Effect of human chorionic gonadotropin on luteal function and reproductive performance of high-producing lactating Holstein dairy cows 1,2 J. E. P. Santos* 3 , W. W. Thatcher†, L. Pool*, and M. W. Overton* *Veterinary Medicine Teaching and Research Center, University of California-Davis, Tulare 93274 and †Department of Animal Sciences, University of Florida, Gainesville 32611 ABSTRACT: The objectives were to evaluate effects of human chorionic gonadotropin (hCG) (3,300 IU i.m.) administered on d 5 after AI on CL number, plasma progesterone concentration, conception rate, and preg- nancy loss in high-producing dairy cows. Following the synchronization of estrus and AI, 406 cows were in- jected with either hCG or saline on d 5 after AI in a randomized complete block design. Blood sampling and ovarian ultrasonography were conducted once between d 11 and 16 after AI. Pregnancy diagnoses were per- formed on d 28 by ultrasonography and on d 45 and 90 after AI by rectal palpation. Treatment with hCG on d 5 resulted in 86.2% of the cows with more than one CL Key Words: hCG, Conception Rate, Dairy Cows, Pregnancy, Progesterone 2001 American Society of Animal Science. All rights reserved. J. Anim. Sci. 2001. 79:2881–2894 Introduction Human chorionic gonadotropin has an activity simi- lar to that of LH and binds to LH receptors on the membrane of small luteal cells to activate a second messenger, which enhances progesterone synthesis. Corpus luteum formation and development is highly responsive to LH, and small luteal cells synthesize more progesterone when stimulated by LH (Hoyer and Nis- wender, 1985). The administration of hCG during the early luteal phase induces ovulation of the first-wave 1 The authors wish to thank David Frea and the staff of Souza Dairy (Fresno, CA) for their assistance during this project. We want to express our appreciation to Pat Sweeney and Thomas Overbay of Forth Dodge Animal Health, Inc., and to Pharmacia Upjohn Company for generously providing Factrel and Lutalyse, respectively. We are grateful to Marie-Joelle Thatcher for running the progesterone assays and to Cristiana Santos, William Sischo, and Rob Atwill for their comments and suggestions during data analyses. 2 This is Journal Series No. R-07799 of the Florida Agric. Exp. Sta. Research was supported partially by the Binational Agricultural Research and Development grant no. IS-3000-98R and the Florida Milk Checkoff Program. 3 Correspondence: 18830 Road 112 (phone: 559/688-1731; fax:559/ 686-4231; E-mail: [email protected]). Received September 29, 2000. Accepted May 15, 2001. 2881 compared with 23.2% in controls. Plasma progesterone concentrations were increased by 5.0 ng/mL in hCG- treated cows. The presence of more than one CL in- creased progesterone concentration in hCG-treated cows but not in controls. Conception rates were higher for hCG-treated cows on d 28 (45.8 > 38.7%), 45 (40.4 > 36.3%), and 90 (38.4 > 31.9%) after AI. Treatment with hCG improved conception rate in cows losing body condition between AI and d 28 after AI. Pregnancy losses were similar between treatment groups. Treat- ment with hCG on d 5 after AI induces accessory CL, enhances plasma progesterone concentration, and im- proves conception rate of high-producing dairy cows. dominant follicle and formation of a functional acces- sory CL. Schmitt et al. (1996a) observed that most of the increase of progesterone production after hCG injection was due to the formation of accessory CL. Furthermore, cows treated with hCG on d 5 after estrus had a greater increase in plasma progesterone from d 6 to 13 and greater circulating progesterone on d 13 of the estrous cycle. Therefore, it is possible that hCG induction of an accessory CL on d 5 of the estrous cycle may increase plasma progesterone and enhance embryo survival, which would improve conception rates in dairy cows. In addition, hCG treatment increases the occurrence of three-wave follicular cycles, in which the emergence of the third-wave dominant follicle is delayed (Diaz et al., 1998). Such an alteration in follicular dynamics may enhance conception rate because a greater number of cows with three follicular waves after insemination con- ceived compared with cows having two follicular waves (Ahmad et al., 1997). The objectives of the current study were to evaluate the effect of injecting 3,300 IU of hCG i.m. on d 5 after AI on CL number and plasma progesterone concentra- tion. In addition, effects of hCG on conception rate at 28, 45, and 90 d after AI were evaluated to determine whether CL number and plasma progesterone after AI impacted the conception rate of dairy cows. Lastly, be- cause the injection of hCG may affect follicular turnover
Transcript
Effect of human chorionic gonadotropin on luteal function and reproductiveperformance of high-producing lactating Holstein dairy cows1,2
J. E. P. Santos*3, W. W. Thatcher†, L. Pool*, and M. W. Overton*
*Veterinary Medicine Teaching and Research Center, University of California-Davis, Tulare 93274 and†Department of Animal Sciences, University of Florida, Gainesville 32611
ABSTRACT: The objectives were to evaluate effectsof human chorionic gonadotropin (hCG) (3,300 IU i.m.)administered on d 5 after AI on CL number, plasmaprogesterone concentration, conception rate, and preg-nancy loss in high-producing dairy cows. Following thesynchronization of estrus and AI, 406 cows were in-jected with either hCG or saline on d 5 after AI in arandomized complete block design. Blood sampling andovarian ultrasonography were conducted once betweend 11 and 16 after AI. Pregnancy diagnoses were per-formed on d 28 by ultrasonography and on d 45 and 90after AI by rectal palpation. Treatment with hCG on d5 resulted in 86.2% of the cows with more than one CL
2001 American Society of Animal Science. All rights reserved. J. Anim. Sci. 2001. 79:2881–2894
Introduction
Human chorionic gonadotropin has an activity simi-lar to that of LH and binds to LH receptors on themembrane of small luteal cells to activate a secondmessenger, which enhances progesterone synthesis.Corpus luteum formation and development is highlyresponsive to LH, and small luteal cells synthesize moreprogesterone when stimulated by LH (Hoyer and Nis-wender, 1985). The administration of hCG during theearly luteal phase induces ovulation of the first-wave
1The authors wish to thank David Frea and the staff of SouzaDairy (Fresno, CA) for their assistance during this project. We wantto express our appreciation to Pat Sweeney and Thomas Overbay ofForth Dodge Animal Health, Inc., and to Pharmacia Upjohn Companyfor generously providing Factrel and Lutalyse, respectively. We aregrateful to Marie-Joelle Thatcher for running the progesterone assaysand to Cristiana Santos, William Sischo, and Rob Atwill for theircomments and suggestions during data analyses.
2This is Journal Series No. R-07799 of the Florida Agric. Exp.Sta. Research was supported partially by the Binational AgriculturalResearch and Development grant no. IS-3000-98R and the FloridaMilk Checkoff Program.
Received September 29, 2000.Accepted May 15, 2001.
2881
compared with 23.2% in controls. Plasma progesteroneconcentrations were increased by 5.0 ng/mL in hCG-treated cows. The presence of more than one CL in-creased progesterone concentration in hCG-treatedcows but not in controls. Conception rates were higherfor hCG-treated cows on d 28 (45.8 > 38.7%), 45 (40.4> 36.3%), and 90 (38.4 > 31.9%) after AI. Treatmentwith hCG improved conception rate in cows losing bodycondition between AI and d 28 after AI. Pregnancylosses were similar between treatment groups. Treat-ment with hCG on d 5 after AI induces accessory CL,enhances plasma progesterone concentration, and im-proves conception rate of high-producing dairy cows.
dominant follicle and formation of a functional acces-sory CL. Schmitt et al. (1996a) observed that most of theincrease of progesterone production after hCG injectionwas due to the formation of accessory CL. Furthermore,cows treated with hCG on d 5 after estrus had a greaterincrease in plasma progesterone from d 6 to 13 andgreater circulating progesterone on d 13 of the estrouscycle. Therefore, it is possible that hCG induction of anaccessory CL on d 5 of the estrous cycle may increaseplasma progesterone and enhance embryo survival,which would improve conception rates in dairy cows.
