DNA damaging properties of single walled carbon nanotubes in humancolon carcinoma cells
Joanna Pelka1,2, Helge Gehrke1, Anja Rechel2, Manfred Kappes3,4, Frank Hennrich4, Christian G. Hartinger5, &Doris Marko1,2
1Department of Food Chemistry and Toxicology, University of Vienna, Vienna, Austria, 2Karlsruher Institut für Technologie,Institut für Angewandte Biowissenschaften, Karlsruhe, Germany, 3Karlsruher Institut für Technologie, Institut für PhysikalischeChemie II, Karlsruhe, Germany, 4Karlsruher Institut für Technologie, Institut für Nanotechnologie, Karlsruhe, Germany and5Institute of Inorganic Chemistry, University of Vienna, Vienna, Austria
AbstractSingle walled carbon nanotubes were studied with respect tocytotoxic and genotoxic properties in cells of the gastrointestinaltract as exemplified for the human colon carcinoma cell lineHT29. No effect on cell growth in the sulphorhodamine B assaywas observed after 24 h of incubation, whereas growthinhibitory properties were found after 48 and 72 h. After 24 hincubation a decrease of mitochondrial activity (WST-1) wasmeasured (‡0.1 mg/ml), whereas membrane integrity (lactatedehydrogenase) was not affected. In cytotoxic concentrations,the formation of reactive oxygen species and a slight increase oftotal glutathione and nuclear Nrf2 were observed. However,already in subcytotoxic concentrations substantial DNAdamaging effects were found in the alkaline comet assay, whichwere not associated with enhanced formation offormamidopyrimidine-DNA-glycosylase-sensitive sites. Inaddition, an increase of kinetochore-negative micronuclei (V79)and phosphorylation of the tumour suppressor proteinp53 (HT29) underlined the genotoxic potential of thesenanostructures.
Keywords: DNA integrity, comet assay, oxidative stress,micronuclei, glutathione
Introduction
Nanosciences have already invaded various fields of dailylife, such as computer electronics, communication, energyproduction or medicine. Carbon nanotubes (CNTs) haveattracted a great deal of attention due to their uniquestructural, electrical and mechanical properties. For exam-ple, the unique absorption of CNTs in the near infraredallows their development for biological sensing or cancertherapy (Kam et al. 2005). Beside the applications in a broadspectrum of technology and health related products,
nanotechnology can also lead to several innovations inthe packaging of food products, also incorporating CNTsamong other things (Sozer & Kokini 2009; Chaudhry et al.2008). The rapid development of novel fields of applicationsand the increasing production raise questions on the envi-ronmental and health impact of CNTs.
In most studies, CNTs are discussed predominantly as aninhalation hazard (Donaldson et al. 2006). Maynard et al.(2004) reported that handling of CNTs during productionstages may result in aerosol release of these materials inrespirable size into the surroundings. Even though theinvestigations on the potential of single walled carbon nano-tubes (SWCNTs) to become airborne showed quite a lowrelease of this material during handling (<53 mg/m3), theprospective widespread use of this class of nanomaterial mayincrease environmental exposure considerably. Severalin vivo and in vitro studies on inhalative and dermal expo-sure to CNTs have been published concerning induction ofoxidative stress, inflammation, cell damage, adverse effectson cell performance and, in a long-term perspective, effectslike fibrosis and cell wall thickening, etc. (Fiorito et al. 2006;Kagan et al. 2006; Jia et al. 2005; Monteiro-Riviere et al. 2005;Muller et al. 2005; Lam et al. 2006; Helland et al. 2007;Shvedova et al. 2003, 2005; Warheit et al. 2004). Other studyreports demonstrate, however, minor toxic response or evena lack of substantial toxic properties (Belyanskaya et al. 2009;Pulskamp et al. 2007; Thurnherr et al. 2009).
In contrast to nanoparticle uptake by inhalation, little isknown so far about the toxicological relevance of SWCNTstaken up orally. In terms of application of CNTs in foodpackaging, these nanomaterials may migrate into the foodand consequently reach the gastrointestinal tract (GIT).Another way of entry of CNTs into the GIT may arisefrom potential use of this nanomaterial for encapsulationof pharmaceuticals against GIT diseases. As a future sce-nario, it appears also to be imaginable that enhanced usage
Correspondence: Doris Marko, Department of Food Chemistry and Toxicology, University of Vienna, Währingerstrasse 38, A-1090 Vienna, Austria,Tel: +43 1 4277 70800. Fax: +43 1 4277 70899. E-mail: [email protected]
(Received 10 May 2011; accepted 16 September 2011)
Nan
otox
icol
ogy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
ity o
f M
elbo
urne
on
03/0
1/13
For
pers
onal
use
onl
y.
of CNTs in multiple fields of applications might result in theentry of the food chain as a contaminant. So far, CNTs werediscussed as one of the least biodegradable materials everdevised. As they are lipophilic by nature (Wu et al. 2006;Helland et al. 2007), they are totally insoluble in waterin pristine form (Lam et al. 2004; Helland et al. 2007).However, recent studies reported the enzymatic degradationof SWCNTs by the plant-derived enzyme horse radish per-oxidase (Allen et al. 2008) or by neutrophil myeloperoxidase(Kagan et al. 2010). Here, carboxyl groups on the surfaceof the CNTs seem to be of particular importance. Hence,biopersistence of these nanomaterials should be investigatedin-depth in future studies, particularly in view of theircharacteristics and functionalisation.
It is known that biopersistent and lipophilic chemicalsmay accumulate along the food chain, which might indeedbe of relevance for some CNT species. SWCNTs have beenshown to be bioavailable to aquatic organisms (OberdörsterE et al. 2005). Water-solubilised and insolubilised SWCNTswere detected in the faecal material collected from thedigestive tract of exposed fish. Furthermore, there is concernthat CNTs might be taken up by microbial communities androots (Oberdörster et al. 2006) thus accumulating in planttissue (Helland et al. 2007). However, a recent study con-cluded by means of quantitative modelling that CNTs willnot enter the food chain as most of this material is burnt inwaste incineration plants (Gottschalk et al. 2009). Takentogether, CNTs deserve comprehensive studies regardingthe contamination of the environment, food chain and thepossibly resulting oral uptake.
Beside the direct oral uptake of nanotubes by food analternative route of entry into the GIT after inhalation is alsopossible. Inhaled nanotubes can enter the GIT after differentphysical clearance mechanisms. Normally, particles smallerthan 10 mm are absorbed by the mucus of the bronchi andtrachea and subsequently are removed by cilia from therespiratory tract. In contrast, particles smaller than 2.5 mmare able to invade the alveoli. In the alveoli, macrophagesphagocytise the insoluble particles and move toward themucociliary escalator and are thereby cleared from therespiratory tract. Subsequently, the mucus-bounded parti-cles can be swallowed and thus enter the GIT.
The formation of reactive oxygen species (ROS) withincells exposed to particulate matter is considered to be amajor contributor to their toxicological effects (Donaldsonet al. 2005). SWCNTs are routinely synthesised using a metalcatalyst (for example, Co, Ni, Mo, Fe). Since completepurification from residual catalysts, which are located insidethe CNTs or outside of these cylinders, is unlikely, thecatalysts can contribute to their toxicological potential, espe-cially to the modulation of the cellular redox status. SWCNTsare hydrophobic tubular nanostructures with diametersvarying between about 0.7 and 3 nm (Jorio et al. 2001;Donaldson et al. 2006). Accordingly, they are smaller thanthe nuclear pore complex and may enter the cell nucleus.Porter et al. (2007) reported the localisation of SWCNTswithin the cell nuclei of macrophages. However, in humanmesenchymal stem cells and HeLa cells strikingly highintracellular concentrations of SWCNTs were observed
with preferential localisation in the cytosol of the cells(Holt et al. 2011).
With the focus on possible oral uptake of CNTs, thepresent study addresses the question of the biological rele-vance of SWCNTs in cells originating from the GIT (humancolon carcinoma cell line, HT29 cells). The objective of ourstudy was the evaluation of potential cytotoxic properties ofSWCNTs in this cell type, with special emphasis on effectsmodulating the cellular redox status and the maintenance ofDNA integrity.
Methods
SWCNTs and CBSWCNTs were synthesised with a pulsed laser vaporisationsetup similar to that originally developed by Guo et al.(1995): an unfocused Nd:YAG laser (Continuum Powerlite,Q-switched, 1064 nm, 0.5 J/pulse, 30 Hz) irradiated the sidesurface of a rotating and axially translating composite carbonrod (Toyo Tanso, Japan) doped with 1 at.% Ni and Co andpositioned in a Ø 70 mm T-like quartz tube inside a hingeoven (Linn High Therm) (Lebedkin et al. 2002). The appa-ratus can run without interruption until the target is fullyconsumed. Laser vaporisation was performed at an oventemperature of 1150�C in 0.5 bar Ar flowing at ~ 80 sccm.SWCNT material was collected on a filter downstream of theoven, along the cold downstream region of the quartz tubeand also along the tubing to the filter. Typical vaporisationconditions yielded 0.1–0.3 g/h of raw SWCNT soot with anSWCNT content of 50–70 wt.% (the remaining materialconsisted of fullerenes, amorphous carbon and catalystparticles) (Hennrich et al. 2003). The SWCNTs have a diam-eter distribution between 1 and 1.4 nm. There are about50 metallic and semiconducting tubes differing in helicityand corresponding chiral index (n, m) in this size range(Lebedkin et al. 2003). In the raw SWCNT soot, the tubeswere found to form close packed bundles generally 10–100 nm in diameter. The carbon black (CB) preparation(Printex 90, Evonik Industries) used as a reference materialin this study was a kind gift by the Institute of Toxicology andGenetics of the Karlsruher Institut für Technologie. For thisspecific CB batch, the manufacturer specified a nominalprimary particle diameter of 14 nm and a surface area of300 m2/g (particle surface area measurements; aka Brunner-Emmet-Teller). Further characteristics of this specific CBbatch were published in the final report of the Nano Careproject (Kuhlbusch et al. 2009).
SWCNT suspensionTo suppress the formation of aggregates or rather bundles inthe aqueous system of the cell culture medium, severalcommercially available surfactants, such as sodium dodecylsulphate (SDS), sodium cholate (SC), amylose, polyvinylpyr-olidone (PVP), lecithin, gum arabica and b-cyclodextrinwere tested for their efficiency to disperse the nanotubesoot produced into a suspension of individual nanotubes.From studies in which a wide variety of surfactants havebeen investigated with respect to their efficiency in dissol-ving/dispersing isolated SWCNTs, it is known that in
SWCNT toxicity in human colon cells
Nan
otox
icol
ogy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
ity o
f M
elbo
urne
on
03/0
1/13
For
pers
onal
use
onl
y.
particular bile salts are found to be exceptionally effective atindividualising SWCNTs. This has been evidenced by atomicforce microscopy (AFM), optical absorption, fluorescenceand Raman spectroscopy which allow estimating the degreeof solubilisation (Wenseleers et al. 2004; Moshammer et al.2009). For the present study, Raman/fluorescence spectros-copy was used to confirm the presence of individualisedSWCNTs in suspension and to compare the dispersionabilities of the different surfactants. The spectra takenwith an excitation wavelength of 785 nm revealed distinctRaman peaks at Stokes-Raman shifts of 1500–1600 cm-1,corresponding to characteristic G modes of SWCNT. Inaddition, photoluminescence (PL) from smaller diametertubes can also be detected in the spectra corresponding to“Raman shifts” of 2000–4000 cm-1. By comparing G modeintensities to PL intensities we get information on the quan-tum of individualised SWCNTs present indicating the dis-persion properties of the different surfactants. SDS and SCwere found to be the most effective surfactants for SWCNTs.Parallel experiments examining the cytotoxic properties ofSC and SDS on HT29 revealed SC, in contrast to SDS, tobe cell compatible up to a concentration of 0.2 mg/ml.Thereupon, a SWCNT–SC stock suspension was prepared.The SWCNT raw material was purified similar to the pro-cedure as described in Moshammer et al. (2009). An amountof 10 mg of SWCNTs raw material was suspended in 12 ml ofD2O with 1 wt.% SC (Sigma) using a tip sonicator (Bandelin,200 W maximum power, 20 kHz, in pulsed modus with100 ms pulses) applied for 2 h at 20% power. The dispersionwas then filtered through a Sephacryl S-200 gel filtrationmedium (Amersham Biosciences) in a glass columnof ~ 2 cm length and 2 cm inner diameter to remove largeragglomerates prior to density gradient centrifugation.
