Digital control circuitry of cancer cell and its apoptosis
R. M. Ardito Marretta∗ and G. Barbaraci†
Abstract: This study, through a typicalaerospace systems architecture, suggests an engi-neering design of a human cancer cell circuitry inwhich a digital optimal control matrix is assignedto repair the DNA damage level and/or to triggerits apoptosis.
Here, the conceived machinery is proposed takinginto account the state of the art in cancer investi-gation. However, it could be further generalized.
The most recent studies on cancer pathologiesgive a predominant role to the oncosuppressorprotein p53 and its antagonist, the oncogeneMdm2.
Experimental and theoretical approaches are inagreement in deducing a “digital” response of thep53 when genomic integrity is damaged. OnceDNA damage is present, the mutual influence ofp53 and its antagonist, the Mdm2 oncogene, isclosed in a feedback loop.
In this work, starting from these current results,a novel molecular mechanism is proposed, basedon a digital optimal control law, whereby p53 andMdm2 proteins activities can be represented byappropriate circuitry and governed by the optimalcontrol law of digital systems.
This procedure obtains a real-time sequence eval-uation of protein oscillations and an unexpectedand relevant acceleration in the DNA repairingwhen suitable digital control matrix is imple-mented.
Those effects suggest interesting perspectives forfuture scientific investigations.
First of all, the proposed digital circuitry, receiv-
∗ Correspondence author. Dipartimento di Ingegneria Strut-turale e Geotecnica, Università di Palermo, Edificio 8,90128, Palermo, (Italy); Tel.: 091-6459912, Fax: 091-485439; Email: [email protected]
† Department of Mechanical Engineering, University ofPalermo, 90128, Palermo, (Italy)
ing the p53 signal from a damaged cell, is ableto repair the current level of genomic alteration.Moreover, the cell fate is newly conceived andbound by the modified pulsing mechanism of p53.
1 Introduction
Since its discovery, the oncosuppressor p53 pro-tein seems to play a prominent role in the evolu-tion of a cancer cell.
Activation and high concentration of p53 are theresponse of the cell “system” to aberrant onco-gene signals.
This protein is capable of inducing the tran-scription of genes in charge of the cell-cycle ar-rest, DNA repair and apoptosis (see Ma, Wag-ner, Rice, Hu, Levine and Stlovitzky (2005);Geva-Zatorsky, Rosenfeld, Itzkovitz, Milo, Si-gal, Dekel, Yarnitzky, Liron, Polak, Lahav et al.(2006); Shangary and Wang (2008); Ciliberto,Novak and Tyson (2005); Zhang, Brazhnik andTyson (2007)).
At this point, an engineering frame of mindwas adopted by the present authors (whose dis-ciplines lie inside aerospace technologies) whenthey faced the acknowledgement of the p53 pro-tein inner mechanism and its role in the humansingle-cell architecture.
Let us view the protein p53 as a battle array inwhich 393 brigades (residues) form divisions (do-mains) having a specific biological role. Then, thefunctional domains of p53 are distinguished intotheir roles and residues; those from 1 to 42 foractivation domain AD1, from 43 to 63 for anotheractivation domain AD2, from 64 to 91 for praline-rich domain PRD, up to 364 to 393 for C-terminalbasic domain BD.
An outstanding recent work (see Harms and Chen(2005)), not only confirms that the above men-
tioned domain BD of p53 is inhibitory in vivo butalso demonstrates that activation of AD2 is re-quired for the pro-apoptotic target gene bindingprotein IGFBP3. The IGFBP3 seems to play adominant role in the apoptosis induced by N- andC- terminally truncated p53.
These conclusions seem to be related to a previousstudy of Bell, Klein, Muller, Hansen and Buch-ner (2002), in which their analysis showed that,at physiological temperatures, wild-type p53 wasmore than 50% unfolded with a 75% loss in DNA-binding activity. Moreover, those results foundthat full-length p53 contained large unstructuredregions in its N- and C-terminal parts.
Although other interesting works could be con-sidered milestones from a purely biological pointof view about p53 behaviour (see Reich, Orenand Levine (1983); Ventura, Kirsch, McLaugh-lin, Tuveson, Grimm, Lintault, Newman, Reczek,Weissleder and Jacks (2007); Bates, Phillips,Clark, Stott, Peters, Ludwig and Vousden (1998);Bar-Or, Maya, Segel, Alon, Levine and Oren(2000)), they remain far from being used math-ematically; for two reasons: first, their adoptedprocedures make use of chemicals, reagents andenzymes and, secondly, they become inappropri-ate if one decides to translate them into unsteadymathematical simulations and computational pre-dictions.
The guidelines of the present paper are thosegiven by Ma, Wagner, Rice, Hu, Levine andStlovitzky (2005); Geva-Zatorsky, Rosenfeld,Itzkovitz, Milo, Sigal, Dekel, Yarnitzky, Liron,Polak, Lahav et al. (2006); Shangary andWang (2008); Ciliberto, Novak and Tyson (2005);Zhang, Brazhnik and Tyson (2007); Bar-Or,Maya, Segel, Alon, Levine and Oren (2000);Lahav, Rosenfeld, Sigal, Geva-Zatorsky, Levine,Elowitz and Alon (2004); Loewer and Lahav(2006); Batchelor, Mock, Bhan, Loewer and La-hav (2008) in which theoretical and experimen-tal studies about oscillations of the p53/Mdm2feedback loop support the chance to mathemati-cally describe this human single-cell proteins ac-tivity and their inner forms dynamics. Mean-while, the methodological approaches, taken asframes of mind, are those of Tsourkas and
Weissleder (2004); Deguchi, Ohashi and Sato(2005); Mooney (2006); Zhou, Chen and Zhang(2007).
