Our understanding of the biology of cytokines, garnered over the past 30 years, owes much to the use of endotoxins and lipopolysaccharides (LPS). This has given rise to the concept that endotoxin/LPSis the main cytokine-stimulating component of bacteria, in spite of the fact that Gram-positive bacte-ria can also stimulate cytokine synthesis. However, during the past decade a growing number of reportshave demonstrated that a range of bacterial components and secreted products also have the capacity to stimulate cytokine gene transcription. In this review the nature of these ‘non-LPS’ cytokine-inducing molecules will be described and their activitywill be related to the currently understood mechanismof action of endotoxin/LPS.
CYTOKINE INDUCTION BY LIPOPOLYSACCHARIDES
LPS is a potent stimulator of a range of cells inducing the synthesis of many cytokines including thepro-inflammatory IL-1, IL-8, TNF and anti-inflamma-tory/immunomodulatory cytokines such as IL-10. Thecytokine-inducing actions of LPS will be dealt withbriefly to give a mechanistic foundation for the discus-sion of the actions of the many other cytokine-inducingmolecules from bacteria. The reader is referred to arange of recent reviews on various aspects of the biol-ogy of endotoxin/LPS.1–6
LPS: definition and composition
The literature on endotoxin/LPS reveals many contradictory claims about potency and efficacy. Thisrelates to the general confusion regarding nomencla-ture, purity and the physical state of LPS. The nomen-clature of this cell wall component has been clarified byHitchcock et al.7 and can be summarized as follows. Theterm lipopolysaccharide (LPS) “should be reserved forpurified bacterial extracts which are reasonably free ofdetectable contaminants. particularly protein.” Suchpreparations may be obtained using the Westphalextraction procedure as a starting point.8 In contrast,the term endotoxin should be used to refer to “prod-ucts of extraction procedures which result in macro-molecular complexes of LPS, protein and phospho-lipid”. Complexes of this type can be obtained byextraction of bacteria with trichloroacetic acid, butanolor EDTA.
The purity of the LPS preparations used by inves-tigators often remains inadequately specified or evenignored. Endotoxin is a mixture of components, there-fore purity is not an issue although its compositionshould be specified by investigators. Unlike most othercompounds, it is difficult to define a ‘pure’ prepara-tion of LPS using conventional criteria. For example,molecular weight cannot be used because within a bac-terial population, and even within an individual cell,LPS is present in a number of forms, due to variationin the length of the O-antigenic side chain. However,this does not relieve investigators from the onus ofensuring that the preparations are free, as far as possible, of contaminants such as proteins, nucleic acids,polysaccharides and lipids. The presence of such compounds not only introduces unknown (possibly biologically-active) components into the assay systembut also results in an inaccurate assessment of the true potency of the ‘LPS’ preparation. Many investigators obtain their LPS from commercial suppliers and there remains an unwarranted acceptanceof the purity of such preparations. Analyses carried outin our laboratories have shown that some can containup to 60% protein.
CYTOKINE INDUCTION BY BACTERIA: BEYONDLIPOPOLYSACCHARIDE
Brian Henderson1 and Michael Wilson2
From 1Maxillofacial Surgery Research Unit and 2Department ofMicrobiology, Eastman Dental Institute for Oral Health CareSciences, University of London, 256 Gray’s Inn Road, LondonWC1X 8LD
Correspondence to: Professor Brian HendersonReceived 14 June 1995; revised and accepted for publication 23
LPS generally has a low solubility and Galanos etal.9 have shown that the biological activities of LPS aregreatly affected by its physical state. In the case ofSalmonella spp., for example, extracted LPS exists as anegatively-charged molecule bound to amines such asputrescine and spermine. Removal of these bases byelectrodialysis followed by neutralization with triethyl-amine (TEA) or metal ions results in LPS with differ-ent solubilities and biological activities. For example,the TEA salt of Sal. abortus-equi LPS is more toxic tomice and has greater pyrogenicity in rabbits than thecorresponding partially-soluble calcium salt. The cal-cium salt, however, is more toxic to rats.9 The effects ofdifferent salt forms of LPS on cytokine induction hasreceived little attention.
LPS receptors
The nature of the cell surface receptor for LPSremains to be discovered. Lei and Morrison,10 usingphotoaffinity crosslinking, described a 70 kDa LPSreceptor on the surfaces of murine lymphocytes andmacrophages and prepared hamster monoclonals tothis protein which activated macrophages and pro-tected mice against endotoxin lethality.11,12 However,Dziarski13 has recently reported that this receptor is, infact, cell bound albumin. Other workers using similarcrosslinking strategies have described a range of LPS-binding cell surface proteins.6 Unfortunately, none ofthese proteins have been shown to have kinetic or sig-nalling properties consonant with being LPS receptors.
Currently, two proteins, CD1414,15 and the β2-inte-grin CD11c/CD1816 are believed to function as LPSreceptors. The former is a 55–60 kDa glycosylphos-phatidylinositol (GPI)-anchored protein expressedstrongly on myelomonocytic cells but not on mes-enchymal cells. While CD14 will bind to LPS, the affin-ity of binding is significantly increased by interactionwith a serum acute phase protein – LPS-binding pro-tein (LBP).17 Trans-fection of the murine pre-B cell70Z/3, which only responds to high concentrations ofLPS, with CD14 increases responsiveness 1000-fold.18
However, two problems exist: (1) CD14 does not havean intracellular signalling domain and (2) the affinity ofbinding of LPS to CD14 is low (1027 to 108 M21).19
However, neutralizing antibodies to CD14 will inhibitthe activity of LPS and can be used to define if cell acti-vation is dependent on this protein.20 Soluble CD14 ispresent in relatively high concentrations in blood andcan form a complex with LPS-LBP which can bind andactivate cells lacking membrane CD14.21 It can also actas an antagonist with myelomonocytic cells whichexpress CD14.22 There is increasing evidence that theinteractions between LPS/LBP/CD14 is designed bothto enhance cellular responses to LPS and to remove andinactivate LPS.23
LPS-induced intracellular signalling
LPS induces a multitude of cellular changes andonly the early responses to LPS, which include induc-tion of cytokine gene transcription, will be dealt with.As one might expect with an agonist able to induce suchprofound cellular activation, most intracellular sig-nalling pathways have been claimed to be activated byLPS/endotoxin. Of course, as many studies have beendone with endotoxin which, as has been detailed, con-tains many bioactive proteins in addition to LPS it isnot clear how many of the activated pathways are LPS-specific. The two most consistent findings are that LPS/endotoxin activates the inositol trisphos-phate/protein kinase C pathway24,25 and stimulates protein tyrosine kinases.26 LPS/endotoxin has also been shown to activate the Raf-1/MAP kinase pathway.27 The possible importance of tyrosine kinasesin LPS toxicity has been shown by the protective effect of a selective tryphostin-type tyrosine kinaseinhibitor in mice receiving lethal levels of endotoxin.28
However, to date no LPS-specific tyrosine kinases havebeen recognized.
BACTERIAL CYTOKINE-STIMULATINGCOMPONENTS OTHER THAN LPS
Table 1 lists the components and products of bac-teria which have been reported to stimulate cytokine synthesis. Many of these molecules are constituents of the bacterial cell wall and the structures of the Gram-positive and Gram-negative cell walls are shownin Figure 1. Unless rigorous purification procedures areemployed any bacterial constituent may be contami-nated with LPS. Even when active components havebeen derived from Gram-positive bacteria, the pres-ence of contaminating LPS must be considered a pos-sibility because of its ubiquity and chemical and physicalstability. The minimum requirement for demonstratingthat any particular activity is not due to contaminatingLPS would be its abrogation by heat (60–100°C) or pro-teases (in the case of proteinaceous components) or bypolymyxin B or neutralizing antibodies to CD14(although high LPS concentrations will overcome thisblockade). Unfortunately, Laude-Sharp et al. haveshown that a negative reaction in the Limulus assaydoes not exclude the presence of LPS.29
Constituents of Gram-positive bacteria
Many of the major pathogenic bacteria of man areGram-positive, including mycobacteria, staphylococciand streptococci. Surprisingly little attention has beenpaid to the capacity of these organisms to stimulatecytokine production, in spite of the fact that the diseasesthey cause—tuberculosis, leprosy and septic shock-like
270 / Henderson and Wilson CYTOKINE, Vol. 8, No. 4 (April 1996: 269–282)
conditions are clearly inflammatory lesions involvingcytokine network interactions.
Lipoarabinomannan (LAM)LAM is a major constituent of Mycobacterial cell wallsand, along with structural analogues (lipomannan [LM]and phosphatidylinositol mannosides [PIM] areregarded as functional equivalents of LPS. They appearto exhibit immunoregulatory and anti-inflammatoryactions which favour the survival of the mycobacteria,including suppression of T lymphocyte proliferationthrough interference with antigen processing,30 inhibi-tion of macrophage activation by γ-interferon31,32 andscavenging of oxygen-derived free radicals.33 Morenoand coworkers30,34 were the first to report that LAMcould stimulate monocytes/macrophages to releaseTNF-α at concentrations as low as 100 ng/ml.Surprisingly, polymyxin B bound to and inactivatedLAM, demonstrating a similarity in structure betweenLAM and LPS and giving a clue to the LAM receptor.Treatment of the LAM with dilute alkali significantlydiminished the TNF-stimulating activity suggestingthat the o-acyl groups are responsible for stimulation
of cytokine synthesis.34 Barnes et al.35 demonstrated thatLAM stimulated human PBMCs to transcribe mRNAfor cytokines normally thought of as being macrophageproducts—IL-1α and β, IL-6, IL-8, GM-CSF, TNF andIL-10 — but not for T cell ‘lymphokines’ IFN-γ, IL-2, IL-3 or IL-4 although whole M. tuberculosis was cap-able of inducing ‘lymphokine’ transcription. ModifiedLAM (LM and PIM) also stimulated the same cytokineprofile; however, deacylation of LAM almost totallyinhibited its capacity to induce cytokine synthesisrevealing that the activity was associated with the phos-phatidylinositol end of the molecule.