Purification of the suspension by density gradientcentrifugationIn a typical experiment, centrifuge tubes were first filled with5 ml of 60 wt.% iodixanol (purchased as OptiPrep, SigmaAldrich) plus 1 wt.% SC. Then the tube was filled with 3 ml ofthe starting SWCNTs suspension, sealed and centrifuged.Density gradient centrifugation runs were performed for18–20 h at 15�C and 45,000 rpm corresponding to centripetalaccelerations of ~ 103,000 and ~ 140,000 g at the middle andbottom of the centrifuge tube, respectively. This resulted in aself-generated density gradient of iodixanol. After centrifu-gation, different coloured regions – approximately 5 ml of thewhole 8 ml from bottom to top – were visible. Of these, onlythe last (topmost) ~ 0.5 ml contained pure SWCNTs. This lastfraction was then isolated and characterised by AFM withrespect to SWCNT size distribution.
Determination of SWCNT size distribution by AFMAFM samples were prepared and measured as described in(Hennrich et al. 2007). Samples were prepared by spincoating of SWCNT starting suspensions onto silicon wafersand rinsing with water and acetone. Intermittent contactmode AFM images were recorded with a Digital Instru-ments Multimode SPM with NSC15 silicon cantilevers(MikroMasch).
The primary preparation contained 10 mg SWCNTs perml SC solution. A first serial dilution was performed in 1 wt.% SC solution, achieving a concentration range of SWCNTsfrom 0.0005 to 10 mg/ml (with 1 wt.% SC still uniformly ineach diluted sample). Subsequently, each concentrationwas diluted 1:50 with cell culture medium (containing 10%foetal calf serum (FCS)). Thus, a final SC concentration of0.02% was achieved in all final dilutions, covering a SWCNTconcentration range from 0.00001 to 0.2 mg/ml, which wasthen applied in the biological experiments. Previous toeach biological experiment the dilutions were freshlyprepared, mixed thoroughly and immediately used forcell incubation. Furthermore, SWCNT concentrations forall assays were calculated as mg/cm2 corresponding to thearea of incubation dishes in the comet assay (area:21.4 cm2) to assure comparable incubation conditions inthe different biological assays dealing with suspensionsinstead of solutions.
Determination of the cobalt and nickel contents of theSWCNTs by ICP-MSThirty-four percent nitric acid (1 ml; HNO3 puriss p.a.,Sigma Aldrich, further purified with a quartz sub-boilingsystem from Milestone-MLS GmbH, Leutkirch, Germany;H2O 18.2 MW, Synergy 185 UV Ultrapure, Millipore, France)and H2O2 (50 ml) were added to the samples (50 ml ofsuspensions containing 10 mg/ml CNTs) and kept at roomtemperature for 24 h. Afterwards, they were digested in amicrowave system for 3� half an hour to complete disso-lution (up to 180�C; Ethos 1600, MLS Milestone, Germany).The samples were diluted to 5 ml and indium (1 ng/ml, CPIInternational, Santa Rosa, CA) was added as an internalstandard. The samples were analysed with an Agilent7500ce inductively coupled plasma mass spectrometer(ICP-MS) in no-gas mode. The MS was equipped with a100 ml/min microflow nebuliser and nickel cones. The RFpower was adjusted to 1500 W and the flow rate of thecarrier gas was set to 0.90 l/min and the makeup gas flowwas 0.22 l/min. The spray chamber temperature was keptconstant at 2�C. 59Co, 60Ni and 115In were monitored withan integration time of 0.1 sec. Samples were delivered viaan autosampler (ASX-520, CETAC) located in a laminar-flow box (FBS 37, Spetec).
Standards were prepared by diluting 10 mg/ml (± 30 mg/ml) single-element cobalt and nickel stocksolutions (CPI international, USA). The calibration rangewas 0.01–3 mg/l. The detection and quantification limits ofthe ICP-MS were determined as 2.8 and 5.9 ng/l for Co andas 32 and 107 ng/l for Ni, respectively.
Cell cultureHT29 cells and the male Chinese hamster V79 lung fibro-blasts were cultured in humidified incubators (37�C, 5%CO2) in Dulbecco’s modified Eagle’s medium (DMEM).The HT29 culture medium was supplemented with 10%(v/v) heat inactivated FCS and 1% (v/v) penicillin–strepto-mycin (PS), the V79 culture medium was supplemented with10% (v/v) heat inactivated FCS, 2% (v/v) PS and 2% (v/v)L-glutamine.
J. Pelka et al.
Nan
otox
icol
ogy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
ity o
f M
elbo
urne
on
03/0
1/13
For
pers
onal
use
onl
y.
The cell line HT29 was obtained from the GermanCollection of Microorganisms and Cell Cultures (DSMZ,Germany). The male Chinese hamster V79 lung fibroblastswere kindly provided by L. Lehmann (Institut für Pharmazieund Lebensmittelchemie, Würzburg, Germany). DMEM andthe supplements were purchased from Invitrogen� LifeTechnologies (Karlsruhe, Germany) and Sigma Aldrich.
Cells were tested routinely and found to be negative formycoplasm contamination.
Toxicological studiesControl of cell viability by trypan blue exclusionIn the comet assay, 3� 105 HT29 cells were seeded into Petridishes (Ø 5.5 cm); in the glutathione (GSH) assay, 1.2 � 106
HT29 cells were seeded into Petri dishes (Ø 10 cm) andallowed to grow for 48 h. Cells were incubated for 24 h withthe SWCNT–SC suspension and thereafter collected andmixed with a trypan blue solution. Cell viability was deter-mined as percentage of living cells.
SRB assayThe assay was performed according to a modified methodof Skehan et al. (1990). Briefly, cells were seeded into 24-wellplates (3 � 104, 2 � 104 for and 0.8 � 104 cells for 24, 48 and72 h of incubation, respectively) and allowed to grow for 48 hprior to treatment. Cells were incubated with SC as solventcontrol and the SWCNT–SC suspension for 24, 48 and 72 h inserum-containing medium. Incubation was terminated byaddition of trichloroacetic acid (50% v/v solution). After 1 hat 4�C, plates were washed four times with water and stainedwith a 0.4% (w/v) solution of sulphorhodamine B (SRB).The dye was eluted with Tris-buffer (10 mM, pH 10.5) andquantified photometrically (570 nm). Cytotoxicity was deter-mined as percent survival, given by the number of treatedover control cells � 100% (T/C).
Cell proliferation and cytotoxicity (WST-1) assayA total of 1 � 104 HT29 cells in 100 ml serum containingmedium (10% FCS) per well were seeded into 96-wellplates and cultured for 48 h. Cells were treated for 24 hwith the SWCNT–SC suspension, the solvent control SC andquercetin (500 mM) as a positive control. The assay wasperformed according to the manufacturers’ protocol(Colorimetric assay WST-1 (2-[4-iodophenyl]-3-[4-nitro-phenyl]-5-(2,4-disulphophenyl)-2H-tetrazolium) basedfor the nonradioactive quantification of cell proliferation,cell viability and cytotoxicity, Roche). After incubation, thesupernatant medium was replaced by WST-1 diluted1:10 (v/v) with culture medium in order to avoid interfer-ence of the nanotubes with the WST-1 reagent and incu-bated for at least 30 min in a humidified atmosphere (37�C,5% CO2).
The WST-1 reagent is cleaved to formazan by mitochon-drial dehydrogenases. Thus, the amount of formed formazan(red) in the assay correlates to the number of metabolicallyactive cells. After incubation time and shaking for 1 min, thedye was measured photometrically at 480 nm and a refer-ence wavelength of 633 nm. The results are given relative tothe untreated control in T/C.
LDH assayIn all, 4� 104 HT29 cells in 100 ml serum containing medium(1% FCS) per well were seeded into 96-well plates andcultured for 48 h. Cells were treated for 24 h with theSWCNT–SC suspension, the solvent control SC and TritonX-100 as a positive control for maximal cell membranedamage. The assay was performed according to the man-ufacturers’ protocol (Cytotoxicity Detection Kit (lactate dehy-drogenase; LDH), Roche). After incubation, 100 ml culturesupernatant were removed cell-free and transferred intowells of a clean 96-well flat bottommicroplate. Subsequently,100 ml freshly prepared reaction mixture (included in theCytotoxicity Detection Kit (LDH), Roche) were added andincubated for 5 min at room temperature.
An increase in the amount of dead or plasma membrane-damaged cells results in an increase of the LDH enzymeactivity in the culture supernatant. Released LDH reducesNAD+ to NADH and H+ by oxidation of lactate to pyruvate. Inthe second enzymatic reaction, 2 H+ are transferred to theyellow tetrazolium salt 2-[4-iodophenyl]-3-[4-nitrophenyl]-5-phenyltetrazolium chloride. The amount of formed for-mazan (red) in the assay is proportional to the number oflysed cells and can be measured photometrically at 492 nm.Cytotoxicity was calculated as percent survival, determinedby the number of treated over control cells � 100% (T/C).
DCF assayThe 2¢,7¢-dichlorofluorescein (DCF) assay was performedusing dark 96-well culture plates seeded with HT29 cellsat a density of 4 � 104 in 100 ml serum containing mediumper well which were subsequently allowed to grow for 48 h.Afterwards, the medium was removed from the well plates,cells were washed two times with 100 ml phosphate bufferper well and incubated with 100 ml 2¢,7¢-dichlorofluorescindiacetate solution (DCFH-DA, 50 mM) for 30 min at 37�C.
During this incubation time, the non-fluorescent andnon-polar DCFH-DA passes the cell membrane and is de-acetylated by cellular esterases to the non-fluorescent butpolar DCFH, which therefore remains inside the cell.
After incubation, the dye-containing supernatant wasremoved and the cells were washed twice with phosphatebuffer. For SWCNT exposure, 200 ml of the respectiveSWCNT–SC suspension in increasing concentrations wasadded to the cells. Therefore, SWCNT–SC suspension wasdispersed in serum-containing (1% FCS) but achromaticmedium (without phenol red) by vortexing. Menadione(MEN) was included in the testing as a positive controlfor the induction of ROS.
In the presence of ROS within the cell, the non-fluorescent DCFH is oxidised to the fluorescent DCF. Theincrease in fluorescence intensity was measured with afluorometer using 485 nm excitation and 535 nm emissionfilters over a period of 3 h at regular intervals. Between themeasurement intervals, plates were stored in the incubatorat 37�C. Tests were carried out four times using cells fromfour different passages. Each SWCNT concentration wasmeasured six times resulting in eighteen replicates. Themean values of the slopes of the linear fluorescence intensityincrease were computed for each replicate followed by the
SWCNT toxicity in human colon cells
Nan
otox
icol
ogy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
ity o
f M
elbo
urne
on
03/0
1/13
For
pers
onal
use
onl
y.
subtraction of the corresponding plate’s mean blank value.Subsequently, the average was calculated over all indepen-dent experiments. For each concentration, the resultingslope was then normalised with the corresponding slopefor the SC-exposed cells (negative control) and expressed asrelative fluorescence units (%).
GSH assayThe cellular GSH level was determined according to themethod of Tietze (1969). HT29 cells (1.2 � 106 in 10 ml ofserum-containing medium) were spread into Petri dishes(Ø 10 cm) and allowed to grow to 80% confluence. Subse-quently, cells were incubated for 3 and 24 h with theSWCNT–SC suspension. Therefore, the suspension was dis-persed in serum-containing medium by vortexing. Tert-butylhydroquinone (t-BHQ) was used as positive controlfor the 3 h incubation whereas L-buthionine-sulphoximine(BSO), a synthetic amino acid, served as positive control forthe 24 h incubation. t-BHQ produces ROS by redox cyclingafter autoxidation to tert-butylquinone. BSO is an irreversibleinhibitor of the g-glutamylcysteine-synthetase, which resultsin a decrease of the GSH biosynthesis.
Cells were collected and the viability was determined bytrypan blue exclusion. After several washing steps with ice-cold phosphate buffer, cells were centrifuged at 420 g for10 min at 4�C. The resulting cell pellet was resuspended in1.6 ml A/B-buffer (15 ml of buffer A (125 mM KH2PO4, 6 mMNa2EDTA) and 85 ml of buffer B (125 mM K2HPO4, 6 mMNa2EDTA)) and centrifuged at 420 g for 10 min at 4�C.Thereafter, 370 ml of A/B-buffer were added to the resultingcell pellet, 2 � 10 ml of this cell suspension were used forprotein quantification whereas the remaining cell suspen-sion wasmixed with 350 ml of 10% sulphosalicylic acid for celllysis. Subsequently, the suspension was centrifuged at fullspeed for 10 min at 4�C to remove the protein precipitate.
For quantitative determination of total (reduced andoxidised) glutathione (tGSH), 10 ml of the supernatantwere mixed with 190 ml of the tGSH mixture (164 ml A/B-buffer, 20 ml 6 mM 5,5¢-dithio-bis(2-nitrobenzoic acid), 4 ml20 mM nicotinamide adenine dinucleotide phosphate(NADPH), 2 ml glutathione reductase (GR) solution(50 U/ml)).
