The Behavior Analyst 1995, 18, 51-68 No. 1 (Spring) Tutorial Stimulus Control: Part I James A. Dinsmoor Indiana University In his effort to distinguish operant from respondent conditioning, Skinner stressed the lack of an eliciting stimulus and rejected the prevailing stereotype of Pavlovian "stimulus-response" psy- chology. But control by antecedent stimuli, whether classified as conditional or discriminative, is ubiquitous in the natural setting. With both respondent and operant behavior, symmetrical gradients of generalization along unrelated dimensions may be obtained following differential reinforcement in the presence and the absence of the stimulus. The slopes of these gradients serve as measures of stimulus control, and they can be steepened without applying differential reinforcement to any two points along the test dimension. Increases and decreases in stimulus control occur under the same conditions as those leading to increases and decreases in observing responses, indicating that it is the increasing frequency and duration of observation (and perhaps also of attention) that pro- duces the separation in performances during discrimination learning. Key words: stimulus, discrimination, generalization, gradients, observing In the analysis of behavior, a great deal of emphasis has been placed on the control of responding by stimuli that follow the response (e.g., reinforc- ing stimuli), but comparatively little at- tention has been given to control by stimuli that precede the response (e.g., discriminative stimuli). Control by stimuli that are already present before the response occurs plays a much more significant role in everyday life than it is currently accorded in our research or even in our textbooks. It may be that the neglect of this top- ic can be traced back to Skinner's ef- forts to distinguish between the type of conditioning that he had begun to study in the laboratory and the older type that had previously been studied by Pavlov (Skinner, 1937, 1938). When a dog was to be trained to sali- vate to a conditional stimulus, as in Pavlov's laboratory (Pavlov, 1927/ 1960), the response was already under a form of stimulus control. In a hungry dog, salivation could regularly and re- liably be induced by presenting some dried biscuit or meat powder or a mild Send correspondence to James Dinsmoor, De- partment of Psychology, Indiana University, Bloomington, Indiana 47405. solution of acid. No prior training was necessary. Because few special condi- tions or restrictions seemed to be at- tached to this method of eliciting sali- vation, Pavlov referred to the stimulus as an unconditional stimulus, the re- sponse to that stimulus as an uncondi- tional response, and the relation be- tween the two as an unconditional re- flex. When some other stimulus, like a light or a tone, was repeatedly present- ed shortly before the meat powder, the dog began to salivate to the new stim- ulus prior to the delivery of the food. As the effectiveness of this stimulus was conditional upon the pairing pro- cedure, Pavlov called the new stimulus a conditional stimulus, the new re- sponse a conditional response, and the new reflex a conditional reflex. (In En- glish translations, the words "condi- tioned" and "unconditioned" have of- ten been used, but the original meaning is clear; see Pavlov, 1927/1960, p. 5.) Furthermore, because in this type of conditioning the food was paired with a stimulus, Skinner gave it the label Type S conditioning. When it was bar pressing that was to be conditioned, however, there was no unconditional stimulus available to 51
Transcript
The Behavior Analyst 1995, 18, 51-68 No. 1 (Spring)
TutorialStimulus Control: Part I
James A. DinsmoorIndiana University
In his effort to distinguish operant from respondent conditioning, Skinner stressed the lack of aneliciting stimulus and rejected the prevailing stereotype of Pavlovian "stimulus-response" psy-chology. But control by antecedent stimuli, whether classified as conditional or discriminative, isubiquitous in the natural setting. With both respondent and operant behavior, symmetrical gradientsof generalization along unrelated dimensions may be obtained following differential reinforcementin the presence and the absence of the stimulus. The slopes of these gradients serve as measuresof stimulus control, and they can be steepened without applying differential reinforcement to anytwo points along the test dimension. Increases and decreases in stimulus control occur under thesame conditions as those leading to increases and decreases in observing responses, indicating thatit is the increasing frequency and duration of observation (and perhaps also of attention) that pro-duces the separation in performances during discrimination learning.Key words: stimulus, discrimination, generalization, gradients, observing
In the analysis of behavior, a greatdeal of emphasis has been placed onthe control of responding by stimulithat follow the response (e.g., reinforc-ing stimuli), but comparatively little at-tention has been given to control bystimuli that precede the response (e.g.,discriminative stimuli). Control bystimuli that are already present beforethe response occurs plays a much moresignificant role in everyday life than itis currently accorded in our research oreven in our textbooks.
It may be that the neglect of this top-ic can be traced back to Skinner's ef-forts to distinguish between the type ofconditioning that he had begun tostudy in the laboratory and the oldertype that had previously been studiedby Pavlov (Skinner, 1937, 1938).When a dog was to be trained to sali-vate to a conditional stimulus, as inPavlov's laboratory (Pavlov, 1927/1960), the response was already undera form of stimulus control. In a hungrydog, salivation could regularly and re-liably be induced by presenting somedried biscuit or meat powder or a mild
Send correspondence to James Dinsmoor, De-partment of Psychology, Indiana University,Bloomington, Indiana 47405.
solution of acid. No prior training wasnecessary. Because few special condi-tions or restrictions seemed to be at-tached to this method of eliciting sali-vation, Pavlov referred to the stimulusas an unconditional stimulus, the re-sponse to that stimulus as an uncondi-tional response, and the relation be-tween the two as an unconditional re-flex.When some other stimulus, like a
light or a tone, was repeatedly present-ed shortly before the meat powder, thedog began to salivate to the new stim-ulus prior to the delivery of the food.As the effectiveness of this stimuluswas conditional upon the pairing pro-cedure, Pavlov called the new stimulusa conditional stimulus, the new re-sponse a conditional response, and thenew reflex a conditional reflex. (In En-glish translations, the words "condi-tioned" and "unconditioned" have of-ten been used, but the original meaningis clear; see Pavlov, 1927/1960, p. 5.)Furthermore, because in this type ofconditioning the food was paired witha stimulus, Skinner gave it the labelType S conditioning.When it was bar pressing that was
to be conditioned, however, there wasno unconditional stimulus available to
51
52 JAMES A. DINSMOOR
elicit the response. Insertion of the lev-er into the animal's chamber was a nec-essary but not a sufficient condition forpressing to occur. The experimenterhad to wait for the animal to performthe desired action before the pellet offood could be delivered as a reinforcer.Because in this case the food waspaired with a response, Skinner calledit Type R conditioning.
Skinner also distinguished two typesof behavior that corresponded to thetwo types of conditioning. Obviously,Pavlov's procedure could be appliedonly to behavior that could be elicited,prior to the training, by a specific stim-ulus. Because it occurred in responseto a stimulus, Skinner called this typeof behavior respondent. But for behav-ior like bar pressing, no equivalentstimulus could be found. Because itoperated on the surrounding environ-ment to produce the reinforcing con-sequence, he called this form of behav-ior operant. At the beginning of train-ing, then, respondent behavior wascharacterized by an eliciting stimulusand operant behavior by its absence.During the initial process of condition-ing, the only stimulus that seemed tobe of any importance for operant be-havior was the one that came after theresponse and caused it to be repeatedon future occasions (reinforcing stim-ulus).
Although contemporary evidencesuggests that Pavlovian conditioning isbroad in its application and complex inits functional relations (e.g., Rescorla,1988), this was not commonly recog-nized in the early years of Skinner'swork. Perhaps it was to contrast hisviews with the prevailing stereotype ofPavlovian conditioning and to insulatehimself against the charge of adoptingan overly atomistic approach that Skin-ner rejected the label of a stimulus-re-sponse psychologist. In his later writ-ings he continued to stress the rele-vance of other factors to operant be-havior and to downplay the importanceof the stimulus (e.g., preface to the1966 printing of The Behavior of Or-ganisms).
Nonetheless, even in the 1937 paperon the two types of conditioning hefound it necessary to note that "it isthe nature of [operant] behavior ...that after the initial conditioning "dis-criminative stimuli are practically in-evitable" (1972, p. 491). And in a nat-ural setting stimulus control is alwayspresent. It can be seen at any time,anywhere we look. All behavior is un-der the exquisitely detailed control ofsurrounding stimuli, some impingingfrom outside the organism, others aris-ing from within its boundaries.
Although it is not always obvious inthe way we talk and write about thesubject, behavior does not occur asrandom strings of unrelated responsesbut in organized sequences, calledchains, in which each successive re-sponse produces the stimuli, internal orexternal, that determine what comesnext. And when the time arrives to ini-tiate a new sequence of behavior, that,too, is signaled by a change in stimulusconditions.
