Intern. J. Neuroscience, 117:971–983, 2007 Copyright C 2007 Informa Healthcare ISSN: 0020-7454 / 1543-5245 online DOI: 10.1080/00207450600934085 EFFECT OF INGESTED SULFITE ON HIPPOCAMPUS ANTIOXIDANT ENZYME ACTIVITIES IN SULFITE OXIDASE COMPETENT AND DEFICIENT RATS VURAL KUCUKATAY MELEK BOR-KUCUKATAY PIRAY ATSAK Pamukkale University Faculty of Medicine Department of Physiology Kinikli, Denizli, Turkey AYSEL A ˘ GAR Akdeniz University Faculty of Medicine Department of Physiology Antalya, Turkey Animal tissues are exposed to sulfite used as a preservative in food and drugs, and generated from the catabolism of sulfur-containing amino acids. Sulfite, which is a very reactive and potentially toxic molecule, is detoxified by the enzyme sulfite oxidase (SOX). Laboratory animals can be made deficient in SOX by the administration of a high-tungsten/low molybdenum regimen. It has been suggested that SOX deficient rats might be used as a model for the prediction of sulfite toxicity in humans. The aim of this study was to investigate the effects of ingested sulfite Received 23 March 2006. This study was carried out as part of a Ph.D. thesis by V. Kucukatay presented to Akdeniz University Health Sciences Institute. The work was supported by a grant from Akdeniz University Research Fund 2002.0122.04. Address correspondence to Vural Kucukatay, Pamukkale University Faculty of Medicine, Department of Physiology, Kinikli, 20020 Denizli, Turkey. E-mail: [email protected]971 Int J Neurosci Downloaded from informahealthcare.com by University of Newcastle on 03/17/14 For personal use only.
EFFECT OF INGESTED SULFITEON HIPPOCAMPUS ANTIOXIDANT ENZYMEACTIVITIES IN SULFITE OXIDASE COMPETENTAND DEFICIENT RATS
VURAL KUCUKATAYMELEK BOR-KUCUKATAYPIRAY ATSAK
Pamukkale UniversityFaculty of MedicineDepartment of PhysiologyKinikli, Denizli, Turkey
AYSEL AGAR
Akdeniz UniversityFaculty of MedicineDepartment of PhysiologyAntalya, Turkey
Animal tissues are exposed to sulfite used as a preservative in food and drugs,and generated from the catabolism of sulfur-containing amino acids. Sulfite, whichis a very reactive and potentially toxic molecule, is detoxified by the enzymesulfite oxidase (SOX). Laboratory animals can be made deficient in SOX by theadministration of a high-tungsten/low molybdenum regimen. It has been suggestedthat SOX deficient rats might be used as a model for the prediction of sulfite toxicityin humans. The aim of this study was to investigate the effects of ingested sulfite
Received 23 March 2006.This study was carried out as part of a Ph.D. thesis by V. Kucukatay presented to Akdeniz
University Health Sciences Institute. The work was supported by a grant from Akdeniz UniversityResearch Fund 2002.0122.04.
Address correspondence to Vural Kucukatay, Pamukkale University Faculty of Medicine,Department of Physiology, Kinikli, 20020 Denizli, Turkey. E-mail: [email protected]
971
Int J
Neu
rosc
i Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
ity o
f N
ewca
stle
on
03/1
7/14
For
pers
onal
use
onl
y.
972 V. KUCUKATAY ET AL.
on hippocampus superoxide dismutase (SOD), catalase (CAT), and glutathioneperoxidase (GPx) activities in SOX competent and deficient rats. HippocampusSOD, CAT and GPx activities were found to be significantly increased by sulfitetreatment in SOX competent groups. On the other hand, exposure to sulfitehad no effect on antioxidant status in hippocampus of SOX deficient rats. Inconclusion, these results suggest that hippocampus antioxidant capacity wheredefense mechanism against the oxidative challenge is up regulated by sulfite inSOX competent rats. This up regulation mechanism in antioxidant enzymes againstto sulfite related oxidative stress is not observed in SOX deficient rats and remainsto be explained.
