University of Connecticut University of Connecticut OpenCommons@UConn OpenCommons@UConn Honors Scholar Theses Honors Scholar Program Spring 5-20-2021 Development of Physiological Markers for Tetrabenazine-Induced Development of Physiological Markers for Tetrabenazine-Induced Motivational Dysfunctions Using Electroencephalography in Male Motivational Dysfunctions Using Electroencephalography in Male and Female Rats and Female Rats Stephanie Akosa [email protected]Follow this and additional works at: https://opencommons.uconn.edu/srhonors_theses Recommended Citation Recommended Citation Akosa, Stephanie, "Development of Physiological Markers for Tetrabenazine-Induced Motivational Dysfunctions Using Electroencephalography in Male and Female Rats" (2021). Honors Scholar Theses. 762. https://opencommons.uconn.edu/srhonors_theses/762
Transcript
University of Connecticut University of Connecticut
OpenCommons@UConn OpenCommons@UConn
Honors Scholar Theses Honors Scholar Program
Spring 5-20-2021
Development of Physiological Markers for Tetrabenazine-Induced Development of Physiological Markers for Tetrabenazine-Induced
Motivational Dysfunctions Using Electroencephalography in Male Motivational Dysfunctions Using Electroencephalography in Male
Follow this and additional works at: https://opencommons.uconn.edu/srhonors_theses
Recommended Citation Recommended Citation Akosa, Stephanie, "Development of Physiological Markers for Tetrabenazine-Induced Motivational Dysfunctions Using Electroencephalography in Male and Female Rats" (2021). Honors Scholar Theses. 762. https://opencommons.uconn.edu/srhonors_theses/762
Development of Physiological Markers for Tetrabenazine-
Induced Motivational Dysfunctions Using Electroencephalography in Male and Female Rats
Stephanie Akosa Dr. Salamone’s Lab
BS: Physiology and Neurobiology Minor: Psychological Sciences
University of Connecticut, Class of 2021
Abstract
Depression is a mental illness that is increasingly rampant in our society. With its
prevalence, various drugs have become commonplace for treatment. Many of these drugs are
serotonin reuptake inhibitors, or SSRIs. These drugs are able to mitigate symptoms of depression
such as rumination and anxiety. However, they are not very successful in treating the
amotivation and anergia that are seen in these patients. In order to investigate which drugs will
be the most successful in treating the symptoms of depression, it is important to develop an
animal model that can accurately represent the motivational symptoms of depression in humans.
The goal of the present study was to use female and male rats to develop electroencephalography
(EEG) markers that can be readily translatable to the pathophysiology of female and male
patients with depression. In this study, tetrabenazine (TBZ), a vesicular monoamine transporter-2
(VMAT-2) inhibitor, or a vehicle (VEH) control, was administered to a group of 8 female rats
and 7 male rats prior to measuring EEG in the home cage. Recordings were taken from each
treatment condition in the medial prefrontal cortex (mPFC), motor (M1/M2), and medial parietal
(mParietal) cortices to investigate the effect of TBZ on these cortices and to determine whether
they are comparable to the neurological markers associated with depression in humans. It was
found that male rats demonstrated a predominant peak frequency at baseline in EEG activity
across all three regions in the alpha/theta frequency range (4-12 Hz, with a sharp peak at 6-8.5
Hz frequency range). The sharp 6-8.5 Hz peak was suppressed when TBZ was administered in a
1.0 mg/kg dose. The EEG recordings of the female rats were not significantly affected. These
findings can be used as a foundation to develop further EEG studies in behaving rats that can be
translated to treatments for clinical depression within humans.
Introduction
Major depressive disorder affects a large portion of the population, with over 264 million
people being diagnosed with this across the globe (WHO, 2020). As stated by the National
Institutes of Health, depression is “a common but serious mood disorder. It causes severe
symptoms that affect how you feel, think, and handle daily activities” (Depression 2018). As the
number of individuals affected by depression has increased over the years, more people have had
to grapple with the cognitive, emotional, and physical symptoms that come with this
psychopathology (Stahl, 2006; Treadway and Zald, 2011; Fava et al. 2014; Salamone et al.
