Progress in Neuro-Psychopharmacology & Biological Psychiatry 46 (2013) 1–12
Contents lists available at ScienceDirect
Progress in Neuro-Psychopharmacology & BiologicalPsychiatry
j ourna l homepage: www.e lsev ie r .com/ locate /pnp
Different effects of the NMDA receptor antagonists ketamine, MK-801,and memantine on postsynaptic density transcripts and theirtopography: Role of Homer signaling, and implications for novelantipsychotic and pro-cognitive targets in psychosis
Andrea de Bartolomeis a,⁎, Chiara Sarappa a, Elisabetta F. Buonaguro a, Federica Marmo a, Anna Eramo b,Carmine Tomasetti a, Felice Iasevoli a
a Laboratory of Molecular and Translational Psychiatry, Department of Neuroscience, University School of Medicine “Federico II”, Naples, Italyb Medical Affairs & Phase IV Clinical Affairs, Lundbeck Pharmaceutical Services LLC, Deerfield, IL, United States
Abbreviations: ACC, anterior cingulate cortex; ANOVA, oactivity-related cytoplasmic protein; BDNF, brain-derived nthe nucleus accumbens; DEPC, diethylpyrocarbonate; dlCPdmCP, dorsomedial caudate putamen; EDTA, ethylene-diheat shock protein of 70 kDa; IC, insular cortex; MAC, medcortex; mGluR5, type 5 metabotropic glutamate receptorsPBS, phosphate buffered saline; PSD, postsynaptic density;standard error mean; shell, shell of the nucleus accumbenssaline sodium citrate solution; TdT, terminal deoxytransfeputamen; vmCP, ventromedial caudate putamen.⁎ Corresponding author at: Department of Neuroscience,
Napoli “Federico II”, Via Pansini, 5, Edificio n.18 3rd floor, 807463673 (office), +39 081 7463884 (lab), +39 3662747462378.
Administration of NMDA receptor antagonists, such as ketamine andMK-801, may induce psychotic-like behaviorsin preclinical models of schizophrenia. Ketamine has also been observed to exacerbate psychotic symptoms inschizophrenia patients. However, memantine, a non-competitive NMDA receptor antagonist approved forAlzheimer's disease and proposed for antipsychotic augmentation, may challenge this view. To date, the molecularmechanisms by which these NMDA receptor antagonists cause different neurochemical, behavioral, and clinicaleffects are still amatter of debate. Here,we investigated bymolecular imagingwhether these agents could different-ly modulate gene expression and topographical distribution of glutamatergic postsynaptic density (PSD) proteins.We focused on Homer1a/Homer1b/PSD-95 signaling network, which may be implicated in glutamate-dependentsynaptic plasticity, as well as in psychosis pathophysiology and treatment.Ketamine (25 and 50 mg/kg) and MK-801 (0.8 mg/kg) significantly induced the transcripts of immediate-earlygenes (Arc, c-fos, and Homer1a) in cortical regions compared to vehicle, whereas they reduced Homer1b andPSD-95 expression in cortical and striatal regions. Differently,memantine (5 mg/kg) did not increaseHomer1a signalcompared to vehicle, whereas it induced c-fos in the somatosensory and in themedial agranular cortices. Moreover,memantine did not affect Homer1b and PSD-95 expression. When compared to ketamine andMK-801, memantinesignificantly increased the expression of c-fos, Homer1b and PSD-95. Overall, ketamine and MK-801 prominentlyincreased Homer1a/Homer1b expression ratio, whereas memantine elicited the opposite effect.These data may support the view that ketamine, MK-801 and memantine exert divergent effects on PSDtranscripts, which may contribute to their partially different behavioral and clinical effects.
University School of Medicine of131, Napoli, Italy. Tel.: +39 0815592 (mobile); fax: +39 081
is).
rights reserved.
1. Introduction
A growing body of evidence implies dysfunctions of glutamatergicneurotransmission in psychosis pathophysiology (Fallgatter et al., 2010;Kantrowitz and Javitt, 2012; Timms et al., 2013). Perturbation of NMDAreceptor-mediated signaling by administration of NMDA receptorantagonists, such as ketamine, has been described to cause psychoticsymptoms in humans and to elicit psychotic-like behaviors in animals(Coyle et al., 2012; de Oliveira et al., 2011; Javitt et al., 2012; Kantrowitzand Javitt, 2010; Morgan et al., 2004; Umbricht et al., 2000). The acuteand chronic administration of ketamine or MK-801, the latter beinganother NMDA receptor antagonist of preclinical use only, has beenregarded as a valuable animal model of psychosis (Iasevoli et al., 2012a;Lipska andWeinberger, 2000; Moghaddam and Krystal, 2012). However,despite sharing similar pharmacological properties, not all NMDA recep-tor antagonists display psychotomimetic propensity.
2 A. de Bartolomeis et al. / Progress in Neuro-Psychopharmacology & Biological Psychiatry 46 (2013) 1–12
Memantine is a non-competitive NMDA receptor antagonist, whichshows moderate affinity for the Mg+ receptor site, channel voltage- anduse-dependency, and rapid unblocking kinetics (Danysz and Parsons,2012). Moreover, preclinical studies have demonstrated that memantineat high concentrations targets a number of receptors and neurotransmit-ter systems, including serotonin receptors, nicotinic acetylcholinereceptors, sigma-1 receptors and serotonin and dopamine uptake (for areview, see: Johnson and Kotermanski, 2006).
In mice, memantine has been found to dose-dependently disruptprepulse inhibition (PPI) of the acoustic startle response (ASR), abehavioral measure that resembles sensorimotor gating deficits ofschizophrenia patients (Nakaya et al., 2011). Nevertheless, adminis-tration of memantine (10 or 15 mg/kg) has been also described tosignificantly restore schizophrenia-like symptoms in a ketamine-induced social withdrawal model in rats (Uribe et al., 2013).
Memantine has been reported to prevent NMDA receptor-dependentexcitotoxicity and to promote neurogenesis in cortical and hippocampalneurons (Maekawa et al., 2009; Wei et al., 2012), while ketamine andMK-801 in animal studies dose-dependently impair cognitive functions(Neill et al., 2010) and cause neurotoxic damage (Liu et al., 2011). In pre-clinical studies, memantine facilitated memory consolidation (Samartgiset al., 2012) and rescued neurochemical abnormalities, as well asbehavioral and cognitive deficits in animal models of neurodegeneration(Borre et al., 2012;Wenk et al., 1997; Zajaczkowski et al., 1996). Howev-er, contrasting evidence exists on these issues. High dose memantine(i.e. 20 mg/kg) may impair, rather than ameliorate, cognitive func-tioning (Misztal and Danysz, 1995; Reus et al., 2008). Low-dosememantine (i.e. 5 mg/kg) has been observed to disrupt memoryand locomotor behavior in adult rats (Creeley et al., 2006); howeverthe same dose has been described to enhance long-term spatialmemory and cognitive functioning in several similar paradigms(Ihalainen et al., 2011; Reus et al., 2008; Zoladz et al., 2006).
Memantine is indicated for the treatment of patients withmoderate-to-severe Alzheimer's disease (Wilkinson, 2012). More-over, it has been reported that memantine administration to patientswith moderate to severe Alzheimer's disease receiving stable doses ofdonepezil resulted in better outcomes than placebo on measures ofcognition, behavior and global outcome (Tariot et al., 2004). Thecompound has also been proposed as an add-on and a pro-cognitiveagent in treatment-resistant psychiatric disorders (de Bartolomeiset al., 2012a; Koukopoulos et al., 2010; Sani et al., 2012; Zdanys andTampi, 2008).
However, the molecular mechanisms by which different NMDAreceptor antagonists induce different neurochemical, behavioral andclinical effects are yet to be completely elucidated. Here, we aimedat verifying the hypothesis that ketamine, MK-801 and memantinecould differently modulate the expression of PSD transcripts implicat-ed in glutamatergic signaling and synaptic plasticity.
PSD is a complex protein mesh located at the dendritic spines ofpost-glutamatergic neurons, whose core components are representedby NMDA receptors (de Bartolomeis et al., 2005). PSDmolecules, suchas the inducible and the constitutive Homer1 isoforms (Homer1a andHomer1b, respectively), Arc, or PSD-95, participate in postsynapticglutamatergic signaling and contribute to the cross-talk betweenthe glutamatergic and the dopaminergic system (de Bartolomeisand Tomasetti, 2012; de Bartolomeis et al., 2013a). Moreover, PSDproteins have been implicated in the pathophysiology of severalneuropsychiatric disorders. Specifically, beta-amyloid accumulationdisrupts PSD-clusters in Alzheimer's disease (Roselli et al., 2009);alterations involving PSD molecules have been described inschizophrenic patients (Hahn et al., 2006; Iasevoli et al., 2013; Toroand Deakin, 2005); gene expression of PSD molecules is significantlyaffected by acute and chronic antipsychotic agents (de Bartolomeiset al., 2013b; Iasevoli et al., 2010, 2011; Tomasetti et al., 2011).
Initial evidence has also been provided that NMDA receptorantagonists affect PSD molecules. MK-801 attenuated the increased
PSD-95 expression in the spinal cord of morphine-tolerant rats(Huang et al., 2012). Ketamine induced Homer1a expression in ratstriatum (Iasevoli et al., 2007) and increased Arc and PSD-95 proteinlevels in rat prefrontal cortex (Li et al., 2010).
It has been proposed that the ratio of Homer1a/Homer1b levels mayimpact the downstream signaling bymGluR5 and affect cognitive perfor-mances (Kammermeier, 2008;Menard andQuirion, 2012). Homer1b andPSD-95 interact to promote dendritic spine growth and enlargement(Sala et al., 2005). Homer1a has been shown to alter mGluR5 signalingin a model of fragile X mental retardation (Ronesi et al., 2012), to exerta negative effect on synapses growth (Sala et al., 2003), and to impaircognitive functioning (Klugmann et al., 2005; Lominac et al., 2005).
