Plant Biotechnology Program Science Research Centre Queensland University of Technology Australia and The Regional Germplasm Centre Secretariat of the Pacific Community Suva, Fiji Development and Application of Virus Indexing Protocols for the International Movement of Taro Germplasm QUT Brisbane December 2003
Transcript
Plant Biotechnology Program Science Research Centre
Queensland University of Technology Australia
and
The Regional Germplasm Centre
Secretariat of the Pacific Community Suva, Fiji
Development and Application of Virus Indexing
Protocols for the International Movement of Taro Germplasm
QUT Brisbane December 2003
i
Table of Contents 1. INTRODUCTION 1 2. THE AGREEMENT TO SHARE 2 3. THE VIRUSES AND THEIR DISEASES 3
4. THE METHODS FOR VIRUS DETECTION 11 4.1 The laboratory and its standards 11 4.2 Methods of indexing 11
4.2.1 General 11 4.2.2 The use of controls 12
5. THERAPY AND INDEXING 13 5.1 Eliminating viruses 13
5.1.1 Post-entry quarantine, Australia 13 5.1.2 Testing for viruses, QUT 14
6. INDEXING RESULTS 14 7. THE SURVEY 17
7.1 Results from the survey 18 8. COMMENTS ON THE RESULTS 19
8.1 How important are the viruses found in taro? 19 8.2 Can we be sure of the distribution of the viruses from the survey? 20 8.3 How sensitive are the indexing methods? 21 8.4 Have the methods been peer reviewed? 21 8.5 Were the same plants tested over time and were the results consistent? 21 8.6 Is it possible to audit the system, in case something goes wrong? 21 8.7 Should the taro be grown in PEQ in the importing country? 21 8.8 What is known about the taro varieties? 22 8.9 What to do next? 22
9. BIBLIOGRAPHY 24
ii
Abbreviations AFFA Agriculture, Fisheries and Forestry, Australia ACIAR Australian Centre for International Agricultural Research AQIS Australian Quarantine Inspection Service AusAID Australian Agency for International Development CBDV Colocasia bobone disease virus CIRAD Centre de coopération internationale en recherche agronomique
pour le Développement DNA Deoxyribonucleic acid DsMV Dasheen mosaic virus dsRNA Double-stranded ribonucleic acid EC European Commission ELISA Enzyme-linked immunosorbent assay FAO Food and Agriculture Organization of the United Nations FSM Federated States of Micronesia HRI Horticulture Research International IBPGR International Board of Plant Genetic Resources INCO-DC International Cooperation with Developing Countries IPPC International Plant Protection Convention ISPM International standards for phytosanitary measures LIPI Lembaga Ilmu Pengetahuan Indonesia N/A Not applicable New Cal New Caledonia NGO Non-government organisation N/T Not-tested PCR Polymerase chain reaction PEQ Post-entry quarantine PHALPS Permanent Heads of Agriculture and Livestock Production
Services PNG Papua New Guinea QUT Queensland University of Technology RGC Regional Germplasm Centre SPC Secretariat of the Pacific Community TaBV Taro bacilliform virus TANSAO Taro Network for Southeast Asia and Oceania TaroGen Taro Genetic Resources: Conservation and Utilisation TaRV Taro reovirus TaVCV Taro vein chlorosis virus UNDP United National Development Programme WTO World Trade Organization
iii
Executive Summary This report summarises five years research into the development of indexing methods so that taro, Colocasia esculenta, can be transferred between countries with the minimum of risk. It is the result of collaboration between countries of the Pacific and Asia, the sources of traditional germplasm and breeders’ lines, the Regional Germplasm Centre of the Secretariat of the Pacific Community, where lines are maintained in vitro, and the Plant Biotechnology Program within the Science Research Centre at Queensland University of Technology, which has developed sensitive virus detection methods, and applied them, creating the opportunity for international exchange. The work has been supported under two projects: TaroGen – Taro Genetic Resources and Conservation, and Virus Indexing and DNA Fingerprinting for the International Movement and Conservation of Taro Germplasm, financed by AusAID and ACIAR, respectively. Agreement for the exchange of germplasm between countries of the Pacific was made under a Code of Conduct developed at the TaroGen Planning Workshop, 3-4 September 1998. The members of TaroGen – the Pacific Island countries as well as regional and international organisations and research institutes associated with the project are obligated under the Agreement to freely share their taro genetic resources for research purposes. Sharing of germplasm across international borders requires that the germplasm is free from pathogens, viruses in particular. The five viruses (Dasheen mosaic virus, DsMV; Taro bacilliform virus, TaBV; Taro vein chlorosis virus, TaVCV; Colocasia bobone disease virus, CBDV; and Taro reovirus, TaRV) that infect taro in the Pacific Islands are described in detail in this report. TaRV is newly discovered, and a second TaBV-like sequence has been detected that might be integrated within the genetic material of the taro. The descriptions of the viruses follow the short, concise style of the FAO/IPGRI Technical Guidelines for the Safe Movement of Germplasm in anticipation that they will update an earlier version published in 1999. For each virus, details are provided on its significance, symptoms (in word and illustration), distribution, methods of transmission, and detection. Highly specific DNA sequences (primers) were developed to detect the presence of each virus in cell sap, and where the work has been published in refereed journals (DsMV and TaBV), the sequences are given. Brief notes describe the laboratory at QUT and the procedures for importing and testing (indexing) taro germplasm, with detailed methodology specified in an appendix. Before indexing, plants are grown from shoot-tips in an attempt to eliminate viruses if present. This is done at the RGC. Large shoot-tips are removed from field-grown plants and grown in vitro. Later, when plantlets form, small shoot-tips are removed and cultured. When these have regrown, and suckers have formed, they are sent to QUT for testing. The suckers are retained in the RGC. Each plant is tested twice for viruses according to the following protocol. Tissue cultured plantlets are sent from the RGC to Brisbane where they enter quarantine under the supervision of AQIS, the Australian Quarantine Inspection Service. The plantlets are deflasked, acclimatized and grown for approximately one crop cycle in a
iv
quarantine planthouse. Leaf samples for virus indexing are taken from the plants at approximately three and six months after planting. In some cases, leaf samples for testing are also taken from the tissue-cultured plants prior to deflasking. As of 10 December 2003, 159 plants – traditional cultivars from the Pacific and the TANSAO collection, and breeders’ lines from breeding programmes in Papua New Guinea and Samoa – have been tested according to agreed protocols described above. Of these 159 plants, 110 tested negatively for five viruses. Other accessions from the same collections held at the RGC are growing in quarantine awaiting indexation. Results from taro surveys carried out in 10 Pacific Island countries to complement the indexing at QUT, did not detect any new viruses, and confirmed the distribution of those already known. DsMV was found in all countries, as was TaBV and the putative integrated sequence, whereas TaVCV and TaRV had more restricted distributions. The survey confirmed that CBDV, which is associated with alomae, a lethal disease, is present only in Papua New Guinea and Solomon Islands. It is expected that countries will wish to import taro from the RGC collections that have been shoot-tip cultured at the RGC and tested for viruses by QUT. Before this can be done, it is expected that some form of risk analysis will be carried out according to accepted international protocols. It is hoped that the information provided in this report will assist with this. In addition, and in order to help further with the decision making process, the report poses a number of questions that government authorities may ask in relation to the taro that are now available from the RGC. The importance of the viruses and their distribution, the sensitivity and consistency of the detection methods (as well as comments by fellow scientists, ie peer reviews), whether the entire procedure can be audited, and the need for post-entry quarantine, are discussed. In a final short section, the report asks the question: What’s next? Here reference is again made to import risk analysis and the obligations of countries under WTO Agreements and the international guidelines and standards of the IPPC to prevent or control the entry, establishment and spread of pests and diseases that have the potential to damage human health, animals, plants or the environment. A bibliography is provided for further reading; contact the SPC library ([email protected]) for assistance in obtaining the references cited.