Briefly, some of these studies come to convergingconclusions, i.e., p53 can be expressed into a se-ries of discrete pulses after DNA damage and thewhole p53/Mdm2 network system is constrainedby a feedback loop and theoretically expressed bya digital scheme. However, these works show dif-ferences in the analysis of results depending onthe sign of the claimed feedback loops.
As a consequence, although these proposed math-ematical models and experiments capture the os-cillating characteristics of p53 and Mdm2, theyshow different and almost conflicting points ofview when linked to the interpretation of resultsin terms of output amplitudes, frequencies, in-terpulses and other oscillation parameters of thep53/Mdm2 dynamic responses. In short, a nega-tive feedback loop generates damped oscillations,while schemes such as positive feedback loops ofp53 may enhance undamped dynamics.
Lahav, Rosenfeld, Sigal, Geva-Zatorsky, Levine,Elowitz and Alon (2004) confirm the p53/Mdm2negative feedback loop oscillations once func-tional p53-CFP (cyan fluorescent protein) andMdm2-YFP (yellow fluorescent protein) fusionproteins are observed through time-lapse fluores-cent microscopy. After γ-irradiation at a 20minresolution during 16h of growth, these authorsmeasured the total fluorescence in the nuclei ofover 200 different cells through a movie tech-nique. Their conclusions regarding the p53 dig-ital behaviour consist of different fractions ofcells showing zero, one, two or more pulsesas a function of γ-irradiation dose; the widthof each pulse was 350±160min, the timing ofthe first pulse maximum being rather variable(360±240min) after damage; the time betweenthe maxima of two consecutive pulses is more pre-cise, i.e., 340±100min. Thus, they found that inthe p53/Mdm2 feedback loop system, the numberof pulses, but not the size or shape of each pulse,depends on the level of the input signal.
An important contribution for understanding thedynamics and variability of p53/Mdm2 systemis given by Geva-Zatorsky, Levine, Elowitz and
Digital control circuitry of cancer cell and its apoptosis 177
Alon (2006). They evaluate the amplitude andwidth of each peak of nuclear Mdm2-YFP andcalculate the average of these properties. Ul-timately, those authors obtained prolonged un-damped oscillations in the p53/Mdm2 system.The onset of oscillations was synchronized withthe DNA damage signal and cells gradually lostsynchrony with each other due to variations in os-cillation frequencies. The characteristic oscilla-tion frequency in each cell was found by Fourieranalysis.
It should be noted that if undamped oscillationsof p53 are obtained, one has to evaluate the be-haviour – in the same bandwidth – of its an-tagonist (and its forms) for taking into account(if present) the cross-correlation factors and fre-quency aliasing distortion.
Batchelor, Mock, Bhan, Loewer and Lahav(2008) draw the conclusion that the p53/Mdm2negative feedback loop is composed of interactionof two different timescales: one, a slow positivetranscriptional arm and a fast negative protein-protein interaction arm. Their results show thatthe p53/Mdm2 feedback loop does not by itselfdrive sustained p53 oscillations. Thus, they iden-tify the wild type p53-induced phosphatase 1,Wip1, as the central element mediating a second(negative) feedback loop for chaining p53 and theupstream signalling proteins.
For the present aim of this paper, two recent arti-cles of Ciliberto, Novak and Tyson (2005); Zhang,Brazhnik and Tyson (2007) will be useful for im-proving a cellular circuitry which is capable ofdigitally processing the p53/Mdm2 system dy-namics and giving a subject for discussion aboutits influence on apoptosis. Both these papers dealwith the mechanisms for triggering p53 pulses inresponse to a DNA damage.
These authors elegantly showed how to obtainsustained p53 oscillations when the p53/Mdm2negative feedback can be supplemented and in-tegrated by a positive loop. Although negativefeedback is necessary for triggering oscillationsin the p53/Mdm2 system, it is not yet sufficient.In fact, if one considers a negative feedback loopwith only two elements (p53→Mdm2 7→ |p53),it cannot oscillate.
Moreover, in the paper of Zhang, Brazhnik andTyson (2007), observations aimed at the employ-ment of the p53 negative feedback loop and its ob-served oscillations address some important ques-tions about the roles of positive feedback loopsin generating and stabilizing oscillations and howapoptosis may be triggered by repeated pulses ofp53.
It is our firm belief that understanding the promi-nent role of the p53/Mdm2 loop and its sub-system dynamics offers a more than promising av-enue for effective cancer therapy.
Now making reference to the previous studiesof Ardito Marretta and Marino (2007); ArditoMarretta, Marino and Bianchi (2008), regardingdigital aerospace well-suited active control mod-els and computer active control systems, ad hocdigital cellular circuitry is built up to pointwiserecognize the cell damage checkpoint, the time-dependent p53 levels and the triggering of pulses.Moreover, a more complex digital cell schemecan determine different number of pulses when re-lated to the expression of downstream genes andtheir evolution to give the chance – in spin-off –to assign gradients of considered species with de-sired levels of p53 and/or leading to cell death.
Then we propose an integration of the proposedmodels whatever type of feedback loop is em-ployed. The digital control system theory, mathe-matically expressed in terms of state-space theory,could unify the disparate observations and offerthe possibility of investigating apoptosis once thedynamics of inner protein forms is considered.