If we were to monitor the stream ofbehavior issuing from a single individ-ual, moment by moment throughoutthe day, control by antecedent stimuliwould be much easier to identify thancontrol by the organism's history of re-inforcement. One form of control liesin the present, the other in the past.Another way of describing the relationbetween the two forms of control is tosay that stimulus control is a transmis-sion device: It is nature's way of bring-ing past reinforcement to bear on cur-rent behavior. To highlight the pointone more time, ask yourself how youwould arrange for someone seated nextto you to pass the salt. Would you re-inforce successive approximations, orwould you present an appropriate stim-ulus?
DISCRIMINATION TRAINING
Although Skinner was not the firstperson to study the formation of a dis-crimination, it was his work, more thanthat of any other person, that broughtthe process into experimental focus.
STIMULUS CONTROL 53
The beauty of Skinner's approach layin its simplicity. He stripped the pro-cess of discrimination down to its bareessentials. The minimum requirementswere two stimuli, so that there wassome difference to discriminate, and asingle response, which could vary inits probability or some other character-istic. In his initial experiment, Skinner(1933, 1938) turned on a light insidethe conditioning chamber and waitedfor the rat to press the lever. When itdid so, Skinner reinforced the responseby delivering a pellet of food. Next, heturned the light off for 5 min, duringwhich time no more presses were re-inforced. The cycle was then repeated,with reinforcement of a single responseeach time the light was turned on andsubsequent extinction in the dark.Eventually, the rat was observed topress the bar promptly whenever thelight came on but not to press very of-ten when the light was not there. Todemonstrate that the relation betweenthe stimulus and the response was aproduct of the experimental contingen-cies and not of some intrinsic connec-tion between a particular stimulus andthe pressing response, Skinner alsotrained some of his animals on the re-verse discrimination, reinforcing thepressing that occurred in the dark butnot the pressing that occurred in thelight. In accord with this reversal of therelation between stimulus and reinforc-er, the rats learned to press the leverwhen in the dark but not when in thepresence of the light.
Discrimination in Applied Settings
To develop the ability to apply theseconcepts to new situations, it is nec-essary to put aside the concrete stimuliand responses used in the conditioninglaboratory and to concentrate on some-thing more abstract, the pattern ofevents. It is the relations among thestimuli and the responses that defineconcepts like discrimination training,observing, and stimulus generalization.The process of learning to identifythese patterns in new settings may be
facilitated by the consideration of someillustrative examples.
In 1941, when the United States en-tered World War II, the Army SignalCorps was faced with the task of train-ing a large number of new recruits asquickly as possible to receive and tosend messages in Morse code. That is,the trainees had to learn to transcribesequences of long and short auditorysignals (commonly represented in printby dashes and dots) into letters of thealphabet and to produce such sequenc-es by operating a switching device.Called upon to devise an efficienttraining program, Fred S. Keller, him-self a former telegrapher, recognizedthat learning to read the incoming sig-nals was a problem in discriminationtraining. Ultimately, the trainees had torespond rapidly and accurately to anumber of auditory patterns thatsounded very much alike to the un-trained ear. He proposed a techniqueknown as the "code-voice method"(Keller, 1943), which was based di-rectly on the procedure Skinner hadused with the albino rat.Up to that time, telegraph and radio-
telegraph operators had begun theirlearning of Morse code by memorizingthe alphabetical equivalents of the se-quences of dashes and dots publishedin visual form, but Keller suspectedthat this form of training did little tohelp the learner discriminate amongthe different patterns of sound. He be-gan immediately with the stimuli to bediscriminated.As is common when laboratory pro-
cedures developed with other animalsare adapted for our highly verbal spe-cies, Keller trained his students on anumber of responses (i.e., a number ofdifferent letters of the alphabet) at thesame time. After a given signal hadbeen presented through earphones orover a loudspeaker system, the traineeentered his (in those days, the traineeswere all male) best guess as to the let-ter or digit it represented in a row ofboxes on a standardized answer sheet.Shortly afterward, the instructor pre-sented the correct response, coded as
54 JAMES A. DINSMOOR
Able, Baker, Charlie, and so on. If thecorrect answer matched what the train-ee had just written down, the fact thathis response was correct presumablyreinforced that response. Note that thiswas an immediate reinforcement, act-ing upon a specific response to a spe-cific signal, as contrasted with the usu-al educational practice of posting anoverall score, which does not pinpointindividual stimuli or individual re-sponses, on the following day.
If the trainee had not been able tocome up with the correct answer, henow entered it in a second row of box-es, below that reserved for his originalresponses. In this case, it still provideda prompt that might help him on thenext presentation of the same signal.(This effect was not suggested by Skin-ner's work but nevertheless appears tohave empirical support, at least forbrief intervals between prompt andtest; Buchwald, 1969.)From learning to translate sequences
of long and short sounds into letters ofthe alphabet, it is but a short jump con-ceptually to learning to translate print-ed letters and groups of letters into thesounds of speech. By the time we be-gin learning to read, most of us are fac-ile speakers of our native tongue; thatis, the responses are well established,and all that is needed is to place theseresponses under the control of the vi-sual stimuli. Even in English there is asubstantial correspondence between in-dividual letters and specific phonemes,so that the learner can often sound outthe complete word, letter by letter, untilthe resemblance is recognized betweenthe assembled fragments and the spo-ken word. At this stage, the learnermay experience difficulty in distin-guishing between similar-appearingletters like b and d or p and q. ("Mindyour ps and qs." )
Training is also given in recognizingthe word as a whole. I can still remem-ber the difficulty experienced by myelder son, who was trained exclusivelyby the whole word method, when theword "jump" appeared for the firsttime at the beginning of a sentence.
Because the word began with a prom-inent capital J, he identified it as theonly word he had previously seen withsuch a configuration, the proper name"Jack."At a higher level of complexity, re-
sponding in accord with verbal instruc-tions is also an example of the controlof behavior by discriminative stimuli.The pigeon can readily learn this skillat an elementary level if pecking is re-inforced in the presence of the stimulus"peck" displayed on the key but is notreinforced in the presence of the stim-ulus "don't peck." In applied situa-tions, a series of stimuli may be em-ployed to guide a sequence of re-sponses. Or verbs and grammatical ob-jects, for example, may be presented innovel combinations, leading to the pro-duction of novel patterns of behavior.This transfer of actions to different ob-jects and similar features of humanverbal behavior may underlie the in-tuitive distinction the beginning stu-dent is apt to draw between "under-standing" the instructions in the hu-man case and "simple conditioning"in the case of the pigeon. But funda-mentally, both illustrate the same pro-cess.
Intermittent Reinforcement inDiscrimination Training
Skinner's technique for studying theformation of a discrimination provideda useful model for things that happenin a natural setting, but from a techni-cal point of view it left something tobe desired. When the behavior in thepresence of the positive stimulus wasmeasured in terms of the time that ittook before the rat pressed the lever(latency) and the behavior in the pres-ence of the negative stimulus was mea-sured in terms of the slope of the cu-mulative record (rate of pressing),there was no way to compare the sub-ject's reactions to the two stimuli.Without a common measure, there wasno way to calculate a single index thatcould express the relation between thetwo performances as the discrimination
STIMULUS CONTROL 55
developed. Even a rate of respondingin the positive stimulus would not bevery revealing if obtained under con-tinuous reinforcement, because itwould be determined mainly by thetime required by the animal to con-sume the successive pellets of food. Toprovide a rate that could be comparedwith the rate in the presence of the neg-ative stimulus, it was necessary to re-sort to an intermittent schedule of re-inforcement (Dinsmoor, 195 1). Then,by dividing the positive rate by thesum of the two rates, for example, anindex could be calculated that wouldbe independent of variations in theoverall level of activity (Dinsmoor,1952). Intermittent reinforcement alsoproduced a response that persisted fora long time without further reinforce-ment, making it useful for comparisonsof the subject's response to differentvalues along some stimulus dimension,as in studies of generalization (Gutt-man & Kalish, 1956). From the time ofthese experiments, most studies ofstimulus control have used alternatingperiods of variable-interval reinforce-ment in the presence of one stimulusand nonreinforcement in the presenceof the other.