1
1. Introduction TaroGen, the SPC/AusAID Taro Genetic Resources: Conservation and Utilisation project, began in 1998 to overcome the threat of taro leaf blight to Pacific Island countries and the loss of taro genetic resources. Breeding programmes were established in Papua New Guinea and Samoa, collections of traditional cultivars were assembled, described and compared, and from these a small number was chosen to represent the diversity of the region. These breeders’ lines and the traditional cultivars are now stored at SPC, in the Regional Germplasm Centre, a laboratory established for the purpose. Collections at the RGC not only include those from the Pacific. They also include a core collection from TANSAO – Taro Network for Southeast Asia and Oceania. The Network funded by EC INCO-DC, also began in 1998, with five Asian and two Pacific partners, and technical support from CIRAD and the Department of Plant Breeding, Wageningen University and Research Centre. Its objectives were similar to those of TaroGen: to collect, describe, rationalise, conserve and exchange taro germplasm. At the outset, countries in TaroGen and TANSAO agreed to share the results – the breeders’ lines and the core collections. There were several reasons for this collaboration: countries that have taro leaf blight wanted varieties with resistance; those yet free from the disease wanted to remove the vulnerability by importing resistant lines in case the disease spread; all countries would like to test the core collections under their unique agro-ecological environments; and where breeding programmes exist they want to use selections from the Asia/Pacific genepools to broaden the genetic base of existing stocks. Before taro can be moved internationally, however, countries have to be satisfied that the plants are not infected by viruses and other internally borne organisms. Several viruses are present in the Asia/Pacific region, distributions differ, and some are associated with severe diseases. FAO/IBPGR guidelines, developed in 1989 for the safe movement of edible aroid germplasm, suggest that vegetative propagating material should be transferred as sterile cultures derived from meristem-tips, and tested for microbial contamination and viruses. In 1998, sensitive tests to detect taro viruses were not available. Consequently, TaroGen sought assistance from ACIAR, and a year later a project, Virus Indexing and DNA Fingerprinting for the International Movement and Conservation of Taro Germplasm, began, with the virus component contracted to the Plant Biotechnology Program, Science Research Centre, Queensland University of Technology. The project has investigated the four taro viruses known at the start of the project, plus another newly discovered. Highly sensitive diagnostic techniques are now available. In addition, the virus distributions are better known after surveys in ten Pacific Island countries. This report summarises the research of the last five years. It describes the new diagnostic techniques, and reports on their application to the breeders’ lines and core collections from the Asia/Pacific region. In doing this, it is hoped that the information will be useful to countries when making decisions on the importation of taro germplasm.
2
2. The agreement to share Pacific Island countries, meeting at the first TaroGen Project Planning Workshop, 3-4 September 1998, at the Horticulture and Food Research Institute of New Zealand Ltd., Auckland, formulated a Code of Conduct for the sharing of taro germplasm. Under this Code of Conduct, TaroGen partners agreed that: 1. Taro germplasm acquired under TaroGen is for research purposes only and remains
the property of the original source country. 2. Germplasm acquired under TaroGen will be freely exchanged between the
participants of the project. 3. Any material acquired in the course of the TaroGen project will not be transferred
beyond the participants listed under Section D of this Code of Conduct without Prior Informed Consent of the original source country and without the use of Material Transfer Agreements.
Further, the countries decided that acquisition and transfer of taro germplasm amongst TaroGen partners would be under the following conditions: • Taro germplasm obtained through TaroGen will be maintained at the Regional
Germplasm Centre (RGC) within the Secretariat of the Pacific Community (SPC), Suva, Fiji, and acquired under Germplasm Acquisition Agreements. The SPC will be responsible for germplasm held in the RGC.
• Taro germplasm constitutes all forms of genetic materials such as seeds, vegetative
propagules, tissue-cultured plantlets and DNA. As the pests and diseases of quarantine concern may be associated with this germplasm, TaroGen will follow FAO/IBPGR Technical Guidelines for the Safe Movement of Edible Aroid Germplasm (1989) for all genetic materials transferred between participants.
The countries and institutions agreeing to exchange germplasm under the Code of Conduct are the following: Ministry of Agriculture, Cook Islands; Ministry of Agriculture, Fisheries and Forestry, Fiji; Department of Agriculture and Livestock, Papua, New Guinea; Ministry of Agriculture, Forestry, Fisheries and Meteorology, Samoa; Ministry of Agriculture and Fisheries, Solomon Islands; Ministry of Agriculture, Tonga; Ministry of Agriculture, Forestry, Fisheries and Livestock, Vanuatu; National Agricultural Research Institute, Papua New Guinea; Agricultural Development in the American Pacific; Australian Centre for International Agricultural Research; International Plant Genetic Resources Institute; Secretariat of the Pacific Community; The Horticulture and Food Research Institute of New Zealand Ltd.; and the University of the South Pacific, Samoa. Countries associated with TANSAO – Taro Network for Southeast Asia and Oceania agreed to share germplasm with the Pacific without an agreement or code of conduct.
3
3. The viruses and their diseases Viruses from four taxonomic groups had been recorded from the Pacific. Dasheen mosaic virus (DsMV) is everywhere; Taro bacilliform virus (TaBV) occurs throughout the region, often without producing symptoms; two rhabdoviruses occur, with Taro vein chlorosis virus (TaVCV) in both south and north of the Pacific, and Colocasia bobone disease virus (CBDV) confined to Papua New Guinea and Solomon Islands, associated with different diseases depending upon the variety; and Taro reovirus (TaRV), with wide distribution in the Pacific. Some of the viruses occur as latent infections, and all can be found in various combinations; this means that it is difficult to ascribe symptoms to infection by individual viruses. It means, too, that is it difficult to diagnose field symptoms, especially where viruses are likely to be present as complexes. 3.1 Dasheen mosaic virus (DsMV) DsMV has been well characterised; however, the studies used isolates mostly from the USA and Taiwan. DsMV is a potyvirus, with filamentous, flexuous, particles, approximately 750 nm long. 3.1.1 Significance DsMV is an important virus, affecting taro wherever it is grown. It also affects other edible aroids as well as ornamentals in the family Araceae. Loss of corm yield of around 20 per cent has been reported in taro, more in the related ornamental species. It is difficult to estimate the effects of the virus in the field, as it is difficult to keep plants free from virus in the field for comparative purposes. 3.1.2 Symptoms Plants show a variety of mosaic patterns: small, irregular, scattered, grey/green (sometime white) patches along or between the major veins (Fig. 1) or brilliant white or yellow feather-like patterns along the veins, sometimes throughout the leaf blade (Fig. 2). The diversity of symptoms has sometimes caused confusion, even to the extent that different pathogens were thought to be responsible, until more detailed analyses showed otherwise. Invariably, plants recover from the symptoms, producing leaves healthy in appearance. By contrast, plants in French Polynesia have been reported with small, stunted and severely distorted leaves. Some leaves are reduced to strap-like structures without leaf blades. These plants often fail to recover.
4
Fig. 1. Dispersed mosaic patterns caused by DsMV (Fiji)
Fig. 2. DsMV causing feather-like mosaic patterns throughout the leaf (New Caledonia)
3.1.3 Hosts The edible aroids (Alocasia, Amorphophallus, Cyrtosperma, Xanthosoma), and the ornamentals (Caladium, Dieffenbachia, Zantedeschia), and other Araceae are hosts. 3.1.4 Geographical distribution Worldwide. There is evidence of a more severe strain in French Polynesia. 3.1.5 Transmission Mechanically, and by aphids (non-persistently) under natural conditions. The common aphid on taro in the Asia/Pacific region is the cotton aphid, Aphis gossypii. Other aphids (Aphis craccivora and Myzus persicae) are reported as vectors, but not the banana aphid, Pentalonia nigronervosa. Seed and pollen transmission is not thought to occur. 3.1.6 Detection At QUT, diagnostic tests have been developed that use both serological (ELISA) and molecular (PCR) methods. These have proven to be both sensitive and robust. For PCR, degenerate primers were designed based on the sequences of numerous virus isolates collected throughout the Pacific, to ensure they amplified virus sequences from all the countries surveyed.