The cellular receptor(s) for LAM have not beenidentified although it has been shown that the phago-cytosis of mycobacteria is dependent on binding to man-nose and receptors for complement and the leukocyte β2-integrins.36 Given the inhibitory effect of polymyxinB, the possibility also exists that LAM acts via CD14-LPS receptors. It has been demonstrated that LAM andLPS activate human monocyte IL-6 gene expression byan identical pathway involving NF-κB and NF-IL-6.37
LAM preparations from virulent or attenuated strains
Bacterial cytokine-inducing factors / 271
TABLE 1. Bacterial factors capable of stimulating cytokine synthesis
Gram-positive componentsLipoarabinomannan, lipomannans and phosphatidylinositol
mannosidesPurified protein derivative (PPD)Mycobacterial heat shock proteinsProtein ALipoteichoic acidStaphylococcal superantigens
Common to all bacteria: Cell Wall ComponentsCell surface proteinsFimbriae and piliLipopeptidesLipoproteinsMuramyl dipeptidePeptidoglycanPolysaccharides
Common to all Bacteria: Extracellular componentsToxinsProteasesSuperantigens
Figure 1. Diagrammatic representation of a cross-section throughthe wall of a typical Gram-positive (a) or Gram-negative (b) bacterium
CM 5 cytoplasmic membrane, PEP 5 pepidoglycan, LTA 5 lipo-teichoic acid, TA 5 teichoic acid, LPS 5 lipopolysaccharide, POR 5 porin, OMP 5 outer membrane protein, PL 5 phospho-lipid, LP 5 lipoprotein, PP 5 periplasmic protein.
of mycobacteria differ markedly in their biologicalactions with the former being unable to stimulatemurine resident peritoneal macrophages to releaseTNF-α .38 The LAM from an avirulent mycobacterium,strain H37Ra, is able to stimulate the transcription ofthe immediate early response genes c-fos, JE and KCand the production of TNF-α. In contrast LAM from avirulent strain, Erdman, fails to trigger these genes andthe production of TNF-α.39,40 This capacity to produceLAM which is unable to stimulate a macrophageresponse is now seen as a key determinant in mycobac-terial virulence and the structural basis of the potencyof LAMs is under investigation.
Other components of mycobacteria stimulatecytokine synthesis. These include a 58 kDa protein ableto stimulate human monocyte TNF-α synthesis,41 puri-fied protein derivative (PPD) capable of stimulating arange of cytokines including IL-1, IL-6, TNF and GM-CSF42 and the 65 kDa mycobacterial heat shock proteinwhich stimulates TNF production.43 In addition, PPDfrom M. tuberculosis has been reported to preferen-tially stimulate a Th1 pattern of cytokine productionfrom human peripheral blood.44
Protein AThis extracellular protein from Staph. aureus stimulateshuman mononuclear cells to release IL-1, IL-4, IL-6,TNF-α and IFN-γ over the concentration range 1–10 µg/ml and was more active than other cell wall constituents including muramyl dipeptide, muramicacid and teichoic acid.45
Teichoic acidsThe teichoic acids are major components of the cellwalls of Gram-positive bacteria, are highly antigenicand have been proposed to be virulence factors.Riesenfeld-Orn et al.46 reported that lipoteichoic acid(LTA) from pneumococci stimulated human mono-cytes to release IL-l but not TNF. In contrast, the LTAfrom Streptococcus faecalis stimulated murine mono-nuclear cells to release both cytokines.47 In this latterreport, deacylation of the LTA abolished the cytokine-stimulating activity which was retained by the phos-phatidylglycolipid fraction derived by acid hydrolysis.In comparative studies of the LTAs from a range ofGram-positive bacterial species, Bhakdi and col-leagues48 reported major differences in their capacityto stimulate human monocytes to produce cytokines(IL-1β, IL-6, TNF-α). LTAs from several enterococcalspecies were as efficacious, but less potent, than a crudepreparation of E. coli LPS. In contrast, LTAs fromorganisms such as Staph. aureus, Strep. mutans andLeuconostoc mesenteroides were inactive. The additionof polycations (poly-L-arginine or poly-L-lysine) tomonocytes stimulated with LTA inhibited cytokineinduction although they had no effect on the cytokine-
stimulating activity of LPS. Analysis of the kinetics ofcytokine stimulation revealed that very short exposureof cells to LTA (5–30 min) was sufficient to triggercytokine production. These results suggest that the cel-lular receptor for LTA is distinct from that of LPS andmay possibly be the macrophage scavenger receptor.4
Blockade of CD14 also failed to inhibit LTA-inducedcytokine induction.49 The cytokine-stimulating LTAswere also shown to be able to stimulate murine bonemarrow macrophages to release NO.50 While Bhakdiand coworkers48 failed to demonstrate that Staph.aureus LTA stimulated cytokine production, Standifordet al.51 reported that LTA from this organism, and fromStrep. pyogenes, stimulated human peripheral bloodmonocytes to produce IL-8.
Teichoic acids have been subject to comparativelydetailed study and the conclusions appear to be thatthey are only weak activators of cytokine synthesiswhich probably act by a distinct mechanism to LPS.
Staphylococcal superantigensMost superantigens51 are produced by the Gram-positive bacterium Staph. aureus and by streptococcalspecies. Among the superantigens released by Staph. aureus are the group of enterotoxins consistingof six different serotypes (staphylococcal enterotoxinsA-E), toxic shock syndrome toxin 1 (TSST-1) and an exfoliative toxin.53 Strep. pneumoniae54 producespyrogenic exotoxins (SPEA, B and C). Other bacteria produce superantigens but, to date, most studies have concentrated on the superantigens fromStaph. aureus.
Injection of mice with a superantigen such as SEB resulted in the rapid appearance of a variety of cytokines such as TNF, IL-1, IL-6 and IFN-λ in the serum and, in D-galactosamine sensitized mice, the injection of 2 µg SEB per mouse resulted in 50%mortality which could be blocked by neutralizing TNF-α/β55 or by administering IL-10.56 The mechanismof cytokine generation in vivo is obviously complex,involving antigen-presenting cells, B lymphocytes andT lymphocytes. However, in cell culture, it appears thatbinding of the superantigens to class II molecules is thetrigger for cytokine production57 and that the subse-quent synthesis of cytokines such as IL-1 is dependenton protein tyrosine phosphorylation.58 The involve-ment of the MHC is clearly seen when class II-deficientmice are used, these animals being resistant to the effects of staphylococcal enterotoxins.59 200 ng/mlof SEA was sufficient to induce maximal production of TNF-α by cultured human monocytes with an IC50
of ,50 ng/ml.60
Synergistic interactions between superantigensand endotoxins which could be important in the induc-tion of tissue pathology have been reported (e.g. Refs59–62).
272 / Henderson and Wilson CYTOKINE, Vol. 8, No. 4 (April 1996: 269–282)
Constituents from Gram-negative bacteria
Gram-negative bacteria have a much thinner pep-tidoglycan layer than Gram-positive organisms andexternal to this is a complex structure termed the outermembrane which contains, on its outer surface,lipopolysaccharide (Fig. 1). In addition, the outer mem-brane of E. coli contains three or four major, and 50 orso less abundant, proteins which are termed outer mem-brane proteins. These include structural proteins suchas lipid A-associated proteins, pore-forming porins andvarious other proteins and lipoproteins.
Lipid A-associated proteinsIn 1968 Sultzer63 reported a spontaneous mutation inthe C3H mouse strain which rendered animals refrac-tory to LPS. However, it was later reported that thishyporesponsiveness depended on the method used to prepare the LPS. Mild extraction resulted in LPS preparations which were mitogenic for C3H/HeJ lym-phocytes but more extensive extraction, yielding pro-tein-free LPS, resulted in loss of activity.64 Theexplanation for these findings was the presence of anumber of proteins which co-purified with the LPS andwhich appeared to be associated with the lipid A moi-ety. These lipid A-associated proteins (LAP) weredemonstrated to be the B cell mitogen in butanolextracts of LPS, being lost during more extensiveextraction.65,66 LAP appears to be an important viru-lence factor, as immunization of Salmonella-hypersus-ceptible mice with LAP-LPS complexes, but not withLPS, protects against the lethality of Sal. typhimuriuminfection.67 Furthermore, LAP from Salmonella specieshave been shown to have potent adjuvant activity cap-able of enhancing murine antibody responses both invitro and in vivo.68–70 Hogan and Vogel71 have claimedthat LAP represent a ‘second signal’ for macrophageactivation.
McCabe et al.72 have claimed that LAP prepara-tions from Sal. typhimurium have IL-1-like properties,injection of LAP inducing the acute phase reactant,serum amyloid A (SAA) in mice. However, LAP failedto stimulate macrophages to produce SAA-inducingactivity (believed to be IL-1) in culture. In this study,LAP were also found to be active in the lymphocyte-activating factor (LAF) assay and to act as a co-stimu-latory factor for the proliferation of resting human Tlymphocytes. LAP was also demonstrated to induce theformation of granulopoietic colonies when added tohuman peripheral blood and bone marrow progenitorcells which had been depleted of accessory cells (mono-cytes, T and B lymphocytes).73 The stability of LAP toheat and proteolysis was also shown in this study. Poratet al.74 isolated LAP from E. coli and largely confirmedMcCabe’s72,73 studies, however, they found that T cellspartially mediated the effect of the LAP and that the formation of granulocyte/macrophage colonies
induced by LAP could be blocked by a neutralizing anti-serum to IL-1β . These workers also reported that theactive protein in the LAP preparation had a molecularweight of 17 kDa. Mangan and coworkers75 comparedthe ability of LPS and LAP from Sal. typhimurium tostimulate IL- 1 synthesis. Significant amounts of mRNAfor IL-1β were induced in human monocytes exposedto 1ng/ml LAP and the LAP induced a greater maxi-mal response than the LPS. We compared the ability ofLAP and protein-free LPS from the oral bacteriumActinobacillus actinomycetemcomitans to stimulate therelease of IL-1β, IL-6 or TNF-α from various humancells. In all instances the LAP was several log ordersmore potent than the LPS.76 The mechanism of intra-cellular signal transduction in B lymphocytes exposedto LAP is complex, with inhibitors of PKC and tyrosinekinases, as well as pertussis toxin, inhibiting lympho-cyte mitogenesis,77 a similar picture to that found withLPS.