Based on these observations, in the present study we investigatedwhether: i) ketamine and MK-801 might impair the Homer1b/PSD-95-mediated signaling, by decreasing their expression and/or increasing theHomer1a/Homer1b ratio; ii) memantine would have different effect onthe expression of the above-mentioned PSD transcripts; iii) thesemolecular changes induced byNMDA receptor antagonistsmay different-ly occur in cortical and striatal regions.
We also evaluated Arc expression, since this gene has been founddisrupted in neuropsychiatric diseases and has been implicated inglutamate-dependent plasticity (Beique et al., 2011; Rudinskiy et al.,2012). Finally, the immediate-early gene c-fos was examined as anoverall marker of neuronal activation (Kovacs, 2008).
2. Materials and methods
2.1. Animals
Male Sprague–Dawley rats sixty days of age (mean weight 250 g ±10 g) were obtained from Charles-River Labs (Lecco, Italy). The animalswere housed and let to adapt to human handling in a temperature(22 °C) and humidity controlled colony room with 12/12 h light–darkcycle (lights on from 6:00 a.m. to 6:00 p.m.) with ad libitum access tolaboratory chow and water. All procedures were conducted in accor-dance with the NIH Guide for Care and Use of Laboratory Animals(NIH Publication No. 85-23, revised 1996) and were approved by localAnimal Care and Use Committee. All efforts weremade tominimize an-imal number and suffering.
2.2. Drug treatment
Memantine hydrochloride powder (gently supplied by H. LundbeckA/S, Copenhagen, Denmark), ketamine hydrochloride powder and(+)-MK-801 hydrogen maleate powder (Sigma-Aldrich, St. Louis, MO,USA) were all dissolved in saline solution (NaCl 0.9%). All solutionswere adjusted to physiological pH value and injected i.p. at a finalvolume of 1 ml/kg.
Rats were randomly assigned to one of the following treatmentgroups (n = 5 for each treatment group): vehicle (NaCl 0.9%, VEH);ketamine 25 mg/kg (KET25); ketamine 50 mg/kg (KET50); MK-8010.8 mg/kg (MK-801); or memantine 5 mg/kg (MEM).
All drugs were given at behaviorally active doses. Memantine dose isconsistent with a dose giving therapeutically relevant plasma levels andsignificant NMDA receptor occupancy, and that has been found not toimpair learning and to be neuroprotective (Camarasa et al., 2010;Canever et al., 2010; More et al., 2008). Ketamine has been given at lowor intermediate subanesthetic doses, which are considered to providean animal model of psychosis (Lipska and Weinberger, 2000). However,these two doses have been demonstrated to cause divergent neurochem-ical and behavioral effects (Iasevoli et al., 2012b;Moghaddamet al., 1997;Verma and Moghaddam, 1996). In this study, we administered either alow or an intermediate subanesthetic dose to investigate whetherreported discrepancy might be also observed in the paradigm studied.MK-801has beengiven at a dose known to causepsychotic-like behaviors(Andine et al., 1999; Pinault, 2008).
Fig. 1. Regions of interest (ROIs) for mRNA expression quantitation. Here aredepicted the ROIs where mRNA expression has been assessed. dmCP: dorsomedialcaudate putamen. dlCP: dorsolateral caudate putamen. vmCP: ventromedial caudateputamen. vlCP: ventrolateral caudate putamen. core: core of the nucleus accumbens.shell: shell of the nucleus accumbens. ACC: anterior cingulate cortex. MAC: medialagranular cortex. MC: motor cortex. SS: somatosensory cortex. IC: insular cortex.
3A. de Bartolomeis et al. / Progress in Neuro-Psychopharmacology & Biological Psychiatry 46 (2013) 1–12
Animals were sacrificed by decapitation 90 min after administra-tion, the brains were rapidly removed, quickly frozen on powdereddry ice and stored at −70 °C prior to sectioning. Serial coronalsections of 12 μm were cut on a cryostat at −18 °C through the fore-brain at the level of the middle-rostral striatum (approx. from Bregma1.20 mm to 1.00 mm), using the rat brain atlas by Paxinos andWatson (1997) as an anatomical reference. Care was taken to selectidentical anatomical levels of treated and control sections usingthionin-stained brain sections. Sections were thaw-mounted ontogelatin-coated slides, and stored at −70 °C for subsequent analysis.
2.3. Probes
Probes used for radioactive in situ hybridization were oligodeoxy-ribonucleotides complementary to base sequence of target genemRNAs. The Homer1a probe is complementary to bases 2527–2574(GenBank #U92079; MWG Biotech, Firenze). The Arc probe iscomplementary to bases 789–833 (GenBank #NM019361; MWGBiotech, Firenze). The c-fos probe is complementary to bases 111–158 (GenBank #AY780203; MWG Biotech, Firenze). The Homer1bprobe was a 48-base oligodeoxyribonucleotide complementary tobases 1306–1354 of the rat Homer1b/c mRNA (GenBank # AF093267;MWG Biotech; Firenze, Italy). The PSD-95 probe was a 45-baseoligodeoxyribonucleotide complementary to bases 225–269 of the ratPSD-95 mRNA (GenBank # M96853; MWG Biotech; Firenze, Italy). Allprobes were designed from GenBank sequences and checked withBLAST in order to avoid cross-hybridization.
2.4. Probe radiolabeling
For each probe a 50 μl labeling reactionmixwas prepared on ice usingDEPC treated water, 1× tailing buffer, 7.5 pmol/μl of oligo, 125 units ofTdT and 100 mCi 35S-dATP. The mix was incubated 20 min at 37 °C.The unincorporated nucleotides were separated from radiolabeled DNAusing ProbeQuant G-50 Micro Columns (Amersham-GE HealthcareBiosciences; Milano, Italy). As an assessment of the probe specificity, theautoradiographic signal distribution was compared and found to beconsistent with previous in situ hybridization studies (Iasevoli et al.,2009). The specificity of each probe was also tested by pilot controlexperiment using the corresponding sense oligodeoxyribonucleotide.
2.5. In situ hybridization
Sections were processed for radioactive in situ hybridizationaccording to previously published protocols (Ambesi-Impiombatoet al., 2003). All solutions were prepared with sterile double-distilled water. The sections were fixed in 4% formaldehyde in0.12 M PBS (pH 7.4), quickly rinsed three times with 1× PBS, andplaced in 0.25% acetic anhydride in 0.1 M triethanolamine/0.9%NaCl, pH 8.0, for 10 min. Next, the sections were dehydrated in 70%,80%, 95% and 100% ethanol, delipidated in chloroform for 5 min,rinsed again in 100% and 95% ethanol and air-dried.
Sections were hybridized with 0.4–0.6 × 106 cpm of radiolabeledoligonucleotide in buffer containing 50% formamide, 600 mM NaCl,80 mM Tris–HCl (pH 7.5), 4 mM EDTA, 0.1% pyrophosphate,0.2 mg/ml heparin sulfate, and 10% dextran sulfate. Slides were cov-ered with coverslips and incubated at 37 °C in a humid chamber for22–24 h. After hybridization the coverslips were removed in 1× SSCand the sections were washed 4 × 15 min in 2× SSC/50% formamideat 43–44 °C, followed by two 30 min washes with 1× SSC at roomtemperature. The slides were rapidly rinsed in distilled water andthen in 70% ethanol.
The sectionswere dried and exposed toKodak-BiomaxMRAutoradio-graphic film (Sigma-Aldrich, Milano, Italy). A slide containing a scale of16 known amounts of 14C standards (ARC-146C, American RadiolabeledChemical, Inc., St. Louis, MO, USA) was co-exposed with the samples.
The autoradiographic films were exposed in a time range of 14–30 days. The optimal time of exposure was chosen to maximizesignal-to-noise ratio and to prevent optical density from approachingthe limits of saturation. Film development protocol included a 1.5 mindip in the developer solution and 3 min in the fixer.
2.6. Image analysis
The quantitation of the autoradiographic signal was performedusing a computerized image analysis system including: a transparen-cy film scanner (Microtek Europe B. V., Rotterdam, The Netherlands),an Apple PowerPC G4, and ImageJ software (v. 1.46v, Rasband, W.S.,http://rsb.info.nih.gov/ij/). The original characteristics of the scannedimages (i.e. contrast, brightness, resolution) were preserved. Eachslide contained 3 adjacent sections of a single animal. All hybridizedsections used for comparative statistical analysis were exposed onthe same sheet of X-ray film. Signal intensity analysis was carriedout on digitized autoradiograms measuring mean optical densitywithin outlined ROIs in correspondence of the cortex, caudate puta-men and nucleus accumbens (Fig. 1). An oval template, proportionalto the dimensions of the anatomical subregion, was used for comput-erized quantitations in each one of the ROIs depicted.
Sections were quantitated blind to the treatment conditions. Inorder to test for inter-observer reliability, an independent quantita-tion was performed by a second investigator. Results obtained bythe first investigator were considered reliable, and then reported,only when they were quantitatively comparable, in terms of consis-tency of the statistically significant effects found, to that obtained bythe second investigator.
2.7. Data processing
Measurements of mean optical density within ROIs were convertedusing a calibration curve based on the standard scale co-exposed to the
4 A. de Bartolomeis et al. / Progress in Neuro-Psychopharmacology & Biological Psychiatry 46 (2013) 1–12
sections. 14C standard values from 4 through 12 were previouslycross-calibrated to 35S brain paste standards, in order to assign adpm/mg tissue wet weight value to each optical density measure-ment through a calibration curve. For this purpose a “best fit” 3rddegree polynomial was used. For each animal, measurements fromthe 3 adjacent sections were averaged and the final data werereported in relative dpm as mean ± S.E.M. ANOVA was used toanalyze treatment effects. The Tukey's post hoc test was used todetermine the locus of effects in any significant ANOVA.