5
The sequence of the DsMV primers is: DsMV 3F 5’ AGTACAAACCTGARCAGCGTGAYA 3’ DsMV 3R 5’ TTYGCAGTGTGCCTYTCAGGT 3’ 3.2 Taro bacilliform virus (TaBV) 3.2.1 Significance The virus is commonly found in taro throughout the Asia/Pacific region, and appears to occur as numerous strains. Alone, it appears to have minimal effect on plant growth. It has been associated with CBDV in Solomon Islands and PNG, causing alomae, a lethal disease (see 3.4). TaBV is a bacilliform virus, approximately, 130 x 30 nm, and has recently been characterised and classified as a species in the genus Badnavirus. A TaBV-like sequence has also been identified from all countries of the Pacific where tests have been made. It has been found in plants without symptoms as well as in those known to be infected with TaBV. This sequence is 50 per cent similar to TaBV, but to date there is no evidence that it represents a new virus. 3.2.2 Symptoms The virus is often latent, present in plants without causing symptoms. Where symptoms occur, they are seen soon after planting, as intermittent, indistinct areas of vein chlorosis, often near the leaf margin (Fig. 3). Fig. 3 TaBV with faint vein chlorotic patterns, more obvious near the leaf margin (Samoa)
Fig. 4 TaBV showing distinct vein chlorosis on leaf curled under at margins (Samoa)
6
Frequently, the leaf blades are bent backwards, and sometimes puckered. Rarely, vein chlorosis occurs throughout the leaf (Fig. 4). Plants may be stunted, but eventually recover, producing apparently healthy leaves. 3.2.3 Hosts In Colocasia, but also in Xanthosoma and Alocasia. There is an unconfirmed report in Commelina benghalensis in Samoa (Macanawai, 2003). 3.2.4 Geographical distribution TaBV is widespread in the Pacific, and occurs in some TANSAO lines of Asian origin. The TaBV-like sequence has been found in taro from the Federated States of Micronesia, Fiji, New Caledonia, Niue, PNG, Samoa, Solomon Islands, Tonga and Vanuatu. It has also been detected in TANSAO lines from Indonesia and Malaysia grown in Vanuatu. 3.2.5 Transmission In Samoa, transmitted by the mealybug, Pseudococcus solomonensis. Also spreads in seed and pollen (Macanawai, 2003). Earlier, in Solomon Islands, the mealy bugs, Planococcus citri and Pseudococcus longispinus (Gollifer et al., 1977) were also thought to be vectors. 3.2.6 Detection At QUT, primers (TSBV 1 and TSBV 4) for a PCR-based diagnostic test have been designed based on sequences of TaBV isolates collected throughout the Pacific. Although Yang et al. (2003b) observed considerable sequence variability in the genome of TaBV isolates from the region, the chosen degenerate1 primers have proven to be robust and detected TaBV in all the countries studied. The sequence of the TaBV primers is: TSBV-1 5’ CKSTGYAARSAACATGGTCTTG 3’ TSBV-4 5’ TAATCAAGYGGWGGGAGYTTCTC 3’ In these tests, there are two controls: a TaBV-positive sample, to ensure that the diagnostic primers are working effectively; and an internal PCR control using two primers based on chloroplast DNA. Control 1F 5’ GAGCCGTAGCGAAAGCGAGTC 3’ Control 1R 5’ GGCATTTCACCCCTAACCACAAC 3’
1 more than one nucleotide synthesised at a given position on the primer; ie an "r" in the primer denotes either an "a" or a "g" at that position
7
Another PCR test is available for the TaBV-like sequences thought to be integrated into taro chromosomes of plants and seeds. The primers are: PNG-BadnaF 5' GCAAAAAGAATGGATTGATACTGAGCAG3' and PNG-BadnaR 5' CATTTTTCTTTCTCCAGTTGGGCTTAC 3.3 Taro vein chlorosis virus (TaVCV) 3.3.1 Significance TaVCV is found in several Pacific countries, but its effect on plant growth appears to be minimal. It is a rhabdovirus, approximately 210 x 70 nm. 3.3.2 Symptoms Symptoms are commonly seen on plants when they are at maximum growth, rather than after planting or at maturity as with other taro viruses. Leaves show a distinct vein chlorosis, more pronounced than TaBV (Figs. 5 and 6). As the leaves age, the chlorosis spreads between the veins. Large, newly emerged, leaves may droop at the margins, giving an umbrella effect (Fig. 5). The number of leaves showing symptoms varies, but in most cases it is three or four. Leaves formed subsequently appear healthy. By contrast to infection from CBDV, which is also a rhabdovirus, galls are not present on the leaf blades and petioles, and plants are not stunted. Fig. 5 Young leave with ‘umbrella’ appearance from infection by TaVCV (Vanuatu)
Fig. 6 Young (right) and old (left) leaves infected with TaVCV showing spread of chlorosis with age (Vanuatu)
8
3.3.3 Hosts Found only in Colocasia. 3.3.4 Geographical distribution Recorded from the Federated States of Micronesia, Fiji, PNG, Republic of Palau, Solomon Islands, Tuvalu, Vanuatu, New Caledonia, Federated States of Micronesia and the Philippines. 3.3.5 Transmission Not known, but most likely by Tarophagus proserpina, or related planthoppers (by analogy with CBDV). Not likely by seed or pollen. 3.3.6 Detection At QUT, a diagnostic test has been developed using specific primers and PCR to amplify virus nucleic acid. The primers detected virus throughout the Pacific Islands, but the sequences have yet to be published. 3.4 Colocasia bobone disease virus (CBDV) 3.4.1 Significance CBDV occurs in only two countries and is associated with bobone disease in certain varieties of taro and alomae in others. It is often found with TaBV and TaVCV. CBDV is a rhabdovirus, approximately 300 x 50 nm. 3.4.2 Symptoms In those varieties susceptible to bobone, severe stunting occurs, especially after planting, with distorted, thickened, brittle leaves (Fig. 7). Galls may be present on the petioles and sometimes on the larger veins (Fig. 8). Plants recover after 3-5 affected leaves, with about 25% loss in corm yield. Sometimes symptoms reoccur after recovery. In those varieties susceptible to alomae there are two types of symptoms: a) leaves show green wrinkled patches mostly between the major veins (Fig. 9), or b) early symptoms are similar to bobone, but new leaves stay rolled, become chlorotic, rot and die (Fig. 10); this disease is known as alomae (meaning taro dies, in the local language where the disease was first described).
9
Fig. 7 Stunted plant with curled, twisted, leaves with bobone disease and infected by CBDV (PNG)
Fig. 9 CBDV infection equivalent to bobone, in a plant susceptible to alomae (Solomon Islands)
Fig. 8 Plant with bobone showing galls on the petiole (PNG)
Fig. 10 Plant with young tightly rolled leaves, typical of alomae, and associated with infection by CBDV and other viruses (Solomon Islands)
3.4.3 Host Found only in Colocasia.
10
3.4.4 Geographical distribution Only recorded in taro from PNG and Solomon Islands. 3.4.5 Transmission By the planthopper, Tarophagus proserpina. Not mechanically, nor thought likely by seed or pollen. 3.4.6 Detection At QUT, a diagnostic test has been developed using specific primers and PCR to amplify virus nucleic acid. The primers detected virus throughout Solomon Islands and PNG. The sequences have yet to be published. 3.5 Taro reovirus (TaRV) 3.5.1 Significance There is no information on the significance of TaRV. It appears to be a reovirus based on the isolation of 10 segments of dsRNA from infected taro plants. Although virions have not yet been purified, they are typically spherical particles, approximately 65-75 nm in diameter. 3.5.2 Symptoms No symptoms have been associated with infection by TaRV. The virus has only been detected together with other viruses. However, taro virus surveys invariably chose plants with virus-like symptoms; thus symptoms of TaRV may have been obscured by those of other viruses. 3.5.3 Hosts Found in Colocasia. 3.5.4 Geographical distribution TaRV has been recorded from PNG, Solomon Islands and Vanuatu. 3.5.5 Transmission Unknown in taro, but plant reoviruses are transmitted by phloem-feeding insects, eg leafhoppers and planthoppers, not mechanically, and not by seed.
