Indian Journal of Experimental Biology Vol. 37, April 1999, pp. 391-395 Effect of ammonium and nitrate ratio on glucose oxidase activity during gluconic acid fermentation by a mutant strain of Aspergillus niger Sima Ray & A K Banik· Department of Chemical Engineering, University of Calcutta, 92, A.P.e. Road, Calcutta 700 009, India Received 6 October 1997; revised 14 December 1998 Of the factors tested, the source and concentration of carbon and nitrogen in the medium exerted maximum effect on growth and acid production. Glucose (15%) and urea (0.14%) induced glucose oxidase synthesis and optill1um yield of cal- cium gluconate. Potassium dihydrogen phosphate (0.2%) and magnesium sulphate (0.06%) stimulated glucose oxidase ac-. tivity and calcium gluconate production. Borax at a concentration of 1.5 gi L induced maximum glucose oxidase and cal- cium gluconate production with increased glucose utilization. Glucose oxidase is of considerable industrial impor- tance because of its wide use as an analytical tool i.e., in the production of gluconic acid I, determination of D-glucose in biological fluids such as blood and fer- mentation products Z , and also as a means of remov- ing residual oxygen and sugar from foods and bever- ages). Gluconic acid producing strains of Aspergillus and Penicillium are used for commercial production of the enzyme 4 In histochemical studies, Van Dijken and Veenhuis 5 have found the enzyme in microbodies of the cells, whereas Mischak et al. 6 have reported it as being extracellular. Elimination of chemical nutri- ents, KH z P0 4 and MgS0 4 .7H z O, from the culture so- lutions resulted in the formation of extremely scanty mycelIa and no production of acid. Addition of low concentration of KH 2 P0 4 and MgS0 4 .7H z O stimu- lated acid production 7 . Low concentration of KH 2 P0 4 and MgS04 .7 H 2 0 were also found to stimulate glu- cose oxidase activity in A. nigerB. Reports are also available that addition of boron compounds like boric acid or borax to the fermentation medium increase the solubility of gluconate i.e., increase in production of calcium gluconate with increase in glucose utiliza- tion. This shows that in the design of industrial pro- duction process the strains resIstant to the action of boron must be employed in order to achieve optimal production of the enzyme. Exo- as well as endocel- lular glucose oxidase is shown to contribute almost equally to overall gluconic acid production 9 . · Co rre sponde nt auth or. Materials and Methods Culture of the mutant strains of A. niger AB-The parent culture of A. niger AB was selected from 250 isolates of the fungus from North Bengal soil. Spore suspension of this parent strain (2.6 x 10 7 spores per ' ml) was treated with ethylene imine and UV-rays in the laboratory and 1,976 mutant strains were isolated. Out of these mutants, A. niger AB 1801 which pro- duced the maximum amount (120 gIL) of calcium gluconate, was selected for further studies 10. Medium and cultural conditions--A. niger AB 1801 was maintained on solid medium consisted of malt extract, yeast extract and. agar at 4°C. . The mentation medium consisted of KH 2 P0 4 , 0.1 %; MgS0 4 ·7H 2 0 , 0. 05% (PH 6.5) and CaCO), Ig as car- bon source (4gllg of CaCO )) in the medium. The carbon and nitrogen sources and CaCO) were steril- ized seperately and added just prior to inoculation. A well sporulated 7 days old slant culture at 28°C was used. The spore density was adjusted to 2.6 x 10 7 spores/ml of the suspension. Surface culture fermen- tation was carried out using 1 'L flat flask, each con- taining 150 ml of fermentation medium. The flasks were incubated at 30°C for 9 days' o. Preparation of enzyme e xtracts-The mycelial mat was harvested after 9 days, washed with ice-cold water, press dried and weighed. The mat was placed in a porcelain mortar and ground for 5 min with sea sand. Sodium acetate buffer (0.5 M: pH 5. 1) was used for preparation of extract containing endocellular glucose oxidase. The extract was centrifuged at 500 g
Transcript
Indian Journal of Experimental Biology Vol. 37, April 1999, pp. 391-395
Effect of ammonium and nitrate ratio on glucose oxidase activity during gluconic acid fermentation by a mutant strain of Aspergillus niger
Sima Ray & A K Banik·
Department of Chemical Engineering, University of Calcutta, 92, A.P.e. Road, Calcutta 700 009, India
Received 6 October 1997; revised 14 December 1998
Of the factors tested, the source and concentration of carbon and nitrogen in the medium exerted maximum effect on growth and acid production. Glucose (15%) and urea (0.14%) induced glucose oxidase synthesis and optill1um yield of calcium gluconate. Potassium dihydrogen phosphate (0.2%) and magnesium sulphate (0.06%) stimulated glucose oxidase ac-. tivity and calcium gluconate production. Borax at a concentration of 1.5 gi L induced maximum glucose oxidase and calcium gluconate production with increased glucose utilization.