To calculate the Homer1a/Homer1b ratio we first divided the meanvalue of Homer1a and Homer1b mRNA expression to the correspon-dent value of vehicle mRNA expression in each region. Thenormalized Homer1a value of expression was then divided fornormalized Homer1b value of expression. A result N1 indicated prom-inent Homer1a over Homer1b expression; a result b1 indicated prom-inent Homer1b over Homer1a expression.
Signal distribution was assessed considering the measurementsfrom each treatment group as the dependent variable, and the ROIsin which expression was measured as the independent variable(i.e.: measurements were analyzed per region effect).
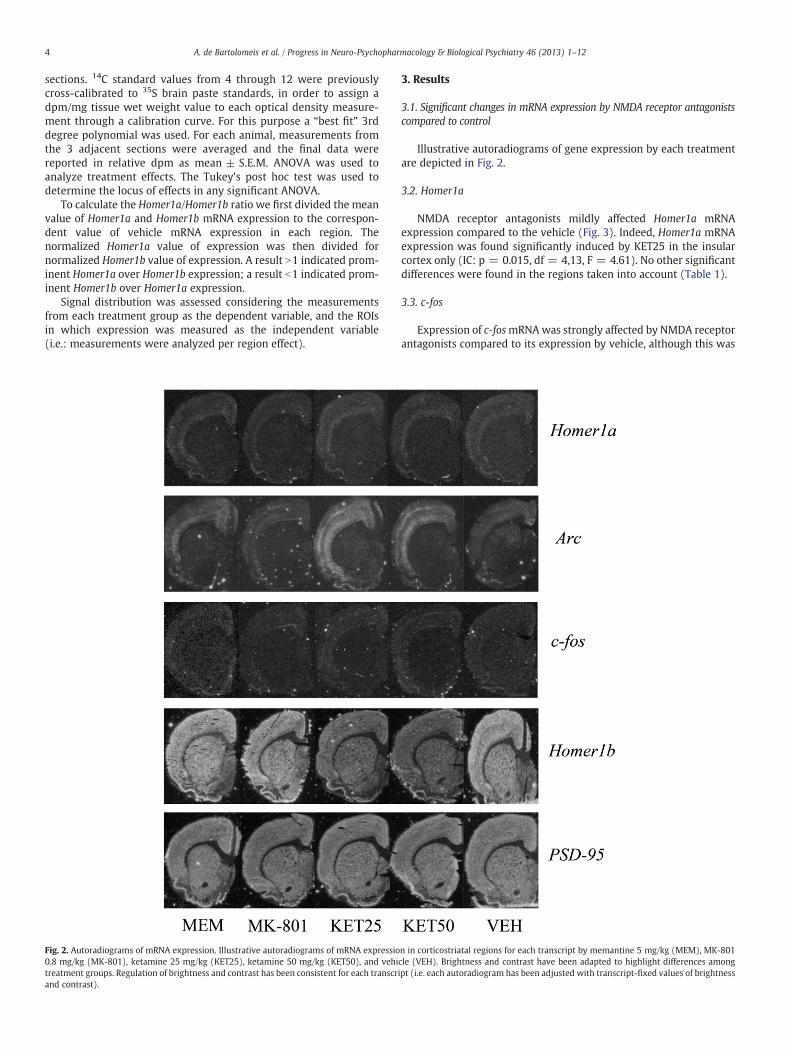
Fig. 2. Autoradiograms of mRNA expression. Illustrative autoradiograms of mRNA expressio0.8 mg/kg (MK-801), ketamine 25 mg/kg (KET25), ketamine 50 mg/kg (KET50), and vehitreatment groups. Regulation of brightness and contrast has been consistent for each transcrand contrast).
3. Results
3.1. Significant changes in mRNA expression by NMDA receptor antagonistscompared to control
Illustrative autoradiograms of gene expression by each treatmentare depicted in Fig. 2.
3.2. Homer1a
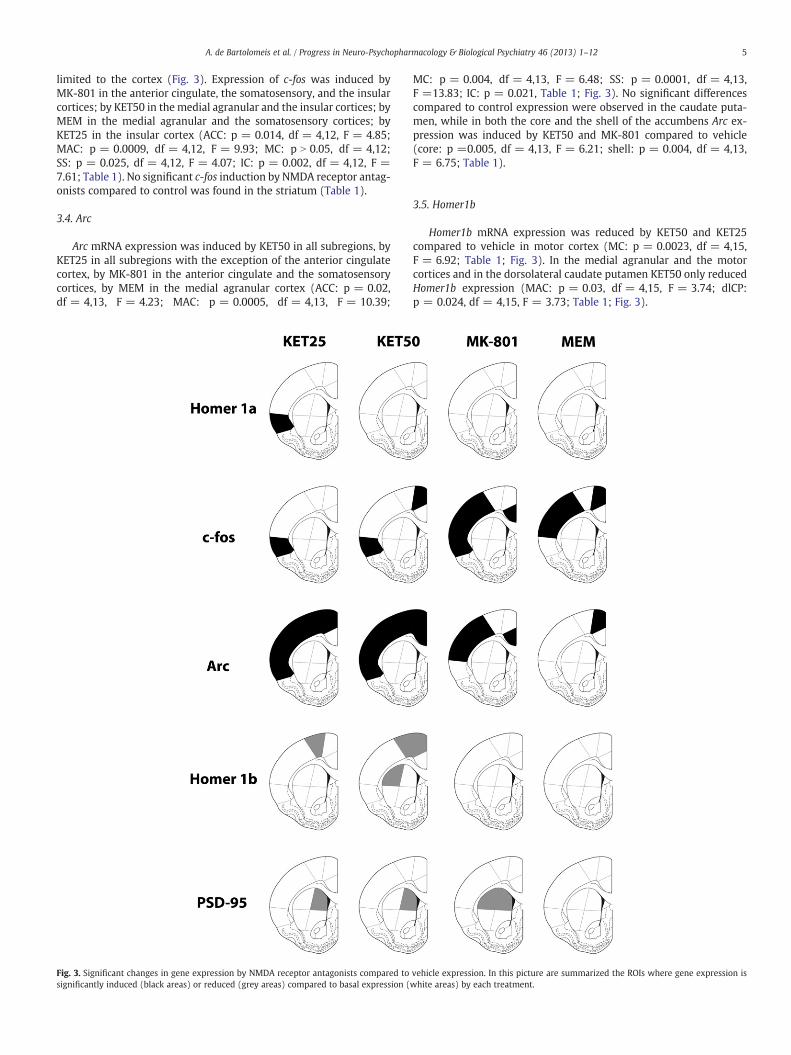
NMDA receptor antagonists mildly affected Homer1a mRNAexpression compared to the vehicle (Fig. 3). Indeed, Homer1a mRNAexpression was found significantly induced by KET25 in the insularcortex only (IC: p = 0.015, df = 4,13, F = 4.61). No other significantdifferences were found in the regions taken into account (Table 1).
3.3. c-fos
Expression of c-fosmRNAwas strongly affected by NMDA receptorantagonists compared to its expression by vehicle, although this was
n in corticostriatal regions for each transcript by memantine 5 mg/kg (MEM), MK-801cle (VEH). Brightness and contrast have been adapted to highlight differences amongipt (i.e. each autoradiogram has been adjusted with transcript-fixed values of brightness
5A. de Bartolomeis et al. / Progress in Neuro-Psychopharmacology & Biological Psychiatry 46 (2013) 1–12
limited to the cortex (Fig. 3). Expression of c-fos was induced byMK-801 in the anterior cingulate, the somatosensory, and the insularcortices; by KET50 in the medial agranular and the insular cortices; byMEM in the medial agranular and the somatosensory cortices; byKET25 in the insular cortex (ACC: p = 0.014, df = 4,12, F = 4.85;MAC: p = 0.0009, df = 4,12, F = 9.93; MC: p N 0.05, df = 4,12;SS: p = 0.025, df = 4,12, F = 4.07; IC: p = 0.002, df = 4,12, F =7.61; Table 1). No significant c-fos induction by NMDA receptor antag-onists compared to control was found in the striatum (Table 1).
3.4. Arc
Arc mRNA expression was induced by KET50 in all subregions, byKET25 in all subregions with the exception of the anterior cingulatecortex, by MK-801 in the anterior cingulate and the somatosensorycortices, by MEM in the medial agranular cortex (ACC: p = 0.02,df = 4,13, F = 4.23; MAC: p = 0.0005, df = 4,13, F = 10.39;
Fig. 3. Significant changes in gene expression by NMDA receptor antagonists compared tosignificantly induced (black areas) or reduced (grey areas) compared to basal expression (
MC: p = 0.004, df = 4,13, F = 6.48; SS: p = 0.0001, df = 4,13,F =13.83; IC: p = 0.021, Table 1; Fig. 3). No significant differencescompared to control expression were observed in the caudate puta-men, while in both the core and the shell of the accumbens Arc ex-pression was induced by KET50 and MK-801 compared to vehicle(core: p =0.005, df = 4,13, F = 6.21; shell: p = 0.004, df = 4,13,F = 6.75; Table 1).
3.5. Homer1b
Homer1b mRNA expression was reduced by KET50 and KET25compared to vehicle in motor cortex (MC: p = 0.0023, df = 4,15,F = 6.92; Table 1; Fig. 3). In the medial agranular and the motorcortices and in the dorsolateral caudate putamen KET50 only reducedHomer1b expression (MAC: p = 0.03, df = 4,15, F = 3.74; dlCP:p = 0.024, df = 4,15, F = 3.73; Table 1; Fig. 3).
vehicle expression. In this picture are summarized the ROIs where gene expression iswhite areas) by each treatment.
