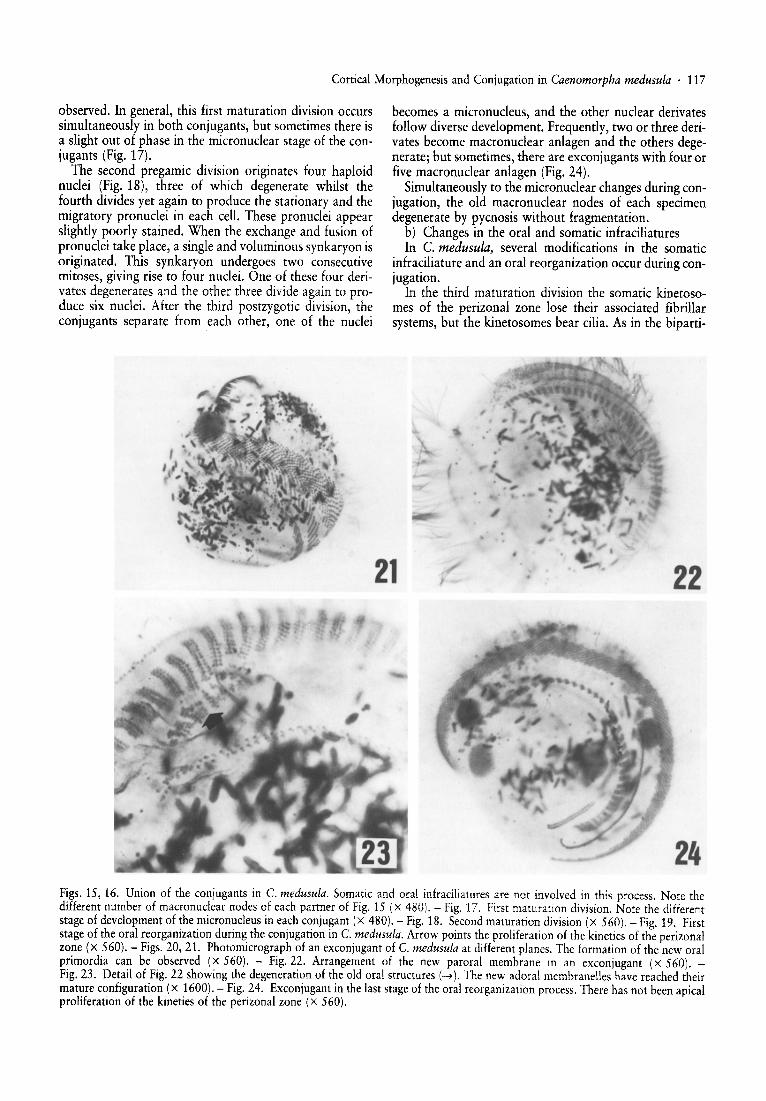
observed . In general, this first maturation division occurssimultaneously in both conjugants, but sometimes there isa slight out of phase in the micronuclear stage of the conjugants (Fig. 17).
The second pregamic division originates four haploidnuclei (Fig. 18), three of which degenerate whilst thefourth divides yet again to produce the sta tionary and themigratory pronuclei in each cell. These pronuclei appearslightly poorly stained. When the exchange and fusion ofpronuclei take place, a single and voluminous synkaryon isoriginated. This synkaryon undergoe s two consecutivemitoses, giving rise to four nuclei. One of these four derivates degenerates and the other three divide again to produce six nuclei. After the third postzygotic division, theconjugants separate from each other, one of the nuclei
becomes a micronucleus, and the other nuclear derivatesfollow diverse development. Frequently , two or three derivates become macronuclear anlagen and the other s degenerate; but sometimes, there are excon jugants with four orfive macronuclear anlagen (Fig. 24).
Simultaneously to the micronuclear changes during conjugation, the old macronuclear nodes of each specimendegenerate by pycnosis without fragmentation.
b) Changes in the oral and somati c infraciliaturesIn C. medusula, several modifications in the somatic
infraciliature and an oral reorganization occur dur ing conjugation.
In the third maturation division the somati c kinetosomes of the perizonal zone lose their associated fibrillarsystems, but the kinetosomes bear cilia. As in the biparti-
,I / .