Glucose oxidase is of considerable industrial importance because of its wide use as an analytical tool i.e., in the production of gluconic acid I, determination of D-glucose in biological fluids such as blood and fermentation productsZ
, and also as a means of removing residual oxygen and sugar from foods and beverages). Gluconic acid producing strains of Aspergillus and Penicillium are used for commercial production of the enzyme4 In histochemical studies, Van Dijken and Veenhui s5 have found the enzyme in microbodies of the cell s, whereas Mischak et al.6 have reported it as being extracellular. Elimination of chemical nutrients, KHzP04 and MgS04.7HzO, from the culture solutions resulted in the formation of extremely scanty mycelIa and no production of acid. Addition of low concentration of KH 2P04 and MgS04.7HzO stimulated acid production7
. Low concentration of KH2P04
and MgS04 .7H20 were also found to stimulate glucose oxidase activity in A. nigerB. Reports are also available that addition of boron compounds like boric acid or borax to the fermentation medium increase the solubility of gluconate i.e., increase in production of calcium gluconate with increase in glucose utilization. Thi s shows that in the design of industrial production process the strains resIstant to the action of boron must be employed in order to achieve optimal production o f the enzyme . Exo- as well as endocellular glucose oxidase is shown to contribute almost equally to overall gluconic acid production9
.
· Correspondent auth or.
Materials and Methods Culture of the mutant strains of A. niger AB-The
parent culture of A. niger AB was selected from 250 isolates of the fungus from North Bengal soil. Spore suspension of this parent strain (2 .6 x 107 spores per ' ml) was treated with ethylene imine and UV-rays in the laboratory and 1,976 mutant strains were isolated. Out of these mutants, A. niger AB 1801 which produced the maximum amount (120 gIL) of calcium gluconate, was selected for further studies 10.
Medium and cultural conditions--A. niger AB 1801 was maintained on solid medium consisted of malt extract, yeast extract and. agar at 4°C. . The f~rmentation medium consisted of KH2P04, 0.1 %; MgS04·7H20 , 0 .05% (PH 6.5) and CaCO), Ig as carbon source (4gllg of CaCO)) in the medium. The carbon and nitrogen sources and CaCO) were sterilized seperately and added just prior to inoculation. A well sporulated 7 days old slant culture at 28°C was used. The spore density was adjusted to 2.6 x 107
spores/ml of the suspension. Surface culture fermentation was carried out using 1 'L flat flask, each containing 150 ml of fermentation medium. The flasks were incubated at 30°C for 9 days' o.
Preparation of enzyme extracts-The mycelial mat was harvested after 9 days, washed with ice-cold water, press dried and weighed . The mat was placed in a porcelain mortar and ground for 5 min with sea sand . Sodium acetate buffer (0.5 M : pH 5.1) was used for preparation of extract containing endocellular glucose oxidase . The extract was centrifuged at 500 g
392 INDIAN 1. EXP BIOL., APRIL 1999
for 10 min and again at 20,000 g for 10 min to remove the bulk of the sea sand and cellular debris and the supernatant fluid was used fop endocellular enzyme activity. The temperature was kept below 4°C throushout the operation .
Detf!~mination of calcium gluconate-At the end of fermentation, contents of the flasks were filtered and mycelium harvested were thoroughly dried at 80°C to a constant weight to measure the growth as per the method of Chaturvedi et al .. Culture filtrate was neutrali zed to pH 7.0 by a saturated solution of Ca(OH)2 and filtered. The filtrate was analysed for estimating calcium gluconate by EDT A titration 10,
a fter adding 10% NaOH solution with the help of Patton and Readers indicator. The end point was indicated by a faint blue colour. A blank titration for saturated Ca(OH)2 solution was also performed.
