Abstract Dystonia is a movement disorder in whichinvoluntary or intermittent muscle contractions causetwisting and repetitive movements, abnormal postures,or both. Excessive co-contraction and abnormalities inthe time course of reciprocal inhibition between antag-onist groups of muscles are considered to be cardinalfeatures of some types of dystonia and reduced speedof movement is often attributed to involuntary activa-tion of antagonist muscles about a joint. In the presentstudy we describe muscle activity during unconstrainedmulti-joint reaching movements. Children diagnosedwith arm dystonia due to cerebral palsy (CP) or pri-mary dystonia (n = 7, 4–16 years, 4 with CP, 3 primary)and similar age healthy subjects pointed alternately totwo targets as fast as possible. The children with dysto-nia showed decreased speed, greater variability, andpauses at targets compared with controls. Decreasedspeed was mostly due to diYculty in reversing reachingdirection, and increased variability was associated withlarge Xuctuations in the duration of the pauses at tar-gets, rather than with variations in the Xexion/exten-sion velocity proWles. Surface electromyographic(EMG) activities were examined to assess if the abnor-malities observed in the children with dystonia couldbe explained in terms of increased levels of co-contrac-tion. Unexpectedly, we found that the children withdystonia showed lower levels of co-contraction thanthe controls during movement, and the pauses at tar-
gets were associated with reduced levels of activationrather than with excessive activity in antagonist groupsof muscles. Therefore reduced speed of movementduring unconstrained reaching may not be due toinvoluntary activation of the antagonist muscle, andco-contraction of opposing muscles about a joint is notan obligatory feature of multi-joint movement in chil-dren with dystonia.
Dystonia in children has been deWned as “a movementdisorder in which involuntary sustained or intermit-tent muscle contractions cause twisting and repetitivemovements, abnormal postures, or both” (Fahn et al.1987; Sanger et al. 2003). Dystonia is considered to be“primary” when it is the dominant manifestation of adeWned or presumed genetic disorder, otherwise it isconsidered to be “secondary”. Dystonia is thought tobe associated with three physiological phenomena; co-contraction of antagonist muscles, overXow of electro-myographic (EMG) activity onto uninvolved musclesduring voluntary movement, and involuntary activa-tion of muscles during passive shortening (Marsden1984). The hypothesis that in dystonia abnormalmotor patterns arise as a consequence of dysfunctionin the basal ganglia (Berardelli et al. 1998) hasreceived empirical as well as theoretical support (seeSanger 2003, for review). According to one model ofbasal ganglia function, dystonia arises from a netdecrease in the Wring of inhibitory neurons projecting
N. Malfait · T. D. Sanger (&)Department of Neurology and Neurological Sciences, Stanford University, 300 Pasteur Dr, A347, Stanford, CA 94305-5235, USAe-mail: [email protected]
123
Exp Brain Res (2007) 176:206–216 207
from the internal segment of the globus pallidus (GPi)to the ventral lateral thalamus, thus increasing theactivity of excitatory thalamocortical projections tomotor and premotor regions of the frontal cortex(Vitek 2002). Consequently, dystonia is sometimesconsidered to result from excessive motor cortical out-Xow with involuntary activation of muscles not nor-mally involved in the desired movement or posture,and it is thus assumed that the result of the basalganglia abnormality is an increase in activation ofmuscles.
Dystonia is an abnormality of both movement andposture (Marsden 1984; Sanger et al. 2003). The eVectof dystonia on active movement has been diYcult tocharacterize. While Wxed dystonic postures are fre-quently observed and movements are slow, inaccurate,variable, and awkward (Sanger et al. 2005), the relationbetween dystonic postures that are seen at rest andabnormalities of movement has not been investigated.It is tempting to hypothesize that dystonic posturesintrude during movement and are reXected by involun-tary and inappropriate muscle activation. Co-contrac-tion of opposing muscles and overXow onto musclesthat antagonize movement may be a cause of theabnormalities in trajectory and the reduced speed ofmovement. However, these hypotheses have not beentested.
Most studies of the kinematics of children withmotor disorders have focused on the lower extremitiesin order to understand and improve ambulatory func-tion (Damiano and Abel 1996; O’Byrne et al. 1998;Thelen et al. 2003). With few exceptions (McPhersonet al. 1991; Fetters and Kluzik 1996; Utley and Sugden1998; Chang et al. 2005; van der Heide et al. 2005a)studies of upper extremity function have mostly beenlimited to constrained single-joint movement (Harris1991; Duque et al. 2003). Therefore, impairment in theupper limbs has not yet been well described. In thepresent study, we examined abnormalities in elbowjoint kinematics observed during unconstrained multi-joint reaching movements in children with eitherprimary or secondary dystonia. In agreement with pre-vious clinical and experimental observations, weexpected to observe decreased speed of movement andincreased variability in the joint kinematics (Sangeret al. 2005; Van Der Heide et al. 2005b). Our goal wasto describe the nature of these two characteristic fea-tures in relation to the EMG activity recorded in theantagonist biceps/triceps muscle groups. In particular,we wanted to determine if movement slowness couldbe attributed to abnormal patterns of co-contractionand involuntary activation of muscles opposing move-ment.
Methods
Subjects
Seven children aged 4–16 years (mean 11.1) diagnosedwith dystonia by an experienced child neurologist(TDS) were recruited from the Stanford PediatricMovement Disorders Clinic. (an eighth child was alsorecruited but was not able to perform a suYcient num-ber of trials to be included in the analyses.) Subjectswith dystonia were included only if they exhibitedabnormal dystonic postures of the arm during forwardreaching movements. Three subjects (2, 5, and 6) wereclassiWed as “primary” dystonia due to lack of an iden-tiWed cause, normal early development with subse-quent appearance of dystonia, and normal MRIWndings (all were negative on testing for the DYT1gene). The remaining four subjects (1, 3, 4, and 7) wereclassiWed as “secondary” and carry a diagnosis of cere-bral palsy (CP) due to known prenatal or peri-natalinjury (see Table 1). None of the subjects showed evi-dence of spasticity (a spastic catch or elevated tendonreXexes) in the elbow Xexors or extensors. Seven con-trol subjects aged 5–16 years (mean 10.6) withoutmotor disorders were recruited from a conveniencesample of children. All healthy children were righthand dominant by parent report. Informed consentwas obtained from parents consistent with a protocolapproved by the Stanford University InstitutionalReview Board. Authorization for analysis, storage, andpublication of protected health information wasobtained from parents according to the Health Infor-mation Portability and Accountability Act (HIPAA).All subjects were rated at the time of testing on theGross Motor Function ClassiWcation System (GMFCS)(Palisano et al. 1997), the upper extremity subscale ofthe Burke–Fahn–Marsden dystonia rating scale (BFM)(Burke et al. 1985), the Barry–Albright dystonia ratingscale (BAD) (Barry et al. 1999), and the UniWed Dys-tonia Rating Scale (UDRS) (Comella et al. 2003). Atthe time of testing, subject 4 was taking trihexyphen-idyl and baclofen. The remaining subjects were on nomedications. This information is summarized inTable 1.