2016a).
One major cognitive impairment of depression is amotivation. Specifically, when given
assessments for effort-related decision making, humans with depression are less likely to choose
the option that requires more effort (Treadway et al. 2012a; Yang et al. 2014). In humans, this
lack of motivation can severely reduce the quality of one’s life, affecting one’s ability to work,
attend school, and maintain relationships (WHO 2020). To combat these symptoms and improve
quality of life, it is important to develop an understanding of how depression works and
determine which drug treatments are the most effective. This necessitates animal models for
research, which allow for the use of controlled settings as well as tools and methods that could
not be ethically used in humans (Hitzemann 2000). Through the use of animal models,
neurochemical changes that lead to effort-related motivation dysfunctions have been well-
established. For example, interference in the dopamine transmission of the ventral striatum
lowers participation in high-effort instrumental activities elicited by conditioned stimuli, whereas
low-effort instrumental activities are unaffected (Aberman and Salamone 1999; Salamone and
Correa 2012). Within our laboratory, we have used the effort-based choice model on rats to study
these motivational dysfunctions in psychopathology. We have conducted investigations in which
tetrabenazine (TBZ) was administered to rats. TBZ works by inhibiting the vesicular monoamine
transporter 2 (VMAT-2) to deplete dopamine, altering effort-based choice behavior in rats. In
comparison with administration of a vehicle control, TBZ reduced the number of lever presses,
while the consumption of concurrently available laboratory chow increased (Randall et al. 2014;
Yohn et al. 2016a). Thus, similarly to humans, rats demonstrate a low-effort bias when subjected
to the pharmacological conditions that have also shown to produce a low-effort bias in humans
(Salamone et al. 2016a,b,c; Yohn et al. 2016a,b,c). Moreover, administration of TBZ can induce
depressive symptoms in people, including fatigue and amotivation (Salamone et al. 2016a).
In terms of the neural basis of depression, there is evidence demonstrating that
mesolimbic DA circuits in the forebrain in conjunction with the ventral striatum, prefrontal
cortex, and ventral pallidum play a significant role in the selection of high-effort activities
(Salamone et al. 1991, 1994; 2007, 2016a,b, 2018; Walton et al. 2003; Floresco and Ghods-
Sharifi 2007; Mingote et al. 2008; Farrar et al. 2008, 2010; Winstanley and Floresco 2016). Still,
more research is needed to properly identify and describe the psychopathology of neural circuits
in effort-related dysfunctions.
Recent studies have suggested that cortical electroencephalographic markers may differ
between healthy and depressed individuals. There has been particular emphasis on the frontal
cortex, which is primarily responsible for cognitive effort tasks, or CET performance
(Winstanley and Floresco 2016). In a study of 37 depressed and 35 nondepressed individuals, the
frontal alpha EEG asymmetry of the depressed individuals at rest was significantly increased
(Gollan et al. 2014). Further, evidence also shows that depressed people have a reduction of EEG
markers in this cortex that are present during the performance of motivated behaviors and in the
anticipation of reinforcement (Nelson et al. 2018; Gheza et al. 2019). Specifically, it has been
determined that bilateral asymmetry of EEGs in the frontal cortex in humans is an important
marker for effort-related motivation (Allen, Coan, & Nazarian, 2004; Coan and Allen 2004).
In addition to EEGs, imaging has also been utilized to investigate the neural basis of
anergia, psychomotor retardation (Hickie et al. 1999; Capuron et al. 2007), and effort-related
decision making (Wardle et al. 2011; Schouppe et al. 2014; Huang et al. 2016; Hogan et al.
2018; Aridan et al. 2019). The results have been consistent with animal studies in that the
striatum and frontal cortex are important for effort-based decision making as well as approach
motivation, or motivation which is widely prompted by positive stimuli (Winstanley and
Floresco 2016; Harmon-Jones et al. 2013).