In this reaction, oxidised glutathione (GSSG) is reducedto GSH by GR and NADPH. Finally, the total GSH level ofthe cell (reduced GSSG and cellular GSH) is measuredphotometrically (l = 405 nm) due to the reaction ofEllman’s reagent (5,5¢-dithio-bis(2-nitrobenzoic acid) to5-thio-2-nitrobenzoate).
For measurement of the GSSG level, the thiol-scaveningreagent 2-vinylpyridinium (2-VP) was used to prevent theparticipation of the reduced form (GSH) in the enzymaticassay. Therefore, 500 ml of the supernatant, 20 ml 2-VP and100 ml 50% triethanolamine solution were mixed andallowed to react at 900 rpm for 1 h at 26�C in a thermo-mixer. Subsequently, 180 ml of the GSSG mixture (154 mlA/B-buffer, 20 ml 6 mM 5,5¢-dithio-bis(2-nitrobenzoic acid),4 ml 20 mM NADPH, 2 ml GR solution (50 U/ml)) wereadded to 20 ml of the derivatised supernatant. The GSSGlevel was determined photometrically due to the reaction of
Ellman’s reagent as described before. The tGSH contentwas determined as nmol tGSH/mg protein. The results aregiven as percent relative to the tGSH content of the controlcells (SC control) set to 100%.
BCA assayCellular protein of the cell suspension in the GSH assay wasquantified with the bicinchoninic acid (BCA) assay. Theprinciple of the BCA assay relies on the formation of aCu2+–protein complex under alkaline conditions, followedby reduction of Cu2+ to Cu+. The amount of reduction isproportional to the protein present. The assay was per-formed according to the protocol of Sigma Aldrich (Bicinch-oninic Acid Protein Assay Kit). Absorbance was measured at592 nm.
Single cell gel electrophoresis (comet assay)Single cell gel electrophoresis was performed under alkalineand neutral electrophoresis conditions according to themethod of Tice et al. (2000). HT29 cells (3 � 105 in 5 mlof serum containing medium) were spread into Petri dishes(Ø 5.5 cm) and allowed to grow for 48 h. Subsequently,HT29 cells were treated for 3 and 24 h with the SWCNT–SCsuspension. Therefore, SWCNT–SC dilutions were dispersedin serum containing medium by vortexing. In the cometassay under alkaline conditions, UV irradiation was includedin the testing as a positive control for the induction ofoxidative stress. Etoposide (ETO), a topoisomerase II poison,was used at a concentration of 50 mM as a positive control inthe comet assay under neutral electrophoresis conditions.Cells were collected and the viability was determined bytrypan blue exclusion. Thereafter, aliquots corresponding to30,000 cells were centrifuged at 425 g for 10 min. Theresulting cell pellet was resuspended in 65 ml of low meltingagarose and distributed onto a frosted glass microscopeslide, precoated with a layer of normal melting agarose.The slides were coverslipped and kept at 4�C for 10 minto allow solidification of the agarose. After the cover glasswas removed, slides were immersed over night at 4�C in lysissolution (89 ml of lysis stock solution: 2.5 M NaCl, 100 mMEDTA, 10 mM Tris HCl, pH 10 (pH 7.5 for neutral electro-phoresis conditions); 1% (w/v) N-laurylsarcosylsodium salt,10 ml DMSO and 1 ml Triton-X-100).
In the comet assay under alkaline conditions, additionaltreatment of the samples with the DNA repair enzymeformamidopyrimidine-DNA-glycosylase (FPG) allows thedetection of FPG-sensitive sites after DNA unwinding. There-fore, cell-slides treated under alkaline conditions werewashed three times in FPG buffer (40 mM HEPES, 0.1 MKCl, 0.5 mM EDTA, pH 8; 0.2 mg/ml BSA (bovine serumalbumin)). The samples were incubated with a 1:3000 FPGsolution for 30 min. Subsequently, DNA was allowed tounwind for 20 min at 4�C (300 mM NaOH, 1 mM EDTA,pH 13.5) followed by horizontal gel electrophoresis at 4�C for20 min (25 V, 300 mA). For the comet assay under neutralconditions, the electrophoresis buffer was adjusted to pH7.5 (90 mM Tris, 90 mM boric acid, 2 mM EDTA). Thereafter,the slides were washed three times with 0.4 M Tris HCl, pH7.5 and stained with ethidium bromide (40 ml per coverslip,
J. Pelka et al.
Nan
otox
icol
ogy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
ity o
f M
elbo
urne
on
03/0
1/13
For
pers
onal
use
onl
y.
20 mg/ml). Fluorescence microscopy was performed with aZeiss Axioskop (lex = 546 ± 12 nm; lem ‡590 nm). Slides weresubjected to computer-aided image analysis (Comet AssayIII System, Perceptive Instruments, Suffolk, Great Britain),scoring 50 images per slide randomly picked from eachelectrophoresis. For each concentration of SWCNTs, twoslides were independently processed and analysed. Theresults were parameterised with respect to tail intensity(intensity of the DNA in the comet tail calculated as per-centage of overall DNA intensity in the respective cell). Thesequantitative data were derived from at least four indepen-dent sets of experiments and from the evaluation of 100 indi-vidual cells per concentration (50/slide) in each experiment.
Micronucleus assayA total of 1 � 105 V79 cells were seeded per slide in 5 mlserum containing medium in quadriperms and allowed togrow for 24 h. Thereafter, cells were treated with theSWCNT–SC suspension, SC as solvent control and the pos-itive control nitroquinolineoxide (0.3 mM, positive control forkinetochore negative micronuclei (MN)) for 24 h in serum-containing medium. Immediately after compound treat-ment, the cells were postincubated compound-free for 3 hand fixed with methanol at �20�C. Briefly, non-specificbinding was blocked by incubation with goat serum for1 h at 37�C. Slides were then incubated with a 1% BSA–phosphate-buffered saline (PBS)-CMF solution containingCREST antibody (diluted 1:100, Antibodies Incorporated,Davis, CA, USA) and monoclonal mouse anti-a-tubulinantibody (diluted 1:500, Sigma Aldrich) for 1 h at 37�C. Afterwash steps in PBS-CMF solution, incubation for 1 h at 37�Cwith a 1% BSA–PBS-CMF solution containing the secondaryantibodies goat polyvalent anti-human antibody (FITC con-jugated, diluted 1:200, Sigma Aldrich) and goat anti-mouseantibody (CY3 conjugated, diluted 1:125, Dianova, Hamburg,Germany) followed. Finally, slides were treated with anantifade solution containing 1 mg DAPI/ml antifade solution.On each slide, cell nuclei and MN were visualised using UVexcitation and the microtubes and kinetochore signals wereanalysed under green and blue illumination. Additionally, allsignals could be visualised simultaneously by means of atriple band-pass filter. At least 2000 cells per slide werescored with respect to MN formation.
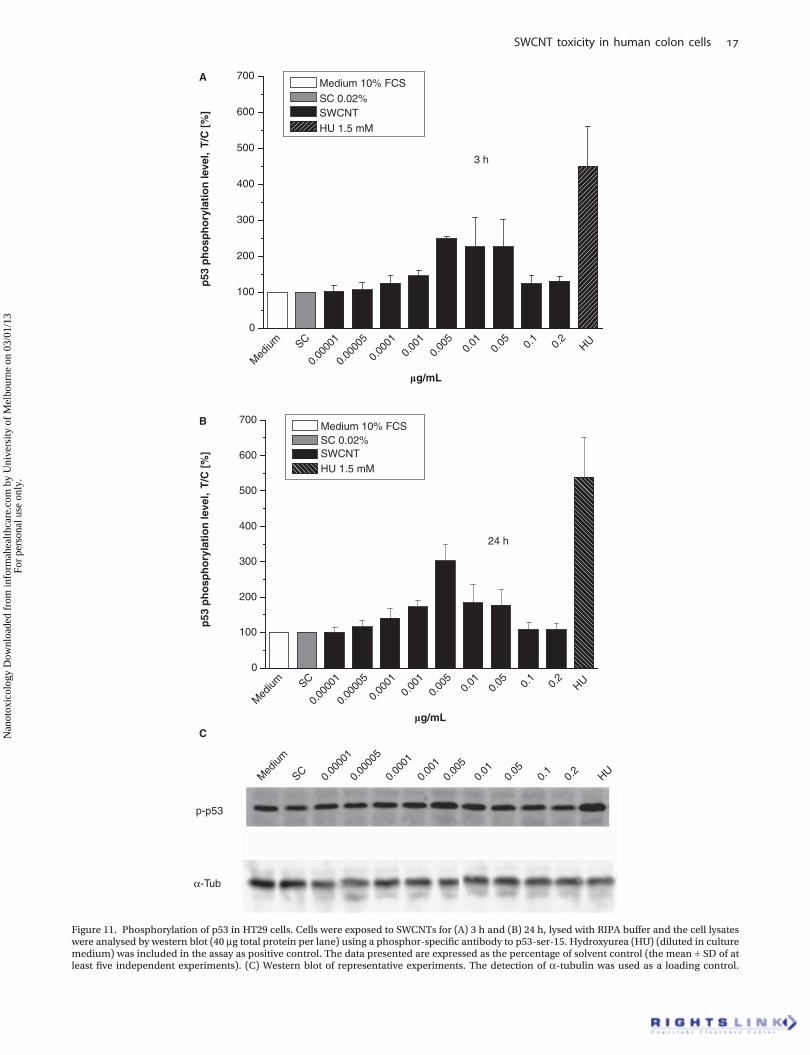
Western blot analysisPhosphorylation of the tumour suppressor protein p53. A totalof 1.2� 106 HT29 cells were seeded in 10 ml per Petri dish (Ø10 cm) and allowed to grow for 48 h. Thereafter, cells weretreated with the SWCNT–SC suspension and the control SCfor 3 and 24 h in serum-containing medium. Cell incubationwith hydroxyurea (HU, 1.5 mM) for 3 h served as positivecontrol. Subsequently, cells were washed twice with ice-cold phosphate buffer and lysed at 4�C in 0.2 ml RIPA buffer(50 mM Tris/HCl, pH 7.4, 250 mM NaCl, 1 mM EDTA, 1 mMNaF, 1% v/v Igepal; 1 mM PMSF, 1 mM sodium orthova-nadate and 2% v/v protease inhibitor cocktail (Roche Diag-nostics, Mannheim, Germany) were freshly added to RIPAbuffer). Thereafter, the lysate was homogenised thoroughlyand subsequently centrifuged for 10 min (20,000 g, 4�C). The
proteins were separated by SDS-PAGE (7% polyacrylamidegel) and transferred onto a nitrocellulose membrane. West-ern blot was performed using a rabbit polyclonal antibodyagainst human p53 phosphorylation (p-p53) (53 kDa; SantaCruz, Heidelberg, Germany) and an anti-rabbit IgG perox-idase conjugate (Santa Cruz, Heidelberg, Germany) as asecondary antibody. a-Tubulin was used as a loading con-trol. The respective chemoluminescent signals (Lumi-GLO,Cell Signaling Technology) were analysed using the LAS3000 system with the AIDA Image Analyzer 3.52 softwarefor quantification (Raytest, Straubenhardt, Germany). Arbi-trary Light Units were plotted as test over control (T/C) (%).
Nuclear translocation of the transcription factor Nrf2. In all,4.5 � 106 HT29 cells were seeded per Petri dish (Ø 15 cm) inDMEM medium supplemented with FCS (10%) and 1% PSand allowed to grow for 48 h. Subsequently, cells weretreated for 1 and 3 h with SWCNT–SC suspension. Chloro-genic acid (100 mM) was used as positive control due to itsantioxidative properties. After the respective incubationtime, the culture medium was removed and the cells wererinsed with ice-cold PBS and abraded on ice with 0.7 ml ofPBS. Thereafter, the lysate was centrifuged for 5 min (800 g,4�C). The pellet was resolved in 200 ml of buffer A (10 mMHepes, pH 7.9, 10 mM KCl, 0.1 mM EDTA, 0.1 mM EGTA,1 mM DTT and 1 mM protease inhibitor cocktail (RocheDiagnostics, Mannheim, Germany) freshly added to bufferA). After incubation for 15 min, 17 ml of Nonidet-P40 (10%)was added, and the lysate was vortexed for 15 sec. Toseparate the nuclear from the cytosolic fraction, the lysatewas repeatedly centrifuged for 10 min (800 g, 4�C). Thesupernatant (cytosolic fraction) was stored on ice, and thepellet was resuspended in 60 ml of buffer B (20 mM Hepes,pH 7.9, 0.4 MNaCl, 1 mMEDTA, 1 mMEGTA, 1 mMDTT and1 mM protease inhibitor cocktail (Roche Diagnostics) freshlyadded to buffer B) and incubated on ice for 15 min.During the incubation time, the lysate was vortexed for10–15 sec every 2 min. After a final centrifugation step of10 min (20,000 g, 4�C), the proteins of the nuclear (super-natant) and the cytosolic fraction were separated by SDS-PAGE (10% polyacrylamide gel) and transferred ontoa nitrocellulose membrane. Western blot was perfor-med using rabbit polyclonal antibody against humanNrf2 (H-300, Santa Cruz, Heidelberg, Germany) and ananti-rabbit IgG peroxidase conjugate (Santa Cruz) as sec-ondary antibody. a-Tubulin was used as a loading control.Lamin B was used as separation control of the cytosolic andnuclear fraction. The respective chemoluminescent signals(Lumi-GLO, Cell Signaling Technology) were analysed asdescribed above.