In addition, hCG treatment increases the occurrenceof three-wave follicular cycles, in which the emergenceof the third-wave dominant follicle is delayed (Diaz etal., 1998). Such an alteration in follicular dynamics mayenhance conception rate because a greater number ofcows with three follicular waves after insemination con-ceived compared with cows having two follicular waves(Ahmad et al., 1997).
The objectives of the current study were to evaluatethe effect of injecting 3,300 IU of hCG i.m. on d 5 afterAI on CL number and plasma progesterone concentra-tion. In addition, effects of hCG on conception rate at28, 45, and 90 d after AI were evaluated to determinewhether CL number and plasma progesterone after AIimpacted the conception rate of dairy cows. Lastly, be-cause the injection of hCG may affect follicular turnover
Santos et al.2882
and promote more three-wave cycles, the potential im-pact of hCG treatment on conception rate was evaluatedduring the subsequent estrous cycle for those cows thatwere open after the first AI.
Materials and Methods
The experiment was conducted on a commercial dairyfarm in south Central Valley of California (Souza Dairy,Fresno, CA). Four hundred and six high-producing(43.0 ± 0.45 kg/d, mean milk ± SE at the time of AI)Holstein dairy cows were housed in free-stall barns andmilked twice daily. Multiparous and primiparous cowswere housed separately. Cows were fed a total mixedration (Table 1) throughout the experiment twice dailyfor ad libitum consumption to meet or exceed the re-quirements for a lactating cow that weighs 650 kg andproduces 45 kg of milk with 3.5% fat (NRC, 1989).
The study was conducted during two periods in 1999and 2000. The first period was from May 14 to Septem-ber 16, 1999, which coincided with the hot months of theyear, when the average daily maximum temperatureswere 28.8, 30.8, 33.8, 33.1, and 32.6°C, in May, June,July, August, and September, respectively (referred toherein as the warm period). The second period coincidedwith the cold months of the year, from October 5, 1999
Table 1. Dietary ingredients and nutrientcomposition of diet
aCalcium salts of long-chain fatty acids; EnerGII (calcium salts ofpalm fatty acids; Bioproducts, Inc., Fairlawn, OH).
bContent: 2.8% Ca, 3.5% P, 1.0% K, 0.45% S, 22% Na, 2.0% CL,and (per kg) 1,800 mg of Zn, 420 mg of Cu, 1,950 mg of Mn, 14 mgof Se, 6 mg of Co, 29 mg of I, 240,000 IU of vitamin A, 51,000 IU ofvitamin D, 1,800 IU of vitamin E.
cCalculated based on NRC (1989).dNFC = Nonfibrous carbohydrates [NFC = DM − (CP + NDF + EE
+ ash).
until March 2, 2000, when the average daily maximumtemperatures were 28.6, 20.6, 15.1, 14.9, 16.8, and19.6°C, in October, November, December, January,February, and March, respectively (cool period) (Han-ford Municipal Airport, Official Certified Local Climato-logical Data). One hundred and eighty-two cows wereenrolled during the warm period and 224 cows duringthe cool period.
Once a week, a group of 30 to 60 cows at between 40and 145 d in milk had their estrous cycle synchronized(Thatcher et al., 1989) with an i.m. injection of 100�g of GnRH (gonadorelin; Factrel, Fort Dodge AnimalHealth Inc., Fort Dodge, IA) followed 7 d later by an i.m.injection of 25 mg of PGF2α (dinoprost tromethamine;Lutalyse, Pharmacia Upjohn Company, Kalamazoo,MI). Estrus was detected once daily, from 2 to 5 d afterthe PGF2α injection, both visually and by tail chalking(Macmillan et al., 1988) using paintstiks (All-weatherPaintstick, LA-CO Industries, Chicago, IL). Cows foundin estrus were assigned to initiate the experiment basedon lactation number and days in milk. Utilizing theprotocol described, we were able to detect in estrus 72%of the cows enrolled in the synchronization program.Therefore, about 20 to 40 cows were enrolled in thestudy every week. All cows detected in estrus were arti-ficially inseminated once, in the morning, by the sametechnician throughout the experimental period. Semenfrom five proven sires was used, and a similar numberof inseminations within each sire was used for eachtreatment group. Cows found open at d 28 after theinitial AI received an injection of 25 mg of PGF2α andwere artificially inseminated as they were detected inestrus within 5 d of the PGF2α treatment. For analysesof second-service conception rate, only cows insemi-nated in the subsequent estrus after the initial AI (18to 24 d after the initial AI) and those that came intoestrus after the PGF2α injection at d 28 were used.
On d 5 after the initial AI, cows received an i.m.injection in the neck area of either 3,300 IU of hCG(Chorulon, INTERVET, Inc., Millsboro, DE) (hCGgroup) or 3 mL of saline solution (control group).
One blood sample (7 mL) was collected from all cowsbetween d 11 and 16 after AI by coccygeal venipuncture,using Vacutainer tubes (Becton & Dickinson Vacu-tainer Systems, Rutherford, NJ) with sodium EDTA.The samples were placed immediately on ice and latercentrifuged at 2,000 × g for 20 min for separation ofplasma. Plasma samples were frozen at −75°C untillater analysis. Progesterone was analyzed by radioim-munoassay according to Knickerbocker et al. (1986). Atthe same time as blood sampling, the ovaries of all cowswere scanned by ultrasound (Ultrascan 50, AllianceMedical, Smithville, MO) equipped with a 5.0-MHz lin-ear transducer to evaluate presence, location (ovary),size (area and volume), and number of corpora lutea(Ginther, 1998). The location of the CL in cows withmultiple ovulations that were diagnosed as carrying asingle pregnancy on d 28 (contralateral or ipsilateralovary to the horn of pregnancy) was determined, and
Human chorionic gonadotropin for dairy cows 2883
the maintenance or regression of accessory CL waslater evaluated.
The diagnosis of pregnancy was performed by ultra-sonography on d 28 ± 1 after AI. During ultrasonogra-phy, a cow was determined pregnant when an embry-onic vesicle with a viable embryo (presence of heartbeat)was detected. At the same time, ovaries of pregnantcows were scanned to determine the presence and loca-tion of the CL. Pregnant cows at d 28 were reconfirmedfor pregnancy by rectal palpation on d 45 ± 1 and againon d 90 ± 2 after AI.
At the time of AI and on d 28 ± 1, 45 ± 1, and 90 ±2 after AI, the same person evaluated the cows’ bodycondition score (BCS) (Ferguson et al., 1994). Previouswork at the University of Florida (Moreira et al., 2000)observed that cows with a BCS lower than 2.5 re-sponded poorly to a timed AI protocol and that concep-tion rates were low. To determine the effects of BCS atbreeding on the outcome variable, we divided cows intotwo groups, those with BCS at AI equal to or lowerthan 2.75 and those with a score greater than 2.75. ABCS of 2.75 was chosen to divide the two groups becauseonly a few cows in this study had a BCS at AI equal toor lower than 2.5. For BCS change from AI to pregnancydiagnosis, we divided cows into three groups: those thatlost BCS, had no change, and those that gained BCSfrom AI to pregnancy diagnosis. The mean (± SEM)change in BCS for cows that lost and gained BCS were−0.28 (± 0.008) and 0.30 (± 0.005), respectively. Milkproduction from the test date closest to the day of AI(day of AI ± 10 d) was divided into four quartiles or intotwo groups (above or below the mean, 43.0 kg/d) andwere used to test the effects of milk yield on the outcomevariables analyzed. The ranges in milk production forthe first, second, third, and fourth quartiles were 20.4to 36.3 kg/d, 36.3 to 41.7 kg/d, 41.7 to 49.9 kg/d, and49.9 to 69.0 kg/d, respectively.