In food and beverages, sulfur dioxide (SO2) and various sulfite salts such assodium metabisulfite are widely used for a variety of preservative properties,including delaying undesirable changes in color and preventing bacterial growth(Taylor et al., 1986). It has been shown that both sulfite salts and SO2 formsulfite ion in body fluids and in aqueous solutions (Gunnison & Palmes, 1973).Additionally, sulfite ions are also continuously formed as a consequence ofbody’s normal processing of sulfur containing amino acids and other sulfur-containing compounds (Cooper, 1983). Sulfite, being a toxic molecule needs tobe detoxified by sulfite oxidase (SOX), which catalyzes the oxidation of sulfiteto sulfate (Cohen & Fridovich, 1971). An extreme example is a congenitaldisease of SOX deficiency. Genetic deficiency of SOX results in a debilitatingdisease characterized by severe neurological abnormalities, seizures, mentalretardation, and dislocation of the ocular lenses that often leads to death ininfancy (Mudd et al., 1967). Thus, sulfite levels in the body must be firmlyregulated. The mechanism of sulfite toxicity is not well known but is thoughtto be through the induction of oxidative stress by oxygen- and sulfur-centeredfree radicals (Ozawa & Hanaki, 1987; Shi, 1994; Abedinzadeh, 2001). Theseradicals were shown to be produced by either non-enzymatic oxidation of sulfiteor by various peroxidases (Mottley et al., 1982).
The brain may be considered to be among the most susceptible organs tooxidative stress that results from the generation of free radicals (Freitas et al.,2001) because it contains relatively high levels of polyunsaturated fatty acidsthat predispose it to lipid peroxidation, a complex process in which membranelipids react with free radicals to produce lipid hydroperoxides (Brand et al.,2000; Schafer et al., 2000). Moreover, neuronal cells are known to have low
Int J
Neu
rosc
i Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
ity o
f N
ewca
stle
on
03/1
7/14
For
pers
onal
use
onl
y.
ACTIVITIES IN SULFITE OXIDASE COMPETENT AND DEFICIENT RATS 973
SOX activity to protect sulfite toxicity (Woo et al., 2003). The central nervoussystem is protected against detrimental effects of free radicals by a numberof antioxidant enzymes such as superoxide dismutase (SOD), catalase (CAT),and glutathione peroxidase (GSH-PX) (Taniguchi, 1992; Michiels et al., 1994;Hayes & McLellan, 1999). It is well known that, there is a delicate balancebetween the rates of free radical formation and their removal by antioxidantenzymes. Thus, oxidant injury in the body may be either due to a decline inantioxidant enzyme capacity or to an increment in oxidative stress (Freeman &Crapo, 1982).
Although several studies have indicated that exposure to sulfite by SO2
inhalation leads to a decrement in antioxidant status in many tissues includingthe brain (Gumuslu et al., 1998; Yargicoglu et al., 2001), it is not clear whetherthere is also an ingested sulfite induced alteration in the activity of antioxidantenzymes in mammals. The present authors have recently reported that sulfiteingestion impairs active avoidance learning in SOX deficient rats but not in com-petent rats. Interestingly, sulfite had no effect on TBARS levels of hippocampusin SOX competent groups but, hippocampus TBARS levels were found to besignificantly increased upon sulfite exposure in SOX deficient rats (Kucukatayet al., 2005). These findings may be due to altered antioxidant enzymeactivities. There are no reports concerning ingested sulfite related changes ofantioxidant enzyme activities in hippocampus of SOX competent and deficientrats. Therefore, this study examined the effects of ingested sulfite on SOD,CAT, and GPx activities in hippocampus of SOX competent and deficient rats.
MATERIALS AND METHODS
Animals and Dietary Treatments
Forty adult male albino rats weighing 180–200 g were used throughout allexperiments. They were bred in the Akdeniz University Animal Unit, housedfive per cage at 22–25◦C with a 12-h light/dark cycle and randomly divided intotwo so as to form SOX competent (SOXC, n = 20) and SOX deficient groups(SOXD, n = 20). Each group was further divided into two subgroups: control(C) and sulfite (S) for SOXC groups and deficient (D), deficient + sulfite (DS)for SOXD groups. Each subgroup contained ten rats.
Rats in SOXC groups were fed ad libitum with standard rat chow andtap water. Rats in S and DS groups were given sulfite in the form ofsodium metabisulfte (25 mg/kg/day) in drinking water. These treatments werecontinued throughout the experiment (6 weeks). Rats in SOXD groups were
Int J
Neu
rosc
i Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
ity o
f N
ewca
stle
on
03/1
7/14
For
pers
onal
use
onl
y.