Results
Dispersion and characterisation of the SWCNTsuspensionSWCNT dispersionRaman spectra of SWCNT samples dispersed in surfactantssuch as SDS, SC, amylose, PVP, lecithin, gum arabica andb-cyclodextrin revealed SDS and SC to be the most efficient
SWCNT toxicity in human colon cells
Nan
otox
icol
ogy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
ity o
f M
elbo
urne
on
03/0
1/13
For
pers
onal
use
onl
y.
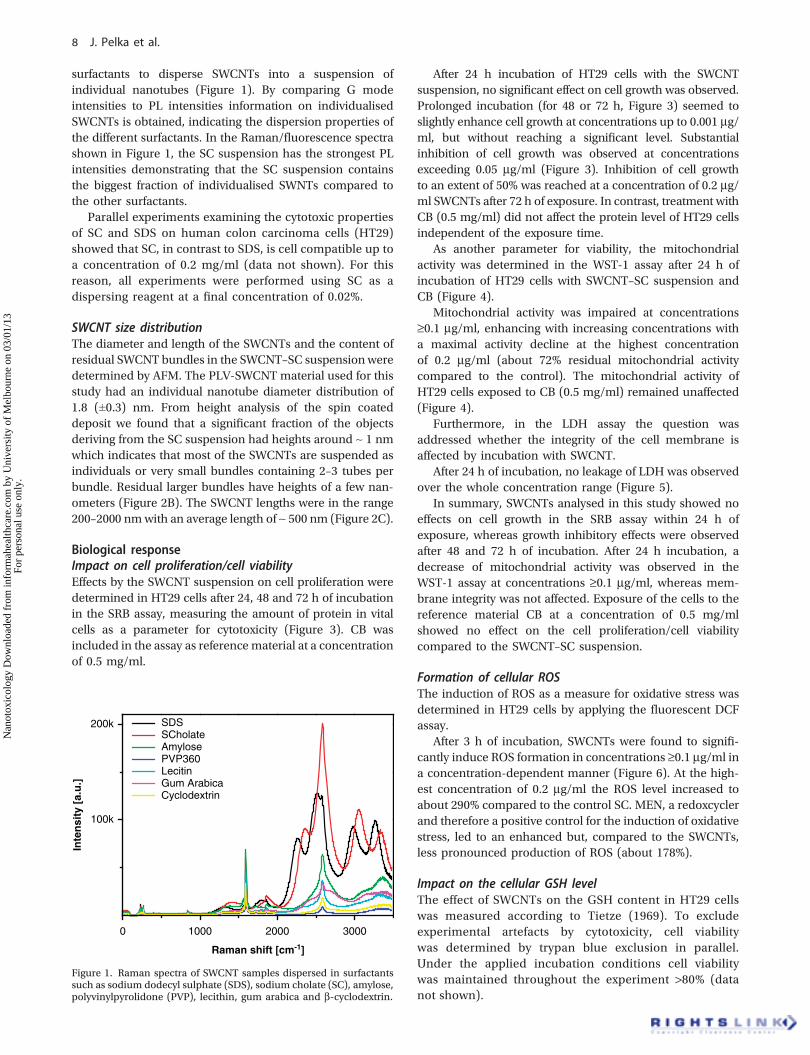
surfactants to disperse SWCNTs into a suspension ofindividual nanotubes (Figure 1). By comparing G modeintensities to PL intensities information on individualisedSWCNTs is obtained, indicating the dispersion properties ofthe different surfactants. In the Raman/fluorescence spectrashown in Figure 1, the SC suspension has the strongest PLintensities demonstrating that the SC suspension containsthe biggest fraction of individualised SWNTs compared tothe other surfactants.
Parallel experiments examining the cytotoxic propertiesof SC and SDS on human colon carcinoma cells (HT29)showed that SC, in contrast to SDS, is cell compatible up toa concentration of 0.2 mg/ml (data not shown). For thisreason, all experiments were performed using SC as adispersing reagent at a final concentration of 0.02%.
SWCNT size distributionThe diameter and length of the SWCNTs and the content ofresidual SWCNT bundles in the SWCNT–SC suspension weredetermined by AFM. The PLV-SWCNT material used for thisstudy had an individual nanotube diameter distribution of1.8 (±0.3) nm. From height analysis of the spin coateddeposit we found that a significant fraction of the objectsderiving from the SC suspension had heights around ~ 1 nmwhich indicates that most of the SWCNTs are suspended asindividuals or very small bundles containing 2–3 tubes perbundle. Residual larger bundles have heights of a few nan-ometers (Figure 2B). The SWCNT lengths were in the range200–2000 nmwith an average length of ~ 500 nm (Figure 2C).
Biological responseImpact on cell proliferation/cell viabilityEffects by the SWCNT suspension on cell proliferation weredetermined in HT29 cells after 24, 48 and 72 h of incubationin the SRB assay, measuring the amount of protein in vitalcells as a parameter for cytotoxicity (Figure 3). CB wasincluded in the assay as reference material at a concentrationof 0.5 mg/ml.
After 24 h incubation of HT29 cells with the SWCNTsuspension, no significant effect on cell growth was observed.Prolonged incubation (for 48 or 72 h, Figure 3) seemed toslightly enhance cell growth at concentrations up to 0.001 mg/ml, but without reaching a significant level. Substantialinhibition of cell growth was observed at concentrationsexceeding 0.05 mg/ml (Figure 3). Inhibition of cell growthto an extent of 50% was reached at a concentration of 0.2 mg/ml SWCNTs after 72 h of exposure. In contrast, treatment withCB (0.5 mg/ml) did not affect the protein level of HT29 cellsindependent of the exposure time.
As another parameter for viability, the mitochondrialactivity was determined in the WST-1 assay after 24 h ofincubation of HT29 cells with SWCNT–SC suspension andCB (Figure 4).
Mitochondrial activity was impaired at concentrations‡0.1 mg/ml, enhancing with increasing concentrations witha maximal activity decline at the highest concentrationof 0.2 mg/ml (about 72% residual mitochondrial activitycompared to the control). The mitochondrial activity ofHT29 cells exposed to CB (0.5 mg/ml) remained unaffected(Figure 4).
Furthermore, in the LDH assay the question wasaddressed whether the integrity of the cell membrane isaffected by incubation with SWCNT.
After 24 h of incubation, no leakage of LDH was observedover the whole concentration range (Figure 5).
In summary, SWCNTs analysed in this study showed noeffects on cell growth in the SRB assay within 24 h ofexposure, whereas growth inhibitory effects were observedafter 48 and 72 h of incubation. After 24 h incubation, adecrease of mitochondrial activity was observed in theWST-1 assay at concentrations ‡0.1 mg/ml, whereas mem-brane integrity was not affected. Exposure of the cells to thereference material CB at a concentration of 0.5 mg/mlshowed no effect on the cell proliferation/cell viabilitycompared to the SWCNT–SC suspension.
Formation of cellular ROSThe induction of ROS as a measure for oxidative stress wasdetermined in HT29 cells by applying the fluorescent DCFassay.
After 3 h of incubation, SWCNTs were found to signifi-cantly induce ROS formation in concentrations ‡0.1 mg/ml ina concentration-dependent manner (Figure 6). At the high-est concentration of 0.2 mg/ml the ROS level increased toabout 290% compared to the control SC. MEN, a redoxcyclerand therefore a positive control for the induction of oxidativestress, led to an enhanced but, compared to the SWCNTs,less pronounced production of ROS (about 178%).
Impact on the cellular GSH levelThe effect of SWCNTs on the GSH content in HT29 cellswas measured according to Tietze (1969). To excludeexperimental artefacts by cytotoxicity, cell viabilitywas determined by trypan blue exclusion in parallel.Under the applied incubation conditions cell viabilitywas maintained throughout the experiment >80% (datanot shown).
Figure 1. Raman spectra of SWCNT samples dispersed in surfactantssuch as sodium dodecyl sulphate (SDS), sodium cholate (SC), amylose,polyvinylpyrolidone (PVP), lecithin, gum arabica and b-cyclodextrin.
J. Pelka et al.
Nan
otox
icol
ogy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
ity o
f M
elbo
urne
on
03/0
1/13
For
pers
onal
use
onl
y.
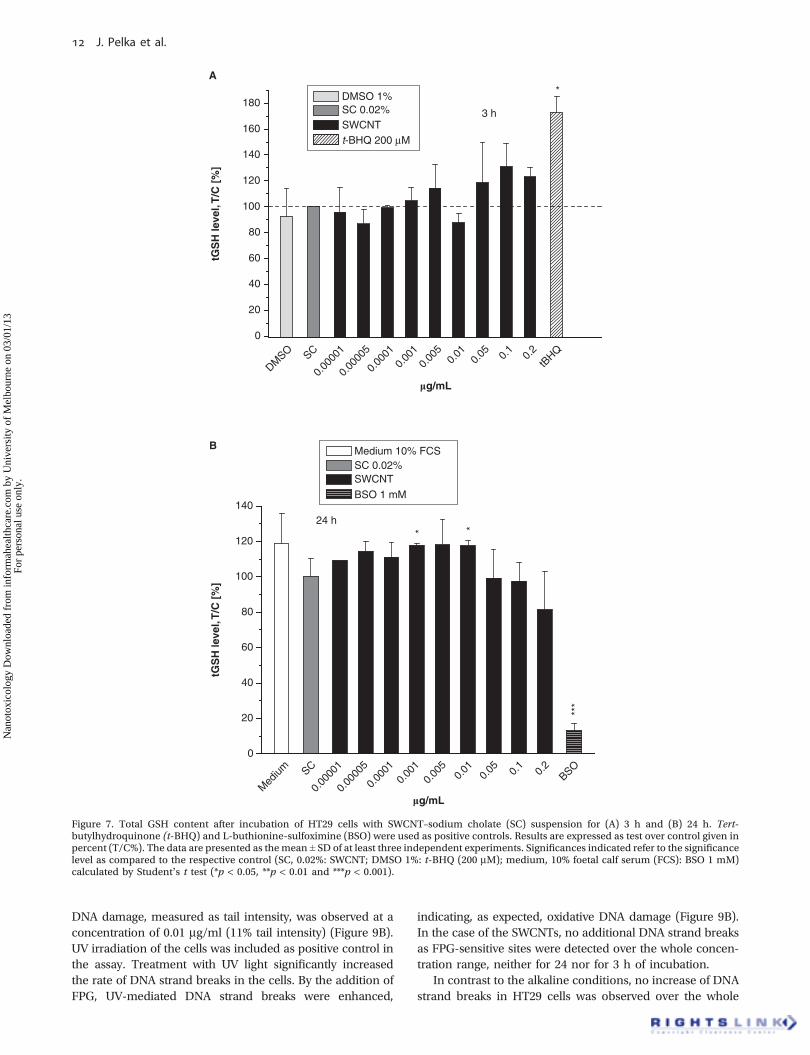
The assay was performed for an incubation time of 3 and24 h over a concentration range of 0.00001–0.2 mg/ml. Theresults are expressed as tGSH (Figure 7A,B).
An exposure to SWCNTs for 3 h resulted in an increase ofthe intracellular tGSH level up to 131% of the negativecontrol SC (0.1 mg/ml) (Figure 7A), albeit no statisticalsignificance was reached. After an incubation time of 24 hthe SWCNT suspension slightly enhanced the intracellulartGSH level in a concentration range of 0.00001–0.01 mg/mlup to 118% (0.005 mg/ml) of the negative control SC.A further increase of the SWCNT concentrations reversedthe effect slightly decreasing the tGSH content to approxi-mately 81%. However, this effect was not statisticallysignificant (Figure 7B).
Nuclear translocation of the transcription factor Nrf2As a measure for the occurrence of oxidative stress, therelease of Nrf2 from the Keap-1–Nrf2 complex and its accu-mulation in the nucleus was determined by western blotanalysis.