Respondent Conditioning as aForm of Discrimination Training
There is a close and interesting par-allel between the procedure Pavlovused to train his dogs to salivate to aconditional stimulus and the procedureSkinner used to train his rats to form adiscrimination. First, consider Skin-ner's procedure. A light (or some otherstimulus) is presented. Skinner calledthis a discriminative stimulus. The ratresponds by pressing the lever. Next, apellet of food is delivered as a rein-forcer. Then a much longer period fol-lows, in which the positive stimulus isabsent and there are no more deliveriesof food. Skinner treated this period asone of negative stimulation and the dif-ference in the rat's behavior as an in-dication of its discrimination betweenthe two stimuli.
Now consider the procedure used inPavlov's laboratory. A light (or otherstimulus) is presented. Pavlov calledthis a conditional stimulus (CS), but itbears the same relation to the overallpattern of events as Skinner's discrim-inative stimulus. The dog responds bysalivating. Next, food is delivered as anunconditional stimulus (the reinforcingagent in this type of conditioning).Then a much longer period follows, inwhich the conditional stimulus is ab-sent and no more food is delivered.Pavlov treated this period simply as aninterval of time between successivepresentations of the conditional stimu-lus, but clearly it is the same thing thatSkinner treated as a period of negativestimulation. Given the basic differencethat distinguishes the two types of con-ditioning-pairing the food with a re-sponse or pairing it with a stimulus-the two procedures are as much alikeas they can possibly be. The similarityis also preserved in more recent work:The critical difference between auto-shaping (a very popular Pavlovian pro-cedure) and operant discriminationtraining based on an intermittentschedule is that in the operant casepecking is required in the presence ofthe positive stimulus to produce a de-livery of the grain, whereas in auto-shaping the grain is delivered at theend of the stimulus period, regardlessof whether any pecking has occurred.The conclusion seems inescapable thatconventional Pavlovian conditioningprocedures are necessarily and inher-ently to be categorized as forms of dis-crimination training.
It is not easy to illustrate the differ-ential performance that develops understandard Pavlovian conditioning pro-cedures, because experimenters work-ing with those procedures do not or-dinarily record the behavior of theirsubjects in the absence of the condi-tional stimulus. Like Pavlov, they viewthat part of the experimental routinesimply as an interval between trialsand take it for granted that nothing ofany consequence is happening there.But Pavlov himself noted that
56 JAMES A. DINSMOOR
when conditioned reflexes are being establishedin dogs for the first time, it is found that thewhole experimental environment, beginningwith the introduction of the dog to the experi-mental room, acquires at first conditioned prop-erties.... Later on, when the special reflex to asingle definite and constant stimulus has ap-peared, all the other elements of the environmentgradually lose their special conditioned signifi-cance. (Pavlov, 1927/1960, p. 115)
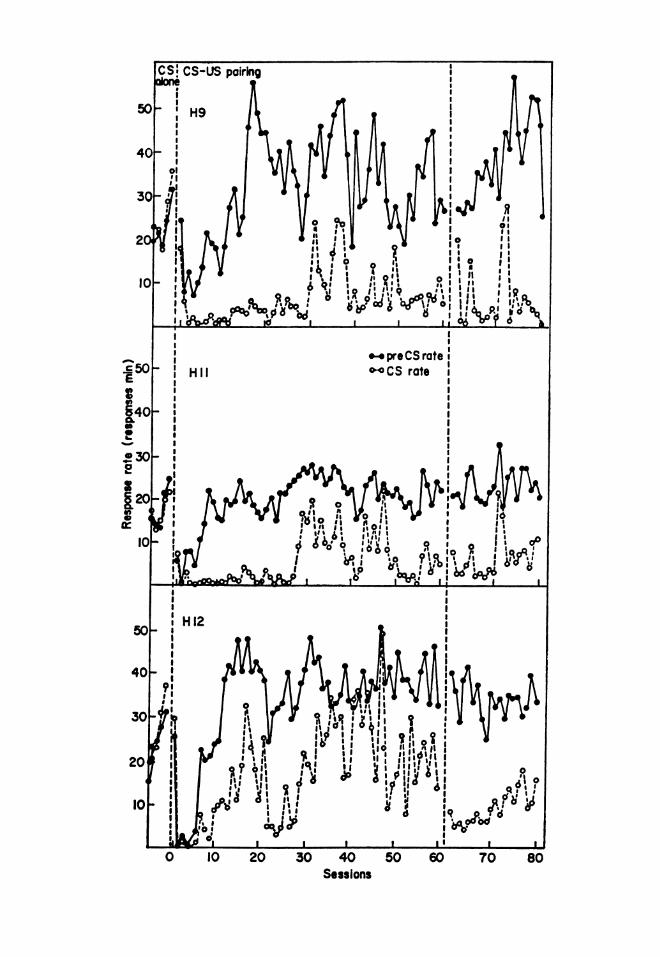
Moreover, some quantitative datashowing the formation of a discrimi-nation between the presence and theabsence of the CS have been madeavailable in a study conducted by Mil-lenson and Dent (1971). With the tech-nique known as conditioned suppres-sion, a baseline rate of pressing a leveror pecking a key is maintained by avariable-interval schedule of nutritivereinforcement. The unconditional stim-ulus is a brief pulse of electric shockapplied at the end of a period of ex-posure to a light or a tone, for example,and the reaction that is monitored (con-ditional response) is a reduction in therate of pressing the lever or peckingthe key in the presence of the light orthe tone (CS). To measure this reduc-tion, the experimenter compares therate in the presence of the conditionalstimulus with the rate during somecontrol period, which is necessarily aperiod when the conditional stimulus isnot present. Usually it is only somequantitative relation between the tworates (suppression ratio) that is report-ed as an index to the progress of theconditioning. In the Millenson andDent study, however, the authorstracked the absolute rates for bothstimuli, independently, for 80 sessionsof training. Their Figure 1 (p. 129; re-produced here) shows that when theelectric shock was first applied, bothrates immediately dropped to muchlower levels, but as training continued,the rate in the absence of the CS re-covered, until it was substantially high-
er than the rate in the presence of theCs.The divergence between the two
rates plotted in this study provides anexcellent illustration of the develop-ment of discriminative control duringPavlovian conditioning. Furthermore,the mere fact that the rate of respond-ing in the presence of the CS has beenreduced, in relation to the control rate,indicates that a similar discriminationhas occurred in other studies of con-ditioned suppression. The suppressionratio used in these studies to monitorthe level of conditioning is in effect anindex to the progress of the discrimi-nation, and the conditional stimulus isa discriminative stimulus. Notwith-standing Skinner's references to "elic-iting" in the one case and "setting theoccasion" in the other, the only dis-tinction seems to lie in the type of be-havior they control. (For a related dis-cussion, see Hearst, 1975.)
STIMULUS GENERALIZATION
One of the most important of the ob-servations coming out of Pavlov's lab-oratory was the discovery that when atactile stimulus had been used as theCS, application of that same stimulusto other places on the dog's anatomyalso elicited a certain amount of sali-vation. In addition, Pavlov noted thatthe nearer this stimulation was to thesite of the original CS, the greater wasthe amount of salivation that was ob-tained. Or, as another example, "If atone of 1000 d.v. [Hz] is established asa conditioned stimulus, many othertones spontaneously acquire similarproperties, such properties diminishingproportionally to the intervals of thesetones from the one of 1000 d.v." (Pav-lov, 1927/1960, p. 113). This declinein the magnitude of the response withincreases in the physical difference
Figure 1. For each of 3 rats, daily rates of pressing in the presence and the absence of a conditionalstimulus ending with electric shock. (Reproduced from Millenson & Dent, 1971, copyright Erl-baum.)
|~~~~~~~~~~- pro CS roteiC0 CSUS,pairHgCSrote
30
0~~~~~
I0L,I
6Pg .I9Ie0 1 e
11#oi'tL
lo 'H 'l I
0 10 20 30 40 50 60 70 80Sessions
58 JAMES A. DINSMOOR
from the original stimulus or increasein magnitude with decreases in thephysical difference-is what is meantby the gradient of generalization. Thegreater the similarity (closer along thephysical dimension), the greater the re-sponse; the less the similarity (fartheralong the physical dimension), thesmaller the response. Indeed, otherthings being equal, the degree of gen-eralization between two stimuli mayserve as a quantitative index to theirsimilarity.