PorinsA number of reports appeared in the late 1970s andearly 1980s to suggest that outer membrane proteinsand lipoproteins from various Gram-negative bacteriacould stimulate immunologically-competent cells, par-ticularly B cells, in in vitro assays.78–81 Porins were alsoshown capable of activating leukocytes.82,83 However,the most extensive studies of porins have been con-ducted by Tufano and coworkers at the University ofNaples. They have shown that porins bind specificallyto leukocytes, but the nature of the receptor(s) has notbeen defined. Porins inhibited the oxidative burst ofPMNs and altered cell morphology,84 but stimulated theproduction of the potent inflammatory mediator,platelet activating factor (PAF).85 Similar effects werereported with porins isolated from Helicobacterpylori.86 The porins from Sal. typhimurium also stimu-lated human vascular endothelial cells to synthesizePAF.87 In addition to having these in vitro actions, injec-tion of 0.3–30 mg purified Sal. typhimurium porins intorat paws induced a dose-dependent oedema whichpeaked after 2–3 h and was still marked after 5 h.Injection of similar concentrations of LPS was withouteffect. Oedema was unaffected in animals which hadbeen decomplemented, could be moderately decreasedby indomethacin and was significantly reduced by dexamethasone. When porins were added to rat peri-toneal cells they induced a concentration-dependentrelease of histamine but had little effect on prostacyclinproduction.88 Porins were also demonstrated to kill D-galactosamine-sensitized LPS-responsive and non-responsive mice. Ten ng of porins killed 20% of animalsand injection of 100 ng produced 80–90% lethality; asimilar dose response being found with the LPS fromSal. typhimurium. The lethal effect of the porin prepa-ration could be completely blocked by preadministra-
Bacterial cytokine-inducing factors / 273
tion of a neutralizing antiserum to TNF-α but was notabrogated by an excess of polymyxin B. Porins were alsofound to be pyrogenic in rabbits and to elicit a localizedSchwartzman reaction when used as the sensitizing andeliciting agent. Both the fever and the Schwartzmanreaction were unaffected by the administration ofpolymyxin B, ruling out any contribution from LPS tothe biological responses.89
Isolated porins from Sal. typhimurium,90 Y. entero-colitica91 or H. pylori86 were shown to stimulate humanmonocytes and lymphocytes to release a range of pro-inflammatory and immunomodulatory cytokinesincluding IL-1, IL-4, IL-6, IL-8, TNF-α, GM-CSF andIFN-γ in a dose-dependent manner. The porins and LPSfrom Sal. typhimurium showed similar linear doseresponses (10 ng/ml to 1 µg/ml) in promoting the syn-thesis of TNF and IL-6. In contrast, the porins were rel-atively inactive in stimulating IL-1α release frommonocytes or IFN-γrelease from human lymphocytes.90
Porins are major components of Gram-negtive bac-teria and their range and potency of actions and theirstability to heat and proteases suggests that they play arole in bacterial virulence. The key unknowns are: (1)what receptors do they trigger; (2) what is the intracel-lular transduction pathway or pathways involved incytokine generation and (3) do porins interact withendotoxins in an additive or synergistic manner.
Miscellaneous outer membrane proteinsOuter membrane proteins of Shigella flexneri stimulatemurine macrophages to release TNF and IL-6.91
Borrelia burgdorferi, a tick-borne spirochaete andcausative agent of Lyme disease, contains two outermembrane lipoproteins termed OspA and OspB whichhave been purified to homogeneity and shown to dose-dependently stimulate the synthesis of both TNF-α andIL-6 by bone marrow-derived macrophages from LPS-unresponsive mice and IL-6 production by humanmonocytes and vascular endothelial cells. IL-6 produc-tion by murine macrophages showed a linear doseresponse over the range 10 ng to 1 µg/ml and additionof IFN-γ enhanced cytokine synthesis two-fold.93,94
The possible interactions between outer membraneproteins and other virulence factors such as LPS remainto be studied. However, a 39 kDa outer membrane pro-tein from Proteus mirabilis which is mitogenic for Bcells95 has been shown to inhibit the LPS-induced acti-vation of macrophage oxygen-derived free radical for-mation as well as the LPS-induced synthesis of IL-1 bymurine macrophages.96 Thus, in addition to inducingcytokine synthesis, at least one outer membrane pro-tein appears to be able to downregulate bacterially-induced cytokine synthesis.
Lipoproteins and lipopeptides from the outermembranes have also been shown to possess immuno-potentiating activity.97–100 Lipoproteins from Bor.
burgdorferi and Treponema pallidum inducedmacrophage TNF-α synthesis as assessed by biologicalactivity and CAT reporter assay. Treatment of theseproteins with proteinase K did not significantly reduceactivity suggesting that lipopeptides may constitute theactive moiety.101 Indeed, the synthetic lipopeptide N-palmitoyl-S-(2,3-bis(palmitoyloxy)-2RS-propyl)-(R)-cysteinyl-alanyl-glycine (Pam3Cys-Ala-Gly), asynthetic analogue of the N-terminus of a bacteriallipoprotein does stimulate the synthesis of IL-1, IL-6and TNF-α by murine macrophages.102 The activelipoprotein from T. pallidum has been cloned and it hasbeen shown that lipid modification of this protein isessential for its ability to stimulate TNF synthesis.103 Thesynthetic lipopeptide CGP 31362 stimulates TNF-α synthesis via protein tyrosine phosphorylation andmitogen-activated protein (MAP) kinases in LPS-unresponsive murine macrophages.104 Thus LPS andlipopeptides both appear to stimulate similar intracel-lular signal transduction pathways.
Constituents produced by both Gram-positive andGram-negative bacteria
Components of the cell wall such as peptidoglycan,muramyl dipeptide, cell wall polysaccharides, pili andfimbriae, proteins loosely associated with the cell, pos-sibly present in the capsule, and true extracellular mol-ecules such as toxins, lysins and pore-forming proteinshave all been reported to induce cytokine synthesis.
Non-proteinaceous cell wall componentsBoth Gram-positive and Gram-negative bacteria havethe complex peptide-containing carbohydrate, pepti-doglycan (PG) in their cell walls. Injection of PG/car-bohydrate complexes from a variety of pathogenic orcommensal bacteria can induce acute and chronic in-flammatory states. Most work has been done with PG-PS complexes from Group A streptococci and these caninduce a variety of local and systemic lesions, depend-ing on the species, strain and route of administration.The pathogenic properties of such complexes have been reviewed by Schwab.105,106 The toxicity of these complexes appears to be due, in part, to the stimulationof secretion of various pro-inflammatory cytokines and associated low molecular weight lipids and free radicals.106
Peptidoglycan. Attention has focused on the syntheticmuramyl dipeptide [N-acetyl-muramyl-L-alanyl-D-isoglutamine] (MDP), the smallest common structuralunit of adjuvant peptidoglycans. MDP and its analoguescan stimulate the synthesis of a wide variety of cytokinesby a range of cell populations and the literature has beenwell reviewed.107–109 However, MDP is a synthetic mol-ecule which is not a product of cellular degradation of
274 / Henderson and Wilson CYTOKINE, Vol. 8, No. 4 (April 1996: 269–282)
peptidoglycan and therefore we feel that resultsobtained with this molecule have no physiological orpathophysiological relevance. There have been fewstudies of the cytokine-stimulating ability of peptido-glycan, probably because of the problems involved indealing with a poorly soluble material with a wide rangeof molecular masses and structures. Riesenfeld-Orn etal.46 were the first to investigate Gram-positive cell wallsas inducers of cytokine synthesis and reported that pep-tidoglycan from Strep. pneumoniae stimulated humanmonocytes to release IL-1 but not TNF. Peptidoglycan-containing teichoic acids were claimed to be the mostpotent inducers of IL-1 synthesis, being 10 000 timesmore potent than E. coli LPS. Keller et al.110 comparedthe ability of peptidoglycan from Moraxella catarrhalis,Staph. aureus and Bacillus subtilis to stimulate rat bonemarrow macrophages to release TNF-α and found allthree were equiactive. In a comparative study of theability of whole Staph. aureus, Staph. epidermidis, andtheir respective peptidoglycans, to stimulate TNF-αrelease from human monocytes, it was demonstratedthat, whole cells were more effective stimuli than theisolated cell wall components. Thus 107 staphylococcalcells, corresponding to 0.1 µg of peptidoglycan, pro-duced the same amount of TNF as 1–10 µg of purifiedpeptidoglycan.111 The explanation for this difference inactivity may relate to degradation of the peptidoglycanduring isolation; to differences in steric interactionbetween whole cell PG and isolated material, or to syn-ergistic interactions between PG and other cell wallcomponents in whole bacteria. Indeed, degradation ofthe PG by enzymes or by sonication decreased its capac-ity to induce TNF synthesis. Water-soluble peptidogly-can fragments from Staph. epidermidis stimulated theproliferation of spleen mononuclear cells from variousstrains of mice and the synthesis of various cytokinesincluding IL-l and GM-CSF. In addition, administrationto animals resulted in the induction of arthritis.112
Natural peptidoglycan breakdown products may have cytokine-inducing actions. Two recent papers report that N-acetylglucosaminyl-1,6-anhydro-N-acetylmuramyl-L-alanyl-D-isoglutamyl-m-diamino-pimelyl-D-alanine [G(Anh)MTetra], a naturallyoccurring breakdown product of peptidoglycan,induces the synthesis of IL-1β, IL-6113 and G-CSF 114 byhuman monocytes at concentrations as low as 50 ng/ml.This confirms the initial study of Dinarello andKrueger.115 The synthesis of all three cytokines can beblocked by inhibitors of protein kinase C but not byinhibitors of protein kinase A or tyrosine kinases, sug-gesting a different mechanism from LPS. Interestingly,the transcriptional control of these three cytokinesshowed distinct characteristics. Using run-on transcrip-tion assays, [G(Anh)MTetra] markedly increased IL-1β transcription, whereas it had much less effect on therate of transcription of the IL-6 gene. The increased
level of mRNA for G-CSF was shown to be due to stabilization of the mRNA transcripts. Using the pro-tein synthesis inhibitor cycloheximide, it was demon-strated that IL-6 mRNA expression depended on thesynthesis of new protein but that this was not the casefor IL-1β or G-CSF.