974 V. KUCUKATAY ET AL.
Table 1. The daily diet regimen of all experimental groups
Groups DietDrinking water
supplementations
Sulfiteoxidasestatus
Control (C) Standard rat chow None NormalSulfite (S) Standard rat chow Sulfite (25 mg/kg) NormalDeficient (D) Mo free diet W (200 ppm) DeficientDeficient + Sulfite (DS) Mo free diet W (200 ppm) + sulfite
(25 mg/kg)Deficient
made SOX deficient by the administration of a low molybdenum diet (AIN 76,Research Dyets Inc., USA) with concurrent addition to their drinking water ofsodium tungstate at the level of 200 ppm tungsten (Table 1). These treatmentswere started 3 weeks before the beginning of sulfite administration. In order tocommence the study, at the end of the 3rd week, SOX activity in the livers ofSOXD rats was measured to confirm that the rats were SOX deficient.
All experimental protocols used in this work were approved by the AkdenizUniversity Animal Care and Usage Committee.
The reagents used during all experimental protocols were obtained fromMerck (Darmstadt, Germany) and Sigma (St. Louis, MO, USA).
Tissue Collection and Preparation for Enzyme Assays
At the end of the 6th week, the animals were killed by exsanguination underurethane anesthesia, and brain and liver tissues were removed immediately.The tissues were stored at −80◦C until use. On the day of the experiment,hippocampi from the removed brains were carefully dissected to avoid tissuedamage according to the anatomic description of Paxinos and Watson (Paxinos& Watson, 1986). Hippocampi and about 1 gram of frozen livers werehomogenized at high speed with the homogenizer placed in the chipped icein 50 mM phosphate buffer. In order to remove precipitates, hippocampalhomogenates were centrifuged at 12,000 g for 10 min at +4◦C and liverhomogenates were centrifuged at 2,100 g for 10 min at +4◦C. The liverswere used for the assay of SOX activity. and the hippocampi were analyzed forantioxidant enzyme activities.
Assays of SOX and Antioxidant Enzyme Activities
SOX Activity Assay. A modification of the procedure of Cohen and Fridovichwas used in order to determine hepatic SOX activity (Cohen & Fridovich, 1971).
Int J
Neu
rosc
i Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
ity o
f N
ewca
stle
on
03/1
7/14
For
pers
onal
use
onl
y.
ACTIVITIES IN SULFITE OXIDASE COMPETENT AND DEFICIENT RATS 975
An aliquot of supernatant was mixed with 5% Triton X-100 and the mixture wasstored on ice. This mixture was further diluted (1:10) with 50 mmol phosphatebuffer (pH 7.4). This diluted mixture was used for measuring SOX activity atroom temperature by monitoring the reduction of cytochrome c at 550 nm. Themixture was added to the cuvette containing 10 mmol sodium sulfite, 0.2 mmolcytochrome c, Triton X-100 (% 5), 100 mmol Tris HCL (pH 8.5), and KCN (10mmol in Tris HCL) in a final volume of 2.5 ml. The slow non-enzymatic rate ofreduction of cytochrome c was first recorded and its rate was subtracted fromthe recorded total rate. One unit of SOX activity was defined as the amount ofenzyme, which caused an absorbance change of 0.1/min under these conditions.The results were expressed as units/mg protein.
Measurement of Hippocampus SOD Activity. SOD activity in hippocam-pus was determined by the method of Misra et al. (Misra & Fridovich, 1972).Briefly, hippocampal supernatants were added to reaction mixture (50 mmolNa2CO3/NaHCO3 buffer, 0,075 mmol EDTA and 3.6 mmol epinephrine HCL)in a final volume of 1.1 ml and SOD activity was measured at 30◦C and 480nm. The unit of activity was defined as the amount of enzyme that inhibits therate of autoxidation of epinephrine by 50% under standard conditions and wasexpressed units/mg protein.
Determination of CAT Activity. Hippocampi CAT activity was measuredaccording to the method of Aebi et al. with hydrogen peroxide as the substrate(Aebi, 1987). The final volume of each enzyme assay was 1.5 ml containing 30mmol H2 O2 and diluted supernatant of hippocampus homogenate. The assaywas performed at 25◦C and 240 nm. Enzyme activity was expressed as k/mgprotein, where k is the rate constant of the 1st order reaction, as defined byAebi.
GPx Activity Assay. A sigma kit (Glutathione Peroxidase Cellular ActivityAssay Kit CGP-1) was used for measuring GPx activity in hippocampus.The assay was based on the oxidation of glutathione (GSH) to oxidizedglutathione (GSSG) catalyzed by GPx coupled to the recycling of GSSG backto GSH utilizing glutathione reductase and NADPH. The decrease in NADPHabsorbance measured at 340 nm during the oxidation of NADPH to NADP isindicative of GPx activity because the enzyme is the rate-limiting factor of thecoupled reactions.