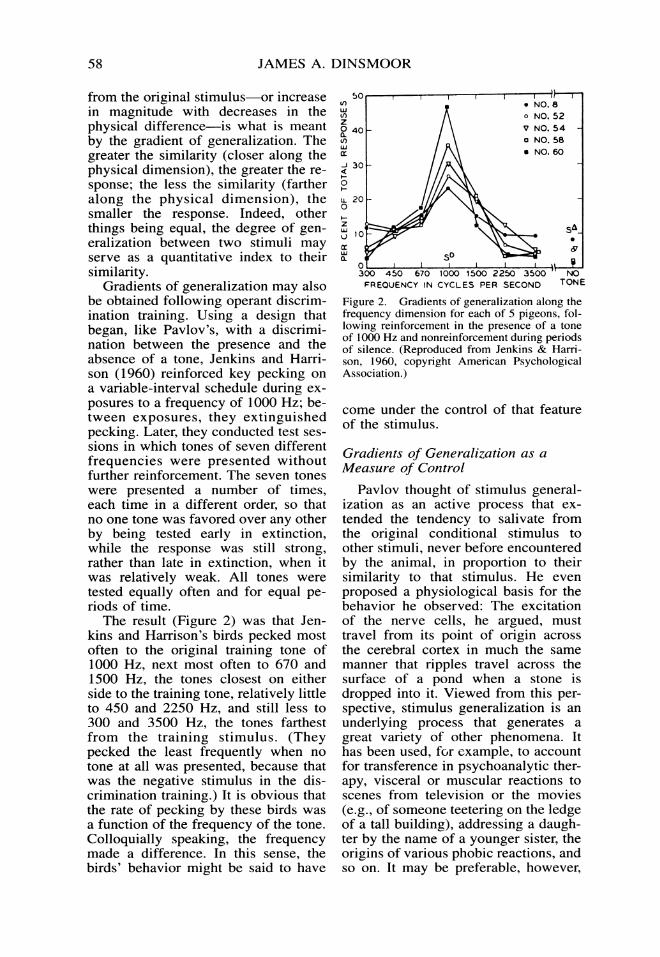
Gradients of generalization may alsobe obtained following operant discrim-ination training. Using a design thatbegan, like Pavlov's, with a discrimi-nation between the presence and theabsence of a tone, Jenkins and Harri-son (1960) reinforced key pecking ona variable-interval schedule during ex-posures to a frequency of 1000 Hz; be-tween exposures, they extinguishedpecking. Later, they conducted test ses-sions in which tones of seven differentfrequencies were presented withoutfurther reinforcement. The seven toneswere presented a number of times,each time in a different order, so thatno one tone was favored over any otherby being tested early in extinction,while the response was still strong,rather than late in extinction, when itwas relatively weak. All tones weretested equally often and for equal pe-riods of time.The result (Figure 2) was that Jen-
kins and Harrison's birds pecked mostoften to the original training tone of1000 Hz, next most often to 670 and1500 Hz, the tones closest on eitherside to the training tone, relatively littleto 450 and 2250 Hz, and still less to300 and 3500 Hz, the tones farthestfrom the training stimulus. (Theypecked the least frequently when notone at all was presented, because thatwas the negative stimulus in the dis-crimination training.) It is obvious thatthe rate of pecking by these birds wasa function of the frequency of the tone.Colloquially speaking, the frequencymade a difference. In this sense, thebirds' behavior might be said to have
50V) ' NO. 8zA 0 NO. 52zg 40 V NO. 54
U, a ~~~~~~~~NO.58cr a~~~~~~~~NO. 60
-J 30
0
u- 20 -ZO U, <z
10
0~~~~S300 450 670 1000 1500 2250 3500 NOFREOUENCY IN CYCLES PER SECOND TONE
Figure 2. Gradients of generalization along thefrequency dimension for each of 5 pigeons, fol-lowing reinforcement in the presence of a toneof 1000 Hz and nonreinforcement during periodsof silence. (Reproduced from Jenkins & Harri-son, 1960, copyright American PsychologicalAssociation.)
come under the control of that featureof the stimulus.
Gradients of Generalization as aMeasure of Control
Pavlov thought of stimulus general-ization as an active process that ex-tended the tendency to salivate fromthe original conditional stimulus toother stimuli, never before encounteredby the animal, in proportion to theirsimilarity to that stimulus. He evenproposed a physiological basis for thebehavior he observed: The excitationof the nerve cells, he argued, musttravel from its point of origin acrossthe cerebral cortex in much the samemanner that ripples travel across thesurface of a pond when a stone isdropped into it. Viewed from this per-spective, stimulus generalization is anunderlying process that generates agreat variety of other phenomena. Ithas been used, for example, to accountfor transference in psychoanalytic ther-apy, visceral or muscular reactions toscenes from television or the movies(e.g., of someone teetering on the ledgeof a tall building), addressing a daugh-ter by the name of a younger sister, theorigins of various phobic reactions, andso on. It may be preferable, however,
STIMULUS CONTROL 59
to think of these phenomena simply asillustrations of incomplete discrimina-tion, rather than as the products ofsome internal process. A favorite illus-tration is the printed stimulus recieve,which in many readers will evoke thesubvocal response "receive."
Clearly, generalization and discrim-ination are opposite numbers. As oneincreases, the other decreases, and viceversa. Although the standard textbookaccount follows Pavlov's conception ofgeneralization, presenting it as theproduct of a hypothetical process ofexcitation or inhibition spreading fromone stimulus to another, there is no rea-son to think of the two of them as con-tending forces battling for control ofthe individual's behavior. It makesgreater sense to think of them as alter-native descriptions-to think of theamount of generalization, or more spe-cifically the slope of the gradient, as anindex to the level of stimulus controlthat has been attained at a given pointin the subject's training. To illustratethis point, consider what would happenif a pigeon were surgically deafened.Because it would have no way of dis-tinguishing among tones of differentfrequency, it would peck at the samerate in the presence of all of them. Thegradient of generalization would be asbroad as possible (i.e., horizontal inslope). Similarly, if several differentvisual stimuli were presented outsideof the conditioning chamber, so thatthe bird could not see them, it wouldpeck at the same rate, regardless ofwhich one was present at a given mo-ment. The maximum amount of gen-eralization would be recorded, butthere is no reason to conjure up anykind of internal process to account forit. A high level of generalization mere-ly indicates a low level of discrimina-tion.
Generalization Decreases,Rather than Increases,with Training
If food is delivered in the presenceof a stimulus but not in its absence, the
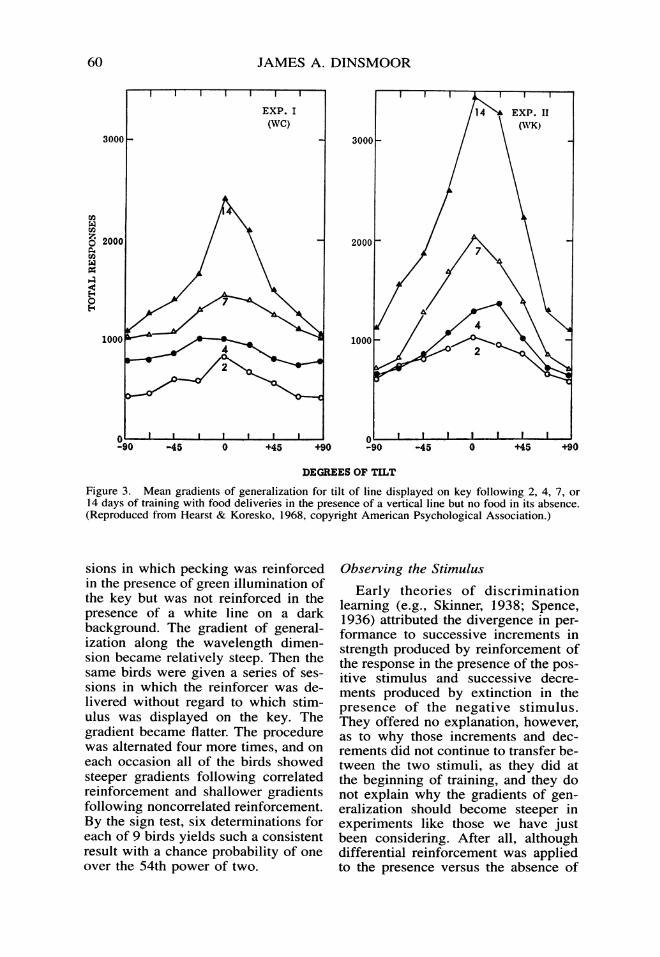
gradient of generalization about thatstimulus becomes narrower, rather thanbroader, as the training continues. Forexample, Hearst and Koresko (1968)plotted average gradients for differentgroups of pigeons after 2, 4, 7, or 14days of training. During the training, awhite line was projected from the rearon an otherwise darkened key for pe-riods of 30 s, interspersed with periodsof 10 s when nothing was projected onthe key. That is, the entire surface ofthe key was dark. During those periodswhen the line was displayed, peckingwas reinforced on a variable-intervalschedule, but when the key was com-pletely dark, no grain was delivered.Then the birds were tested with linesof eight different tilts, ranging from thevertical to the horizontal. After only 2days of training, a modest amount ofpecking occurred in the presence ofeach of the test stimuli, with relativelylittle difference among them (Figure3). The gradient was extremely broad.After longer periods of training, how-ever, the number of responses as afunction of position along the tilt di-mension began to bunch up at the val-ue used as the positive stimulus and todrop off more and more steeply towardthe sides. Generalization was decreas-ing and stimulus control was increas-ing as the training progressed.