The purpose of the present experiments was to further investigate these findings,
comparing the characteristics of the mPFC, M1/M2, and mParietal EEG readings in the left and
right hemispheres of awake male and female rats in baseline and TBZ-induced, DA-depleted
conditions. The mPFC was the focus of this study due to the abundance of studies drawing
connections between executive function, effort-based choice, decision making, and the prefrontal
cortex. Additionally, the mPFC is thought to reflect the effects of TBZ on the ventral striatum
and striatopallidal circuits that are important for effort-based decision making (Rotolo Ph.D.
Dissertation, 2020). EEGs from the mParietal and M1/M2 cortices were recorded to serve as a
point of reference for the mPFC while simultaneously ascertaining whether TBZ affected other
cortical areas. In summation, this investigation was done to work towards creating physiological
markers for altered DA transmission and effort-related dysfunction in preclinical animal models
that are translatable to human male and female studies.
Materials and Methods
Surgery
Rats were anesthetized with 100 mg/kg of ketamine and 10 mg/kg of xylazine prior to being
placed within the stereotaxic device for electrode implantation surgery. Surgical implantation of
8 ¼” stainless-steel screw electrodes were performed on 8 untrained female rats and 7 untrained
male rats (total n=15). The EEG electrodes were placed next to the mPFC, M1/M2, and
mParietal in addition to two screws that served as ground and reference electrodes. Rats were
monitored for pupil reflex, knee-jerk reflex, and breathing throughout the surgery. Recordings
were carried out after a recovery period of 7 days.
Electrophysiological Recording Experiment
A Digital Lynx SX Electrophysiology System (Neuralynx) was used to record the EEG activity
of female and male rats in the awake state. Wide-band activity (1-2000 Hz, 4006 samples/sec)
was recorded using the Neuralynx system and was analyzed offline using MATLAB software
(MathWorks Inc, Natick, MA). Approximately 1 minute of baseline EEG activity was recorded
from each rat after a short period of reading stabilization following the connection of the rat to
the Neuralynx system via cable. The rats were then injected intraperitoneally with either vehicle
or TBZ (1.0 mg/kg). A lead time of 120 minutes elapsed before resuming recordings. The
following week, each rat’s treatment was counterbalanced with either vehicle or TBZ.
Data Preprocessing
Signal processing and related procedures were carried out with the FieldTrip software package
(Oostenveld et al. 2011; http://www.fieldtriptoolbox.org/) and custom scripts in MATLAB (The
MathWorks, Natick, MA). The following steps were used to preprocess the EEG data: 1) data
were mean subtracted; 2) data were divided into 1-s epochs for the purpose of removing the
epochs that contained artifacts; 3) the first epoch and the last epoch were removed; 4) data was
inspected to determine and remove the epochs that contained high-amplitude artifacts according
to Z-score threshold criterion.
Neurophysiological Analysis
Power spectral analysis was carried out by first segmenting the preprocessed EEG data into 1-s
epochs with 0.5-s overlap. We then used Welch’s overlapped averaged periodogram method
(Welch, 1967) with a Hamming window on each epoch to estimate the power spectral density
(PSD) over frequencies from 1 Hz to 100 Hz (0.5 Hz resolution).