Experimental task
Subjects were seated comfortably and unrestrained in anon-metallic chair. Subjects were asked to perform atask directly derived from the “Wnger-to-nose” reach-ing task commonly used in routine neurological evalua-tion; speciWcally subjects were required to pointalternately to a target located at arm’s length in front
123
208 Exp Brain Res (2007) 176:206–216
of them in the sagittal plane and their nose. Location ofthe target was adjusted for each subject such that it wassituated at the same height as the nose of the subjectand at full arms-length distance. Children were askedto perform the task “as fast as possible without miss-ing”. Each child performed two series of 20 completeXexion/extension cycles with each arm. Most of thechildren with dystonia had diYculty maintaining con-tinuous movement for 20 trials in a row. For the analy-ses, we selected series of at least Wve cycles ofmovements that were executed without interruption(interruption was deWned as any pause that lasted morethan 3 s). This criterion resulted in a rejection of 28%of the movement cycles for the subjects with dystoniaand less than 1% for the controls. For several subjectswith dystonia the proportion of rejected cycles wasclose to 50%. Therefore in order to include a compara-ble number of movements for both groups of subjects,for each subject we analyzed only the Wrst 20 move-ment cycles that satisWed the criterion (for one childwith dystonia (subject 1) only 18 trials could beincluded). The preferred arm (less impaired, for thechildren with dystonia and dominant for the controls)was always tested Wrst. Subject 4 was able to completethe task only on the left arm due to severe dystonia onthe right side. Series of trials were self-initiated startingat the distal target.
Signal recording
Kinematic data were recorded using magnetic positionsensors (Polhemus Inc.) attached using either Velcro™straps or medical-grade adhesive to eight points on thebody: the mid-shaft of each upper arm, the dorsum ofeach distal forearm between the radius and ulna, thedorsum of each hand over the mid-shaft of the third
metacarpal bone, the back over the Wrst or second tho-racic vertebra, and the forehead 1–3 cm above the nosein the midline. The location of joint axes relative toeach sensor was measured using one of the sensors as amarker, in accordance with the “digitizing” procedureof commercially available kinematics analysis software(Skill Technologies, Inc.).
EMG activity of four groups of muscles in each armwas recorded using surface electrodes (DE2.3, DelsysInc.). Each active electrode has two parallel bar con-tacts of length 1 cm and width 1 mm, with inter-elec-trode spacing of 1 cm. Internal electronics in theelectrodes provide 1000 § 2% ampliWcation and ana-log Wltering with half-height cutoV between 20 § 5 and450 § 50 Hz and roll-oV of 12 dB per octave. Common-mode rejection ratio (CMRR) is greater than 80 dB at60 Hz. Electrodes were placed over the belly of thebiceps brachii and triceps brachii. Electrodes were alsoplaced over the Xexor carpi radialis and extensor carpiradialis longus, but data from the forearm electrodeswas not used in this experiment since the forearm elec-trodes were not placed over muscles that contribute toelbow motion. EMG signals were sampled at 1,000 Hzand digitized with 16-bit precision (CED Power 1401;Cambridge Electronic Design, Cambridge, UK).
Data analysis
Kinematic data were recorded from each position sen-sor, sampled at 120 Hz, Wltered with a digital low-passWlter (6 dB cutoV at 20 Hz), and numerically diVerenti-ated. In the present paper, analyses are restricted toelbow kinematics. For each movement cycle, onset andoVset of Xexion and extension were deWned using thevelocity proWle zero-crossings. Each Xexion orextension movement was then further divided into
Table 1 Clinical characteristics and testing results for all subjects
ID Age Sex BFM UDRS BAD GMFCS Diagnosis Symptoms Pause duration mean § SD (s)
Co-contraction ‘MIN’ mean § SDR L R L R L
1 4 F 2 2 2 2 1 1 2 Prematurity with perinatal intraventricular hemorrhage
Bilateral arm dystonia and leg spasticity
0.207 § 0.177 0.055 § 0.057
2 8 F 2 0 1.5 0 2 0 2 Primary hemidystonia Right foot and hand dystonia
0.163 § 0.273 0.077 § 0.038
3 9 M 4 9 6 10 2 3 5 Prematurity with perinatal intraventricular hemorrhage
Left > right arm dystonia and leg spasticity
0.434 § 0.324 0.045 § 0.035
4 13 F 9 9 13 12 4 3 5 Perinatal hypoxic injury Generalized dystonia and chorcoathetosis
1.0474 § 0.753 0.061 § 0.054
5 13 M 4 2 1.5 1.5 1 1 2 Primary dystonia Bilateral arm dystonia and writer’s cramp
acceleration and deceleration phases according to thezero-crossings of the acceleration. For the subjects withdystonia, two additional phases were deWned that cor-responded to pauses in the velocity and accelerationproWles that these subjects exhibited at both distal andproximal targets before initiating movement directionreversal. Such pauses were not seen in the control sub-jects. Therefore there were six phases for children withdystonia: acceleration during extension, decelerationduring extension, pause at the distal target, accelera-tion during Xexion, deceleration during Xexion, andpause at the proximal target. Control subjects did notexhibit pauses, and therefore had only four phases. Foreach Xexion/extension cycle, we computed the durationof the complete cycle and the duration of each move-ment phase, as well as the maximum joint angularvelocity and maximum and minimum joint angles. Toassess kinematic variability across trials for each sub-ject, we also calculated the coeYcient of variation (CV)of each measure: the standard deviation over the trialsdivided by the mean. The use of CV allows comparisonof measures of variability between diVerent perfor-mance conditions.