Experimental Design and Statistical Analysis
The experimental design was a randomized completeblock design (Kuhel, 1994). Cows were blocked ac-cording to lactation number and days in milk, and thenrandomly assigned to one of the two treatments on aweekly basis. Continuous data were analyzed by theGLM procedure of the SAS (SAS Inst. Inc., Cary, NC)program using the following linear model:
Yijklm = µ + Ti + PAj + PEk + BCl + Mm + (T × PA)ij
PAj = parity effectPEk = period effectBCl = body condition effect
(T × PA)ij = interaction between Ti and PAj(T × PE)ik = interaction between Ti and PEk(T × BC)il = interaction between Ti and BCl(T × M)im = interaction between Ti and Mm
(T × PA × PE)ijk = interaction between Ti and PAj andPEk
(T × PA × BC)ijl = interaction between Ti and PAj andBCl
(T × PA × M)ijm = interaction between Ti and PAj andMm
(T × PE × BC)ikl = interaction between Ti and PEk andBCl
(T × PE × M)ikm = interaction between Ti and PEk andMm
(T × BC × M)ilm = interaction between Ti and BCl andMm
Bm = block effectsCn = covariate effect
Eijklmn = residual error
When the results of plasma progesterone and numberof CL were analyzed, the number of days after AI whenthe blood sample was collected and ultrasonographyperformed was used for covariate adjustment of thedata. The effect of number of CL on plasma progester-one concentrations was tested. Interactions that werenot significant (P > 0.15) were excluded from the model.
Body condition scores after AI in association withpregnancy statuses were analyzed as analysis of vari-ance for repeated measures using the PROC MIXEDprocedure of SAS (Littell et al., 1998). The model in-cluded observed mean, block, pregnancy status (preg-nant or open), the random experimental error of cowwithin pregnancy status, time when BCS data werecollected, interaction between pregnancy status andtime, and the random experimental error.
Regression analyses were performed to determine thefitted-line plot for total luteal area and plasma proges-terone concentrations (MINITAB, 1996). A simple lin-ear regression analysis was conducted to determine thebest subset regression that predicts plasma progester-one concentrations.
Nonnormally distributed data such as conceptionrate, pregnancy loss, and CL regression were analyzedusing the TLOGISTIC procedure of SAS (SAS, Inst.Inc.). A stepwise regression model was used (Hosmerand Lemeshow, 1989). The model statement in the lo-gistic regression analysis was specified that a variablehad to be significant at the 0.30 level before it could beentered into the model, whereas variables that enteredthe model had to be significant at the 0.20 level in orderfor them to remain in the model, and variables wereentered or removed by the Wald statistic criterion(Hosmer and Lemeshow, 1989). The full logistic regres-sion model included block, treatment, parity, period,number of CL, plasma progesterone, milk production,
Santos et al.2884
Table 2. Effect of treatment on plasma progesterone concentration and corpora lutea number at d 14 after AI
Treatmenta P <b
Item Control-P Control-M hCG-P hCG-M SEM TRT Parity TRT × Parity
aP = primiparous, M = Multiparous.bTRT = treatment, TRT × Parity = interaction between treatment and parity.cCL = corpus luteum.
BCS at breeding, BCS change from AI to pregnancydiagnosis, and the respective two- and three-way inter-actions. For the analysis of CL regression, only thosecows diagnosed pregnant with multiple CL were in-cluded. In these analyses, location of the multiple CL(contralateral or ipsilateral) relative to the pregnanthorn was included in the logistic regression model.
The relative risk and the 95% confidence interval fora cow to have one or more than one CL were determinedusing the frequency distribution of CL for control andhCG-treated cows, and significance was determined bychi-square (Dean et al., 1991). Treatment differenceswith P < 0.05 were considered significant and P < 0.10were considered a tendency.
Results
Days after AI when blood samples were collected andultrasonography was performed did not differ betweentreatments and averaged 13.8 d (P = 0.15). Administer-ing 3,300 IU of hCG on d 5 after AI resulted in 175 ofthe 203 treated cows with more than one CL. Cowstreated with hCG had a six times greater relative riskof having more than 1 CL than controls (RR = 6.03;95% CI: 4.1 < RR < 8.8; P < 0.001). More cows in thehCG-treated group had multiple CL (86.2% vs 23.2%;P < 0.001), which reflected the increase in mean numberof CL per cow (P < 0.0001; Table 2). The number of cowswith no detectable CL at the ultrasonography on d 14was smaller for the hCG-treated cows than for the con-trol group (1.5% vs 6.9%; P = 0.006). An interactionbetween treatment and period was observed for numberof CL. The increase in number of CL for cows treatedwith hCG compared with control cows was greater dur-ing the cool (1.8 vs 2.4) than during the warm (1.0 vs1.3) period (P = 0.02). Number of CL on d 14 was notinfluenced by BCS at breeding. However, the changein BCS between AI and d 28 after insemination tendedto affect CL number (P = 0.07), with cows that lost BCShaving the highest mean number of CL (1.9), followedby those whose BCS did not change (1.6) or those thatgained BCS (1.5) in the same period. Milk productionanalyzed either as a quartile or as two groups, aboveor below the mean milk, had no effect on number of
CL, and no interaction was observed for treatment andmilk yield on CL number. Treatment affected the sur-face area and volume of the total luteal tissue (P =0.001), as well as the largest CL. However, most of theeffect of treatment on total luteal area and volume wascaused by the presence of multiple CL. When the num-ber of CL was included as a covariate in the analysesof luteal area and volume, treatment effect on totalluteal area and volume was eliminated (P = 0.24).
Plasma progesterone concentrations during mid lu-teal phase were increased by 5.0 ng/mL in the hCG-treated cows compared with controls (P < 0.001). Differ-ences in progesterone concentrations between hCG andcontrol cows were +8.7 ng/mL for primiparous cows and+1.2 ng/mL for multiparous cows (P = 0.02). Cows withmore observable CL during ultrasonography had higherprogesterone concentrations (P < 0.001), and an interac-tion between CL number and treatment on plasma pro-gesterone was observed (P < 0.001). Within the hCGgroup, cows with more than one CL had higher plasmaprogesterone than those with only one CL (20.3 vs 10.9ng/mL), an increase of 9.4 ng/mL (Figure 1). Such aneffect of additional CL on progesterone concentrationswas not observed for cows in the control group, in whichprogesterone concentrations were similar (14.9 vs 14.6ng/mL). Body condition score at AI had a tendency toimpact plasma progesterone (P < 0.10). Progesteroneconcentration in cows with low BCS (≤ 2.75) was 1.7ng/mL lower than in those with moderate BCS (≥ 3.0),and an interaction between BCS at AI and treatmentwas observed. Within the hCG-treated cows, BCS atAI (moderate vs low BCS) had no impact on plasmaprogesterone concentrations (18.8 vs 18.9 ng/mL), but,in the control group, cows with moderate BCS had 3.4ng/mL higher progesterone concentration in plasmathan those with low BCS (15.5 vs 12.2 ng/mL; P = 0.05).Changes in BCS from breeding to d 28 did not affectplasma concentrations of progesterone, but milk pro-duction did. When progesterone concentrations of cowswith production above or below 43.0 kg/d were com-pared, higher producing cows tended to have lower pro-gesterone concentrations (17.4 vs 15.2 ng/mL), but thiseffect was observed only for cows in the control group(16.3 > 11.5 ng/mL vs 18.6 < 19.0 ng/ml; P = 0.06).
Human chorionic gonadotropin for dairy cows 2885
Figure 1. Plasma progesterone concentrations in cows with 0, 1, or more than 1 detectable corpus luteum (CL) atultrasound on d 14 after AI. Treatment with 3,300 IU of hCG i.m. was administered on d 5 after AI. Solid black bars,control; white bars with diagonal shading, hCG. The number of CL increased plasma progesterone concentrations (P< 0.001) and an interaction between treatment and CL was observed for progesterone concentrations (P < 0.001). Onecow in the hCG-treated group had a high progesterone concentration, but no detectable CL was observed duringultrasonography of the ovaries. The SEM for progesterone concentrations in control and hCG-treated cows were asfollows: 0 CL, 2.14 and 4.43; 1 CL, 0.61 and 1.51; and 2 CL, 1.21 and 0.62.