Table 1Summarization of mRNA expression amount (expressed in relative disintegrations per minute, d.p.m., as mean ± standard error mean) in each ROI by each treatment group.Significant differences among groups at the ANOVA test have been marked by an asterisk in the corresponding row of the ANOVA column. # denotes significances survivingBonferroni correction.
6 A. de Bartolomeis et al. / Progress in Neuro-Psychopharmacology & Biological Psychiatry 46 (2013) 1–12
3.6. PSD-95
PSD-95 mRNA expression was reduced by KET50, KET25, andMK-801 compared to vehicle in the dorsomedial caudate putamen(dmCP: p = 0.03, df = 4,14, F = 3.81; Fig. 3). PSD-95 expressionwas also reduced by MK-801 compared to control in the dorsolateralputamen (dlCP: p = 0.015, df = 4,14, F = 4.44; Table 1).
3.7. Significant changes in gene expression among treatment groups
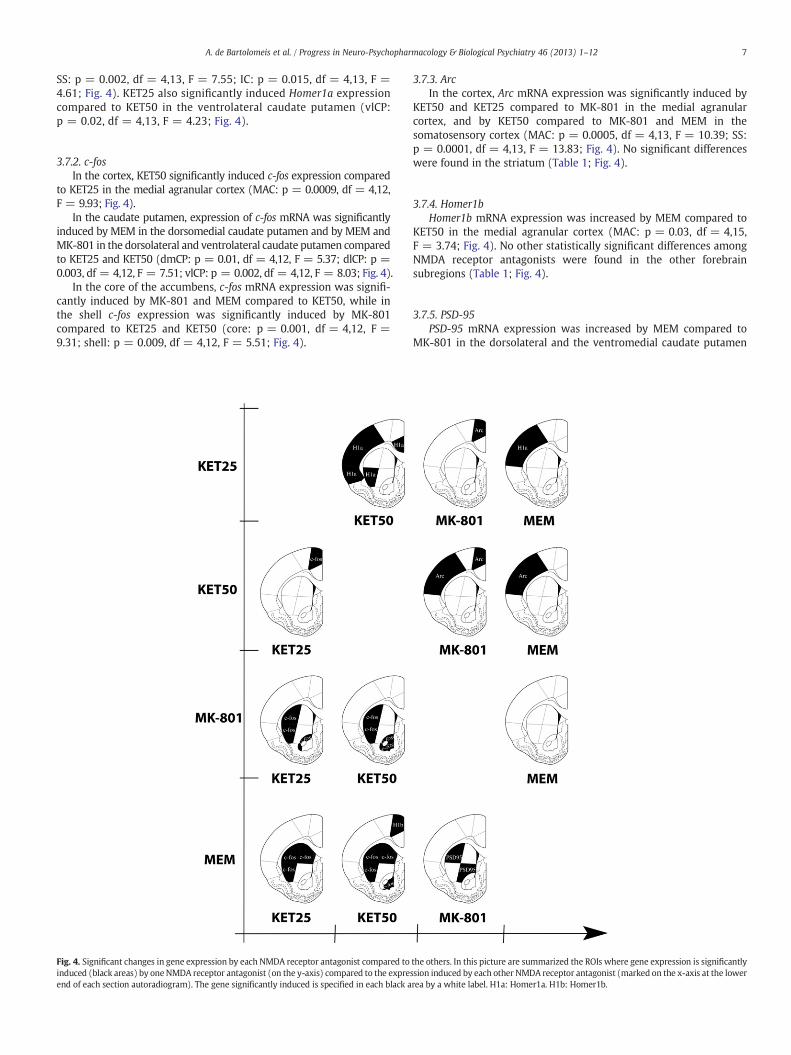
3.7.1. Homer1aIn the cortex, Homer1a mRNA expression was significantly
induced by KET25 compared to KET50 in the anterior cingulate, thesomatosensory, and the insular cortices, and compared to MEM inthe somatosensory cortex (ACC: p = 0.04, df = 4,13, F = 3.41;
7A. de Bartolomeis et al. / Progress in Neuro-Psychopharmacology & Biological Psychiatry 46 (2013) 1–12
SS: p = 0.002, df = 4,13, F = 7.55; IC: p = 0.015, df = 4,13, F =4.61; Fig. 4). KET25 also significantly induced Homer1a expressioncompared to KET50 in the ventrolateral caudate putamen (vlCP:p = 0.02, df = 4,13, F = 4.23; Fig. 4).
3.7.2. c-fosIn the cortex, KET50 significantly induced c-fos expression compared
to KET25 in the medial agranular cortex (MAC: p = 0.0009, df = 4,12,F = 9.93; Fig. 4).
In the caudate putamen, expression of c-fos mRNA was significantlyinduced by MEM in the dorsomedial caudate putamen and by MEM andMK-801 in the dorsolateral and ventrolateral caudate putamen comparedto KET25 and KET50 (dmCP: p = 0.01, df = 4,12, F = 5.37; dlCP: p =0.003, df = 4,12, F = 7.51; vlCP: p = 0.002, df = 4,12, F = 8.03; Fig. 4).
In the core of the accumbens, c-fos mRNA expression was signifi-cantly induced by MK-801 and MEM compared to KET50, while inthe shell c-fos expression was significantly induced by MK-801compared to KET25 and KET50 (core: p = 0.001, df = 4,12, F =9.31; shell: p = 0.009, df = 4,12, F = 5.51; Fig. 4).
Fig. 4. Significant changes in gene expression by each NMDA receptor antagonist compared toinduced (black areas) by one NMDA receptor antagonist (on the y-axis) compared to the expreend of each section autoradiogram). The gene significantly induced is specified in each black a
3.7.3. ArcIn the cortex, Arc mRNA expression was significantly induced by
KET50 and KET25 compared to MK-801 in the medial agranularcortex, and by KET50 compared to MK-801 and MEM in thesomatosensory cortex (MAC: p = 0.0005, df = 4,13, F = 10.39; SS:p = 0.0001, df = 4,13, F = 13.83; Fig. 4). No significant differenceswere found in the striatum (Table 1; Fig. 4).
3.7.4. Homer1bHomer1b mRNA expression was increased by MEM compared to
KET50 in the medial agranular cortex (MAC: p = 0.03, df = 4,15,F = 3.74; Fig. 4). No other statistically significant differences amongNMDA receptor antagonists were found in the other forebrainsubregions (Table 1; Fig. 4).
3.7.5. PSD-95PSD-95 mRNA expression was increased by MEM compared to
MK-801 in the dorsolateral and the ventromedial caudate putamen
the others. In this picture are summarized the ROIs where gene expression is significantlyssion induced by each other NMDA receptor antagonist (marked on the x-axis at the lowerrea by a white label. H1a: Homer1a. H1b: Homer1b.
8 A. de Bartolomeis et al. / Progress in Neuro-Psychopharmacology & Biological Psychiatry 46 (2013) 1–12
(dlCP: p = 0.015, df = 4,14, F = 4.44; vmCP: p = 0.04, df = 4,14,F = 3.27; Fig. 4).
3.8. Homer1a/Homer1b mRNA expression ratio
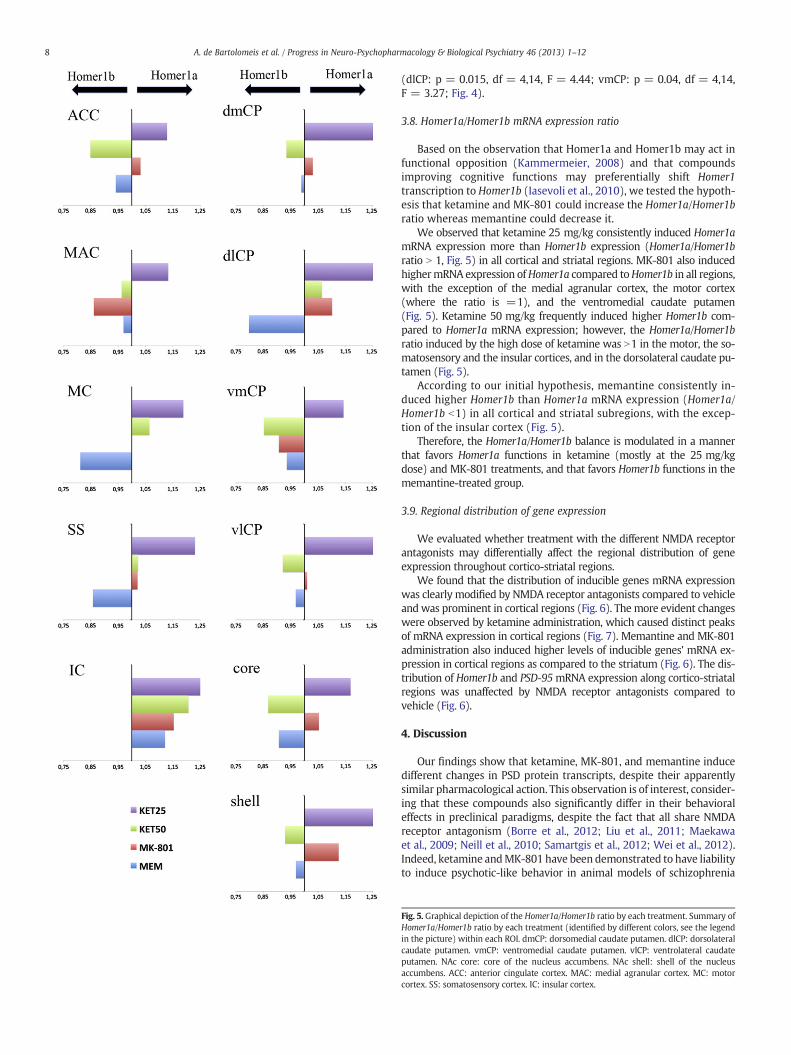
Based on the observation that Homer1a and Homer1b may act infunctional opposition (Kammermeier, 2008) and that compoundsimproving cognitive functions may preferentially shift Homer1transcription to Homer1b (Iasevoli et al., 2010), we tested the hypoth-esis that ketamine and MK-801 could increase the Homer1a/Homer1bratio whereas memantine could decrease it.
