The isolated, volcanic Mascarene Islands of Mauritius (20.3°S, 57.6°E), Réunion (21.1°S, 55.5°E), and Rodrigues (19.7°S, 63.4°E) are situated in the southwestern Indian Ocean, with Madagascar, the nearest large landmass, some 665 km to the east of Réunion. Rodrigues (Fig. 1A) is the smallest of the Mascarene Islands, only 17.7 km long and 8.5 km wide, with a surface area of 104 km². The Mascarenes have been subject to extremely high biota extinction rates
due to direct human impacts and the introduction of invasive species, epitomised by the loss of the iconic dodo, Raphus cucullatus (Linnaeus, 1758), of Mauritius (Cheke and Hume, 2008). A diversity of endemic giant tortoises once inhabited these islands, with Cylindraspis indica (Schneider, 1783) on Réunion featuring highly variable shell shapes, the dome-shelled C. inepta Günther, 1873 and the saddleback-shelled C. triserrata Günther, 1873 on Mauritius, and the dome-shelled C. peltastes (Dumeril & Bibron, 1835) and the saddleback-shelled C. vosmaeri (Suckow, 1798) on Rodrigues (Arnold, 1979; Bour, 1980, 1981; Bour et al., 2014a). Although giant tortoises were abundant on all of the Mascarenes, Rodrigues harboured one of the densest populations of giant tortoises on Earth, estimated at 150,000–200,000 individuals (North-Coombes, 1971).
The importance of giant tortoises as fresh meat and to obtain oil for mariners on long sea voyages resulted in their over-exploitation, and as soon as settlement began on the Mascarenes, thousands were killed indiscriminately (Mondini, 1990; Cheke and Bour, 2014). The Mauritius mainland populations disappeared by 1721, with only small numbers of tortoises surviving
Herpetology Notes, volume 14: 103-116 (2021) (published online on 10 January 2021)
Discovery of the first Mascarene giant tortoise nesting site on Rodrigues Island, Indian Ocean (Testudinidae: Cylindraspis)
Julian P. Hume1,*, Owen Griffiths2, Aurèle Anquetil Andre3, Arnaud Meunier3, and Roger Bour4,†
1 Bird Group, Department of Zoology, Natural History Museum, Akeman Street, Tring, Herts HP23 6AP, United Kingdom.
2 La Vanille Réserve des Mascareignes, Senneville, Rivière des Anguilles, Mauritius.
3 Francois Leguat Giant Tortoise and Cave Reserve, Anse Quitor, Rodrigues Island, Mauritius.
4 Muséum National d’Histoire Naturelle, 57 rue Cuvier, 75005 Paris, France.
Abstract. Five species of giant tortoise (genus Cylindraspis) once occurred in huge abundance on the Mascarene Islands of Mauritius, Réunion, and Rodrigues. They disappeared after colonisation of the island by humans in the 17th and 18th centuries, primarily due to over-hunting and predation of eggs and young by introduced pigs and cats. So rapid was their extinction that virtually nothing is known about their life history, especially nesting and egg-laying behaviour. Here we report the discovery on Rodrigues of the first Mascarene tortoise-nesting site, which contained intact nesting chambers, complete egg clutches and fossil remains of a known native predator of tortoise eggs. We further compare the nesting behaviour with the giant tortoises of Aldabra Atoll in the Seychelles and the Galapagos Archipelago in Ecuador and provide details of the decline and extinction of Mascarene tortoises, most notably those of Rodrigues, for which good historical records exist.
† We dedicate this paper to our friend, colleague, and co-author Roger Bour, a world expert on Mascarene tortoises and a life-long steam train fanatic, whose passing came much too soon.
“There are such plenty of land-turtles in this Isle [Rodrigues], that sometimes you see two or three thousand of them in a flock; so that one may go above a hundred paces on their backs; or, to speak more properly on their carapaces, without setting foot to the ground.” [François Leguat 1708: 65]
Julian P. Hume et al.104
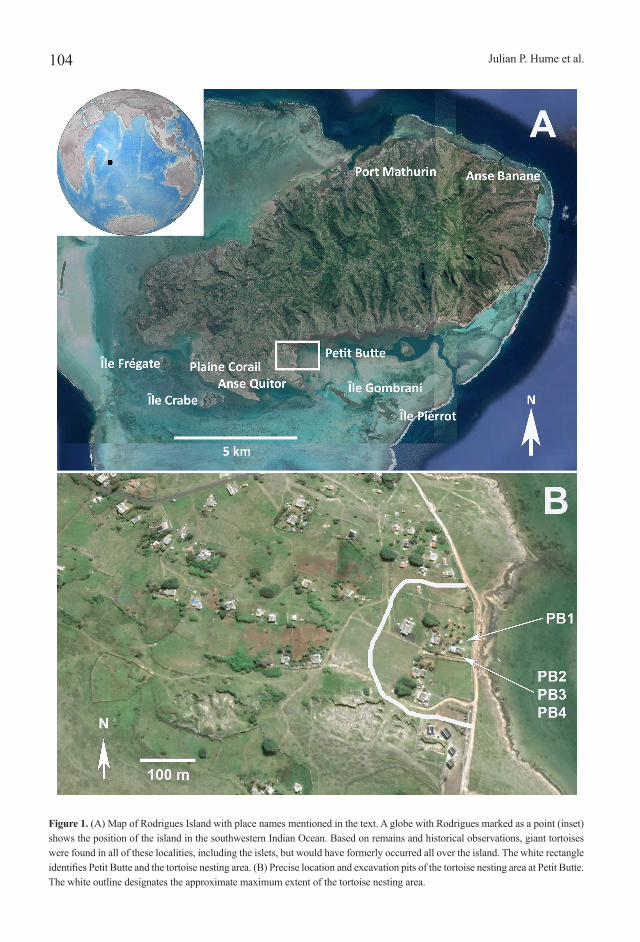
Figure 1. (A) Map of Rodrigues Island with place names mentioned in the text. A globe with Rodrigues marked as a point (inset) shows the position of the island in the southwestern Indian Ocean. Based on remains and historical observations, giant tortoises were found in all of these localities, including the islets, but would have formerly occurred all over the island. The white rectangle identifies Petit Butte and the tortoise nesting area. (B) Precise location and excavation pits of the tortoise nesting area at Petit Butte. The white outline designates the approximate maximum extent of the tortoise nesting area.
on surrounding islets until the 1730s, or possibly until as late as the 1870s on Round Island (Cheke and Hume, 2008; Cheke and Bour, 2014). Réunion tortoises were extirpated by the 1730s in the lowlands (Bory de Saint-Vincent, 1804; Bour, 1979, 1981; Cheke and Bour, 2014) but, remarkably, a small population survived in the remote montane refuge of Cilaos until at least 1840 (Hermann, 1903; Bour et al., 2014a; Cheke and Bour, 2014). Tortoises on Rodrigues remained comparatively unmolested until the early 18th century, but after the extinction of the populations on Mauritius and Réunion, attention turned to the more off-the-beaten-path Rodrigues (North-Coombes, 1994).