Treatment with hCG increased conception rate on d28 after AI (P < 0.01; Table 3). Similar conception rateswere observed during the warm and the cool periods,but a tendency for an interaction between treatment
Table 3. Effect of treatment and number of corpora lutea on d 14 on conception rates(%) at d 28, 45, and 90 after AI for cows receiving hCG or saline on d 5 after AI
(number of pregnant/total number in each group is indicated in parenthesis)
aCL = corpus luteum; TRT × CL = interaction between treatment and number of corpora lutea.
and period was detected (P = 0.09). Control and hCG-treated cows had similar conception rates during thewarm period (44.1 vs 43.3%), but hCG increased concep-tion rate compared with control during the cool period
Santos et al.2886
Table 4. Plasma progesterone (ng/mL) concentrations (± SEM) at d 14 after AI incontrol and hCG-treated cows diagnosed open or pregnant at d 28, 45 or 90 after AI
Pregnancy status
Open Pregnant P <a
Day after AI Control hCG Control hCG PS TRT TRT × PS
aPS = pregnancy status, TRT = treatment; TRT × PS = interaction between treatment and pregnancystatus.
(47.8 vs 34.2%). Cows with more CL had higher concep-tion rates at d 28 (P < 0.001; Table 3). Nevertheless,no treatment × CL interaction was detected (P = 0.34).Although conception rate was numerically higher forcontrol than for hCG cows with more than one CL,the number of control cows with more than one CLrepresented only 23.2% of that group compared with86.2% of the hCG group. Progesterone concentration atd 14 influenced conception rate on d 28 (P = 0.007).Plasma progesterone in pregnant cows was 3.1 ng/mLhigher than in cows that were open at d 28 (P < 0.001;Table 4), and this pregnancy effect was observed forcontrol, as well as hCG-treated cows. Similar effects ofprogesterone were observed for conception rate at 45and 90 d after AI.
When milk production was divided into quartiles,cows with higher production tended to have lower con-
Figure 2. Effect of milk production (quartiles) at the test date closest to the day of AI on conception rates (CR) ofdairy cows on d 28, 45, and 90 after AI. Solid black bars, first quartile (20 to 35.9 kg/d); white bars with horizontalshading, second quartile (36 to 41.9 kg/d); white bars, third quartile (42 to 49.9 kg/d); white bars with vertical shading,fourth quartile (50 to 69 kg/d). Milk production affected conception rate on d 28 (P < 0.09) and 45 (P < 0.03) but hadno effect on d 90 (P > 0.15). No interaction between treatment and milk production was observed for conception ratein any of the days after AI when pregnancy was diagnosed.
ception rates (P = 0.09). Conception rates decreasedfrom 53.4% to 41.4, 38.8, and 35.9% for the lowest,second, third, and highest quartiles of milk yield (Fig-ure 2).
Body condition score also affected conception rate atd 28. Cows with moderate BCS at the time of AI hadhigher conception rates than cows with low BCS (48.7vs 34.4%; P < 0.001), and this effect was similar forcontrol and hCG-treated cows (Figure 3). Similar to theresults observed for BCS at the day of AI, changes inBCS from AI to d 28 also were associated with changesin conception rate. Cows that gained BCS from AI to d28 had higher pregnancy than those that maintainedor lost BCS (47.0 vs 37.4 and 42.7%; P = 0.03). Interest-ingly, an interaction between treatment and BCSchange was determined for pregnancy at d 28 (Figure4). Cows that received lower BCS when treated with
Human chorionic gonadotropin for dairy cows 2887
Figure 3. Effect of body condition score (BCS) at the time of AI on conception rates (CR) of control and hCG-treatedcows on d 28, 45, and 90 after AI. Cows were divided into two groups, those with BCS equal to or lower than 2.75(black solid bars) and those with a BCS greater than 2.75 (white bars with diagonal shading). Body condition scoreat AI affected conception rate (P < 0.0001), but no interaction between treatment and BCS was observed for conceptionrate (P > 0.15).
Figure 4. Effect of body condition score (BCS) changes from AI to d 28 on conception rate (CR) of control and hCG-treated cows at d 28 after AI. Treatment with 3,300 IU of hCG i.m. was administered on d 5 after AI. Solid black bars,control; white bars with diagonal shading, hCG. Body condition score change affected conception rate (P < 0.03), andan interaction between treatment and BCS change was observed for conception rate (P < 0.05).
Santos et al.2888
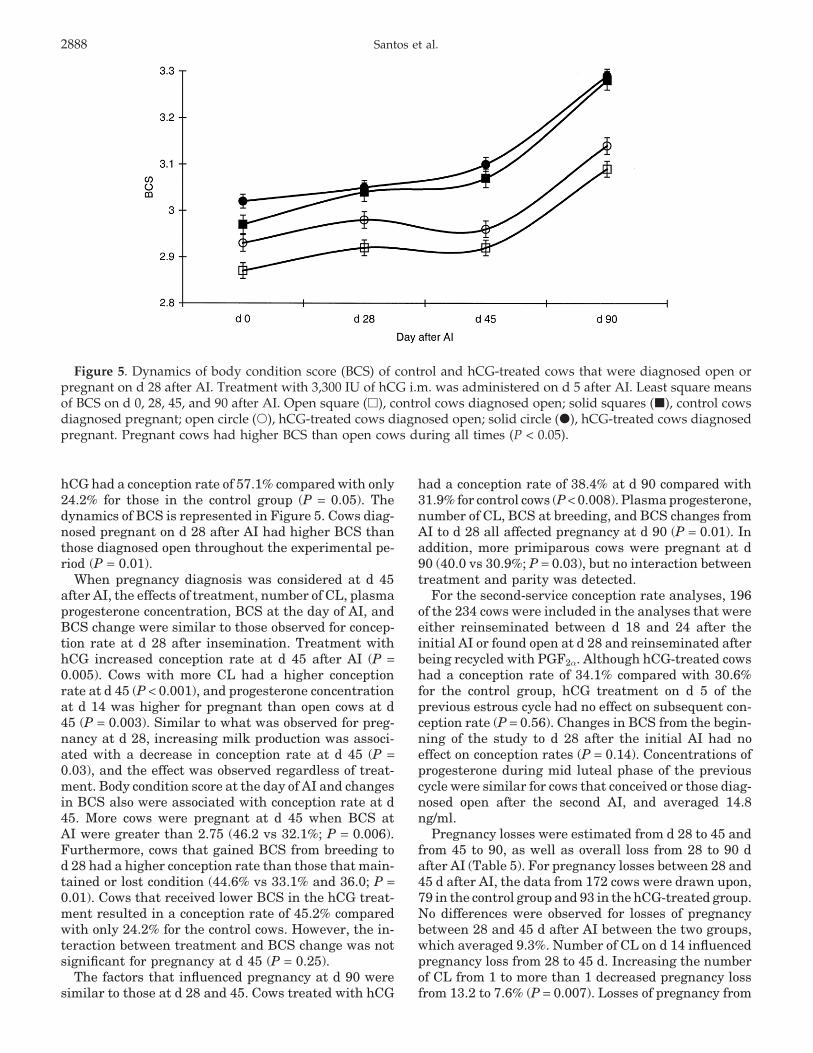
Figure 5. Dynamics of body condition score (BCS) of control and hCG-treated cows that were diagnosed open orpregnant on d 28 after AI. Treatment with 3,300 IU of hCG i.m. was administered on d 5 after AI. Least square meansof BCS on d 0, 28, 45, and 90 after AI. Open square (�), control cows diagnosed open; solid squares (�), control cowsdiagnosed pregnant; open circle (�), hCG-treated cows diagnosed open; solid circle (�), hCG-treated cows diagnosedpregnant. Pregnant cows had higher BCS than open cows during all times (P < 0.05).
hCG had a conception rate of 57.1% compared with only24.2% for those in the control group (P = 0.05). Thedynamics of BCS is represented in Figure 5. Cows diag-nosed pregnant on d 28 after AI had higher BCS thanthose diagnosed open throughout the experimental pe-riod (P = 0.01).