We observed that ketamine 25 mg/kg consistently induced Homer1amRNA expression more than Homer1b expression (Homer1a/Homer1bratio N 1, Fig. 5) in all cortical and striatal regions. MK-801 also inducedhighermRNA expression ofHomer1a compared toHomer1b in all regions,with the exception of the medial agranular cortex, the motor cortex(where the ratio is =1), and the ventromedial caudate putamen(Fig. 5). Ketamine 50 mg/kg frequently induced higher Homer1b com-pared to Homer1a mRNA expression; however, the Homer1a/Homer1bratio induced by the high dose of ketamine was N1 in the motor, the so-matosensory and the insular cortices, and in the dorsolateral caudate pu-tamen (Fig. 5).
According to our initial hypothesis, memantine consistently in-duced higher Homer1b than Homer1a mRNA expression (Homer1a/Homer1b b1) in all cortical and striatal subregions, with the excep-tion of the insular cortex (Fig. 5).
Therefore, the Homer1a/Homer1b balance is modulated in a mannerthat favors Homer1a functions in ketamine (mostly at the 25 mg/kgdose) and MK-801 treatments, and that favors Homer1b functions in thememantine-treated group.
3.9. Regional distribution of gene expression
We evaluated whether treatment with the different NMDA receptorantagonists may differentially affect the regional distribution of geneexpression throughout cortico-striatal regions.
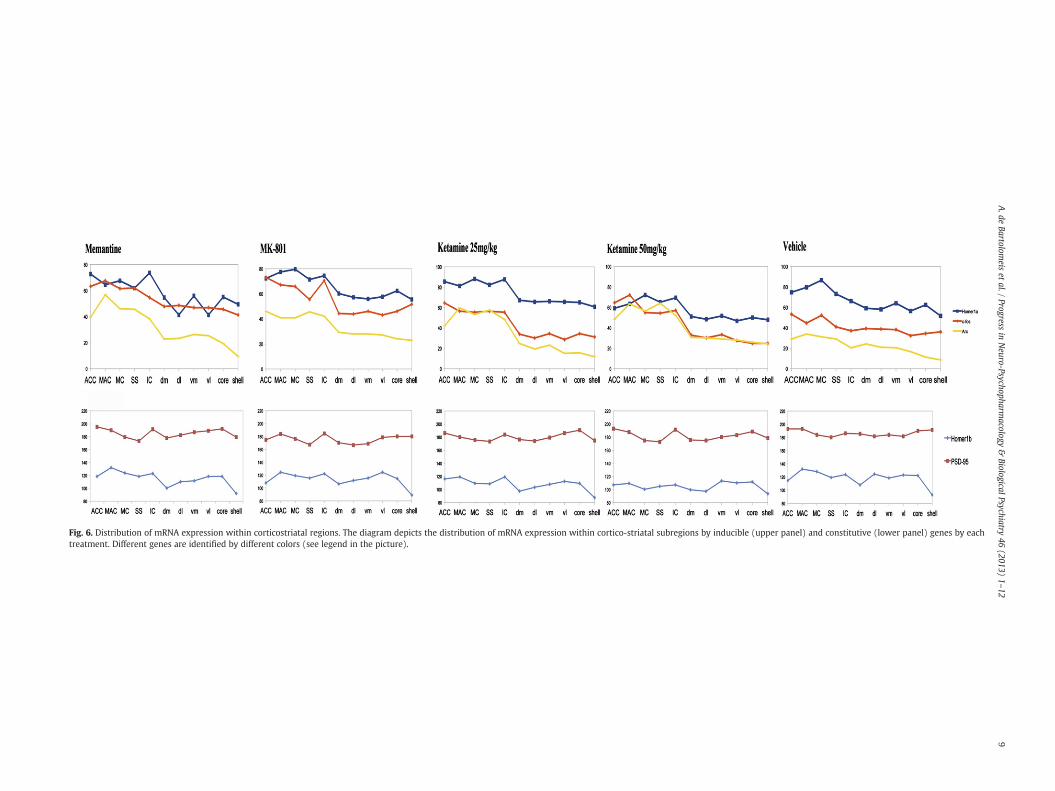
We found that the distribution of inducible genes mRNA expressionwas clearly modified by NMDA receptor antagonists compared to vehicleandwas prominent in cortical regions (Fig. 6). The more evident changeswere observed by ketamine administration, which caused distinct peaksof mRNA expression in cortical regions (Fig. 7). Memantine and MK-801administration also induced higher levels of inducible genes' mRNA ex-pression in cortical regions as compared to the striatum (Fig. 6). The dis-tribution of Homer1b and PSD-95mRNA expression along cortico-striatalregions was unaffected by NMDA receptor antagonists compared tovehicle (Fig. 6).
4. Discussion
Our findings show that ketamine, MK-801, and memantine inducedifferent changes in PSD protein transcripts, despite their apparentlysimilar pharmacological action. This observation is of interest, consider-ing that these compounds also significantly differ in their behavioraleffects in preclinical paradigms, despite the fact that all share NMDAreceptor antagonism (Borre et al., 2012; Liu et al., 2011; Maekawaet al., 2009; Neill et al., 2010; Samartgis et al., 2012; Wei et al., 2012).Indeed, ketamine andMK-801 have been demonstrated to have liabilityto induce psychotic-like behavior in animal models of schizophrenia
Fig. 5. Graphical depiction of the Homer1a/Homer1b ratio by each treatment. Summary ofHomer1a/Homer1b ratio by each treatment (identified by different colors, see the legendin the picture) within each ROI. dmCP: dorsomedial caudate putamen. dlCP: dorsolateralcaudate putamen. vmCP: ventromedial caudate putamen. vlCP: ventrolateral caudateputamen. NAc core: core of the nucleus accumbens. NAc shell: shell of the nucleusaccumbens. ACC: anterior cingulate cortex. MAC: medial agranular cortex. MC: motorcortex. SS: somatosensory cortex. IC: insular cortex.
Fig. 6. Distribution of mRNA expression within corticostriatal regions. The diagram depicts the distribution of mRNA expression within cortico-striatal subregions by inducible (upper panel) and constitutive (lower panel) genes by eachtreatment. Different genes are identified by different colors (see legend in the picture).
10 A. de Bartolomeis et al. / Progress in Neuro-Psychopharmacology & Biological Psychiatry 46 (2013) 1–12
(Coyle et al., 2012; Javitt et al., 2012; Kantrowitz and Javitt, 2010),whereas memantine has been approved for Alzheimer's diseasetreatment and has beenproposed for treatment-resistant schizophreniaand bipolar disorder (de Bartolomeis et al., 2012a; Sani et al., 2012;Wilkinson, 2012).
As a first step in this study, we evaluatedwhether the NMDA receptornon-competitive antagonists ketamine, MK-801, and memantine mightdifferently affect gene expression compared to the vehicle and then weinvestigated whether significant differences in gene expression couldalso be observed among these NMDA receptor antagonists. We predictedthat pro-psychotic agents, but not memantine, might prominently affectHomer1b or PSD-95 compared to Homer1a expression and mightoverexpress Arc.
We mainly found that: i) ketamine induced Homer1a expression inthe insular cortex compared to the vehicle, whereas it reduced Homer1band PSD-95 expression in cortical and striatal regions; ii)MK-801 reducedPSD-95 expression in the caudate putamen; iii) Arc expression wasextensively induced in the cortex by ketamine, moderately by MK-801,and in the medial agranular cortex only by memantine.
Homer1b and PSD-95 are key PSD molecules involved in synapticrearrangements underlying dendritic spine growth and synapticstrength (de Bartolomeis and Szumlinski, 2012; de Bartolomeiset al., 2012b; Iasevoli et al., 2013), and the down-regulation ofHomer1b has been found to attenuate glutamate-mediatedexcitotoxicity in rat cortical neurons (Chen et al., 2012b). It could behypothesized that reduction of Homer1b and PSD-95 expression byketamine andMK-801 may represent a negative feedback mechanismto counteract their “neurotoxic” potential and might contribute to ab-errant synaptic rearrangements.