21 22
24Figs. 15, 16. Union of the conjugants in C. medusula. Somatic and oral infraciliatures are not involved in this process. Note thedifferent number of macronuclear nodes of each partner of Fig. 15 (x 480). - Fig. 17. First maturation division. Note the differentstage of development of the micronucleus in each conjugant (X 480). - Fig. 18. Second maturation division (x 560). - Fig. 19. Firststage of the oral reorganization during the conjugation in C. medusula. Arrow points the proliferation of the kineties of the perizonalzone (x 560). - Figs. 20, 21. Photomicrograph of an exconjugant of C. medusula at different planes. The formation of the new oralprimordia can be observed (X 560). - Fig. 22. Arrangement of the new paroral membrane in an exconjugant (x 560). Fig.23. Detail of Fig.22 showing the degeneration of the old oral structures (..... ). The new adoral membranelles have reached theirmature configuration (X 1600). - Fig.24. Exconjugantin the last stage of the oral reorganization process. There has not been apicalproliferation of the kineties of the perizonal zone (x 560).
118 . A. Martin-Gonzales, S. Serrano and D. Fernandez-Galiano
tion process, the disappearance of the fibrillar systemsassociated to the kinetosomes produces a decrease in themovement of the conjugants that sink to the bottom. Atthis moment the fibrillar structures associated to the adoral zone of membranelles and the paroral membranedisappear. We have not observed changes in the kineties ofthe spine and the bell.
The somatic infraciliature does not undergo any otherchanges during conjugation.
After the formation of the synkaryon, a slow and gradual degeneration of the paroral membrane and the adoralzone of membranelles begins in each conjugant. An antiapical proliferation of the kineties in the perizonal zoneappears during the three synkaryon divisions and this proliferation gives rise to the membranellar primordia(Fig. 19). When the conjugant pairs separate, the oralreorganization continues according to the same pattern asthat in the stomatogenesis of this ciliate, that is, the pararal membrane and the new adoral zone of membranellesare formed and the parental oral structures degenerate(Figs. 20-24).
Discussion
Our observations on the general morphology in Caenomorpha medusula differ on several points with previousdescriptions by Villeneuve-Brachon [40] and Jankowski[19] although we basically agree with the results obtainedby Fernandez-Galiano and Fernandez-Leborans [16].
The somatic infraciliature of this ciliate is very reduced[19] and it is not uniformly distributed as oblique rows onthe dorsal side and as horizontal rows on the ventral surface [40]. However, as pointed out by Fernandez-Galianoand Fernandez-Leborans [16], Jankowski's results presenttwo important erroneous interpretations; in the first place,each kinety of the bell is not formed by eight to ten cirribut has in fact two rows of kinetosomes as Rodrigues deSanta Rosa (30] stated in his brief ultrastructural study. Inthe second place, the perizonal zone is not a polykinetymade up of oblique rows with five kinetosomes: on thecontrary, this zone is composed of numerous obliquerows, each with 20 to 24 pairs of kinetosomes.
With regard to the oral infraciliature, Rodrigues deSanta Rosa [30] indicated that each membranelle has fourrows of kinetosomes whereas Fernandez-Galiano andFernandez-Leborans [16] only observed three rows ofkinetosomes per oral membranelle, each one with tenkinetosomes. Both affirmations are certainly true becausethey are the consequence of a partial observation of theadoral zone of membranelles. In C. medusula this structure consists of 37 to 45 membranelles of different sizes;the most posterior or left ventral ones present four rows ofkinetosomes, three of which have nine kinetosomes whilstthe fourth is very short with only two or three kinetosomeso The other oral membranelles are smaller and eachhas only three rows with five kinetosomes.
The paroral membrane presents two close rows of kinetosomes. Its ultrastructure (30J corresponds to a diplostichomonade [28].
Finally, we have to emphasize that in our strain a ratherhigh percentage of cells appear with three kineties in thespine and not two as described by all the previously mentioned authors. Furthermore, there are two to four macronuclear nodes although in a few cases we have found fivenodes although Jankowski [19] pointed out that C. medusula had three or four macronuclear nodes instead of twonodes as C. lata.
As mentioned above, no study on the morphogeneticalprocess has been made up to now in any of the speciesincluded in the suborder Armophorina. The only referencein the matter denotes that this stomatogenesis "is verylikely of an apokinetal nature" [7]. As can be observedclearly in our photomicrographs, at least in C. medusula,all the new oral structures of the opisthe proceed from theproliferation of the greater part of the somatic kineties ofthe parental perizonal zone and therefore they derive fromthe parental somatic infraciliature.