Results
EEG recordings were taken on 8 untrained female rats and 7 untrained male rats from
their mPFC, M1/M2, and mParietal cortices via implanted EEG electrodes. A PSD was used to
determine the effect of 1.0 mg/kg TBZ versus VEH treatment on EEG activity in awake female
(n=8) (Figure 1) and male (n=7) rats (Figure 2). During baseline home cage EEG recording from
the mPFC, M1/M2, and mParietal cortex of both freely moving male and female groups, the
predominant peak in EEG activity in the lower frequency ranges in all three brain cortices is
distinctly in the alpha/theta frequency range (4-12 Hz, with a sharp peak at 6-8.5 Hz frequency
range). In order to correct for the baseline values, power of pre- and post-injection was
subtracted from the baseline. In other words, the change from pre-injection to post-injection of
TBZ or VEH was determined. The values were then analyzed by analysis of variance (ANOVA)
mixed effects model. Across all three brain cortices, there was a significant sex x drug (vehicle
versus TBZ) effect interaction (F(1,317) = 4.74, p < 0.05) (see Table 1). There was a sharp 6-8.5
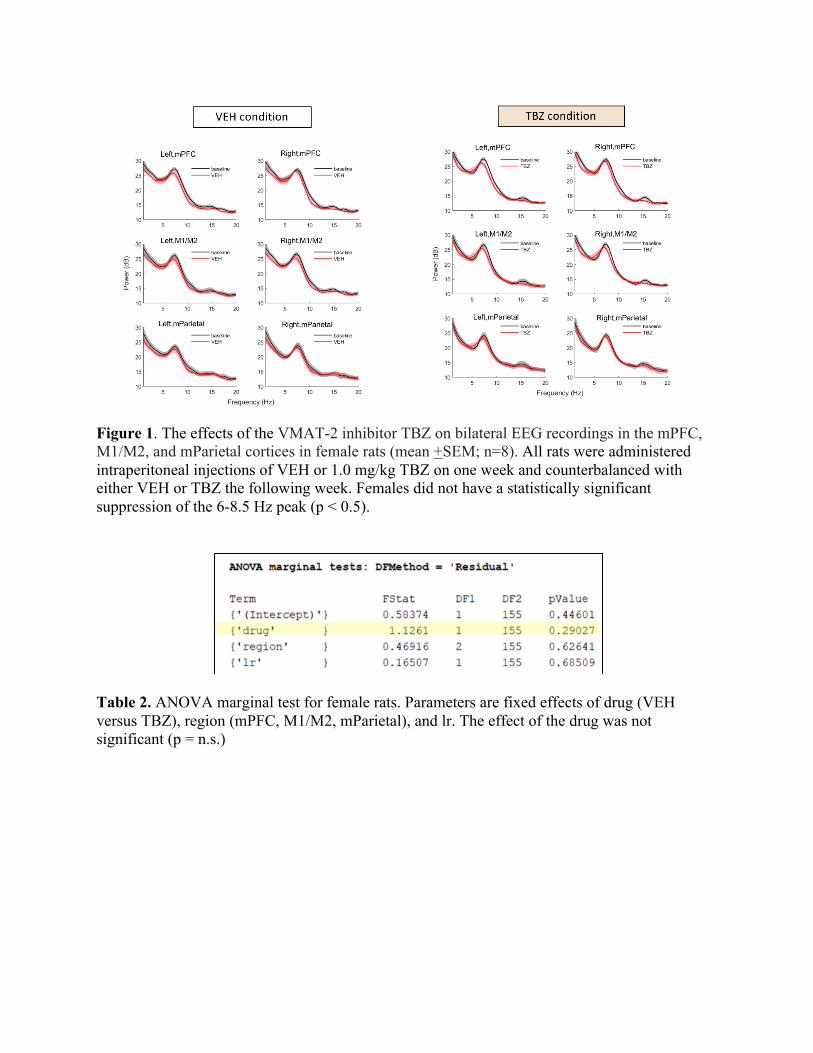
Hz peak that was not blunted in females (F(1,155) = 1.13, p = n.s.) (See Figure 1 and Table 2).
However, the power of this peak was significantly affected by the drug in male rats (F(1,159) =
5.01, p < 0.05) (see Figure 2 and Table 3). Thus, the effect of the drug was different in males
versus females. Specifically, this peak was significantly blunted relative to vehicles by
administration of 1.0 mg/kg TBZ in male rats, but not in females.
Table 1. ANOVA marginal test for fixed effects of sex (male versus female), drug (VEH versus TBZ), region (mPFC, M1/M2, mParietal), (left versus right) lr, and sex:drug (interaction of sex and drug). The effects of sex and drug approached significance (p = 0.08 < 0.5 ; p = 0.06 < 0.5). The interaction between sex and drug was significant, meaning the effect of the drug is different in males than it is in females (p = 0.03 < 0.05).
