This article was downloaded by: [University of Chicago Library] On: 30 September 2013, At: 12:16 Publisher: Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK Journal of Natural History Publication details, including instructions for authors and subscription information: http://www.tandfonline.com/loi/tnah20 Distribution, habitat and food preferences of sympatric high intertidal isopod species Ligia occidentalis and Ligia pallasii (Ligiidae: Oniscidea) Renate Eberl a a Department of Evolution and Ecology, University of California Davis, One Shields Avenue, Storer Hall, Davis, CA, 95616, USA Published online: 16 Jul 2012. To cite this article: Renate Eberl (2012) Distribution, habitat and food preferences of sympatric high intertidal isopod species Ligia occidentalis and Ligia pallasii (Ligiidae: Oniscidea), Journal of Natural History, 46:29-30, 1779-1797, DOI: 10.1080/00222933.2012.700334 To link to this article: http://dx.doi.org/10.1080/00222933.2012.700334 PLEASE SCROLL DOWN FOR ARTICLE Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) contained in the publications on our platform. However, Taylor & Francis, our agents, and our licensors make no representations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of the Content. Any opinions and views expressed in this publication are the opinions and views of the authors, and are not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon and should be independently verified with primary sources of information. Taylor and Francis shall not be liable for any losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoever or howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use of the Content. This article may be used for research, teaching, and private study purposes. Any substantial or systematic reproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in any form to anyone is expressly forbidden. Terms &
Transcript
This article was downloaded by: [University of Chicago Library]On: 30 September 2013, At: 12:16Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registeredoffice: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
Journal of Natural HistoryPublication details, including instructions for authors andsubscription information:http://www.tandfonline.com/loi/tnah20
Distribution, habitat and foodpreferences of sympatric highintertidal isopod species Ligiaoccidentalis and Ligia pallasii (Ligiidae:Oniscidea)Renate Eberl aa Department of Evolution and Ecology, University of CaliforniaDavis, One Shields Avenue, Storer Hall, Davis, CA, 95616, USAPublished online: 16 Jul 2012.
To cite this article: Renate Eberl (2012) Distribution, habitat and food preferences of sympatrichigh intertidal isopod species Ligia occidentalis and Ligia pallasii (Ligiidae: Oniscidea), Journal ofNatural History, 46:29-30, 1779-1797, DOI: 10.1080/00222933.2012.700334
To link to this article: http://dx.doi.org/10.1080/00222933.2012.700334
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the“Content”) contained in the publications on our platform. However, Taylor & Francis,our agents, and our licensors make no representations or warranties whatsoever as tothe accuracy, completeness, or suitability for any purpose of the Content. Any opinionsand views expressed in this publication are the opinions and views of the authors,and are not the views of or endorsed by Taylor & Francis. The accuracy of the Contentshould not be relied upon and should be independently verified with primary sourcesof information. Taylor and Francis shall not be liable for any losses, actions, claims,proceedings, demands, costs, expenses, damages, and other liabilities whatsoever orhowsoever caused arising directly or indirectly in connection with, in relation to or arisingout of the use of the Content.
This article may be used for research, teaching, and private study purposes. Anysubstantial or systematic reproduction, redistribution, reselling, loan, sub-licensing,systematic supply, or distribution in any form to anyone is expressly forbidden. Terms &
Journal of Natural HistoryVol. 46, Nos. 29–30, August 2012, 1779–1797
Distribution, habitat and food preferences of sympatric high intertidalisopod species Ligia occidentalis and Ligia pallasii (Ligiidae: Oniscidea)
Renate Eberl*
Department of Evolution and Ecology, University of California Davis, One Shields Avenue, StorerHall, Davis, CA 95616, USA
(Received 21 June 2011; final version received 1 June 2012; printed 25 July 2012)
The distribution, habitat and food preferences of two partially sympatric high inter-tidal isopods, Ligia pallasii and Ligia occidentalis, were investigated. A survey alongthe Eastern Pacific coast indicated species-specific site preferences as evidenced bya mosaic pattern of distribution with few sites of co-occurrence. Ligia pallasii pre-ferred sites with rocky cliffs and caves that may provide a thermal refuge for thenorthern species, whereas L. occidentalis apparently is a habitat generalist of rockyshores. Differences in reproductive patterns exist: L. occidentalis reproduced onceannually in spring and summer, but gravid females of L. pallasii were observedthroughout the year. Feeding experiments did not indicate any significant differ-ences in diets; both isopod species fed mainly on seaweed and diatoms but preferredrare protein sources when available.
Delimiting factors that determine distribution patterns of animals has been a long-standing aim of ecological research (Elton 1946; Andrewartha and Birch 1954;Hutchinson 1959). Congeneric species often have very similar ecologies and so may beexpected to show intense interspecific competition (Connell 1961; MacArthur 1972;Schoener 1974). Partitioning of resources – e.g. food (Johnson 1986; Genner et al.1999) or space (Connell 1961) – can reduce interspecific competition and allow forthe coexistence of species that are occupying different niches (Vandermeer 1972). Oneof the best known examples of spatial resource partitioning between closely relatedspecies comes from the rocky intertidal, where the barnacle Semibalanus balanoidesoutcompetes Chthamalus stellatus at lower tide heights but is unable to survive athigher tide heights (Connell 1961); this interaction leads to a clear vertical zonation.A combination of biotic, e.g. competition and predation (Connell 1961; Paine 1966),and abiotic factors, e.g. thermal tolerance and desiccation resistance (Somero 2002), isresponsible for these vertical zonation patterns regularly found in intertidal organisms.Habitat segregation at small scales leads to the coexistence of closely related species atlarger scales and thereby enhances diversity. In most cases biotic factors are consideredmore important in the lower intertidal zone, whereas abiotic factors are consideredto play a larger role in the upper intertidal zone (Connell 1961; Paine 1974; Somero2002; Stillman 2002). These studies rarely extend to the highest reaches of the intertidal
zone where conditions are particularly harsh, and only a few organisms, among themlimpets, littorinid snails and isopods, can withstand extended dry periods.
Both terrestrial and aquatic isopod communities often contain sympatric speciesthat share similar available food sources, but may differ in feeding or microhabitatpreferences (Franke and Janke 1998; Zimmer et al. 2000, 2002). Oniscidean isopodsare some of the most abundant macro-decomposers in both terrestrial and intertidalfood chains (Pennings et al. 2000; Zimmer and Topp 2000; Zimmer et al. 2002). Whilethe majority of oniscidean isopods are found in a diversity of terrestrial habitats, thegenus Ligia (Ligiidae, Oniscidea) inhabits the higher portion of the marine intertidalzone, the same sea–land transition zone where terrestrial isopods are believed to haveevolved from ligiid-like ancestors (Abbott 1939; Carefoot 1998).
Most Ligia inhabit rocky shorelines from the splash zone to a few meters aboveand feed on washed up seaweed (wrack) and diatoms in the intertidal zone (Penningset al. 2000; Colombini and Chelazzi 2003). Wrack deposited in the intertidal zonerepresents an important subsidy to marine intertidal habitats and serves as food formany intertidal invertebrates, prominent among them isopods (Pennings et al. 2000).
In the supralittoral zone of the Eastern Pacific Ligia pallasii and Ligia occidentalisoccur in sympatry over a portion of their respective ranges (Schmalfuss 2003) and canbe morphologically distinguished in the field (Figure 1; Brusca et al. 2001). Ligia pal-lasii has a range that extends from the Aleutian Islands, Alaska to central California,L. occidentalis can be found from central California to Mexico (Morris et al. 1980;Schmalfuss 2003). However, little is known about the exact ranges and the patternsof abundance of these two species. Ligia spp. abundances may change across distribu-tional ranges. For example, according to the “abundant centre hypothesis” (Sagarinand Gaines 2002) species are expected to be more abundant toward the centre of therange with numbers declining toward range edges.