In accordance with standard practice for analysis ofEMG signals (for example Hogan and Mann 1980;Clancy and Hogan 1994; Miscellaneous 1996), any con-stant DC oVset was removed, and then the EMG sig-nals were full-wave rectiWed and low-pass Wltered at20 Hz to extract the amplitude envelope. The resultingamplitude envelope reXects the temporal changes inpower over the entire sampled spectrum. The choice oflow pass Wlter at 20 Hz was determined by the low-passcutoV of the kinematic measurements in order to allowcomparison. Because the EMG activity levels observedduring unconstrained reaching may be very diVerentfrom those observed during isometric maximum volun-tary contraction (MVC), rectiWed and Wltered EMG foreach muscle was normalized to the muscle’s maximumrectiWed and Wltered EMG amplitude observed duringthe entire experiment, rather than to MVC.
To compare average levels of co-contraction duringeach of the six phases of movement, we used two diVer-ent methods of time binning. The Wrst method usedWxed 50 ms time bins. In this case, the Wnal bin of eachmovement phase could be less than 50 ms long. Thesecond method divided each movement phase intoWve-bins of equal length. This resulted in variable-length bins between 33 and 108 ms for the childrenwith dystonia, and between 25 and 67 ms for the con-trols. Similar results were obtained with both methods,and in the results below we report data for variable-length bins. The rectiWed, low-pass Wltered, and nor-malized EMG was averaged over each time bin.
There is no generally-accepted quantitative measureof co-contraction. We performed our analysis usingtwo diVerent methods, each of which produces a scalarvalue for each time bin. The Wrst method (“DOT”) cal-culates the dot product of the (Wltered, rectiWed, nor-malized, and averaged) EMG between the biceps andtriceps in each time bin, which is equivalent to comput-ing the zero-time-lag covariance. Since rectiWed EMGis always positive, this measure will be zero if and onlyif at most one of the two muscles is active at the sametime. The second method (“MIN”) calculates the mini-mum value of the (Wltered, rectiWed, normalized, andaveraged) EMG between the agonist and antagonistmuscles in each time bin. This measure will also bezero if and only if one of the two muscles is alwayszero. The principal diVerence between the two meth-ods is that in DOT, the value of co-contractiondepends on both the agonist and antagonist activation,while for MIN the value depends only on the antago-nist. The DOT method represents a model of co-con-traction as an abnormal correlation between opposingmuscles, but it suVers from the fact that increasing ago-nist activity will increase the measure of co-contractioneven if the antagonist remains unchanged. The MINmethod represents a model of co-contraction as anyactivity in the normally-silent antagonist muscle. (Notethat although MIN is related to the amount of agonisttorque that is countered by the antagonist, we do notattempt to estimate the resulting torques since thereare not yet widely accepted methods for determiningthe relation between surface EMG and torque duringmovement.) Since EMG is normalized by the maxi-mum level observed during the testing, both methodstake into account possible diVerences in electrodeplacement or muscle morphology.
There are other possible methods of quantifying co-contraction (Damiano et al. 2000). We chose not to usethe ratio of antagonist to agonist activity since the mag-nitude of co-contraction could become very large if theagonist activity is low. We chose not to use the ratio ofthe lesser to the greater of the opposing muscles (whichis equivalent to the MIN value normalized to theopposing muscle’s activity) since this may underesti-mate co-contraction when agonist activity is large.Recently, a new co-contraction index has been pro-posed that (in the elbow) is related to the ratio of thediVerence in muscle activities to the sum of their activi-ties (Yao et al. 2004, 2006). However, this index is notsuitable for our purposes, since it can produce valuesindicating maximal co-activation even when total acti-vation is very low, and it is undeWned when both mus-cles are relaxed as we frequently observed duringmovement.
123
210 Exp Brain Res (2007) 176:206–216
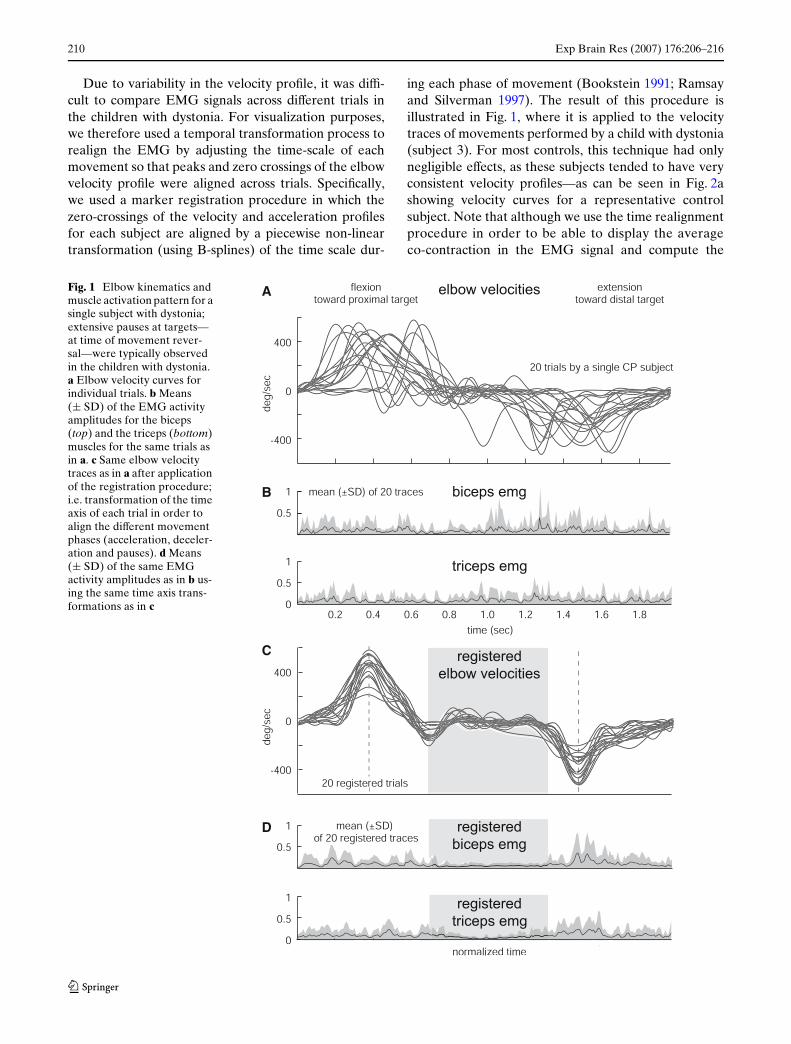
Due to variability in the velocity proWle, it was diY-cult to compare EMG signals across diVerent trials inthe children with dystonia. For visualization purposes,we therefore used a temporal transformation process torealign the EMG by adjusting the time-scale of eachmovement so that peaks and zero crossings of the elbowvelocity proWle were aligned across trials. SpeciWcally,we used a marker registration procedure in which thezero-crossings of the velocity and acceleration proWlesfor each subject are aligned by a piecewise non-lineartransformation (using B-splines) of the time scale dur-
ing each phase of movement (Bookstein 1991; Ramsayand Silverman 1997). The result of this procedure isillustrated in Fig. 1, where it is applied to the velocitytraces of movements performed by a child with dystonia(subject 3). For most controls, this technique had onlynegligible eVects, as these subjects tended to have veryconsistent velocity proWles—as can be seen in Fig. 2ashowing velocity curves for a representative controlsubject. Note that although we use the time realignmentprocedure in order to be able to display the averageco-contraction in the EMG signal and compute the