Figure 1. The effects of the VMAT-2 inhibitor TBZ on bilateral EEG recordings in the mPFC, M1/M2, and mParietal cortices in female rats (mean +SEM; n=8). All rats were administered intraperitoneal injections of VEH or 1.0 mg/kg TBZ on one week and counterbalanced with either VEH or TBZ the following week. Females did not have a statistically significant suppression of the 6-8.5 Hz peak (p < 0.5).
Table 2. ANOVA marginal test for female rats. Parameters are fixed effects of drug (VEH versus TBZ), region (mPFC, M1/M2, mParietal), and lr. The effect of the drug was not significant (p = n.s.)
Figure 2. The effects of the VMAT-2 inhibitor TBZ on bilateral EEG recordings in the mPFC, M1/M2, and mParietal cortices in male rats (mean +SEM; n=7). All rats were administered intraperitoneal injections of VEH or 1.0 mg/kg TBZ on one week and counterbalanced with either VEH or TBZ the following week. Males had a statistically significant suppression of the 6-8.5 Hz peak (p < 0.5).
Table 3. ANOVA marginal test for male rats. Parameters are fixed effects of drug (VEH versus TBZ), region (mPFC, M1/M2, mParietal), and lr. The effect of the drug was significant (p < 0.05).
Discussion
Our lab conducted these studies to determine the neurophysiological effects of TBZ on
untrained, freely moving male and female rats. This involved using EEG recordings that focused
on power spectral density analyses of signals from the bilateral mPFC, M1/M2, and mParietal
cortices. The use of animal models to study effort and motivation is valid as the data collected
from previous animal studies correlates to what is seen in patients with major depressive
disorder. This investigation is one the first in a series that will lead to the studies of the
electrophysiology of trained animals carrying out effort-related choice tasks. Performing
investigations in trained animals is important to allow for the discovery of more sensitive
electrophysiological biomarkers in males and females (Rotolo Ph.D. Dissertation, 2020). Thus,
the present studies are necessary in order to develop the initial EEG markers that are readily
translatable to humans.
In this investigation, 8 female rats and 7 male rats were each surgically implanted with
cortical EEG electrodes to record from the mPFC, M1/M2, and mParietal cortices. Based on
recordings post-injection of TBZ compared to baseline along with a vehicle control, statistically
significant results were found for male rats. The results demonstrated a predominant peak in
EEG activity at baseline in all three brain cortices clearly in the alpha/theta frequency range (4-
12 Hz, with a sharp peak at 6-8.5 Hz frequency range). Using the ANOVA mixed effects model,
the data shows that collapsed across all three brain regions, there was a significant sex x drug
interaction (F(1,317) = 4.74, p < 0.05) (see Table 1). This peak remained unaffected in female
rats (F(1,155) = 1.13, p = n.s.) (See Figure 1 and Table 2). In male rats, there was a significant
suppression of the sharp 6-8.5 Hz peak (F(1,159) = 5.01, p < 0.05) (See Figure 2 and Table 3). In
conclusion, the effect of the drug on the observed EEG power spectra was different in males
versus females. Specifically, the peak frequency band was significantly blunted relative to
vehicle by administration of 1.0 mg/kg TBZ in male rats, but not in females. This has potential
implications that DA depletion seen in depressed individuals does not affect the male and female
brain in the same manner. Additionally, a limitation of this investigation was that only one
dosage amount (1.0 mg/kg TBZ) was used. For future studies, administering rats various dosages
would allow for a deeper investigation of how the male and female brain are affected by different
degrees of DA depletion.
