ORIGINAL RESEARCH Early Response of Bone Marrow Osteoprogenitors to Skeletal Unloading and Sclerostin Antibody Mohammad Shahnazari • Thomas Wronski • Vivian Chu • Alyssa Williams • Alicia Leeper • Marina Stolina • Hua Zhu Ke • Bernard Halloran Received: 24 February 2012 / Accepted: 16 April 2012 / Published online: 27 May 2012 Ó Springer Science+Business Media, LLC 2012 Abstract Sclerostin functions as an antagonist to Wnt signaling and inhibits bone-forming activity. We studied the effects of skeletal unloading and treatment with sclerostin antibody (Scl-Ab) on mesenchymal stem cell, osteoprogenitor and osteoclast precursor pools, and their relationship to bone formation and resorption. Male C57BL/6 mice (5-months-old) were hind limb unloaded for 1 week or allowed normal ambulation and treated with Scl- Ab (25 mg/kg, s.c. injections on days 1 and 4) or placebo. Unloading decreased the serum concentration of bone formation marker P1NP (-35 %), number of colony- forming units (CFU) (-38 %), alkaline phosphatase– positive CFUs (CFU-AP?)(-51 %), and calcified nodules (-35 %); and resulted in a fourfold increase in the number of osteoclast precursors. The effects of Scl-Ab treatment on unloaded and normally loaded mice were nearly identical; Scl-Ab increased serum P1NP and the number of CFU, CFU-AP?, and calcified nodules in ex vivo cultures; and increased osteoblast and bone mineralizing surfaces in vivo. Although the marrow-derived osteoclast precursor population increased with Scl-Ab, the bone osteoclast surface did not change, and the serum concentration of osteoclast activity marker TRACP5b decreased. Our data suggest that short-term Scl-Ab treatment can prevent the decrease in osteoprogenitor population associated with skeletal unloading and increase osteoblast surface and bone mineralizing surface in unloaded animals. The anabolic effects of Scl-Ab treatment on bone are preserved during skeletal unloading. These findings suggest that Scl-Ab treatment can both increase bone formation and decrease bone resorption, and provide a new means for prevention and treatment of disuse osteoporosis. Keywords Bone Á Osteoblast Á Osteoclast Á Sclerostin Á Skeletal unloading Targeting the Wnt signaling pathway to augment bone formation has been the focus of numerous recent studies [1–4]. Wnt pathways are involved in coordinating proper bone development, formation, and growth, both before and after birth [5, 6]. Signaling by Wnt proteins is antagonized by sclerostin, which is expressed mainly by osteocytes and functions to inhibit bone formation [7, 8]. Targeted dele- tion of the sclerostin gene increases the osteoblast surface (Ob.S/BS), mineralizing surface (MS/BS), bone formation rate (BFR), and bone volume (BV/TV) [9]. Treatment of adult rats with sclerostin antibody (Scl-Ab) for 3–5 weeks is reported to increase MS/BS and BFR [10] and of estrogen-deficient rats to increase Ob.S/BS, MS/BS, BFR, and BV/TV [11]. These data overwhelmingly demonstrate the anabolic effect of decreasing sclerostin activity. The effects of blocking sclerostin activity on bone resorption are less clear; the ratio of ratio of osteoclast surface to bone Thomas Wronski received research funding from Amgen Inc. Marina Stolina and Hua Zhu Ke are employed by Amgen. Marina Stolina, Hua Zhu Ke, and Thomas Wronski have stock ownership in Amgen. Other authors have stated that they have no conflict of interest. M. Shahnazari Á V. Chu Á B. Halloran (&) Division of Endocrinology, Veterans Affairs Medical Center, University of California, San Francisco, CA 94121, USA e-mail: [email protected]T. Wronski Á A. Williams Á A. Leeper Department of Physiological Sciences, College of Veterinary Medicine, University of Florida, Gainesville, FL 32610-0125, USA M. Stolina Á H. Z. Ke Department of Metabolic Disorders, Amgen Inc., One Amgen Center Drive, Thousand Oaks, CA 91320, USA 123 Calcif Tissue Int (2012) 91:50–58 DOI 10.1007/s00223-012-9610-9