The Huguenot refugee François Leguat had reported the abundance of tortoises during his stay in 1691–93 (Leguat, 1708), and in 1725 a reconnaissance party was sent from Réunion to ascertain the importance of then uninhabited Rodrigues to French colonial strategy, which included ships’ mate Julien Tafforet (North-Coombes, 1971; Dupon, 1974). Tafforet became marooned on Rodrigues for eight months and subsequently wrote a detailed report on the island and confirmed the abundance of giant tortoises (Tafforet, 1725–26). As a direct result, a tortoise hunting station was set up on Rodrigues in 1735 to provide meat and oil to the burgeoning colonies on Mauritius and Réunion (Herbert and Nichelson, 1780; Cheke and Hume, 2008; Hume, 2019). A holding pound was installed on Rodrigues to prepare tortoises for shipment, with an equivalent facility at Port Louis, the capital of Mauritius, to receive them (Cheke and Hume, 2008; Cheke and Bour, 2014). North-Coombes (1994) estimated that over 280,000 tortoises were butchered on Rodrigues or exported alive between ca. 1732 and 1771, with around 10,000 taken annually for the first two decades. It was only due to the collapse of the giant tortoise populations in the 1770s that the slaughter ended, as it was then no longer viable to collect them (North-Coombes, 1971). A single live tortoise was collected in 1786 on the Plaine Corail, southwestern Rodrigues (Hume et al., 2015), with the last record of two seen around 1795 in inaccessible gorges by the Civil Administrator of Rodrigues, Philibert Marragon (Bour et al., 2014b). Such was the speed of their extinction that virtually nothing was recorded about the life history of the Mascarene giant tortoises, especially nesting and egg-laying behaviours. Reports that the tortoises sought out dry coastal areas for laying, with the eggs incubated by the sun, were the only exceptions (Leguat, 1708; Mundy in Temple, 1914; Lougnon, 2006).
Freycinet (1825) reported the discovery of the first fossil bones and eggs of Cylindraspis tortoises on Mauritius (the present whereabouts of this material is unknown), which had been buried in volcanic tuff and could be dated to the Middle Pleistocene, since the last eruption on the island occurred around 31,000 YBP (Moore et al., 2011; Cheke et al., 2016). The only other discovery of Cylindraspis eggs in the Mascarenes was made by Bour et al. (2014a), who described four incomplete eggs of C. indica buried in soft sediment and excavated by Jean-Pierre Esmiol and Roger Bour near Boucan-Canot, northwestern Réunion. Unusually for extant giant tortoises, the eggs were ellipsoidal and not spherical (but see below).
During an excavation on the right side of their garden to insert a septic tank at Petit Butte, Plaine Corail, southwestern Rodrigues, in 2017, the Milazar family noticed a continuous layer of broken eggshells at approximately 400 mm depth (identified hereafter as PB1). Destruction of the site during excavation prompted Denis Claude Milazar to start a second small excavation on the left side of the garden (PB2). Careful removal of sediment revealed almost complete tortoise eggs and a possible nesting chamber, but the discovery lacked contextual data. AAA and AM returned to the site for further analysis and examined the eggs and the locality. Once the importance of the discovery was realised, a scientific team led by JPH and OG excavated the site in 2018 in a community effort that included participation of local landowners, staff of the François Leguat Cave and Giant Tortoise Reserve (FLR), Mauritius Wildlife Foundation (MWF), and members of the Rodrigues Regional Assembly (RRA). This process proved extremely productive, with fossil remains found over a substantial area adjacent to the coast (Fig. 1B).
Here we report on the excavation of the first recorded nesting site of the endemic Mascarene giant tortoises, which includes the description of eggs and egg chambers, and the discovery of unhatched young and an associated natural predator. We further compare the nesting behaviour of Rodrigues Cylindraspis giant tortoises with those of other giant tortoises and discuss the reasons for their decline and extinction, most notably for those on Rodrigues, for which historical data exists.
Materials and Methods
Only part of the left side of the Milazar family garden remained pristine, so excavation of two further sites (PB3 and PB4) began to the west (PB3) and to the south
Discovery of the First Cylindraspis Giant Tortoise Nesting Site on Rodrigues Island 105
(PB4) of PB2 (Fig. 1B). PB3 and PB4 measured 3 x 2.5 m and were divided into 1-m quadrants with string lines. A datum was set at 600 mm above surface between PB2 and PB4. Removal of the surface vegetation and humic layer exposed the underlying sandy soil layer. Excavation of 30 mm layers, each prefixed with PB (= Petit Butte), were taken to a maximum depth of 600 mm, which was about 200 mm below the visible horizon of fossil-bearing sediments. Careful excavation revealed potential egg chambers, and where these were still intact, removal of sediment occurred around and below the chamber in order to preserve as much detail as possible. As the sediment was dry, sieving of each excavated 30 mm level occurred on site. Sieving was undertaken using three sieve sizes, 15 mm, 10 mm, and 2 mm, with all finds deposited away from the nesting site. Precise locations of all fossil material were recorded with pencil on graph paper, including the egg chambers and their relationship to each other. All fossil material was photographed in-situ, with particular emphasis placed on intact tortoise incubation chambers that still contained eggs and egg clutches. Intact eggs and a unique clutch of eggs (PB47) were hardened with Paraloid B72 before removal. All fossil material was bulk labelled, individually packed and sent for analysis at the research facilities at FLR. Once excavation was complete, the entire fossil locality was back-filled, with the excavated boundaries lined with sheet plastic to allow continued research in the future. All fossil material is stored at FLR for curation.
Plaine Corail is a vast calcarenite outcrop, which contains pockets of soil, intermixed with large expanses of exposed limestone without sediment. Using a 50-m tape, perimeter measurements were done to record the entire soft-sediment area at Petit Butte, with the surface examined for eggshell deposits.
Results
The maximum size of the nesting area, defined as the area that included a suitable depth of soil (> 300 mm), was 90 x 133 m. It is bordered on all sides by exposed calcarenite with no sediment (Fig. 1B). No other soil deposits occur near Petit Butte, the next nearest at Anse Quitor to the west (Fig. 1A). The lagoon to the south created a natural border, with the closest nesting site (PB3) situated 8 m to the north, and the furthest soil deposit 55 m inland, both measurements taken from the shoreline. Separating the nesting area from the lagoon edge is a garden wall and access track (Fig. 1B). The nesting area is 3.5 m above present sea level (PSL) in the lagoon, with
a 1-m, twice-daily sea level rise and fall. Examination of the surface area, some of which was under cultivation, revealed disturbed eggshell fragments extending to all sides of the soil boundary. Covering much of the surface area is Sand Couch, Sporobolus virginicus (L.) Kunth, a grass species introduced in the 19th century (Hubbard and Vaughan, 1940), with roots and a disturbed humic layer descending to a soil depth of 80–100 mm. Directly underneath the humic layer, undisturbed sediments were dry and loose with good drainage, comprising undifferentiated < 1 mm sub-angular to rounded coral sand and light brown silt, which remained uniform to at least 600 mm depth. The sediments contained large terrestrial snails, predominantly Tropidophora articulata (Gray, 1834), T. fimbriata rodriguesensis (Crosse, 1873), and the small Omphalotropis littorinula Crosse, 1873 (see Griffiths and Florens, 2006), and local marine molluscs, with variably sized fragments of calcarenite. The base of the nesting area comprised a layer of around 30 mm containing broken, compacted eggshell, naturally hardened by tortoise urine that appeared translucent white depending on the angle of light (Fig. 2).