After an incubation time of 1 h, the SWCNT–SC sus-pension showed a marginal influence on the Nrf2 translo-cation by increasing the nuclear Nrf2 level up to 138 ± 22%(0.2 mg/ml) of the control SC (Figure 8A). An exposure toSWCNTs for 3 h did not lead to any accumulation ofthe transcription factor in the nucleus over the wholeconcentration range (data not shown).
Induction of DNA strand breaksThe effect of the SWCNT suspension on DNA integrity wasmeasured as DNA strand breaks in HT29 cells by single cell gelelectrophoresis (comet assay) under alkaline as well as neutralelectrophoresis conditions. In addition to the standard cometassay protocol, treatment with the DNA repair enzyme FPGwas included in the assay (under alkaline conditions) as anindication for oxidative DNA damage. In parallel to all experi-ments, cell viability was determined by trypan blue exclusionto exclude experimental artefacts by cytotoxicity. Under theapplied incubation conditions, cell viability was maintainedthroughout the experiment at >80% (data not shown).
0 1 2 3 4
0
20
40
60 Fraction 2(1.8 ± 0.3) nm
Co
un
t
Height [nm]
7.5
5.0
2.5
010.0
μm 0.0 nm
1.0 nm
2.0 nm
7.55.02.50
10.0
0 1000 2000 30000
10
Fraction 2(530 ± 200) nm
Co
un
t
Length [nm]
A
B C
Figure 2. Diameter and length of the sodium cholate (SC) dispersed SWCNTs determined by atomic force microscopy (AFM). Representative AFMpicture (A), diameter (B) and length distribution (C).
SWCNT toxicity in human colon cells
Nan
otox
icol
ogy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
ity o
f M
elbo
urne
on
03/0
1/13
For
pers
onal
use
onl
y.
SC
0.00
005
0.00
010.
001
0.00
50.
010.
05 0.1
0.2
CB0
20
40
60
80
100
120
140
* ***
***
***
24 h
48 h 72 h
Cel
l gro
wth
, T/C
[%
]
mg/mL
Figure 3. Effects of SWCNTs on the cell growth of HT29 cells after 24, 48 and 72 h of incubation determined by the sulphorhodamine B (SRB)assay. Carbon black (CB, 0.5 mg/ml) was included in the assay as reference material. Growth modulation was calculated as survival of treatedcells over control cells � 100 (T/C%). The data are presented as the mean ± SD of at least three independent experiments. Significancesindicated refer to the significance level as compared to the control sodium cholate (SC) (0.02%) calculated by Student’s t test (*p < 0.05,**p < 0.01 and ***p < 0.001).
Med
ium SC
0.00
005
0.00
010.
001
0.00
50.
010.
05 0.1
0.2
CB0
20
40
60
80
100
120
140
160
**
Mit
och
on
dri
al a
ctiv
ity,
T/C
[%
]
Medium 10% FCS
SC 0.02%
SWCNT Quercetin 500 μM
mg/mL
24 h
***
Querc
etin
Figure 4. Mitochondrial activity of HT29 after 24 h of incubation determined in the WST-1 assay. Quercetin (solvent control: DMSO 1%)was included in the assay as a positive control. Carbon black (CB, 0.5 mg/ml) was used as reference material. Mitochondrial activity isexpressed as test over control given in percent (T/C%). The data are presented as the mean ± SD of at least six independentexperiments each performed sixfold. Significances indicated refer to the significance level as compared to the respective control (sodiumcholate (SC), 0.02%: SWCNT, CB; medium, 10% foetal calf serum (FCS): quercetin, 500 mM) calculated by Student’s t test (*p < 0.05 and***p < 0.001).
J. Pelka et al.
Nan
otox
icol
ogy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
ity o
f M
elbo
urne
on
03/0
1/13
For
pers
onal
use
onl
y.
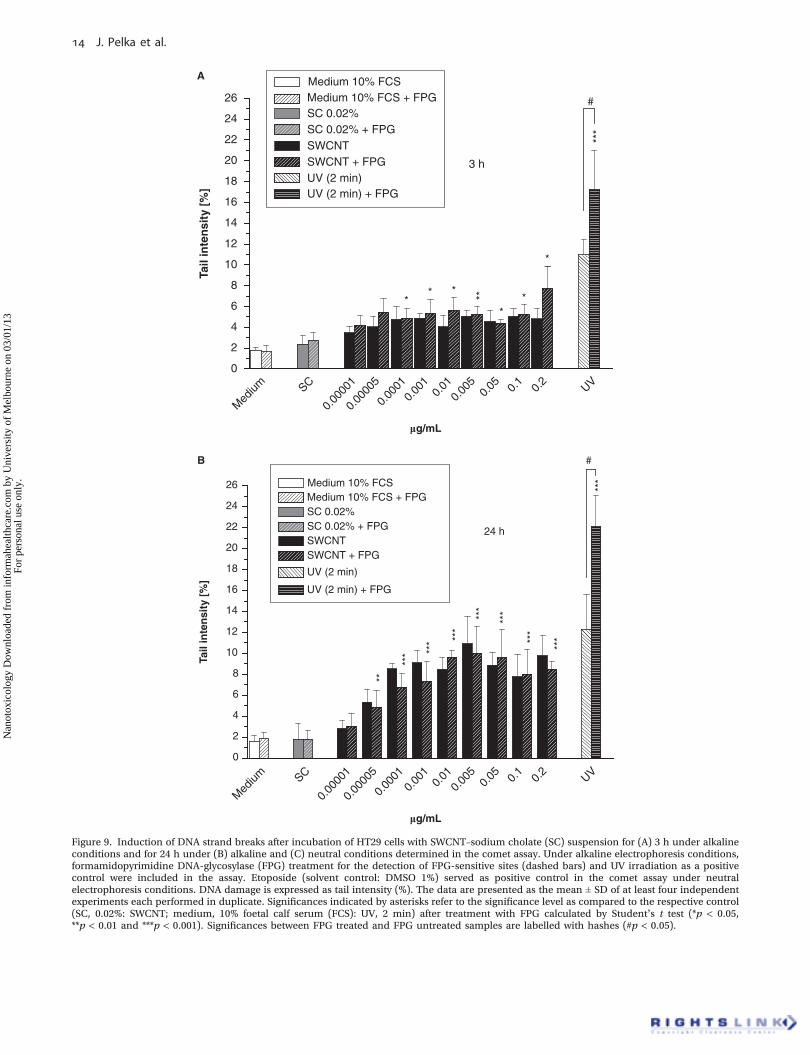
Comet assay under alkaline electrophoresis conditionswas performed after 3 and 24 h of exposure to SWCNT–SCsuspension. After an incubation time of 3 h only slight butsignificant DNA strand breaking properties were observed atconcentrations ‡0.0001 mg/ml (Figure 9A).
After 24 h of incubation the SWCNT–SC suspension wasfound to further increase the rate of DNA strand breaks inHT29 cells (Figure 9B).
These DNA strand breaking properties occurred alreadyat a concentration of 0.00005 mg/ml (Figure 9B). The highest
Med
ium SC
0.00
010.
001
0.00
50.
010.
05 0.1
0.2
Trito
n X 1
% CB0
20
40
60
80
100
120
140600
800
1000
1200
1400
1600
Mem
bra
ne
inte
gri
ty, T
/C [
%]
Medium 1% FCSSC 0.02%
SWCNTTriton X-100 1%
mg/mL
24 h
***
Figure 5. Membrane integrity of HT29 after 24 h of exposure to SWCNT–sodium cholate (SC) suspension determined in the lactate dehydrogenase(LDH) assay. Triton X-100 was used as a positive control for maximal cell membrane damage. Carbon black (CB, 0.5 mg/ml) was included in theassay as reference material. The data presented are expressed as the percentage of solvent control, representing the mean ± SD of at least fiveindependent experiments each performed sixfold. Significances indicated refer to the significance level as compared to the respective control (SC,0.02%: SWCNT, CB; medium, 10% foetal calf serum (FCS): Triton X-100, 1%) calculated by Student’s t test (*p < 0.05, **p < 0.01, ***p < 0.001).
SC
0.00
005
0.00
010.
001
0.00
50.
010.
05 0.1
0.2
MEN
0
50
100
150
200
250
300
350
400
450
**
***
RO
S g
ener
atio
n, R
FU
[%
]
SC 0.02% SWCNT
MEN 20 μM
µg/mL
3 h
***
Figure 6. Reactive oxygen species (ROS) formation in HT29 cells induced by incubation with SWCNT–sodium cholate (SC) suspension for 3 hdetermined in the DCF assay. The redox cycler menadione (MEN (20 mM), solvent control: DMSO 1%) was included in the assay as a positivecontrol. ROS generation is expressed as relative fluorescence units (RFU (%)). The increase in fluorescence intensity was measured with afluorimeter using 485 nm excitation and 535 nm emission filters. The data are presented as the mean ± SD of at least four independent experimentseach performed sixfold. Significances indicated refer to the significance level as compared to the respective control calculated by Student’s t test(*p < 0.05, **p < 0.01 and ***p < 0.001).
SWCNT toxicity in human colon cells
Nan
otox
icol
ogy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
ity o
f M
elbo
urne
on
03/0
1/13
For
pers
onal
use
onl
y.
DNA damage, measured as tail intensity, was observed at aconcentration of 0.01 mg/ml (11% tail intensity) (Figure 9B).UV irradiation of the cells was included as positive control inthe assay. Treatment with UV light significantly increasedthe rate of DNA strand breaks in the cells. By the addition ofFPG, UV-mediated DNA strand breaks were enhanced,
indicating, as expected, oxidative DNA damage (Figure 9B).In the case of the SWCNTs, no additional DNA strand breaksas FPG-sensitive sites were detected over the whole concen-tration range, neither for 24 nor for 3 h of incubation.
In contrast to the alkaline conditions, no increase of DNAstrand breaks in HT29 cells was observed over the whole
DMSO SC
0.00
001
0.00
005
0.00
010.
001
0.00
50.
010.
05 0.1
0.2
tBHQ
0
20
40
60
80
100
120
140
160
180
tGS
H le
vel,
T/C
[%
]
mg/mL
DMSO 1%SC 0.02%SWCNTt-BHQ 200 μM
*
3 h
Med
ium SC
0.00
001
0.00
005
0.00
010.
001
0.00
50.
010.
05 0.1
0.2
BSO0
20
40
60
80
100
120
140
* *
***
tGS
H le
vel,
T/C
[%
]
mg/mL
Medium 10% FCS SC 0.02% SWCNT
BSO 1 mM
24 h
A
B
Figure 7. Total GSH content after incubation of HT29 cells with SWCNT–sodium cholate (SC) suspension for (A) 3 h and (B) 24 h. Tert-butylhydroquinone (t-BHQ) and L-buthionine-sulfoximine (BSO) were used as positive controls. Results are expressed as test over control given inpercent (T/C%). The data are presented as themean ± SD of at least three independent experiments. Significances indicated refer to the significancelevel as compared to the respective control (SC, 0.02%: SWCNT; DMSO 1%: t-BHQ (200 mM); medium, 10% foetal calf serum (FCS): BSO 1 mM)calculated by Student’s t test (*p < 0.05, **p < 0.01 and ***p < 0.001).
J. Pelka et al.
Nan
otox
icol
ogy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
ity o
f M
elbo
urne
on
03/0
1/13
For
pers
onal
use
onl
y.