Although there are occasional find-ings in the literature on stimulus con-trol that raise questions of interpreta-tion, the important factor determiningwhether gradients of generalization be-come steeper with continued trainingappears to be the presence or absenceof a correlation in time between thepresence of the stimulus and the deliv-ery of the reinforcer. Differential rein-forcement with respect to a stimulusleads to steeper gradients on either sideof that stimulus. Nondifferential rein-forcement leads to a flattening andbroadening of those gradients. Thereare a number of experiments that sup-port this rule (see Dinsmoor, 1985), butthe most dramatic is the one conductedby Lyons and Thomas (1967). First theauthors gave their birds a series of ses-
60 JAMES A. DINSMOOR
EXP. I 14 EXP. II(WC) / :(WK)
3000 -3000
zo 2000 2000
0
-90 -45 0 +45 +90 -90 -45 0 +45 +90
DEGREES OF TILT
Figure 3. Mean gradients of generalization for tilt of line displayed on key following 2, 4, 7, or14 days of training with food deliveries in the presence of a vertical line but no food in its absence.(Reproduced from Hearst & Koresko, 1968, copyright American Psychological Association.)
sions in which pecking was reinforcedin the presence of green illumination ofthe key but was not reinforced in thepresence of a white line on a darkbackground. The gradient of general-ization along the wavelength dimen-sion became relatively steep. Then thesame birds were given a series of ses-sions in which the reinforcer was de-livered without regard to which stim-ulus was displayed on the key. Thegradient became flatter. The procedurewas alternated four more times, and oneach occasion all of the birds showedsteeper gradients following correlatedreinforcement and shallower gradientsfollowing noncorrelated reinforcement.By the sign test, six determinations foreach of 9 birds yields such a consistentresult with a chance probability of oneover the 54th power of two.
Observing the Stimulus
Early theories of discriminationlearning (e.g., Skinner, 1938; Spence,1936) attributed the divergence in per-formance to successive increments instrength produced by reinforcement ofthe response in the presence of the pos-itive stimulus and successive decre-ments produced by extinction in thepresence of the negative stimulus.They offered no explanation, however,as to why those increments and dec-rements did not continue to transfer be-tween the two stimuli, as they did atthe beginning of training, and they donot explain why the gradients of gen-eralization should become steeper inexperiments like those we have justbeen considering. After all, althoughdifferential reinforcement was appliedto the presence versus the absence of
STIMULUS CONTROL 61
the positive stimulus, it was not ap-plied to any two points along the testdimension. Where is the differentialstrengthening between those twopoints? The only plausible explanationfor the growing difference in responseto different points along the test di-mension is that under this kind of train-ing the subject learns to observe (e.g.,look at) and presumably to attend tothat and similar stimuli.To repeat a point raised earlier, note
that neither the subject in an experi-ment nor the ordinary person goingabout his or her daily business can dis-criminate between stimuli with whichhe or she has no sensory contact. Totake an extreme case, a person who iscurrently located in London cannot re-act to the color displayed on a trafficsignal located in Tokyo. Similarly,even if the signal is within the rangeof his or her vision, this same personcannot react to it unless he or she looksin the right direction to see that signal.The rays of light never impinge on thecells of the retina. Accommodation ofthe lens may also be necessary to bringthe image into a sharp focus. Readingglasses or a magnifying device may benecessary to discriminate fine detail. Adeaf person turns to the speaker to readthe speaker's lips. In the supermarket,a shopper cannot judge the flavor of anewly offered brand of candy or cook-ies unless he or she is given a sampleto place in his or her mouth. Until then,the constituent chemicals never reachthe taste buds. To detect the presenceof a herd of wildebeest, the lion sniffsthe wind, bringing the odorous mole-cules into contact with the sensorymembranes within its nasal passages,and to detect the approach of the lion,the wildebeest do the same. A dog ap-proaches and sniffs the body of a per-son or another dog. We judge the thick-ness and texture of a piece of cloth byrubbing it between two fingers, we de-tect fever in a child by holding a handagainst his or her forehead, and wesniff the carton of milk in our refrig-erator to determine whether it is stillfresh or has become soured. As a col-
lective category, the responses thatbring about these contacts betweenstimulus energies and receptor cells areknown, after Wyckoff (1952), as ob-serving responses.
In the laboratory, the natural observ-ing responses are difficult to record. In-strumentation to determine where thepigeon is looking from moment to mo-ment would be extremely cumbersome,and fixing the position of the bird'shead would interfere with its ability topeck the key. Experimental contingen-cies can be set up, however, that permitother, more readily recorded responsesto mediate the subject's contact withthe discriminative stimuli. Becausethese substitute responses have thesame function as natural observing re-sponses, they are classified as artificialobserving responses. For purposes ofexperimental control, it is desirablethat such responses have no effect onthe scheduling of primary reinforce-ment, and Wyckoff made this a part ofthe standard definition. In his experi-mental work, Wyckoff (1969) used anelectric circuit controlled by a pedal ly-ing on the floor of the pigeon's condi-tioning chamber. In combination withother switching circuitry, this circuitcontrolled the color displayed on thekey. When the bird stood on the pedal(observing), the key was either red orgreen, depending on whether theschedule was one of intermittent rein-forcement or one of no reinforcement;when the bird was off the pedal (notobserving), however, the key was al-ways white, regardless of the scheduleof reinforcement. Although the color ofthe key was sometimes affected by theschedule of reinforcement, note thatthe schedule of reinforcement was nev-er affected by whether the bird was onor off the pedal: It was controlled byan entirely separate timing circuit.Wyckoff considered this arrange-
ment to be equivalent to natural behav-ior like looking at the key and seeingthe red or the green or looking else-where (e.g., the wall of the chamber)and not seeing a discriminative stimu-lus. He found that when the red pre-
62 JAMES A. DINSMOOR
Positive StimulusNegative Stimulus,
Left KeyRight Key
r I I I
I I I I I
Seconds
L~~~~~~~~~~~~~~f It l f,t ft ft
III
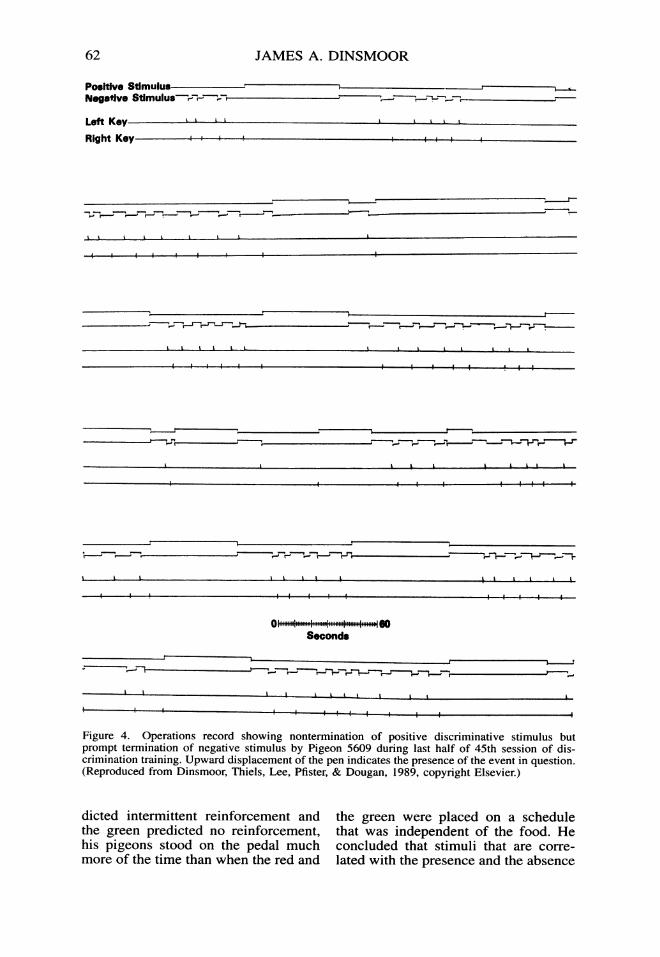
Figure 4. Operations record showing nontermination of positive discriminative stimulus butprompt termination of negative stimulus by Pigeon 5609 during last half of 45th session of dis-crimination training. Upward displacement of the pen indicates the presence of the event in question.(Reproduced from Dinsmoor, Thiels, Lee, Pfister, & Dougan, 1989, copyright Elsevier.)
dicted intermittent reinforcement andthe green predicted no reinforcement,his pigeons stood on the pedal muchmore of the time than when the red and
the green were placed on a schedulethat was independent of the food. Heconcluded that stimuli that are corre-lated with the presence and the absence
STIMULUS CONTROL 63
of a schedule of primary reinforcementbecome conditioned reinforcers andtherefore strengthen and maintain theresponse that produces them.