In other studied species included in the different suborders of the order Heterotrichida, stomatogenesis is apokinetal or parakinetaI. In the suborder Heterotrichina, themost detailed studies are centred on the genera Blepbarisma [26, 32], Condylostoma [3J, Climacostomum [10]and Stentor [26]. In all of these genera the oral primordium originates from the left lateral proliferation of somepostoral somatic kineties which constitute an outstandingstomatogenic territory. This territory coincides with or isplaced to the right of the discontinuity zone or area "V".As a result of this proliferation a kinetosomal field isformed which later breaks into two unequal parts: a leftone which always gives rise to the new adoral zone ofmembranelles and a right one which originates the paroralmembrane in all the species and other structures of paroralcomplex, if the cell has them, but not the frontal field ofStentor which has a somatic origin [26J.
In the suborder Coliphorina stomatogenesis is parakinetal whereas the members in the suborder Licnophorinapresent an apokinetal stomatogenesis [7, 21J.
In Nyctotherus ovalis (31], included in the suborderClevelandellina, stomatogenesis begins at the zone of discontinuity originating an orderly suboral kinetosomalfield which represents the primordium of the infundibularadoral zone of membranelles. The new peristomial adoralzone of membranelles results from the proliferation of the8-10 most ventral kineties of the proliferation zone. Theparoral membrane is formed by proliferation of the somatic kineties which are placed over the proliferation furrow.
If we compare our results in C. medusula with stomatogenic patterns of other members of the order Heterotrichida, many differences are made evident. Firstly, thesomatic infraciliature is very scant in C. medusula and thevegetative cell has neither a suboral outstanding territorynor is it originated during stomatogenesis although manysomatic kineties of the perizonal zone (included the dorsalones) take part in this process of stomatogenesis. On theother hand, the formation gradient of the adoral membranelles is postero-anterior and extends from the left ventralzone towards the right ventral zone. In contrast, in otherheterotrichous ciliates, for instance in Blepharisma (26,32J and Stentor [26J, the arrangement gradient of the
Cortical Morphogenesis and Conjugation in Caenomorpha medusula . 119
membranellar primordia is antero-posterior and in Nyctotherus ovalis [31] this gradient is dorso-ventral.
Stomatogenesis in Nyctotherus ovalis [31] presents twosimilarities with C. medusula. (1) The oral primordiumderives from a longitudinal proliferation of the somatickineties exclusively and (2) this oral primordium alwaysappears ordered.
As can be verified, at least in C. medusula, stomatogenesis-isnot apokinetal as Corliss [7] presupposed; rather, theoral structures of the opisthe derive directly from thesomatic kineties of the parental perizonal zone. However,the stomatogenic pattern of this species can not be included in any of the two types of stomatogenesis with somaticorigin defined by Corliss [6, 7] and Corliss and Lorn [8]. Inthis ciliate, stomatogenesis is not telokinetal because theposterior extremes of the parental perizonal zone areinvolved and not the anterior extremes of the somatickineties. Furthermore, it is not parakinetal because thestomatogenic kineties are not postoral neither they are allventral and proliferation is not lateral but longitudinal.
In Caenomorpha medusula the union of the conjugantsoccurs in the anterior and middle ventral zone of each cell.This process has been studied in only a few species ofheterotrichs in which the conjugant union takes place atthe peristomial areas, which can be unciliated as in Blepharisma [22] and Spirostomum (personal observations)or ciliated as in Stentor [4].
With regard to the nuclear phenomena present duringconjugation, the heterotrichous ciliates are not a homogeneous group; there are differences even amongst the species of the same genus as can be expected if one considerstheir high variety of nuclear number. In C. medusula thenumber of macronuclei does not seem to bear relationshipwith the mating type; consequently we have observedpairs of conjugants in which both cells have the samenumber of nodes and others with an unequal number ofmacronuclear nodes.
In our species, as in Spirostomum ambiguum [35], Blepharisma americanum [1], Stentor polymorphus andS. coeruleus [23] and many other species of ciliates, thereare three maturation divisions but only one nuclear derivate of the second pregamic division undergoes the thirdmaturation division. However, in other heterotrichs suchas Blepharisma tropicum [36] and Fabrea salina [12] twoor three to six nuclei undergo this third division so that thepronuclei of each conjugant can not be genetically identical.