Spatial niche segregation is thought to occur in areas of range overlap withL. pallasii occupying space higher in the supralittoral zone than L. occidentalis (Abbott
A B C
Figure 1. Ligia occidentalis (A) and Ligia pallasii (B, C) can be distinguished by the distancebetween the eyes and the shape of the caudal peduncle of the uropod. Sexual dimorphism isonly present in L. pallasii (B, female; C, male). All scale bars represent 10 mm. Photos: J. Sones.
Dow
nloa
ded
by [
Uni
vers
ity o
f C
hica
go L
ibra
ry]
at 1
2:16
30
Sept
embe
r 20
13
Journal of Natural History 1781
1939; Morris et al. 1980; Ricketts and Calvin 1985); this proposition appears to havebeen made based on local observations rather than any extensive surveys of Ligia spp.distributions. It is unknown how often both species co-occur within the same area andwhether or not they share similar food sources. The natural diet and food preferencesof L. pallasii have been the subject of a few studies (Carefoot 1989; Pennings et al.2000; Zimmer et al. 2002), but the diet of L. occidentalis has never been investigated.How these two species use resources (food and space) in areas of range overlap is notknown.
This study is the first large-scale survey along the Eastern Pacific coast that inves-tigates the distribution of L. pallasii and L. occidentalis. Monthly monitoring of sitesin the centre of range overlap was combined with the large-scale survey to determinetemporal stability of distribution patterns. A series of feeding experiments were con-ducted to determine potential differences in feeding preference and differences in foodchoice between the two Ligia species.
Materials and methods
Large-scale survey of Ligia spp. distribution in the Eastern PacificTo identify potential survey sites and estimate areas of range overlap betweenL. pallasii and L. occidentalis, museum data bases (California Academy of Sciences,Smithsonian Museum of Natural History, Santa Barbara Museum of NaturalHistory), literature (Richardson 1905; Morris et al. 1980; Schmalfuss 2003) as wellas online resources (OBIS, www.iobis.org) were searched.
Field surveys were conducted between December 2006 and January 2010 alongroughly 1500 km of the Eastern Pacific coastline from Northern Oregon (Escola StatePark, 45.92◦ N, 123.97◦ W) to Southern California (Ventura, 34.25◦ N, 119.27◦ W).Based on the limited records obtained from literature and museum database searches,the area of range overlap between L. pallasii and L. occidentalis was estimated to occurbetween Cape Blanco (42.84◦ N, 124.56◦ W) and Monterey (36.61◦ N, 121.90◦ W) andso the majority of survey effort (78% of a total of 132 sites) was focused there. Initialsites were selected according to records of species occurrence (see above); additionalsites were chosen based on safety and ease of access, as well as suitability of habitat(presence of rocky substrate). Geographical coordinates were obtained with a hand-held Global Positioning System receiver (ETrex, Garmin Ltd, Olathe, KS, USA), andeach site was assessed for the following physical characteristics: presence or absence ofa cliff, caves or deep cracks, freshwater drip, as well as for the presence and abundanceof algal wrack, common high intertidal algae (Enteromorpha intestinalis, Ulva spp.,Pelvetiopsis spp.), and invertebrate groups (grapsid crabs, staphilinid beetles, mites andlittorine snails). Then, a survey was performed of 50–100 m of shoreline in a 15-minutetimed interval for the presence of L. pallasii and L. occidentalis, and the number ofindividuals in each species found during this timeframe was recorded. At each site,each Ligia species was noted as either absent (no individuals found after 15 minutes),present (1–20 individuals) or abundant (> 20 individuals). Although habitat charac-teristics in the field often determined the speed of movement of the observer, everyattempt was made to survey the whole extent of the chosen coastal stretch evenly andto survey the entire supralittoral zone as far as was possible. As both species escapepredation and heat stress by hiding under rocks and in crevices, the numbers observed
Dow
nloa
ded
by [
Uni
vers
ity o
f C
hica
go L
ibra
ry]
at 1
2:16
30
Sept
embe
r 20
13
1782 R. Eberl
probably underestimate the true number of individuals present. A single observerconducted surveys at all sites to maximize consistency of observations between sites.A subset of sites (n = 49) was sampled multiple times to assess consistency of speciesdistribution over time, and scores of abundance from the multiple visits were averaged.
Sites were analysed based on the presence, absence or coexistence of L. pallasiiand L. occidentalis. The relationship between different site characteristics (presence orabsence of rocky cliffs, caves, freshwater seeps) and the occurrence of L. pallasii (LP)and L. occidentalis (LO) within their respective ranges were evaluated with Pearson’schi-square tests (Sokal and Rohlf 1995). Correlation between abundance of specieswith latitude was determined to detect patterns in accordance with the abundant centrehypothesis. Only a portion of the range was surveyed for each species: for L. pallasii,the northernmost survey sites are roughly equivalent to the centre of the range, whereasfor L. occidentalis, the southernmost survey sites are roughly equivalent to the centreof the range. In the case of decline of abundance towards the range edges, decline inspecies presence and abundance would be found at southern sites for L. pallasii and atnorthern sites for L. occidentalis.
Monitoring of sites in Bodega BayTo determine persistence of species distribution and variation of abundance over time,two sites were selected with different habitat characteristics in the centre of the rangeoverlap: Mussel Point (MP) and Bodega Harbor (BH), both at the Bodega MarineReserve (38◦18′ N, 123◦03′ W). MP and BH had been previously indentified as sitesoccupied by L. pallasii and L. occidentalis, respectively, and were monitored monthlyfrom May 2007 to December 2010. MP is a semi-protected area at the outer coastthat is characterized by steep rocky cliffs with several small caves; BH is a protectedsite within Bodega Bay and consists of small rocks and pebbles on sandy to muddysubstrate and lacks cliffs.
Due to the different types of habitat, different surveying methods were deemednecessary. At MP areas of rock wall (inside and outside the caves) were subdividedinto six sections of roughly similar size, for each section the percent algal coverage wasestimated and 2-minute timed counts of Ligia were performed for each section. At BHthe area was divided into 10 equal plots of 40 × 2 m length (lower bound roughly athigh tide line) and three plots were chosen at random for monitoring. Within theseplots five 0.25-m2 squares were sampled haphazardly, avoiding areas with rocks largerthan the square. After removal of algal wrack, rocks were carefully turned over andthe Ligia were counted. At both sites, temperature loggers (i-buttons DS1921G, DallasSemiconductor; Maxim Integrated Products, San Jose, CA, USA) measured temper-ature at hourly intervals. For each site the mean average temperature per month aswell as the monthly average of the daily maximum and minimum temperatures werecalculated and plotted.