Later studies have refined this con-clusion: We now realize that it is thestimulus that is positively correlatedwith the primary reinforcement (S+)that becomes a conditioned reinforcerand maintains the observing behavior,whereas the stimulus that is negativelycorrelated with the primary reinforce-ment (S-) actually becomes aversiveto the subject. (For reviews of the ob-serving literature up to their respectivedates of publication, see Dinsmoor,1983; Fantino, 1977.) As an illustra-tion, consider a detailed record of stim-uli and responses obtained by Dins-moor, Thiels, Lee, Pfister, and Dougan(1989) during the second half of one oftheir experimental sessions (Figure 4).The pigeon could turn a discriminativestimulus on by pecking a key to theright of the food key (observing) andcould turn it off by pecking one to theleft (equivalent to turning away fromthe stimulus). The record shows thatwhen the stimulus that came on as aresult of a peck on the observing keywas positive, the pigeon left it on in-definitely; but when the stimulusturned out to be negative, within thenext few seconds the pigeon usuallyturned it off.
In most studies of the formation ofa discrimination, the experimenter doesnot record the subject's observing be-havior. But any time reinforcers are de-livered in the presence of one memberof a pair of stimuli and not in the pres-ence of the other, the conditions havebeen met for such behavior to be ac-quired. Presumably this behavior is, infact, acquired and leads to an increasein the frequency and/or duration of thesubject's sensory contact with the rel-evant stimuli. It is the increasing con-tact with the stimuli that is responsiblefor the increasing slope of the gradientof generalization.
There have been a number of exper-iments in which there seems to be noother explanation (see Dinsmoor,
1985). In these experiments, pigeonshave been trained to discriminate be-tween the presence and the absence ofa specific stimulus (e.g., line, pattern,wavelength) displayed on the key andhave then been tested with variationsin some feature of that stimulus. Al-though they did share one point incommon-the positive stimulus it-self-the dimension along which thetest stimuli were arranged was other-wise completely independent of (i.e.,perpendicular to) the dimension sepa-rating the stimuli used in the training.Nevertheless, the slope of the gradientbecame steeper with continued train-ing. Recall, for example, the previous-ly cited experiment by Hearst and Ko-resko (1968). Pecking was reinforcedin the presence of a vertical line, butno food was delivered in its absence(blank key). Then lines of several dif-ferent tilts were presented during thetest period, without further reinforce-ment. With the exception of the verti-cal line, note that the pigeon had neverseen any of these stimuli before andhad not been trained to distinguishamong them; that is, pecking had neverbeen increased by reinforcement or de-creased by nonreinforcement in thepresence of any other member of theseries. The increasing slope of the gra-dient could not be attributed to differ-ential reinforcement of responding tolines of different tilt. It was based on acharacteristic of the stimulus, the de-gree to which each tilt in the series re-sembled the one used in the training,but that characteristic had been broughtto bear by training applied to a dimen-sion quite independent of the one onwhich the test was conducted. Some-thing had been carried over from thetraining on one dimension to the test-ing on the other, and the most plausiblesuggestion is that what had been trans-ferred was the behavior of looking atthe relevant stimulus.
Gradients of Generalization Aroundthe Negative Stimulus
In most experiments of this sort, ithas been the presence of the critical
64 JAMES A. DINSMOOR
*- I Dayo-o 2 Days
160 4-A8Days A-- ~~8 Days A-_ A 400
14 0l--0-1 16 Days,
1 20 300oZU
LU 0, 80' , 200,
60 'L'U /
40 " ~ 100
20 - 7~0 Z 0-90 -60 -30 0 +30 +60 +90 Blank
DEGREES FROM S-
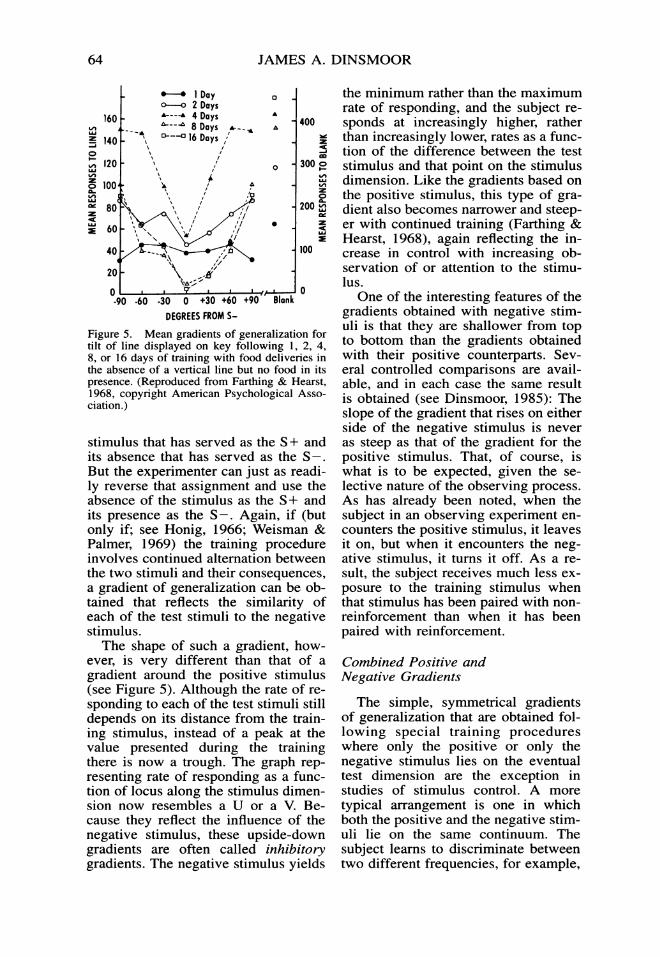
Figure 5. Mean gradients of generalization fortilt of line displayed on key following 1, 2, 4,8, or 16 days of training with food deliveries inthe absence of a vertical line but no food in itspresence. (Reproduced from Farthing & Hearst,1968, copyright American Psychological Asso-ciation.)
stimulus that has served as the S + andits absence that has served as the S -.But the experimenter can just as readi-ly reverse that assignment and use theabsence of the stimulus as the S+ andits presence as the S -. Again, if (butonly if; see Honig, 1966; Weisman &Palmer, 1969) the training procedureinvolves continued alternation betweenthe two stimuli and their consequences,a gradient of generalization can be ob-tained that reflects the similarity ofeach of the test stimuli to the negativestimulus.The shape of such a gradient, how-
ever, is very different than that of agradient around the positive stimulus(see Figure 5). Although the rate of re-sponding to each of the test stimuli stilldepends on its distance from the train-ing stimulus, instead of a peak at thevalue presented during the trainingthere is now a trough. The graph rep-resenting rate of responding as a func-tion of locus along the stimulus dimen-sion now resembles a U or a V. Be-cause they reflect the influence of thenegative stimulus, these upside-downgradients are often called inhibitorygradients. The negative stimulus yields
the minimum rather than the maximumrate of responding, and the subject re-sponds at increasingly higher, ratherthan increasingly lower, rates as a func-tion of the difference between the teststimulus and that point on the stimulusdimension. Like the gradients based onthe positive stimulus, this type of gra-dient also becomes narrower and steep-er with continued training (Farthing &Hearst, 1968), again reflecting the in-crease in control with increasing ob-servation of or attention to the stimu-lus.One of the interesting features of the
gradients obtained with negative stim-uli is that they are shallower from topto bottom than the gradients obtainedwith their positive counterparts. Sev-eral controlled comparisons are avail-able, and in each case the same resultis obtained (see Dinsmoor, 1985): Theslope of the gradient that rises on eitherside of the negative stimulus is neveras steep as that of the gradient for thepositive stimulus. That, of course, iswhat is to be expected, given the se-lective nature of the observing process.As has already been noted, when thesubject in an observing experiment en-counters the positive stimulus, it leavesit on, but when it encounters the neg-ative stimulus, it turns it off. As a re-sult, the subject receives much less ex-posure to the training stimulus whenthat stimulus has been paired with non-reinforcement than when it has beenpaired with reinforcement.