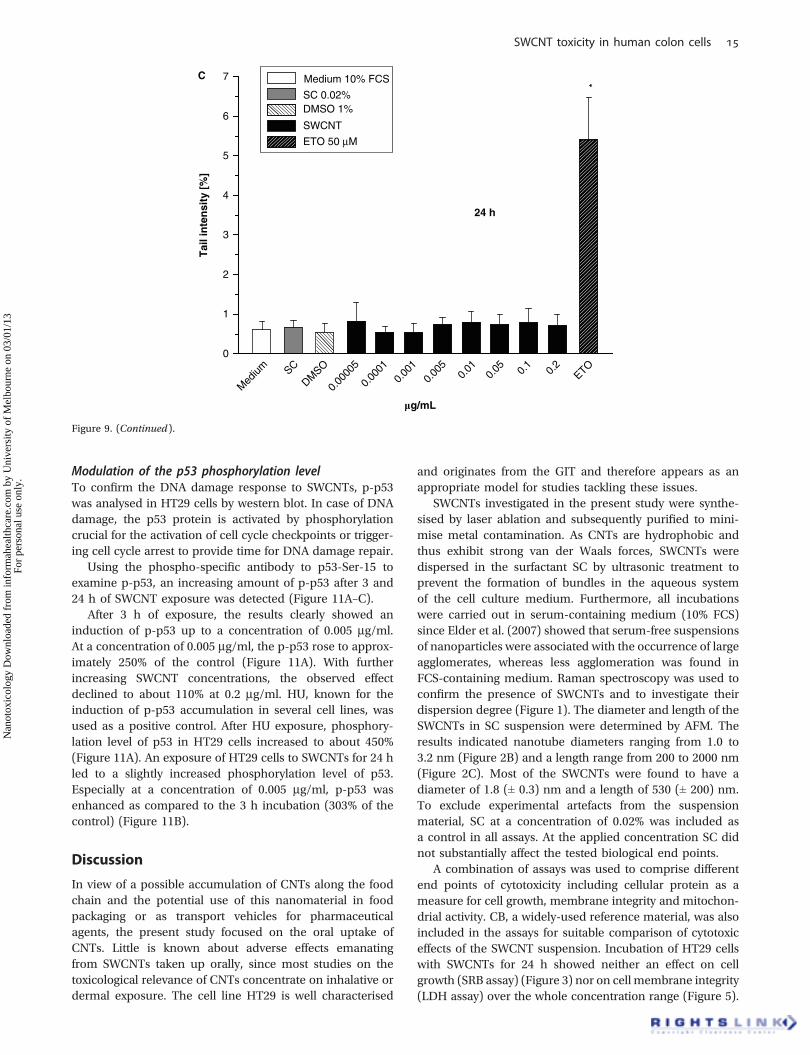
concentration range under neutral electrophoresis condi-tions (Figure 9C) indicating no DNA double strand breakinduction after 24 h of exposure to the SWCNT–SCsuspension.
ETO, a topoisomerase II poison and therefore a referencecompound for the induction of DNA double strand breaks,was included in the assay as a positive control. ETO signif-icantly increased the rate of DNA strand breaks at a con-centration of 50 mM after 24 h of incubation up to 5.5% tailintensity (Figure 9C).
Induction of MNTo investigate whether enhanced levels of DNA strandbreaks correlate with the induction of chromosome andgenome mutations (i.e., MN containing DNA fragmentsand MN containing whole chromosomes), cells treatedwith SWCNTs were analysed with respect to MN formation.The male Chinese hamster V79 lung fibroblasts werechosen for studies on MN formation as they are charac-terised by short generation time and high cloning efficiency.
Furthermore, the morphology of the microtubules of V79cells allows a clear identification of MN.
V79 cells were treated for 24 h with SWCNT–SC followedby a compound-free postincubation period of 3 h. Theoccurrence of micronucleated cells was expressed as numberof MN per 1000 cells. MN containing whole chromosomeswere detected by immunochemical staining of kinetochoreproteins (Figure 10).
The spontaneous frequency of micronucleated cellscontaining whole chromosomes and DNA fragments inthe applied V79 cell line was 1 ± 0 and 3 ± 1, respectively,after treatment with SC alone for 24 h (Figure 10).After exposure to SWCNTs, the number of micronuc-leated cells containing DNA fragments (dashed bars)increased slightly in a concentration range of 0.00005–0.2 mg/ml, with a maximum of 11 ± 2 at a concentra-tion of 0.05 mg/ml. However, the number of MNcontaining whole chromosomes remained nearly at thelevel of SC, indicating no induction of kinetochore-positive MN.
0
20
Nrf2
α-Tub
40
60
80
100
120
140
160
180
200
220
CGASC
Med
ium 0.2
0.1
0.05
0.01
0.00
50.
001
0.00
01
0.00
005
Nu
clea
r N
rf2
leve
l, T
/C [
%]
mg/mL
Medium 10% FCSSC 0.02%SWCNTCGA 100 μM
0.00
001
CGASCM
edium
0.2
0.1
0.05
0.01
0.00
50.
001
0.00
01
0.00
005
0.00
001
1 hA
B
Figure 8. Nuclear translocation of Nrf2 in HT29 cells. Cells were exposed to SWCNTs for (A) 1 h, cytosolic and nuclear fraction were separatedand both analysed by western blot (60 mg total protein per lane) using a rabbit polyclonal antibody against human Nrf2. Chlorogenic acid(CGA) (diluted DMSO 1%) was included in the assay as positive control. The data presented are expressed as the percentage of solvent control(the mean ± SD of three independent experiments). (B) Western blot of representative experiments. The detection of a-tubulin was used as aloading control.
Figure 9. Induction of DNA strand breaks after incubation of HT29 cells with SWCNT–sodium cholate (SC) suspension for (A) 3 h under alkalineconditions and for 24 h under (B) alkaline and (C) neutral conditions determined in the comet assay. Under alkaline electrophoresis conditions,formamidopyrimidine DNA-glycosylase (FPG) treatment for the detection of FPG-sensitive sites (dashed bars) and UV irradiation as a positivecontrol were included in the assay. Etoposide (solvent control: DMSO 1%) served as positive control in the comet assay under neutralelectrophoresis conditions. DNA damage is expressed as tail intensity (%). The data are presented as the mean ± SD of at least four independentexperiments each performed in duplicate. Significances indicated by asterisks refer to the significance level as compared to the respective control(SC, 0.02%: SWCNT; medium, 10% foetal calf serum (FCS): UV, 2 min) after treatment with FPG calculated by Student’s t test (*p < 0.05,**p < 0.01 and ***p < 0.001). Significances between FPG treated and FPG untreated samples are labelled with hashes (#p < 0.05).
J. Pelka et al.
Nan
otox
icol
ogy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
ity o
f M
elbo
urne
on
03/0
1/13
For
pers
onal
use
onl
y.
Modulation of the p53 phosphorylation levelTo confirm the DNA damage response to SWCNTs, p-p53was analysed in HT29 cells by western blot. In case of DNAdamage, the p53 protein is activated by phosphorylationcrucial for the activation of cell cycle checkpoints or trigger-ing cell cycle arrest to provide time for DNA damage repair.
Using the phospho-specific antibody to p53-Ser-15 toexamine p-p53, an increasing amount of p-p53 after 3 and24 h of SWCNT exposure was detected (Figure 11A–C).
After 3 h of exposure, the results clearly showed aninduction of p-p53 up to a concentration of 0.005 mg/ml.At a concentration of 0.005 mg/ml, the p-p53 rose to approx-imately 250% of the control (Figure 11A). With furtherincreasing SWCNT concentrations, the observed effectdeclined to about 110% at 0.2 mg/ml. HU, known for theinduction of p-p53 accumulation in several cell lines, wasused as a positive control. After HU exposure, phosphory-lation level of p53 in HT29 cells increased to about 450%(Figure 11A). An exposure of HT29 cells to SWCNTs for 24 hled to a slightly increased phosphorylation level of p53.Especially at a concentration of 0.005 mg/ml, p-p53 wasenhanced as compared to the 3 h incubation (303% of thecontrol) (Figure 11B).
Discussion
In view of a possible accumulation of CNTs along the foodchain and the potential use of this nanomaterial in foodpackaging or as transport vehicles for pharmaceuticalagents, the present study focused on the oral uptake ofCNTs. Little is known about adverse effects emanatingfrom SWCNTs taken up orally, since most studies on thetoxicological relevance of CNTs concentrate on inhalative ordermal exposure. The cell line HT29 is well characterised
and originates from the GIT and therefore appears as anappropriate model for studies tackling these issues.
SWCNTs investigated in the present study were synthe-sised by laser ablation and subsequently purified to mini-mise metal contamination. As CNTs are hydrophobic andthus exhibit strong van der Waals forces, SWCNTs weredispersed in the surfactant SC by ultrasonic treatment toprevent the formation of bundles in the aqueous systemof the cell culture medium. Furthermore, all incubationswere carried out in serum-containing medium (10% FCS)since Elder et al. (2007) showed that serum-free suspensionsof nanoparticles were associated with the occurrence of largeagglomerates, whereas less agglomeration was found inFCS-containing medium. Raman spectroscopy was used toconfirm the presence of SWCNTs and to investigate theirdispersion degree (Figure 1). The diameter and length of theSWCNTs in SC suspension were determined by AFM. Theresults indicated nanotube diameters ranging from 1.0 to3.2 nm (Figure 2B) and a length range from 200 to 2000 nm(Figure 2C). Most of the SWCNTs were found to have adiameter of 1.8 (± 0.3) nm and a length of 530 (± 200) nm.To exclude experimental artefacts from the suspensionmaterial, SC at a concentration of 0.02% was included asa control in all assays. At the applied concentration SC didnot substantially affect the tested biological end points.
A combination of assays was used to comprise differentend points of cytotoxicity including cellular protein as ameasure for cell growth, membrane integrity and mitochon-drial activity. CB, a widely-used reference material, was alsoincluded in the assays for suitable comparison of cytotoxiceffects of the SWCNT suspension. Incubation of HT29 cellswith SWCNTs for 24 h showed neither an effect on cellgrowth (SRB assay) (Figure 3) nor on cell membrane integrity(LDH assay) over the whole concentration range (Figure 5).
Med
ium SC
DMSO
0.00
005
0.00
010.
001
0.00
50.
010.
05 0.1
0.2
ETO0
1
2
3
4
5
6
7
*
Tai
l in
ten
sity
[%
]
mg/mL
Medium 10% FCS
SC 0.02% DMSO 1%
SWCNT
ETO 50 μM
24 h
C
Figure 9. (Continued ).
SWCNT toxicity in human colon cells
Nan
otox
icol
ogy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
ity o
f M
elbo
urne
on
03/0
1/13
For
pers
onal
use
onl
y.
Similar results were obtained in the trypan blue exclusionassay, performed in parallel to the comet and the GSHassays (data not shown). However, a decrease in mitochon-drial activity was observed (Figure 4). Compared to this,treatment with CB showed no influence on the investigatedbiological end points.
Yang et al. (2009) also reported an impact of SWCNTs onmitochondrial activity in PMEF cells (primary mouse embryofibroblasts) using the WST-1 assay. The activity of the mito-chondrial dehydrogenases decreased to about 74% of thecontrol at the highest concentration used, comparable to theactivity decline detected in the present study. However,beside a different cell line, SWCNT concentrations weremuch higher (5–100 mg/ml) and the investigated nanotubesdiffered considerably in diameter and length (Ø 8 nm, length<5 mm). Apparently, comparison between the existingresearch data concerning cytotoxicity of CNTs is limiteddue to the use of different CNT species, cell model systemsand procedure protocols with in part incomplete materialcharacterisation. Thus, although a spectrum of publicationsreported cytotoxic effects of CNTs (Cui et al. 2005; Jia et al.2005; Shvedova et al. 2003, 2005; Soto et al. 2005; Wick et al.2007), several studies did not detect substantial cytotoxicproperties (Bianco 2004; Chlopek et al. 2006; Flahaut et al.2006; Yang et al. 2009).
Furthermore, SWCNTs were found to strongly interactwith a large range of molecular species from dyes to poly-saccharides and other bio-molecules (Casey et al. 2005;Wörle-Knirsch et al. 2006; Yang et al. 2009). Casey et al.(2008) reported that SWCNTs might induce cytotoxicity in
A549 cells indirectly by alteration of the cell culture mediumwhich potentially results in false positive toxic effects.SWCNTs were found to adsorb nutrient constituents ofthe culture medium causing changes in cell viability. Inthe present study, such interference appears to be quiteunlikely as results of the trypan blue exclusion test andthe LDH assay showed no effects on cell viability ofHT29 cells after 24 h exposure to SWCNTs. Furthermore,potential interactions between organic indicator dyes andSWCNTs in the LDH or WST-1 assay were excluded by cell-free experiments (data not shown).
Manufactured SWCNTs usually contain significantamounts of metal catalysts primarily in the form of nano-particles like Ni, Co, Mo or Fe (up to 40 wt.%) (Morjan et al.2002), which are used in the production process for con-trolled CNT growth. These metal contents play an impor-tant role for the toxicological response to respectivepreparations (Pulskamp et al. 2007). SWCNTs used inthe present study were produced by the laser ablationmethod incorporating Ni0 and Co0 as metal catalysts andwere subsequently purified by gradient centrifugation for24 h. Analysis by ICP-MS indicates a residual catalystamount of 3.1 ± 0.2 wt.% Co, corresponding to 0.1 mM,and Ni <6 wt.% (max. 0.2 mM).
Pulskamp et al. (2007) investigated the SWCNT-mediatedmodulation of the intracellular ROS level in dependenceon the purity level of this nanomaterial. They found anenhanced generation of ROS only for the less purifiedSWCNTs with a residual metal content of 8 wt.% whereasthe purified SWCNTs (metal residue: 0.23 wt.%) showed no
0
5
10
15
20
25
30
35
40
SWCNT mg/mL
Kinetochore positive MNKinetochore negative MN
Mic
ron
ucl
eate
d V
79 c
ells
/100
0 ce
lls
0.2
0.1
0.05
0.00
001
0.00
1
0.00
010.