Cell surface carbohydrates. Capsular polysaccharidesplay an important role in protecting bacteria from hostdefences such as phagocytosis and enable adhesion tohost tissues. The mucoid exopolysaccharide from Ps. aeruginosa116 and a capsular-like, serotype-specificpolysaccharide antigen from A. actinomycetemcomi-tans117 both stimulated murine myelomonocytic cells torelease IL-1 in the presence of excess polymyxin B.Benabdelmoumene et al.118 showed that a purified poly-saccharide from Strep. mutans was capable of stimulat-ing human monocytes to produce both IL-1 and TNF-αover the dose range 10 to 75 µg/ml activity not beingblocked by polymyxin B. A similar dose response wasseen with a carbohydrate antigen of Actinomyces vis-cosus which stimulated IL-1, IL-6 and TNF synthesis bymurine macrophages.119 The relationship between thechemical structure of defined polysaccharides andcytokine synthesis was reported by Espevik et al.120 whoshowed that certain polysaccharides were as potent as(albeit fairly inactive) E. coli LPS and concluded thatβ1-4-linked polyuronic acids probably bind to the samereceptor as LPS. Injection of a type-specific polysac-charide from type II group B streptococci has beenshown to induce TNF-α in the serum and, at very highlevels, to induce lethality which can be blocked by a neutralizing antiserum to TNF.121
Proteinaceous cell surface componentsA number of proteins and proteinaceous structures (pili and flagella) are associated with the bacterial cellsurface.
Pili. These structures, also known as fimbriae, are oneof the means by which bacteria attach to surfaces,including eukaryotic cells.122 The leukocyte adhesionmolecules CDl l and CD18 are receptors for the type 1-fimbriated (mannose specific) E. coli.123 Injection ofisolated E. coli pili into mice elicited a local stimulationof IL-6 release.124 In in vitro studies, pili from E. coliwere shown to stimulate epithelial cell lines to releaseIL-6,125–127 although high concentrations of pili wererequired. In contrast, fimbriae from the oral periodon-topathogen Porphyromonas gingivalis stimulated IL-1gene expression and protein production by murinemacrophages at much lower concentrations (2–4 µg/ml:Ref. 128). Such fimbriae also induced the transcriptionof the KC gene, which encodes a neutrophil chemo-attractant in mouse peritoneal macrophages.129
Attachment of Sal. typhimurium to monolayers of
Bacterial cytokine-inducing factors / 275
human intestinal epithelium resulted in the trans-epithelial migration of human PMNs and the synthesisof the potent chemoattractant, IL-8. However, neutral-ization of this cytokine did not block PMN transmigra-tion, the authors suggesting that a novel transcellularchemotactic factor is induced by interaction of bacteriawith the epithelial cells.130 The invasin protein of Y.pestis, which is utilized by this organism to allow it toinvade cells, interacts with β1 integrins and acts as a co-stimulatory signal for the proliferation of CD4 lym-phocytes, a process accompanied by the synthesis ofTNF-α and IFN-γ.131
Miscellaneous cell surface-associated proteins. Numerousproteins appear to be loosely-associated with the bac-terial cell surface and can be easily removed by gentlesolubilization. The authors have shown that gentlesaline extraction of certain bacteria releases a pro-teinaceous fraction (which we have termed surface-associated material, SAM) which has potent biologicalactions. The SAM from a number of bacteria are ableto stimulate bone breakdown in vitro by mechanismsdependent on IL-1 and/or TNF synthesis.132–134 SAMfrom a number of oral bacteria has subsequently beenfound to be capable of stimulating the release of sev-eral cytokines (IL-1, TNF, IL-6 and IL-8) from varioushuman cell populations including monocytes, gingivalfibroblasts, PBMCs and neutrophils.76,135 The SAMsfrom some oral bacteria were considerably more potentcytokine inducers than the corresponding LPSs andwere indeed as potent as LPS from E. coli. For ex-ample, SAM from A. actinomycetemcomitans was ableto stimulate IL-6 production by human PBMCs at a concentration of 10 pg/ml.76,135 We have isolated a 2 kDapeptide from this bacterium which can, in contrast toLPS, directly induce the synthesis of IL-6 in an IL-1 andTNF-independent manner (unpublished data). Otherworkers have also shown that soluble surface-associ-ated proteins have the capacity to activate myelomono-cytic cells and to induce cytokine synthesis. Forexample, Mai et al.136 reported that H. pylori, an organ-ism associated with gastric lesions, could stimulatehuman monocytes to express HLA-DR, IL-2 receptorand synthesize both IL-1 and TNF. As few as 1000 bac-teria were able to induce significant cytokine synthesis.Extraction of LPS-free surface-associated proteins andLPS revealed that the former fraction was both signif-icantly more potent and efficacious in stimulatingcytokine synthesis. There has been increasing interestin this organism in recent years and a number of recentreports have suggested that H. pylori releases factorsthat: (1) can stimulate epithelial cells to produceleukotrienes and IL-8; (2) are directly chemotactic forleukocytes.137 The surface-associated protein fractioncontains a number of distinct proteins and thus thepotency of the active molecule is probably high. A
55 kDa cell surface protein from Prevotella intermediainduced the dose-dependent formation of IL-1α and β,IL-6, IL-8 and TNF-α by human PBMCs, and IL-1 andIL-6 production by human vascular endothelial cells andgingival fibroblasts. The potency of this protein wasequivalent to, or greater than, the LPS purified from thisbacterium and significant release of cytokines was foundat a concentration of l 00 ng/ml. In the case of IL-8 pro-duction, 100 ng/ml (2 nM) gave a maximal response. The55 kDa protein also stimulated macrophages fromC3H/HeJ mice showing that it was acting by a distinctmechanism from LPS.138 The SR protein of Strep. mutansalso stimulates human monocytes to release IL-1, IL-6and TNF even in the presence of polymyxin B.139
Extracellular proteins
Bacteria are known to produce a wide range ofextracellular proteins, some of which function as viru-lence factors. Examples include the wide variety of tox-ins that produce the symptoms of diseases such astetanus, cholera, food poisoning etc. In the past fewyears it has been discovered that certain bacterial tox-ins are extremely potent stimulators of cytokine syn-thesis exhibiting activity at picomolar concentrations.
Bacterial toxinsA commonly-encountered bacterial toxin is E. colihaemolysin, an example of a pore-forming toxin. Thisprotein is believed to insert into cell membranes as amonomer to generate a transmembrane lesion of 1–2 nm diameter. Surprisingly, this toxin is not particu-larly effective at causing lysis of erythrocytes. However,it is cytocidal to human monocytes. A key finding is thatsub-cytocidal concentrations (peak concentration 10 ng/ml) of this toxin caused the rapid release of largeamounts of IL-1β from cultured monocytes but notfrom freshly isolated cells. This appears to be due to thestimulation of the processing and export of pre-formedcytokine.140 Monocytes did not release TNF-α in re-sponse to this toxin. The ability of very low amounts ofthis haemolysin to rapidly stimulate IL-1 release couldcontribute to the pathological effects of this toxin. Asimilar finding was reported with the pore-formingstaphylococcal α-toxin.141 A recent report has high-lighted the synergism that exists between LPS and E. coli haemolysin.142 Newly discovered toxins such asthe mitogenic factor (MF) from streptococci also havethe capacity to stimulate cytokine synthesis.143
Listeriolysin O, a 58 kDa thiol-activated haemolysin,is the best characterized virulence factor of Listeriamonocytogenes and is necessary for the intracellular survival of this organism. This Gram-positive bacteriumcan cause severe infections in newborns and the immuno-compromised. Two reports have established that thistoxin can potently induce murine macrophages to syn-thesize both IL-1 mRNA transcripts and protein.144,145
276 / Henderson and Wilson CYTOKINE, Vol. 8, No. 4 (April 1996: 269–282)
A toxin from Bord. pertussis termed pertussis toxin,which is a protein, has also been shown to enhance IL-4 production which may explain the ability of thistoxin to upregulate IgE responses.146
Two of the most potent stimulators of pro-inflam-matory cytokine synthesis are pneumolysin from Strep. pneumoniae and toxin B from Clostridium difficile an organism causing pseudomembranous coli-tis. Three pg/ml of pneumolysin was sufficient to inducethe synthesis of both IL-1 and TNF by human mono-cytes and 10 pg/ml produced .50% of the maximalresponse. With a molecular weight of 56 kDa this isequivalent to a molar concentration of approximately200 femtomolar (200 3 10215 M). Pneumolysin was alsoextremely efficacious and, at a concentration of 1 ng/ml,induced cells to release 3–4 times the amount ofcytokine that a dose of 50 ng/ml LPS was capable ofinducing.147 Purified toxin B stimulated human mono-cytes to produce IL-1, TNF and IL-6, the maximal stim-ulation of TNF and IL-6 synthesis was seen with 300 pg/ml. With a molecular weight of 269 kDa this isequivalent to a molar concentration of 10212 M. ToxinB and LPS showed synergistic interactions in the induc-tion of the synthesis of IL-α, IL-6 and TNF. A secondtoxin from this organism, toxin A, also induced cytokinerelease but was approximately 1000 times less potentthan toxin B.148 The receptor for toxin A has been iden-tified as a galactose- and N-acetylglucosamine-con-taining glycoprotein coupled to a G-protein.149
ProteasesBacterial and viral proteases have recently been foundto interact with the cytokine network in both positiveand negative ways. Thus exotoxin B, a conserved extra-cellular cysteine protease of Strep. pyogenes, cleavesinactive IL-1β precursor between His-115 and Asp-116to produce biologically active IL-1.150 The naturalmonocyte protease IL-1β converting enzyme (ICE)cleaves pro-IL-1β between Asp-116 and Ala-117.151
Exotoxin B from Strep. pyogenes cleaves the inactiveIL-1β precursor between His-115 and Asp-116 andtherefore could stimulate the production of biologi-cally-active IL-1β locally. In contrast, vaccinia virus con-tains a gene (CRM A) for a 38 kDa serpin which inhibitsthe activity of ICE thus inhibiting IL-1β production.152
Ps. aeruginosa produces an alkaline protease and elas-tase both of which can inactivate human IFN-γ andTNF-α153 and a protease from Legionella pneumophiliahas been reported to inactivate IL-2.154
BACTERIAL INVASION AND CYTOKINEPRODUCTION
A number of bacteria enter and survive withinmammalian cells and there is increasing evidence that
cytokines and/or their receptors are involved in theprocess of cell invasion. For example, the invasion ofintestinal epithelial cells by Sal. typhimurium requiresthe phosphorylation of the epidermal growth factorreceptor.155 The invasion of fibroblasts by Sh. flexneriinduced the production of IFN-β. In contrast, isogenicvariants which were not invasive failed to stimulatecytokine synthesis. This suggests that interferon pro-duction may be required for bacterial invasion.156
Similarly Eckmann et al.157 concluded, from their studies of the invasion of epithelial cells by Salmonellaspp and Listeria monocytogenes, that the process of cellentry, and not simply the presence of the bacteria, wasthe signal inducing invaded cells to synthesize andsecrete IL-8. As epithelial cells are the first site of entryof bacteria, the activation of IL-8 synthesis would actas an early warning system at a time when bacterialproducts are unavailable to stimulate circulating leuko-cytes.