Three intact egg chambers, one each in PB2, PB3, and PB4, ranged from 360–700 mm in length and 150–300 mm in depth (Table 1), with the base consistently reaching a depth of ~400 mm (300 mm below the upper 100-mm humic layer (Fig. 3). PB2 sat 2 m east of PB3, with PB4 2.5 m further inland and to the west. One chamber (PB2) was ‘sock-shaped’, with a plug of soil extending from the surface to its ‘heel’ (Fig. 4). Two of the chambers contained outlines of eggs, with clutches of eight (PB21) and nine (PB314), which proved impossible to remove intact (Fig. 2). However, a complete clutch in PB4, removed intact from the chamber, contained 13 eggs (PB47) (Fig. 5). The excavation revealed five incomplete hatchling tortoise bones (three humeri and two scapula-coroacoids; PB22), with the most complete humerus measuring 15 mm in total length; a complete phalanx of the Rodrigues Rail Erythromachus leguati Milne-Edwards, 1874 was also recovered (PB23). A Cylindraspis carapace fragment (PB35), fibula fragment (PB38), and scapula-coracoid (PB41) were the only other bones found. Two eggs were extremely small, at only 11 x 12.5 mm these were measuring approximately one-fifth the size of the larger-sized specimens (Table 1; Fig. 6C) but were collected at the same depth and in the same nesting cavity of PB3. Most of the eggs recovered from PB2, PB3, and PB4 were spherical (n = 10), averaging 51 mm in diameter (Fig. 6B), but one (PB32) was oval-shaped, measuring 40 x 55 mm (Table 1; Fig. 6A).
Julian P. Hume et al.106
Comparison of giant tortoise nesting behaviour. The nesting behaviour of Aldabra and Galapagos giant tortoises is comparatively well known, but it is in complete contrast to what our study reveals about Mascarene tortoises. Only a few observers mention any details, but all agree that coastal sandy areas were essential for laying and the sun necessary for incubating the eggs.
Mascarenes.—In 1673, Hubert Hugo, commander of Mauritius from 1673–77 (Moree, 1998), described the carnage that introduced pigs caused among turtle and tortoise eggs by devouring them on the shore as they were laid, and that they also took eggs laid in hollow trees exposed to the sun (Pitot, 1905; Cheke, 1987). This suggests that not all tortoises buried their eggs, which appears to be in direct contrast with those on Rodrigues. However, it is more likely that this egg-laying behaviour was atypical, perhaps a result of high competition for a suitable sandy-soil medium. The explorer Peter Mundy (in Temple, 1914) reported that “They lay their eggs in the sand, which are hatched by the sunne,” as do the tortoises on Aldabra and Galapagos.
On Réunion, Sieur Luillier (in Lougnon, 2006) stated in 1703 that “There is a season when they lay their eggs, but it is the sun that incubates them…,” and that “for months each year they neither eat or drink, while during the remaining eight months they lay their eggs and take what sustenance they need for the four others.” Melet (in Cheke and Bour, 2014) reported in 1671 that herds of hundreds of tortoises descended from the mountains every day, and that a single tortoise may contain three or four hundred eggs. The daily migration may be in reference to the tortoises travelling to the coast to breed, as the heavily forested mountain slopes were much cooler and presumably lacked suitable nesting localities. Melet’s description of a dissected tortoise containing hundreds of eggs is almost certainly in reference to a mature ovary in which hundreds of egg follicles are stored until optimum conditions induce ovulation, which would also include atrophied follicles and remains of mature follicles after the expulsion of the ripe ovum (Bourn, 1977; Swingland and Coe, 1978).
Discovery of the First Cylindraspis Giant Tortoise Nesting Site on Rodrigues Island 107
Figure 2. Dorsal view of the Cylindraspis vosmaeri egg chamber in quadrant PB3 on Rodrigues Island. The egg outlines sit in a mixture of tortoise mucus and urine, which is still visible as translucent white patches on the surface. Scale bar = 100 mm.
Leguat (1708), who was marooned on Rodrigues from 1691–93, provided the most detailed account of the Mascarene tortoises and remarked that “The land-turtles lay theirs in the sand, and cover them, that they may be hatch’d.” Leguat stayed in the north, where the capital Port Mathurin is now (Fig. 1A), so he presumably witnessed tortoises breeding on the north coast. However, sandy soil deposits predominate in the north, west, and east of the island, and especially on the Plaine Corail, where caverns have produced hundreds of hatchling/juvenile tortoise bones (Hume, 2014a). Accurate estimations can be made of the Rodrigues tortoise population density, because of detailed shipment records taken during the peak of tortoise exploitation (North-Coombes, 1994). Based on Aldabra tortoise density, woodland areas on Aldabra support 20 animals per hectare (= 2000 per km2; Aldabra has an area of 155.4 km²) (Bourn and Coe, 1979; Cheke and Bour, 2014), which is also a reasonable estimate for Rodrigues (Cheke and Hume, 2008). Rodrigues is larger, covering 109 km2, and has a higher average rainfall (1200 mm per year) compared with Aldabra (946 mm per year) (Coe et al., 1979). Vast numbers of Rodrigues tortoise fossil remains have been found in caverns on the Plaine Corail, primarily due to the excellent depositional preservation environment limestone caves provide (Hume, 2005), but individual bones have been found elsewhere (Fig. 1A), confirming that tortoises occurred all over the island. The reports of Leguat (1708), Tafforet (1725–26), and Pingré (1763) show that the tortoises were numerous on the lagoonal islets, particularly Ȋle Frégate and Ȋle Gombrani (Fig. 1A). Based on distribution, a total population of 218,000 was possible, but this would have been fluctuating drastically, with the limit set by drought years rather than an average (Cheke and Bour, 2014). Rodrigues may also have had a higher carrying capacity due to increased rainfall and that the population included two ecologically separated species (see Cheke and Hume, 2008).
Aldabra.—Around 100,000–150,000 tortoises occur on Aldabra (Bourn, 1977; Gerlach, 2014). The nesting season takes place in June–September, with a peak in July and coinciding with the dry season (Swingland and Coe, 1978). Females locate the nesting site by detecting the odour of urine from previous nesting attempts, and the same female returns to the same site in subsequent years (Swingland and Coe, 1978); this also helps prevent females excavating other nests. At least 300 mm depth of soil is essential, and with some shade, which provides nest exposure to direct sun for at least 5 h of the day (Swingland and Coe, 1978; Gerlach, 2004). The 300-
Julian P. Hume et al.108
Figure 3. A profile of the Cylindraspis vosmaeri nesting area in quadrant PB1 at Petit Butte, Rodrigues Island. The solid line represents the original surface, with the dotted line around 300 mm below the surface line marking the top of the nesting chamber. The 100 mm scale bar is directly above the tortoise eggshell layer. Note the sandy composition of the soil.