Unlike ketamine and MK-801, memantine did not reduce theexpression of Homer1b and PSD-95 compared to control, consistentwith the putative neuroprotective action of this compound. In agree-ment with our results, memantine has been previously described toregulate the expression of genes implicated in neurotoxicity and/orneuroprotection (Chen et al., 2012a; Rosi et al., 2009). Specifically,memantine has been found to up-regulate BDNF mRNA and proteinexpression to inhibit HSP70 gene expression in hypoxic ischemia, as
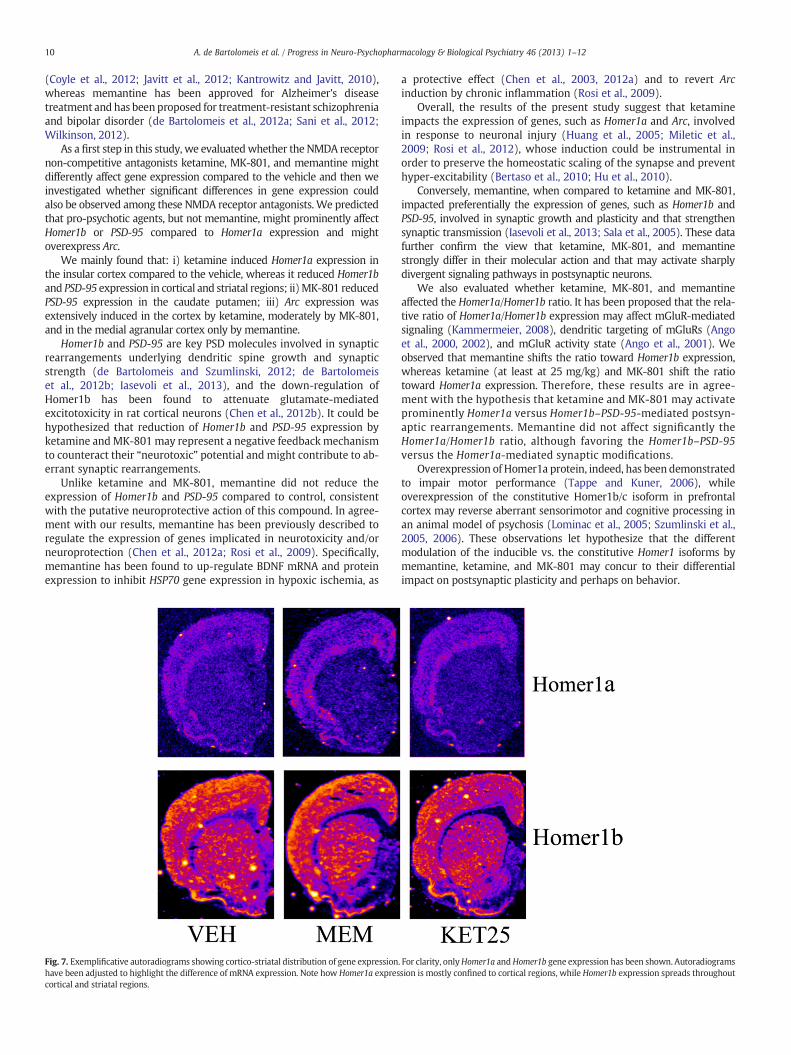
Fig. 7. Exemplificative autoradiograms showing cortico-striatal distribution of gene expressionhave been adjusted to highlight the difference of mRNA expression. Note how Homer1a exprescortical and striatal regions.
a protective effect (Chen et al., 2003, 2012a) and to revert Arcinduction by chronic inflammation (Rosi et al., 2009).
Overall, the results of the present study suggest that ketamineimpacts the expression of genes, such as Homer1a and Arc, involvedin response to neuronal injury (Huang et al., 2005; Miletic et al.,2009; Rosi et al., 2012), whose induction could be instrumental inorder to preserve the homeostatic scaling of the synapse and preventhyper-excitability (Bertaso et al., 2010; Hu et al., 2010).
Conversely, memantine, when compared to ketamine and MK-801,impacted preferentially the expression of genes, such as Homer1b andPSD-95, involved in synaptic growth and plasticity and that strengthensynaptic transmission (Iasevoli et al., 2013; Sala et al., 2005). These datafurther confirm the view that ketamine, MK-801, and memantinestrongly differ in their molecular action and that may activate sharplydivergent signaling pathways in postsynaptic neurons.
We also evaluated whether ketamine, MK-801, and memantineaffected the Homer1a/Homer1b ratio. It has been proposed that the rela-tive ratio of Homer1a/Homer1b expression may affect mGluR-mediatedsignaling (Kammermeier, 2008), dendritic targeting of mGluRs (Angoet al., 2000, 2002), and mGluR activity state (Ango et al., 2001). Weobserved that memantine shifts the ratio toward Homer1b expression,whereas ketamine (at least at 25 mg/kg) and MK-801 shift the ratiotoward Homer1a expression. Therefore, these results are in agree-ment with the hypothesis that ketamine and MK-801 may activateprominently Homer1a versus Homer1b–PSD-95-mediated postsyn-aptic rearrangements. Memantine did not affect significantly theHomer1a/Homer1b ratio, although favoring the Homer1b–PSD-95versus the Homer1a-mediated synaptic modifications.
Overexpression of Homer1a protein, indeed, has been demonstratedto impair motor performance (Tappe and Kuner, 2006), whileoverexpression of the constitutive Homer1b/c isoform in prefrontalcortex may reverse aberrant sensorimotor and cognitive processing inan animal model of psychosis (Lominac et al., 2005; Szumlinski et al.,2005, 2006). These observations let hypothesize that the differentmodulation of the inducible vs. the constitutive Homer1 isoforms bymemantine, ketamine, and MK-801 may concur to their differentialimpact on postsynaptic plasticity and perhaps on behavior.
. For clarity, onlyHomer1a andHomer1b gene expression has been shown. Autoradiogramssion is mostly confined to cortical regions, while Homer1b expression spreads throughout
11A. de Bartolomeis et al. / Progress in Neuro-Psychopharmacology & Biological Psychiatry 46 (2013) 1–12
As a third aimof thiswork,we evaluatedwhether the different NMDAreceptor antagonists induced gene expression with regional selectivity.Indeed, perturbation of glutamate transmission by ketamine or othersimilar agents has been observed to cause region-specific changes andmicrocircuit dysfunctions prominently in the cortex (Murray et al., inpress; Roopun et al., 2008). We observed that the administration ofNMDA receptor antagonists was correlated to a re-distribution of induc-ible gene expression, with more prominent expression in cortical regionscompared to striatal ones. In agreement with this result, mRNA expres-sion of c-fos, a marker of neuronal activity, was significantly induced byall NMDA receptor antagonists in cortical regions but not in the striatum,at least at the doses used in our experimental paradigm.
These results strengthen the view that the cortex represents a majorsite of action for NMDA receptor antagonists and may provide furtherevidence for previous behavioral observations on cortex-mediated cogni-tive functions by ketamine, MK-801, and memantine (Nagakura et al.,2012; Neill et al., 2010; Tikhonravov et al., 2010).
When analyzing the results of this study, it should be taken intoaccount that only few significant differences were conserved afterapplication of a very stringent post-hoc test (i.e. Bonferroni correc-tion). However, according to expert opinion in the field (Perneger,1998), we chose to adopt the Tukey's post-hoc test, that we considera reliable and conservative analysis, which allows minimizing the riskof both type I (false positives) and type II (false negatives) errors.
5. Conclusions
In conclusion, the results of our study support the view thatmemantine—at least at the doses used herein and prominently inthe cortex—differently modulates key PSD molecules compared toketamine and MK-801. This different modulation on PSD signalingnetworks may contribute, at least partially, to the different actionon neuroplasticity exerted by these compounds and may representone putative molecular substrate for their behavioral effects.
Acknowledgments
AdB conceived the experimental design. FI, CS, EFB, FM, and CTcarried out the experiments. AdB FI, CS, EFB, FM, CT and AE analyzedthe results. FI, CT, and AdB wrote the manuscript. All authors revisedand approved the draft in its final version. We also wish to thank Dr.Rodolfo Rossi and Dr. Gianmarco Latte for their help in realizing thefigures of this work.
This work has been made in part with the contribution of anunrestricted grant by H. Lundbeck A/S to the Department of Neuro-science of the University of Naples “Federico II”. The authorsdisclose any actual or potential conflict of interest within threeyears of beginning the submitted work that could inappropriatelyinfluence, or be perceived to influence, their work.
References
Ambesi-Impiombato A, D'Urso G, Muscettola G, de Bartolomeis A. Method for quantita-tive in situ hybridization histochemistry and image analysis applied for Homer1agene expression in rat brain. Brain Res Brain Res Protoc 2003;11:189–96.
Andine P, Widermark N, Axelsson R, Nyberg G, Olofsson U, Martensson E, et al.Characterization of MK-801-induced behavior as a putative rat model of psychosis.J Pharmacol Exp Ther 1999;290:1393–408.
Ango F, Pin JP, Tu JC, Xiao B, Worley PF, Bockaert J, et al. Dendritic and axonal targetingof type 5 metabotropic glutamate receptor is regulated by homer1 proteins andneuronal excitation. J Neurosci 2000;20:8710–6.
Ango F, Prezeau L, Muller T, Tu JC, Xiao B,Worley PF, et al. Agonist-independent activationof metabotropic glutamate receptors by the intracellular protein Homer. Nature2001;411:962–5.
Ango F, Robbe D, Tu JC, Xiao B, Worley PF, Pin JP, et al. Homer-dependent cell surface ex-pression of metabotropic glutamate receptor type 5 in neurons. Mol Cell Neurosci2002;20:323–9.
Beique JC, Na Y, Kuhl D, Worley PF, Huganir RL. Arc-dependent synapse-specifichomeostatic plasticity. Proc Natl Acad Sci U S A 2011;108:816–21.
Bertaso F, Roussignol G, Worley P, Bockaert J, Fagni L, Ango F. Homer1a-dependent crosstalkbetween NMDA and metabotropic glutamate receptors in mouse neurons. PLoS One2010;5:e9755.
Borre Y, Bosman E, Lemstra S, Westphal KG, Olivier B, Oosting RS. Memantine partlyrescues behavioral and cognitive deficits in an animal model of neurodegeneration.Neuropharmacology 2012;62:2010–7.
Camarasa J, Rodrigo T, Pubill D, Escubedo E. Memantine is a useful drug to prevent thespatial and non-spatial memory deficits induced by methamphetamine in rats.Pharmacol Res 2010;62:450–6.
Canever L, Oliveira L, D'Altoe de Luca R, Correa PT, de BFD, Matos MP, et al. A rodentmodel of schizophrenia reveals increase in creatine kinase activity with associatedbehavior changes. Oxid Med Cell Longev 2010;3:421–7.
Chen H, Liu Z, Zhou Z, Jiang M, Qian L, Wu S. The regulatory effect of memantine onexpression and synthesis of heat shock protein 70 gene in neonatal rat modelswith cerebral hypoxic ischemia. Chin Med J (Engl) 2003;116:558–64.