2 Adopted method
To design suitable digital circuitry for a hu-man single-cell, we start by taking into ac-count the protein forms of p53 (p53 mono-ubiquitinated, p53 poly-ubiquitinated and p53 to-tal, i.e., p53U , p53UU and p53tot , respectively)and Mdm2 (nuclear, cytoplasmic and phospho-rilated, i.e., Mdm2nuc, Mdm2cyt and Mdm2Pcyt ,respectively) with their time-dependency from aset of equations (see Ciliberto, Novak and Tyson(2005)). A powerful tool for resolving the prob-lem consists of processing the set of equations in
Figure 1: Human cell circuitry with control matrix
[P53 ] Rate State Vector1
[p53 ]
[Mdm 2nuc ]
[p53 U ]
[p53 UU ]
[p53 U ] '
[p53 ]
[p53 U ]
[p53 UU ]
[p53 ] '
[p53 U ] '
[p53 UU ] '
([ p53 tot ] ''; [p53 U ]''; [p53 UU ] '' )
[Mdm 2nuc ]
[p53 U ]
[p53 UU ]
[p53 UU ] '
[p53 ]
[p53 U ]
[p53 UU ]
[p53 U ] '
[p53 UU ] '
[p53 tot ] '
m
emu
[Mdm 2nuc ]2
[P53 ] State Vector1
Figure 2: Cell dynamic matrix
a computational space, i.e., the state-space whichmathematically represents all the possible condi-tions and combinations among the variables of theproblem. Once the state-space contains the nec-essary equations, the numerical solution will bethe task of matrix calculus. In this case, througha matrix representation in the state-space of themutual p53 and Mdm2 dynamics, a control ma-trix acting in positive feedback can be obtainedand processed.
Control theory is widely employed in many engi-
neering designs and several digital control strate-gies are currently applied to suppress or amplifyinstabilities and/or margins of safety. For digi-tally design of such control computer-aided sys-tems, devoted algorithms must be developed andprocessed in appropriate circuitries. Similarly totypical digital control system design, a convenientset of mathematical equations may be availablefor processing the biological system parametersand their mutual influences. For the single cellcircuitry design, we digitally transform the math-
Digital control circuitry of cancer cell and its apoptosis 179
ks53 pp
ks53 pp
ks2pp
ks2pp
[p53 ], [Mdm 2nuc ], [DNAdam ]
[P53 ] Vector Component
[Mdm 2] Vector Components
[DNAdam ], kd2
Subsystem 2
[P53]
[P21]
[P53AIP1]
[APAF1]
[cytoc ]
[Apoptosoma ]
[DNAdam ] [DNAdam ]
Subsystem
[P53] B([P53])
Subsystem
[Mdm2cyt ] B([Mdm2cyt ])
Scope
Integrator
1s
Integrator
1s
Dynamic System of [P53 ] Protein
[P53] State Vector
[P53]
[Mdm2nuc]
A*[P53] State Vector
Dynamic System of [Mdm 2] Protein
[Mdm2] State Vector
[DNAdam ]
A*[Mdm2] State Vector
kd2
m
m
[0;ks2p;0;0]
[ks53 p;0]
Figure 3: Cell digital circuitry (model #1 Zhang, Brazhnik and Tyson)
ematical model proposed by Ciliberto, Novak andTyson (2005) (see figs 1 and 2) and the model #1of those employed by Zhang, Brazhnik and Tyson(2007) (see fig. 3).
Also, we digitally connect the control circuitriesobtained from those models with a wiring dia-gram of apoptosis (fig. 4), as given by Zhang,Brazhnik and Tyson (2007).
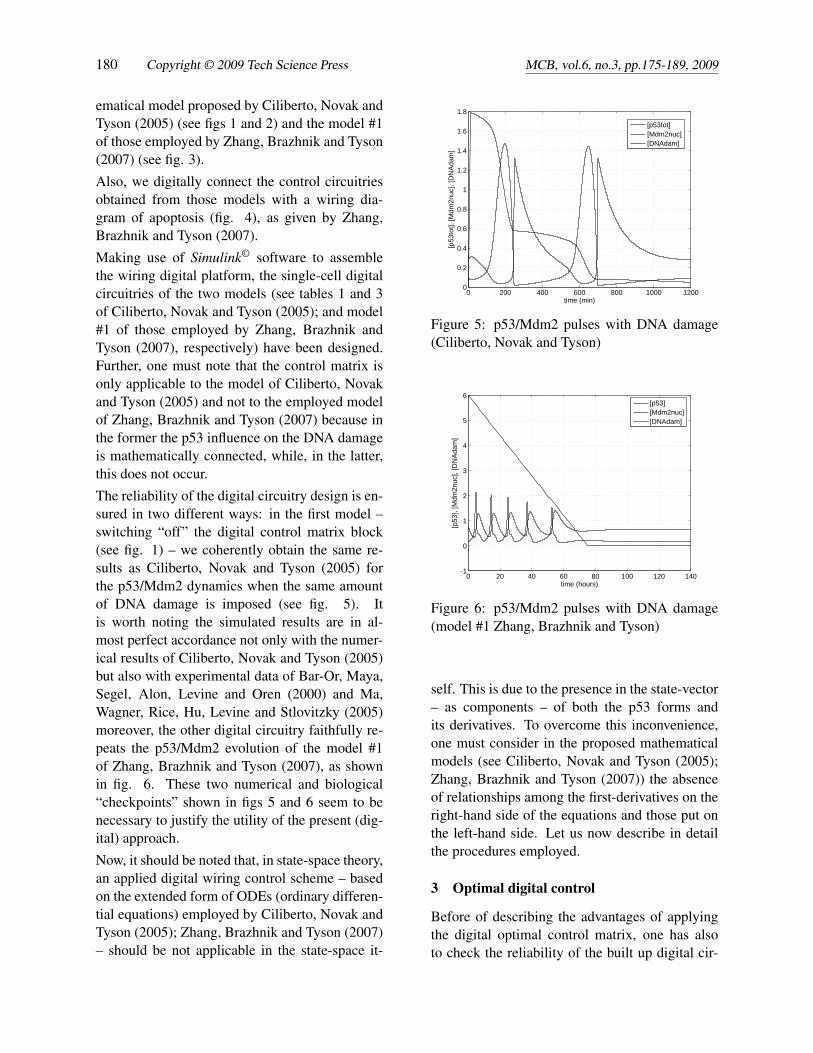
The reliability of the digital circuitry design is en-sured in two different ways: in the first model –switching “off” the digital control matrix block(see fig. 1) – we coherently obtain the same re-sults as Ciliberto, Novak and Tyson (2005) forthe p53/Mdm2 dynamics when the same amountof DNA damage is imposed (see fig. 5). Itis worth noting the simulated results are in al-most perfect accordance not only with the numer-ical results of Ciliberto, Novak and Tyson (2005)but also with experimental data of Bar-Or, Maya,Segel, Alon, Levine and Oren (2000) and Ma,Wagner, Rice, Hu, Levine and Stlovitzky (2005)moreover, the other digital circuitry faithfully re-peats the p53/Mdm2 evolution of the model #1of Zhang, Brazhnik and Tyson (2007), as shownin fig. 6. These two numerical and biological“checkpoints” shown in figs 5 and 6 seem to benecessary to justify the utility of the present (dig-ital) approach.