11
3.5.6 Detection At QUT, a diagnostic test has been developed using specific primers and PCR to amplify virus nucleic acid. Comparisons were made with other reoviruses and primers were made to conserved regions of the virus dsRNA. They were later refined after examination of nucleotide variability of numerous virus isolates throughout the Pacific to ensure that they amplified virus sequences from the countries surveyed. The sequences have yet to be published. 4. The methods for virus detection This section provides details of the laboratory at QUT where the indexing procedures were developed as well as the methods used for indexing taro germplasm for the viruses described in section 3. 4.1 The laboratory and its standards The Plant Biotechnology Group within the Science Research Centre, QUT, was selected by ACIAR because of its leadership in the field of plant virus diagnostics, especially those of tropical and subtropical crops. . The laboratories are modern with state-of-the-art equipment. Further, the group already had extensive overseas experience, particularly in Southeast Asia, through ACIAR projects The Centre is approved by AQIS to import and analyse plant material (permit Q0327). It is inspected annually by AQIS to ensure it complies with the requirements of the permit: all material is stored correctly (locked cupboards or freezers); appropriately labelled (yellow quarantine stickers); and that all samples listed on the permit can be traced and have been recorded on a database. In this way, if an audit is required it can be done with the accuracy required. 4.2 Methods of indexing 4.2.1 General All the tests carried out to detect taro viruses in leaf tissues, seed or pollen were done on nucleic acid extracts. After extraction, the nucleic acids were subjected to PCR with primers specifically designed to increase (amplify) a portion of the viral genetic material, if present. PCR involves rapid heating and cooling of DNA in the presence of virus-specific sequences (primers), a polymerase enzyme, nucleotides and buffer. With 2-3 hours a sufficient quantity of DNA is produced and this appears as a band on an agarose gel. By manipulating the heating and cooling temperature of the PCR program and through the
12
use of specific primers, it is possible to amplify only the viral sequences. The size of the product amplified is defined by the distance between the primers where they bind to the viral genome. For example, if two primer sequences are 500 nucleotides apart on a viral genome, the resultant PCR product will be 500 nucleotides in length. The technique is extremely sensitive and, in theory, one molecule of DNA can be amplified many millions of times following PCR. In practice, the technique is not as sensitive as this, as plant DNA/RNA extracts contain chemicals that inhibit the PCR reaction. Another problem is that viruses are often unevenly distributed throughout a plant. Thus, samples need to be taken from a number of different leaves to ensure the best possible chance of detecting the virus of concern. 4.2.2 The use of controls It is very important to have appropriate controls in the PCR reaction, as interpretation of PCR results can sometimes be difficult, and on some occasions non-specific bands, amplified from plant genomic DNA, may appear on the gel. AT QUT, three controls are used: • The first is a DNA sample from a known infected plant (positive control). This shows
us that the PCR test is working and produces a band of the correct size against which all other samples are compared.
• The second is a sample from a plant not infected with the virus being tested (healthy control). This should not amplify a band of the same size as the positive control, but will confirm if any non-viral bands are amplified.
• The third is a water-only control, to confirm that none of the reagents are contaminated. No bands should be amplified from the water control.
• For the TaBV test, we also have an “internal control”. That is, using primers that amplify nucleic acid from chloroplast DNA in every sample, we are able to show that the PCR test is functioning properly, even if virus is not detected in the samples. Unfortunately, for the RNA virus tests (CBDV, DsMV, TaRV, TaVCV) we are unable to include this control, but this will be developed in the future.
The details for each of the tests are given in Appendix 1. On some occasions, bands obtained from test samples may be very faint, or slightly different in size to the predicted virus band. In these cases, the bands are analysed further using a technique called Southern hybridization. The definitive viral PCR band is labeled with a chemical and this labeled band is called a “probe”. The DNA bands from the PRC reactions on an agarose gel are transferred to a nylon membrane, which is then “probed”. Only virus-specific bands are visible following exposure to x-ray film, enabling confirmation of the PCR result. The probe labeling technique used is DIG-labeling and the full protocol is available in the Roche Diagnostics manual. For an explanation of Southern hybridisation in general, see Sambrook et al. (1989).
13
5. Therapy and indexing The SPC RGC maintains large collections of taro that have undergone shoot-tip culture and indexation for viruses. These collections are made up of the following: • TaroGen core collections from 10 Pacific Island countries. • TaroGen breeders’ lines from PNG and Samoa TLB resistance breeding programmes. • TANSAO core collection. • Elite varieties from several countries obtained prior to TaroGen (and indexed by HRI,
Littlehampton, UK) • Selected accessions from the Federated States of Micronesia, Palau, Philippines, New
Caledonia and Vanuatu All the accessions in the TaroGen and TANSAO collections were cultured from meristems at the RGC, and sent as in vitro plantlets to AQIS, Australia, where they were grown in post-entry quarantine and then indexed by QUT. The next section describes the methods. 5.1 Eliminating viruses All of the accessions selected for virus indexing are established in tissue culture from shoot-tips (approximately (0.8mm), according to established techniques. This was done either in the country of origin or, where this was not possible (collections from PNG, Solomon Islands and Vanuatu) at the School of Land and Food Sciences, University of Queensland. The Fiji collection was cultured at the RGC, and the Niue collection in Tonga. The TANSAO lines of the Asia core collection were sent from LIPI, Indonesia to the RGC as plantlets growing in tissue culture. They had been indexed for DsMV by the Department of Plant Breeding, Wageningen University and Research Centre using a commercial antiserum. They contained accessions that tested positive for DsMV. The plantlets were re-established from shoot-tips at the RGC. Each shoot-tip is given a unique number at the RGC, which is retained by the plantlet when sent to PEQ, Brisbane, and when indexed by QUT. Numbers given to shoot-tips/plantlets that died are not reassigned. Details of all accessions are entered into a database that describes treatments, tests and transfers, with relevant dates. 5.1.1 Post-entry quarantine, Australia When plantlets grown from shoot-tips reach an appropriate size (about 6 months) they are sent to Australia to be grown in PEQ, and subsequently indexed for viruses by QUT. It is important to note that the plantlet grown from the original shoot-tip (not its suckers) is sent for indexing. This is done in case plants are infected by virus, but the particles are not distributed uniformly. If no virus is detected in the plant grown from the original
14
shoot-tip, is it reasonable to assume that its suckers will be free of virus, too. But the reverse is not necessarily true: freedom of virus in the suckers does not mean that the plant grown from a shoot-tip is free from infection. Upon arrival in Australia as tissue cultures, the plantlets are held over in quarantine for approximately 12 hours and then released for planting in the AQIS PEQ facility at Eagle Farm, Brisbane, under the supervision of the Principal Technical Officer, DPI. There they are de-flasked, transferred to a perlite/peat/sand mix (50:30:20), and kept under high humidity in a Perspex cabinet to acclimatise before transfer to larger pots. They remain at the facility for 9 months, or approximately one crop cycle. During that time, they are observed frequently for virus symptoms and tested for all known taro viruses on two occasions. This is necessary because some viruses are known to occur at very low concentrations in tissue cultured plants and may remain undetected. The concentration of virus in plants is expected to rise considerably when they are growing vigorously. 5.1.2 Testing for viruses, QUT Each plantlet is indexed for viruses at least twice at QUT, using the protocols outlined in Section 3 and detailed in Appendix 1. Leaf samples for virus indexing are taken from the taro plants growing in PEQ at approximately three and six months after planting. On each occasion, tissue is sampled from 2-3 leaves, in case there is uneven distribution of virus inside the plants. In some cases, leaf samples for testing are also taken from the tissue cultured plants prior to deflasking. 6. Indexing results All 159 accessions sent to QUT were tested for five viruses (DsMV, TaBV, TaVCV, TaRV, CBDV) on two occasions while growing in PEQ. Because of the importance of CBDV, Southern hybridisation tests were carried out to check PCR results. In some cases, leaves for virus testing were also taken from tissue-cultured plants prior to de-flasking. Of the 159 accessions, 110 were negative for all five viruses (Table 1). Five of the 49 that were positive contained DsMV, the remainder, TaBV.