Fig. 1 Elbow kinematics and muscle activation pattern for a single subject with dystonia; extensive pauses at targets—at time of movement rever-sal—were typically observed in the children with dystonia. a Elbow velocity curves for individual trials. b Means (§ SD) of the EMG activity amplitudes for the biceps (top) and the triceps (bottom) muscles for the same trials as in a. c Same elbow velocity traces as in a after application of the registration procedure; i.e. transformation of the time axis of each trial in order to align the diVerent movement phases (acceleration, deceler-ation and pauses). d Means (§ SD) of the same EMG activity amplitudes as in b us-ing the same time axis trans-formations as in c
123
Exp Brain Res (2007) 176:206–216 211
variable bin widths for EMG analysis, analysis of thevelocity proWle for variability, speed, and pauses wasdone without modiWcation of the time scale.
In order to examine diVerences between groups onmultiple potentially correlated measures, we ran multi-variate analyses using the SAS GLM (Generalized Lin-ear Model) procedure (SAS v8.02; SAS Institute, Cary,NC). Univariate repeated measures analyses of vari-ance were run using the SAS MIXED procedure (ineach case, the Akaike Information Criterion (AIC)statistic was used to choose the covariance matrixstructure). For post hoc comparisons, P values werecompared to levels adjusted by using a Bonferroni-Holm sequential procedure. The primary outcomemeasure was the average co-contraction over Xexionand extension compared between children with dysto-nia and controls. Secondary measures included analysisof EMG activation levels during individual (accelera-tion/deceleration and pause) phases, the duration ofcomplete movement cycles and of each phase, the Xex-ion/extension velocity peaks, the maximum elbow rota-tion, and the coeYcient of variation for each of thesemeasures.
Results
Preliminary statistical tests
We performed a set of preliminary analyses to deter-mine the signiWcance of diVerences between the twoarms (more vs. less impaired for the children with dys-tonia; dominant vs. non-dominant for the controls).Using observations on both arms, we ran a one wayrepeated measures MANOVA on the duration of thecycle of movement (Xexion/extension) and its variabil-ity (CV) that included group and arm as between andwithin subject factors. For the children with dystonia,
we opposed the less impaired to the most impairedlimb, and for the healthy controls we contrasted thedominant to the non-dominant arm. (The subject withdystonia (subject 5) for whom data from only the rightarm was available was excluded from this part of theanalysis.) As expected, the children with dystoniashowed longer and more variable cycle duration thanthe healthy subjects [Wilks’ Lambda, F(2,10) = 11.37,P = 0.0027]. Associated univariate tests revealed thatthe two groups diVered the most in their degree of var-iability [F(1,11) = 23.34, P = 0.0005, for CV of cycleduration; and F(1,11) = 4.63, P = 0.0546, for cycle dura-tion]. There was an interaction between arm and group[Wilks’ Lambda, F(2,10) = 6.43, P = 0.0160; univariatetests: F(1,11) = 5.39, P = 0.0405, for duration; andF(1,11) = 8.36, P = 0.0147, for CV], suggesting a diVer-ent eVect of changing the arm on performance of thesubjects with and without dystonia. Complementaryunivariate repeated measures ANOVAs were runwhose post-hoc comparisons showed that subjects withdystonia were signiWcantly slower and more variablewith the most impaired arm than with the less impairedone (for cycle duration: 1.991 § 0.241 (mean § SE) vs.1.570 § 0.246 s, P = 0.0098; for CV of cycle duration:0.213 § 0.01 vs. 0.134 § 0.01%; P = 0.0010) while thecontrols performed comparably well with the dominantand non-dominant arm (for cycle duration:1.011 § 0.147 vs. 1.020 § 0.104 s, P = 0.9416; for CV ofcycle duration: 0.087 § 0.008 vs. 0.084 § 0.007%;P = 0.8690). In order to evaluate the eVects of dystoniccompared with unimpaired arms, analyses will focus oncomparisons between data collected on the mostimpaired arm of the subjects with dystonia and obser-vations on the dominant arm of the healthy subjects(for subject 5, we used the data collected on the lessimpaired arm). In this way we hope to accentuate themagnitude of any diVerences between dystonic andcontrol subjects.
Fig. 2 Elbow kinematics and muscle activation pattern for a single control subject; smooth transition between the Xexion and extension phases was typi-cally observed in the controls. Some controls exhibited very high levels of antagonist mus-cle coactivation. a Elbow velocity proWles for individual trials. b Means (§ SD) of the EMG activity amplitudes for the biceps (top) and the triceps (bottom) muscles correspond-ing to the trials shown in a
123
212 Exp Brain Res (2007) 176:206–216
Given the fact that our subjects were seated unre-strained (which was necessitated by the abnormal trun-cal postures in some children with dystonia), and thushad the possibility to move their trunk toward the dis-tal target, a necessary preliminary was to ensure thatreaches by subjects of both groups involved compara-ble elbow excursions. Rotation amplitude was87.78 § 5.91° (mean § SE) for the subjects with dysto-nia and 92.39 § 4.79° for the controls, and rotationamplitude CV was 15.9 § 2.8% for subjects with dysto-nia and 11.4 § 2.7% for the healthy subjects; these val-ues were not signiWcantly diVerent [mutivariate test:Wilks’ Lambda, F(2,11) = 0.61, P = 0.5632; univariatetests: F(1,12) = 0.37, P = 0.5552, for amplitude;F(1,12) = 1.31, P = 0.2741, for CV of amplitude].