Mesocosm experimentFeeding of the two Ligia species on algal biofilm was investigated in a mesocosmexperiment. Small mesocosms (15 × 15 × 25 cm, ntotal = 48) were placed atan angle of 30◦ in two outdoor plastic tanks with flow-through seawater. Eachmesocosm was covered with a window screen (to avoid the escape of isopods
Dow
nloa
ded
by [
Uni
vers
ity o
f C
hica
go L
ibra
ry]
at 1
2:16
30
Sept
embe
r 20
13
Journal of Natural History 1783
and to provide shade). Seawater entered each mesocosm from the top and exitedthrough four holes (5-cm diameter) covered with window screen. Before the startof the experiment, seawater was allowed to flow for 10 days to allow for thedevelopment of algal biofilm. Mesocosms were assigned randomly to one of fourtreatments (12 replicates each): 10 L. pallasii (LP), 10 L. occidentalis (LO), fiveL. occidentalis + five L. pallasii (LO + LP) or no isopods (none). Mesocosmswere checked daily. After 21 days isopods were removed and the algal biofilm ineach container was collected by careful scraping with a spatula, dried at 60◦C for72 hours and dry weight was determined on a Mettler AE240 analytical balance(Mettler-Toledo Inc., Columbus, OH, USA). A one-way analysis of variance (ANOVA)was used to determine significant differences in algal dry weight between thetreatments.
Food choice experiment IFood preference had been previously determined for L. pallasii from Vancouver Island(Pennings et al. 2000). To determine preferences of L. occidentalis and L. pallasii andto ascertain that feeding preferences for L. pallasii do not change with location, a feed-ing preference experiment was conducted following methods described in Pennings etal. (2000) with a subset of seaweed species used in their study. The food choices in thepresent study consisted of Nereocystis luetkeana, Costaria costata, Ulva spp., Mazaellaflaccida and Fucus gardneri, species that are readily available either fresh or as algalwrack to both Ligia species around the Bodega Marine Reserve. Since it has been pre-viously shown that Ligia prefers algal wrack over fresh seaweed (Pennings et al. 2000),wrack was collected near the Bodega Marine Reserve. Seaweeds were standardized bycutting out 16-mm diameter discs with a hole punch. Ligia were kept individually inplastic cylindrical containers (95 mm in diameter and 75 mm in height) with loose fit-ting lids. Fresh seawater was available in a 5-ml dish and replaced daily. Assays werecontinued for up to 3 days, and discs were progressively removed when ≥ 70% of a discwas eaten to determine the rank order of preference for all five seaweeds. Replicates inwhich animals did not feed within 3 days were dropped.
Food choice experiment IIThe experiment described above is limited to algae that have a blade width of at least16 mm, so a second experiment with four different food types: diatoms, green algae(Ulva spp.), brown algae (Pelvetiopsis limitata) and fish (Shaved Bonito flakes; Ninben,Nihonbashi, Japan) was performed. Diatoms and the two macroalgae represent threelower quality foods that are available constantly, albeit at varying amounts, to bothLigia species; the fish flakes represent an occasional input of high protein in the formof fish carcasses. Ligia pallasii and L. occidentalis were collected by hand at BodegaMarine Reserve and kept without food for 3 days in the laboratory before the startof the experiment. Diatoms were collected from a sea table at Bodega Marine Reserve,Ulva spp. and P. limitata were collected from the intertidal zone around Bodega MarineReserve. Diatoms and algae were dried at 60◦C for 48 hours, and all foods were groundto a fine powder. Gels of the different food types containing 3 g molecular biologygrade agarose (Denville Scientific, Metuchen, NJ, USA), 2 g powdered dried food and200 ml deionized water were poured; a control contained only agarose and water. All
Dow
nloa
ded
by [
Uni
vers
ity o
f C
hica
go L
ibra
ry]
at 1
2:16
30
Sept
embe
r 20
13
1784 R. Eberl
gels were cut into 6-mm squares, kept on moist filter paper to mimic conditions duringthe experiment, and weighed on a Mettler AE240 analytical balance (Mettler-ToledoInc., Columbus, OH) before and at the end of the experiment.
The bottoms of 60 plastic cylindrical containers (95-mm in diameter and 75-mm inheight) were fitted with Whatman filter paper and divided into two sections, on one halfthe five gel squares (four different food types and control) were randomly placed in fiveequally spaced positions, the other half contained a small Petri dish with 2 ml seawaterthat was replaced daily. Individual L. pallasii and L. occidentalis (n = 25 per species)were randomly assigned to containers (one individual per container), 10 containerswithout Ligia were used as controls. Containers were observed 11 times over a periodof 70 hours and it was determined on what food type, if any, they were feeding. Ananimal was considered to be feeding if its head was on top of a square of food at thetime the container was checked. Containers were cleaned daily by pipetting seawaterand removing dissolved fecal pellets. At the end of the experiment each individual foodsquare was blotted dry and weighed. The amount of food eaten for each food type wasdetermined by subtracting the final from the initial weight. Welch ANOVA and Tukey–Kramer post hoc tests were used to test for statistical differences in the amount of eachfood group consumed for each Ligia species.
Results
Large-scale surveyThe survey revealed a mosaic pattern of distribution for both L. pallasii and L. occi-dentalis and a few sites where both L. pallasii and L. occidentalis occurred togetherand were abundant. Patterns of species occurrence showed temporal consistency. Ofthe 104 sites (out of a total of 132) located between South of Cape Blanco (42.83◦ N,124.56◦ W) and Monterey (36.61◦ N, 121.90◦ W), the area of presumed range overlap,40% (n = 49) were visited more than once and none showed a reversal in species occur-rence during the survey period (December 2006 to January 2010). Ligia pallasii and L.occidentalis were each found at 59 of the 132 sites surveyed and occurred together atonly 17 sites (16.3% of sites within the presumed range overlap). In 14 of these sitesthe abundance of L. pallasii was greater than that of L. occidentalis, sometimes oneto two orders of magnitude greater. However when the relative abundances of the twospecies were compared, both species scored approximately evenly (L. occidentalis ninesites and L. pallasii eight sites with greater relative abundance). At 22 of all sites nei-ther Ligia species was found despite the fact that the habitat at these sites was deemedsuitable.
Each species was encountered only in areas within its presumed respective range,and both species were only found between Cape Blanco and Monterey Bay. Of the15 sites that were visited north of Cape Blanco, no Ligia occidentalis were encoun-tered, but 13 sites were occupied by L. pallasii. Of the 13 sites south of MontereyHarbor, no L. pallasii were found, but 10 of these sites were occupied by L. occidentalis(Table 1).
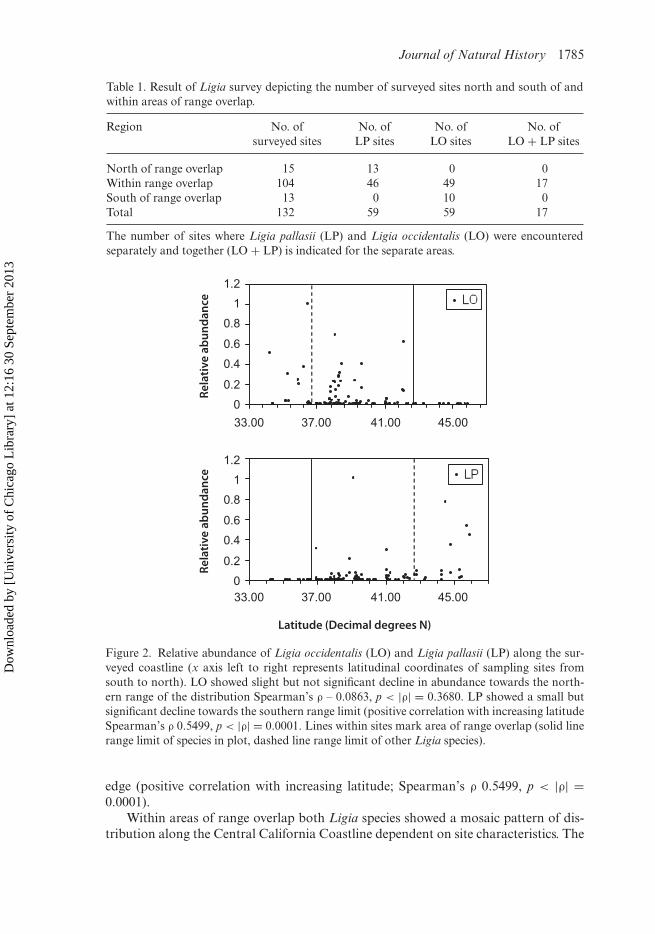
Both L. pallasii and L. occidentalis showed weak patterns of decline in abundancefrom the centre of the range towards the one respective range margin included in thissurvey (Figure 2). Ligia occidentalis showed a slight, but not significant, decline inabundance towards the northern range edge (Spearman’s ρ – 0.0863, p < |ρ| = 0.3680).Ligia pallasii populations decreased slightly in abundance towards the southern range
Dow
nloa
ded
by [
Uni
vers
ity o
f C
hica
go L
ibra
ry]
at 1
2:16
30
Sept
embe
r 20
13
Journal of Natural History 1785
Table 1. Result of Ligia survey depicting the number of surveyed sites north and south of andwithin areas of range overlap.