Now, it should be noted that, in state-space theory,an applied digital wiring control scheme – basedon the extended form of ODEs (ordinary differen-tial equations) employed by Ciliberto, Novak andTyson (2005); Zhang, Brazhnik and Tyson (2007)– should be not applicable in the state-space it-
0 200 400 600 800 1000 12000
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
1.8
time (min)
[p53
tot],
[Mdm
2nuc
], [D
NA
dam
]
[p53tot][Mdm2nuc][DNAdam]
Figure 5: p53/Mdm2 pulses with DNA damage(Ciliberto, Novak and Tyson)
0 20 40 60 80 100 120 140-1
0
1
2
3
4
5
6
time (hours)
[p53
], [M
dm2n
uc],
[DN
Ada
m]
[p53][Mdm2nuc][DNAdam]
Figure 6: p53/Mdm2 pulses with DNA damage(model #1 Zhang, Brazhnik and Tyson)
self. This is due to the presence in the state-vector– as components – of both the p53 forms andits derivatives. To overcome this inconvenience,one must consider in the proposed mathematicalmodels (see Ciliberto, Novak and Tyson (2005);Zhang, Brazhnik and Tyson (2007)) the absenceof relationships among the first-derivatives on theright-hand side of the equations and those put onthe left-hand side. Let us now describe in detailthe procedures employed.
3 Optimal digital control
Before of describing the advantages of applyingthe digital optimal control matrix, one has alsoto check the reliability of the built up digital cir-
Digital control circuitry of cancer cell and its apoptosis 181
cuitries according to the mathematical aspects ofthe adopted models and their translation into thestate-space theory.
A) Model of Ciliberto, Novak and Tyson – Tak-ing into account their system of non-linear/firstorder differential equations (ODEs), one can de-scribe both p53 and Mdm2 dynamics as well asthe evolution of their forms. This mathematicalapproach is capable of deducing the mutual influ-ence among these concentrations. The digital cir-cuitry shall be able to reproduce the experimentalbasis of this model, i.e.:
a) Mdm2 and p53 are mainly degraded in thecell nucleus; b) Mdm2 is the activator of a re-action for degrading p53 in a ubiquitin-manner;c) Mdm2 attaches only two ubiquitins of p53(p53U and p53UU ); d) three forms of p53 (p53U ,p53UU , and p53tot) induce transcription of Mdm2in nonphosphorilated and cytoplasmic forms; e)for translocating into the nucleus, Mdm2cyt needsto be phosphorilated (Mdm2cyt→Mdm2Pcyt); f)the phosphorilated cytoplasmic Mdm2Pcyt movesfreely into and out of the nucleus; g) phosphorila-tion of Mdm2cyt is inhibited by p53tot in looping.
In the present study, to achieve the optimal con-trol scheme – based on an LQR-type (LinearQuadratic Regulator) kernel – a suitable manip-ulation of the previous set of ODEs has been donefor writing and using them from the extendedform into the state-space representation.
Following this procedure, a dynamic matrix rep-resentation of the p53/Mdm2 system is obtainedin which the time-dependent and mutual influenceof these proteins can be fully described. Fromnow on, the inner mechanisms of degradation offorms of cell proteins and/or phosphorilation aremarginally mentioned, the purpose of the presentwork being a global mathematical procedure todesign a cellular circuitry and the post-processingof the output.
In agreement with to but differently from Cilib-erto, Novak and Tyson (2005), in the present pro-cedure, a non linear ODEs system has been em-ployed to write the set of equations with respect tothe p53 protein and not to the total p53. In moredetail, the state-vector for p53 and its concentra-
where the overdot represents the time-derivativeand the subscripts U , UU identify the first-, thepoly-ubiquitin protein forms, respectively; whilethe vector
b =[ks53 0 0 0 0 0
]Tcontains the coefficient of Ciliberto, Novak andTyson (2005). The vector b is defined once thestate-space representation is implemented for de-scribing the input-state relationships.
We consider the level of [Mdm2nuc(t)] as a timevariable and a matrix P for the dynamic activityof p53 (time-variant). Mdm2 dynamics can be ex-pressed by a vector in the state-space.