Calcium gluconate present in the broth was estimated from the data according to the calculation;
0.1 M EDT A == 0.04484 g of calcium gluconate.
Assay of glucose oxidase-The culture filtrate was used for exoce llul ar enzyme activity and the cell-free enzyme ex tract was used for endocellular activity. Both the activities were determined following the method of Bergmeyer", using peroxide coupled indicator reaction in the presence of a colour dye. The intensity of colour was measured colorimetrically as the dye O-dianisidine is oxidized to a highly colour product at Amax = 460 nm .
Effect of different carbon sources-Di fferent carbon sources were sterili zed and added aseptically to the rest o f the steri le basal medium in IS% concentration , (Nl-I4)2 S04 in 0.2S% concentration was added as the ll itrogen source. During the experiment ISO mI of fermenta tion medium was taken, in I L fl at flasks, inoculated with 7.S ml of cell suspension and incubated at 30°C for 9 days.
Results and Discussion Gl ucose ( 174.0 giL of calcium gluconat~; 7.0 gIL
of ce llular growth; 0.90a and 0.29b units/ml of glucose ox idase activity) was found better source of carbon than sucrose ( 106.0 giL of calcium gluconate; 7.4 giL
• exocc llul ar g lucose oxidase acti vity from 1.0 ml of culture fil trate. b endoccllular g lucose oxidase activity [1.0 gm wet weight eq ui valent to 0. 12 gm dry weight crushed, centrifuged and make a volume of 20 ml. From this volume, 1.0 ml was taken for the ac ti vi ty determination].
of cellular growth; 0.80' and 0.08b units/ml of glucose oxidase activity) and starch (47.0 g IL of calciw11 gluconate; 7.2 gIL of cellular growth; 0.09" and 0.03b
units/ml of glucose OXIdase activity) for calcium gluconate production and glucose oxidase activity.
There was no production of calcium gluconate and no glucose oxidase activity in the medium containing either of galactose, lactose, sodium acetate, sodium citrate, xylose and arabinose and hence no cell growth was detected. Glucose is a monosaccharide, directly oxidized to gluconic acid by A . . niger. Sucrose is a disaccharide, hydrolyzed to a monosaccharide glucose, which is then oxidized to gluconic acid. Since starch is a polysaccharide, it involves two step hydrolysis (starch ~ sucrose ~ glucose)before being oxidized to gluconic acid.
Determination of optimum concentration of glu
cose-As glucose was found to be the best carbon source for calcium gluconate production by A.niger AB 1801 , different concentrations of glucose were tested for calcium gluconate and glucose oxidase production.
Production of calcium gIuconate and glucose oxidase was observed at 10% of glucose concentration (110.2 giL; 0.20a and 0.09b units/mIl which was optimum at IS% of glucose (0.90a and 0.29t. units/ml ; 174.0 gi l), however, higher concentration of glucose (16%) showed inhibitory effect on these contents (0 .8S' and O.ISb units/ml ; 160.0 giL). Cellular growth was not vigorous at 8% (6.0 gIL) and 10% (6.2 giL) of glucose solution. Earlier reports have also suggested that IS% glucose concentration gives the maximum amount of calcium gluconate7
.,z
.:J.
Effect of different nitrogen sources-Different nitrogen sources were examined for their effect on efficient fermentation· by A. niger AB 1801. The basal medium supplemented with IS% glucose was used as the carbon source for studying the effect of different nitrogen sources. To this medium was added a given nitrogen source at a concentration equivalent to 0.OS3 g nitrogen (N) per 100 m!.
Urea was observed as a superior nitrogen source (17S.S gIL; 1.20' and 0.40b units/rnl) for calcium gluconate and glucose oxidase production by A. niger AB 1801. Other sources of nitrogen taken for the present study were moderate stimulators, but ammonium sulphate(174.0 gIL; 0.90' and 0 .09b units/ml) and ammonium phosphate (1S1.0 giL ; 0.70' and 0.12b units/ml) were inhibitory to some extent. Previous
RA Y & BANIK: EFFECf OF AMMONIUM AND NITRATE RATIO ON GLUCOSE OXIDASE 393
• .... ~ 7 ~200
01 ... - c -- 0
190 01 ... U
~ j ... "0
~ 0
D ~ 180 ... '" 0\6 +' 0 ... c
~ e 170 ~ .3 -;; (!)