Chen SL, Tao PL, Chu CH, Chen SH, Wu HE, Tseng LF, et al. Low-dose memantineattenuated morphine addictive behavior through its anti-inflammation andneurotrophic effects in rats. J Neuroimmune Pharmacol 2012a;7:444–53.
Chen T, Fei F, Jiang XF, Zhang L, Qu Y, Huo K, et al. Down-regulation of Homer1b/c attenuatesglutamate-mediated excitotoxicity through endoplasmic reticulum and mitochondriapathways in rat cortical neurons. Free Radic Biol Med 2012b;52:208–17.
Coyle JT, Basu A, Benneyworth M, Balu D, Konopaske G. Glutamatergic synapticdysregulation in schizophrenia: therapeutic implications. Handb Exp Pharmacol2012:267–95.
Creeley C, Wozniak DF, Labruyere J, Taylor GT, Olney JW. Low doses of memantinedisrupt memory in adult rats. J Neurosci 2006;26:3923–32.
Danysz W, Parsons CG. Alzheimer's disease, beta-amyloid, glutamate, NMDA receptorsand memantine—searching for the connections. Br J Pharmacol 2012;167:324–52.
de Bartolomeis A, Szumlinski KK. Group 1 metabotropic receptors and schizophrenia.WIREs Membr Transp Signal 2012;1:94–103.
de Bartolomeis A, Tomasetti C. Calcium-dependent networks in dopamine–glutamateinteraction: the role of postsynaptic scaffolding proteins. Mol Neurobiol 2012;46:275–96.
de Bartolomeis A, Fiore G, Iasevoli F. Dopamine–glutamate interaction and antipsychoticsmechanism of action: implication for new pharmacological strategies in psychosis.Curr Pharm Des 2005;11:3561–94.
de Bartolomeis A, Sarappa C, Magara S, Iasevoli F. Targeting glutamate system for novelantipsychotic approaches: relevance for residual psychotic symptoms and treat-ment resistant schizophrenia. Eur J Pharmacol 2012a;682:1–11.
de Bartolomeis A, Tomasetti C, Cicale M, Yuan PX, Manji HK. Chronic treatment with lithiumor valproate modulates the expression of Homer1b/c and its related genes Shank andInositol 1,4,5-trisphosphate receptor. Eur Neuropsychopharmacol 2012b;22:527–35.
de Bartolomeis A, Buonaguro EF, Iasevoli F. Serotonin–glutamate and serotonin–dopamine reciprocal interactions as putative molecular targets for novel anti-psychotic treatments: from receptor heterodimers to postsynaptic scaffoldingand effector proteins. Psychopharmacology (Berl) 2013a;225:1–19.
de Bartolomeis A,Marmo, Buonaguro EF, Rossi R, Tomasetti C, Iasevoli F. Imaging brain geneexpression profiles by antipsychotics: region-specific action of amisulpride on postsyn-aptic density transcripts compared to haloperidol. Eur Neuropsychopharmacol 2013b.
de Oliveira L, Fraga DB, De Luca RD, Canever L, Ghedim FV, Matos MP, et al. Behavioralchanges and mitochondrial dysfunction in a rat model of schizophrenia induced byketamine. Metab Brain Dis 2011;26:69–77.
Fallgatter AJ, Ehlis AC, Herrmann MJ, Hohoff C, Reif A, Freitag CM, et al. DTNBP1(dysbindin) gene variants modulate prefrontal brain function in schizophrenicpatients—support for the glutamate hypothesis of schizophrenias. Genes BrainBehav 2010;9:489–97.
Hahn CG, Wang HY, Cho DS, Talbot K, Gur RE, Berrettini WH, et al. Altered neuregulin1-erbB4 signaling contributes to NMDA receptor hypofunction in schizophrenia.Nat Med 2006;12:824–8.
Hu JH, Park JM, Park S, Xiao B, Dehoff MH, Kim S, et al. Homeostatic scaling requiresgroup I mGluR activation mediated by Homer1a. Neuron 2010;68:1128–42.
Huang WD, Fei Z, Zhang X. Traumatic injury induced homer-1a gene expression incultured cortical neurons of rat. Neurosci Lett 2005;389:46–50.
Huang YN, Tsai RY, Lin SL, Chien CC, Cherng CH, Wu CT, et al. Amitriptyline attenuatesastrocyte activation and morphine tolerance in rats: role of the PSD-95/NR1/nNOS/PKCgamma signaling pathway. Behav Brain Res 2012;229:401–11.
Iasevoli F, Polese D, Ambesi-Impiombato A, Muscettola G, de Bartolomeis A.Ketamine-related expression of glutamatergic postsynaptic density genes: possibleimplications in psychosis. Neurosci Lett 2007;416:1–5.
Iasevoli F, Tomasetti C, Ambesi-Impiombato A, Muscettola G, de Bartolomeis A. Dopaminereceptor subtypes contribution to Homer1a induction: insights into antipsychoticmolecular action. Prog Neuropsychopharmacol Biol Psychiatry 2009;33:813–21.
Iasevoli F, Tomasetti C, Marmo F, Bravi D, Arnt J, de Bartolomeis A. Divergent acute andchronic modulation of glutamatergic postsynaptic density genes expression by the an-tipsychotics haloperidol and sertindole. Psychopharmacology (Berl) 2010;212:329–44.
Iasevoli F, Ambesi-Impiombato A, Fiore G, Panariello F, Muscettola G, de Bartolomeis A. Pat-tern of acute induction of Homer1a gene is preserved after chronic treatmentwithfirst-and second-generation antipsychotics: effect of short-term drug discontinuation andcomparison with Homer1a-interacting genes. J Psychopharmacol 2011;25:875–87.
Iasevoli F, Latte G, Avvisati L, Sarappa C, Aloj L, de Bartolomeis A. The expression ofgenes involved in glucose metabolism is affected by N-methyl-D-aspartatereceptor antagonism: a putative link between metabolism and an animal modelof psychosis. J Neurosci Res 2012a;90:1756–67.
Iasevoli M, Giantin V, Voci A, Valentini E, Zurlo A, Maggi S, et al. Discussing end-of-lifecare issues with terminally ill patients and their relatives: comparisons amongphysicians, nurses and psychologists. Aging Clin Exp Res 2012b;24:35–42.
12 A. de Bartolomeis et al. / Progress in Neuro-Psychopharmacology & Biological Psychiatry 46 (2013) 1–12
Iasevoli F, Tomasetti C, de Bartolomeis A. Scaffolding proteins of the post-synapticdensity contribute to synaptic plasticity by regulating receptor localization anddistribution: relevance for neuropsychiatric diseases. Neurochem Res 2013;38:1–22.
Ihalainen J, Sarajarvi T, Rasmusson D, Kemppainen S, Keski-Rahkonen P, Lehtonen M,et al. Effects of memantine and donepezil on cortical and hippocampalacetylcholine levels and object recognition memory in rats. Neuropharmacology2011;61:891–9.
Javitt DC, Zukin SR, Heresco-Levy U, Umbricht D. Has an angel shown the way?Etiological and therapeutic implications of the PCP/NMDA model of schizophrenia.Schizophr Bull 2012;38:958–66.
Johnson JW, Kotermanski SE. Mechanism of action of memantine. Curr Opin Pharmacol2006;6:61–7.
Kammermeier PJ. Endogenous homer proteins regulate metabotropic glutamatereceptor signaling in neurons. J Neurosci 2008;28:8560–7.
Kantrowitz JT, Javitt DC. Thinking glutamatergically: changing concepts of schizophreniabased upon changing neurochemical models. Clin Schizophr Relat Psychoses 2010;4:189–200.
Kantrowitz J, Javitt DC. Glutamatergic transmission in schizophrenia: from basicresearch to clinical practice. Curr Opin Psychiatry 2012;25:96–102.
Klugmann M, Symes CW, Leichtlein CB, Klaussner BK, Dunning J, Fong D, et al.AAV-mediated hippocampal expression of short and long Homer 1 proteinsdifferentially affect cognition and seizure activity in adult rats. Mol Cell Neurosci2005;28:347–60.
Koukopoulos A, Reginaldi D, Serra G, Sani G. Antimanic and mood-stabilizing effect ofmemantine as an augmenting agent in treatment-resistant bipolar disorder.Bipolar Disord 2010;12:348–9.
Kovacs KJ. Measurement of immediate-early gene activation—c-fos and beyond.J Neuroendocrinol 2008;20:665–72.
Li N, Lee B, Liu RJ, Banasr M, Dwyer JM, Iwata M, et al. mTOR-dependent synapseformation underlies the rapid antidepressant effects of NMDA antagonists. Science2010;329:959–64.
Lipska BK, Weinberger DR. To model a psychiatric disorder in animals: schizophreniaas a reality test. Neuropsychopharmacology 2000;23:223–39.
Liu F, Paule MG, Ali S, Wang C. Ketamine-induced neurotoxicity and changes in geneexpression in the developing rat brain. Curr Neuropharmacol 2011;9:256–61.
Lominac KD, Oleson EB, Pava M, Klugmann M, Schwarz MK, Seeburg PH, et al. Distinctroles for different Homer1 isoforms in behaviors and associated prefrontal cortexfunction. J Neurosci 2005;25:11586–94.
Maekawa M, Namba T, Suzuki E, Yuasa S, Kohsaka S, Uchino S. NMDA receptorantagonist memantine promotes cell proliferation and production of maturegranule neurons in the adult hippocampus. Neurosci Res 2009;63:259–66.
Menard C, Quirion R. Successful cognitive aging in rats: a role for mGluR5 glutamatereceptors, homer 1 proteins and downstream signaling pathways. PLoS One2012;7:e28666.