Combined Positive andNegative Gradients
The simple, symmetrical gradientsof generalization that are obtained fol-lowing special training procedureswhere only the positive or only thenegative stimulus lies on the eventualtest dimension are the exception instudies of stimulus control. A moretypical arrangement is one in whichboth the positive and the negative stim-uli lie on the same continuum. Thesubject learns to discriminate betweentwo different frequencies, for example,
STIMULUS CONTROL 65
GROUP E -
30- - GROUP IA A
GROUP D -----
sn25- AOOz --A.-
20 5
z15-
z
490 510 520 530 540 550 560 570 580 590 610
WAVELENGTH ( NM)
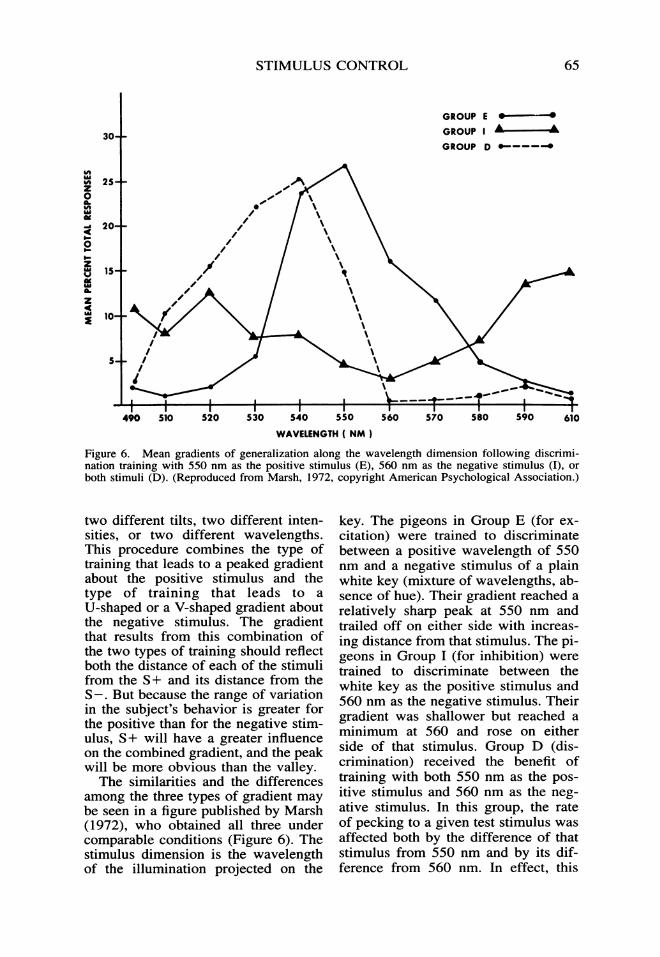
Figure 6. Mean gradients of generalization along the wavelength dimension following discrimi-nation training with 550 nm as the positive stimulus (E), 560 nm as the negative stimulus (I), orboth stimuli (D). (Reproduced from Marsh, 1972, copyright American Psychological Association.)
two different tilts, two different inten-sities, or two different wavelengths.This procedure combines the type oftraining that leads to a peaked gradientabout the positive stimulus and thetype of training that leads to aU-shaped or a V-shaped gradient aboutthe negative stimulus. The gradientthat results from this combination ofthe two types of training should reflectboth the distance of each of the stimulifrom the S+ and its distance from theS-. But because the range of variationin the subject's behavior is greater forthe positive than for the negative stim-ulus, S+ will have a greater influenceon the combined gradient, and the peakwill be more obvious than the valley.The similarities and the differences
among the three types of gradient maybe seen in a figure published by Marsh(1972), who obtained all three undercomparable conditions (Figure 6). Thestimulus dimension is the wavelengthof the illumination projected on the
key. The pigeons in Group E (for ex-citation) were trained to discriminatebetween a positive wavelength of 550nm and a negative stimulus of a plainwhite key (mixture of wavelengths, ab-sence of hue). Their gradient reached arelatively sharp peak at 550 nm andtrailed off on either side with increas-ing distance from that stimulus. The pi-geons in Group I (for inhibition) weretrained to discriminate between thewhite key as the positive stimulus and560 nm as the negative stimulus. Theirgradient was shallower but reached aminimum at 560 and rose on eitherside of that stimulus. Group D (dis-crimination) received the benefit oftraining with both 550 nm as the pos-itive stimulus and 560 nm as the neg-ative stimulus. In this group, the rateof pecking to a given test stimulus wasaffected both by the difference of thatstimulus from 550 nm and by its dif-ference from 560 nm. In effect, this
66 JAMES A. DINSMOOR
gradient combined the effects of theother two types of training.The quantitative relations among
these three types of gradient have beenexamined most carefully in a studyconducted by Hearst (1968, 1969). Forone group of pigeons, Hearst used avertical line as his positive stimulus toobtain gradients of excitation along thetilt dimension; for a second group, heused a line of a different tilt as his neg-ative stimulus to obtain gradients of in-hibition; and for a third group, he usedboth of these stimuli to obtain a com-bined gradient. Altogether, he replicat-ed this same basic experimental designthree times, using three different de-grees of separation between S- andS +. There were nine groups in all. Af-ter correcting for the possible effects ofbehavioral contrast, Hearst concludedthat there was a relatively satisfactoryfit between the sum of the gradientsobtained with separate S+ training andS- training and the gradient obtainedfollowing combined training with bothof these stimuli. This finding impliesthat it is appropriate to account for thedevelopment of the combined gradient,following normal discrimination train-ing, with the same principles as thoserequired to explain the changes in thepositive and the negative gradients thatmade it up. That is, discrimination be-tween two stimuli lying along the samecontinuum reflects the combined influ-ences of the subject's observing of theS - and its observing of the S+.
IMMEDIATE DISCRIMINATION
Early theorists like Skinner (1933,1938) and Spence (1936) saw suchconventional measures of "strength"as latency, rate, proportion of re-sponses, and resistance to extinctionseparate during discrimination training.When, as was often the case, there wasa progressive increase in S+ and a pro-gressive decrease in S-, it was naturalfor them to conclude that reinforce-ment of the response in the one stim-ulus and its extinction in the other werethe factors responsible for the separa-
tion of the two performances. Yet un-der other circumstances, where there isa high level of observing from the be-ginning of the discrimination training,the separation is already evident assoon as the response is shaped, beforeany extinction has occurred. Nostrengthening or weakening seems tobe required.
Consider the case in which the twostimuli are alternative colors displayedon the pigeon's key. As the color is dis-persed evenly over the entire area ofthe key, occupying a large visual anglefrom the bird's point of view, it wouldseem to be difficult for the subject tomiss seeing it as it pecks that key. Ininitial training conducted in my labo-ratory prior to a discrimination basedon punishment (Dinsmoor, Flint,Smith, & Viemeister, 1969), we foundthat after pecking had been shaped andintermittently reinforced with the keylighted red, it was already very diffi-cult to get the bird to peck that samekey lighted with green. The birdstopped as soon as the green appearedand did not resume pecking duringlong periods of exposure to that colorinterspersed with further exposures toreinforcement in red. Terrace (1963)has documented a similar phenome-non, although it was based on a morecomplex sequence of procedures (seeDinsmoor, 1985, for further discus-sion). Despite Keller and Schoenfeld'sdictum (1950, p. 119), extinction is notthe hallmark of discrimination. The al-ternative schedules of reinforcementdetermine the nature of the final per-formance under either stimulus, as inwhat are known as multiple schedules,but the independence of the scheduleeffects requires a different explanation.