u ~ 160 u C u
150 5 ~
003 0·04 0·05 0·06 0-{)7 G·l0 . 0·1'5 0·20 I
M9504 ' 7H20 Conl:e'ntration(%l
-
0'30
o
I 1-3 ~
o )(
~ >-
1-1 -> -;::. u o
0·9 ~ o "0 )(
o 0·7 '"
III o U j
0-5 2i
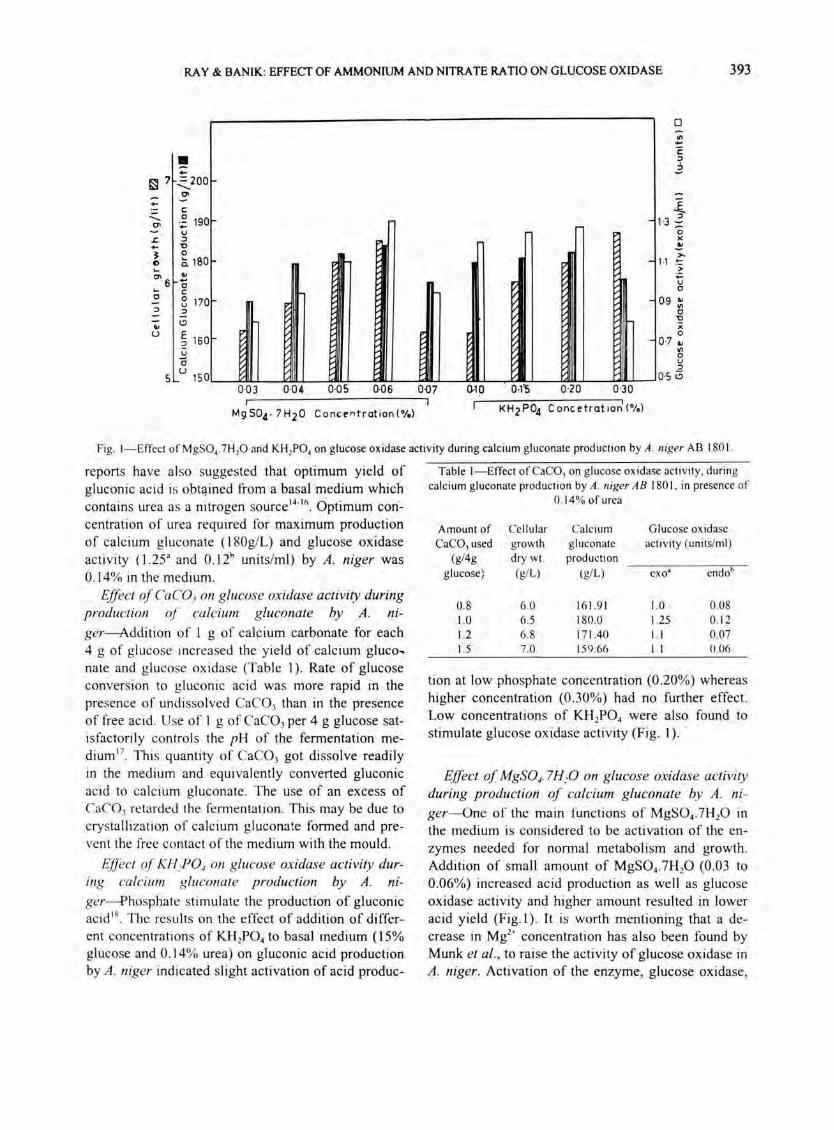
Fig. I-Effect of MgSO •. 7Hp and KHIPO. on glucose oxidase activity during calcium gluconate production by A. niger AS 180 I.
reports have also suggested that optimum yield of gluconic acid is obt~ined from a basal medium which contains urea as a nitrogen source I4
-16
. Optimum con'centration of urea required for maximum production of calcium gluconate (180g/L) and glucose oxidase activity (1.25" and 0. 12b units/ml) by A. niger was 0.14% in the medium.
Effect oj CaCO ! on glucose oxidase activity during production oj calcium gluconate by A. ni
ger--Addition of I g of calcium carbonate for each 4 g of glucose increased the yield of calcium gluco .. nate and glucose oxidase (Table I) . Rate of glucose conversion to gluconic acid was more rapid in the presence of undissolved CaC03 than in the presence of free acid. Use of 1 g of CaC03 per 4 g glucose satisfactorily controls the pH of the fermentation medium 17. This quantity of CaC03 got dissolve readily in the medium and equivalently converted gluconic acid to calcium gluconate. The use of an excess of CaC0 3 retarded the fermentation . Thi s may be due to crystallization of calcium gluconate formed and prevent the free contact of the medium with the mould.