Int J
Neu
rosc
i Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
ity o
f N
ewca
stle
on
03/1
7/14
For
pers
onal
use
onl
y.
976 V. KUCUKATAY ET AL.
Determination of Liver and Hippocampus Protein Concentrations
The protein concentration of liver and hippocampus samples was determinedaccording to the method of Lowry et al. (1951) using bovine serum albumin asa standard.
Statistical Analysis
Results were expressed as means ± SE. Differences between mean values inthe study groups were evaluated by 1-way ANOVA followed by TUKEY posthoc test. p values <.05 were accepted as statistically significant.
RESULTS
Animal Health and Survival
All animals appeared generally healthy during the experimental period. Therewere no signs of toxicity and body-weight gain and survivals were similar forall experimental groups.
Hepatic SOX Activity
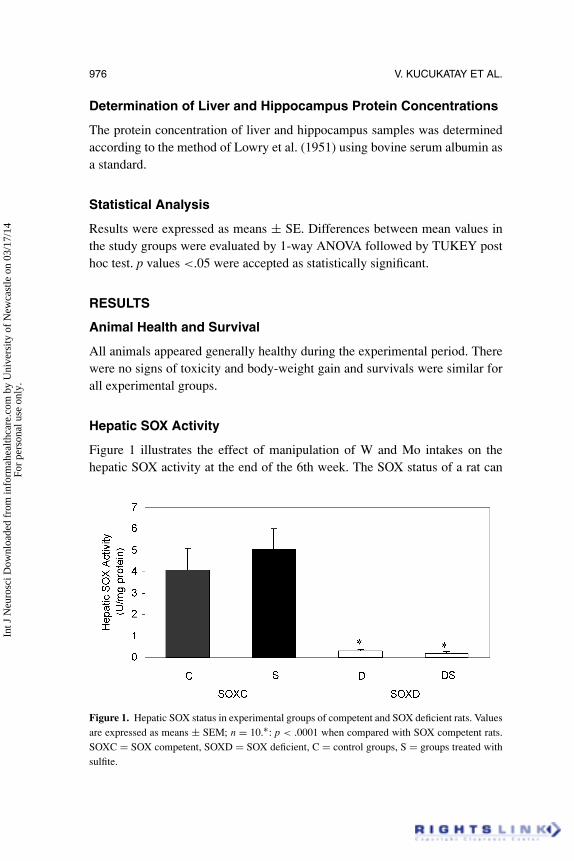
Figure 1 illustrates the effect of manipulation of W and Mo intakes on thehepatic SOX activity at the end of the 6th week. The SOX status of a rat can
Figure 1. Hepatic SOX status in experimental groups of competent and SOX deficient rats. Valuesare expressed as means ± SEM; n = 10.∗: p < .0001 when compared with SOX competent rats.SOXC = SOX competent, SOXD = SOX deficient, C = control groups, S = groups treated withsulfite.
Int J
Neu
rosc
i Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
ity o
f N
ewca
stle
on
03/1
7/14
For
pers
onal
use
onl
y.
ACTIVITIES IN SULFITE OXIDASE COMPETENT AND DEFICIENT RATS 977
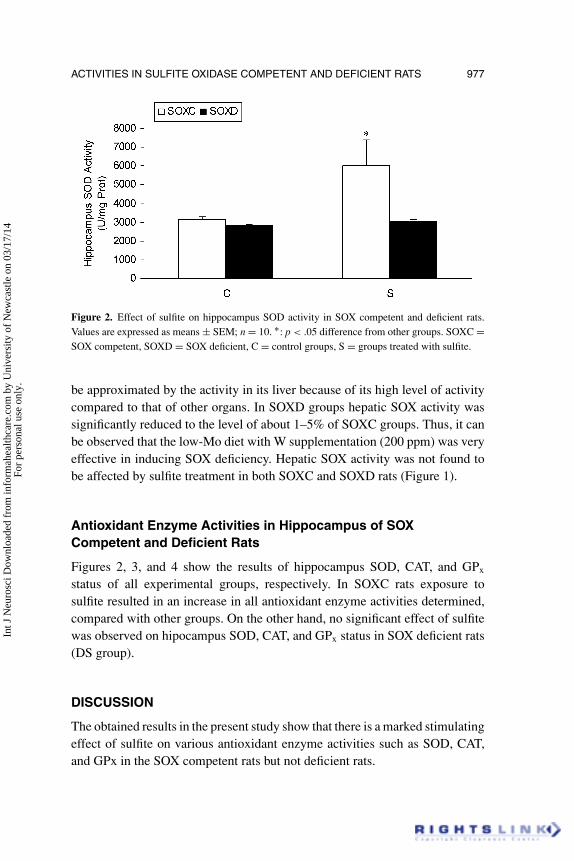
Figure 2. Effect of sulfite on hippocampus SOD activity in SOX competent and deficient rats.Values are expressed as means ± SEM; n = 10. ∗: p < .05 difference from other groups. SOXC =SOX competent, SOXD = SOX deficient, C = control groups, S = groups treated with sulfite.