CONCLUSIONS
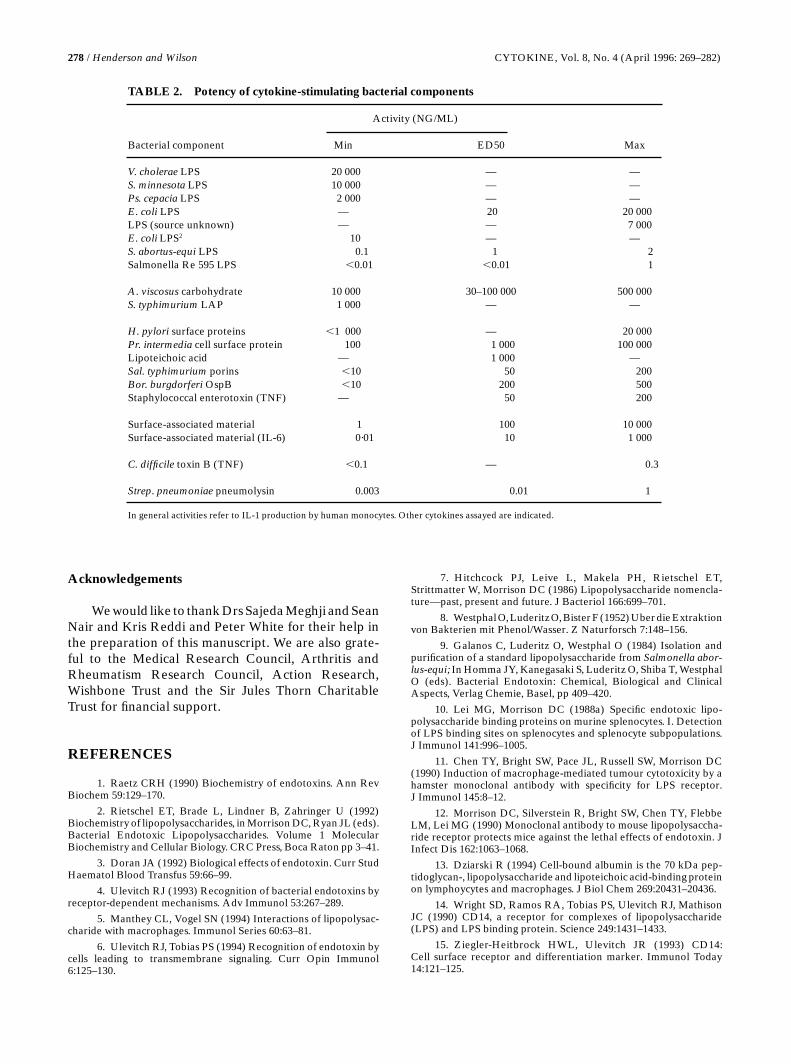
It is now clear that, in addition to LPS, bacteria con-tain, or produce, a wide range of cytokine-inducing pro-teins, carbohydrates and lipids, and most of thesemolecules have only been discovered during the pastdecade. Many of these molecules are very potentcytokine inducers (Table 2). The protein, pneumolysinis active at femtomolar concentrations and a recentlydescribed amphiphilic lipid-like compound fromMycoplasma fermentans stimulates macrophage IL-6synthesis with an ED50 of 50 pM.158 It is not clear howmany more cytokine-inducing molecules bacteria pos-sess, nor is it clear what their functions are. It seemsunlikely that all these molecules evolved simply to stim-ulate eukaryotic cytokine synthesis and perhaps theyform part of a network of interactions between theprokaryotic and eukaryotic worlds. Recent transgenicdeletion mutants (knockouts) have suggested that thereis a complex interplay between commensal bacteria and the cytokine network. Thus IL-2159 and IL-10160
homozygous knockouts develop colitis and enterocoli-tis respectively and, in the case of the former, if animalsare maintained under gnotobiotic conditions then thecolitis does not develop. Thus it is possible that the fail-ure to develop inflammation in response to commensalflora (which contain LPS and many other proinflam-matory molecules) is an active cytokine-dependentprocess involving the synthesis of immunomodulatorycytokines such as IL-10, TGF-β1 etc which have thecapacity to downregulate inflammation. Whatever theirfunction, the discovery that bacteria contain such a widerange of molecules able to induce cytokine synthesismust change our view of the nature of the interactionsbetween bacteria and host cells.
Bacterial cytokine-inducing factors / 277
Acknowledgements
We would like to thank Drs Sajeda Meghji and SeanNair and Kris Reddi and Peter White for their help inthe preparation of this manuscript. We are also grate-ful to the Medical Research Council, Arthritis andRheumatism Research Council, Action Research,Wishbone Trust and the Sir Jules Thorn CharitableTrust for financial support.
REFERENCES
1. Raetz CRH (1990) Biochemistry of endotoxins. Ann RevBiochem 59:129–170.
2. Rietschel ET, Brade L, Lindner B, Zahringer U (1992)Biochemistry of lipopolysaccharides, in Morrison DC, Ryan JL (eds).Bacterial Endotoxic Lipopolysaccharides. Volume 1 MolecularBiochemistry and Cellular Biology. CRC Press, Boca Raton pp 3–41.
3. Doran JA (1992) Biological effects of endotoxin. Curr StudHaematol Blood Transfus 59:66–99.
8. Westphal O, Luderitz O, Bister F (1952) Uber die Extraktionvon Bakterien mit Phenol/Wasser. Z Naturforsch 7:148–156.
9. Galanos C, Luderitz O, Westphal O (1984) Isolation andpurification of a standard lipopolysaccharide from Salmonella abor-lus-equi; In Homma JY, Kanegasaki S, Luderitz O, Shiba T, WestphalO (eds). Bacterial Endotoxin: Chemical, Biological and ClinicalAspects, Verlag Chemie, Basel, pp 409–420.
10. Lei MG, Morrison DC (1988a) Specific endotoxic lipo-polysaccharide binding proteins on murine splenocytes. I. Detectionof LPS binding sites on splenocytes and splenocyte subpopulations.J Immunol 141:996–1005.
11. Chen TY, Bright SW, Pace JL, Russell SW, Morrison DC(1990) Induction of macrophage-mediated tumour cytotoxicity by ahamster monoclonal antibody with specificity for LPS receptor. J Immunol 145:8–12.
12. Morrison DC, Silverstein R, Bright SW, Chen TY, FlebbeLM, Lei MG (1990) Monoclonal antibody to mouse lipopolysaccha-ride receptor protects mice against the lethal effects of endotoxin. JInfect Dis 162:1063–1068.
13. Dziarski R (1994) Cell-bound albumin is the 70 kDa pep-tidoglycan-, lipopolysaccharide and lipoteichoic acid-binding proteinon lymphoycytes and macrophages. J Biol Chem 269:20431–20436.
14. Wright SD, Ramos RA, Tobias PS, Ulevitch RJ, MathisonJC (1990) CD14, a receptor for complexes of lipopolysaccharide(LPS) and LPS binding protein. Science 249:1431–1433.
15. Ziegler-Heitbrock HWL, Ulevitch JR (1993) CD14: Cell surface receptor and differentiation marker. Immunol Today14:121–125.
278 / Henderson and Wilson CYTOKINE, Vol. 8, No. 4 (April 1996: 269–282)
TABLE 2. Potency of cytokine-stimulating bacterial components
18. Lee JD, Kato K, Tobias PS, Kirkland TN, Ulevitch RJ (1992)Transfection of CD 14 into 70Z/3 cells dramatically enhances the sen-sitivity to complexes of lipopolysaccharide (LPS) binding protein. JExp Med 175:1697–1705.
19. Kirkland TN, Finley F, Leturcq D, Moriarty A, Lee J-D,Ulevitch RJ, Tobias PS (1993) Analysis of lipopolysaccharide bind-ing by CD14. J Biol Chem 268:24818–24823.
20. Lee J-D, Kravchenko V, Kirkland TN, Han J, Mackman N,Moriarty A, Leturcq D, Tobias PS, Ulevitch RJ (1993) GPI-anchoredor integral membrane forms of CD14 mediate identical cellularresponses to endotoxin. Proc Natl Acad Sci USA 90:9930–9934.
21. Bazil V, Horejsi V, Baudys M, Kristofova H, Strominger J,Kostra W, Hilgert I (1986) Biochemical characterization of a solubleform of the 53kDa monocyte surface antigen. Eur J Immunol16:1583–1589.
22. Haziot A, Rong G-W, Bazil V, Silver J, Goyert SM (1994)Recombinant soluble CD14 inhibits LPS-induced tumor necrosis fac-tor-a production by cells in whole blood. J Immunol 152:5868–5876.
23. Wurfel MM, Hailman E, Wright SD (1995) Soluble CD14acts as a shuttle in the neutralization of LPS by LPS-binding proteinand reconstituted high density lipoprotein. J Exp Med 181:1743–1754.
24. Raetz CRH (1993) Bacterial endotoxins: extraordinarylipids that activate eukaryotic signal transduction. J Bacteriol175:5745–5753.
25. Shapira L, Takabashi S, Champagne C, Amar S, Van DykeTE (1994) Involvement of protein kinase C and protein tyrosinekinase in lipopolysaccharide-induced TNF-α and IL-1β productionby human monocytes. J Immunol 153:1818–1824.