The findings of this investigation will also serve as a good foundation for future research
on how frontal EEG recordings change during the performance of effort-related tasks. During the
PROG lever pressing task, rats must press a lever a certain number of times to deliver favorable,
carbohydrate-rich pellets that act as reinforcers for this behavior. As time goes on, the ratio of
presses to released pellets progressively increases. During this task, it has been shown that with
an increase in the lever pressing ratio, rats reach a break point during which they stop lever
pressing and switch to consumption of the readily-accessible, but less preferable lab chow
(Cordony 2019). According to Pizzagalli et al. (2005), Nelson et al. (2018), and Gheza et al.
(2019), it is hypothesized that there will be alterations in frontal EEG activity when rats reach
their break point. It has also been hypothesized that the administration of TBZ will result in a
decrease in lever pressing and an increase in chow intake, which will also lead to changes in
frontal EEG activity. More knowledge of EEG activity and electrophysiological biomarkers
would allow for further investigation of neural circuitry via anterograde and retrograde tracing.
Given the anatomical connections between the ventral striatum and ventral pallidum via the
thalamus to the mPFC, and the role of this circuitry in effort-related decision making, it is
important to investigate how different neural pathways are affected by TBZ (Fareri et al. 2017).
Such findings would significantly deepen our understanding of the circuit mechanisms that are
responsible for effort-based aspects of motivation.
References
Aberman JE, Salamone JD (1999) Nucleus accumbens dopamine depletions make rats more sensitive to high ratio requirements but do not impair primary food reinforcement. Neuroscience 92(2):545-552. doi:10.1016/s0306-4522(99)00004-4.
Allen JJ, Coan JA, Nazarian M. (2004) Issues and assumptions on the road from raw signals to metrics of frontal EEG asymmetry in emotion. Biol Psychol 67(1-2):183-218. doi:10.1016/j.biopsycho.2004.03.007 .
Aridan N, Malecek NJ, Poldrack RA, Schonberg T (2019) Neural correlates of effort-based valuation with prospective choices. Neuroimage 185:446-454.
Capuron L, Pagnoni G, Demetrashvili MF, Lawson DH, Fornwalt FB, Woolwine B, Berns GS, Nemeroff CB, Miller AH (2007) Basal ganglia hypermetabolism and symptoms of fatigue during interferon - therapy. Neuropsychopharmacology 32: 2384-2392.
Cordony, M.B.L., Chow, J.Y.L. & Boakes, R.A. Motivation to run measured by progressive ratio tests: Failure to support the addiction hypothesis for rats. Learn Behav 47: 131–140 (2019).
Salamone JD and Correa M (2012) The mysterious motivational functions of mesolimbic dopamine. Neuron 76:470–85.
Demyttenaere K, De Fruyt J, Stahl SM (2005) The many faces of fatigue in major depressive disorder. Int J Neuropsychopharmacology 8, 93-105 doi: 10.1017/S1461145704004729.
Depression (2020) World Health Organization. https://www.who.int/news-room/fact-sheets/detail/depression.
Fareri, D. S., Gabard-Durnam, L., Goff, B., Flannery, J., Gee, D. G., Lumian, D. S., Caldera, C., & Tottenham, N. (2017). Altered ventral striatal-medial prefrontal cortex resting-state connectivity mediates adolescent social problems after early institutional care. Development and psychopathology, 29(5): 1865–1876.
Farrar AM, Font L, Pereira M, Mingote S, Bunce JG, Chrobak JJ, Salamone JD (2008) Forebrain circuitry involved in effort-related choice: Injections of the GABAA agonist muscimol into ventral pallidum alter response allocation in food-seeking behavior. Neuroscience 152:321-330.
Farrar AM, Segovia KN, Randall PA, Nunes EJ, Collins LE, Stopper CM, Port RG, Hockemeyer J, Müller CE, Correa M, Salamone JD (2010) Nucleus accumbens and effort-related functions: behavioral and neural markers of the interactions between adenosine A2A and dopamine D2 receptors. Neuroscience 166: 1056-1067.
Fava M, Ball S, Nelson JC, Sparks J, Konechnik T, Classi P, et al. (2014) Clinical relevance of fatigue as a residual symptom in major depressive disorder. [Review] Depress Anxiety; 31: 250–7.