Table 1. Egg and clutch characteristics of the Giant Rodrigues Saddleback Tortoise Cylindraspis vosmaeri (n = 10). Parameters are provided with means ± standard deviations and ranges. Measurements are given in millimeters.
Species Egg Diameter (n = 10)
Clutch Size (n = 4)
Chamber Length (n = 3)
Chamber Depth (n = 3)
Cylindraspis vosmaeri 50–53 51.1 ± 1.5
8–13 10 ± 2.50
360–700 546.6 ± 172.4
150–300 210 ± 79.3
Discovery of the First Cylindraspis Giant Tortoise Nesting Site on Rodrigues Island 109
Figure 5. A complete clutch of Cylindraspis vosmaeri eggs removed intact from PB4 on Rodrigues Island. The clutch contains 13 eggs. Scale bar = 100 mm.
Figure 4. Lateral view of a Cylindraspis vosmaeri tortoise egg chamber in PB2 on Rodrigues Island. Note the ‘sock-shaped’ chamber with a ‘heel’ to the right and a connection to the surface. The original plug that sealed the chamber has fallen away.
mm depth of soil is essential to maintaining a steady nest temperature (Swingland and Coe, 1978).
The nest chamber is dug with each hind foot operating alternately, and large quantities of urine (1–6 litres) and mucus are released when the hole is around 200 mm deep to prevent collapse (Swingland and Coe, 1978). The nesting cavity is ‘sock-shaped’, with the clutch glued together with mucus in the ‘heel’ of the sock to prevent eggs rolling into the ‘toe,’ a cavity left open for the hatchling tortoises to crawl into (Swingland and Coe, 1978). One hind foot gently spreads the eggs, which can form multiple egg layers (Swingland and Coe, 1978; Gerlach, 2004). The chamber is then back-filled with sediment (nest plug) (Swingland and Coe, 1978). The high mucus content of the urine soaks the area around the nesting chamber and forms a concrete-hard encasement protecting the eggs from predators and also as a control over humidity and temperature, which remains between 27–30°C (Swingland and Coe, 1978); any increase in temperature would potentially produce all female clutches (Swingland, 1991). Most nesting chambers (82%) are at a depth of 300–400 mm, as deep as the full hind leg length (Swingland and Coe, 1978; Gerlach, 2004). The parents play no further part in the process.
Hatchlings remain underground for 10–20 days or up to four weeks after hatching, relying on the yolk sac for nutrition for 5–7 days before it is absorbed, and break through to the surface at night by digging (Bourn, 1977; Swingland and Coe, 1978). Hatching occurs just prior to the first rains, which prevents drowning of nests and fungal attack due to increased humidity and provides new plant growth for the emergent hatchlings (Swingland and Coe, 1978; Gerlach, 2004, 2014). Depending on environmental conditions, females may lay more than one clutch per season (Gerlach, 2004), but recruitment can cease if the population density reaches capacity (Swingland, 1989b).
Galapagos.—Divided into 15 species or subspecies, with four now extinct, Galapagos tortoises have a combined present day population of ~19,000 animals, reduced to just over 3000 by 1974, compared with an estimated population of around 250,000 upon discovery in the early 17th century (Swingland, 1989a; Caccone et al., 1999, 2002; Ciofi et al., 2006). The nesting behaviour of Galapagos tortoises is similar to that of Aldabra tortoises. The egg-laying season occurs in July–November, when females may travel several kilometres to reach a suitable nesting area on a dry, sandy coast, which must have at least 300 mm depth of sandy soil (de Vries, 1984; Swingland, 1989a). The
hind legs excavate a deep cylindrical chamber, and the tortoise lays up to 17 eggs within (Swingland, 1989a; Rostal et al., 1998). The digging process is facilitated by wetting the soil with urine and, after laying, the cavity is plugged with urine-saturated soil and sealed by pressing down with the plastron (Macfarland et al., 1974). Galapagos tortoise nest in beach sand and in a small area, concentrating eggs and hatchlings, which makes them more vulnerable to predation (Macfarland et al., 1974).
Egg and clutch size. Rodrigues tortoise eggs are spherical and almost all uniform in size (Table 1), but with a few exceptions. As seen in C. indica eggs (Bour et al., 2014a), one of the Rodrigues eggs is ellipsoidal, while another two are anomalies, being extremely small (Fig. 6C). However, a number of tortoise species produce these abnormal eggs (Cooper, 1983). As two
Julian P. Hume et al.110
Figure 6. Three variations of egg shapes and sizes of Cylindraspis vosmaeri found at Petit Butte on Rodrigues Island. A single egg (FB32) is ellipsoid (A), with almost all other eggs spherical in shape (B). Two extremely small eggs (C) are abnormalities. Scale bar = 10 mm.
sympatric Cylindraspis tortoise species inhabited Rodrigues, a comparison of physical remains of the animals themselves is necessary to identify the eggs. The Rodrigues Saddleback, C. vosmaeri, is known from a stuffed male (MNHN 1883.558), the only known skin specimen of an adult Cylindraspis tortoise, and there are a number of carapaces and plastrons referable to both species that originate from slaughtered specimens or those that died in captivity (Gerlach, 2004; Bour, 2005; Bour et al., 2014a). Two hatchlings (species indeterminate) are also preserved in spirit (Bour et al. 2014a). Subfossil specimens comprise complete carapaces, including some with scutes, as well as huge quantities of bones, almost all from caverns on the Plaine Corail (Hume, 2005, 2014a). Cylindraspis vosmaeri reached 1100 mm in straight carapace length (a linear measurement taken from the proximal nuchal scute and distal supracaudal scute) in the male, whereas the female was much smaller at 560–680 mm (Gerlach, 2004; Bour et al., 2014a), a common sexually dimorphic character in giant tortoises (Fritts, 1984; Gerlach, 2004; Bonin et al., 2006; Stanford, 2010). Cylindraspis peltastes was around half the size of C. vosmaeri, ranging from 390–410 mm in straight carapace length
(Gerlach, 2004; Bour et al., 2014a). Aldabra tortoises are similar in straight carapace length (450–1230 mm) (Gerlach, 2004) to C. vosmaeri, as is the size range of eggs: Cylindraspis 51 mm, Aldabrachelys 48–55 mm (Table 1; Terahara and Moriyama, 1998). We therefore consider the eggs found at Petit Butte most likely referable to C. vosmaeri. Clutch size in Aldabra tortoises ranges from 5–15 in the wild (Swingland and Coe, 1978), with Galapagos species laying from 5–17 eggs (Swingland, 1989a). The clutch size of C. vosmaeri ranged from 8–13 in the three nest cavities discovered. Aldabra tortoises lay eggs on top of each other, and there is evidence to suggest that the top layer hatches first (Swingland and Coe, 1978), which may have also been the case for the multi-layered egg clutches of C. vosmaeri.