When pregnancy diagnosis was considered at d 45after AI, the effects of treatment, number of CL, plasmaprogesterone concentration, BCS at the day of AI, andBCS change were similar to those observed for concep-tion rate at d 28 after insemination. Treatment withhCG increased conception rate at d 45 after AI (P =0.005). Cows with more CL had a higher conceptionrate at d 45 (P < 0.001), and progesterone concentrationat d 14 was higher for pregnant than open cows at d45 (P = 0.003). Similar to what was observed for preg-nancy at d 28, increasing milk production was associ-ated with a decrease in conception rate at d 45 (P =0.03), and the effect was observed regardless of treat-ment. Body condition score at the day of AI and changesin BCS also were associated with conception rate at d45. More cows were pregnant at d 45 when BCS atAI were greater than 2.75 (46.2 vs 32.1%; P = 0.006).Furthermore, cows that gained BCS from breeding tod 28 had a higher conception rate than those that main-tained or lost condition (44.6% vs 33.1% and 36.0; P =0.01). Cows that received lower BCS in the hCG treat-ment resulted in a conception rate of 45.2% comparedwith only 24.2% for the control cows. However, the in-teraction between treatment and BCS change was notsignificant for pregnancy at d 45 (P = 0.25).
The factors that influenced pregnancy at d 90 weresimilar to those at d 28 and 45. Cows treated with hCG
had a conception rate of 38.4% at d 90 compared with31.9% for control cows (P < 0.008). Plasma progesterone,number of CL, BCS at breeding, and BCS changes fromAI to d 28 all affected pregnancy at d 90 (P = 0.01). Inaddition, more primiparous cows were pregnant at d90 (40.0 vs 30.9%; P = 0.03), but no interaction betweentreatment and parity was detected.
For the second-service conception rate analyses, 196of the 234 cows were included in the analyses that wereeither reinseminated between d 18 and 24 after theinitial AI or found open at d 28 and reinseminated afterbeing recycled with PGF2α. Although hCG-treated cowshad a conception rate of 34.1% compared with 30.6%for the control group, hCG treatment on d 5 of theprevious estrous cycle had no effect on subsequent con-ception rate (P = 0.56). Changes in BCS from the begin-ning of the study to d 28 after the initial AI had noeffect on conception rates (P = 0.14). Concentrations ofprogesterone during mid luteal phase of the previouscycle were similar for cows that conceived or those diag-nosed open after the second AI, and averaged 14.8ng/ml.
Pregnancy losses were estimated from d 28 to 45 andfrom 45 to 90, as well as overall loss from 28 to 90 dafter AI (Table 5). For pregnancy losses between 28 and45 d after AI, the data from 172 cows were drawn upon,79 in the control group and 93 in the hCG-treated group.No differences were observed for losses of pregnancybetween 28 and 45 d after AI between the two groups,which averaged 9.3%. Number of CL on d 14 influencedpregnancy loss from 28 to 45 d. Increasing the numberof CL from 1 to more than 1 decreased pregnancy lossfrom 13.2 to 7.6% (P = 0.007). Losses of pregnancy from
Human chorionic gonadotropin for dairy cows 2889
Table 5. Effect of treatment on pregnancy losses (%)between d 28 and 45, 45 and 90, and 28
d 28 to 45 of gestation were not affected by changes inBCS (P = 0.15), and cows that gained, maintained, orlost BCS experienced a 15.6, 11.3, and 5.1% pregnancyloss, respectively.
Data from 156 cows (74 control and 82 hCG) wereused to analyze pregnancy losses between d 45 and 90after AI. Percentages of cows that lost pregnancy werenot affected by hCG (P = 0.11). Body condition score atAI also influenced pregnancy losses between d 45 and90. Only 1.8% of cows pregnant at d 45 with moderateBCS at AI were found not pregnant at d 90, but 11.1%for those that had a low BCS lost their pregnancy be-tween d 45 and 90 (P = 0.06). Overall, pregnancy lossesfrom d 28 to d 90 were similar between hCG and controlcows and averaged 17.4%.
For the analyses of CL regression in cows with morethan one CL, data were collected only during the secondperiod of the study (cool season). A total of 71 cows (17controls and 54 hCG) with more than one CL werediagnosed with a single pregnancy on d 28 after AI.More additional CL were located in the ovary contralat-eral to the horn bearing the pregnancy (47 vs 24). Loca-tion of the additional CL (ipsilateral or contralateral tothe horn bearing the pregnancy) did not influence thenumber of CL that regressed by d 28 (P = 0.18; Table6). Additional CL regressed in seven (14.9%) and inone (4.2%) of the cows with double ovulation in thecontralateral and ipsilateral ovary, respectively. Treat-ment with hCG tended to reduce the number of preg-nant cows that had CL regression (23.5 vs 7.4%; P =0.07). However, no interaction between treatment andCL location on regression of additional CL was ob-served, and all pregnant cows that experienced CL re-
Table 6. Effect of treatment and accessory corpus luteum (CL) location on accessoryCL regression in cows with double ovulation at d 14
aC-C = control contralateral; C-I = control ipsilateral; hCG-C = human chorionic gonadotropin contralateral;hCG-I = human chorionic gonadotropin ipsilateral.
bTRT = treatment; L = location of accessory CL relative to the pregnant horn; TRT × L = interactionbetween treatment and location of accessory CL.
gression (eight cows) had milk production below themean milk yield (16.7 vs 0%; P = 0.04).
Discussion
Although there are numerous studies regarding theeffects of hCG on follicular development and CL func-tion (Schmitt et al., 1996a; Sianangama and Rajama-hendran, 1996; Diaz et al., 1998), few utilized a rela-tively large number of cows to assess the effectivenessof this hormonal treatment on conception rate and preg-nancy loss of high-yielding dairy cows under field condi-tions (Eduvie and Seguin, 1982).
Administering 3,300 IU of hCG i.m. on d 5 after AIresulted in 86.2% of the cows with more than one CLand the risk of hCG cows having more than one CL wasincreased by sixfold. A model of follicular developmentin dairy cows proposed by Ginther et al. (1996) indicatedthat deviation of the dominant follicle takes place 2to 3 d after a new cohort of follicles is recruited. Thedominant follicle acquires LH receptors or gene expres-sion for LH receptors between 2 and 4 d after waveemergence, when it achieves 8 to 10 mm in diameter(Xu et al., 1995; Ginther et al., 1996). Therefore, on d 5of the estrous cycle, dairy cows should have a dominantfollicle responsive to LH that is capable of ovulatingwhen ovulation is induced by hCG. Some cows in thehCG-treated group might have had a spontaneous dou-ble ovulation during estrus prior to hCG treatment,since multiple CL were observed in 23.2% of the con-trol cows.
Multiple ovulation previously was associated withthe level of milk production (Fricke and Wiltbank,1999). However, the current study did not observe aneffect of milk production, analyzed either as quartilesor as mean milk yield, on the incidence of multipleovulations in cows in any of the treatment groups. Wilt-bank et al. (2000) suggested that high-producing cowsmight select more than one dominant follicle, whichresults in double ovulation. When the data were ana-lyzed in a similar manner as described by Fricke andWiltbank (1999), the number of CL for cows with pro-duction above or below the mean milk (43.0 kg/d) weresimilar, and averaged 1.54 and 1.68, respectively, and
Santos et al.2890
no interaction between treatment and milk yield on CLnumber was observed.
Fourteen cows (6.9%) in the control and three (1.5%)in the hCG group had no detectable CL at ultrasonogra-phy on d 14 after AI, which indicates that either ovula-tion did not occur after estrus, detection of estrus wasinaccurate, CL were missed by ultrasonography, or theCL regressed prematurely. It is possible that the 13.8%of the hCG-treated cows with a single and not two CLat ultrasonography may have resulted from one or moreof the factors mentioned previously. This would explainthe low conception rate for hCG-treated cows with onlyone CL (12.0%). The effect of hCG on CL number wasmore pronounced during the cool season. Heat stresscan suppress follicular dominance in dairy cows, andthe dominant follicle might lose ovulatory capacitywhen cows are exposed to high ambient temperatures.Wolfenson et al. (1995) observed that heat-stressedcows had a 2 to 3 d earlier emergence of the second-wave dominant follicle. Perhaps, by d 5 after AI, thefirst-wave dominant follicle of some cows might havelost dominance and that would decrease the responseto hCG during the warm period.