26. Weinstein SL, Sanghera JS, Lemke K, DeFranco AL, PelechSL (1992) Bacterial lipopolysaccharide induces tyrosine phosphoryl-ation and activation of mitogen-activated protein kinases inmacrophages. J Biol Chem 267:14955–14961.
27. Reimann T, Buscher D, Hipskind RA, Krautwald S,Lohmann-Matthes M-L, Baccarini M (1994) Lipopolysaccharideinduces activation of the Raf-1/MAP kinase pathway. J Immunol 153:5740–5749.
28. Novogrodsky A, Vanichkin A, Patya M, Gazit A, OsherovN, Levitzki A (1994) Prevention of lipopolysaccharide-induced lethaltoxicity by tyrosine kinase inhibitors. Science 264:1319–1322.
29. Laude-Sharp M, Haeffner-Cavaillon N, Caroff M,Lantreibecq F, Pusineri C, Kazatchkine MD (1990) Dissociationbetween the interleukin-1-inducing capacity and Limulus reactivity oflipopolysaccharides from Gram-negative bacteria. Cytokine 2:253–258.
30. Moreno C, Mehlert A, Lamb J (1988) The inhibitory effectsof mycobacterial lipoarabinomannan and polysaccharides upon poly-clonal and monoclonal human T cell proliferation. Clin Exp Immunol74:206–210.
32. Sibley LD, Adams LB, Krahenbuhl JL (1990) Inhibition ofinterferon-gamma-mediated activation in mouse macrophagestreated with lipoarabinomannan. Clin Exp Immunol 80:141–148.
33. Chan J, Fan X, Hunter SW, Brennan PJ, Bloom BR (1991)Lipoarabinomannan, a possible virulence factor involved in persis-tence of Mycobacterium tuberculosis within macrophages. InfectImmun 59:1755–1761.
34. Moreno C, Taverne J, Mehlert A, Bate CAW, Brealey RJ,Meager A, Rook, GAW, Playfair JHL (1988) Lipoarabinomannanfrom Mycobacterium tuberculosis induces the production of tumournecrosis factor from human and murine macrophages. Clin ExpImmunol 76:240–245.
35. Barnes PF, Chatterjee D, Abrams JS, Lu S, Wang E,Yamamura M, Brennan PJ, Modlin RL (1992) Cytokine productioninduced by Mycobacterium tuberculosis lipoarabinomannan:Relationship to chemical structure. J Immunol 149:541–547.
36. Schlesinger LS, Hull SR, Kaufman TM (1994) Binding ofthe terminal mannosyl units of lipoarabinomannan from a virulentstrain of Mycobacterium tuberculosis to human macrophages. J Immunol 152:4070–4079.
37. Zhang Y, Broser M, Rom WN (1994) Activation of the inter-leukin-6 gene by Mycobacterium tuberculosis and lipopolysaccharideis mediated by nuclear factors NF-IL-6 and NF-κB. Proc Natl AcadSci USA 91:2225–2229.
38. Adams LB, Fukutomi Y, Krahenbuhl JL (1993) Regulationof murine macrophage effector function by lipoarabinomannan frommycobacterial strains with different degrees of virulence. InfectImmun 61:4173–4181.
39. Roach TIA, Barton CH, Chaterjee D, Blackwell JM (1993)Macrophage activation: Lipoarabinomannan from avirulent and vir-ulent strains of Mycobacterium tuberculosis differentially induces theearly genes c-fos, KC, JE, and tumour necrosis factor-α. J Immunol150:1886–1896.
40. Roach TIA, Chaterjee D, Blackwell JM (1994) Induction ofearly response genes KC and JE by mycobacterial lipoarabinoman-nans: Regulation of KC expression in murine macrophages byLsh/Ity/Bcg (Candidate Nramp). Infect Immun 62:1176–1184.
41. Wallis RS, Paranjape R, Phillips M (1993) Identification bytwo-dimensional gel electrophoresis of a 58 kilodalton tumor necro-sis factor-inducing protein of Mycobacterium tuberculosis. InfectImmun 61:627–632.
42. Ausiello CM, Urbani F, Gessani S, Spagnoli GC, Gomez MJ,Cassone A (1993) Cytokine gene expression in human peripheralblood mononuclear cells stimulated by mannoprotein constituentsfrom Candida albicans. Infect Immun 61:4105–4111.
43. Friedland JS, Shattock R, Remick DG, Griffin GE (1993)Mycobacterial 65-kD heat shock protein induces release of pro-inflammatory cytokines from human monocytic cells. Clin ExpImmunol 91:58–62.
44. Del Prete GF, De Carli M, Mastromauro C, Biagiotti R,Macchia D, Falagiani P, Rici M, Romagnani S (1991) Purified proteinderivative of Mycobacterium tuberculosis and excretory-secretoryantigen(s) of Toxocara canis expand in vitro human T cells with stable and opposite (type I T helper or type 2 T helper) profile ofcytokine production. J Clin Invest 88:346–350.
45. Tufano MA, Cipollaro de L’Ero G, Ianniello R, GaldieroM, Galdiero F (1991) Protein A and other surface components ofStaphylococcus aureus stimulate production of IL-1α, IL-4, IL-6, TNFand IFN-γ. Eur Cytokine Netw 2:361–366.
46. Riesenfeld-Orn I, Wolpe S, Garcia-Bustos JF, HoffmannMK, Tuomanen E (1989) Production of interleukin-l but not tumornecrosis factor by human monocytes stimulated with pneumococcalcell surface components. Infect Immun 57:1890–1893.
47. Tsutsui O, Kikeguchi S, Matsumara, Kato K (1991)Relationship of the chemical structure and immunobiological activ-ities of lipoteichoic acid from Streptococcus faecalis (Enterococcushirae) ATCC 9790. FEMS Microbiol Immunol 76:211–218.
48. Bhakdi S, Klonisch T, Nuber P, Fischer W (1991) Stimulationof monokine production by lipoteichoic acids. Infect Immun59:4614–4620.
49. Heumann D, Barras C, Severin A, Glauser MF, Tomasz A(1994) Gram-positive cell walls stimulate synthesis of tumor necro-sis factor alpha and interleukin-6 by human monocytes. Infect Immun62:2715–2721.
50. Keller R, Fischer W, Keist R, Bassetti S (1992) Macrophageresponse to bacteria: Induction of marked secretory and cellularactivities by lipoteichoic acids. Infect Immun 60:3664–3672.
51. Standiford TJ, Arenberg DA, Danforth JM, Kunkel SL,VanOtteren GM, Strieter RM (1994) Lipoteichoic acid induces secre-tion of interleukin-8 from human blood monocytes: A cellular andmolecular analysis. Infect Immun 62:119–125.
Bacterial cytokine-inducing factors / 279
52. White J, Herman A, Pullen AM, Kubo R, Kappler JW,Marrack P (1989) The Vβ-specific superantigen staphylococcalenterotoxin B: Stimulation of mature T cells and clonal deletion inneonatal mice. Cell 56:27–35.
53. Marrack P, Kappler J (1990) The staphylococcal entero-toxins and their relatives. Science 248:705–711.
54. Betley MJ, Borst DW, Regassa LB (1992) Staphylococcalenterotoxins, toxic shock syndrome toxin and streptoccal pyrogenicexotoxins: A comparative study of their molecular biology. ChemImmunol 55:1–35.
55. Miethke T, Wahl C, Echtenacher B, Krammer P, Heeg K,Wagner H (1992) T cell-mediated lethal shock triggered in mice bythe superantigen staphylococcal enterotoxin B: Critical role of tumornecrosis factor. J Exp Med 175:91–98.
57. Fuleihan R, Mourad W, Geha RS, Chatila T (1991)Engagement of MCH-class II molecules by staphylococcal exotoxinsdelivers a comitogenic signal to human B cells. J Immunol146:1661–1666
58. Scholl PR, Trede N, Chatila TA, Geha RS (1992) Role ofprotein tyrosine phosphorylation in monokine induction by thestaphylococcal superantigen toxic shock syndrome toxin-1. J Immunol 148:2237–2241.
59. Stiles BG, Bavari S, Krakauer T, Ulrich RG (1993) Toxicityof staphylococcal enterotoxins potentiated by lipopolysaccharide:Major histocompatability complex class II molecule dependency andcytokine release. Infect Immun 61:5333–5338.
60. Grossman D, Lamphear JG, Mollick JA, Betley MJ, RichRR (1992) Dual roles for class II major histocompatibility complexmolecules in staphylococcal enterotoxin-induced cytokine produc-tion and in vivo toxicity. Infect Immun 60:5190–5196.
61. Beezhold DH, Best GK, Bonventre PF, Thompson M (1987)Synergistic induction of IL-1 by endotoxin and toxic shock syndrometoxin-1 using rat macrophages. Infect Immun 55:2865–2869.
62. Henne E, Campbell WH, Carlson E (1991) Toxic shock syn-drome toxin 1 enhances synthesis of endotoxin-induced tumor necro-sis factor in mice. Infect Immun 59:2929–2933.
63. Sultzer BM (1968) Genetic control of leucocyte responsesto endotoxin. Nature 219:1253–1255.
64. Skidmore BJ, Morrison DC, Chiller JM, Weigle WO (1975)Immunologic properties of bacterial LPS. II. The unresponsivenessof C3H/HeJ spleen cells to LPS-induced mitogenesis is dependent onthe methods used to extract LPS. J Exp Med 142:1488–1508.
65. Sultzer BM, Goodman GW (1976) Endotoxin protein: A Bcell mitogen and polyclonal activator of C3H/HeJ lymphocytes. J ExpMed 144:821–827.
66. Goodman GW, Sultzer BM (1979) Endotoxin protein is amitogen and polyclonal activator of human B lymphocytes. J ExpMed 149:713–723.
67. Killion JW, Morrison DC (1986) Protection of C3H/HeJmice from lethal Salmonella typhimurium LT2 infection by immu-nization with lipopolysaccharide-lipid-A-associated protein com-plexes. Infect Immun 54:1–8.