Tortoise egg and hatchling predators. A phalanx (III-I) of the flightless Rodrigues Rail Erythromachus leguati Milne-Edwards, 1874, a known predator of tortoise eggs (Hume, 2019), was found in tortoise eggshell layer PB3 (25–30 cm), confirming its association with them (Fig. 7). Tafforet (1625–26) gave the only description of the rail’s diet and stated:
Discovery of the First Cylindraspis Giant Tortoise Nesting Site on Rodrigues Island 111
Figure 7. A reconstruction of Rodrigues giant tortoises on the Plaine Corail, based on a unique stuffed Saddleback Tortoise Cylindraspis vosmaeri male (MNHN 1883.558; centre left) and a complete carapace of Domed Tortoise C. peltastes (MNHN 7831; front) (see Bour et al. 2014a). The Rodrigues rail Erythromachus leguati, a predator of tortoise eggs and young, forages amongst the tortoises. Illustration by JPH.
“There is another kind of bird, the size of a young hen, which has the beak and legs red. […] They generally feed on the eggs of the land tortoises, which they take from the ground, and it makes them so fat that they often have difficulty running.”
[translation from Hume, 2019]
The rail was likely an opportunistic predator, feeding on tortoise eggs and young during the tortoise-breeding season and on invertebrates and sea bird colonies at other times of the year (Hume, 2019). The rail disappeared from Rodrigues between 1750 and 1760, around a decade after the introduction of cats, which are lethal predators of birds, especially flightless island species (Hume, 2017, 2019).
Other possible terrestrial predators of tortoise eggs and young were the extinct Rodrigues Starling, Necropsar rodericanus Günther & Newton, 1879, which certainly scavenged tortoise carcasses (Tafforet, 1625–26; Hume, 2014b), and the Rodrigues Night Heron, Nycticorax megacephalus Milne-Edwards, 1874, a predator of the large, 23-cm long Newton’s Day Gecko, Phelsuma edwardnewtoni J. Vinson & J.-M. Vinson, 1969, and clearly capable of taking hatchling tortoises. Two species of frigatebird, Greater Frigatebird Fregata minor (Gmelin, 1789) and Lesser Frigatebird F. ariel (G.R. Gray, 1845) were once resident on Rodrigues (Cheke & Hume, 2008). Both species prey on marine turtle hatchlings (Safford and Hawkins, 2013), and have been reported to do so on Rodrigues (Leguat, 1708: 66–67). Tortoise hatchlings exposed on the coast may have been equally vulnerable.
Terrestrial crabs that predominantly or temporarily inhabit dry habitats on Mauritius and Rodrigues are now almost extirpated on the mainland, but prior to human settlement there was a great density and diversity of species (Hume, 2005). On Rodrigues, the large, carnivorous and scavenging species Geograpsus grayi (Milne-Edwards, 1853), Ocypode cordimanus Latreille, 1818, and the hermit crab Coenobita rugosa Milne-Edwards, 1837 once occurred in woods and at some distance from the shore (Miers, 1879). Their diet consists of other crabs, tortoise and turtle young, and they also scavenge carcasses (Alexander, 1979; Bourn and Coe, 1979), so they would likely have been predators of hatchling Rodrigues tortoises.
Decline and extinction. The wasteful extermination of tortoises in the Mascarenes is testament to human greed and short-sightedness, but over-hunting itself may not have been the coup de grace for the tortoises, especially on Rodrigues. Introduced pigs that became feral on Mauritius and lowland Réunion wreaked havoc
on tortoise eggs and young (Pitot, 1905; Lougnon, 2006; Cheke and Hume, 2008), as they do in the Galapagos (Hoeck, 1984; Cruz et al., 2005), but this was not the case on Rodrigues. Pigs were first introduced to the island around 1795, when the tortoises were all but extinct (Cheke and Hume, 2008; Bour et al., 2014a). Black Rat, Rattus rattus (Linnaeus, 1758), are particularly devastating to Galapagos tortoises, preying on hatchlings and juveniles 1–2 years old (Swingland, 1989a), but they are less of a threat on Aldabra, although they will take tortoise hatchlings (Swingland, 1989b). On Rodrigues, rats were abundant in 1691 (Leguat, 1708), so they were introduced much earlier (Hume, 2019), but they do not appear to have overly affected the tortoise population. Atkinson (1985, 2000) showed that on islands where land crabs are abundant, ground-nesting birds commonly withstand rats, as they are pre-adapted to this functionally similar predator; Rodrigues rail survived alongside rats for decades (Hume, 2019). Therefore, tortoise hatchlings may have been similarly adapted. On Aldabra, female tortoises take around 17–23 years to reach maturity (Swingland and Coe, 1979), so a reproductive crash and noticeable decline in numbers of Mascarene tortoises would take five or ten years (Cheke and Hume, 2008). The vast numbers of tortoises reported by Tafforet in 1725–26 and later collectors up to the 1760s (Pingré, 1763; North-Coombes, 1994; Cheke and Hume, 2008) confirm that the majority of young grew to maturity.
Although it was primarily over-collecting for meat that decimated the tortoises on Rodrigues (North-Coombes, 1994), recruitment may still have continued throughout this period and replenished the population, with juveniles taking advantage of space made available after removal of adults (Cheke and Hume, 2008). As on Aldabra (Swingland, 1989b), recruitment may have ceased once the carrying capacity of tortoises was reached. However, after the establishment of cats around 1750 or earlier, survival of hatchling tortoises drastically diminished (Cheke and Hume, 2008). Both Pingré (1763) and Marragon (in Bour et al., 2014b) blamed cats entirely for the demise of the Rodrigues tortoises. Furthermore, Cylindraspis tortoises had extremely thin shells (Arnold, 1979), which would have made them more vulnerable to cat predation. Once recruitment stopped, and with the large scale, continual harvesting of surviving adult tortoises, the population crashed. After the settlement began in 1735, tortoise collecting was still lucrative by 1761 (Pingré, 1763), with 30,000 tortoises exported in four vessels in 1759 alone (Günther, 1898; North-Coombes, 1994). Due to
Julian P. Hume et al.112
their thin shells and hideous transportation conditions, with tortoises stacked one on top of the other, of 21,000 tortoises taken in six voyages during 1759–61 just over 50% survived the voyage alive (Milne-Edwards, 1875). The horticulturist and botanist Pierre Poivre (Froberville, 1848; Stoddart and Peake, 1979) noted that 4000–5000 animals per year in the period 1750–70 were exported, each animal with an average mass of 9 kg. By 1767, Commander Yves Julienne and five slaves remained on the island, but with the continued shortage of fresh meat on Mauritius, the Rodrigues settlement increased to 23 with orders from Mauritius, specifically to collect tortoises (North Coombes, 1971). The men scoured the island for the remaining animals, and a consignment of 1215 big carosses, the local name for the large saddleback species C. vosmaeri, was shipped to Mauritius (North-Coombes, 1971, 1994). This was the last mention of the saddleback. Just a year later, it was difficult to collect just a few hundred animals for each of two shipments, so in 1769 the settlement was withdrawn, bar a soldier and a few slaves left to husband the surviving tortoises (North-Coombes, 1971, 1994). Although data are lacking about the last remaining individuals, North-Coombes (1994) estimated the collection of around 2000 tortoises in 1768–71, adding a few subsequent smaller shipments to the total and any left alive on Rodrigues. Numbers must have been very small, as a solitary individual collected on the Plaine Corail in 1786 is the penultimate record (Hume et al., 2015). In 1791, the few remaining settlers returned to Mauritius and left the island temporarily abandoned (North-Coombes, 1971), but the tortoise population on Rodrigues was by now beyond recovery.