SUMMARY
This tutorial is addressed to readerswho are not as conversant as theywould like to be with basic principlesin the control of behavior by antece-dent stimuli. Its purpose is to providea brief overview and a theoretical in-tegration of some carefully selected re-
STIMULUS CONTROL 67
search findings. The present install-ment deals primarily with the acquisi-tion of control by discriminative stim-uli; a second installment will extendthe coverage to more specialized topicsthat are also important to a systematicanalysis of behavior.The reader is warned, however, that
the present analysis departs from stan-dard textbook accounts in several re-spects:
1. In contrast to Skinner's rejectionof "stimulus-response psychology," itis here suggested that antecedent stim-uli exert a pervasive influence on op-erant, as well as respondent, behavior.They serve as a crucial link betweencurrent behavior and past reinforce-ment. They are the direct and imme-diate determinants of what the individ-ual does at any given moment.
2. As Skinner (1938, pp. 176-177,241) but few others have recognized,the procedure by which Pavlov trainedhis dogs to salivate to a conditionalstimulus (respondent conditioning) isinherently a form of discriminationtraining. In this sense, all conditionalstimuli are discriminative stimuli, andthe distinction in terminology appearsto rest more on the type of behaviorthat is under consideration and howthat behavior is measured than on anydifference in the role of the stimulus.
3. The Pavlovian view that stimulusgeneralization represents an active pro-cess in which new and different stimulibecome effective because of the spreadof excitation from one point to anotheris rejected. The gradient of generaliza-tion becomes narrower, rather thanbroader, as training continues. In thepresent account, discrimination andgeneralization are treated simply as al-ternative ways of looking at the samephenomenon, and the slope of the gra-dient is used as a way of measuring it.
4. In discrimination training, thepositive stimulus serves as a condi-tioned reinforcer of observing behav-ior, leading to an increase in sensorycontact with that stimulus; increasedcontact with the negative stimulus is anincidental but inevitable consequence
of the increase in observing. The de-creasing transfer of the effects of re-inforcement to behavior in the pres-ence of S - and of extinction to behav-ior in the presence of S+ is attributedto more frequent and more prolongedobservation of the two stimuli. This re-fers both to the time at which rein-forcement is or is not delivered and tothe time at which the performance issubsequently measured. Although re-moved from direct experimental obser-vation, attention may play a similarrole (Sutherland & Mackintosh, 1971).The respective schedules of reinforce-ment interact with the increasing sep-aration of the two performances to de-termine their eventual levels and tem-poral patterns.
REFERENCES
Buchwald, A. M. (1969). Effects of "right" and"wrong" on subsequent behavior: A new in-terpretation. Psychological Review, 76, 132-143.
Dinsmoor, J. A. (1951). The effect of periodicreinforcement of bar-pressing in the presenceof a discriminative stimulus. Journal of Com-parative and Physiological Psychology, 44,354-361.
Dinsmoor, J. A. (1952). The effect of hungeron discriminated responding. Journal of Ab-normal and Social Psychology, 47, 67-72.
Dinsmoor, J. A. (1983). Observing and condi-tioned reinforcement. Behavioral and BrainSciences, 6, 693-704.
Dinsmoor, J. A. (1985). The role of observingand attention in establishing stimulus control.Journal of the Experimental Analysis of Be-havior, 43, 365-381.
Dinsmoor, J. A., Flint, G. A., Smith, R. F, &Viemeister, N. F (1969). Differential rein-forcing effects of stimuli associated with thepresence or absence of a schedule of punish-ment. In D. P. Hendry (Ed.), Conditioned re-inforcement (pp. 357-384). Homewood, IL:Dorsey.
Dinsmoor, J. A., Thiels, E., Lee, D. M., Pfister,J., & Dougan, J. D. (1989). Selective observ-ing: Pigeons turn discriminative stimuli onand off by pecking separate keys. In N. W.Bond & D. A. T. Siddle (Eds.), Proceedings,XXIVth International Congress of Psycholo-gy: Vol. 6. Psychobiology: Issues and appli-cations (pp. 443-456). Amsterdam: Elsevier.
Fantino, E. (1977). Conditioned reinforcement:Choice and information. In W. K. Honig & J.R. Staddon (Eds.), Handbook of operant be-havior (pp. 313-339). Englewood Cliffs, NJ:Prentice-Hall.
68 JAMES A. DINSMOOR
Farthing, G. W., & Hearst, E. (1968). General-ization gradients of inhibition after differentamounts of training. Journal of the Experi-mental Analysis of Behavior, 11, 743-752.
Guttman, N., & Kalish, H. I. (1956). Discri-minability and stimulus generalization. Jour-nal of Experimental Psychology, 51, 79-88.
Hearst, E. (1968). Discrimination learning asthe summation of excitation and inhibition.Science, 162, 1303-1306.
Hearst, E. (1969). Excitation, inhibition anddiscrimination learning. In N. J. Mackintosh& W. K. Honig (Eds.), Fundamental issues inassociative learning (pp. 1-41). Halifax, NovaScotia: Dalhousie University Press.
Hearst, E. (1975). The classical-instrumentaldistinction: Reflexes, voluntary behavior, andcategories of associative learning. In W. K.Estes (Ed.), Handbook of learning and cog-nitive processes: Vol. 2. Conditioning and be-havior theory (pp. 181-223). Hillsdale, NJ:Erlbaum.
Hearst, E., & Koresko, M. B. (1968). Stimulusgeneralization and amount of prior training onvariable-interval reinforcement. Journal ofComparative and Physiological Psychology,66, 133-138.
Honig, W. K. (1966). The role of discriminationtraining in the generalization of punishment.Journal of the Experimental Analysis of Be-havior, 9, 377-384.
Jenkins, H. M., & Harrison, R. H. (1960). Ef-fects of discrimination training on auditorygeneralization. Journal of Experimental Psy-chology, 59, 246-253.
Keller, F. S. (1943). Studies in internationalMorse code: I. A new method of teachingcode reception. Journal of Applied Psycholo-gy, 27, 407-415.
Keller, F S., & Schoenfeld, W. N. (1950). Prin-ciples of psychology. New York: Appleton-Century-Crofts.
Lyons, J., & Thomas, D. R. (1967). Effects ofinterdimensional training on stimulus gener-alization: II. Within-subject design. Journal ofExperimental Psychology, 75, 572-574.
Marsh, G. (1972). Prediction of the peak shift
in pigeons from gradients of excitation andinhibition. Journal of Comparative and Phys-iological Psychology, 81, 262-266.
Millenson, J. R., & Dent, J. G. (1971). Habit-uation of conditioned suppression. QuarterlyJournal ofExperimental Psychology, 23, 126-134.
Pavlov, I. P. (1960). Conditioned reflexes: Aninvestigation of the physiological activity ofthe cerebral cortex (G. V. Anrep, Trans.). NewYork: Dover. (Original work published 1927)
Rescorla, R. A. (1988). Pavlovian conditioning:It's not what you think it is. American Psy-chologist, 43, 15 1-160.
Skinner, B. F (1933). The rate of establishmentof a discrimination. Journal of General Psy-chology, 9, 302-350.
Skinner, B. F (1937). Two types of conditionedreflex: A reply to Konorski and Miller. Jour-nal of General Psychology, 16, 272-279.
Skinner, B. F (1938). The behavior of organ-isms: An experimental analysis. New York:Appleton-Century-Crofts.
Skinner, B. F (1972). Cumulative record: A se-lection ofpapers (3rd ed.). New York: Apple-ton-Century-Crofts.
Spence, K. W. (1936). The nature of discrimi-nation learning in animals. Psychological Re-view, 43, 427-449.
Sutherland, N. S., & Mackintosh, N. J. (1971).Mechanisms of animal discrimination learn-ing. New York: Academic Press.
Terrace, H. S. (1963). Discrimination learningwith and without "errors." Journal of the Ex-perimental Analysis of Behavior, 6, 1-27.
Weisman, R. G., & Palmer, J. A. (1969). Fac-tors influencing inhibitory stimulus control:Discrimination training and prior non-differ-ential reinforcement. Journal of the Experi-mental Analysis of Behavior, 12, 229-237.
Wyckoff, L. B. (1952). The role of observingresponses in discrimination learning. Part 1.Psychological Review, 66, 68-78.
Wyckoff, L. B. (1969). The role of observingresponses in discrimination learning. In D. PHendry (Ed.), Conditioned reinforcement (pp.237-250). Homewood, IL: Dorsey.