Gheza D, Bakic J, Baeken C, De Raedt R, Pourtois G (2019) Abnormal approach-related motivation but spared reinforcement learning in MDD: Evidence from fronto-midline Theta oscillations and frontal Alpha asymmetry. Cogn Affect Behav Neurosci. 2019 Jun;19(3):759-777.
Harmon-Jones, E., Harmon-Jones, C., & Price, T. F. (2013). What is Approach Motivation? Emotion Review, 5(3), 291–295.
Hickie I, Ward P, Scott E, Haindl W, Walker B, Dixon J, Turner K (1999) Neo-striatal rCBF correlates of psychomotor slowing in patients with major depression. Psychiatry Res 92: 75-81.
Hitzemann R. (2000). Animal models of psychiatric disorders and their relevance to alcoholism. Alcohol research & health : the journal of the National Institute on Alcohol Abuse and Alcoholism, 24(3), 149–158.
Hogan PS, Galaro JK, Chib VS (2018) Roles of Ventromedial Prefrontal Cortex and Anterior Cingulate in Subjective Valuation of Prospective Effort. Cereb Cortex [Epub ahead of print].
Huang J, Yang XH, Lan Y, Zhu CY, Liu XQ, Wang YF, Cheung EF, Xie GR, Chan RC (2016) Neural substrates of the impaired effort expenditure decision making in schizophrenia. Neuropsychology 30(6):685-696.
Jackie K. Gollan, Denada Hoxha, Dietta Chihade, Mark E. Pflieger, Laina Rosebrock, John Cacioppo (2014) Frontal alpha EEG asymmetry before and after behavioral activation treatment for depression. Biological Psychology 99;198-208.
Mingote S, Font L, Farrar AM, Vontell R, Worden LT, Stopper CM, Port RG, Sink KS, Bunce JG, Chrobak JJ, Salamone JD (2008) Nucleus accumbens adenosine A2A receptors regulate exertion of effort by acting on the ventral striatopallidal pathway. J Neurosci 28:9037-9046.
Nelson BD, Kessel EM, Klein DN, Shankman SA (2018) Depression symptom dimensions and asymmetrical frontal cortical activity while anticipating reward. Psychophysiology 55(1).
Pizzagalli DA, Sherwood RJ, Henriques JB, Davidson RJ (2005) Frontal brain asymmetry and reward responsiveness: A source-localization study. Psychological Science, 16(10):805– 813.
Randall, P. A., Lee, C. A., Nunes, E. J., Yohn, S. E., Nowak, V., Khan, B., Shah, P., Pandit, S., Vemuri, V. K., Makriyannis, A., Baqi, Y., Müller, C. E., Correa, M., & Salamone, J. D. (2014). The VMAT-2 inhibitor tetrabenazine affects effort-related decision making in a progressive ratio/chow feeding choice task: reversal with antidepressant drugs. PloS one, 9(6): e99320.
Robert Oostenveld, Pascal Fries, Eric Maris, and Jan-Mathijs Schoffelen. FieldTrip: Open Source Software for Advanced Analysis of MEG, EEG, and Invasive Electrophysiological Data. Computational Intelligence and Neuroscience, vol. 2011, Article ID 156869, 9 pages, 2011. doi:10.1155/2011/156869.
Rotolo, Renee A. (2020) An Investigation of Motivational Dysfunctions in a Rat Model of Effort-Related Choice Behavior: Behavioral and Neurochemical Evidence for Possible Novel Pharmacological Treatments. Doctoral Dissertations. 2575.
Salamone JD, Correa M, Farrar A, Mingote SM (2007) Effort-related functions of nucleus accumbens dopamine and associated forebrain circuits. Psychopharmacology (Berl) 191:461-482.
Salamone JD, Correa M, Ferrigno S, Yang JH, Rotolo RA, Presby RE (2018) The Psychopharmacology of Effort-Related Decision Making: Dopamine, Adenosine, and Insights into the Neurochemistry of Motivation. Pharmacol Rev 70(4):747-762.