be approximated by the activity in its liver because of its high level of activitycompared to that of other organs. In SOXD groups hepatic SOX activity wassignificantly reduced to the level of about 1–5% of SOXC groups. Thus, it canbe observed that the low-Mo diet with W supplementation (200 ppm) was veryeffective in inducing SOX deficiency. Hepatic SOX activity was not found tobe affected by sulfite treatment in both SOXC and SOXD rats (Figure 1).
Antioxidant Enzyme Activities in Hippocampus of SOXCompetent and Deficient Rats
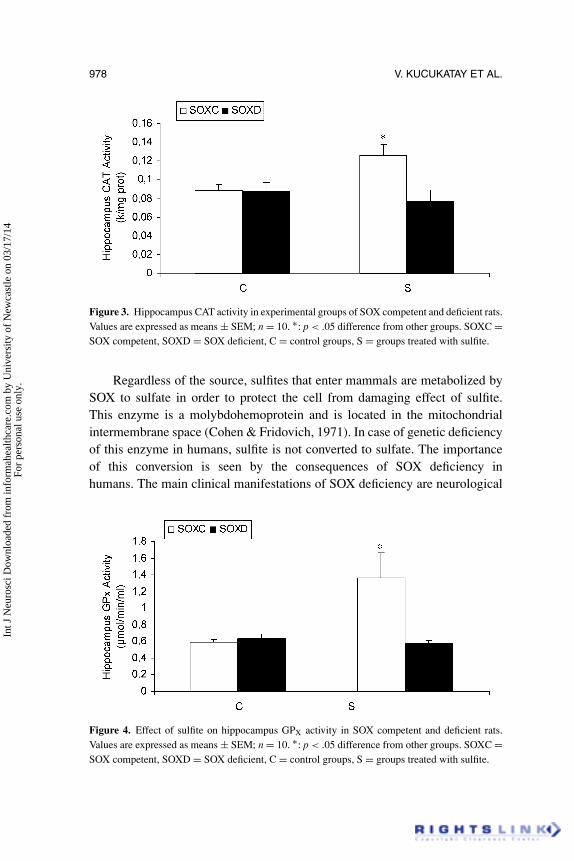
Figures 2, 3, and 4 show the results of hippocampus SOD, CAT, and GPx
status of all experimental groups, respectively. In SOXC rats exposure tosulfite resulted in an increase in all antioxidant enzyme activities determined,compared with other groups. On the other hand, no significant effect of sulfitewas observed on hipocampus SOD, CAT, and GPx status in SOX deficient rats(DS group).
DISCUSSION
The obtained results in the present study show that there is a marked stimulatingeffect of sulfite on various antioxidant enzyme activities such as SOD, CAT,and GPx in the SOX competent rats but not deficient rats.
Int J
Neu
rosc
i Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
ity o
f N
ewca
stle
on
03/1
7/14
For
pers
onal
use
onl
y.
978 V. KUCUKATAY ET AL.
Figure 3. Hippocampus CAT activity in experimental groups of SOX competent and deficient rats.Values are expressed as means ± SEM; n = 10. ∗: p < .05 difference from other groups. SOXC =SOX competent, SOXD = SOX deficient, C = control groups, S = groups treated with sulfite.
Regardless of the source, sulfites that enter mammals are metabolized bySOX to sulfate in order to protect the cell from damaging effect of sulfite.This enzyme is a molybdohemoprotein and is located in the mitochondrialintermembrane space (Cohen & Fridovich, 1971). In case of genetic deficiencyof this enzyme in humans, sulfite is not converted to sulfate. The importanceof this conversion is seen by the consequences of SOX deficiency inhumans. The main clinical manifestations of SOX deficiency are neurological
Figure 4. Effect of sulfite on hippocampus GPX activity in SOX competent and deficient rats.Values are expressed as means ± SEM; n = 10. ∗: p < .05 difference from other groups. SOXC =SOX competent, SOXD = SOX deficient, C = control groups, S = groups treated with sulfite.