005
0.01
0.00
005
NQO
DMSOSC
Med
ium
24 h
Figure 10. Micronuclei in V79 cells after incubation with SWCNTs (negative control sodium cholate (SC) 0.02%) for 24 h. Nitroquinolineoxide(NQO) (0.3 mM, solvent control DMSO 1%) served as a control for the induction of chromosome fragments (kinetochore negativemicronuclei). Datarepresent mean of four independent experiments ± SD. Cell nuclei and micronuclei were visualised using UV excitation and microtubes andkinetochore signals were analysed under green and blue illumination. Dashed bars represent kinetochore negative micronuclei, and barskinetochore positive micronuclei. The occurrence of micronucleated cells is expressed as number of micronuclei cells per 1000 cells.
J. Pelka et al.
Nan
otox
icol
ogy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
ity o
f M
elbo
urne
on
03/0
1/13
For
pers
onal
use
onl
y.
Med
ium SC
0.00
001
0.00
005
0.00
010.
001
0.00
50.
010.
05 0.1
0.2
HU 0
100
200
300
400
500
600
700
mg/mL
p53
ph
osp
ho
ryla
tio
n le
vel,
T/C
[%
]
Medium 10% FCS
SC 0.02% SWCNT
HU 1.5 mM
3 h
Med
ium SC
0.00
001
0.00
005
0.00
010.
001
0.00
50.
010.
05 0.1
0.2
HU 0
100
p-p53
α-Tub
200
300
400
500
600
700
mg/mL
p53
ph
osp
ho
ryla
tio
n le
vel,
T/C
[%
]
Medium 10% FCS SC 0.02% SWCNT HU 1.5 mM
24 h
Med
ium
SC 0.00
001
0.00
005
0.00
01
0.00
10.
005
0.01
0.05
0.1
0.2
HU
A
B
C
Figure 11. Phosphorylation of p53 in HT29 cells. Cells were exposed to SWCNTs for (A) 3 h and (B) 24 h, lysed with RIPA buffer and the cell lysateswere analysed by western blot (40 mg total protein per lane) using a phosphor-specific antibody to p53-ser-15. Hydroxyurea (HU) (diluted in culturemedium) was included in the assay as positive control. The data presented are expressed as the percentage of solvent control (the mean ± SD of atleast five independent experiments). (C) Western blot of representative experiments. The detection of a-tubulin was used as a loading control.
SWCNT toxicity in human colon cells
Nan
otox
icol
ogy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
ity o
f M
elbo
urne
on
03/0
1/13
For
pers
onal
use
onl
y.
modulating activity. Furthermore, there are studies reportingthat ROS formation after treatment with nanomaterials likecolloidal fullerene nanoparticles (nC60) may be due to otherimpurities like solvents (e.g., tetrahydrofuran) or oxidativeby-products in nanomaterial preparations (Spohn et al. 2008;Xia et al. 2010).
In contrast, there are some studies documenting aninduction of ROS formation by carbon-containing nano-structures like CB regardless of the respective impuritylevel (Brown et al. 2004; Gilmour et al. 1996; Sayes et al.2005; Sun et al. 2004). Also, Yang et al. (2009) found astrong increase in ROS production after 24 h of exposure toSWCNTs in PMEF cells using the DCF assay in a concen-tration range of 10–100 mg/ml. In addition, a depletion ofthe cellular GSH level was reported. Of note, CNT dimen-sions and concentrations applied by Yang et al. comparedto the present study differ strongly, whereas the puritylevels of the SWCNT preparations were even higher(about 99.9% purity), although detailed information onthe residual metal content are not available (Yang et al2009).
Furthermore, the SWCNT preparation investigated in thisstudy affected the cellular redox status. Enhanced ROS for-mation was observed at concentrations ‡0.1 mg/ml (Figure 6),thus in the concentration range resulting in cytotoxicresponse (Figure 4). Likewise, a slight increase in intracellulartGSH level was observed at similar concentrations (Figure 7A)accompanied by a slight activation of the Nrf2 signallingpathway (Figure 8A,B). The translocation of Nrf2 is the crucialstep to a successful Nrf2 mediated antioxidative signalling. Anincreased abundance of Nrf2 in the nucleus leads down-stream to an onset of antioxidant phase II enzyme expression.Regarding these results, a SWCNT-mediated induction ofoxidative stress can be postulated.
Indeed, these effects decreased or disappeared completelywith prolonged incubation time. Thus, a slight, albeit notsignificant, decrease of the intracellular tGSH level (Figure7B) and no influence on the nuclear Nrf2 accumulation(data not shown) were observable. To which extent theresidual metal content contributes to the observed effectscannot be said with certainty, especially since existing dataare inconsistent.
It is of note that well below the cytotoxic concentrationsDNA damaging properties of SWCNTs were already detectedunder alkaline pH conditions with enhanced tail intensityalready at the lowest concentration of 0.00005 mg/ml(Figure 9B). These effects significantly increased withprolonged incubation time (Figure 9A,B). However, no addi-tional FPG-sensitive sites were detectable over the wholeconcentration range indicating that oxidative stress doesnot play a crucial role for the genotoxic properties ofthese SWCNTs. In contrast to the alkaline comet assay, noDNA double strand breaking properties were observed inthe comet assay under neutral pH conditions over thewhole concentration range (Figure 9C). These resultssuggest that DNA double strand breaks were not the pre-dominant genotoxic lesion induced by the applied SWCNTpreparation. Zhu et al. (2007) reported an increasedexpression of the double strand break repair protein
Rad 51 and the phosphorylation of gH2AX histone, amarker protein for the induction of double strand breakswithin 2 h of exposure to multiwalled CNTs. Furthercomprehensive studies are needed to associate structuralproperties of CNTs with the ability to induce DNA doublestrand breaks.
However, it has also to be considered that single-stranded DNA (ssDNA) is effectively used for dispersionof bundled SWCNTs. Zheng et al. (2003) provided evidencefor dispersion of SWCNTs in water by their sonication inthe presence of ssDNA as binding free energy of ssDNA toCNTs rivals that of two nanotubes for each other. Theysuggested by means of molecular modelling that ssDNAcan bind to CNTs through p-stacking, resulting in helicalwrapping to the surface. Such an interaction of SWCNTsand intact DNA during assay procedure might adulteratethe results of the alkaline comet assay. Thus, as additionalparameters for the occurrence of DNA damage the p-p53 aswell as MN formation were determined. In the presence ofDNA damage, the p53 protein is activated by phosphory-lation, triggering cell cycle arrest to provide time forDNA repair. Supporting the results of the alkaline cometassay, substantially enhanced p-p53 in HT29 cells wasobserved increasing with time by 3 and 24 h of incubation(Figure 11A,B).
In V79 cells, a standard cell line for the detection of MNformation, a slight but significant increase in kinetochore-negative MN was observed at subcytotoxic concentrationsfurther underlining the DNA damaging potential of theapplied SWCNT preparation (Figure 10). Comparable tothe findings in the comet assay, maximal effects were foundin the concentration range of 0.005–0.05 mg/ml. The lesspronounced effect in the MN test compared to the rate ofDNA strand breaks detected in the comet assay may bedue to the fact that theMN test detects only genotoxic lesionsthat persist without repair at least one mitotic cycle (vanGoethem et al. 1997; Kisin et al. 2007).
A contribution of the metal residues to the genotoxiceffects of SWCNTs seems unlikely for the following reasons:Ni and Co mediated biological effects are primarilybased on the production of ROS (Colognato et al. 2008;Oller et al. 1997). However, substantial DNA damag-ing properties already occurred well below the concentra-tions where a modulation of the intracellular ROS levelwas observed. In addition, no FPG sensitive sites weredetectable over the whole concentration range. Further-more, Ni/Co mediated genotoxic effects documented inthe literature occurred at much higher concentrations thandetermined for the SWCNTs of the present study.Colognato et al. (2008) detected Co-NP-induced DNAbreaks and MN at concentrations exceeding 40 mM. Inthe case of nickel nanoparticles, very few studies existexamining their biological activity. An exception are thein vivo studies of Zhang et al. (1998, 2003) and Ispas et al.(2009), which reported biological effects of Ni-NP atconcentrations exceeding 800 mM. Thus, regarding theresidual metal content of the SWCNTs investigated inthis study (about 0.1 mM), a metal mediated mechanismof action can most likely be excluded.
J. Pelka et al.
Nan
otox
icol
ogy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
ity o
f M
elbo
urne
on
03/0
1/13
For
pers
onal
use
onl
y.
Conclusion
In summary, the applied SWCNT–SC suspension was foundto mediate slight cytotoxic effects in HT29 cells. At cytotoxicconcentrations the formation of ROS and a slight increase ofthe cellular tGSH and the nuclear Nrf2 levels were observed.However, substantial DNA damaging effects were alreadyobserved at subcytotoxic concentrations in the comet assayunder alkaline condition, which were not associated withenhanced formation of FGP-sensitive sites suggesting thatthe DNA damaging effect is not primarily based on theonset of oxidative stress. Furthermore, exposure to SWCNTsenhanced the formation of kinetochore-negative MN inV79 cells as well as the phosphorylation of the tumoursuppressor protein p53 in HT29 cells, underlining the geno-toxic potential of these nanostructures.
These results suggest that careful scrutiny of the adversehealth effects of CNTs is needed in view of the rapiddevelopment of novel fields of applications, an increasedproduction and thus increased exposure of human beingsand a possible accumulation along the food chain.
Acknowledgements
This work was partly funded by the DFG (Center for Func-tional Nanostructures (CFN), E1.1 and C3.2). The authorsalso gratefully acknowledge support by the HelmholtzAssociation under POF NanoMikro and Anna Bytzek andNorbert Kandler (University of Vienna, Institute of InorganicChemistry) for their help in collecting the ICP-MS data.
Declaration of interest
The authors report no conflicts of interest. The authors aloneare responsible for the content and writing of the paper.
ReferencesAllen BL, Kichambare PD, Gou P, Vlasova II, Kapralov AA,
Konduru N, et al. 2008. Biodegradation of single-walled carbonnanotubes through enzymatic catalysis. Nano Lett 8:3899–3903.
Belyanskaya L, Weigel S, Hirsch C, Tobler U, Krug HF Wick P. 2009.Effects of carbon nanotubes on primary neurons and glial cells.Neurotoxicology 30:702–711.
Bianco A. 2004. Carbon nanotubes for the delivery of therapeuticmolecules. Expert Opin Drug Deliv 1:57–65.
Brown DM, Donaldson K, Borm PJ, Schins RP, Dehnhardt M,Gilmour P, et al. 2004. Calcium and ROS-mediated activation oftranscription factors and TNF-alpha cytokine gene expression inmacrophages exposed to ultrafine particles. Am J Physiol Lung CellMol Physiol 286:L344–L353.
Casey A, Farrell GF, McNamara M, Byrne HJ, Chambers G. 2005.Interaction of carbon nanotubes with sugar complexes. SyntheticMetals 153:357–360.
Casey A, Herzog E, Lyng FM, Byrne HJ, Chambers G, Davoren M. 2008.Single walled carbon nanotubes induce indirect cytotoxicity bymedium depletion in A549 lungs cells. Toxicol Lett 179:78–84.
Chaudhry Q, Scotter M, Blackburn J, Ross B, Boxall A, Castle L, et al.2008. Applications and implications of nanotechnologies for thefood sector. Food Addit Contam 25:241–258.
Chlopek J, Czajkowska B, Szaraniec B, Frackowiak E, Szostak K,Béguin F. 2006. In vitro studies of carbon nanotubes biocompati-bility. Carbon 44:1106–1111.
Colognato R, Bonelli A, Ponti J, Farina M, Bergamaschi E, Sabbioni E,Migliore L. 2008. Comparative genotoxicity of cobalt nanoparticlesand ions on human peripheral leucocytes in vitro. Mutagen23:377–382.
Cui D, Tian F, Ozkan CS, Wang M, Gao H. 2005. Effect of single wallcarbon nanotubes on human HEK293 cells. Toxicol Lett 155:73–85.
Donaldson K, Aitken R, Tran L, Stone V, Duffin R, Forrest G,Alexander A. 2006. Carbon nanotubes: a review of their propertiesin relation to pulmonary toxicology and workplace safety. Toxicol Sci92:5–22.
Donaldson K, Tran L, Jimenez LA, Duffin R, Newby DE, Mills N, et al.2005. Combustion-derived nanoparticles: a review of their toxicol-ogy following inhalation exposure. Part Fibre Toxico 2:10.