15
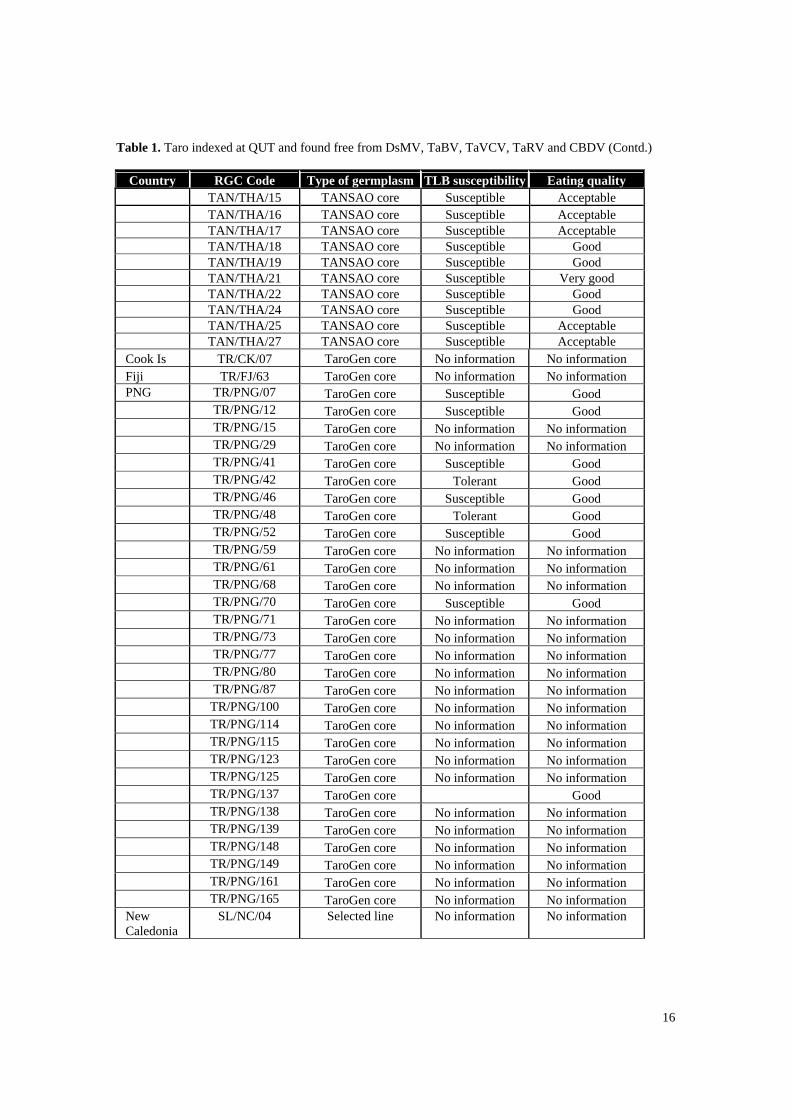
Table 1. Taro indexed at QUT and found free from DsMV, TaBV, TaVCV, TaRV and CBDV
Country RGC Code Type of germplasm TLB susceptibility Eating quality Indonesia TAN/IND/06 TANSAO core Susceptible Good TAN/IND/07 TANSAO core Susceptible Good TAN/IND/08 TANSAO core Susceptible Good TAN/IND/09 TANSAO core Tolerant Good TAN/IND/10 TANSAO core Susceptible Good TAN/IND/13 TANSAO core Tolerant Very good TAN/IND/14 TANSAO core Susceptible Good TAN/IND/15 TANSAO core Susceptible Good TAN/IND/16 TANSAO core Susceptible Good TAN/IND/17 TANSAO core Susceptible Good TAN/IND/19 TANSAO core Tolerant Very good TAN/IND/20 TANSAO core Tolerant Very good TAN/IND/22 TANSAO core Susceptible Good TAN/IND/23 TANSAO core Tolerant Good TAN/IND/24 TANSAO core Susceptible Poor TAN/IND/27 TANSAO core Susceptible Good TAN/IND/30 TANSAO core Susceptible Good TAN/IND/31 TANSAO core No information No information Malaysia TAN/MAL/02 TANSAO core Immune Very poor TAN/MAL/03 TANSAO core Immune Very poor TAN/MAL/06 TANSAO core Immune Very poor TAN/MAL/07 TANSAO core Resistant Very good TAN/MAL/08 TANSAO core Resistant Very good
TAN/MAL/09 TANSAO core Immune Very good TAN/MAL/10 TANSAO core Immune Very good TAN/MAL/11 TANSAO core Resistant Very good TAN/MAL/12 TANSAO core Resistant Very good TAN/MAL/13 TANSAO core Immune Good TAN/MAL/14 TANSAO core Resistant Poor Philippines TAN/PHL/02 TANSAO core Tolerant Very good TAN/PHL/05 TANSAO core Tolerant Very good TAN/PHL/06 TANSAO core Tolerant Good TAN/PHL/09 TANSAO core Tolerant Acceptable TAN/PHL/10 TANSAO core Tolerant Not edible TAN/PHL/13 TANSAO core Tolerant Acceptable TAN/PHL/14 TANSAO core Tolerant Good TAN/PHL/15 TANSAO core Tolerant Excellent PNG TAN/PNG/02 TANSAO core No information No information TAN/PNG/13 TANSAO core Susceptible Excellent TAN/PNG/14 TANSAO core No information No information Thailand TAN/THA/01 TANSAO core Susceptible Good TAN/THA/02 TANSAO core Susceptible Good TAN/THA/03 TANSAO core Susceptible Good TAN/THA/04 TANSAO core Susceptible Acceptable TAN/THA/07 TANSAO core Susceptible Good (eddoe) TAN/THA/09 TANSAO core Susceptible Acceptable TAN/THA/10 TANSAO core Susceptible Acceptable TAN/THA/13 TANSAO core Susceptible Acceptable
16
Table 1. Taro indexed at QUT and found free from DsMV, TaBV, TaVCV, TaRV and CBDV (Contd.)
Country RGC Code Type of germplasm TLB susceptibility Eating quality TAN/THA/15 TANSAO core Susceptible Acceptable TAN/THA/16 TANSAO core Susceptible Acceptable TAN/THA/17 TANSAO core Susceptible Acceptable TAN/THA/18 TANSAO core Susceptible Good TAN/THA/19 TANSAO core Susceptible Good TAN/THA/21 TANSAO core Susceptible Very good TAN/THA/22 TANSAO core Susceptible Good TAN/THA/24 TANSAO core Susceptible Good TAN/THA/25 TANSAO core Susceptible Acceptable
TAN/THA/27 TANSAO core Susceptible Acceptable Cook Is TR/CK/07 TaroGen core No information No information Fiji TR/FJ/63 TaroGen core No information No information PNG TR/PNG/07 TaroGen core Susceptible Good TR/PNG/12 TaroGen core Susceptible Good TR/PNG/15 TaroGen core No information No information TR/PNG/29 TaroGen core No information No information
TR/PNG/41 TaroGen core Susceptible Good TR/PNG/42 TaroGen core Tolerant Good TR/PNG/46 TaroGen core Susceptible Good TR/PNG/48 TaroGen core Tolerant Good TR/PNG/52 TaroGen core Susceptible Good TR/PNG/59 TaroGen core No information No information TR/PNG/61 TaroGen core No information No information TR/PNG/68 TaroGen core No information No information TR/PNG/70 TaroGen core Susceptible Good TR/PNG/71 TaroGen core No information No information TR/PNG/73 TaroGen core No information No information TR/PNG/77 TaroGen core No information No information TR/PNG/80 TaroGen core No information No information TR/PNG/87 TaroGen core No information No information TR/PNG/100 TaroGen core No information No information TR/PNG/114 TaroGen core No information No information TR/PNG/115 TaroGen core No information No information TR/PNG/123 TaroGen core No information No information TR/PNG/125 TaroGen core No information No information TR/PNG/137 TaroGen core Good TR/PNG/138 TaroGen core No information No information TR/PNG/139 TaroGen core No information No information TR/PNG/148 TaroGen core No information No information TR/PNG/149 TaroGen core No information No information TR/PNG/161 TaroGen core No information No information TR/PNG/165 TaroGen core No information No information
New Caledonia
SL/NC/04 Selected line No information No information
17
Table 1. Taro indexed at QUT and found free from DsMV, TaBV, TaVCV, TaRV and CBDV (Contd.)