Int J
Neu
rosc
i Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
ity o
f N
ewca
stle
on
03/1
7/14
For
pers
onal
use
onl
y.
ACTIVITIES IN SULFITE OXIDASE COMPETENT AND DEFICIENT RATS 979
abnormalities such as severe mental retardation, seizures, spastic quadriparesis,dislocated lenses, progressive destruction of brain tissue, and early death(Mudd et al., 1967). These findings highlight the potential neurotoxicity ofsulfite.
Six sulfite compounds (sulfur dioxide, sodium sulfite, sodium andpotassium bisulfite, and sodium and potassium metabisulfite) have been listedas “Generally Recognized as Safe” (GRAS) for use in foods and drugs.The acceptable daily intake (ADI) for exogenous sulfite was established as0–0.7 mg/kg/body weight by The Joint FAO/WHO Expert Committee onFood Additives (FAO/WHO, 1974). However, the daily intake of sulfite is notagreement with this value. Studies have shown that it is possible to consume180–200 mg/body weight from foods and beverages in a single day or meal(Taylor et al., 1986). Hence, the authors chose higher level of sulfite in thedrinking water than ADI value (25 mg/kg).
The study also used a SOX deficent rat model to evaluate sulfite effecton hippocampus antioxidant status. There are significant differences amongspecies in their SOX activity (Tejnorova, 1978). Most notable is the differencebetween rat and man. It was shown that rat liver has about a 20-fold greaterSOX activity than human liver (Johnson & Rajagopalan, 1976). Studies havealso shown an inverse correlation between SOX levels in liver and the sulfitetoxicity in several species of laboratory animals (Tejnorova, 1978). One canspeculate that these species may not be the most appropriate model for theprediction of sulfite toxicity in man. Therefore, it has been suggested that SOXdeficient rats might be used as a model for the prediction of sulfite toxicity inhuman (Johnson et al., 1974).
It was shown that there are toxic effects of exogenous sulfite on manycellular components (Hayatsu & Miller, 1972; Meng et al., 2002). Althoughthere is little information about the mechanism of sulfite toxicity on neuron,sulfur-, and oxygen-centered free radicals induced brain damage might playan important role in the development of these abnormalities in SOX deficiency(Ozawa & Hanaki, 1987; Shi, 1994; Abedinzadeh, 2001). Indeed, it was shownthat sulfite is a reductant that can undergo one- or two-electron oxidationforming a sulfite radical (SO−.
3) and sulfate (SO−24), respectively (Mottley
et al., 1982). The sulfite radical can further react with molecular oxygen forminga sulfite peroxyl radical (SO3 OO.) and a sulfate radical (SO−.
4) (Mottleyet al., 1982). These sulfur centered free radicals can react with many cellularcomponent including lipids, proteins, and DNA. These interactions betweenradicals and cell components are one of the major damaging pathways inbiological systems.
Int J
Neu
rosc
i Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
ity o
f N
ewca
stle
on
03/1
7/14
For
pers
onal
use
onl
y.
980 V. KUCUKATAY ET AL.
In the present study, it was found that there were significantly increasedSOD, CAT, and GPx activities in hippocampus of sulfite-exposed non-deficientgroups (CS group). These significant increments of hippocampus antioxidantcapacity in CS group may be an adaptive response to sulfite dependent oxidativestress. It has been shown that SOD, CAT, and GPx are primary enzymes toprotect the brain against damage caused by ROS (Taniguchi, 1992; Michielset al., 1994; Hayes & McLellan, 1999). Furthermore, these enzymes activitiescan be induced by oxidative stress in some circumstances such as exercise(Ji, 1995). Under normal conditions, a delicate balance exists between rates offree radical formation and their removal by antioxidant enzymes (Freeman &Crapo, 1982). Although an up regulation of one of the antioxidant enzymesmay have a protective role, this is only true when adequate activity of the othersexists (Freitas et al., 2001). In agreement with the present results, there are a lotof studies that performed an enhancement of cell tolerance to oxidative stressby modifying the cell antioxidant capacity (Ji, 1995; Lee, 2002). In addition,it is believed that the changes in antioxidant enzymes induced by oxidativestress might be due to synthesis of new isozymes or enhancement of activitiesof existing antioxidant enzymes for metabolism of free radicals (Lee, 2002).Therefore, the elevated level in antioxidant enzymes may be also related tosulfite-based oxidative stress in the present study. It remains to be seen whetherthese alterations in enzyme activities of this study are due to alterations in genetranscription and de novo synthesis, or are due to posttranslational modificationsof pre-existing enzyme.