68. Sultzer BM, Craig JP, Castagna R (1985) The adjuvant effectof pertussis endotoxin protein in modulating the immune responseto cholera toxoid in mice. Dev Biol Stand 61:225–232.
69. Sultzer BM, Craig JP, Castagna R (1986) Endotoxin-associated proteins and their polyclonal and adjuvant activities. In Szentivanyi A, Friedman H (eds). Immunobiology and Immuno-pharmacology of Bacterial Endotoxin; Plenum; New York,pp 435–447.
70. Sultzer BM, Craig JP, Castagna R (1987) Immuno-modulation by outer membrane proteins associated with the endo-toxin of Gram-negative bacteria. Prog Leukocyte Biol 6:113–123.
71. Hogan MH, Vogel SN (1987) Lipid A-associated proteinsprovide an alternative ‘second signal’ in the activation of recombi-
nant interferon-γ-primed, C3H/HeJ macrophages to a fully tumori-cidal state. J Immunol 139:3697–3702.
73. Bjornson BH, Agura E, Harvey JH, Johns M, Andrews RG,McCabe WR (1988) Endotoxin-associated protein: a potent stimu-lus for human granulocytopoietic activity which may be accessory cellindependent. Infect Immun 56:1602–1607.
74. Porat R, Yanoov M, Johns MA, Shibolet S, Michalevicz R(1992) Effects of endotoxin-associated protein on hematopoiesis.Infect Immun 60:1756–1760.
75. Mangan DF, Wahl SM. Sultzer BM, Mergenhagen SE(1992) Stimulation of human monocytes by endotoxin-associatedprotein: Inhibition of programmed cell death (apotosis) and poten-tial significance in adjuvanticity. Infect Immun 60:1684–1686.
76. Reddi K, Wilson M, Poole S, Meghji S, Henderson B (1994)Relative cytokine-stimulating activities of surface components of theoral periodontopathic bacterium Actinobacillus actinomycetem-comitans. Cytokine 7:534–541.
77. Bandekar JR, Castagna R, Sultzer BM (1992) Roles of pro-tein kinase C and G proteins in activation of murine resting B lym-phocytes by endotoxin-associated protein. Infect Immun 60:231–236.
78. Bessler WG, Ottenbreit BP (1977) Studies on the mitogenicprinciple of the lipoprotein from the outer membrane of Escherichiacoli. Biochem Biophys Res Commun 76:239–246.
79. Goldman RC, White D, Leive L (1981) Identification ofouter membrane proteins, including known lymphocyte mitogens, asthe endotoxin protein of Escherichia coli 0111. J Immunol127:1290–1294.
80. Mohri S, Watanabe T, Nariuchi H (1982) Studies of theimmunological activities of the outer membrane protein fromEscherichia coli. Immunology 46:271–280.
81. Vodermeier M, Stab K, Bessler WG (1986) A defined frag-ment of bacterial protein (OmpF) is a polyclonal B cell activator.Infect Immun 51:233–239.
83. Takada H, Ogawa T, Yoshimura F, Otsuka K, Kokeguchi S,Kato K, Ememoto T, Kotani S. (1988) Immunobiological activities ofa porin fraction isolated from Fusobacterium nucleatum ATCC 10953.Infect Immun 56:855–863.
84. Tufano M, Ianniello R, Galdiero M, De Martino L, Galdiero F(1989) Effect of Salmonella typhimurium porins on biological activitiesof human polymorphonuclear leukocytes. Microb Pathog 7:337–346.
85. Tufano MA, Tetta C, Biancone L, Iorio EL, Baroni A,Giovane A, Camussi G (1992) Salmonella typhimurium porins stim-ulate platelet-activating factor synthesis by human polymorpho-nuclear neutrophils. J Immunol 149:1023–1030.
90. Galdiero F, Cipollaro de L’Ero G, Bendetto N, Galdiero M,Tufano MA (1993) Release of cytokines induced by Salmonellatyphimurium porins. Infect Immun 61:155–161.
280 / Henderson and Wilson CYTOKINE, Vol. 8, No. 4 (April 1996: 269–282)
91. Tufano MA, Rossano F, Catalanotti P, Liguori G, MarinelliA, Baroni A, Marinelli P (1994) Properties of Yersinia enterocoliticaporins: Interference with biological functions of phagocytes, nitricoxide production and selective cytokine release. Institut Pasteur ResMicrobiol 145:297–307.
92. Czarny A, Witkowska D, Mulczyk M (1993) Induction oftumor necrosis factor and interleukin-6 by outer membrane proteinsof Shigella in spleen cells and macrophages of mice. Achiv ImmunolTher Exp 41:153–157.
93. Ma Y, Weis JJ (1993) Borrelia burgdorferi outer surfacelipoproteins OspA and OspB possess B-cell mitogenic and cytokine-stimulatory properties. Infect Immun 61:3843–3853.
94. Tai K-F, Ma T, Weis JJ (1994) Normal human B lympho-cytes and mononuclear cells respond to the mitogenic and cytokine-stimulatory activities of Borrelia burgdorferi and its lipoproteinOspA. Infect Immun 62:520–528.
95. Korn A, Kroll H-P, Berger H-P, Kahler A, Hebler R,Brauburger J, Muller K-P, Nixdorf K (1993) The 39-kilodalton outermembrane protein of Proteus mirabilis is an OmpA protein and mito-gen for murine B lymphocytes. Infect Immun 61:4915–4918.
96. Weber G, Link F, Ferber E, Munder PG, Zeitter D, BartlettRR, Nixdorff K (1993) Differential modulation of the effects oflipopolysaccharide on macrophages by a major outer membrane pro-tein of Proteus mirabilis. J Immunol 151:415–424.
97. Melchers F, Braun V, Galanos C (1975) The lipoprotein ofthe outer membrane of Escherichia coli: A B lymphocyte mitogen. JExp Med 142:473-482.
98. Bessler WG, Cox M, Lex A, Suhr B, Weismuller K-H, JungG (1985) Synthetic lipoprotein analogues of bacterial lipoprotein arepotent polyclonal activators for murine B lymphocytes. J Immunol135:1900–1908.
99. Lex A, Wiesmuller K-H, Jung G, Bessler WG (1986) A synthetic analogue of Escherichia coli lipoprotein; tripalmitoyl pentapeptide constitutes a potent immune adjuvant. J Immunol 137:2676–2681.
100. Deres K, Schild H, Wiesmuller K-H, Jung G, RammenseeH-G (1989) In vivo priming of virus-specific cytotoxic T lymphocyteswith synthetic lipopeptide vaccine. Nature 342:561–564.
101. Radolf JD, Norgard MV, Brandt ME, Isaacs RD, ThompsonPA, Beutler B (1991) Lipoproteins of Borrelia burgdorferi andTreponema pallidum activate cachectin/tumor necrosis factor synthesis:Analysis using a CAT reporter construct. J Immunol 147:1968–1974.
102. Hauschildt S, Hoffmann P, Beuscher H-U, Dufhues G,Heinrich P, Wiesmuller K-H, Jung G, Bessler WG (1990) Activationof bone marrow-derived mouse macrophages by bacterial lipopep-tide: cytokine production, phagocytosis and Ia expression. Eur JImmunol 20:63–68.
103. Akins DR, Purcell BK, Mitra MM, Norgard MV, RadolfJD (1993) Lipid modification of the 17-kilodalton membraneimmunogen of Treponema pallidum determines macrophage activa-tion as well as amphiphilicity. Infect Immun 61:1202–1210.
104. Dong Z, Qi X, Fidler IJ (1993) Tyrosine phosphorylationof mitogen-activated protein kinases is necessary for activation ofmurine macrophages by natural and synthetic bacterial products. JExp Med 177:1071–1077.
105. Schwab J (1993) Phlogistic properties of peptidoglycan-polysaccharide polymers from cell walls of pathogenic and normalflora bacteria which colonize humans. Infect Immun 61:4535–4539.
106. Schwab J (1995) Bacterial cell-wall-induced arthritis:Models of chronic recurrent polyarthritis and reactivation of mono-articular arthritis. In Henderson B, Edwards JCW, Pettipher ER (eds). Models and Mechanisms in Rheumatoid Arthritis. AcademicPress) London, pp 439–454.
107. Cavaillon JM, Haeffner-Cavaillon N (1990) Signalsinvolved in interleukin-1 synthesis and release by lipopolysaccharide-stimulated monocytes/macrophages. Cytokine 2:313–329.
108. Chedid L (1983) Muramyl peptides as possible endoge-nous immunopharmacological mediators. Microbiol Immunol27:723–732.
109. O’Reilly T, Zac O (1992) Enhancement of the effective-ness of antimicrobial therapy by muramyl peptide immunomodula-tors. Clin Infect Dis 14:1100–1109.
110. Keller R, Gustafson JE, Keist R (1992) The macrophageresponse to bacteria. Modulation of macrophage functional activityby peptidoglycan from Moraxella (Branhamella) catarrhalis. Clin ExpImmunol 89:384–389.
111. Timeerman CP, Matsson E, Martinez-Martinez L, DeGraf L,Van Strijp JAG, Verbrugh HA, Verhoef J, Fleer A (1993) Induction ofrelease of tumor necrosis factor from human monocytes by staphylococciand staphylococcal peptidoglycans. Infect Immun 61:4167–4172.
112. Onta T, Sashida M, Fujii N, Sugawara S, Rikiishi H,Kumagai K (1993) Induction of acute arthritis in mice by peptido-glycan derived from gram-positive bacteria and its possible role incytokine production. Microbiol Immunol 37:573–582.
113. Dokter WHA, Dijkstras A, Koopmans SB, Stulp BK, KeckW, Halie MR, Vellenga E (1994a) G(Anh)MTetra, a natural bacter-ial cell wall breakdown product, induces interleukin-1β and inter-leukin-6 expression in human monocytes. J Biol Chem 269:4201–4206.
115. Dinarello CA, Krueger JM (1986) Induction of inter-leukin-1 by synthetic and naturally occurring muramyl peptides. FedProc 45:2545–2548.
116. Daley L, Pier GB, Liporace JD, Eardley DD (1985)Polyclonal B cell stimulation and interleukin-1 induction by themucoid exopolysaccharide of Pseudomonas aeruginosa associatedwith cystic fibrosis. J Immunol 134:3089–3093.