Human chorionic gonadotropin treatment on d 5 afterestrus increased luteal weight of the accessory CL re-moved from the ovary on d 13 of the cycle comparedwith treatment with GnRH, but no effect was observedfor the weight of the original CL removed on d 17(Schmitt et al., 1996a). Diaz et al. (1998) observed asimilar pattern of early development of the spontaneousCL between control heifers and heifers receiving hCGon d 5 after ovulation. Sianangama and Rajamahen-dran (1996) measured CL of heifers receiving hCG ond 7 of the estrous cycle and observed no difference indiameter between the spontaneous CL in control andtreated cows up to d 12 of the estrous cycle. However,the same group (Rajamahendran and Sianangama,1992) showed that cows receiving hCG on the day ofAI had larger CL on d 7 after AI. Although most of theeffect of hCG on total luteal area and volume in thecurrent study was caused by the presence of multipleCL, treatment with hCG may also have increased thesize of the CL originating from the spontaneous ovula-tion. When we analyzed surface area and volume, onlyof the largest luteal structure present in the ovaries byultrasound on d 14 after AI, cows treated with hCGhad a CL with greater surface area and volume thanthat of control cows. Schmitt et al. (1996a) observed anincrease in size of small and large luteal cells for cowstreated with hCG on d 5 of the estrous cycle comparedwith those receiving saline. In ewes, administration of300 IU i.v. of hCG on d 5 and 7.5 of the estrous cycleincreased luteal weight and decreased the proportionof small to large luteal cells (Farin et al., 1988).
As reported previously (Diaz et al., 1998; Schmitt etal., 1996a), treatment of cows with hCG on d 5 of theestrous cycle resulted in greater plasma progesteroneconcentrations during mid luteal phase than those inthe controls. Schmitt et al. (1996a) observed that in
Figure 6. Regression of total corpus luteum (CL) area(cm2) and plasma progesterone concentration (ng/mL)on d 14 after AI (all cows included). Progesterone, ng/mL = 1.74262 + 3.47352X − 0.150919X2, where X is thetotal luteal area in cm2 (r2 = 0.46).
vitro production of progesterone by the spontaneousCL removed on d 17 of the estrous cycle did not differbetween hCG and control cows. Furthermore, no differ-ence in plasma progesterone concentrations was ob-served after removal of the induced CL on d 13 of thecycle. Therefore, it is likely that the increase in plasmaprogesterone concentrations observed in hCG-treatedcows at d 5 is associated with the increased numberof CL and not increased steroidogenic activity of thespontaneous CL.
A simple linear regression analysis was conductedto determine the best subset regression that predictsplasma progesterone concentrations. The predictorsused were largest CL area, total luteal area, and num-ber of CL. Largest CL area resulted in the lowest ad-justed coefficient of determination, and it explainedonly 19% of the variation in plasma progesterone on d14. The other two predictors, total CL area and numberof CL, explained 37 and 31% of the variation in plasmaprogesterone, respectively. Another regression analysiswas performed to determine the fitted-line plot, and aquadratic relationship between total luteal area andprogesterone concentrations was determined (Figure6): Progesterone, ng/mL = 1.74262 + 3.47352X −0.150919X2, where X is the total luteal area in squarecentimeters (adjusted r2 = 0.46).
The role of progesterone in fertility and pregnancymaintenance is undisputed. Poor luteal activity hasbeen associated with infertility in cattle (Wiebold,1998). Cows that have lower progesterone concentra-tions in the estrous cycle, either before or after breeding,have lower conception rates. Some studies have indi-
Human chorionic gonadotropin for dairy cows 2891
cated that peripheral concentrations of progesteroneare higher in pregnant cows as early as 6 d after AI(Shelton et al., 1990). Luteal cells from CL of cows con-sidered subfertile had decreased progesterone secretionper unit of luteal tissue when compared with cells fromcyclic and pregnant heifers (Shelton et al., 1990). Lowerprogesterone concentrations during the estrous cycleprevious to breeding may affect LH pulse frequency andalter the maintenance of the dominant follicle, whichmight reduce fertility (Mihm et al., 1994). Cattle withthree-wave cycles, which are associated with greaterprogesterone concentrations, may have higher concep-tion rates (Ahmad et al., 1997). Moreover, low circulat-ing progesterone after breeding may impact embryodevelopment and maternal recognition of pregnancy(Mann et al., 1999). Because hCG induces the formationof accessory CL and increases concentrations of proges-terone in plasma during mid luteal phase, it has beensuggested that hCG administration after breeding mayincrease conception rate (Sianangama and Rajamahen-dran, 1992) and reduce the incidence of early embryonicmortality in cattle (Rajamahendran and Sianangama,1992). Human chorionic gonadotropin also has beenshown to promote three-wave estrous cycles (Diaz etal., 1998). Inducing accessory CL and promoting three-wave cycles may delay the occurrence of preovulatoryfollicles around maternal recognition of pregnancy,which might positively impact CL lifespan and em-bryo survival.
Pregnant cows on d 28, 45, and 90 had greater concen-trations of plasma progesterone on d 14 after AI thanthose diagnosed open, and this effect was detected re-gardless of treatment. These results indicate thathigher mid luteal phase progesterone concentrationsfavor pregnancy during the first trimester. However,pregnancy losses during the first 90 d after insemina-tion were not affected by progesterone concentrationson d 14. The number of CL affected progesterone con-centrations but only when induced by hCG. This sug-gests that spontaneous double ovulations do not resultin higher plasma progesterone concentrations than asingle ovulation. However, when an accessory CL wasinduced by hCG, progesterone increased, indicatingthat steroidogenesis was enhanced. Body conditionscore at the day of AI influenced plasma concentrationsof progesterone differently in control and hCG-treatedcows. Treatment with hCG increased progesterone con-centrations of cows with a low BCS. A low BCS at breed-ing is associated usually with reduced fertility (Moreiraet al., 2000) and this may be mediated partially by lowerplasma progesterone concentrations. Utilizing hCG insuch cows may benefit conception rate through higherprogesterone concentrations. Similar to BCS at AI, milkyield also tended to influence plasma progesterone con-centrations and an interaction between treatment andmilk yield was observed. Control cows with productionabove the mean had lower progesterone concentrations,but no effect was observed for hCG-treated cows. Milkfat contains progesterone and higher producing cows
might eliminate more progesterone through milk. Inaddition, a high correlation is observed between milkyield and feed intake. Because higher DM intakes in-crease the flow of blood through the splanchnic tissues,it is possible that progesterone clearance by the liveris enhanced in cows with higher yields of milk. Vas-concelos et al. (1998) observed that bouts of feed intakedecreased serum progesterone relative to the prefeed-ing basal concentration in pegnant cows. Similar re-sults were observed for open cows with no CL that werecarrying a progesterone intravaginal device (Wiltbanket al., 2000).
The number of CL observed on d 14 after AI influ-enced conception rate on d 28, 45, and 90 after AI.Although no interaction between CL number and treat-ment on conception rate was observed, it is possiblethat the mechanism by which number of CL influencedpregnancy was distinct between the two groups. Cowswith more than one CL in the hCG-treated group hadhigher plasma progesterone concentrations, and thatmight have mediated the increase in conception rate.However, cows in the control group with more than oneCL, which originated from spontaneous double ovula-tions, had progesterone concentrations similar to thosewith only one CL. It is possible that the increase inconception rate for the few control cows with spontane-ous double ovulations may have been caused by an in-crease in the number of fertilized eggs. Fricke and Wilt-bank (1999) also observed an increase in conceptionrate for cows exhibiting double ovulations after a timed-AI protocol.
The incidence of spontaneous double ovulation was23.2% for control cows. This number was similar toresults observed by Kaim and Bor (2000) but higherthan previously reported for high-producing Holsteinafter a timed-AI protocol (Fricke and Wiltbank, 1999).Although 29 out of 47 cows in the control group withmultiple CL were pregnant, only 2 of them had twinpregnancies observed by ultrasound on d 28 or by rectalpalpation on d 45 after AI. This indicates that the major-ity of pregnant cows with spontaneous multiple ovula-tions do not carry a twin pregnancy. Fricke and Wilt-bank (1999) observed that only 3 of the 11 cows exhib-iting double ovulation after a timed-AI protocol calvedtwins. Because embryonic mortality is higher duringthe early stages of embryo development (Ginther, 1998;Vasconcelos et al., 1997), it is possible that the lack oftwin pregnancy in cows with spontaneous multiple CLwas caused by early embryonic death. Previous workby Day et al. (1995) observed that cows carrying twinshad a higher incidence of embryonic loss and abortionduring gestation. Moreover, the lack of twin pregnancyin control cows bearing more than one CL could alsoindicate a failure to fertilize the additional oocyte.