In contrary to the SOX competent group, no significant alteration inantioxidant enzymes was found in SOX deficient rats treated with sulfite.The reason for this observation may be a decrease in production of reducingpower for antioxidant enzyme activities due to sulfite treatment. The activityof antioxidant enzymes requires a constant supply of reducing power(NADH/NADPH) through two major sites, the Krebs cycles in mitochondriaand the pentose phosphate pathway in the cytosol (Kletzien et al., 1994). Thesesites of production of reducing power in the brain may be affected negativelyby the sulfite. Therefore, the unchanged activity of antioxidant enzymes in thehippocampus of SOX deficient rats treated with sulfite may result from to bedecreased capacity of reducing power by sulfite. In addition, whether SOXdeficiency protocol also affects antioxidant activities remains to be elucidated.
In summary, the present results suggest that sulfite treatment may causeoxidative stress and competent animals in SOX cope with this stressfulcondition whereas SOX deficent rats, which are an exaggerated model forthe normal human situation, can not handle this possible sulfite dependent
Int J
Neu
rosc
i Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
ity o
f N
ewca
stle
on
03/1
7/14
For
pers
onal
use
onl
y.
ACTIVITIES IN SULFITE OXIDASE COMPETENT AND DEFICIENT RATS 981
oxidative stress. This is a conclusion that needs further work to elucidate themechanisms of these observations.
REFERENCES
Abedinzadeh, Z. (2001). Sulfur-centered reactive intermediates derived from theoxidation of sulfur compounds of biological interest. Canadian Journal ofPhysiology and Pharmacology, 79, 166–170.
Aebi, B. E. (1987). Catalase. In Bergmeyer, H. U. (Ed.), Methods of enzymatic analysis.Vol. III. (pp. 273–286). Germany: VCH Verlagsgesellschaft mbh.
Brand, A., Gil, S., & Yavin, E. (2000). N-methyl bases of ethanolamine prevent apoptoticcell death induced by oxidative stress in cells of oligodendroglia origin. Journalof Neurochemistry, 74, 1596–1604.
Cohen, H. J., & Fridovich, I. (1971). Hepatic sulfite oxidase. Purification and properties.Journal of Biological Chemistry, 246, 359–366.
Cooper, A. J. (1983). Biochemistry of sulfur-containing amino acids. Annual Review ofBiochemistry, 52, 187–222.
FAO/WHO. (1974). Expert Committee on Food Additives. Toxicological Evaluation ofCertain Food Additives with a Review of General Principle and of Specification,17th report. Rome, Food an Agriculture Organization.
Freeman, B. A., & Crapo, J. D. (1982). Biology of disease: Free radicals and tissueinjury. Laboratory Investigation, 47, 412–426.
Freitas, J. J., Pompeia, C., Miyasaka, C. K., & Curi, R. (2001). Walker-256 tumorgrowth causes oxidative stress in rat brain. Journal of Neurochemistry, 77, 655–663.
Gumuslu, S., Akbas, H., Aliciguzel, Y., Agar, A., Kucukatay, V., & Yargicoglu, P.(1998). Effects of sulfur dioxide inhalation on antioxidant enzyme activities in raterythrocytes. Industrial Health, 36, 70–73.
Gunnison, A. F., & Palmes, E. D. (1973). Persistence of plasma S-sulfonates followingexposure of rabbits to sulfite and sulfur dioxide. Toxicology and AppliedPharmacology, 24, 266–278.
Hayatsu, H., & Miller, R. C. (1972). The cleavage of DNA by the oxygen-dependentreaction of bisulfite. Biochemical and Biophysical Research Communications, 46,120–124.
Hayes, J. D., & McLellan, L. I. (1999). Glutathione and glutathione-dependent enzymesrepresent a co-ordinately regulated defence against oxidative stress. Free RadicalResearch, 31, 273–300.
Ji, L. L. (1995). Exercise and oxidative stress: Role of the cellular antioxidant systems.Exercise and Sport Sciences Reviews, 23, 135–166.
Johnson, J. L., & Rajagopalan, K. V. (1976). Purification and properties of sulfite oxidasefrom human liver. The Journal of Clinical Investigation, 58, 543–550.
Int J
Neu
rosc
i Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
ity o
f N
ewca
stle
on
03/1
7/14
For
pers
onal
use
onl
y.