Country RGC Code Type of germplasm TLB susceptibility Eating quality Vanuatu TR/VAN/27 TaroGen core No information No information Palau PAL/02 Selected line No information No information
PAL/03 Selected line Resistant Acceptable PAL/06 Selected line No information No information PAL/20 Selected line Resistant Good
PNG BL/PNG/06 Breeders’ line Highly resistant Good PNG BL/PNG/10 Breeders’ line Resistant Acceptable PNG BL/PNG/11 Breeders’ line Resistant Acceptable PNG BL/PNG/12 Breeders’ line Resistant Acceptable PNG BL/PNG/13 Breeders’ line Resistant Acceptable Samoa BL/SM/10 Breeders’ line Tolerant Good Samoa BL/SM13 Breeders’ line Tolerant Good
1Information on the characteristics of the varieties was collected in the country of derivation of the cultivars: there is no guarantee that the same TLB resistance or corm quality will be maintained when the plants are grown elsewhere. 7. The Survey The 4th meeting of TaroGen, 28-29 November 2000, recommended a regional survey of taro viruses, to provide information that would complement the results of virus indexing. This was endorsed by the 14th PHALPS, Nadi, Fiji, 30 April-4 May 2001. Surveys for taro viruses were last done in the region under the FAO/UNDP root crops project in the 1980s. Since then, hitherto unrecognised viruses have been discovered, some of those already identified had spread, and detection methods improved. Thus, an intensive survey in countries wishing to share germplasm under TaroGen would clarify virus distributions. It would provide information that would complement the results from virus indexing, and when taken together allow countries to make informed decisions on the importation of taro germplasm. It was also seen as a check on the presence of other viruses that might be present, but, hitherto, had escaped detection. The survey began in Vanuatu (15-22 April 2002) and, over the next 10 months, visits were made to Fiji (20-29 May), Tonga (2-6 July), Samoa (6-11 July), American Samoa (8-10 July), Solomon Islands (5-13 November), Papua New Guinea (13-25 October) and New Caledonia (8-14 December). In each country, a selection of the major taro producing areas was visited, plants with virus symptoms found, photographed, and leaves collected. From the leaves, pieces of tissue with symptoms were removed, shredded and dried over silica gel. Leaves that appeared healthy were treated the same way. Fresh leaves showing symptoms of viruses of interest were also collected for virus purification and the development or refinement of indexing protocols. Leaves of Alocasia, Amorphophallus and Xanthosoma were also examined and collected if virus-like symptoms were present. In each country, visits were made with government staff from research, extension or quarantine divisions, and occasionally staff of NGOs.
18
Taro leaves were also sent to QUT from the Cook Islands, the Federated States of Micronesia, French Polynesia and Marshall Islands. 7.1 Results from the survey The results from the survey are given in Table. 2. Overall, they confirmed the known distributions of the taro viruses. The exception being TaRV, which was unknown prior to the research at QUT. The distribution of TaBV was more widespread than realised previously. Table 2. Taro viruses found in ten Pacific Island countries during a survey in 2002
NB. TaRV was not detected in New Caledonia during the survey, but was present in samples sent by Michel Grisoni in June 2000. In other countries where TaRV was detected during the survey, it was never found alone, whereas all other viruses were detected in single as well as in mixed infections where more than one virus was present. †Samples sent from French Polynesia in March 2000, and tested by ELISA and western hybridisation. DsMV: found to be distributed throughout the Pacific. Typical spots, blotches and feathery mosaic patterns were found on plants where diagnostic tests recorded DsMV alone, and on plants which contained DsMV and other viruses. DsMV was also detected in a large number of plants that appeared healthy, confirming that infections can be latent. TaBV: found widely distributed, but symptoms are still unclear. Many of the plants that were positive for this virus either appeared healthy or were infected by other viruses. Only in Samoa were faint vein-clearing symptoms consistently associated with infection. These were most clearly seen in cultivar PSB-G2 from the Philippines. An interesting find from the survey was a TaBV-like sequence in all taro plants tested, including symptomless plants and those indexed and found free from TaBV using the primers TSBV 1and TSBV 4. Analysis of the sequences from PNG and Vanuatu showed that they are approximately 50% similar to TaBV. It is not yet known whether it is part of a second badnavirus or it exists within the taro DNA, in which case it is said to be
19
integrated into the plant genome. Integration is known to occur in other badnaviruses, eg Banana streak virus. The common occurrence of the sequence in taro suggests that it is yet another example of the phenomenon. It appears from the survey that the TaBV-like sequence is present in the TANSAO core collection. Two tested positive for the sequence – IND66 and MAL9 – but these were sampled from plants in the field, so it is possible they were infected from a local source. CBDV: found only in PNG and Solomon Islands. The result confirms previous findings and provides further evidence that it is the cause of bobone, and a component of alomae. TaVCV: found widely distributed in the Pacific. Partial analysis of the genomes of TaVCV and CBDV indicates that these viruses are definitive, yet distinct, rhabdoviruses. Plants infected by TaVCV showed distinctive vein chlorosis as previously reported. Overall, plants in Vanuatu showed more intense chlorosis than those elsewhere, but it was not known if this was due to varietal differences or other factors. TaRV: a previously undescribed virus infecting taro found in three countries. Symptoms are unknown, since the virus was detected mostly in plants infected with other viruses. As such, the impact of this virus on taro is unknown, but thought to be slight. Of the other aroids tested, Xanthosoma was found infected with DsMV and TaBV (Tonga, Solomon Islands and Vanuatu). Alocasia had infections of DsMV and TaBV (Tonga), and TaBV (Samoa). 8. Comments on the results The report describes the procedures used to test taro and other edible aroids for the presence of viruses that might be of concern to quarantine authorities. While is it not possible to say categorically that the taro that tested negatively are free from viruses, it is possible to state that the plants have been subjected to the most sophisticated and sensitive tests currently available. All methods of testing involve analysis for virus nucleic acids, involving PCR and virus-specific primers. In order to be helpful to quarantine and other authorities of Pacific Island governments as they make decisions on the importation of the indexed taro now available from the RGC, Fiji, this section considers some frequently asked questions in relation to the taro and the tests made on them. 8.1 How important are the viruses found in taro? The viruses differ greatly in their effect on taro. On their own, or together in various combinations, four of the five viruses (DsMV, TaBV, TaVCV and TaRV) appear to have little impact on yield, with, perhaps, the exception of DsMV.
20
Attempts to measure the effect of DsMV on yield in other parts of the world have found a c.20% loss in corm weight following infection (or the expression of symptoms), but none of the reports is convincing. There is, however, a severe disease associated with DsMV in French Polynesia. Once infected, the plants do not recover, although there is no report that they die. This disease is unusual, and may be due to a severe form of the virus. The plants seen with the severe symptoms in French Polynesia are the same as those that occur in other parts of Polynesia, where DsMV is present, but such severe symptoms have not been reported. Therefore, it is unlikely that they are due to the varieties in French Polynesia being more susceptible to infection by DsMV. None of the taro now available from the RGC originated from French Polynesia. Of the other three viruses, TaBV and TaVCV produce mild symptoms – chlorosis without distortions – and TaBV is often latent. No symptoms have yet been associated with TaRV, which may also be latent. The virus with the greatest potential to impact on taro production is CBDV. On its own it causes bobone, a non-lethal leaf distortion in some (perhaps 10) varieties. In the remainder, and that is the majority, it causes a mild leaf crinkle, either alone or with TaBV. But where it occurs with TaBV (and perhaps also with TaVCV), the result is a lethal disease – alomae. The etiology of alomae is far from clear, but it does seem likely that CBDV is the key component. CBDV is spread by plant hoppers, Tarophagus spp. Three species are now recognised in the Pacific, and it is likely (although not proven) that each could spread the virus. However, as the virus is persistent in the host, with several days between acquisition and transmission, it can be controlled by insecticides. (This is not the case with DsMV, which is non-persistent in its aphid vector, and lost soon after acquisition.) However, insecticides are unlikely to be used on taro grown as a subsistence crop, so every attempt must be made to prevent the further spread of CBDV. 8.2 Can we be sure of the distribution of the viruses from the
survey? The surveys carried out in eight Pacific Island countries looked at many thousands of taro in areas where it is a major crop. Large numbers of plants were sampled and these were analysed for all the viruses of concern. In a majority of cases, the results of the analyses were in keeping with the symptoms observed. Where there were disparities they related to viruses that are latent in taro or symptoms were complicated by the complexity of viruses present. Although, it is thought unlikely that viruses were overlooked, it is difficult to be categorical about this. Where visits were not made, and instead samples were sent for analysis, that is, Cook Island, the Federated States of Micronesia and Marshall Islands, there is a possibility that viruses were overlooked. However, the analyses from these countries agreed with surveys made previously.