Previous research (Eduvie and Seguin, 1982; Helmerand Britt, 1986; Sianangama and Rajamahendran,1992) observed conflicting results on conception rate ofcows receiving hCG. Sianangama and Rajamahendran(1992) observed a remarkable increase in conception
Santos et al.2892
rate for cows receiving hCG on d 7 after AI (62% vs47%). However, two previous reports (Eduvie and Seg-uin, 1982; Helmer and Britt, 1986) showed no differ-ences in conception rate between control and hCG-treated cows. In the latter studies (Eduvie and Seguin,1982; Helmer and Britt, 1986), the conception rate ofcontrol cows (conception rate > 59%) was markedlyhigher than those currently observed for high-produc-ing Holstein cows (Butler, 1998) and those observed inthis study. It is likely that such highly fertile groupsof cows might not benefit from hCG treatment. In addi-tion, a limited number of cows and different protocolsand dosage regimens were utilized, which might haveaffected the ability to detect any difference betweentreatment groups. Helmer and Britt (1986) injectedcows with hCG at breeding, and this would not inducethe formation of an accessory CL. In such case, anyincrease in progesterone concentration would reside inenhancing the development of the ovulatory CL. Be-cause the response in progesterone concentration tohCG treatment is mostly associated with the formationof an accessory CL (Diaz et al., 1998; Schmitt et al.,1996a), it is possible that the lack of response to hCGtreatment might have been partly due to the fact thatno accessory CL was induced. In the current study,treatment with hCG on d 5 after AI significantly in-creased the conception rate of high-producing dairycows when pregnancy was determined on d 28, 45, or90 after AI. These results suggest that hCG inductionof multiple CL improves conception rates in high-pro-ducing dairy cows.
Summer thermal stress is an important factor dimin-ishing fertility in lactating dairy cows (Hansen and Are-chiga, 1999). The effects of heat stress on fertility areassociated with alterations in patterns of follicular de-velopment (Wolfenson et al., 1995), changes in hormoneprofiles, and suppressed progesterone production,which negatively impacts estrus expression and concep-tion rate, and increases pregnancy losses (Vasconceloset al., 1997; Hansen and Arechiga, 1999). A study con-ducted at the University of Florida (Schmitt et al.,1996b) did not show improvement in conception ratefollowing the induction of an accessory CL by a singleinjection of 3,000 IU of hCG on d 5 after AI duringsummer heat stress. We observed a tendency for inter-action between treatment and period on conception ratein the current study (P = 0.09). Human chorionic gonad-otropin had no effect on conception rate during the hotperiod (43.3 vs 44.1%), but improved conception rateduring the cool period (47.8 vs 34.2%). Cattle embryosare most susceptible to thermal stress during the first2 to 3 d after fertilization, and, when cows are exposedto high ambient temperatures, embryos generally dieduring the first days after conception (Hansen and Are-chiga, 1999). Therefore, similar to the findings bySchmitt et al. (1996b), it is unlikely that inducing anaccessory CL by treatment with hCG on d 5 after AIwould improve conception rate of heat-stressed dairycows.
The effects of BCS on the fertility of cattle are welldocumented (Ferguson, 1991). Cows with a low or de-clining body condition are more likely to be in anestrousand to have lower fertility (Ferguson, 1991; Moreira etal., 2000). We observed that cows with a BCS greaterthan 2.75 at the day of AI had a higher conception ratethan those with a BCS equal to or lower than 2.75 (48.7vs 34.4%; P < 0.001) and treatment did not affect thisresponse. The dynamics of BCS from d 0 to d 90 afterAI clearly demonstrate that BCS influences pregnancyin dairy cows. Cows diagnosed pregnant on d 28 afterAI had a higher BCS throughout the study than thatof those diagnosed open. Interestingly, an interactionbetween treatment and BCS change was detected forpregnancy at d 28. Cows that lost BCS when treatedwith hCG had a conception rate of 57.1%, comparedwith only 24.2% for controls. This demonstrates thathCG has the potential to improve conception rate incows losing body condition.
Human chorionic gonadotropin had no influence onthe decrease in conception rate caused by higher milkyields. Butler (1998) observed that conception rate hasdeclined during the last 45 yr for Holstein dairy cattle inthe U.S. and that milk production has simultaneouslyincreased steadily. Macmillan et al. (1996) observedthat cows managed under intensive systems, in whichhigh milk production is achieved, have shorter periodsfrom calving to first ovulation but lower conception rateduring the breeding period when compared with cowsmanaged for lower levels of production. These remark-able differences between groups of cows may be associ-ated with follicular development and concentrations ofplasma progesterone (Bilby et al., 1998). Higher produc-ing cows generally consume more feed DM, and feedintake can affect plasma progesterone concentrationsof dairy cows (Vasconcelos et al., 1998; Wiltbank etal., 2000).
In a recent study involving a large number of preg-nancy determinations (Vasconcelos et al., 1997), lossesof pregnancy in lactating dairy cows were 10.5, 6.3, 1.7,and 1.7% between d 28 and 42, 42 and 56, 56 and 70,and 70 and 90 after AI, respectively. These losses weresimilar to those observed in the current study. Preg-nancy losses were similar between the two treatmentgroups during the first 90 d of gestation.
Implications
The administration of 3,300 IU of human chorionicgonadotropin (hCG) i.m. on d 5 after AI in high-produc-ing dairy cows consistently induced the formation of anaccessory corpus luteum and increased plasma proges-terone concentrations during mid luteal phase. Sponta-neous double ovulations had no impact on plasma pro-gesterone concentrations. Increased progesterone con-centration was associated with pregnancy regardlessof treatment. The increase in the number of corporalutea and plasma progesterone concentrations causedby treatment with hCG was associated with greater
Human chorionic gonadotropin for dairy cows 2893
conception rates when pregnancy was diagnosed on d28, 45, and 90 after artificial insemination. Most ofthe benefit on conception was observed during the coolseason. Furthermore, pregnancy in cows losing bodycondition score benefited from hCG treatment. In con-clusion, treatment with hCG offers an opportunity toinduce the formation of an accessory CL, increaseplasma concentrations of progesterone, and improveconception rates in lactating dairy cows.
Literature Cited
Ahmad, N., E. C. Townsend, R. A. Dailey, and E. K. Inskeep. 1997.Relationships of hormonal patterns and fertility to occurrenceof two or three waves of ovarian follicles, before and after breed-ing, in beef cows and heifers. Anim. Reprod. Sci. 49:13–28.
Bilby, C. R., K. L. Macmillan, G. A. Vekerk, J. A. Peterson, A. T.Koenigsfeld, and M. C. Lucy. 1998. A comparative study of ovar-ian function in American (US) Holstein and New Zealand (NZ)Friesian lactating diary cows. J. Dairy Sci. 81(Suppl.1):222.(Abstr.)
Butler, W. R. 1998. Influence of protein nutrition on ovarian anduterine physiology in dairy cattle. J. Dairy Sci. 81:2533–2539.
Day, J. D., L. D. Weaver, and C. E. Franti. 1995. Twin pregnancydiagnosis in Holstein cows: Discriminatory powers and accuracyof diagnosis by transrectal palpation and outcome of twin preg-nancies. Can. Vet. J. 36:93–97.
Dean, A. G., J. A. Dean, A. H. Burton, and R. C. Dicker. 1991. EpiInfo: A general purpose microcomputer program for health infor-mation systems. Am. J. Prev. Med. 7:178–182.
Diaz, T., E. J.-P. Schmitt, R. L. De la Sota, M.-J. Thatcher, andW. W. Thatcher. 1998. Human chorionic gonadotropin-inducedalterations in ovarian follicular dynamics during the estrouscycle of heifers. J. Anim. Sci. 76:1929–1936.
Eduvie, L. O., and B. E. Seguin. 1982. Corpus luteum function andpregnancy rate in lactating dairy cows given human chorionicgonadotropin at middiestrus. Theriogenology 17:415–422.