The reconstruction of the nuclear apparatus does notoccur in any of the subtypes described by Raikov [29]when the synkaryon divides three times. In C. medusulaone of the derivates from the second postzygotic divisiondegenerates and the number of macronuclear anlagenvaries. The variability of nuclear phenomena in the exconjugant has been previously described in other species ofciliated protozoa, for instance Didinium nasutum [27] andFabrea salina [12].
Oral reorganisation during conjugation was reported byMaupas (1889). This reorganisation begins when the conjugants are still united as in Paramecium tetraurelia [25]and Urocentrum turbo [34]. In these species oral reorgani-
sation begins soon after the pairs unite and finishes whenthe exconjugants are formed, but in C. medusula the process begins during the exchange of pronuclei and concludes in the exconjugants. In Nyctotheroides cordiformis(= Nyctotherus cordiformis) [41] only the adoral zone ofmembranelles degenerates and disappears during the firstmaturation division in order to be substituted by anotherof new formation.
It must be pointed out that in C. medusula the fibrillarderivates associated to the kinetosomes disappear duringthe bipartition and conjugation processes. In both stagesof the biological cycle this phenomenon seems to be anecessary and preceding step to the beginning of the longitudinal proliferation of the kinetosomes, and it bears withthe stop of the ciliary movement.
According to Grain [18] the fibrillar and microtubularderivates provide mechanical anchorage for the kinetosomes, but other authors [24,39] consider that these structures determine the direction of the effective stroke of ciliarybeat.
The shortening or overlapping of fibrillar derivates asthe kinetodesmal fibers during the bipartition in ciliateshave been observed in Paramecium [11, 14, 15]. Fernandez-Galiano [14] thinks that the kinetodesmal fibers mightbe a protein reserve that is mobilised during division whilstCohen et al. [5] suggest that the likely role of these fibers isskeletal and this role is assumed by the cytospindle duringbipartition of the cell.
Some considerations on the systematic position of theorder Heterotrichida
In the classifications made by Corliss [7] and by Levineet al. [21], the order Heterotrichida is included within theorders Odontostomatida, Oligotrichida and Hypotrichidain the subclass Spirotrichia in the class Polyhymenophorea. The order Heterotrichida comprises the subordersHeterotrichina, Clevelandellina, Armophorina, Coliphorina, Plagiotomina and Licnophorina.
Small and Lynn [37], accepting in part the hypothesis bySeravin and Gerassimova [33], divide the phylum Ciliophora into three subphyla: Postciliodesmatophora, Rhabdophora and Cyrtophora. The first includes the classesKaryorelictea and Spirotrichea, and the class Spirotricheacomprises the orders Heterotrichida, Armophorida, Plagiotomida, Licnophorida, Clevelandellida and Odontostomatida. The suborder Coliphorina is included in the orderHeterotrichida. In 1985, Small and Lynn divided the classSpirotrichea into three different subclasses: Heterotrichia,Choreotrichia and Stichotrichia and included in the firstone the orders aforementioned, the order Odontostornatida and the new order Phacodiniida.
Taking into account our results on the general morphology, morphogenesis of bipartition and conjugation inC. medusula as well as the ultrastructural data of this species [30] we are inclined to agree with Small and Lynn [37,38] that the suborder Armorphorina (including C. medusula) should be considered as an order.
According to Fernandez-Leborans [17] the members ofsuborder Armophorina come from the suborder Heterotrichina. This evolutionary transition is caused by several
120 . A. Martin-Gonzales, S. Serrano and D. Fernandez-Galiano
types of processes (Blepharisma~ Bothrostoma~ Metopus~ Brachonella~ Cirranter~ Caenomorpha). In thefirst place there would have to be a process of perizonation[19]; in other words, several specialized somatic kinetiesor kinetal fragments would contribute to the oral infraciliature in the process of nutrition. In the second place,there is a torsion on the oral structures which would leadto a progressive horizontal location of the peristome, andfinally a reduction of the posterior somatic infraciliaturewould take place.
Unfortunately, there are no ultrastructural and stomatogenic data on the intermediate genera between Blepharisma and Caenomorpha supporting this hypothesis, andtherefore it is not possible to establish comparative relations at these two essential levels. Consequently, the studies by Bohatier [3] and Pelvat and de Haller [26] on otherheterotrichous ciliates which conclude that the phylogenetical line Blepharisma ~ Climacostomum ~ Stentor [19]is doubtful if one bears in mind the results obtained onmorphogenesis, and it is highly probable that these ciliatescome from different evolutionary lines.