Transcript
ORIGINAL RESEARCH
Early Response of Bone Marrow Osteoprogenitors to SkeletalUnloading and Sclerostin Antibody
Mohammad Shahnazari • Thomas Wronski •
Vivian Chu • Alyssa Williams • Alicia Leeper •
Marina Stolina • Hua Zhu Ke • Bernard Halloran
Received: 24 February 2012 / Accepted: 16 April 2012 / Published online: 27 May 2012
� Springer Science+Business Media, LLC 2012
Abstract Sclerostin functions as an antagonist to Wnt
signaling and inhibits bone-forming activity. We studied
the effects of skeletal unloading and treatment with
sclerostin antibody (Scl-Ab) on mesenchymal stem cell,
osteoprogenitor and osteoclast precursor pools, and their
relationship to bone formation and resorption. Male
C57BL/6 mice (5-months-old) were hind limb unloaded for
1 week or allowed normal ambulation and treated with Scl-
Ab (25 mg/kg, s.c. injections on days 1 and 4) or placebo.
Unloading decreased the serum concentration of bone
formation marker P1NP (-35 %), number of colony-
forming units (CFU) (-38 %), alkaline phosphatase–
positive CFUs (CFU-AP?) (-51 %), and calcified nodules
(-35 %); and resulted in a fourfold increase in the number
of osteoclast precursors. The effects of Scl-Ab treatment on
unloaded and normally loaded mice were nearly identical;
Scl-Ab increased serum P1NP and the number of CFU,
CFU-AP?, and calcified nodules in ex vivo cultures; and
increased osteoblast and bone mineralizing surfaces in
vivo. Although the marrow-derived osteoclast precursor
population increased with Scl-Ab, the bone osteoclast
surface did not change, and the serum concentration of
osteoclast activity marker TRACP5b decreased. Our data
suggest that short-term Scl-Ab treatment can prevent the
decrease in osteoprogenitor population associated with
skeletal unloading and increase osteoblast surface and bone
mineralizing surface in unloaded animals. The anabolic
effects of Scl-Ab treatment on bone are preserved during
skeletal unloading. These findings suggest that Scl-Ab
treatment can both increase bone formation and decrease
bone resorption, and provide a new means for prevention
and treatment of disuse osteoporosis.
Keywords Bone � Osteoblast � Osteoclast � Sclerostin �Skeletal unloading
Targeting the Wnt signaling pathway to augment bone
formation has been the focus of numerous recent studies
[1–4]. Wnt pathways are involved in coordinating proper
bone development, formation, and growth, both before and
after birth [5, 6]. Signaling by Wnt proteins is antagonized
by sclerostin, which is expressed mainly by osteocytes and
functions to inhibit bone formation [7, 8]. Targeted dele-
tion of the sclerostin gene increases the osteoblast surface
(Ob.S/BS), mineralizing surface (MS/BS), bone formation
rate (BFR), and bone volume (BV/TV) [9]. Treatment of
adult rats with sclerostin antibody (Scl-Ab) for 3–5 weeks
is reported to increase MS/BS and BFR [10] and of
estrogen-deficient rats to increase Ob.S/BS, MS/BS, BFR,
and BV/TV [11]. These data overwhelmingly demonstrate
the anabolic effect of decreasing sclerostin activity. The
effects of blocking sclerostin activity on bone resorption
are less clear; the ratio of ratio of osteoclast surface to bone
Thomas Wronski received research funding from Amgen Inc. Marina
Stolina and Hua Zhu Ke are employed by Amgen. Marina Stolina,
Hua Zhu Ke, and Thomas Wronski have stock ownership in Amgen.
Other authors have stated that they have no conflict of interest.