Analysis of kinematics
We compared the two groups of subjects on the dura-tion of the movement cycles and the maximum angularelbow velocities reached during Xexion and extension,whose means (§ SE) are presented in Fig. 3a. A oneway MANOVA on these three variables was not sig-niWcant [Wilks’ Lambda, F(3,10) = 2.02, P = 0.1754],but univariate analyses showed that while the subjectswith dystonia performed movement cycles of longerduration than the controls [F(1,12) = 5.85, P = 0.0323],the maximum elbow velocities they reached during
Xexion and extension were not reliably lower[F(1,12) = 3.06, P = 0.1056, for Xexion; F(1,12) = 1.35,P = 0.2680, for extension]. This is easily explainedwhen one examines the velocity proWles that character-ized the reaching cycles by the children with dystonia.As an example, Fig. 1a and c shows the elbow angularvelocity traces for trials by a single subject with dysto-nia (subject 3); one may see how this subject pauses attargets before reversing movement direction. In con-trast, smooth transition between Xexion and extensionwas typically observed in the healthy children; as illus-trated in Fig. 2a that shows velocity proWles for individ-ual trials by a representative control subject. In fact,the diVerence in movement duration between groupsdid not remain signiWcant after subtracting the pausesat the targets from the movement durations of the chil-dren with dystonia [t(12) = 0.57, P = 0.577]. Durationof the pauses was 0.358 § 0.104 s (mean § SE) at thedistal and 0.336 § 0.062 s at the proximal targets.
Analysis of kinematic variability
The mean values (§ SE) of the CV for the duration ofthe movement cycles as well as for the maximum angu-lar elbow velocities reached during Xexion and exten-sion are shown in Fig. 3b. A one way MANOVA onthese variables showed that the subjects with dystoniaexhibited substantially increased variability relative to
Fig. 3 Kinematic and kinematic variability measures for the twogroups of subjects. a From left to right, means (§ SE) across sub-jects for duration of the complete Xexion/extension movement cy-cles, and Xexion and extension velocity peaks. b From left to right,
coeYcients of variation (CV) for complete movement durationand for Xexion and extension velocity peaks. c CV for the dura-tion of pauses and the magnitude of velocity peaks during Xexionand extension (*P < 0.05, **P < 0.01, ***P < 0.001)
123
Exp Brain Res (2007) 176:206–216 213
the controls [Wilks’ Lambda, F(3,10) = 8.30,P = 0.0046]. Univariate tests revealed further that thisgroup eVect was mostly due to diVerences in cycleduration variability [F(1,12) = 28.90, P = 0.0002]. Infact, while the velocities reached by the subjects withdystonia were more variable than those reached by thecontrols during the Xexion phase [F(1,12) = 11.93,P = 0.0048], both groups exhibited comparable level ofvariability during the extension phase [F(1,12) = 2.75,P = 0.1230]. For the children with dystonia, a compari-son between CV of pauses and CV of velocity peaks—whose mean values are presented in Fig. 3c—showedthat these subjects were substantially more variable inthe duration of the pauses they observed at targetsthan in the velocity peaks they reached [repeated mea-sures ANOVA, F(1,6) = 39.96, P = 0.0007], suggestingthat much of the variability in cycle duration was dueto variability in the duration of pauses at the targets.
Analysis of EMG
The patterns of EMG activities underlying the diVer-ences between subjects with dystonia and controlswere examined using three diVerent measures. Toassess the level of antagonist muscle co-activation, weused DOT and MIN (see Methods); and to describeEMG activity modulation across reaching cycles, weused the average EMG amplitude observed for eachmuscle during each movement phase.
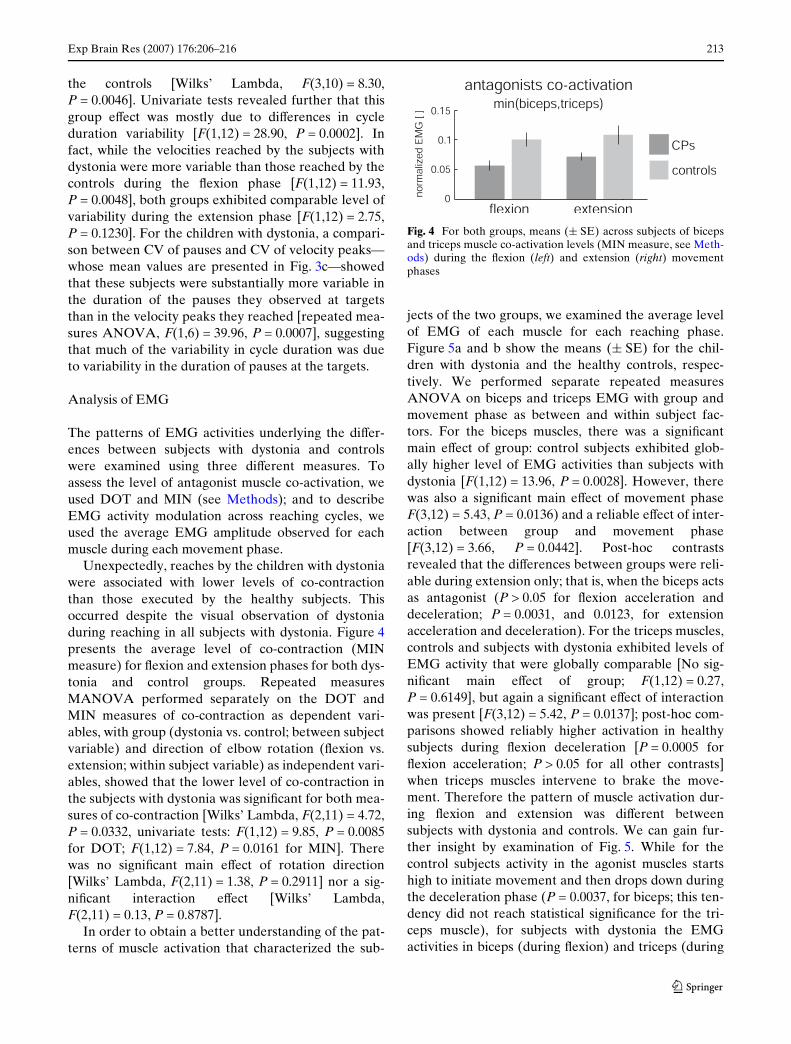
Unexpectedly, reaches by the children with dystoniawere associated with lower levels of co-contractionthan those executed by the healthy subjects. Thisoccurred despite the visual observation of dystoniaduring reaching in all subjects with dystonia. Figure 4presents the average level of co-contraction (MINmeasure) for Xexion and extension phases for both dys-tonia and control groups. Repeated measuresMANOVA performed separately on the DOT andMIN measures of co-contraction as dependent vari-ables, with group (dystonia vs. control; between subjectvariable) and direction of elbow rotation (Xexion vs.extension; within subject variable) as independent vari-ables, showed that the lower level of co-contraction inthe subjects with dystonia was signiWcant for both mea-sures of co-contraction [Wilks’ Lambda, F(2,11) = 4.72,P = 0.0332, univariate tests: F(1,12) = 9.85, P = 0.0085for DOT; F(1,12) = 7.84, P = 0.0161 for MIN]. Therewas no signiWcant main eVect of rotation direction[Wilks’ Lambda, F(2,11) = 1.38, P = 0.2911] nor a sig-niWcant interaction eVect [Wilks’ Lambda,F(2,11) = 0.13, P = 0.8787].