Acknowledgements
This work was supported by the "Comision Asesorade Investigaci6n Cientffica y Tecnica" (Proyecto 0754/81).
References
1 Bhandary A. V. (1960): Conjugation in Blepharisma undulans americanum. ]. Protozool., 7, 250-255.
2 Bohatier]. (1978): Contribution a l'etude de la morphogenese de regeneration chez les protozoaires cilies des genresDileptus et Condylostoma. These de l'Universite de Clermont-Ferrand.
3 Bohatier]., Tuffrau M. and Tuffrau H. (1976): Morphogenese de regeneration dans Ie genre Condylostoma (ciliesheterotriches). Protistologica, 12, 295-306.
4 Burchill B. R. (1967): Conjugation in Stentor coeruleus. J.Protozool., 14, 683-687.
5 Cohen]., Adoutte A., Grandchamp S., Houdebine L. M. andBeisson]. (1982): Immunocytochemical study of microtubular structures throughout the cell cycleof Paramecium. Biol.Cell., 44, 35-44.
6 Corliss J. O. (1973): Evolutionary trends in patterns ofstomatogenesis in the ciliated protozoa. ]. Protozool., 20(suppl.), 506.
7 Corliss]. O. (1979): The ciliated protozoa: characterization,classification and guide to the literature. 2. ed., p. 455. Pergamon Press.
8 Corliss J. O. and Lorn ]. (1985): An annoted glossary ofprotozoological terms. In: Hutner S. H., Lee j. j. and BoveeE. C. (eds.): An illustrated guide to the protozoa,pp. 393-575. Allen Press, Kansas, USA.
9 Dryl S. (1959): Antigenic transformation in Parameciumaurelia after homologous antiserum treatment during autogamy and conjugation.]. Protozool., 6 (suppl.), 25.
10 Dubochet C. F., Peck R. K. and Haller G. de. (1979): Morphogenesis in the heterotrich ciliate Climacostomum virens.I. Oral development during cell division. J. Protozool., 26,218-226.
11 Ehret C. F. and MacArdle E. W. (1974): The structure ofParamecium as viewed from its constituent levelsof organiza-
tion. In: Van Wagtendonk W. J., (ed.): Paramecium: a current survey, pp. 263-337. Elsevier, Amsterdam, New Yorkand London.
12 Ellis I. M. (1937): The morphology, division, and conjugation of the salt-marsh ciliate Fabrea salina Henneguy. Univ.Calif. Publ. Zool., 41, 343-388.
13 Fernandez-Galiano D. (1976): Silverimpregnation of ciliatedprotozoa: procedure yielding good results with the pyridinated silver carbonate method. Trans. Amer. Micros. Soc.,95, 557-560.
14 Fernandez-Galiano D. (1977): Le comportement descinetodesmes pendant la division de Paramecium caudatum.XVI Reunion Ann. G. P. L. F. Inst. Univ. Biol. Marine. Bordeaux I.
15 Fernandez-Galiano D. (1978): Le comportement descinetodesmes pendant la division de Paramecium caudatum.Protistologica, 15,291-294.
16 Fernandez-Galiano D. and Fernandez-Leborans G. (1980):Caenomorpha medusula Perty, 1852 (Heterotrichida,Armophorina): nouvelles donnees sur la ciliature et l'infraciliature. Protistologica, 16, 5-10.
17 Fernandez-Leborans G. (1981): Sur la phylogenie desHeterotrichina Stein, 1867 des Armophorina Jankowski,1964 (Ciliophora, Heterotrichida). C. R. Acad. Sc. Paris,292, 821-824.
18 Grain J. (1969): Le cinetosome et ses derives chez les cilies.Ann. Biol., 8, 53-97.
19 Jankowski A. W. (1964): Morphology and evolution ofCiliophora. III. Diagnosis and phylogenesis of 53 sapropelebionts, mainly of the order Heterotrichida. Arch. Protistenk., 107, 185-294.