M. Shahnazari � V. Chu � B. Halloran (&)
Division of Endocrinology, Veterans Affairs Medical Center,
University of California, San Francisco, CA 94121, USA
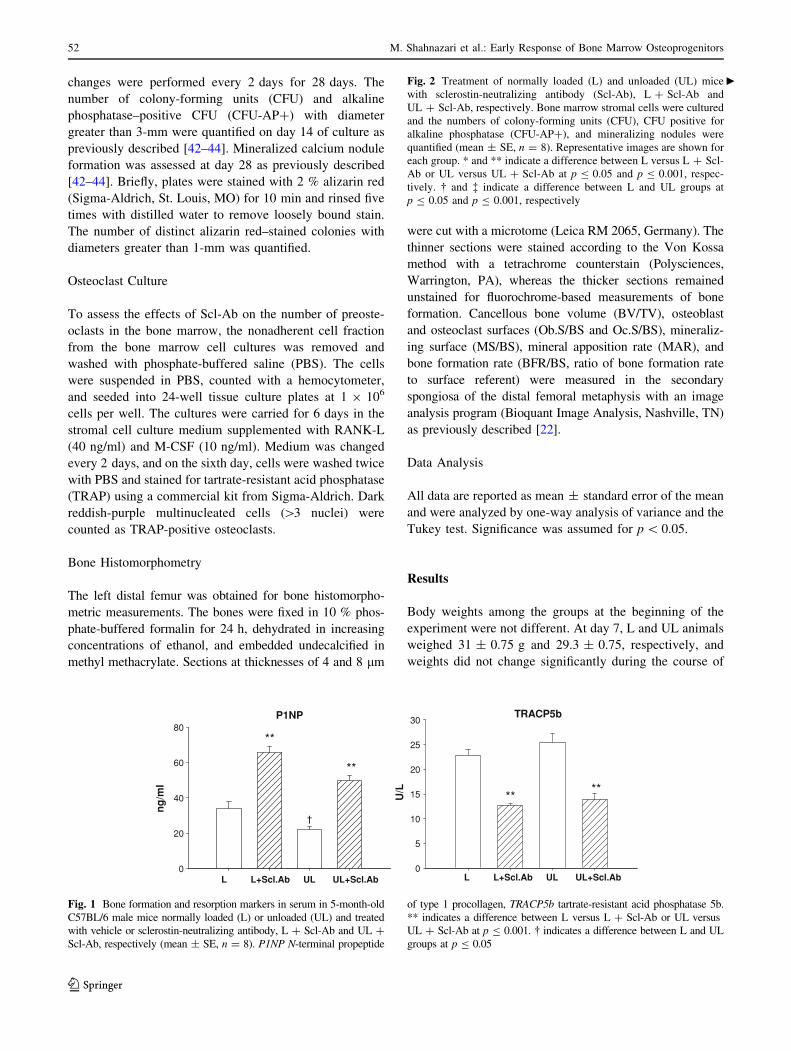
L loaded, UL unloaded, Scl-Ab sclerostin neutralizing antibody, BV/TV ratio of bone volume to total volume, Tb.Th. trabecular thickness, Tb.N.trabecular number, Tb.Sp. trabecular separation, MS/BS ratio of mineralizing surface to bone surface, MAR mineral apposition rate, BFR/BS ratio
of bone formation rate to surface referent, Oc.S/BS ratio of osteoclast surface to bone surface, Ob.S/BS ratio of osteoblast surface to bone surface
Difference between L versus L ? Scl-Ab or UL versus UL ? Scl-Ab is shown for * p B 0.05 and ** p B 0.001
M. Shahnazari et al.: Early Response of Bone Marrow Osteoprogenitors 55
123
did not expect to find a Scl-Ab effect after only 1 week of
treatment.