Region No. ofsurveyed sites
No. ofLP sites
No. ofLO sites
No. ofLO + LP sites
North of range overlap 15 13 0 0Within range overlap 104 46 49 17South of range overlap 13 0 10 0Total 132 59 59 17
The number of sites where Ligia pallasii (LP) and Ligia occidentalis (LO) were encounteredseparately and together (LO + LP) is indicated for the separate areas.
Rela
tive
abun
danc
eRe
lativ
e ab
unda
nce
Latitude (Decimal degrees N)
33.00
0
0.2
0.4
0.6
0.8
1
1.2
0
0.2
0.4
0.6
0.8
1
1.2
37.00 41.00 45.00
33.00 37.00 41.00 45.00
Figure 2. Relative abundance of Ligia occidentalis (LO) and Ligia pallasii (LP) along the sur-veyed coastline (x axis left to right represents latitudinal coordinates of sampling sites fromsouth to north). LO showed slight but not significant decline in abundance towards the north-ern range of the distribution Spearman’s ρ – 0.0863, p < |ρ| = 0.3680. LP showed a small butsignificant decline towards the southern range limit (positive correlation with increasing latitudeSpearman’s ρ 0.5499, p < |ρ| = 0.0001. Lines within sites mark area of range overlap (solid linerange limit of species in plot, dashed line range limit of other Ligia species).
edge (positive correlation with increasing latitude; Spearman’s ρ 0.5499, p < |ρ| =0.0001).
Within areas of range overlap both Ligia species showed a mosaic pattern of dis-tribution along the Central California Coastline dependent on site characteristics. The
Dow
nloa
ded
by [
Uni
vers
ity o
f C
hica
go L
ibra
ry]
at 1
2:16
30
Sept
embe
r 20
13
1786 R. Eberl
Table 2. Relationship between different environmental variables and the occurrence of Ligiapallasii and Ligia occidentalis within their respective ranges.
A significant Likelihood χ 2 value indicates that the presence or high abundance of a variable issignificantly correlated with a high abundance of the respective Ligia species.
presence of cliffs, caves or large cracks and freshwater seeps were important to siteswhere L. pallasii were found (χ2
p < 0.0001; χ2FW, 113,1 = 30.352, p < 0.0001), but not L. occidentalis (χ2
cliff, 113,1 =1.099, p = 0.2946; χ2
caves, 109,1 = 0.912, p = 0.3396; χ2FW, 111,1 = 0.359, p = 0.5489).
There was a positive relationship between the amount of wrack at a site for L. occiden-talis (χ2
wrack, 94,2 = 11.696, p < 0.005), but not for L. pallasii (χ2wrack, 96,2 = 0.0443, p =
0.801), the opposite was true for the presence of the algae E. intestinalis, Pelvetiopsisspp. and Ulva spp., which showed a significantly positive relationship with L. pallasiibut not with L. occidentalis (Table 2).
Temporal variation of abundanceMonthly monitoring revealed that site occupancy showed consistency with respectto species occurrence but predictably the number of individuals varied dependingon the season (Figure 3: L. pallasii, Figure 4: L. occidentalis). Throughout the 41-month monitoring period only L. occidentalis and never L. pallasii were found at BH.At MP L. pallasii was the only Ligia species found, except for a single adult maleL. occidentalis discovered on 2 July 2007.
Abundances and percentage of different age classes varied with time for bothspecies. Ligia occidentalis showed a single reproductive period each spring as evidencedby the presence of gravid females and young < 5 mm in length (Figure 3), whereasgravid individuals of L. pallasii as well as young were also present during the winterand no clear periodic pattern was observed (Figure 4).
Temperatures changed predictably with the seasons and were lower at MP thanat BH (Figure 5A, B) both for the mean average temperature (MP: 11.4 ± 0.2◦C,BH: 13.14 ± 0.2◦C) as well as for the mean daily minimum (MP: 10.95 ± 0.2◦C, BH:9.18 ± 0.3◦C) and maximum (MP: 11.88 ± 0.2◦C, BH: 20.31 ± 0.3◦C) temperatures.
Dow
nloa
ded
by [
Uni
vers
ity o
f C
hica
go L
ibra
ry]
at 1
2:16
30
Sept
embe
r 20
13
Journal of Natural History 1787
Num
ber
of L
igia
per
squ
are
met
er
Figure 3. Monthly survey of Ligia occidentalis at Bodega Harbor from March 2007 toDecember 2010. Overall abundance is represented as mean number of individuals per squaremetre and is composed of numbers from four different size classes (see key).
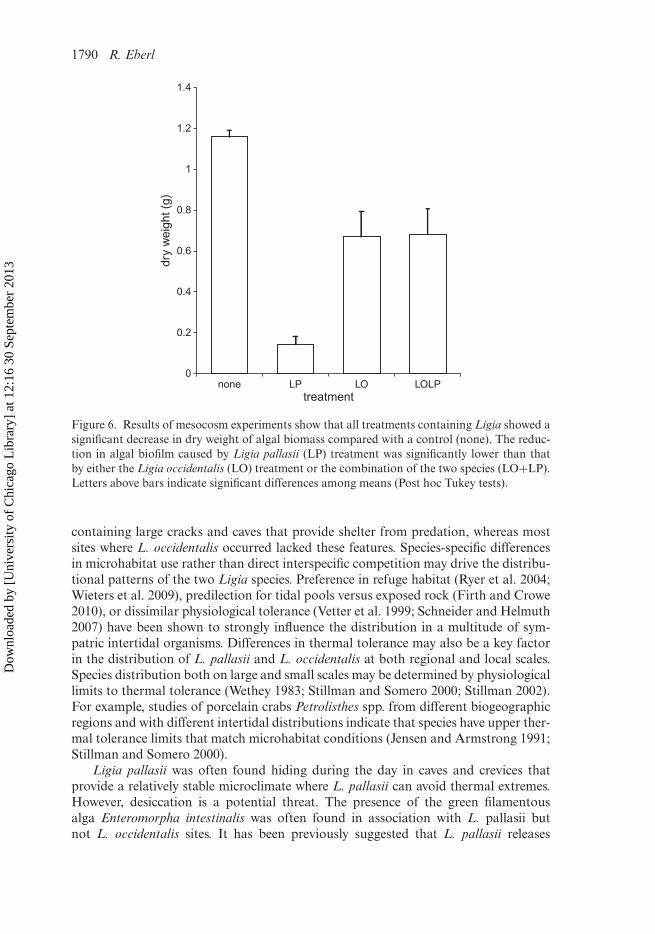
Mesocosm experimentsBoth Ligia species fed on algal biofilm mostly composed of several species of diatomsand small filamentous green algae. The dry weight of algal biofilm differed significantlyacross the four treatments – Welch ANOVA F(3, 22.3) = 141.55, p < 0.001. Tukeypost hoc comparisons of the four groups indicate that the dry weight of the controlgroup (M = 1.16 ± 0.03 g) was significantly greater than those for any other treat-ment. Algal weight in the L. pallasii treatment (0.145 ± 0.003 g) was significantly lowerthan in the L. occidentalis (0.67 ± 0.12 g) or L. occidentalis plus L. pallasii treatment(0.681 ± 0.13 g). Differences between L. occidentalis and L. occidentalis plus L. pallasiiwere not statistically significant (p > 0.05, Figure 6).