20 Kirby H. (1950): Materials and methods in the study of protozoa, pp. 1-72. Univ, California Press, Berkeley.
21 LevineN. D., Corliss]. 0., Cox F. E. G., Deroux G., GrainJ.,Honigberg B. M., Leedale G. F., Loeblich A. R. III, Lorn J.,Lynn D., Merinfeld E. G., Page F. c., Poljansky G., SpragueV., Vavra]. and Wallace F. G. (1980): A newly revised classification of the protozoa. ]. Protozool., 27, 37-58.
22 Miyake A. (1981): Physiology and biochemistry of conjugation in ciliates. In: Levandowsky M. and Hutner S. H. (eds.):Biochemistry and physiology of protozoa, pp. 126-233.Academic Press.
23 Muslow W. (1913): Die Conjugation von Stentor coeruleusund Stentorpolymorphus. Arch. Protistenk., 28, 363-388.
24 Ng S. F. and Williams R.]. (1977): An ultrastructural investigation of 1800 rotated ciliary meridians in Tetrahymenapyriformis. ]. Protozool., 24, 257-263.
25 Ng S. F. and Newman A. (1984): The role of the micronucleus in stomatogenesis in sexual reproduction of Parameciumtetraurelia: micronuclear and stomatogenic events. Protistologica, 20, 43-69.
26 Pelvat B. and Haller G. de. (1979). - La regeneration del'appareil oral chez Stentorcoeruleus: etude au protargol etessai de morphogenese comparee. Protistologica, 15,369-386.
27 Prandtl H. (1906): Die Konjugation von Didinium nasutumO. F. M. Arch. Protistenk., 7,229-258.
28 Puytorac P. de and Grain]. (1976): Ultrastructure du cortexbuccal et evolution chez les cilies, Protistologica, 12, 49-67.
29 Raikov I. B. (1972): Nuclear phenomena during conjugationand autogamy in ciliates. In: Chen T.-T. (ed.): Research inprotozoology, vol. 4, pp. 147-289. Pergamon Press, Londonand New York.
30 Rodrigues de Santa Rosa M. (1976): Observations sur l'ultrastructure du cilie heterotriche Armophorina:Caenomorpha medusula Perty, 1852. J. Protozool., 23(suppl.), 19A.
Cortical Morphogenesis and Conjugation in Caenomorpha medusula . 121
31 Santos S. M., Guinea A. and Fernandez-Galiano D. (submitted): Etude de l'infraciliature et du processus de bipartitionchez Nyctotherus oualis Leidy, 1850 (Ciliophora, Heterotrichida).
32 Sawyer H. R. and Jenkins R. A. (1977): Stomatogenic eventsaccompanying binary fission in Blepharisma. J. Protozool.,24, 140-149.
33 Seravin L. N. and Gerassimova Z. P. (1978): A new macrosystem of ciliates. Acta Protozool., 17, 399-318.
34 Serrano S., Martin-Gonzalez A. and Fernandez-Galiano D.(1986): Nuclear phenomena and oral reorganization duringthe conjugation of Urocentrum turbo O. F. M. Arch. Protistenk. (in press).
35 Seshachar B. R. and Bai A. R. K. (1968): Conjugation inSpirostomum ambiguum Ehrbg. Acta Protozool., 6, 51-56.
36 Seshachar B. R. and Bhandary A. V. (1962): Observations onthe life-cycle of a new race of Blepbarisma undulans fromIndia. J. Protozool., 9, 265-270.
37 Small E. B. and Lynn D. H. (1981): A new macrosystem forthe phylum Ciliophora Doflein, 1901. Bio-Systems, 14,387-401.
38 Small E. B. and Lynn D. H. (1985): Phylum Ciliophora. In:Hutner S. H., Lee J. J. and Bovee E. C. (eds.): An illustratedguide to the protozoa, pp. 393-575. Allen Press, Kansas,USA.
39 Tamm S. L.,Sonneborn T. M. and Dippell R. V. (1975): Therole of cortical orientation in the control of the direction ofciliary beat in Paramecium. J. Cell. BioI., 64, 98-112.
40 Villeneuve-Brachon S. (1940): Recherches sur les ciliesheterotriches: cinetome, argyrome, myonemes, formesnouvelles on peu connues. Arch. Zool. Exp. Gen., 82, 1-180.
41 Wichterman R. (1937): Division and conjugation in Nyctotherus cordiformis (Ehr) Stein (Protozoa, Ciliata) with special reference to the nuclear phenomena. J. Morphol., 60,563-611.