Elder A, Yang H, Gwiazda R, Teng X, Thurston S, He H,Oberdörster G. 2007. Testing nanomaterials of unknown toxicity:An example based on platinum nanoparticles of different shapes.Adv Mater 19:3124–3129.
Fiorito S, Serafino A, Andreola F, Bernier P. 2006. Effects of fullerenesand single-wall carbon nanotubes on murine and human macro-phages. Carbon 44:1100–1105.
Flahaut E, Durrieu MC, Remy-Zolghadri M, Bareille R, Baquey C. 2006.Study of the cytotoxicity of CCVD carbon nanotubes. J Mater Sci41:2411–2416.
Gilmour PS, Brown DM, Lindsay TG, Beswick PH, MacNee W,Donaldson K. 1996. Adverse health effects of PM10 particles:involvement of iron in generation of hydroxyl radical. Occup Envi-ron Med 53:817–822.
Gottschalk F, Sonderer T, Scholz RW, Nowack B. 2009. Modelledenvironmental concentrations of engineered nanomaterials (TiO2,ZnO, Ag, CNT, Fullerenes) for different regions. Environ Sci Technol43:9216–9222.
Guo T, Nikolaev P, Thess A, Colbert DT, Smalley RE. 1995. Catalyticgrowth of single walled nanotubes by laser vaporization. Chem PhysLett 243:49–54.
Helland A, Wick P, Koehler A, Schmid K, Som C. 2007. Reviewing theEnvironmental and Human Health Knowledge Base of CarbonNanotubes. Environ Health Perspect 115:1125–1131.
Hennrich F, Krupke R, Arnold K, Rojas Stuetz JA, Lebedkin S,Koch T, et al. 2007. The Mechanism of Cavitation-InducedScission of Single-Walled Carbon Nanotubes. J Phys Chem B111:1932–1937.
Hennrich F, Wellmann R, Malik S, Lebedkin S, Kappes MM. 2003.Reversible modification of the absorption properties of single-walledcarbon nanotube thin films via nitric acid exposure. Phys ChemChem Phys 5:178–183.
Holt BD, Dahl KN, Islam MF. 2011. Quantification of uptake andlocalization of bovine serum albumin-stabilized single-wall carbonnanotubes in different human cell types. Small [ePub May 31, 2011]DOI: 10.1002/smll.201100437.
Ispas C, Andreescu D, Patel A, Goia DV, Andreescu S,Wallace KN. 2009. Toxicity and developmental defects of differentsizes ans shape nickel nanoparticles in zebrafish. Environ SciTechnol 43:6349–6356.
Jorio A, Saito R, Hafner JH, Lieber CM, Hunter M, McClure T, et al.2001. Structural (n, m) determination of isolated single-wallcarbon nanotubes by resonant Ramanscattering. Phys Rev Lett 86(6):1118–1121.
Jia G, Wang H, Yan L, Wang X, Pei R, Yan T, et al. 2005. Cytotoxicity ofcarbon nanomaterials: single-wall nanotube, multi-wall nanotube,and fullerene. Environ Sci Technol 39:1378–1383.
Kagan VE, Konduru NV, Feng W, Allen BL, Conroy J, Volkov Y, et al.2010. Carbon nanotubes degraded by neutrophil myeloperoxidaseinduce less pulmonary inflammation. Nat Nanotechnol 5:354–359.
Kagan VE, Tyurina YY, Tyurin VA, Konduru NV, Potapovich AI,Osipov AN, et al. 2006. Direct and indirect effects of single walledcarbon nanotubes on RAW 264.7 macrophages: role of iron. ToxicolLett 165:88–100.
Kam NWS, O’Connell M, Wisdom JA, Dai H. 2005b. Carbon nanotubesas multifunctional biological transporters and near-infrared agentsfor selective cancer cell destruction. Proc Natl Acad Sci USA102:11600–11605.
Kisin ER, Murray AR, Keane MJ, Shi XC, Schwegler-Berry D,Gorelik O, et al. 2007. Single-walled carbon nanotubes: geno-and cytotoxic effects in lung fibroblast V79 cells. J Toxicol EnvironHealth A 70:2071 – 2079.
Kuhlbusch TAJ, Krug HF, Nau K. 2009. NanoCare Health relatedAspects of Nanomaterials. Final Scientific Report. DECHEMA e.V.ISBN: 978-3-89746-108-6.
Lam C-W, James JT, McCluskey R, Arepalli S, Hunter RL. 2006. A reviewof carbon nanotube toxicity and assessment of potential occupa-tional and environmental health risks. Crit Rev Toxicol 36:189–217.
SWCNT toxicity in human colon cells
Nan
otox
icol
ogy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
ity o
f M
elbo
urne
on
03/0
1/13
For
pers
onal
use
onl
y.
Lam C-W, James JT, McCluskey R, Hunter RL. 2004. Pulmonary toxicityof single-wall carbon nanotubes in mice 7 and 90 days after intra-tracheal instillation. Toxicol Sci 77:126–134.
Lebedkin S, Arnold K, Hennrich F, Krupke R, Renker B,Kappes MM. 2003. FTIR-luminescence mapping of dispersedsingle-walled carbon nanotubes. New J Phys 5:140.1–140.11.
Lebedkin S, Schweiss P, Renker B, Malik S, Hennrich F, Neumaier M,et al. 2002. Single-wall carbon nanotubes with diameters approach-ing 6 nm obtained by laser vaporization. Carbon 40:417–423.
Maynard AD, Baron PA, Foley M, Shvedova AA, Kisin ER,Castranova V. 2004. Exposure to carbon nanotube material: aerosolrelease during the handling of unrefined single-walled carbonnanotube material. J Toxicol Environ Health A 67:87–108.
Monteiro-Riviere NA, Nemanich RJ, Inman AO, Wang YY,Riviere JE. 2005. Multi-walled carbon nanotube interactions withhuman epidermal keratinocytes. Toxicol Lett 155:377–384.
Morjan RE, Nerushev OA, Sveningsson M, Falk LKL, Rohmund F,Campbell EEB. 2002. Parametric study of nanotube growth fromC2H2 and C60 on supported iron catalyst particles. Am Inst PhysicsConf Proc 633:186.
Moshammer K, Hennrich F, Kappes MM. 2009. Selective suspension inaqueous sodium dodecyl sulfate according to electronic structuretype allows simple separation of metallic from semiconductingsingle-walled carbon nanotubes. Nano Res 2:599–606.
Muller J, Huaux F, Moreau N, Misson P, Heilier J-F, Delos M, et al.2005. Respiratory toxicity of multi-wall carbon nanotubes. ToxicolAppl Pharmacol 207:221–231.
Oberdörster E, Ortiz-Acevedo A, Xie H, Pantano P, Baughman RH,Musselman IH, et al. 2005. Exposure of fathead minnow to fullereneand single-walled carbon nanotubes [Abstract]. Off J Soc Toxicol 84(Suppl 1):325.
Oberdörster E, Zhu S, Blickley TM, McClellan-Green P, Haasch ML.2006. Ecotoxicology of carbon-based engineered nanoparticles:effects of fullerene (C60) on aquatic organisms. Carbon 44:1112–1120.
Oller AR, Costa M, Oberdörster G. 1997. Carcinogenicity assessment ofselected nickel compounds. Toxicol Appl Pharm 143(1):152–166.
Porter AE, Gass M,Muller K, Skepper JN, Midgley PA, WellandM. 2007.Direct imaging of single-walled carbon nanotubes in cells. NatNanotechnol 2:713–717.
Pulskamp K, Diabaté S, Krug HF. 2007. Carbon nanotubes show nosign of acute toxicity but induce intracellular reactive oxygen speciesin dependence on contaminants. Toxicol Lett 168:58–74.
Sayes CM, Gobin AM, Ausman KD, Mendez J, West JL, Colvin VL. 2005.Nano-C60 cytotoxicity is due to lipid peroxidation. Biomaterials26:7587–7595.
Shvedova AA, Castranova V, Kisin ER, Schwegler-Berry D, Murray AR,Gandelsman V, et al. 2003. Exposure to carbon nanotube material:assessment of nanotube cytotoxicity using human keratinocyte cells.J Toxicol Environ Health A 24:1909–1926.
Shvedova AA, Kisin ER, Mercer R, Murray AR, Johnson VJ,Potapovich AI, et al. 2005. Unusual inflammatory and fibrogenicpulmonary responses to single-walled carbon nanotubes in mice.Am J Physiol Lung C 289:L698–L708.
Skehan P, Storeng R, Scudiero D, Monks A, McMahon J, Vistica D, et al.1990. New colorimetric assay for anticancer-drug screening. J NatlCancer Inst 82:1107–1112.
Soto KF, Carrasco A, Powell TG, Garza KM, Murr LE. 2005. Compar-ative in vitro cytotoxicity assessment of some manufactured nano-particulate materials characterized by transmission electronmicroscopy. J Nanopart Res 7:145–169.
Sozer N, Kokini JL, 2009. Nanotechnology and its applications in thefood sector. Trends Biotechnol 27(2):82–89.
Spohn P, Hirsch C, Hasler F, Bruinink A, Krug HF, Wick P. 2008.C60 fullerene: A powerful antioxidant or a damaging agent? Theimportance of an in-depth material characterization prior to toxicityassays. Environ Pollut 157:1134–1139.
Sun T, Jia Z, Xu Z. 2004. Different hydroxyl radical scavenging activityof water-soluble [beta]-alanine C60 adducts. Bioorg Med Chem Lett14:1779–1781.
Thurnherr T, Su DS, Diener L, Weinberg G, Manser P, Pfaender N, et al.2009. Comprehensive evaluation of in vitro toxicity of threelarge-scale produced carbon nanotubes on human JurkatT cells and a comparison to crocidolite asbestos. Nanotoxicology3:319–338.
Tice RR, Agurell E, Anderson D, Burlinson B, Hartmann A,Kobayashi H, et al. 2000. Single cell gel/comet assay: guidelinesfor in vitro and in vivo genetic toxicology testing. Environ MolMutagen 35:206–221.
Tietze F. 1969. Enzymic method for quantitative determina-tion of nanogram amounts of total an oxidized glutathion:Application to mammalian blood and other tissues. Anal Biochem27:502–522.
Van Goethem F, Lison D, Kirsch-Volders M. 1997. Comparative eval-uation of the in vitro micronucleus test and the alkaline single cellgel electrophoresis assay for the detection of DNA damaging agents:genotoxic effects of cobalt powder, tungsten carbide and cobalt-tungsten carbide. Mutat Res 392:31–43.
Wenseleers W, Vlasov II, Goovaerts E, Obraztsova ED, Lobach AS,Bouwen A. 2004. Efficient isolation and solubilization of pristinesingle-walled nanotubes in bile salt micelles.Advanced FunctionalMaterials 14(11):1105–1112.
Wick P, Manser P, Limbach LK, Dettlaff-Weglikowska U, Krumeich F,Roth S, et al. 2007. The degree and kind of agglomeration affectcarbon nanotube cytotoxicity. Toxicol Lett 168:121–131.
Wörle-Knirsch JM, Pulskamp K, Krug HF. 2006. Oops they did it again!Carbon nanotubes hoax scientists in viability assays. Nano Lett6:1261–1268.
Wu Y, Hudson J, Lu Q, Moore J, Mount A, Rao A, et al. 2006. Coatingsingle-walled carbon nanotubes with phospholipids. J Phys Chem B110:2475–2478.
Xia XR, Monteiro-Riviere NA, Riviere JE. 2010. Intrinsic biologicalproperty of colloidal fullerene nanoparticles (nC60): Lack of lethalityafter high dose exposure to human epidermal and bacterial cells.Toxicol Lett 197:128–134.
Yang H, Liu C, Yang D, Zhang H, Xi Z. 2009. Comparative study ofcytotoxicity, oxidative stress and genotoxicity induced by four typicalnanomaterials: the role of particle size, shape and composition.J Appl Toxicol 29:69–78.
Zhang Q, Kusaka Y, Sato K, Mo Y, Fukada M, Donaldosn K. 1998.Toxicity of ultrafine nickel particles in lungs after intratrachealinstillation. J Occup Health 40:171–176.
Zhang Q, Kusaka Y, Zhu X, Sato K, Mo Y, Kluz T,Donaldosn K. 2003. Comparative toxicity of standard nickel andultrafine nickel in lungs after intratracheal instillation. J OccupHealth 45:23–30.
Zheng M, Jagota A, Semke ED, Diner BA, McLean RS, Lustig SR, et al.2003. DNA-assisted dispersion and separation of carbon nanotubes.Nat Mater 2:338–342.
Zhu L, Chang DW, Dai L, Hong Y. 2007. DNA damage induced bymultiwalled carbon nanotubes in mouse embryonic stem cells.Nano Lett 7:3592–3297.