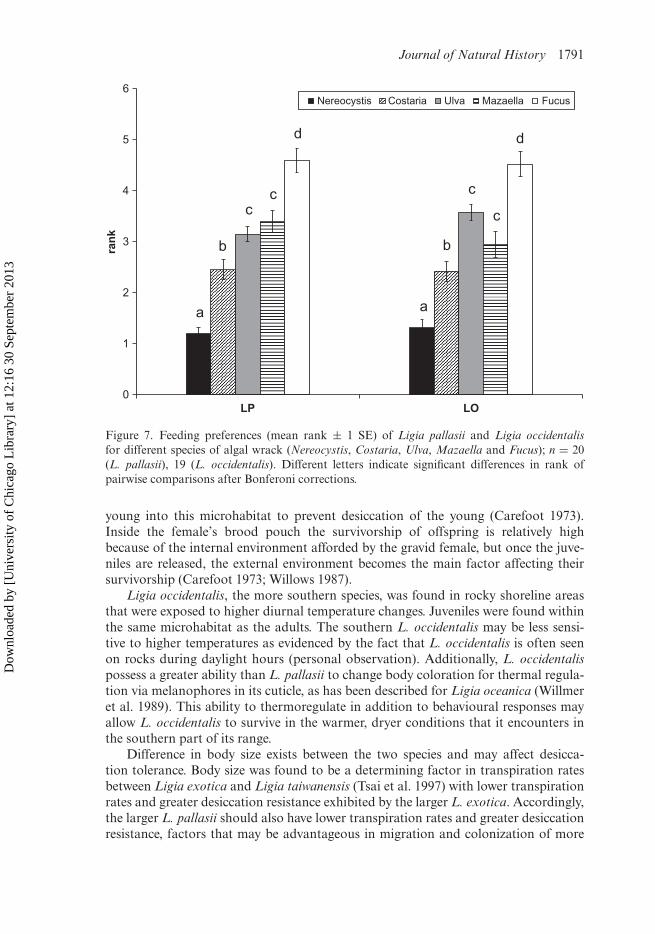
Food choice experiments I and IILigia occidentalis and L. pallasii had similar feeding preferences for different algalwrack species. Nereocystis luetkeana ranked highest in preference, C. costata, Ulva spp.and M. flaccida were in the middle, and F. gardneri ranked lowest (Figure 7).
Isopods were rarely observed feeding (L. pallasii: 20/275, L. occidentalis: 31/275)in this experiment. Ligia pallasii were observed eight times to feed on fish and green
Dow
nloa
ded
by [
Uni
vers
ity o
f C
hica
go L
ibra
ry]
at 1
2:16
30
Sept
embe
r 20
13
1788 R. Eberl
Figure 4. Monthly survey of Ligia pallasii at Mussel Point from March 2007 to December2010. Overall abundance is represented as mean number of individuals per square metre andis composed of numbers from five different size classes (see key).
algae and never on the agar-only square. Ligia occidentalis were observed to feed eighttimes on the green algae, brown algae and fish and sampled the non-food square twice.Both. L. pallasii and L. occidentalis showed a preference for fish, which was consumedmore than any other food (Figure 8). Whereas L. pallasii fed equally on diatoms andthe two algal species, L. occidentalis preferred diatoms to the two algal species offered(Figure 8).
Discussion
Ligia pallasii and L. occidentalis displayed a mosaic pattern of distribution in areasof sympatry between southern Oregon and Monterey Bay where both species onlyrarely occurred at the same site. Temperature generally decreases toward northernlatitudes along the eastern Pacific coast and the more northern L. pallasii tends toencounter cooler temperatures along its range than the more southern L. occidentalis.However, complex interactions of local factors (e.g. differences in time of low tide orthe topographic orientation of a site) with regional patterns can cause differences inenvironmental conditions at varying scales (Underwood and Chapman 1996; Helmuthet al. 2002, 2006). Differences in thermal patterns between L. pallasii and L. occiden-talis sites at Bodega Marine Reserve in this study suggest that microclimatic differenceson local scales may partially explain the observed distributional pattern in areas ofsympatry.
Dow
nloa
ded
by [
Uni
vers
ity o
f C
hica
go L
ibra
ry]
at 1
2:16
30
Sept
embe
r 20
13
Journal of Natural History 1789
0.00
5.00
10.00
15.00
20.00
25.00
Apr-08
Jun-08
Jul-08
Sep-08
Nov-08
Dec-08
Feb-09
Apr-09
May-09
Jul-09
Sep-09
Oct-09
Dec-09
Feb-10
Mar-10
May-10
Jul-10
Aug-10
Oct-10
Nov-10
average max min
0.00
5.00
10.00
15.00
20.00
25.00
Apr-08
Jun-08
Jul-08
Sep-08
Nov-08
Dec-08
Feb-09
Apr-09
May-09
Jul-09
Sep-09
Oct-09
Dec-09
Feb-10
Mar-10
May-10
Jul-10
Aug-10
Oct-10
Nov-10
A
B
Figure 5. Temperature records at Mussel Point cave (A) and Bodega Harbor (B) represented asmean temperature per month (average) and the monthly average of the daily maximum (max)and minimum (min) temperatures.
Differences in habitat preferences, shown for other intertidal organisms (Jensenand Armstrong 1991; Franke and Janke 1998), rather then a strict vertical niche seg-regation within the intertidal zone as has been previously suggested for L. pallasii andL. occidentalis (Morris et al. 1980; Ricketts and Calvin 1985) appear to drive the dis-tribution of the two Ligia species within areas of range overlap. Whereas vertical nichesegregation is a pattern frequently encountered in the rocky intertidal zone in a multi-tude of closely related organisms (Connell 1961; Collins et al. 1996; Menge et al. 2007;Donahue et al. 2009), L. pallasii and L. occidentalis did not show a clear pattern ofvertical niche segregation as evidenced by a lack of sites where both species occurtogether. Ligia pallasii and L. occidentalis showed microhabitat preferences withinthe intertidal. Ligia pallasii was frequently found in areas with cliffs and rock walls
Dow
nloa
ded
by [
Uni
vers
ity o
f C
hica
go L
ibra
ry]
at 1
2:16
30
Sept
embe
r 20
13
1790 R. Eberl
0
0.2
0.4
0.6
0.8
1
1.2
1.4
none LP LO LOLPtreatment
dry
wei
ght (
g)
Figure 6. Results of mesocosm experiments show that all treatments containing Ligia showed asignificant decrease in dry weight of algal biomass compared with a control (none). The reduc-tion in algal biofilm caused by Ligia pallasii (LP) treatment was significantly lower than thatby either the Ligia occidentalis (LO) treatment or the combination of the two species (LO+LP).Letters above bars indicate significant differences among means (Post hoc Tukey tests).