21
8.3 How sensitive are the indexing methods? The indexing methods (PCR and Southern hybridization) are the most sensitive available. Great effort has been taken to ensure that the primers used in the PCR tests detect as many different isolates as possible, hence many of the primer pairs used in this study are degenerate primers, based on more than one viral genome. It has not been possible to design degenerate primers as yet for TaVCV and CBDV, as the viral sequence has only been obtained for one isolate and variability studies are yet to be completed. However, the primers used, do detect virus isolates from a range of countries, although it is impossible to say that any primer set, be they degenerate or specific, will detect all virus sequences. 8.4 Have the methods been peer reviewed? Research papers and theses have been published describing the virus indexing methods for TaBV, DsMV and TaRV (Dowling, 2002; Macanawai, 2003; Yang, 2003a &b), and primer sequence data have been provided. For the other viruses, papers are in preparation. A list of the publications is given in Section 9. 8.5 Where the same plants tested over time and where the results
consistent? To date, 159 plants have been tested on thee occasions, the majority as tissue cultures, and then all at 3 and 6 months in PEQ. The results were consistent over the three tests. Only two plants were found to be negative in tissue culture and then positive for DsMV in PEQ. This was not an unexpected result. Virus may be in low concentration in tissue cultures and/or unequally distributed in the leaves. It is for this very reason that plants are grown in PEQ, and tested twice more during the growing cycle. During the growth of the plants, the viruses multiply, and this improves the chance of detection. (Because the tests on tissue cultures are not reliable and often there is insufficient leaf material to index for all five viruses, they will not be done in the future.) 8.6 Is it possible to audit the system, in case something goes wrong? Yes, the protocols for shoot-tip culture and virus indexing established by SPC and QUT are documented in detail, as are the PEQ activities of AQIS. If necessary, an audit can be done on any accession, tracing its origins from the RGC to PEQ in Australia. All the accessions are given a dedicated number, and everything that is done to any accession is recorded in lab notebooks and on a database. 8.7 Should the taro be grown in PEQ in the importing country? This is a decision for the quarantine authorities of the importing country. Our recommendation is that no further closed PEQ is necessary. However, we strongly
22
recommend that all material be inspected for any abnormalities during the multiplication stage by suitably trained personnel. 8.8 What is known about the taro varieties? TaroGen is producing a Tarologue, similar to that developed by INIBAP for banana varieties. In general, the varieties that are available are either part of the core collection of the Pacific and Asia, and thus represent wide diversity useful in selection and breeding programmes, or are from current breeding programmes in PNG and Samoa and are resistant to taro leaf blight. Table 1 provides brief notes on the varieties that are now available. 8.9 What to do next? National authorities concerned with agricultural development in Pacific Islands can now consider the importation of taro from the Pacific and Asia genepools knowing that they have been indexed by the most advanced technologies available. They can be obtained as tissue cultures from the SPC RGC with supporting documentation verifying the results of the testing. In making decisions on the importation of the taro now available, it is expected that authorities will do so with reference to the WTO Agreement on the Application of Sanitary and Phytosanitary Measures (SPS Agreement). The risks will be considered and measures applied to reduce the risks in a manner considered appropriate under the Agreement to protect “human, animal or plant life or health”, based on scientific principles. In order to do this, authorities may wish to consult international guidelines and standards on import risk analysis developed under the International Plant Protection Convention2.
(IPPC) such as ISPM 2 and ISPM 11. The ISPM 11 Pest Risk Analysis for Quarantine Pests is relevant in this context: Section 2.3 Assessment of Potential Economic Consequences states that:
In many instances, detailed analysis of the estimated economic consequences is not necessary if there is sufficient evidence or it is widely agreed that the introduction of a pest will have unacceptable economic consequences (including environmental consequences). In such cases, risk assessment will primarily focus on the probability of introduction and spread. It will, however, be necessary to examine economic factors in greater detail when the level of economic consequences is in question, or when the level of economic consequences is needed to evaluate the strength of measures used for risk management or
2 ISPMs are the standards, guidelines and recommendations recognized as the basis for phytosanitary measures applied by members of the World Trade Organization under the Agreement on the Application of Sanitary and Phytosanitary Measures. They are developed by the Commission of Phytosanitary Measures under the IPPC. They can be found at: http://www.ippc.int/servlet/CDSServlet?status=ND0xMzM5OS4xNjE0MiY3PWVuJjYxPXB1YmxpY2F0aW9ucyY2NT1pbmZv
23
in assessing the cost-benefit of exclusion or control.
In the present context, the “probability of introduction” might be considered extremely low, considering the treatments that the taro have undergone, the shoot-tip culture and the extensive testing using sensitive molecular techniques. Further, the Agreement on the Application of Sanitary and Phytosanitary Measures states, in relation to consequences, that:
Members shall take into account as relevant economic factors; the potential damage in terms of loss of production or sales in the event of entry, establishment or spread of a pest or disease; the costs of control or eradication in the territory of the importing Member; and the relative cost-effectiveness of alternative approaches to limiting risks.
The IPPC, in ISPM no. 11, differentiates between the ‘direct’ and ‘indirect’ effects of a pest. Direct effects are that those relating to host damage, crop losses and the environment, the biology of the pest, including its adaptability and virulence, spread and control, as well as the impact of the pest on present production practices. Indirect effects concern the socio-economic impact of the pest incursion, on trade (domestic and export markets), cost of control and further research, and the impact of control measures on the environment. However, there are acknowledged difficulties for Pacific Island countries in coming to terms with the intentional standards and agreements. The greatest difficulty is in relation to limited management capabilities to address quarantine pests identified during any risk analysis3. To overcome this, countries have recommended that major quarantine pests be considered ‘Critical’, and that they should be managed offshore. This is an important consideration. Countries may conclude that the taro now available from the SPC RGC are in accord with that recommendation: they have been through a lengthy period of off-shore quarantine, in Australia under the supervision of AQIS, and tested thoroughly by QUT for all known taro viruses over a period of 9 months. This may obviate the need for PEQ in the importing countries. Finally, for some countries the analysis of risk associated with the importation of in vitro, virus-indexed taro will need to be weighed against the economic consequences of exclusion. For instance, those countries yet vulnerable to taro leaf blight, may take the view that the economic consequences of taro leaf blight, should it be introduced, are so great that they far outweigh the risk that may be associated with the import of indexed taro varieties resistant to the disease. They might decide to protect the economy of the country by importing resistant germplasm for distribution to growers ahead of the event. Even those countries where the disease exists, may wish to consider the economic impact of controlling the taro leaf blight using the resistant varieties now available.
3 Import Risk Analysis Workshop Manual (In preparation). A Manual prepared by Robert Ikin on behalf of the Secretariat of the Pacific Community, Fiji.
24
9. Bibliography Attoui, H. Billoir, F. Cantaloube, J.F. Biagini, F. de Micco, P. and X. de Lamballerie.
(2000). Strategies for the sequence determination of viral dsRNA genomes. J Virol Meth 89, 147-158.
Brunt, A.A. Crabtree, K. and A.J. Gibbs. (1990) Viruses of Tropical Plants. CABI, Wallingford UK, pp. 242-243.