Salamone JD, Correa M, Yohn S, Lopez-Cruz L, San Miguel N, and Alatorre L (2016b) The pharmacology of effort-related choice behavior: dopamine, depression, and individual differences. Behav Processes 127, 3-17. doi: 10.1016/j.beproc.2016.02.008.
Salamone JD, Cousins MS, Bucher S (1994) Anhedonia or anergia? Effects of haloperidol and nucleus accumbens dopamine depletion on instrumental response selection in a T-maze cost/benefit procedure. Behav Brain Res 65:221-229.
Salamone JD, Steinpreis RE, McCullough LD, Smith P, Grebel D, Mahan K (1991) Haloperidol and nucleus accumbens dopamine depletion suppress lever pressing for food but increase free food consumption in a novel food choice procedure. Psychopharmacology (Berl) 104(4):515‐521. doi:10.1007/BF02245659.
Salamone JD, Yohn SE, Lopez-Cruz L, San Miguel N, Correa M (2016a) Activational aspects of motivation: neural mechanisms and implications for psychopathology. Brain 139: 1325- 1347.
Schouppe N, Demanet J, Boehler CN, Ridderinkhof KR, Notebaert W (2014) The role of the striatum in effort-based decision-making in the absence of reward. J Neurosci 34(6):2148-2154.
Treadway MT, Bossaller NA, Shelton RC, and Zald DH (2012) Effort-based decision-making in major depressive disorder: a translational model of motivational anhedonia. J Abnorm Psychol 121(3), 553-558.
Treadway MT, Zald DH (2011) Reconsidering anhedonia in depression: lessons from translational neuroscience. Neurosci Biobehav Rev 35: 537–55.
Yang XH, Huang J, Zhu CY, Wang YF, Cheung EF, Chan RC, Xie GR (2014) Motivational deficits in effort-based decision making in individuals with subsyndromal depression, first-episode and remitted depression patients. Psychiatry Res 220(3), 874-882. doi: 10.1016/j.psychres.2014.08.056.
Yohn SE, Collins SL, Contreras-Mora HM, Errante EL, Rowland MA, Correa M, Salamone JD (2016a) Not all antidepressants are created equal: differential effects of monoamine uptake inhibitors on effort-related choice behavior. Neuropsychopharmacology 41(3), 686-694. doi: 10.1038/npp.2015.188.
Yohn SE, Lopez-Cruz L, Hutson PH, Correa M, and Salamone JD (2016b) Effects of lisdexamfetamine and s-citalopram, alone and in combination, on effort-related choice behavior in the rat. Psychopharmacology 233(6), 949-960. doi: 10.1007/s00213-015- 4176-7.
Yohn SE, Yumna A, Haley A, Tripodi G, Baqi Y, Muller CE, San Miguel N, Correa M, Salamone JD (2016c) Effort-related motivational effects of the pro-inflammatory cytokine interleukin-6: pharmacological and neurochemical characterization. Psychopharmacology.
Walton ME, Bannerman DM, Alterescu K, Rushworth MF (2003) Functional specialization within medial frontal cortex of the anterior cingulate for evaluating effort-related decisions. J Neurosci 23(16):6475-6479. doi:10.1523/JNEUROSCI.23-16-06475.2003.
Wardle MC, Treadway MT, Mayo LM, Zald DH, de Wit H (2011) Amping up effort: effects of d-amphetamine on human effort-based decision-making. J Neurosci 31(46): 16597-16602.
Welch, Peter D. “The Use of Fast Fourier Transform for the Estimation of Power Spectra: A Method Based on Time Averaging Over Short, Modified Periodograms.” IEEE® Transactions on Audio and Electroacoustics. Vol. AU-15, 1967, pp. 70–73
Winstanley CA, Floresco SB (2016) Deciphering Decision Making: Variation in Animal Models of Effort- and Uncertainty-Based Choice Reveals Distinct Neural Circuitries Underlying Core Cognitive Processes. J Neurosci 36(48):12069-12079.