containing large cracks and caves that provide shelter from predation, whereas mostsites where L. occidentalis occurred lacked these features. Species-specific differencesin microhabitat use rather than direct interspecific competition may drive the distribu-tional patterns of the two Ligia species. Preference in refuge habitat (Ryer et al. 2004;Wieters et al. 2009), predilection for tidal pools versus exposed rock (Firth and Crowe2010), or dissimilar physiological tolerance (Vetter et al. 1999; Schneider and Helmuth2007) have been shown to strongly influence the distribution in a multitude of sym-patric intertidal organisms. Differences in thermal tolerance may also be a key factorin the distribution of L. pallasii and L. occidentalis at both regional and local scales.Species distribution both on large and small scales may be determined by physiologicallimits to thermal tolerance (Wethey 1983; Stillman and Somero 2000; Stillman 2002).For example, studies of porcelain crabs Petrolisthes spp. from different biogeographicregions and with different intertidal distributions indicate that species have upper ther-mal tolerance limits that match microhabitat conditions (Jensen and Armstrong 1991;Stillman and Somero 2000).
Ligia pallasii was often found hiding during the day in caves and crevices thatprovide a relatively stable microclimate where L. pallasii can avoid thermal extremes.However, desiccation is a potential threat. The presence of the green filamentousalga Enteromorpha intestinalis was often found in association with L. pallasii butnot L. occidentalis sites. It has been previously suggested that L. pallasii releases
Dow
nloa
ded
by [
Uni
vers
ity o
f C
hica
go L
ibra
ry]
at 1
2:16
30
Sept
embe
r 20
13
Journal of Natural History 1791
0
1
2
3
4
5
6
LP LO
ran
kNereocystis Costaria Ulva Mazaella Fucus
a
b
cc
d
a
b
c
c
d
Figure 7. Feeding preferences (mean rank ± 1 SE) of Ligia pallasii and Ligia occidentalisfor different species of algal wrack (Nereocystis, Costaria, Ulva, Mazaella and Fucus); n = 20(L. pallasii), 19 (L. occidentalis). Different letters indicate significant differences in rank ofpairwise comparisons after Bonferoni corrections.
young into this microhabitat to prevent desiccation of the young (Carefoot 1973).Inside the female’s brood pouch the survivorship of offspring is relatively highbecause of the internal environment afforded by the gravid female, but once the juve-niles are released, the external environment becomes the main factor affecting theirsurvivorship (Carefoot 1973; Willows 1987).
Ligia occidentalis, the more southern species, was found in rocky shoreline areasthat were exposed to higher diurnal temperature changes. Juveniles were found withinthe same microhabitat as the adults. The southern L. occidentalis may be less sensi-tive to higher temperatures as evidenced by the fact that L. occidentalis is often seenon rocks during daylight hours (personal observation). Additionally, L. occidentalispossess a greater ability than L. pallasii to change body coloration for thermal regula-tion via melanophores in its cuticle, as has been described for Ligia oceanica (Willmeret al. 1989). This ability to thermoregulate in addition to behavioural responses mayallow L. occidentalis to survive in the warmer, dryer conditions that it encounters inthe southern part of its range.
Difference in body size exists between the two species and may affect desicca-tion tolerance. Body size was found to be a determining factor in transpiration ratesbetween Ligia exotica and Ligia taiwanensis (Tsai et al. 1997) with lower transpirationrates and greater desiccation resistance exhibited by the larger L. exotica. Accordingly,the larger L. pallasii should also have lower transpiration rates and greater desiccationresistance, factors that may be advantageous in migration and colonization of more
Dow
nloa
ded
by [
Uni
vers
ity o
f C
hica
go L
ibra
ry]
at 1
2:16
30
Sept
embe
r 20
13
1792 R. Eberl
agardiatomsUlvaPelvetiopsisfish
Figure 8. Differences in food preference for the two Ligia species [Ligia occidentalis (LO):F (4,53.45) = 15.918, p = 0.0001; Ligia pallasii (LP): F (4,53,45) = 15.928, p = 0.0004] as shown formean dry weight consumed for each food type (error bars are ± 1 SE). Changes in weight of thedifferent food types without isopods (none: F (4,22.15) = 0.699, p = 0.601) show minimal changesin weight possibly because of microbial activity. Values of p are based on analysis of variance.Letters above bars indicate significant differences among means (Post hoc Tukey tests).
terrestrial habitats. While a habitat preference for sites with cliffs affords L. pallasiigreater upward mobility, the geographic range of L. pallasii is restricted to areaswith a cool humid marine climate with frequent occurrence of coastal summer fog(Johnstone and Dawson 2010). The interplay of species-specific differences in physi-ological tolerance and vertical migration patterns should be experimentally tested infuture studies.
The northern L. pallasii showed a decline in abundance towards the southern endof its range. Although older records show the occurrence of L. pallasii in Monterey,California (including at Hopkins Marine Station), no specimens were found south ofCapitola in this study. Change in thermal conditions as well as disturbance by humans(e.g. high frequency of visitors in the intertidal, sample collection by researchers) mayhave either drastically reduced species abundance at these southern sites or caused anorthern range shift altogether. Substantial changes in intertidal community composi-tion in Monterey over a time frame of 60 years when the same transects were sampledfrom 1931–1933 and 1993–1994 were recorded as well as an average increase of 2.2◦Cin sea surface temperature during the summer (Barry et al. 1995; Sagarin et al. 1999).Unfortunately, this data set does not include records of L. pallasii.
Monthly monitoring of sites in the centre of range overlap showed that speciesabundance varied during the year for both species. Differences in breeding phenologywere found between the two Ligia species. While L. occidentalis showed a distinctive
Dow
nloa
ded
by [
Uni
vers
ity o
f C
hica
go L
ibra
ry]
at 1
2:16
30
Sept
embe
r 20
13
Journal of Natural History 1793
breeding cycle with one reproductive event during the late spring and early summer,L. pallasii appears capable of almost year-round reproduction. The pattern observedin L. pallasii could be the result of cohort splitting. Different growth rates in juvenilescould lead to differential timing of maturity and subsequently early and late reproduc-ing females; this was not specifically investigated in this study. This differential repro-ductive pattern was described in detail in L. oceanica (Willows 1987) and could con-tribute to the stability of populations in some isopod species (Al-Dabbagh and Block1981; Willows 1987; Zimmer 2004). Plasticity in life history traits between populationsof L. exotica are apparently correlated with habitat differences (Tsai and Dai 2001).Populations of L. exotica in Taiwan from sites further away from the coast in a creekbed that are exposed to greater aridity showed delayed maturity of females and theproduction of fewer but larger offspring. The observed differences between supralit-toral and freshwater L. exotica populations could however be attributed to the exis-tence of cryptic species that frequently occur in Ligia spp. (Jung et al. 2008; Hurtadoet al. 2010; Markow and Pfeiler 2010) rather than plasticity in life history traits.