Farin, C. E., C. L. Moeller, H. Mayan, F. Gamboni, H. R. Sawyer,and G. D. Niswender. 1988. Effect of luteinizing hormone andhuman chorionic gonadotropin on cell populations in the ovinecorpus luteum. Biol. Reprod. 38:413–421.
Ferguson, J. D. 1991. Nutrition and reproduction in dairy cows. TheVeterinary Clinics of North America—Food Animal Practice7:483–508.
Ferguson, J. D., D. T. Galligan, and N. Thomsen. 1994. Principaldescriptors of body condition score in Holstein cows. J. DairySci. 77:2695–2703.
Fricke, P. M., and M. C. Wiltbank. 1999. Effect of milk production onthe incidence of double ovulation in dairy cows. Theriogenology52:1133–1143.
Ginther, O. J. 1998. Ultrasonic Imaging and Animal Reproduction:Cattle. Book 3. pp 29–71. Equiservices Publishing, CrossPlains, WI.
Ginther, O. J., M. C. Wiltbank, P. M. Fricke, J. R. Gibbons, and K. Kot.1996. Selection of the dominant follicle in cattle. Biol. Reprod.55:1187–1194.
Hansen, P. J., and C. F. Arechiga. 1999. Strategies for managingreproduction in the heat-stressed dairy cow. J. Dairy Sci.82(Suppl. 2):36–50.
Helmer, S. D., and J. H. Britt. 1986. Fertility of dairy cattle treatedwith human chorionic gonadotropin (hCG) to stimulate proges-terone secretion. Theriogenology 26:683–695.
Hosmer, D. W., and S. Lemeshow. 1989. Model-building strategiesand methods for logistic regression. In: Applied Logistic Regres-sion. pp 82–135. John Wiley and Sons Inc., New York.
Hoyer, P. B., and G. D. Niswender. 1985. The regulation of steroido-genesis is different in the two types of ovine luteal cells. Can.J. Physiol. Pharmacol. 63:240–248.
Kaim, M., and A. Bor. 2000. Incidence of unilateral or bilateral doubleovulation and its effects on fertility in dairy cows in summerand winter. In Proc. 14th Intern. Congr. Anim. Reprod. Vol. 1.Stockholm, Sweden. p 288 (Abstr.).
Knickerbocker, J. J., W. W. Thatcher, F. W. Bazer, M. Drost, D. H.Barron, K. B. Fincher, and R. M. Roberts. 1986. Proteins secretedby day 16 to 18 conceptuses extend corpus luteum function incows. J. Biol. Reprod. 77:381–391.
Kuhel, R. 1994. Complete block designs. In: Statistical Principles ofResearch Design and Analysis. pp 256–305. Wadsworth Publish-ing Co., Belmont, CA.
Littell, R. C., P. R. Henry, and C. B. Ammerman. 1998. Statisticalanalysis of repeated measures data using SAS procedures. J.Anim. Sci. 76:1216–1231.
Macmillan, K. L., I. J. Lean, and C. T. Westwood. 1996. The effectsof lactation on the fertility of dairy cows. Austral. Vet. J.73:141–147.
Macmillan, K. L., V. K. Taufa, D. R. Barnes, A. M. Day, and R. Henry.1988. Detecting estrus in synchronized heifers using tailpaintand aerosol raddle. Theriogenology 30:1099–1114.
Mann, G. E., G. E. Lamming, R. S. Robinson, and D. C. Wathes. 1999.The regulation of interferon-τ production and uterine hormonereceptors during early pregnancy. J. Reprod. Fertil.54(Suppl.):317–328.
Mihm, M., A. Baguisi, M. P. Boland, and J. F. Roche. 1994. Associationbetween the duration of the dominance of the ovulatory follicleand pregnancy rate in beef heifers. J. Reprod. Fertil.102:123–130.
MINITAB. 1996. MINITAB� Reference Manual Release II. MINITABInc., State College, PA.
Moreira, F., C. Risco, M. F. A. Pires, J. D. Ambrose, M. Drost, M.DeLorenzo, and W. W. Thatcher. 2000. Effect of body conditionon reproductive efficiency of lactating dairy cows receiving atimed insemination. Theriogenology 53:1305–1319.
Rajamahendran, R., and P. C. Sianangama. 1992. Effect of humanchorionic gonadotropin on dominant follicles in cows: formationof accessory corpora lutea, progesterone production and preg-nancy rates. J. Reprod. Fertil. 95:577–584.
Schmitt, E. J. P., C. M. Barros, P. A. Fields, M. J. Fields, T. Diaz, J.M. Kluge, and W. W. Thatcher. 1996a. A cellular and endocrinecharacterization of the original and induced corpus luteum afteradministration of a gonadotropin-releasing hormone agonist orhuman chorionic gonadotropin on day five of the estrous cycle.J. Anim. Sci. 74:1915–1929.
Schmitt, E. J. P., T. Diaz, C. M. Barros, R. L. de la Sota, M. Drost, E.W. Fredriksson, C. R. Staples, R. Thorner, and W. W. Thatcher.1996b. Differential response of the luteal phase and fertility incattle following ovulation of the first-wave follicle with humanchorionic gonadotropin or an agonist of gonadotropin-releasinghormone. J. Anim. Sci. 74:1074–1083.
Seguin, B. E., W. D. Oxender, and J. H. Britt. 1977. Effect of humanchorionic gonadotropin and gonadotropin-releasing hormone oncorpus luteum function and estrous cycle duration in dairy heif-ers. Am. J. Vet. Res. 38:1153–1156.
Shelton, K., M. F. Gayerie de Abreu, M. G. Hunter, T. J. Parkinson,and G. E. Lamming. 1990. Luteal inadequacy during early lutealphase of subfertile cows. J. Reprod. Fertil. 90:1–10.
Sianangama, P. C., and R. Rajamahendran. 1992. Effect of humanchorionic gonadotropin administered at specific times followingbreeding on milk progesterone and pregnancy in cows. Therio-genology 38:85–96.
Sianangama, P. C., and R. Rajamahendran. 1996. Characteristicsof corpus luteum formed from the first wave dominant folliclefollowing hCG in cattle. Theriogenology 45:977–990.
Thatcher, W. W., K. L. Macmillan, P. J. Hansen, and M. Drost. 1989.Concepts for regulation of corpus luteum function by the concep-tus and ovarian follicles to improve fertility. Theriogenology31:14–164.
Santos et al.2894
Vasconcelos, J. L. M., K. A. Bungert, S. J. Tsai, F. S. Wechsler, andM. C. Wiltbank. 1998. Acute reduction in serum progesteroneconcentrations due to feed intake in pregnant lactating dairycows. J. Dairy Sci. 81(Suppl. 1):226 (Abstr.).
Vasconcelos, J. L. M., R. W. Silcox, J. A. Lacerda, J. R. Pursley,and M. C. Wiltbank. 1997. Pregnancy rate, pregnancy loss, andresponse to heat stress after AI at 2 different times from ovula-tion in dairy cows. Soc. Study Reprod. 56(Suppl. 1):230 (Abstr.).
Wiebold, J. L. 1998. Embryonic mortality and the uterine environ-ment in first service lactating dairy cows. J. Reprod. Fertil.84:393–399.
Wiltbank, M., P. Fricke, S. Sangsritavong, R. Sartori, and O. J.Ginther. 2000. Mechanisms that prevent and produce doubleovulations in dairy. J. Dairy Sci. 83:2998–3007.
Wolfenson, D., W. W. Thatcher, L. Badinga, J. D. Savio, R. Meidan,B. J. Lew, R. Braw-Tal, and A. Berman. 1995. Effect of heat stresson follicular development during the estrous cycle in lactatingcattle. Biol. Reprod. 52:1106–1113.
Xu, Z., H. A. Garverick, G. W. Smith, M. F. Smith, S. A. Hamilton,and R. S. Youngquist. 1995. Expression of follicle-stimulatinghormone and luteinizing hormone receptors messenger ribo-nucleic acids in bovine follicles during the first follicular wave.Biol. Reprod. 53:951–957.