The last observation of a living Rodrigues tortoise was by Philibert Marragon in 1795, who had started his term as administrator in 1794 (North-Coombes, 1971; Bour et al., 2014b). In a final statement on the matter, he not only bemoaned the fate of the species, but also the loss of a source of fresh meat, the ultimate reason for the tortoise’s demise:
“The tortoises formerly so common seem to be completely destroyed. In more than a year since I have been here I have only seen two, and those in almost inaccessible gorges. This is a great shame for, in addition to their goodness, they would have been a great succour in this land totally without butcher’s meat.”
Discussion
The nesting site at Petit Butte proved to be exceptional in terms of preservation and abundance of fossil material, which adds much to our knowledge about the nesting behaviour of Cylindraspis tortoises. The nesting season runs from June–September on Aldabra and July–November in the Galapagos, coinciding with the dry season and just before the start of the wet season (Swingland and Coe, 1978; Swingland, 1989b). The dry season (austral winter) on Rodrigues, the driest of the Mascarene Islands, extends from May–November, with rainfall averaging from 800 mm on the Plaine Corail in the coastal southwest to > 1600 mm in the central-west uplands (Cheke and Hume, 2008). Temperatures reach a mean summer temperature of 25.9°C and the mean winter temperature is 22.3°C (Mauritius Meteorological Services, 2020). Presumably, Rodrigues tortoises nested during the dry season, with the hatchlings emerging just before or in November, ready for the wetter and warmer austral summer and in time to benefit from new plant growth. As on Aldabra, the hatchlings may have emerged at night to avoid the attention of diurnal predators, which on Rodrigues included a rail, night heron, starling, and two species of frigatebirds.
The location of the nesting site appears to be one chosen specifically for the essential 300 mm depth of sandy, free-draining soil and within close proximity to the shoreline. The choice of coastal sand for nesting may also have been in response to abundant sandy soil deposits, maximising sunlight and taking advantage of associated higher temperatures, as forested areas inland would be cooler; thus, egg temperature would be lower (Cheke and Hume, 2008). Some shade is necessary for Aldabra tortoise nest chambers, so if the same applied to Cylindraspis, the original dense vegetation on Rodrigues provided sufficient protection, albeit stunted on the Plaine Corail (Pingré, 1763; Strahm, 1989). The presence of extinct and endangered terrestrial snails in the sediments that are open, exposed, coastal scrub inhabitants (Griffths and Florens, 2006) strongly confirms that this type of habitat was preferred for nesting. As in Aldabra tortoises (Swingland and Coe, 1978), the same female may have returned to the same nesting site, relocating the precise location by detecting the odour, and judging by the density of eggshell fragments (30 mm depth), these return events probably lasted decades. The nesting chamber was dug to a depth of approximately 300 mm using the hind feet, as in Aldabra and Galapagos tortoises (Swingland and Coe, 1978; Swingland, 1989a; Gerlach, 2004, 2014), and the loose sandy soil intermixed with urine and mucus to make
Discovery of the First Cylindraspis Giant Tortoise Nesting Site on Rodrigues Island 113
a cement to harden the finished chamber. Residue of the urine and cement are still present at the site (Fig. 2). The intact chambers were situated 2 and 2.5 m apart, but the presence of broken eggshell elsewhere from successful hatching suggests that destruction of all other chambers was due to subsequent nest excavation. The nest chamber is sock-shaped and with a plug, as in Aldabra tortoise (Swingland and Coe, 1978) (Fig. 4), and measured from 360–700 mm in length and 150–300 mm in depth, which allowed an underground area for hatchling tortoises to enter; thus, chamber size was variable and dependent on the size of the tortoise. Clutch size falls within those recorded on Aldabra and Galapagos (Swingland and Coe, 1978; Swingland, 1989a,b), ranging from 8–13 in C. vosmaeri, but this may vary depending on season and possibly the age of the adult. The eggs were spherical and averaged 51 mm in diameter (Table 1), but the presence of one elongate example and two abnormal small eggs, shows that the Rodrigues tortoise was subject to extreme egg size and shape variation. In European Testudo species, the elongate shape of eggs is due to smaller-sized females rather than abnormalities (Hailey and Loumbourdis, 1988). That three unhatched clutches occurred in the nesting area demonstrates that they were at times subject to failure. Clutches primarily fail to hatch on Aldabra because of infertility, with 10–20% of all eggs infertile and 20–40% failing to hatch (Swingland and Coe, 1978); nest flooding, temperature deficiency or fungal attack could also be a cause. It is very fortunate that these clutches remained intact, despite the nesting area presumably turned over for decades, as the dense layer of eggshell fragments elsewhere in the nesting site are indicative of breeding success.
The discovery of this remarkable tortoise-nesting site provides an unprecedented insight into the life history of a tortoise species now extinct for more than two centuries. It is very likely that C. peltastes and the other species on Mauritius and Réunion had similar ecologies, but despite the wealth of fossil material discovered on each island (Hume, 2014a), the nesting site at Petit Butte is unique. The Mascarene endemic genus Cylindraspis is now extinct, and its former existence evidenced only by documented accounts, illustrations, and physical remains. Conversely, the tortoises of Aldabra and most of those in the Galapagos survived the massacre that took place in the Mascarenes, solely because the discovery of these islands by humans occurred much later and conservation efforts prevented potential extinctions (Stoddart and Peake 1979; Swingland, 1989a,b; Bourn et al., 1999; Chambers, 2005).
Acknowledgements. We thank Denis Claude Milazar (landowner), Annabelle Agathe and Monica Young Kan Seong (RRA); Stephen Kirsakye, Jenifer César, Anieta Shan-Yu, and Jean Alfred Bégne (MWF), and Rodney Severy (FLR) for their help in excavating the tortoise nesting site, Jayshree Mungur-Medhi for support and permissions during the excavation, and FLR for support on Rodrigues. We further thank an anonymous reviewer for helpful comments.
References
Alexander, H.G.L. (1979): A preliminary assessment of the role of the terrestrial decapod crustaceans in the Aldabran ecosystem. Philosophical Transactions of the Royal Society B, Biological Sciences 286: 241–246.
Arnold, E.N. (1979): Indian Ocean giant tortoises: their systematics and island adaptations. Philosophical Transactions of the Royal Society B, Biological Sciences 286: 127–145.
Atkinson, I.A.E. (1985): The spread of commensal species of Rattus to oceanic islands and their effects on island avifaunas. In: Conservation of Island Birds, p. 35–81. Moors, P.J., Ed., Cambridge, United Kingdom, International Council for Bird Preservation.