Food availability and food quality can have a significant effect on survival and fit-ness of isopods (Brody and Lawlor 1984; Hassall and Dangerfield 1990; Hassall et al.2003). Both L. pallasii and L. occidentalis showed a preference for high-quality protein-rich food in the food choice experiments. Both species were found to eat similar food,mostly diatoms, algal wrack and the occasional rare protein source. While Ligia spp.have long been recognized as important decomposers of algal wrack (Carefoot 1989;Pennings et al. 2000; Zimmer et al. 2000), a recent study demonstrated the impor-tance of L. exotica as a grazer on algal biofilm in a tropical system (Christofolettiet al. 2011). The results from the mesocosm experiment in this study show a similargrazing effect in Ligia spp. from a temperate climate. Both L. pallasii and L. occiden-talis significantly reduced the biomass of biofilm under experimental conditions. Thelarger L. pallasii, as expected, ate more biofilm than L. occidentalis and may there-fore have a greater impact on epilithic biofilm in the rocky intertidal. The presenceof a second Ligia species within mesocosms did not significantly reduce feeding. Thisresult suggests that direct competition for food may not be important in the trophicecology of these two isopods. Furthermore, L. pallasii and L. occidentalis are unlikelyto compete for food in the wild because they were rarely found at the same sites. Formost of the year food seems to be available in sufficient quantities and not limitingto L. pallasii abundance (Carefoot 1998). In fact, it was shown that L. pallasii spentonly a small portion of time feeding (Carefoot 1998). There is no reason to suggestthat conditions of food availability may be significantly different for L. occidentalis. Inaddition to epilithic biofilm, both Ligia species preferred similar species of algal wrack,preferred algal types for both L. pallasii and L. occidentalis in central California weresimilar to those of L. pallasii in British Columbia and preference may be mediated byalgal traits (Pennings et al. 2000). Preference for certain algal types appears to remainconstant over large geographic areas, at least in L. pallasi. In addition to the type ofalgal wrack, colonization of washed up algae by microbes may be important in affect-ing food quality (Zimmer et al. 2002; Ihnen and Zimmer 2008). In a feeding trial withmicrobe-enriched artificial food the amount of microbial colonization increased theattractiveness of a given food source to the terrestrial isopod Porcelio scaber, whichalso showed clear preferences for particular types of microbes on the food (Ihnen andZimmer 2008). Differences of microbial colonization of algae and wrack could influ-ence food choice in Ligia spp. as well and should be investigated in future studies. BothL. pallasii and L. occidentalis probably have varied diets composed of epilithic biofilm,
Dow
nloa
ded
by [
Uni
vers
ity o
f C
hica
go L
ibra
ry]
at 1
2:16
30
Sept
embe
r 20
13
1794 R. Eberl
fresh seaweeds and algal wrack as well as carrion, depending on availability, but donot directly compete for available food.
Ligia pallasii and L. occidentalis have an area of range overlap from southernOregon to Monterey Bay, California. The distribution of L. pallasii and L. occiden-talis shows a pattern of mosaic distribution in areas or range overlap with a paucity ofsites where both species occur and interact and compete with each other. The resultsfrom this study show no distinct pattern of vertical zonation between L. pallasii andL. occidentalis but differences in habitat preference, most notably the occurrence ofL. pallasii at sites with rocky cliffs. Variation in temperature between microhabitatscould explain the differences in species distribution both at regional and local scales.No significant differences in food preferences were found between these species and itis therefore unlikely that competition for food determines the distributional pattern ofthe two isopod species.
Acknowledgements
T. Grosholz and D.R. Strong provided helpful suggestions for improvement of this manuscript.Thanks to A. Newsom for help with isopod collection and maintenance of mesocosms. TheBodega Marine Laboratory provided wet lab. space and outdoor tank facilities. Thanks toJ. Sones for help with site selection at the Bodega Marine Reserve and for supplying Ligia pho-tos used in Figure 1. Funding for this research was provided through UC Davis internal grants,Explorer’s Club Diversa Research Fund and through NSF DGE grant 0841297 to S.L. Williams.This is a contribution of the Bodega Marine Laboratory, University of California Davis.
References
Abbott DP. 1939. Shore Isopods; niches occupied and degrees of transition toward land life withspecial reference to the family Ligydae.7th Pacific Science Congress 3:505–511.
Al-Dabagh KY, Block W. 1981. Population ecology of a terrestrial isopod in 2 Breckland East-Anglican grass heaths. J Anim Ecol. 50:61–78.
Andrewartha HG, Birch LC. 1954. The distribution and abundance of animals. Chicago:University of Chicago Press.
Barry JP, Baxter CH, Sagarin RD, Gilman SE. 1995. Climate-related, long-term faunal changesin a California rocky intertidal community. Science 267:672–675.
Brody MS, Lawlor LR. 1984. Adaptive variation in offspring size in the terrestrial isopod,Armadillidium vulgare. Oecologia 61:55–59.
Brusca RC, Coelho V, Taiti S. 2001. A Guide to the Coastal Isopods of California. Availablefrom: http://tolweb.org/notes/?note_id=3004, Accessed 20 May 2011
Carefoot TH. 1973. Studies of the Growth, Reproduction and Life Cycle of the SupralittoralIsopod Ligia pallasii. Marine Biol. 18:302–311.
Carefoot TH. 1989. Diet and its effect on oxygen consumption in the semiterrestrial isopodLigia. Monitore zool. ital. (N.S) Monogr. 4:193–210.
Carefoot TH. 1998. A day in the life of an isopod: time and energy allocations in thesemiterrestrial Ligia pallasii. Israel J Zool. 44:463–471.
Christofoletti RA, Almeida TVV, Ciotti AM. 2011. Environmental and grazing influence onspatial variability of intertidal biofilm on suptropical rocky shores. Marine Ecol-Prog Ser.424:15–23.
Colombini I, Chelazzi L. 2003. Influence of marine allochthonous input on sandy beachcommunities. p.115–159. Oceanography and Marine Biology, Vol 41. London: Taylor& Francis Ltd.
Connell JH. 1961. Influence of interspecific competition and other factors on distribution ofbarnalce Chthamalus stellatus.Ecology 42:710–723.
Donahue MJ, Nichols A, Santamaria CA, League-Pike PE, Krediet CJ, Perez, KO, ShulmanMJ.. 2009. Predation risk, prey abundance, and the vertical distribution of three brachyurancrabs on Gulf of Maine shores. J Crustacean Biol. 29:523–531.
Elton C. 1946. Competition and the structure of ecological communities. J Anim Ecol. 15:54–68.Firth LB, Crowe TP. 2010. Competition and habitat suitability: small-scale segregation
underpins large-scale coexistence of key species on temperate rocky shores. Oecologia162:163–174.
Franke HD, Janke M. 1998. Mechanisms and consequences of intra- and interspecific interfer-ence competition in Idotea baltica (Pallas) and Idotea emarginata (Fabricius) (Crustacea:Isopoda): a laboratory study of possible proximate causes of habitat segregation. J ExpMarine Biol Ecol 227:1–21.
Hassall M, Dangerfield JM. 1990. Density-dependent processes in the population dynamics ofArmadillidium vulgare (Isopoda, Oniscidea). J Anim Ecol. 59:941–958.
Hassall M, Helden A, Benton T. 2003. Phenotypic plasticity and interpopulation differences inlife history traits of Armadillidium vulgare (Isopoda: Oniscidea). Oecologia 137:85–89.
Helmuth B, Halpin PM, O’Donnell M, et al. 2002. Climate change and latitudinal patterns ofintertidal thermal stress. Science 298:1015–1017.
Helmuth B, Broitman BR, Blanchette CA, et al. 2006. Mosaic patterns of thermal stress in therocky intertidal zone: Implications for climate change. Ecol Monogr. 76:461–479.