In order to obtain a better understanding of the pat-terns of muscle activation that characterized the sub-
jects of the two groups, we examined the average levelof EMG of each muscle for each reaching phase.Figure 5a and b show the means (§ SE) for the chil-dren with dystonia and the healthy controls, respec-tively. We performed separate repeated measuresANOVA on biceps and triceps EMG with group andmovement phase as between and within subject fac-tors. For the biceps muscles, there was a signiWcantmain eVect of group: control subjects exhibited glob-ally higher level of EMG activities than subjects withdystonia [F(1,12) = 13.96, P = 0.0028]. However, therewas also a signiWcant main eVect of movement phaseF(3,12) = 5.43, P = 0.0136) and a reliable eVect of inter-action between group and movement phase[F(3,12) = 3.66, P = 0.0442]. Post-hoc contrastsrevealed that the diVerences between groups were reli-able during extension only; that is, when the biceps actsas antagonist (P > 0.05 for Xexion acceleration anddeceleration; P = 0.0031, and 0.0123, for extensionacceleration and deceleration). For the triceps muscles,controls and subjects with dystonia exhibited levels ofEMG activity that were globally comparable [No sig-niWcant main eVect of group; F(1,12) = 0.27,P = 0.6149], but again a signiWcant eVect of interactionwas present [F(3,12) = 5.42, P = 0.0137]; post-hoc com-parisons showed reliably higher activation in healthysubjects during Xexion deceleration [P = 0.0005 forXexion acceleration; P > 0.05 for all other contrasts]when triceps muscles intervene to brake the move-ment. Therefore the pattern of muscle activation dur-ing Xexion and extension was diVerent betweensubjects with dystonia and controls. We can gain fur-ther insight by examination of Fig. 5. While for thecontrol subjects activity in the agonist muscles startshigh to initiate movement and then drops down duringthe deceleration phase (P = 0.0037, for biceps; this ten-dency did not reach statistical signiWcance for the tri-ceps muscle), for subjects with dystonia the EMGactivities in biceps (during Xexion) and triceps (during
Fig. 4 For both groups, means (§ SE) across subjects of bicepsand triceps muscle co-activation levels (MIN measure, see Meth-ods) during the Xexion (left) and extension (right) movementphases
123
214 Exp Brain Res (2007) 176:206–216
extension) stay essentially at the same level throughoutthe acceleration and deceleration phases (P > 0.05, forboth muscles). Thus subjects with dystonia did notexhibit the same degree of modulation of muscle activ-ity as seen in the controls.
Note that—in contrast to the control subjects—inchildren with dystonia the relative activity of the bicepsand triceps muscles was diVerent at the end of Xexionand the beginning of extension, as well as at the end ofextension and the beginning of Xexion. Thus thesubjects with dystonia did not demonstrate a smoothtransition from the end of the extension phase to thebeginning of the Xexion phase. We conjecture thatthe pause may in part be necessary in order to “reset”the muscle activities prior to the next movement.
Finally, we looked at the relation between our kine-matic and EMG measures and the severity of symp-toms for the children with dystonia. As expected fromthe Wnding that co-contraction is greater in the controlsubjects, there was no signiWcant correlation in the sub-jects with dystonia between co-contraction and severityof dystonia (MIN measure against BFM: corr = 0.2008,P = 0.6659; UDRS: corr = 0.2312, P = 0.6179; BAD:corr = 0.4376, P = 0.3260). Correlations were muchhigher with the duration of the pauses, and approachedbut did not reach signiWcance (duration of pauses andBFM: corr = 0.6378, P = 0.1232; UDRS: corr = 0.7100,P = 0.0738; BAD: corr = 0.5743, P = 0.1774). We sus-pect that with a larger number of subjects this lattercorrelation would achieve signiWcance.
Discussion
We examined unconstrained reaching movements bychildren with primary and secondary dystonia. Wedescribed the kinematics of elbow movement and thesurface EMG activity in the biceps and triceps. Asexpected, the analysis of kinematics showed increasedmovement duration and increased variability in move-
ment duration in the children with dystonia comparedto control subjects. However, the increased duration ofthe reaching cycles by the children with dystonia wasmostly due to extensive pauses that these subjectsexhibited at targets, rather than to reduced elbow rota-tion velocities. The increased variability was mostlydue to variability in the duration of the pauses.