982 V. KUCUKATAY ET AL.
Johnson, J. L., Rajagopalan, K. V., & Cohen, H. J. (1974). Molecular basis of thebiological function of molybdenum. Effect of tungsten on xanthine oxidaseand sulfite oxidase in the rat. Journal of Biological Chemistry, 249, 859–866.
Kletzien, R. F., Harris, P. K., & Foellmi, L. A. (1994). Glucose-6-phosphatedehydrogenase: A “housekeeping” enzyme subject to tissue-specific regulationby hormones, nutrients, and oxidant stress. FASEB Journal, 8, 174–181.
Kucukatay, V., Savcioglu, F., Hacioglu, G., Yargicoglu, P., & Agar, A. (2005). Effectof sulfite on cognitive function in normal and sulfite oxidase deficient rats.Neurotoxicology Teratology, 27, 47–54.
Lee, M. Y. (2002). Effects of Na(2)SO(3) on the activities of antioxidant enzymes ingeranium seedlings. Phytochemistry, 59, 493–499.
Lowry, O. H., Rosenbrough, N. J., Far, A. L., & Randel, R. J. (1951). Proteinmeasurement with folin-phenol reagent. Journal of Biological Chemistry, 193,265–275.
Meng, Z., Sang, N., & Zhang, B. (2002). Effects of derivatives of sulfur dioxide onmicronuclei formation in mouse bone marrow cells in vivo. Bulletin of EnvironmentContamination and Toxicology, 69, 257–264.
Michiels, C., Raes, M., Toussaint, O., & Remacle, J. (1994). Importance of Se-glutathione peroxidase, catalase, and Cu/Zn-SOD for cell survival against oxidativestress. Free Radical Biology and Medicine, 17, 235–248.
Misra, H. P., & Fridovich, I. (1972). The role of superoxide anion in the autoxidationof epinephrine and a simple assay for supeoxide dismutase. Journal of BiologicalChemistry, 247, 3170–3175.
Mottley, C., Trice, T. B., & Mason, R. P. (1982). Direct detection of the sulfurtrioxide radical anion during the horseradish peroxidase-hydrogen peroxideoxidation of sulfite (aqueous sulfur dioxide). Molecular Pharmacology, 22, 732–737.
Mudd, S. H., Irreverre, F., & Laster, L. (1967). Sulfite oxidase deficiency in man:demonstration of the enzymatic defect. Science, 156, 1599–1602.
Ozawa, T., & Hanaki, A. (1987). Spin-trapping of sulfite radical anion, SO3-., by awater-soluble, nitroso-aromatic spin-trap. Biochemical and Biophysical ResearchCommunications, 142, 410–416.
Paxinos, G., & Watson, C. (1986). The rat brain in stereotaxic coordinates, 2nd ed,Academic Press, San Diego.
Schafer, F. Q., Qian, S. Y., & Buettner, G. R. (2000). Iron and free radical oxidations incell membranes. Cellular and Molecular Biology, 46, 657–662.
Shi, X. (1994). Generation of SO3.- and OH radicals in SO3(2-) reactions with inorganicenvironmental pollutants and its implications to SO3(2-) toxicity. Journal ofInorganic Biochemistry, 56, 155–165.
Taniguchi, N. (1992). Clinical significances of superoxide dismutases: Changes in aging,diabetes, ischemia, and cancer. Advances in Clinical Chemistry, 29, 1–59.
Int J
Neu
rosc
i Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
ity o
f N
ewca
stle
on
03/1
7/14
For
pers
onal
use
onl
y.
ACTIVITIES IN SULFITE OXIDASE COMPETENT AND DEFICIENT RATS 983
Taylor, S. L., Higley, N. A., & Bush, R. K. (1986). Sulfites in foods: Uses,analytical methods, residues, fate, exposure assessment, metabolism, toxicity, andhypersensitivity. Advances in Food Research, 30, 1–76.
Tejnorova, I. (1978). Sulfite oxidase activity in liver and kidney tissue in five laboratoryanimal species. Toxicology and Applied Pharmacology, 44, 251–256.
Woo, W. H., Yang, H., Wong, K. P., & Halliwell, B. (2003). Sulphite oxidase geneexpression in human brain and in other human and rat tissues. Biochemical andBiophysical Research Communications, 305, 619–623.
Yargicoglu, P., Gumusluoriob, S., Agar, A., Korgun, D. K., & Kucukatay, V. (2001).Effect of sulfur dioxide inhalation on erythrocyte antioxidant status, food intake,and lipid peroxidation during aging. Archives of Environment Health, 56, 53–57.