117. Takahashi T, Nishihara T, Ishihara Y, Amano K, ShibuyuN, Moro I, Koga T (1991) Murine macrophage interleukin- 1 releaseby capsularlike serotype-specific polysaccharide antigens ofActinobacillus actinomycetemcomitans. Infect Immun 59:18–23.
118. Benabdelmoumene S, Dumont S, Petit C, Poindron P, Wachsman D, Klein JP (1991) Activation of human monocytes byStreptococcus mutans serotype F polysaccharide: Immunoglobulin GFc receptor expression and tumor necrosis factor and interleukin-1pro-duction. Infect Immun 59:3261–3266.
119. Takada H, Kimura S, Hamada S (1993) Induction of inflam-matory cytokines by a solube moiety prepared from an enzyme lysateof Actinomyces viscosus cell walls. J Med Microbiol 38:395–400.
120. Otterlei M, Sundan A, Skjak-Braek G, Ryan L, SmidsrodO, Espevik T (1993) Similar mechanisms of action of defined poly-saccharides and lipopolysaccharides: characterization of binding andtumor necrosis factor alpha induction. Infect Immun 61:1917–1935.
121. Mancuso G, Tomasello F, von Hunolstein C, Orefici G, TetiG (1994) Induction of tumor necrosis factor alpha by the group- andtype-specific polysaccharides from type III group B streptococci.Infect Immun 62:2748–2753.
122. Hultgren SJ, Abraham S, Caparon M, Falk P, St Geme IIIJW, Normark S (1993) Pilus and nonpilus bacterial adhesins:Assembly and function in cell recognition. Cell 73:887–901.
123. Gbarah A, Gahmberg CG, Ofek I, Jacobi U, Sharon N(1991) Identification of the leukocyte adhesion molecules CD11 andCD18 as receptors for type I-fimbriated (mannose-specific)Escherichia coli. Infect Immun 59:4524–4530.
124. Linder H, Engberg I, Hoschutsky H, Mattsby-Baltzer I,Svanborg C (1991) Adhesion dependent activation of mucosal inter-leukin-6 production. Infect Immun 59:4357–4362.
125. Hedges S, Svensson M, Svanborg C (1992) Interleukin-6response of epithelial cell lines to bacterial stimulation in vitro. InfectImmun 60:1295–1301.
126. Kreft B, Bohnet S, Carstensen O, Hacker J, Marre R (1993)Differential expression of interleukin-6, intracellular adhesion mol-ecule I, and major histocompatibility complex class II molecules inrenal carcinoma cells stimulated with S. finbriae of uropathogenicEscherichia coli. Infect Immun 61:3060–3063.
128. Hanazawa S, Murakami Y, Hirose K, Amano S, OhmoriY, Higuchi H, Kitano S (1991) Bacteroides (porphyromonas) gingi-valis fimbriae activate mouse peritoneal macrophages and inducegene expression and production of interleukin-1. Infect Immun59:1972–1977.
129. Hanazawa S, Murakami Y, Takeshita A, Kitami H, OhtaK, Amano S, Kitano S (1992) Porphyromonas gingivalis fimbriaeinduce expression of the neutrophil chemotactic factor KC gene ofmouse peritoneal macrophages: Role of protein kinase C. InfectImmun 60:1544–1549.
130. McCormick BA, Colgan SP, Delp-Archer C, Miller SI,Madara JL (1993) Salmonella typhimurium attachment to humanintestinal epithelial monolayers: transcellular signalling to sub-epithelial neutrophils. J Cell Biol 123:895–907.
131. Brett SJ, Mazurov AV, Charles IG, Tite JP (1993) Theinvasin protein of Yersinia spp provides co-stimulatory activity tohuman T cells through interaction with β1 integrins. Eur J Immunol23:1608–1614.
132. Wilson M, Kamin S, Harvey W (1985) Bone resorbingactivity of purified capsular material from Actinobacillus actino-mycetemcomitans. J Periodont Res 20:484–491.
133. Wilson M, Meghji S, Barber P. Henderson B (1993)Biological activities of surface-associated material from Porphy-romonas gingivalis. FEMS Immunol Med Microbiol 6:147–155.
134. Meghji S, Wilson M, Barber P, Henderson B (1994) Boneresorbing activity of surface-associated material from Actinobacillusactinomycetemcomitans and Eikenella corrodens. J Med Microbiol41:197–203.
135. Reddi K, Wilson M, Poole S, Henderson B (1994)Comparison of the pro-inflammatory-cytokine-stimulating activity ofthe surface-associated material of periodontopathic bacteria. JPeriodont Res (in press).
136. Mai UEH, Perez-Perez GI, Wahl LM, Wahl SM, BlaserMJ, Smith PD (1991) Soluble surface proteins from Helicobacterpylori activate monocytes/macrophages by lipopolysaccharide-inde-pendent mechanism. J Clin Invest 87:894–900.
137. Lee A, Fox J, Hazell S (1993) Pathogenicity of Helicobacterpylori: a perspective. Infect Immun 61:1601–1610.
138. Matsushita K, Nagaoka S, Arakaki R, Kawabata Y, Iki K,Kawagoe M, Takada H (1994) Immunobiological activities of a 55 kilodalton cell surface protein of Prevotella intermedia ATCC25611. Infect Immun 62:2459–2469.
139. Soell M, Holveck F, Scholler M, Wachsman D, Klein J-P(1994) Binding of Streptococcus mutans SR protein to human mono-cytes: Production of tumor necrosis factor, interleukin-1 and inter-leukin-6. Infect Immun 62:1805–1812.
140. Bhakdi S, Muhly M, Korom S, Schmidt G (1990) Effectsof Escherichia coli hemolysin on human monocytes: Cytocidal actionand stimulation of interleukin-1 release. J Clin Invest 85:1746–1753.
141. Bhakdi S, Muhly M, Korom S, Hugo F (1989) Release ofinterleukin-1β associated with potent cytocidal action of staphylo-coccal alpha-toxin on human monocytes. Infect Immun 57:3512–3519.
142. Walmrath D, Ghofrani HA, Rosseau S, Schutte H, CramerA, Kadus W, Grimminger F, Bhakdi S, Seeger W (1994) Endotoxin“priming” potentiates lung vascular abnormalities in response toEscherichia coli hemolysins: An example of synergism between endo-and exotoxin. J Exp Med 180:1437–1443.
143. Norrby-Teglund A, Norgren M, Holm SE, Andersson U,Anderrson J (1994) Similar cytokine induction profiles of a novel
streptococcal exotoxin, MF, and pyrogenic exotoxins A and B. InfectImmun 62:3731–3738.
144. Tsukada H, Kawamura I, Fujimura T, Igarashi K-I,Arakawa M, Mitsuyama M (1992) Induction of macrophage inter-leukin-1 production by Listeria monocytogenes hemolysin. CellImmunol 140:21–30.
145. Yoshikawa H, Kawamura I, Fujita M, Tsukada H,Arakawa M, Mitsuyama M (1993) Membrane damage and inter-leukin-1 production in murine macrophages exposed to listeriolysinO. Infect Immun 61:1334–1339.
146. Mu H-H, Sewell WA (1993) Enhancement of interleukin-4 production by pertussis toxin. Infect Immun 61:2834–2840.
147. Houldsworth S, Andrew PW, Mitchell TJ (1994)Pneumolysin stimulates production of tumor necrosis factor alphaand interleukin-1βby human mononuclear phagocytes. Infect Immun62:1501–1503.
148. Flegel WA, Muller F, Daubener W, Fischer H-G, HaddingU, Northoff H (1991) Cytokine response by human monocytes toClostridium difficile toxin A and toxin B. Infect Immun 59:3659–3666.
149. Pothoulakis C, LaMont JT, Eglow R, Gao N, Rubins JB,Theoharides TC, Dickey BF (1991) Characterization of rabbit ilealreceptors for Clostridium difficile toxin A: Evidence for a receptor-coupled G protein. J Clin Invest 88:119–125.
150. Kapur V, Majesky MW, Li L-L, Black RA, Musser JM(1993) Cleavage of interleukin-1β(IL-1β) precursor to produce activeIL-1β by a conserved extracellular cysteine protease fromStreptococcus pyogenes. Proc Natl Acad Sci USA 90:7676–7680.
151. Wilson KP, Black JF, Thomson JA, Kim EE, Griffith JP,Navia MA, Murcko MA, Chambers SP, Aldape RA, Raybuck SA,Livingstone DJ (1994) Structure and mechanism of interleukin-1βconverting enzyme. Nature 370:270–275.
152. Ray CA, Black RA, Kronheim SR, Greenstreet TA, SleathPR, Salvesen GS, Pickup DJ (1992) Viral inhibition of inflammation:Cowpox virus encodes an inhibitor of the interleukin-1β convertingenzyme. Cell 69:597–604.
153. Parmely MP, Gale A, Clabaugh M, Horvat R, Zhou WW(1990) Proteolytic inactivation of cytokines by Pseudomonas aerug-inosa. Infect Immun 58:3009–3014.
154. Mintz CS, Miller RD, Gutgsell NS, Malek T (1993)Legionella pneumophilia protease inactivates interleukin-2 andcleaves CD4 on human T cells. Infect lmmun 61:3416–3421.
155. Galan JE, Pace J, Hayman MJ (1992) Involvement of theepidermal growth factor receptor in the invasion of cultured mam-malian cells by Salmonella typhimurium. Nature 357:588–589.
156. Hess CB, Niesel DW, Cho YJ, Klimpel GR (1987) Bacterialinvasion of fibroblasts induces interferon production. J Immunol138:3949–3953.
157. Eckmann L, Kagnoff MF, Fierer J (1993) Epithelial cellssecrete the chemokine interleukin-8 in response to bacterial entry.Infect Immun 61:4569–4574.
158. Muhlradt PF, Frisch M (1994) Purification and partial bio-chemical characterization of a Mycoplasma fermentans-derived sub-stance that activates macrophages to release nitric oxide, tumornecrosis factor, and interleukin-6. Infect Immun 62:3801–3807.
159. Sadlack B, Merz H, Schorle H, Schimpl A, Feller AC,Horak I (1993) Ulcerative colitis in mice with a disrupted interleukin-2 gene. Cell 75:253–261.