We assessed if the abnormalities observed in the chil-dren with dystonia could be explained in terms of inap-propriate antagonist muscle activation. Unexpectedly,we found that children with dystonia showed lower lev-els of co-contraction than healthy controls during move-ment. Moreover, the pauses at the targets thatcharacterized the reaching cycles by the subjects withdystonia were associated with low levels of muscle acti-vation, rather than co-contraction. Examination of theEMG patterns throughout the complete movementcycle indicated that the delays observed by subjects withdystonia between movements in opposite directionsseem to arise as a consequence of deWcient patterns ofagonist/antagonist muscle activation. SpeciWcally, inthese children activation of the agonist muscle remainedinappropriately constant throughout elbow displace-ment, and EMG activity in the antagonist muscle wasnot properly built up in order to slow elbow rotation andprepare for smooth movement reversal. As a conse-quence, erratic elbow trajectories were observed at theend of movement and the pauses at targets could beexplained by a necessity for the system to “reset” EMGactivity prior to reversal of movement direction.
Since basal ganglia have been implicated in deWcits oftiming and muscle activation as well as in abnormalitiesof motor pattern, our results are consistent with thehypothesis of deWcits of basal ganglia function in chil-dren with dystonia. However, brain injury in secondarydystonia may be widespread, and we cannot exclude thepossibility that our Wndings are due to a combination ofmotor deWcits, including dystonia, bradykinesia, cho-reoathetosis, weakness, or spasticity. Extensive pauseshave been described in adult patients with hand
Fig. 5 EMG activity levels throughout the diVerent movementphases for both groups of subjects; clearly diVerent muscle activa-tion patterns were observed for the two groups of subjects. a, b
Means (§ SE) of the EMG activity levels for the children withdystonia and the control subjects, respectively
123
Exp Brain Res (2007) 176:206–216 215
dystonia performing repetitive Wnger Xexion/extensionmovements (Curra et al. 2004). But in contrast to whatwe observed in children with dystonia (for whom thepeak velocity during movement was not signiWcantlydiVerent from controls), the adult patients also showedbradykinesia (Curra et al. 2000). Movement “fragmen-tation” is not a feature that is speciWc to dystonia andcan also be observed, for instance, in developing infants(von Hofsten 1991) or in patients following stroke(Krebs et al. 1999). Furthermore, the gradual blendingof submovements observed during development or dur-ing stroke recovery has been interpreted as evidencethat complex movements are composed of discrete bal-listic movements (Rohrer et al. 2002).
In contrast with this hypothesis, other recent Wnd-ings point to the idea that rhythmic arm movementmay be fundamentally diVerent from discrete armmovements (Schaal et al. 2004). In the present context,further work would be needed to determine if thepauses exhibited by the subjects with dystonia aresymptomatic of complex movement fragmentation orwhether they reveal fundamental diVerences in themovement strategy adopted by the children with dysto-nia relative to healthy subjects. For example, childrenwith dystonia might select (perhaps involuntarily) astrategy that includes sequential discrete reachingmovements rather than smooth rhythmic movements.Since the subjects with dystonia and the control sub-jects used a fundamentally diVerent type of trajectoryof movement, our results need to be interpreted withcaution since the diVerences in EMG may be partlyexplained by diVerences in the type of movement. Thispossibility does not weaken our main Wnding ofdecreased co-contraction in dystonia, but it does sug-gest that decreased co-contraction could be at leastpartly due to diVerences in the choice of trajectoryrather than to diVerences in the activation of musclesfor the same trajectory. Further study of this questionwill require experiments in which both groups of sub-jects are required to perform point-to-point move-ments with similar movement trajectories, in order tomatch the kinematic and dynamic properties of themovements as well as possible.
In agreement with the reduced co-contraction shownin our results, MacKinnon and co-workers (2004)reported that in ballistic wrist movements, prolongedagonist activation with co-contraction of the antagonistmuscle could also be observed in healthy controls andtherefore was not a feature speciWc to their adultpatients with primary dystonia (who in fact exhibiteddeWcits in muscle activation after movement onset ratherthan increased EMG activity). In the present study, thehigh levels of co-contraction that could be observed in
controls—as illustrated in Fig. 2a—may have been aconsequence of the biomechanical requirements of thetask we used. In order to achieve smooth and rapid tran-sitions between elbow rotations in opposite directions,the healthy subjects maintained high levels of activationin both agonist and antagonist groups of musclesthroughout the series of repetitive reaching movements.The function of muscles controlling the elbow may havebeen to stabilize against interaction torques, while it ispossible that shoulder muscles were the primary driversof movement. When one examines Fig. 5, this seems tobe especially true for the extension phase where rotationof the elbow might have been mostly determined byshoulder muscle contraction, while the biceps may haveintervened essentially in order to achieve stabilization ofthe limb at the endpoint.
This study has several important weaknesses. In anattempt to characterize movement abnormalities in bothprimary and secondary dystonia, we included childrenfrom both groups and a large range of ages. However,the small total number of subjects does not allow com-parison between groups, nor do we have adequate sub-jects to determine the eVects of age. The study subjectswere homogeneous in the sense of having the symptomof dystonia, but they were heterogeneous in terms of eti-ology. Although there was no signiWcant diVerence inpeak velocity between children with dystonia and con-trol subjects, we did not attempt to match other featuresof the trajectory such as acceleration proWles or handpath, and it is thus possible that the increased co-con-traction in the controls was due to the details of theirmovement trajectories. (For example, since childrenwith dystonia moved more slowly, it is possible that thisreduced the need to counteract elbow interaction torquegenerated by shoulder movement.) This possibility doesnot weaken our conclusion (that co-contraction was notsigniWcant in the children with dystonia) but it suggeststhat future studies should attempt to match the kine-matic features of movement as much as possible.Despite these weaknesses, we feel that this study yieldsan important initial insight into the control of musclesduring movement in children with dystonia. Althoughco-contraction may be present in certain adult dystoniasand it may be present during static postures in childrenwith dystonia, it may not be a dominant feature of activemovement in children with dystonia.
In summary, in the present study, much of the vari-ability in the elbow kinematics in the children with dys-tonia could be explained by abnormalities of the timingof sub-movements. Further, movement timing variabil-ity was mostly unrelated to variability in the transportphase, but instead was related to diYculty in reversingmovement direction. The prolonged pauses observed
123
216 Exp Brain Res (2007) 176:206–216
at targets did not appear to be associated with inappro-priate co-contraction of antagonistic groups of muscles.Children with dystonia did not show higher levels ofco-contraction relative to the healthy children. There-fore we conclude that co-contraction may not be amajor cause of movement abnormality during uncon-strained reaching in childhood dystonia.