Hurtado LA, Mateos M, Santamaria CA. 2010. Phylogeography of Supralittoral RockyIntertidal Ligia Isopods in the Pacific Region from Central California to Central Mexico.Plos One 5.
Hutchinson GE. 1959. Homage to Santa-Rosalia or why there are so many kinds of animals.Am Nat. 93:145–159.
Ihnen K, Zimmer M. 2008. Selective consumption and digestion of litter microbes by Porcellioscaber (Isopoda: Oniscidea). Pedobiologia 51:335–342.
Jensen GC, Armstrong DA. 1991. Intertidal zonation among congeners – factors reulating dis-tribution of porcelain crabs Petrolisthes spp (Anomura, Porcellanidae). Marine Ecol-ProgSer. 73:47–60.
Johnson RA.1986. Intraspecific resource partitioning in the bumble bees Bombus ternarius andBombus pennsylvanicus. Ecology 67:133–138.
Johnstone JA, Dawson TE. 2010. Climatic context and ecological implications of summer fogdecline in the coast redwood region. Proc Natl Acad Sci USA. 107:4533–4538.
Jung J, Eo HS, Rho HS, Kim W. 2008. Two genetic lineages of sea slaters, Ligia (Crustacea:Isopoda) in South Korea: A population genetic approach. Molecules Cells 25:523–530.
MacArthur RH. 1972. Coexistence of species. p.253–259. In Benke J, ed. Challenging BiologicalProblems. New York: Oxford University Press.
Markow TA, Pfeiler E. 2010. Mitochondrial DNA evidence for deep genetic divergences inallopatric populations of the rocky intertidal isopod Ligia occidentalis from the easternPacific. Mol Phylogen Evol. 56:468–473.
Menge BA, Daley BA, Sanford E, Dahlhoff EP, Lubchenco J. 2007. Mussel zonation in NewZealand: an integrative eco-physiological approach. Marine Ecol-Prog Ser. 345:129–140.
Morris RH, Abbott DP, Haderlie EC. 1980. Intertidal invertebrates of California. Stanford(CA): Stanford University Press.
Dow
nloa
ded
by [
Uni
vers
ity o
f C
hica
go L
ibra
ry]
at 1
2:16
30
Sept
embe
r 20
13
1796 R. Eberl
Paine RT. 1966. Food web complexity and species diversity. Am Nat. 100:65–75.Paine RT. 1974. Intertidal community structure: experimental studies on the relationship beteen
a dominant competitor and its principal predator. Oecologia 15:93–120.Pennings SC, Carefoot TH, Zimmer M, Danko JP, Ziegler A. 2000. Feeding preferences of
supralittoral isopods and amphipods. Can. J. Zool. 78:1918–1929.Richardson H. 1905. A monograph of isopods of North America. Bull US Natl Mus. 54:727p.Ricketts EF, Calvin J. 1985. Between Pacific tides. 5th edition. Stanford (CA): Stanford
University Press.Ryer CH, Stoner AW, Titgen RH. 2004. Behavioral mechanisms underlying the refuge value of
benthic habitat structure for two flatfishes with differing anti-predator strategies. Mar EcolProgr Ser. 268:231–243.
Sagarin RD, Barry JP, Gilman SE, Baxter CH. 1999. Climate-related change in an intertidalcommunity over short and long time scales. Ecol Monogr. 69:465–490.
Sagarin RD, Gaines SD. 2002. The ‘abundant center’ distribution: to what extent is it abiogeographical rule? Ecol Lett. 5:137–147.
Schmalfuss H. 2003. World catalog of terrestrial isopods (Isopoda: Oniscidea). Stuttg BeitrNaturk Ser A (Biol). 654:341pp.
Schneider KR, Helmuth B. 2007. Spatial variability in habitat temperature may drive patternsof selection between an invasive and native mussel species. Marine Ecol-Prog Ser. 339:157–167.
Schoener TW. 1974. Resource partitioning in ecological communities. Science 185:27–39.Sokal RR, Rohlf RF. 1995. Biometry. 3rd edition. New York: Freeman and Company.Somero GH. 2002. Thermal physiology and vertical zonation of intertidal animals: Optima,
limits, and costs of living. Integr Compar Biol. 42:780–789.Stillman JH. 2002. Causes and consequences of thermal tolerance limits in rocky intertidal
porcelain crabs, genus Petrolisthes. Integr Compar Biol. 42:790–796.Stillman JH, Somero GN. 2000. A comparative analysis of the upper thermal tolerance limits of
eastern Pacific porcelain crabs, genus Petrolisthes: influences of latitude, vertical zonation,acclimation, and phylogeny. Physiol Biochem Zool. 73:200–208.
Tsai ML, Dai CF, Chen CF. 1997. Variations of reproductive traits of two semi-terrestrialisopods, Ligia exotica and L. taiwanensis (Crustacea: Ligiidae), in southern Taiwan. ZoolStud. 36:33–41.
Tsai ML, Dai CF. 2001. Life history plasticity and reproductive strategy enabling the invasion ofLigia exotica (Crustacea: Isopoda) from the littoral zone to an inland creek. Mar Ecol-ProgSer. 210:175–184.
Underwood AJ, Champman MG. 1996. Scales of spatial patterns of distribution of intertidalinvertebrates. Oecologia 107: 212–224.
Vandermeer JH. 1972. Niche Theory. Annu Rev Ecol System. 3:107–132.Vetter RAH, FrankeHD, Buchholz F. 1999. Habitat-related differences in the responses to oxy-
gen deficiencies in Idotea baltica and Idotea emarginata (Isopoda, Crustacea). J Exp MarineBiol Ecol. 239:259–272.
Wethey DS. 1983. Geographic limits and local zonation – the barnacles Semibalanus (Balanus)and Chthamalus in New England. Biol Bull. 165:330–341.
Wieters EA, Salles E, Januario SM, et al. 2009. Refuge utilization and preferences betweencompeting intertidal crab species. J Exp Marine Biol Ecol. 374: 37–44.
Willmer PG, Baylis M,Simpson CL. 1989. The roles of color-change and behavior in thehygrothermal balance of a littoral isopod Ligia oceanica. Oecologia 78:349–356.
Willows RI. 1987 Population and individual energetics of Ligia oceanica (Crustacea, Isopoda)in the rocky supralittoral. J Exp Marine Biol Ecol. 105:253–274.
Zimmer M. 2004. Effects of temperature and precipitation on a flood plain isopod community:a field study. Eur J Soil Biol. 40:139–146.
Dow
nloa
ded
by [
Uni
vers
ity o
f C
hica
go L
ibra
ry]
at 1
2:16
30
Sept
embe
r 20
13
Journal of Natural History 1797
Zimmer M, Brauckmann HJ, Broll G, Topp W. 2000. Correspondence analytical evalua-tion of factors that influence soil macro-arthropod distribution in abandoned grassland.Pedobiologia 44:695–704.
Zimmer M, Danko JP, Penning SC, Danford AR, Carefoot TH, Ziegler A, Uglow RF. 2002Cellulose digestion and phenol oxidation in coastal isopods (Crustacea: Isopoda). MarineBiol. 140:1207–1213.
Zimmer M, Topp W. 2000. Species-specific utilization of food sources by sympatric woodlice(Isopoda: Oniscidea). J Anim Ecol. 69:1071–1082.



































![Enfraquecimento Do Legislativo Ligia[1]](https://static.documents.pub/doc/80x56/55cf8f80550346703b9d0376/enfraquecimento-do-legislativo-ligia1.jpg)