Effect oj KHPO~ on glucose oxidase activity during calcium gluconate production by A. ni
ger--Phosphate stimulate the production of gluconic acid l8
. The results on the effect of addition of different concentrations of KH ZP04 to basal medium (15% glucose and 0.14% urea) on gluconic acid production by A. niger indicated slight activation of acid produc-
Table I-Effect of CaCO] on glucose oxidase activity, during calcium gluconate production by A. niger AB 180 I , in presence of
0.14% of urea
Amount of Cellul ar Calcium Glucose oxidase CaCO] used growth gluconate activity (unitslml )
(g/4g dry wI. production g lucose) (giL) (giL) exo' endob
tion at low phosphate concentration (0 .20%) whereas higher concentration (0 .30%) had no further effect. Low concentrations of KH ZP04 were also found to stimulate glucose oxidase activity (Fig. I) . .
Effect of MgS04. 7H!O on glucose ox·idase activity during production oj calcium gluconate by A. ni
ger-One of the main functions of MgS04.7HzO in the medium is considered to be activation of the enzymes needed for normal metabolism and growth. Addition of small amount of MgS04 .7H zO (0.03 to 0.06%) increased acid production as well as glucose oxidase activity and higher amount resulted i,n lower acid yield (Fig.l) . It is worth mentioning that a decrease in Mgz+ concentration has also been found by Munk et al., to raise the activity of glucose oxidase in A. niger. Activation of the enzyme, glucose oxidase,
394 INDIAN 1. EXP BIOL., APRIL 1999
z o ~
§ a:: Cl.
400 ____ 1'Sg/L
-6-2 'Sg/L
-0-1 ' og/L
A
w~ ~ ..... . ~:::'300 UOI 3~ t:> ~ ::> U ...J « U
> ... u o
'" III o "0
x o
'" III o U OJ
t:> o '~5----~----~-----~~--~~--~LJ
Concn of Glucose(%)
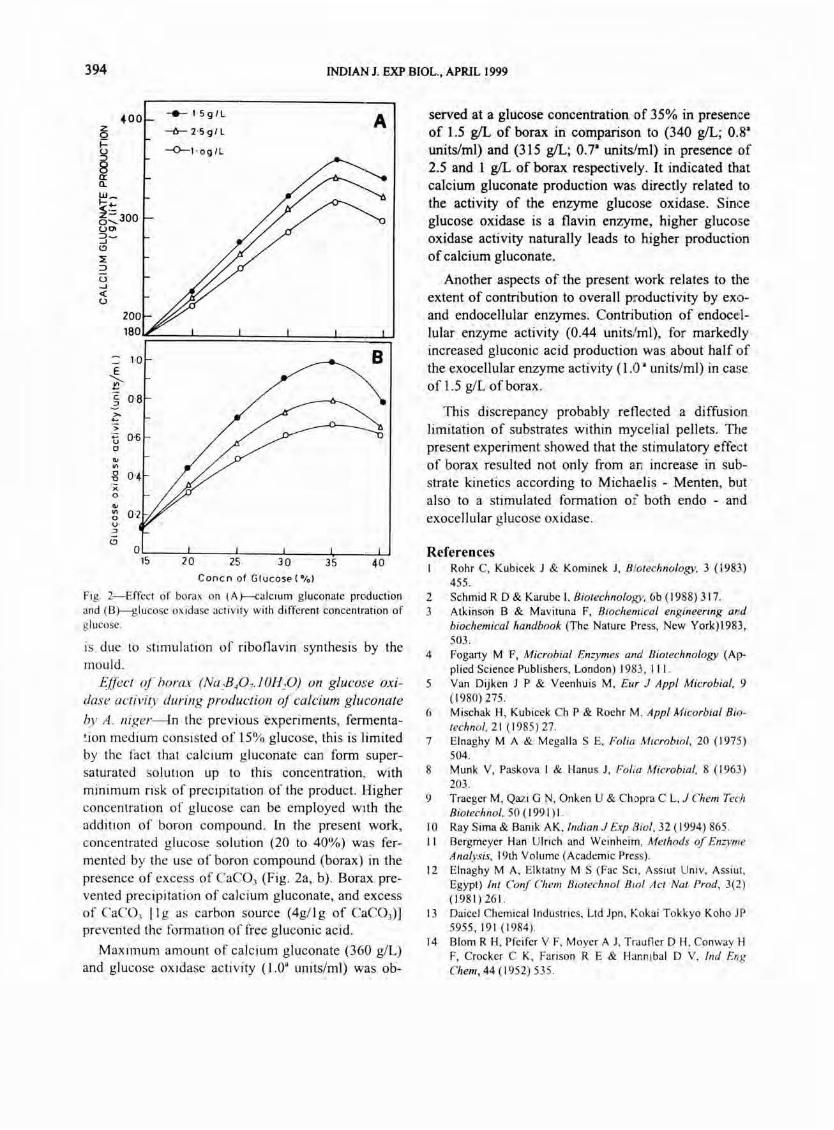
Fi g. 2-Effect of borax on (A}-calcium gluconate production and (8 )--g lu cosc oxidase a'ctivity with different concentration of g lucose.