Atkinson, I.A.E. (2000): Introduced mammals and models for restoration. Biological Conservation 99: 81–96.
Bonin, F., Devaux, B., Dupré, A. (2006): Turtles of the World. Baltimore, Maryland, USA, Johns Hopkins University Press. 416 pp.
Bory de Saint-Vincent, J.B.G.M. (1804): Voyage dans les Quatre Principals Îles des Mers d’Afrique. Paris, France, F. Buisson.
Bour, R. (1979): Histoire de la tortue terrestre de Bourbon. Comptes Rendus de l’Académie des Sciences, Paris 25: 97–147.
Bour, R. (1980): Systématiques des tortues terrestres des Îles Mascareignes: genre Cylindraspis Fitzinger 1835 (Reptilia, Chelonii). Bulletin du Muséum National d’Histoire Naturelle, Paris 4(2A): 895–904.
Bour, R. (1981): Histoire de la tortue terrestre de Bourbon. Bulletin de l’Académie de Île de la Réunion 25: 98–147.
Bour, R. (2005): Type specimens of Testudo rotunda Latreille, 1801. Emys 12(6): 23–27.
Bour, R., Mourer-Chauviré, C., Ribes, S. (2014a): Morphology and palaeontological exploration (up to 2000) of the extinct tortoises of the Mascarene islands. In: Western Indian Ocean Tortoises: Ecology, Diversity, Evolution, Conservation, Palaeontology, p. 121–202. Gerlach, J., Ed., Manchester, United Kingdom, Siri Scientific Press.
Bour, R., Frétey, T., Cheke, A.S. (2014b): Philibert Marragon (1749–1826) and the Mémoire sur l’Isle de Rodrigue (1795). Bibliotheca Herpetologica 10(2): 5–32.
Bourn, D. (1977): Reproductive study of giant tortoises on Aldabra. Journal of Zoology, London 182: 27–38.
Bourn, D., Coe, M.J. (1979): Features of tortoise mortality and decomposition on Aldabra. Philosophical Transactions of the Royal Society B, Biological Sciences 286: 189–193.
Bourn, D.M., Gibson, C., Augeri, D., Wilson, C.J., Church, J., Hay, S.I. (1999): The rise and fall of the Aldabran giant tortoise population. Proceedings of the Royal Society of London B, Biological Sciences 266: 1091–1100.
Julian P. Hume et al.114
Caccone, A., Gentile, G., Gibbs, J.P., Fritts, T.H., Snell, H.L., Betts, J., Powell, J.R. (2002): Phylogeography and history of giant Galapagos tortoises. Evolution 56(10): 2052– 2066.
Caccone, A., Gibbs, J.P., Ketmaier, V., Suatoni, E., Powell, J.R. (1999): Origin and evolutionary relationships of giant Galapagos tortoises. Proceedings of the National Academy of Sciences 96(23): 13223–13228.
Chambers, P. (2005): A Sheltered Life: the Unexpected History of the Giant Tortoise. Oxford, United Kingdom, Oxford University Press. 306 pp.
Cheke, A.S. (1987): An ecological history of the Mascarene Islands, with particular reference to extinctions and introductions of land vertebrates. In: Studies of Mascarene Island Birds, p. 5–89. Diamond, A.W., Ed., Cambridge, United Kingdom, Cambridge University Press.
Cheke, A.S., Bour, R. (2014): Unequal struggle – how humans displaced the dominance of tortoises in island ecosystems. In: Western Indian Ocean Tortoises: Ecology, Diversity, Evolution, Conservation, Palaeontology, p. 31–120. Gerlach, J., Ed., Manchester, United Kingdom, Siri Scientific Press.
Cheke, A.S., Hume, J.P. (2008): Lost Land of the Dodo. London, United Kingdom, T. & A.D. Poyser. 464 pp.
Cheke, A.S., Pedrono, M., Bour, R., Anderson, A., Griffiths, C., Iverson, J.B., Hume, J.P., Walsh, M. (2016): Giant tortoises spread to western Indian Ocean islands by sea drift in pre-Holocene times, not by later human agency – response to Wilmé et al. (2016). Journal of Biogeography 44: 1426–1440.
Ciofi, C., Wilson, G. A., Beheregaray, L.B., Marquez, C., Gibbs, J.P., Tapia, W., et al. (2006): Phylogeographic history and gene flow among giant Galápagos tortoises on southern Isabela Island. Genetics 172(3): 1727–1744.
Coe, M.J., Bourn, D., Swingland, I.R. (1979): The biomass production and carrying capacity of giant tortoises on Aldabra. Philosophical Transactions of the Royal Society B, Biological Sciences 286: 163–176.
Cooper, J.E. (1983): Preliminary studies on the eggs of three species of chelonians. Testudo 2(2): 33–35.
Cruz, F., Donlan, C.J., Campbell, K, Carrion, V. (2005): Conservation action in the Galapagos: feral pig (Sus scrofa) eradication from Santiago Island. Biological Conservation 121: 473–478.
Dupon, J.F. (1974): Relation de l’Isle Rodrigue. Text attributed to Tafforet, circa. 1726. Proceedings of the Royal Society of Arts & Sciences of Mauritius 4(1): 1–16.
Freycinet, L.C.D. (1825): Voyage Autour du Monde Exécuté sur les Corvettes de S.M. l’Uranie et la Physicienne Pendant les Années 1817, 1818, 1819, et 1820. Volume 1. Paris, France, Pillet Ainé. 734 pp.
Fritts, T.H. (1984): Evolutionary divergence of giant tortoises in Galapagos. Biological Journal of the Linnean Society 21(1–2): 165–176.
Froberville, E. de (1848): Rodrigues, Galega, Les Sechelles, Les Almirantes, etc. In: Iles de Afrique. Part 3. Iles Africaines de la Mer des Indes. Volume 2, p. 65–114. Avezac, M.A.P. d’, Ed., Paris, France, Firmin Didot Freres.
Gerlach, J. (2004): Giant Tortoises of the Indian Ocean. Frankfurt am Main, Germany, Chimaira. 207 pp.
Gerlach, J. (2014): Ecology and status of Western Indian Ocean Aldabrachelys giant tortoises. In: Western Indian Ocean Tortoises:
Ecology, Diversity, Evolution, Conservation, Palaeontology, p. 229–251. Gerlach, J., Ed., Manchester, United Kingdom, Siri Scientific Press.
Griffiths, O.L., Florens, V. (2006): A Field Guide to the Non-marine Molluscs of the Mascarene Islands (Mauritius, Rodrigues and Réunion) and the Northern Dependencies of Mauritius. Mauritius, Bioculture Press. 185 pp.
Günther, A. (1898): Recent progress on our knowledge of the distribution of the gigantic land tortoises. Proceedings of the Linnean Society 110: 14–29.
Hailey, A., Loumbourdis, N.S. (1988): Egg size and shape, clutch dynamics, and reproductive effort in European tortoises. Canadian Journal of Zoology 66: 1527–1536.
Herbert, W., Nichelson, W. (1780): A New Directory for the East Indies. Fifth Edition. London, United Kingdom, Henry Gregory. 554 pp.