Once the proposed state-space representationgives the same results as the model of Ciliberto,Novak and Tyson (2005), the digital optimal con-trol law has been implemented and based on theassumption that the matrix is such that [Mdm2nuc]is equal to a constant. Then, the compact expres-sion
P ={
Pi j; i, j = 1,2,3 so that pi j ∈ℜ
}represents the time-invariant dynamic super-matrix. Using the digital scheme, several simu-lations have been performed and they allowed theidentification of the constant value of [Mdm2nuc]equal to 0.1 in such a way as to obtain a rate ofDNA repair much more quickly than was obtainedby Ciliberto, Novak and Tyson (2007).
B) Model #1 of Zhang, Brazhnik and Tyson – Atfirst, the question formulated in this work seemsto be appropriate, i.e., is a negative feedback loop(p53 upregulates Mdm2, which deactivates p53)sufficient to explain the observed oscillation?
In fact, from a purely mathematical point of view,the feedback becomes positive if it is consideredas a part of a complete typical digital system. In-stead, if one looks at the single state-vector quan-tities (in feedback) containing all the forms ofp53, then the feedback loop becomes “hybrid”(positive - negative) in itself, because any element
of the control matrix has opposite sign with eachother. In a schematic representation, one has thecontrol matrix as follows
K =
+k11 −k12 +k13... +k22 −k23
sym · · · +k33
; ki j > 0
The optimal control matrix is derived from thestabilizing solution of the Riccati equation as afunction of the dynamic matrix of the protein p53whose elements are the reaction coefficients ofCiliberto, Novak and Tyson (2005). The aboveki j matrix elements are, in turn, functions of thosecoefficients.
In the model #1 of Zhang, Brazhnik and Tyson(2007), Mdm2 activates p53. Combined withp53-induced Mdm2 transcription, Mdm2 therebyenhances its own synthesis. The values of sta-ble steady state concentrations are obtained oncethe level of the DNA damage is set equal to zero.When this level is different from zero, the degra-dation of nuclear Mdm2 increases and its concen-tration begins to fall. The interesting result of thismodel is that if the damage level is quickly re-paired, the p53/Mdm2 control system develops asingle-pulse response to repair the DNA damageitself. The second pulse occurs if the level of theDNA damage is relatively high. Also in this case,the designed digital circuitry faithfully reproducesthe results as shown in fig. 6.
C) Apoptosis – Zhang, Brazhnik and Tyson(2007) show a wiring diagram of apoptosis andapply it to their model #1 (see fig. 2 of Zhang,Brazhnik and Tyson (2007)). Although the abovementioned models (Ciliberto, Novak and Tyson(2005); Zhang, Brazhnik and Tyson (2007)) givedifferent evolutions of the DNA damage repair-ing, digital control theory in state-space can unifythese models on the basis of the developed controlmatrix. Indeed, a well-suited optimal control lawallows any DNA damage evolution to be assignedto the realized human single-cell circuitry. Then,we take into account the DNA damage level andshape proposed by Zhang, Brazhnik and Tyson(2007) applied to the model of Ciliberto, Novakand Tyson (2005) once it is connected to a wiringdiagram of apoptosis (see figs 1 and 4).
4 Results and discussions
Once the reliability of the adopted cellular digi-tal circuitries has been tested apart from the signof the feedback loop, thanks also to the dynamiccontrol matrix, we study its effects on the proteinssystems of the p53/Mdm2 network in presence ofa DNA damage.
Here, we consider the running digital control pro-cess for the previous selected models of Ciliberto,Novak and Tyson (2005); Zhang, Brazhnik andTyson (2007) and evaluate the p53 dynamics ininteraction with the other protein parameters andthe DNA damage levels. All the recursive rou-tines and circuitries were processed using Mat-lab/Simulink platforms at the Department of Me-chanical Engineering – University of Bath (UK) –under the supervision of doctor Michael Carley.
A) Model of Ciliberto, Novak and Tyson – Look-ing at the simulation output when the digital con-trol matrix is switched “on”, it has been roughlynoted that the p53tot and Mdm2nuc levels increasethroughout the same timescale (see fig. 7) whencompared to the results having the digital controlmatrix switched “off”; since high levels of p53tot
involve a decrease in the DNA damage, we nowswitch the [Mdm2nuc] level to its lowest value insuch a way as to obtain a positive feedback loopaccording to the digital control theory rules.
0 200 400 600 800 1000 12000
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
1.8
time (min)
[p53
tot],
[Mdm
2nuc
], [D
NA
dam
]
[p53tot][Mdm2nuc][DNAdam]
Figure 7: p53/Mdm2 pulses with DNA damage(controlled)
Surprisingly, the results change deeply. Digitaloptimal control is then able to realize remarkable
Digital control circuitry of cancer cell and its apoptosis 183
0 200 400 600 800 1000 12000
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
1.8
time (min)
[DN
Ada
m]
present methodCiliberto et al.
Figure 8: DNA repair comparison
effects: first of all, a DNA damage repair speedfaster than 50% (see fig. 8) and – in cascade– a relevant variation of p53/Mdm2nuc dynam-ics. From the comparison between figures 5 and7, one may deduce that the oscillation parame-ters of p53/Mdm2 have been modified in terms ofamplitudes (higher concentrations) and interpulse(time-shift in the second pulse of about 40min).The initial conditions of Ciliberto, Novak andTyson (2005) being equal, the faster rate of DNAdamage repair can strongly affect the response ofthe whole p53/Mdm2nuc dynamics. Moreover, theoptimal control matrix is able to output sustainedamplitudes of both p53 and Mdm2nuc values.