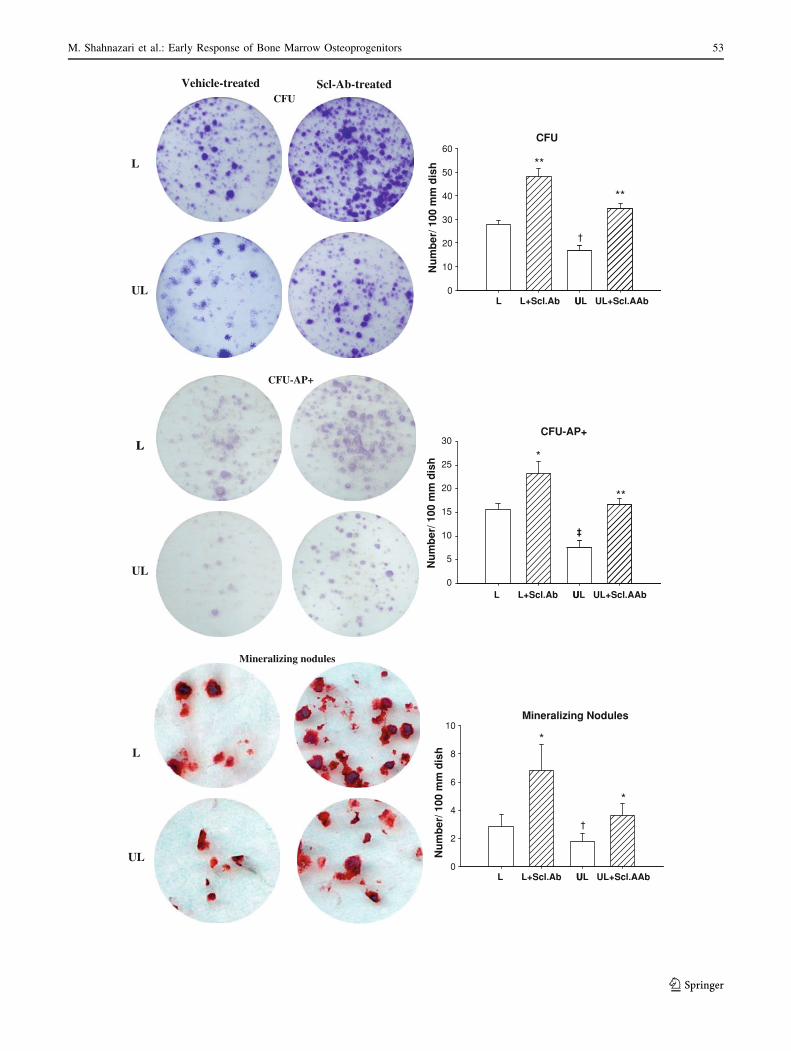
The changes in CFU-AP? and calcium nodule number
induced by Scl-Ab treatment and unloading were propor-
tional to the changes in CFU or MSC number, suggesting
that the increase in osteoprogenitor number in treated mice
may reflect the increase in the MSC number. Scl-Ab
treatment seems to increase the MSC pool size but to have
less of an effect on the propensity of these cells to form
osteoblasts in vitro. In vivo, however, our data do not
exclude the possibility that neutralization of sclerostin
activity may promote recruitment of osteoprogenitors into
the osteoblast lineage. It may also stimulate migration of
osteoblasts to the bone surface and bone-forming activity.
Scl-Ab treatment of normally loaded and unloaded mice
also increased the osteoclast precursor population. This
unexpected result suggests that Wnt signaling may play an
important role in regulation of the osteoclast precursor pool
[13]. Although limited, there are data directly linking
LRP5/6 expression and hematopoietic stem cell function
[48, 49], and canonical Wnt signaling has been shown to
alter the differentiation potential of Lin-CD34?CD1a-
human thymic progenitors [14]. Sclerostin may also act
indirectly to regulate the osteoclast precursor pool.