Hermann, J. (1903): Colonisation de l’Ȋle Bourbon et Foundation du Quartier Saint-Pierre. Paris, France, C. Delagrave. 408 pp.
Hoeck, H.N. (1984): Introduced fauna. In: Key Environments. Galápagos, p. 233–245. Perry, R., Ed., Oxford, United Kingdom, Pergamon Press.
Hubbard, C.E, Vaughan, R.E. (1940): The Grasses of Mauritius and Rodriguez. London, United Kingdom, Waterlow & Sons. 127 pp.
Hume, J.P. (2005): Contrasting taphofacies in ocean island settings: the fossil record of Mascarene vertebrates. Monografies de la Societat d’Història Natural de les Balears 12: 129–144.
Hume, J.P. (2014a): Fossil discoveries on Mauritius and Rodrigues. In: Western Indian Ocean Tortoises: Ecology, Diversity, Evolution, Conservation, Palaeontology, p. 203–228. Gerlach, J., Ed., Manchester, United Kingdom, Siri Scientific Press.
Hume, J.P. (2014b): Systematics, morphology, and ecological history of the Mascarene starlings (Aves: Sturnidae) with the description of a new genus and species from Mauritius. Zootaxa 3849(1): 1–75.
Hume, J.P. (2017): Extinct Birds. Second Edition. London, United Kingdom, Helm. 608 pp.
Hume, J.P. (2019): Systematics, morphology, and ecology of rails (Aves: Rallidae) of the Mascarene Islands, with one new species. Zootaxa 4626(1): 1–107.
Hume, J.P., Steel, L., André, A.A., Meunier, A. (2015): In the footsteps of the bone collectors: nineteenth-century cave exploration on Rodrigues Island, Indian Ocean. Historical Biology 27(2): 265–286.
Leguat, F. (1708): Voyages et Avantures de François Leguat & de Ses Compagnons en Deux Îles Désertes des Indes Orientales. Amsterdam, The Netherlands, Jean Louis de Lorme.
Lougnon, A. (2006): Sous le Signe de la Tortue. Voyage Anciens à l’Ile Bourbon, (1611–1725). Saint-Denis, La Réunion, France, Editions Orphie. 284 pp.
MacFarland, C.G., Villa, J., Toro, B. (1974): The Galápagos giant tortoises (Geochelone elephantopus). I. Status of the surviving populations. Biological Conservation 6(2): 118–133.
Mauritius Meteorological Services (2020): Climate of Rodrigues & Islands. Available at: http://metservice.intnet.mu. Accessed on 10 June 2020.
Miers, E.J. (1879): Crustacea. Philosophical Transactions of the Royal Society of London 168: 485–496.
Discovery of the First Cylindraspis Giant Tortoise Nesting Site on Rodrigues Island 115
Milne-Edwards, A. (1875): Nouveaux documents sur l’époque de la disparition de la faune ancienne de l’Île Rodrigue. Annales des Sciences Naturelles 6S(2): 1–20.
Mondini, E. (1990): Les tortues géantes terrestres et le choc de l’histoire. Cahiers d’Outre Mer 43: 555–559.
Moore, J., White, W.M., Paul, D., Duncan, R.A., Abouchami, W., Galer, S.J. (2011): Evolution of shield-building and rejuvenescent volcanism of Mauritius. Journal of Volcanology and Geothermal Research 207: 47–66.
Moree, P.J. (1998): A Concise History of Dutch Mauritius, 1598–1710. London, United Kingdom, Kegan Paul International. 127 pp.
North-Coombes, A. (1971): The Island of Rodrigues. Mauritius, The Standard Printing Establishment. 337 pp.
North-Coombes, A. (1994): Histoire des Tortues de Terre de Rodrigues. Second Edition. Mauritius, Mauritius Printing Specialists. 100 pp.
Pingré, A.-G. (1763): Voyage à l’Isle Rodrigue. Ms 1804. Bibliothéque Ste-Geneviève, Paris, France. 470 pp. [edited and rearranged version of MS 1803 (1761), a day-to-day diary from 1760–1762].
Pitot, A. (1905): T’Eylandt Mauritius. Esquisses Historiques (1598–1710). Précédés d’une Notice sur la Découverte des Mascareignes et Suivies d’une Monographie du Dodo, des Solitaires de Rodrigue et de Bourbon et de l’Oiseau Bleu. Port Louis, Mauritius, Coignet Frères & Cie. 496 pp.
Rostal, D.C., Robeck, T.R., Grumbles, J.S., Burchfield, P.M., Owens, D.W. (1998): Seasonal reproductive cycle of the Galapagos Tortoise (Geochelone nigra) in captivity. Zoo Biology 17: 505–517.
Safford, R.J., Hawkins, A.F.A. (2013): The Birds of Africa. Volume VIII. The Malagasy Region. London, United Kingdom, Christopher Helm. 1024 pp.
Standford, C.B. (2010): The Last Tortoise: a Tale of Extinction in Our Lifetime. Cambridge, Massachusetts, USA, Belknap Press of Harvard University Press. 240 pp.
Stoddart, D.R., Peake, J.F. (1979): Historical records of Indian Ocean giant tortoise populations. Philosophical Transactions of the Royal Society of London B, Biological Sciences 286: 147–161.
Strahm, W. (1989): Plant Red Data Book for Rodrigues. Königstein, Germany, Koeltz Scientific Books. 241 pp.
Swingland, I.R. (1989a): Geochelone elephantopus. Galapagos giant tortoises. The conservation biology of tortoises. Occasional Papers of the IUCN Species Survival Commission 5: 24–28.
Swingland, I.R. (1989b): Geochelone gigantea. Aldabran giant tortoise. The conservation biology of tortoises. Occasional Papers of the IUCN Species Survival Commission 5: 105–108.
Swingland, I.R. (1991): Environmental sex determination, migration, and regulation in the Aldabran giant tortoise. Proceedings of the Desert Tortoise Council Symposium 1987–1991: 43–50.
Swingland, I.R., Coe, M.J. (1978): The natural regulation of giant tortoise populations on Aldabra Atoll: reproduction. Journal of Zoology, London 186: 285–309.
Swingland, I.R., Coe, M.J. (1979): The natural regulation of giant tortoises on Aldabra Atoll: recruitment. Philosophical Transactions of the Royal Society of London B, Biological Sciences 286: 177–188.
Tafforet, J. (1725-26): Relation de l’Isle Rodrigue. Ms, Archives Nationales, Paris, Fonds des Colonies, Correspondence de I’île de France, C4, Volume 12, no 48: ex Ministère de la Marine, no 29, 1.
Temple, R.C., Ed. (1914): The travels of Peter Mundy in Europe and Asia, 1608–1667. Hakluyt Society SII 35(2): 1–437.
Terahara, M., Moriyama, S. (1998): Breeding the Aldabra giant tortoise. Animals & Zoos 12: 404–407.
Vries, T.J. de. (1984): The giant tortoises: a natural history disturbed by man. In: Key Environments: Galapagos, p. 145–156. Perry, R., Ed., Oxford, United Kingdom, Pergamon Press.