The dual action of the control matrix has dif-ferent damping effects: these are more evidentfor the second pulse of p53 and almost irrelevantfor Mdm2. This different behaviour in the sec-ond pulse is linked to the local gradient of theDNA damage repair pathway; while in the firstpulse, the difference – between the uncontrolledand controlled models – of the DNA damage lev-els is very high, the local pathways and then thegradients are quite superimposable up to 220min.When the second pulse triggers, the DNA localdamage levels are totally different in shape (gra-dient) and amplitudes. In the uncontrolled digi-tal system, the gradient is confined inside a rangeof 80min, while in the controlled one, the rangebecomes wider up to 120min. In any case, as re-gards the p53/Mdm2nuc system, the action of thecontrol matrix implies an overall effect, i.e., an
amplifying of concentrations scattered along thesame timescale. Also, the global interpulse ofthe p53/Mdm2 network is shifted; from a com-parison between figures 5 and 7, one can deducethat its first pulse triggers at the same time in-terval (200÷240min); while, in the digital con-trol system, the second pulse occurs after a delayof 600+48min. Both the uncontrolled and con-trolled digital circuitries show in-phase oscilla-tions of the single protein forms, the frequencyof the [p53tot /Mdm2nuc]uncontr system being equalto 3.78×10−52.5×10−3Hz and 3.3×10−5Hz for[p53tot /Mdm2nuc]contr network.
The use of appropriate cell digital machinery re-veals interesting perspectives for future biologyspin-off. Two of them seem to be easily demon-strated, i.e., robustness and generalization. Accu-racy of results, freedom in choosing any other cellproteins forms, genotypes and/or target genes, cir-cuitry components manipulations and low compu-tational resources yield reason enough for theseconclusions.
B) Model #1 of Zhang, Brazhnik and Tyson – Asmentioned before, this model has been convertedinto a digital platform circuitry for two reasons:one, to check the reliability of the cell digital ma-chinery and, second, for the proposed apoptosiswiring diagram shown and linked by those authorsto this model. As mentioned before – in the lightof the interchangeability of the previous digitalcontrol matrix – we adopt this model and its pro-tein forms to analyse the response of the systemwhen an apoptosis wiring diagram is considered.In this model, Mdm2 activates p53 and, in positivefeedback, p53 and Mdm2cyt abruptly increase. Atthis point, an increasing quantity of Mdm2 mi-grates into the nucleus so giving degradation ofp53 and a decrease in its level. As a consequence,the Mdm2 rate decreases and, accordingly, theMdm2nuc level drops (see fig. 6). In this model,the initial DNA damage is repaired at a constantrate. When a wiring diagram of apoptosis is con-nected to this model, those authors define threeforms of p53 (helper, killer and lurker) and fol-low the evolution of these forms to elegantly de-duce and propose a digital apoptosis mechanismby numbering the p53 pulses. In agreement with
to but differently from those authors, we link theirapoptosis wiring diagram – digitally converted –to cell digital circuitry having a digital control ma-trix (see fig. 1).
C) Apoptosis – the decision on cell fate is nowconceived and bound through the modified puls-ing mechanism of p53 and other consideredspecies when the cellular digital optimal con-trol system is applied to an apoptosis wiring di-agram (see figs 1, 4 and 9). Following Zhang,Brazhnik and Tyson (2007), we take into accounttheir apoptotic model (see table 5 of their men-tioned work). In sequence, switching “on” and“off” the cell digital connection of the controlmatrix, the controlled (“on”) and uncontrolledfeedback (“off”) networks (see fig. 9) give re-sults concerning the time evolution of the apop-totic species (cyclin-dependent kinase inhibitor,p21; p53-regulated apoptosis-inducing protein 1,p53AIP1; apoptotic protease activating factor 1,APAF1; and target gene “cytoc” as a functionalof APAF1 in apoptosome expression), the valueof apoptosome parameter (expressed by the Heav-iside function) being equal to 1 for matching (ornot) the cell death. The species dynamics com-parison – both in the uncontrolled and controllednetworks – sketched in fig. 10 show remarkableinformation. First of all, instabilities of apop-totic species vanish in the digital control systemand, in sequence, variations of cell death param-eter in terms of timescale and triggering occur.In the uncontrolled system, the cell fate triggersat 124.61min and goes on for 32.3min, while inthe digital controlled network cell fate triggers at115.3min and remains for 27.7min. Moreover, in-teresting topics of discussion can be derived fromthe previous results when they are sketched allat once on the same timescale (see figs 11 and12). Both the uncontrolled and controlled sys-tems (although with different triggering and timerange) show apoptotic phase just in correspon-dence with the inflexion point of the consideredspecies pathways. A possible and common (forboth the networks) unification could be mathe-matically expressed: cell death triggers when thelocal gradients of the species (p21 being equal tozero) reach their maximum values at the same in-
stant; apoptotic phase remains until all the con-sidered species match their numerical maximumvalues; then p21 triggers. Once the local gradientof p21 reaches its transient maximum value, thecell death phase ends.
To sum up, we have reason to believe that the evi-dence of designing a cell digital platform could beuseful and representative in studying p53/Mdm2evolution in a cancer cell and its fate. This is afirst step. Limitations of the current digital designcan be found in its application to a single-cell andthe hypothesis of the considered number of apop-totic species (one has to remember that only somecases of Zhang, Brazhnik and Tyson (2007) matchcell death).
Nevertheless, those limitations can become fu-ture qualities when the proposed digital cell cir-cuitry is enhanced for multi-cellular systems andfor suitable feedback of the ionizing radiation sig-nal, which will be studied in future work.