Recruitment of cells into the hematopoietic lineages is a
dynamic process, and if the rate of recruitment from the
osteoclast precursor pool into the osteoclast lineage is
slowed, it is conceivable that this might result in an
abnormal accumulation of preosteoclasts. Stimulation of
Wnt/b-catenin signaling in osteoblasts has been shown to
decrease osteoclast formation by up-regulating osteopro-
tegerin (OPG) and down-regulating RANK-L [12, 13, 15,
16]. Our results are consistent with the idea that inhibition
of sclerostin action through treatment with the Scl-Ab
stimulates Wnt signaling in the osteoblast, thereby
increasing the OPG/RANK-L ratio and inhibiting osteo-
clastogenesis. Sclerostin may also influence migration to
and maintenance of the osteoclast population on the bone
surface. Spencer et al. [13] report that LRP6 is expressed in
mature osteoclasts. If sclerostin impairs migration, reduces
osteoclast stability on the bone surface, or both, it might
account, in part, for the apparent anomaly between osteo-
clast precursor number and Oc.S/BS.
Oc.S/BS has been reported to decrease [11] or remain
unchanged in response to blocking sclerostin [9]. Our cell
data are similar and suggest that Scl-Ab treatment for
1 week has little or no effect on Oc.S/BS. Serum
TRACP5b, however, was decreased, suggesting that
although osteoclast number may not change, osteoclast
activity per cell may decrease. Serum CTX (C-terminal
telopeptides of type I collagen, another marker of bone
resorption) has been reported to be unchanged after Scl-Ab
treatment, a finding consistent with the maintenance of a
normal Oc.S/BS in these studies [10]. The reason for
the discrepancy between the serum concentrations of
TRACP5b and CTX is not clear but may reflect the nature
of these variables. TRAP is expressed by cells other than
osteoclasts and may not in all instances accurately reflect
breakdown of bone.
The dose of Scl-Ab we chose was based on previous
animal studies showing efficacy on bone formation [10, 11,
39]. In future studies, it will be important to determine the
effects of lower and higher doses of Scl-Ab on osteoclast
formation and activity. Therapeutically, there may be
other doses that are more efficacious for regulating bone
resorption.
Collectively, our data suggest that sclerostin may func-
tion not only to regulate the osteoprogenitor pool size and
bone formation, but also to regulate the osteoclast precur-
sor pool size and osteoclast activity. Despite the increase in
osteoclast precursors, osteoclast number on the bone sur-
face remained unchanged and serum TRACP5b activity
clearly decreased, suggesting that Scl-Ab treatment may
act to block resorption by inhibiting recruitment of cells
from the osteoclast precursor population into the OC
lineage, impairing migration to the bone surface, sup-
pressing osteoclast fusion and activity, or causing an
increase in osteoclast turnover. These findings suggest a
new paradigm where the hematopoietic cell population and
lineage allocation are regulated by sclerostin. The benefi-
cial effects of Scl-Ab treatment in patients with low bone
mass may be mediated through increased formation and,
despite an increase in osteoclast precursors, decreased bone
resorption. Future studies are suggested to investigate the
effects of Scl-Ab on osteoclast formation in vitro to
examine the direct effects of inhibiting sclerostin on
osteoclast precursors in addition to the effects poten-
tially mediated through osteoblasts and their associated
cytokines.
In summary, our data suggest that skeletal unloading
causes a rapid decrease in the MSC and osteoprogenitor
populations, and a rapid increase in the osteoclast precursor
population in the bone marrow. Because of the short-term
nature of our studies (7 days), these changes may not be
reflected in changes in BFR/BS and bone volume. Scl-Ab
treatment in both unloaded and normally loaded mice
markedly increased MSC, osteoprogenitor, and osteoclast
precursor numbers and osteoblast surface, but did not affect
osteoclast surface. Serum concentrations of P1NP and
TRACP5b in unloaded and normally loaded, and treated
and untreated mice confirmed the pro-anabolic effects of
Scl-Ab treatment on bone and demonstrated that the ana-
bolic response of bone to Scl-Ab is preserved during
skeletal unloading or loss of weight bearing. These findings
suggest that Scl-Ab treatment may be useful in the pre-
vention and treatment of disuse osteoporosis.
56 M. Shahnazari et al.: Early Response of Bone Marrow Osteoprogenitors
123
Acknowledgments This work was supported by the Veterans
Affairs Merit Review program, NASA, and the Northern California