is due to stimulation of riboflavin synthesis by the mould.
Effect of horax (Na!B40 7·JOH!O) on glucose oxidase activity during production of calcium gluconate
by A. niger---ln the previous experiments, fermenta!ion medium consisted of 15% glucose, this is limited by the fact that calcium gluconate can form supersaturated solution up to this concentration, with minimum risk of precipitation of the product. Higher concentration of glucose can be employed with the addition of boron compound. In the present work, concentrated glucose solution (20 to 40%) was fermented by the use of boron compound (borax) in the presence of excess of CaC03 (Fig. 2a, b). Borax prevented precipitation of calcium gluconate, and excess of CaCO, [1 g as carbon source (4g/ 1 g of CaCOJ ) ]
prevented the formation of free gluconic acid.
Maximum amount of calcium gluconate (360 giL) and glucose oxidase activity (1.0a units/ml) was oh-
served at a glucose concentration of 35% in presence of 1.5 gIL of borax in comparison to (340 gIL; 0.8' units/ml) and (315 gIL; 0.7' units/ml) in presence of 2.5 and 1 gIL of borax respectively. It indicated that calcium gluconate production was directly related to the activity of the enzyme glucose oxidase. Since glucose oxidase is a flavin enzyme, higher glucose oxidase activity naturally leads to higher production of calcium gluconate.
Another aspects of the present work relates to the extent of contribution to overall productivity by exoand endocellular enzymes. Contribution of endoce:llular enzyme activity (0.44 units/ml), for markedly increased gluconic acid production was about half of the exocellular enzyme activity (1 .0' units/ml) in case of 1.5 giL of borax.
This discrepancy probably reflected a diffusion limitation of substrates within mycelial pellets. TIle present experiment showed that the stimulatory effect of borax resulted not only from an increase in substrate kinetics according to Michaelis - Menten, but also to a stimulated formation of both endo - and exocellular glucose oxidase.
14 810m R H, Pfeifer V F, Moyer A J, Traut1er D H, Conway H F, Crocker C K, Farison R E & Hanni bal D V, In d Eng Chem, 44 ( 1952) 535.
RAY & I3A NIK EFFECT OF AMMONIUM AND NITRATE RATIO ON GLUCOSE OXIDASE
15 Jaksa I, Gyuran J, Nemeth S, Trischler F, Udvardy Nagy I Mrs, (R ichter Gedeon Vegyeszeti Gyar Rt) Hung Teljes HU 28, (I ')8:l ) 770 .
I (i Shah D N & Kothari R M, Biotech Let! , 15( I) (1993) 35 . 17 MandaI S K & Chatterjee S P, Res alld IlId, 31 (1986) 52 . 18 Martin S M & Steel R. ('an J Microbiol, I (1954) 470.









![Aluminum tris(dihydrogen phosphate) [Al(H PO ...ipcbee.com/vol6/no2/84-F30013.pdf · Aluminum tris(dihydrogen phosphate) [Al(H2PO4)3]: environmentally friendly catalyst for the preparation](https://static.documents.pub/doc/80x56/5c943a8f09d3f2c2238c0fe8/aluminum-trisdihydrogen-phosphate-alh-po-aluminum-trisdihydrogen-phosphate.jpg)