Miletic G, Driver AM, Miyabe-Nishiwaki T, Miletic V. Early changes in Homer1 proteinsin the spinal dorsal horn are associated with loose ligation of the rat sciatic nerve.Anesth Analg 2009;109:2000–7.
Misztal M, Danysz W. Comparison of glutamate antagonists in continuous multiple-trialand single-trial dark avoidance. Behav Pharmacol 1995;6:550–61.
Moghaddam B, Krystal JH. Capturing the angel in “angel dust”: twenty years of transla-tional neuroscience studies of NMDA receptor antagonists in animals and humans.Schizophr Bull 2012;38:942–9.
MoghaddamB, Adams B, Verma A, Daly D. Activation of glutamatergic neurotransmission byketamine: a novel step in the pathway from NMDA receptor blockade to dopaminergicand cognitive disruptions associated with the prefrontal cortex. J Neurosci 1997;17:2921–7.
More L, Gravius A, Nagel J, Valastro B, Greco S, Danysz W. Therapeutically relevantplasma concentrations of memantine produce significant L-N-methyl-D-aspartatereceptor occupation and do not impair learning in rats. Behav Pharmacol2008;19:724–34.
Morgan CJ, Mofeez A, Brandner B, Bromley L, Curran HV. Ketamine impairs responseinhibition and is positively reinforcing in healthy volunteers: a dose–responsestudy. Psychopharmacology (Berl) 2004;172:298–308.
Murray JD, Anticevic A, Gancsos M, Ichinose M, Corlett PR, Krystal JH, et al. Linkingmicrocircuit dysfunction to cognitive impairment: effects of disinhibition associatedwith schizophrenia in a cortical working memory model. Cereb Cortex 2012. http://dx.doi.org/10.1093/cercor/bhs370 [in press].
Nagakura A, Shitaka Y, Yarimizu J, Matsuoka N. Characterization of cognitive deficits ina transgenic mouse model of Alzheimer's disease and effects of donepezil andmemantine. Eur J Pharmacol 2012.
Nakaya K, Nakagawasai O, Arai Y, Onogi H, Sato A, Niijima F, et al. Pharmacologicalcharacterizations of memantine-induced disruption of prepulse inhibition of theacoustic startle response in mice: involvement of dopamine D2 and 5-HT2Areceptors. Behav Brain Res 2011;218:165–73.
Neill JC, Barnes S, Cook S, Grayson B, Idris NF, McLean SL, et al. Animal models ofcognitive dysfunction and negative symptoms of schizophrenia: focus on NMDAreceptor antagonism. Pharmacol Ther 2010;128:419–32.
Paxinos G, Watson C. The rat brain in stereotaxic coordinates. Sidney, Australia:Academic Press Inc.; 1997.
Perneger TV. What's wrong with Bonferroni adjustments 1998;316:1236–8. [(Clinicalresearch ed.].
Pinault D. N-methyl D-aspartate receptor antagonists ketamine and MK-801 inducewake-related aberrant gamma oscillations in the rat neocortex. Biol Psychiatry2008;63:730–5.
Reus GZ, Valvassori SS, Machado RA, Martins MR, Gavioli EC, Quevedo J. Acute treatmentwith low doses of memantine does not impair aversive, non-associative and recogni-tion memory in rats. Naunyn Schmiedebergs Arch Pharmacol 2008;376:295–300.
Ronesi JA, Collins KA, Hays SA, Tsai NP, GuoW, Birnbaum SG, et al. Disrupted Homer scaf-folds mediate abnormal mGluR5 function in a mouse model of fragile X syndrome.Nat Neurosci 2012;15(S1):431–40.
Roopun AK, Cunningham MO, Racca C, Alter K, Traub RD, Whittington MA.Region-specific changes in gamma and beta2 rhythms in NMDA receptor dysfunc-tion models of schizophrenia. Schizophr Bull 2008;34:962–73.
Roselli F, Hutzler P, Wegerich Y, Livrea P, Almeida OF. Disassembly of shank and homersynaptic clusters is driven by soluble beta-amyloid(1–40) through divergentNMDAR-dependent signalling pathways. PLoS One 2009;4:e6011.
Rosi S, Ramirez-Amaya V, Vazdarjanova A, Esparza EE, Larkin PB, Fike JR, et al. Accuracyof hippocampal network activity is disrupted by neuroinflammation: rescue bymemantine. Brain 2009;132:2464–77.
Rosi S, Belarbi K, Ferguson RA, Fishman K, Obenaus A, Raber J, et al. Trauma-inducedalterations in cognition and Arc expression are reduced by previous exposure to56Fe irradiation. Hippocampus 2012;22:544–54.
Rudinskiy N, Hawkes JM, Betensky RA, Eguchi M, Yamaguchi S, Spires-Jones TL, et al.Orchestrated experience-driven Arc responses are disrupted in a mouse model ofAlzheimer's disease. Nat Neurosci 2012;15:1422–9.
Sala C, Futai K, Yamamoto K, Worley PF, Hayashi Y, Sheng M. Inhibition of dendriticspine morphogenesis and synaptic transmission by activity-inducible proteinHomer1a. J Neurosci 2003;23:6327–37.
Sala C, Roussignol G, Meldolesi J, Fagni L. Key role of the postsynaptic density scaffoldproteins Shank and Homer in the functional architecture of Ca2+ homeostasis atdendritic spines in hippocampal neurons. J Neurosci 2005;25:4587–92.
Samartgis JR, Schachte L, Hazi A, Crowe SF. Memantine facilitatesmemory consolidation andreconsolidation in the day-old chick. Neurobiol Learn Mem 2012;97:380–5.
Sani G, Serra G, Kotzalidis GD, Romano S, Tamorri SM, Manfredi G, et al. The role ofmemantine in the treatment of psychiatric disorders other than the dementias: areview of current preclinical and clinical evidence. CNS Drugs 2012;26:663–90.
Szumlinski KK, Lominac KD, Kleschen MJ, Oleson EB, Dehoff MH, Schwarz MK, et al.Behavioral and neurochemical phenotyping of Homer1 mutant mice: possiblerelevance to schizophrenia. Genes Brain Behav 2005;4:273–88.
Szumlinski KK, Kalivas PW, Worley PF. Homer proteins: implications for neuropsychiatricdisorders. Curr Opin Neurobiol 2006;16:251–7.
Tappe A, Kuner R. Regulation of motor performance and striatal function by synapticscaffolding proteins of the Homer1 family. Proc Natl Acad Sci U S A 2006;103:774–9.
Tariot PN, Farlow MR, Grossberg GT, Graham SM, McDonald S, Gergel I, et al. Memantinetreatment in patients with moderate to severe Alzheimer disease already receivingdonepezil: a randomized controlled trial. JAMA 2004;291:317–24.
Tikhonravov D, Neuvonen T, Pertovaara A, Savioja K, Ruusuvirta T, Naatanen R, et al.Dose-related effects of memantine on a mismatch negativity-like response inanesthetized rats. Neuroscience 2010;167:1175–82.
Timms AE, Dorschner MO, Wechsler J, Choi KY, Kirkwood R, Girirajan S, et al. Supportfor the N-methyl-D-aspartate receptor hypofunction hypothesis of schizophreniafrom exome sequencing in multiplex families. JAMA Psychiatry 2013:1–9.
Tomasetti C, Dell'Aversano C, Iasevoli F, Marmo F, de Bartolomeis A. The acute and chroniceffects of combined antipsychotic-mood stabilizing treatment on the expression ofcortical and striatal postsynaptic density genes. Prog Neuropsychopharmacol BiolPsychiatry 2011;35:184–97.
Toro C, Deakin JF. NMDA receptor subunit NRI and postsynaptic protein PSD-95 inhippocampus and orbitofrontal cortex in schizophrenia and mood disorder.Schizophr Res 2005;80:323–30.
Umbricht D, Schmid L, Koller R, Vollenweider FX, Hell D, Javitt DC. Ketamine-induceddeficits in auditory and visual context-dependent processing in healthy volun-teers: implications for models of cognitive deficits in schizophrenia. Arch GenPsychiatry 2000;57:1139–47.
Uribe E, Landaeta J, Wix R, Eblen A. Memantine reverses social withdrawal induced byketamine in rats. Exp Neurobiol 2013;22:18–22.
Verma A, Moghaddam B. NMDA receptor antagonists impair prefrontal cortex functionas assessed via spatial delayed alternation performance in rats: modulation bydopamine. J Neurosci 1996;16:373–9.
Wei H, Dobkin C, Sheikh AM, Malik M, Brown WT, Li X. The therapeutic effect ofmemantine through the stimulation of synapse formation and dendritic spinematuration in autism and fragile X syndrome. PLoS One 2012;7:e36981.
Wenk GL, Zajaczkowski W, Danysz W. Neuroprotection of acetylcholinergic basal fore-brain neurons by memantine and neurokinin B. Behav Brain Res 1997;83:129–33.
Wilkinson D. A review of the effects of memantine on clinical progression inAlzheimer's disease. Int J Geriatr Psychiatry 2012;27:769–76.
ZajaczkowskiW, Quack G, DanyszW. Infusion of (+) -MK-801 andmemantine—contrastingeffects on radial maze learning in rats with entorhinal cortex lesion. Eur J Pharmacol1996;296:239–46.
Zdanys K, Tampi RR. A systematic review of off-label uses of memantine for psychiatricdisorders. Prog Neuropsychopharmacol Biol Psychiatry 2008;32:1362–74.
Zoladz PR, Campbell AM, Park CR, Schaefer D, Danysz W, Diamond DM. Enhancement oflong-term spatial memory in adult rats by the noncompetitive NMDA receptor antag-onists, memantine and neramexane. Pharmacol Biochem Behav 2006;85:298–306.