5 Outcome reproducibility
Once the initial conditions of stable steady-stateare considered (see table 1), a digital control cir-cuitry design can be performed through the pro-tein [p53] state-space representation (or the equiv-alent [Mdm2] in feedback), i.e.:
x(t) = P([Mdm2nuc (t)]) x(t)+b (1)
z(t) =[[Mdm2nuc] [Mdm2cyt ] [Mdm2Pcyt ]
·[Mdm2nuc]
·[Mdm2cyt ]
·[Mdm2Pcyt ]
]T(2)
z(t) = M ([p53tot (t)] ,kd2 (t)) + c([p53tot (t)])(3)
c = [0 f ([p53tot (t)]) 0000]T (4)
where [Mdm2nuc (t)] defines the time-dependentoncogene concentration rate in nuclear form dur-ing the DNA repairing action; while, [Mdm2cyt ]and [Mdm2Pcyt ] are the time-dependent oncogeneconcentration rates in cytoplasmic and phospho-rilated forms, respectively; kd2 (t) is the rate con-stant for degradation of [Mdm2nuc (t)]; the time-dependent expression f ([p53tot (t)]) representsthe Hill function.
Digital control circuitry of cancer cell and its apoptosis 185
[DNAdam ] Dynamic behaviour
kd2 Variation
[Ionizing Radiation ]
Integrator
Integrator
control super matrix
signal end
direct link betweendigital control matrix and
p53 /Mdm2/DNA damage block
kd2
[P53 ]tot , [Mdm 2nuc ]
[P53 ] Vector Components
[Mdm 2cyt], [Mdm 2Pcyt ]
[Mdm 2] Vector Components
[DNAdam ]ARBAet
[DNAdam ]ARBA
[DNAdam ], kd2
[DNAdam ]
To Workspace
y
[DNAdam ] kd 2
[p53 tot ] u[Mdm 2]
Subsystem PSD
[P53 tot ]
Subsystem Ciliberto
[DNAdam ] Ciliberto
SubsystemApoptosis
[P53 ]
[P21 ]
[P53AIP 1]
[APAF 1]
[cytoc ]
[Apoptosome ]
[p53 tot ]
[ Ionizing Radiation ]
[DNAdam ]
[DNAdam ]
Signal 1
Signal 2
Signal 3
Scope
K
K* uvec
1s
1s
Dynamic System of [P53 ] Protein
[P53 ] State Vector
[Mdm 2nuc ]
[P53 ] Rate State Vector
Dynamic System of [Mdm 2] Protein
[Mdm 2] State Vector
[p53 tot ]
kd 2
[Mdm 2] Rate State Vector
emu
m
emu
[ks53 ;0;0;0;0;0]
Clock
BB
K*uvec
"0"vector
[0;0;0]
Figure 9: Human cell circuitry control matrix switched “off”
One has to note all the blacked quantities abovementioned are denoting time varying scalar quan-tities. Meanwhile, P and M identify the dynamicmatrices for the trascription factor p53 and theMdm2 oncogene, respectively. They are obtainedonce a state-space representation model has beencarried out by the extended expression of theODEs employed by Ciliberto, Novak and Tyson(2005).
The subscripts nuc, cyt and Pcyt identify nuclear,cytoplasmic and phosphorilated cytoplasmic pro-tein forms, respectively; while the vector (4) con-tains the known Hill function f previously usedby Ciliberto, Novak and Tyson (2005) (for the co-efficients, see tables 1-2).
More precisely, some auxiliary matrices havebeen employed as follows:
K = LQR(
P,BB,Q,R,)
(5)
where K is the digital optimal control law matrixobtained by resolving the Riccati’s equation, BBthe input-state transition matrix, while the posi-tive definite matrices Q and R are defined in sucha way their diagonal contains the setting valuesto obtain suitable control law for a desired pro-tein forms dynamics. The pre-multiplying factors
of the matrices Q and R have been imposed insuch a way that the pathways of the componentsof the state-vectors x(t), z(t)and the DNA dam-age are quite similar to those of Ciliberto, Novakand Tyson (2005), while the optimal control ma-trix has to accomplish the task to accelerate theDNA repair process according to the equation:
d [DNAdam]dt
= kDNA[IR]
− kdDNA [p53tot ][DNAdam]
JDNA +[DNAdam](6)
in which IR represents the functional of theimposed radiation dose; while kDNA, kdDNA,[DNAdam], JDNA represent the (direct) rate con-stant linked to ionizing radiation, the direct con-stant rate linking DNA damage to the rate oftranscription factor p53tot , the amount of dam-aged DNA and the Michaelis constant of p53tot-dependent DNA damage, respectively.
The system of differential equations obtainedmust be processed and resolved, i.e.:PT S +SP−
from the first equation of the above system, oneobtains the Riccati stabilizing solution S, and thesecond equation is then solved.
The optimal control matrix terms being a functionof the following parameters
K = ki j ∈ℜ/
ki j = f(qi j,ri j, [Mdm2nuc] ,k f ,k′d53,kd53,kr
)[Mdm2nuc] = const
qi j ∈ Q
ri j ∈ R
(8)
where k f , k′d53, kd53 and kr are the translation ratesof [Mdm2nuc], [p53tot ], [p53UU ] and the transla-tion rate of [p53UU ]-dependent [p53U ], respec-tively.