Acknowledgments This work was supported by grant numberNS041243 from the National Institute of Neurologic Disordersand Stroke to Dr. Sanger, as well as by the Stanford UniversityDepartment of Neurology and Neurological Sciences. Weacknowledge additional funding from the Don and Linda CarterFoundation and the Crowley Carter Foundation. Several peoplecontributed to recruitment of subjects and performance of exper-iments, including Kimberly Murphy, Sara Sherman-Levine, AbeIshihara, Brian Placek, and Jason Kaiser; we are very grateful fortheir eVorts.
References
Barry MJ, VanSwearingen JM et al (1999) Reliability and respon-siveness of the Barry-Albright Dystonia Scale. Dev MedChild Neurol 41(6): 404–411
Berardelli A, Rothwell JC et al (1998) The pathophysiology ofprimary dystonia. Brain 121(Pt 7):1195–1212
Bookstein FL (1991) Morphometric tools for landmark data:geometry and biology. Cambridge University Press, Cam-bridge
Burke RE, Fahn S, Marsden CD, Bressman SB, Moskowitz C,Friedman J (1985) Validity and reliability of a rating scale forthe primary torsion dystonias. Neurology 35:73–77
Chang J-J, Wu T-I et al (2005) Kinematical measure for spasticreaching in children with cerebral palsy. Clin Biomech20(4):381–388
Clancy EA, Hogan N (1994) Single site electromyograph ampli-tude estimation. IEEE Trans Biomed Eng 41(2):159–167
Comella CL, Leurgans S, Wuu J, Stebbins GT, Chmura T (2003)Rating scales for dystonia: a multicenter assessment. MovDisord 18:303–312
Curra A, Agostino R et al (2004) Impairment of individual Wngermovements in patients with hand dystonia. Mov Disord19(11):1351–1357
Curra A, Berardelli A et al (2000) Movement cueing and motorexecution in patients with dystonia: a kinematic study. MovDisord 15(1):103–112
Damiano DL, Abel MF (1996) Relation of gait analysis to grossmotor function in cerebral palsy. Dev Med Child Neurol38:389–396
Damiano DL, Martellotta TL et al (2000) Muscle force produc-tion and functional performance in spastic cerebral palsy:relationship of cocontraction. Arch Phys Med Rehabil81(7):895–900
Duque J, Thonnard JL et al (2003) Correlation between impaireddexterity and corticospinal tract dysgenesis in congenitalhemiplegia. Brain 126:732–747
Fahn S, Marsden CD et al (1987) ClassiWcation and investigationof dystonia. In: Marsden CD, Fahn S (eds) Movement disor-ders 2. Butterworths, London, pp 332–358
Fetters L, Kluzik J (1996) The eVects of neurodevelopmentaltreatment versus practice on the reaching of children withspastic cerebral palsy. Phys Ther 76(4):346–358
Harris SR (1991) Movement analysis—an aid to early diagnosisof cerebral palsy. Phys Ther 71:215–221
Hogan N, Mann RW (1980) Myoelectric signal processing: opti-mal estimation applied to electromyography—Part I: Deri-vation of the optimal myoprocessor. IEEE Trans BiomedEng 27:382–395
Krebs HI, Aisen ML, et al (1999) Quantization of continuous armmovements in humans with brain injury. Proc Natl Acad SciUSA 96(8):4645–4649
MacKinnon CD, Velickovic M et al (2004) Corticospinal excit-ability accompanying ballistic wrist movements in primarydystonia. Mov Disord 19(3):273–284
Marsden CD (1984) The pathophysiology of movement disor-ders. Neurol Clin 2(3):435–459
McPherson JJ, Schild R et al (1991) Analysis of upper extremitymovement in four sitting positions: a comparison of personswith and without cerebral palsy. Am J Occup Ther45(2):123–129
O’Byrne JM, Jenkinson A et al (1998) Quantitative analysisand classiWcation of gait patterns in cerebral palsy using athree-dimensional motion analyzer. J Child Neurol13(3):101–108
Palisano R, Rosenbaum P et al (1997) Development and reliabil-ity of a system to classify gross motor function in childrenwith cerebral palsy. Dev Med Child Neurol 39(4):214–223
Ramsay JO, Silverman BW (1997) Functional data analysis.Springer, Berlin Heidelberg New York
Rohrer B, Fasoli S et al (2002) Movement smoothness changesduring stroke recovery. J Neurosci 22(18):8297–8304
Sanger TD (2003) Pathophysiology of pediatric movement disor-ders. J Child Neurol 18(Suppl 1):S9–S24
Sanger TD, Delgado MR et al (2003) ClassiWcation and deWnitionof disorders causing hypertonia in childhood. Pediatrics111(1):e89–e97
Sanger TD, Kaiser J et al (2005) Reaching movements in child-hood dystonia contain signal-dependent noise. J Child Neu-rol 20(6):489–496
Schaal S, Sternad D et al (2004) Rhythmic arm movement is notdiscrete. Nat Neurosci 7:1136–1143
Thelen DD, Riewald SA et al (2003) Abnormal coupling of kneeand hip moments during maximal exertions in persons withcerebral palsy. Muscle Nerve 27(4):486–493
Utley A, Sugden D (1998) Interlimb coupling in children withhemiplegic cerebral palsy during reaching and grasping atspeed. Dev Med Child Neurol 40:396–404
Van Der Heide J, Fock JM et al (2005a) Kinematic characteristicsof postural control during reaching in preterm children withcerebral palsy. Pediatr Res 58(3):586–593
van der Heide JC, Fock JM et al (2005b) Kinematic characteris-tics of reaching movements in preterm children with cerebralpalsy. Pediatr Res 57(6):883–889
Vitek JL (2002) Pathophysiology of dystonia: a neuronal model.Mov Disord 17(Suppl 3):S49–S62
von Hofsten C (1991) Structuring of early reaching movements: alongitudinal study. J Mot Behav 23:280–292
Yao J, Acosta AM et al (2004) Muscle focus: a new biomechani-cal-based index on the selectivity of EMG activity and itsapplication in quantifying the muscle coactivation patternsduring isometric torque generation at the elbow and shoul-der. In: Proceedings 26th annual international conference onIEEE EMBS, San Francisco
Yao J, Acosta AM et al (2006) A biomechanics-based method forthe quantiWcation of muscle selectivity in a musculoskeletalsystem. J Biomech 39(8):1527–1530