Catia Delmiglio (2003). The effects of virus diseases on the growth and distribution of Sicyos australis, and molecular and morphological characterisation of the species in New Zealand. MSc Thesis: The University of Auckland.
Choi, Y.G. and J.W. Randles. (1997) Microgranular cellulose improves dsRNA recovery from plant nucleic acid extracts. BioTechniques 23, 610-611.
Dowling, M (2002). Partial characterisation of Taro reovirus and Taro vein chlorosis virus using a single primer amplification technique. BSc Honours thesis. School of Life Sciences, QUT.
Gollifer, D.E. and J.F. Brown. (1972) Virus diseases of Colocasia esculenta in the British Solomon Islands. Plant Dis Rep 56, 597-599.
Gollifer, D.E. Jackson, G.V.H. Dabek, A.J. Plumb, R.T. and Y.Y. May. (1977) The occurrence and transmission of viruses of edible aroids in the Solomon Islands and the Southwest Pacific. PANS 23, 171-177.
Jackson, G.V.H. (1980) Diseases and pests of taro. Noumea, New Caledonia: South Pacific Commission, 52 pp.
Jackson, G.V.H. (1978) Alomae and bobone diseases of taro. South Pacific Commission Advisory Leaflet 8. Noumea.
Jackson, G.V.H. and D.E. Gollifer. (1975) Disease and pest problems of taro (Colocasia esculenta L. Schott) in the British Solomon Islands. PANS 21, 45-53.
James, M. Kenten, R.H. and R.D. Woods. (1973) Virus-like particles associated with two diseases of Colocasia esculenta (L.) Schott in the British Solomon Islands. J Gen Virol 21, 145-153.
Lambden, P.R. Cooke, S.J. Caul, E.O. and I.N. Clarke. (1992) Cloning of non-cultivatable human rotavirus by single primer amplification. J. Virol 66, 1817-1822.
Maino, M.K. (2003) The development of a serological-based diagnostic test for Dasheen mosaic potyvirus (DsMV). MSc Thesis, School of Life Sciences, Queensland University of Technology.
Macanawai, A. (2003) The epidemiology of Taro bacilliform virus in Samoa. MSc thesis, USP, Samoa.
Pearson, M.N. Jackson, G.V.H. Saelea, J. and S.G. Morar. (1999) Evidence for two rhabdoviruses in taro (Colocasia esculenta) in the Pacific region. Australasian Plant Pathology 28, 248-253.
Rodoni, B.C. Dale, J.L. and R.M. Harding. (1994) Review of alomae disease of taro. Papua New Guinea Journal of Agriculture, Forestry and Fisheries 37, 14-18.
Sambrook J. Fritsch, E.F. and Maniatis, T. (1989). Molecular Cloning. A laboratory Manual. 2nd edition. Cold Spring Harbour Laboratory Press. USA.
Shaw, E.D. Plumb, R.T. and G.V.H. Jackson. (1979) Virus diseases of taro (Colocasia esculenta) and Xanthosoma spp. in Papua New Guinea. Papua New Guinea Agriculture Journal 30, 71-97.
25
Yang, I.C. Hafner, G.J. Dale, J.L. and R.M. Harding. (2003a) Genomic characterisation of Taro bacilliform virus. Arch Virol 148, 937-949.
Yang, I.C. Hafner, G.J. Revill, P.A. Dale, J.L. and R.M. Harding. (2003b) Sequence diversity of South Pacific isolates of Taro bacilliform virus and the development of a PCR-based diagnostic test. Arch Virol 148, 1957-1968.
Zettler, F.W. and R.D. Hartman. (1986) Dasheen mosaic virus and its control in cultivated aroids. Extension Bulletin, ASPAC Food Fertilizer Technology Center, Taiwan, (No. 233) 13 pp.
Zettler, F.W. and R.D. Hartman. (1987) Dasheen mosaic virus as a pathogen of cultivated aroids and control of the virus by tissue culture. Plant Dis 71, 958-963.
26
Appendix 1
Methods of indexing plant samples for taro viruses Nucleic acid extractions Nucleic acids were extracted from 100 mg of fresh plant tissue or 20 mg of dried tissue using QIAGEN DNeasy (for TaBV) or RNeasy (for DsMV, TaVCV, TaRV and CBDV) kits, according to the manufacturer’s protocol. Purified nucleic acids were stored at –800 until analysed by PCR. Different methods were used to detect the viruses because TaBV has a DNA genome, whereas the others have RNA genomes. Detection of DNA viruses – TaBV The viral DNA was amplified in 25 µl reactions containing 25 ρmol of each TaBV-specific primer (TSBV-1 and TSBV-4), 1 ρmol of each internal control primer (Control 1F and Control 1R), 20 mM Tris-HCl, pH8.4, 50 mM KCl, 1.5 mM MgCl2, 200 µM dNTPs and 1.25 units of PlatinumTM Taq polymerase (Invitrogen). All PCR reactions were set up on ice as follows. TSBV-1 (20 ρmol/µl) 0.5 µl TSBV-4 (20 ρmol/µl) 0.5 µl Control 1F (2 ρmol/µl) 0.5 µl Control 1R (2 ρmol/µl) 0.5 µl dNTPs 0.5 µl 10 X Invitrogen PCR buffer minus MgCl2 2.5 µl MgCl2 (50 mM) 0.75 µl PlatinumTM Taq 0.25 µl dH2O 18 µl DNA (1:20 dilution) 1 µl The reactions were thermal cycled on an MJ Research PTC-200 Peltier Thermal Cycler under the following conditions: 1 cycle of 940 C for 2 min 30 sec; 35 cycles of 940 C for 30 sec, 570 C for 30 sec, 720C for 1 min; and a final extension at 720C for 5 min. The products were analysed by electrophoresis through 2% TAE agarose gels, stained with ethidium bromide and viewed with a UV transilluminator. Detection of RNA viruses- CBDV, DsMV, TaRV and TaVCV All RNA viruses were detected by amplification with a Titan RT-PCR kit (Roche). All PCR reactions were set up on ice. The RNA was amplified in a 25µl reaction, which was set up as 2 separate master mixes according to the manufacture’s instructions. Master Mix 1 (MM1) contained 10 pmol of each virus-specific primer, 200 µM dNTPs, 10 mM DTT, 1 µl RNA and 8.75 µl of RNase-free dH2O. MM1 was denatured at 950C for 2
27
min, quenched on ice, then Master mix 2 (MM2) was added. MM2 consisted of 5 ul of 5 x RT-PCR buffer, 0.5 µl of Titan enzyme mix and 7 µl of RNase-free water. Following the addition of MM2, the reactions were mixed, pulse-centrifuged and placed in a pre-heated thermal cycler at 42OC. MM1 Virus-specific Primer 1 (20 ρmol/µl) 0.5 µl Virus-specific Primer 2 (20 ρmol/µl) 0.5 µl DTT (100 mM) 1.25 µl dNTPs (10 mM) 0.5 µl dH2O 8.75 µl RNA 1 µl 950C for 2 min, quench on ice and add ice-cold MM2. MM2 5 x Titan RT-PCR buffer 5 µl dH20 7 µl Enzyme mix 0.5 µl The PCR conditions differed slightly for each RNA virus detected, as follows: DsMV: 1 cycle of 420C for 35 min, 940C for 2 min; 35 cycles at 940C for 30 sec, 550C for 30 sec and 680C for 1 min; and 1 cycle of 680C for 5 min. TaVCV: 1 cycle of 420C for 35 min, 940C for 2 min; 35 cycles at 940C for 20 sec, 500C for 30 sec and 680C for 1 min; and 1 cycle of 680C for 10 min. CBDV:1 cycle of 420C for 35 min, 940C for 2 min; 35 cycles at 940C for 10 sec, 550C for 30 sec and 680C for 1 min; and 1 cycle of 680C for 10 min. TaRV: 1 cycle of 420C for 35 min, 940C for 2 min; 35 cycles at 940C for 20 sec, 550C for 30 sec and 680C for 1 min; and 1 cycle of 680C for 10 min. The products were analysed by electrophoresis through 2% TAE agarose gels, stained with ethidium bromide and viewed with a UV transilluminator