Easy mathematical manipulation of the equationof p53 yields:
ddt
[p53tot ] =ddt
[p53]+ddt
[p53U ]+ddt
[p53UU ]
(9)
and then one obtains:
ddt
[p53] =
− k′d53 [p53]−(
k′d53 [p53U ]+ddt
[p53U ])
−((
k′d53 + kd53)[p53UU ]+
ddt
[p53UU ])
ddt
[p53U ] = k f [Mdm2nuc] [p53]
−(k′d53 + kr + k f [Mdm2nuc]
)[p53U ]+kr [p53UU ]
ddt
[p53UU ] = k f [Mdm2nuc] [p53U ]
−(k′d53 + kd53 + kr
)[p53UU ] (10)
Now, if the optimal control law is applied, furtherterms belonging to the matrix K must be added,and then the extended form of the final system
of equations is obtained. Rearranging the system(10), one comes to:
ddt
[p53] =−(k′d53 + k f [Mdm2nuc]
)[p53]
+ kr [p53U ]+ ks53 +3
∑n=1
k1,nxn,1
ddt
[p53U ] = k f [Mdm2nuc] [p53]
−(k′d53 + kr + k f [Mdm2nuc]
)[p53U ]
+ kr [p53UU ]+3
∑n=1
k2,nxn,1 (11)
ddt
[p53UU ] = k f [Mdm2nuc] [p53U ]
−(k′d53 + kd53 + kr
)[p53UU ]+
3
∑n=1
k3,nxn,1
For the adopted values of the matrix elements, ki j,see table 2. Once the LQR has been performed,those elements were obtained by a linear combi-nation of the rate constants shown in table 1.
Acknowledgement: The authors wish to thankAndrea Ciliberto and Galit Lahav for their use-ful suggestions and the specific literary material.Marzia Sabella and Notre Dame du Cap de LaMadeleine made this work possible.
References
1. Ma L, Wagner J, Rice J-J, Hu W, Levine AJ,Stlovitzky G (2005) A plausible model forthe digital response of p53 to DNA damage.Proc Natl Acad Sci USA 40:14266-14271.
2. Geva-Zatorsky N, Rosenfeld N, Itzkovitz S,Milo R, Sigal A, Dekel E, Yarnitzky T, LironY, Polak P, Lahav G, Alon U (2006) Os-cillations and variability in the p53 system.Molec Syst Biol 10.1038:1-13.
3. Shangary S, Wang S (2008) Targeting theMdm2-p53 interaction for cancer therapy.Clin Cancer Research 17:5318-5324.
Digital control circuitry of cancer cell and its apoptosis 189
4. Ciliberto A, Novak B, Tyson J (2005) Steadystates and oscillations in the p53/Mdm2 net-work. Cell Cycle 3:488-493.
5. Zhang T, Brazhnik P, Tyson J-J (2007) Ex-ploring mechanisms of the DNA-damage re-sponse. Cell Cycle 1:85-94.
6. Harms K-L, Chen X (2005) The C terminusof p53 family proteins is a cell fate determi-nant. Mol Cell Biol 5:2014-2030.
7. Bell S, Klein C, Muller L, Hansen S, Buch-ner J (2002) p53 contains large unstruc-tured regions in its native state. J Mol Biol322:917-927.
8. Reich C-N, Oren M, Levine A-J (1983) Twodistinct mechanisms regulate the levels of acellular tumor antigen, p53. Mol Cell Biol12:2143-2150.
9. Ventura A, Kirsch D-G, McLaughlin M-E,Tuveson D-A, Grimm J, Lintault L, NewmanJ, Reczek E-E, Weissleder R, Jacks T (2007)Restoration of p53 function leads to tumourregression in vivo. Nature 445:661-665.
10. Bates S, Phillips A-C, Clark P-A, Stott F, Pe-ters G, Ludwig R-L, Vousden K-H (1998)p14ARF links the tumour suppressor RB andp53. Nature 395:124-125.
11. Bar-Or R-L, Maya R, Segel L-A, AlonU, Levine A-J, Oren M (2000) Generationof oscillations by the p53-Mdm2 feedbackloop: a theorethical and experimental study.Proc Natl Acad Sci USA 21:11250-11255.
12. Lahav G, Rosenfeld N, Sigal A, Geva-Zatorsky N, Levine A-J, Elowitz M-B,Alon U (2004) Dynamics of the p53-Mdm2 feedback loop in individual cells.Nature Genetics − published online(www.nature.com/naturegenetics) doi10.1038:1-4.
13. Loewer A, Lahav G (2006) Cellular confer-ence call: external feedback affect cell-fatedecision. Cell 124:1128-1130.
14. Batchelor E, Mock C-S, Bhan I, LoewerA, Lahav G (2008) Recurrent initiation: amechanism for triggering p53 pulses in re-sponse to DNA damage. Molecular Cell30:277-289.
15. Tsourkas A, Weissleder R (2004) Illumi-nating the dynamics of intracellular activitywith “active” molecular reporters. Molecu-lar and Cell Biomechanics 1:136-146.
16. Deguchi S, Ohashi T, Sato M (2005) In-tracellular stress transmission through actinstress fiber network in adherent vascularcells. Molecular and Cell Biomechanics2:205-216.
17. Zhou J, Chen J-K, Zhang Y (2007) Theoreti-cal analysis of thermal damage in biologicaltissues caused by laser irradiation. Molecu-lar and Cell Biomechanics 4:27-40.
18. Mooney D (2006) Materials to regulate cellfate in vitro and in vivo. Molecular and CellBiomechanics 3:169-176.
19. Ardito Marretta R-M, Marino F (2007)Wing flutter suppression enhancement usinga well-suited active control model. J AeroEngng 221:441-453.
20. Ardito Marretta R-M, Marino F, Bianchi P(2008) Computer active control of damp-ing fluid of a racing superbike suspensionscheme for road-safety improvement spin-off. Int J Vehicle Des 46:436-450.