Biol Cell (I 996) 86,67-12 f3 Elsevier, Paris 67 Original article Effects of different vertebrate growth factors on primary cultures of hemocytes from the gastropod mollusc, HaZiotk tubercukzta Jean-Marc Lebel, Wilfrid Giard, Pascal Favrel, Eve Boucaud-Camou L&orato&e de Biobgie et Biotechnobgies Marines, IBBA, IFREMER URM 14, Universit6 de Caen, Esplanade de la Paix, 14032 Caen cedex, France (Received 6 January 1996; accepted 12 March 1996) Summary - A useful experimental system from primary cultures of hemocytes from H&Otis tuberculutu has been established. Six days after initiation of the culture, the viability of hemocytes remained constant as measured by the M’IT assay. In addition, hemocytes showed physiological responsesas judged by protein and DNA synthesesin response to treatment with vertebrate growth factors. Por- cine insulin and human epidermal growth factor (EGF) stimulated [jH]-leucine and [3H]-thymidine incorporation in hemocytes in a dose-dependent manner. No additive effect of insulin and EGF is observed either for PHI-leucine or for [3H]-thymidine incorporation. The response of primary cultures of abalone hemocytes to vertebrate growth factors confirms their growth potential in vitro and provides a suitable model for further studies on regulation of the control of cellular processes such as cell growth, differentiation and migration in invertebrate cells. moliusc / hemocytes / cell culture / growth factors Introduction In vertebrates as well as in insects, primary culture metho- dology has opened up new approaches for studying cellular and molecular events occurring during control of cell pro- liferation, cell differentiation or cell metabolism. Since the sixties many attempts have been made to establish long-term primary cultures of marine mollwcan cells [35]. However, despite numerous investigations testing a wide variety of growth media, supplements and cultural conditions, very lit- tle progress has been made [ 191. A major difficulty concerns the low rate of cell proliferation in vitro, even for tissues with high mitotic potential such as embryonic or larval tis- sues [24]. The lack of appropriate culture media, especially the absence of growth promoting substances, is the main reason of fails to obtain continuous cell multiplication in vitro [22]. Supplementing the basal medium culture with homologous hemolymph or with extracts from cerebral gan- glia or genital glands, perhaps a source of mitotic promoting factors, has not significantly improved results [7, 10, 191. Interestingly, the presence of fetal calf serum as well as ver- tebrate growth factors in the culture medium increased the viability of a wide variety of marine invertebrate cells including marine molluscan cells [1, 7, 241, suggesting that vertebrate growth-promoting factors may be efficient for these cells as is the case for cultured insect cells [ 171. In invertebrates, the presence of substances homologous to vertebrate growth-promoting factors has been strongly suggested, as a result of immunocytochemical and biochem- ical investigations. Some of these growth factors belong to the family of the insulin-like peptides (ILPs) [33]. ILPs, which have been detected in the digestive system and the brain of different rnolluscs, present similarities with mam- malian insulin at both the structural and functional levels [8]. Furthermore, in the pulmonate mollusc Lymnaea stugnuks, cDNAs encoding a protein with characteristics of vertebrate preproinsulins and called molluscan insulin-related peptides (MIPS), have been identified in growth-controlling neurons suggesting a possible role of MIPS in body growth [29, 301. According to these results, the involvement of ILPs in body and shell growth and energy metabolism has been estab- lished in molluscs [28]. Another interesting growth-factor family has also been found in invertebrates. This family is composed of members analogous to vertebrate polypeptidic growth factors such as epidem& growth factor (EGF). The presence of EGF-like peptides in invertebrates was first documented with the identification of specific nucleotide sequences closely homologous to vertebrate EGF and/or EGF-receptor gene in Drosophikz [l& 27, 371, Caenorhub- ditis [13], different sea urchin species [5, 14, 39, 401 and more recently My?& [ 151. In agreement with this hypothe- sis, an EGF-like factor has been found in the tissues of the mussel Mytiks edulis [25]. In addition, Cancre et ul [3], recently demonstrated the presence of an EGF-like sub- stance in hemocytes of the crustacean Palaemon serrutus. Invertebrate hemocytes constitute a population with mor- phological and functional heterogeneity which has been ascribed to their degree of diffe=ntiation [36]. Among their diverse functions is a major role in the defence mechanisms, though they are also involved in wound healing and have the ability to secrete components of the extracellular matrix (EMC) in vitro [32]. Another characteristic of these cells concerns their capacity to divide in viva [6]. Taking into account these characteristics, it seems likely that hemocytes could provide a suitable model to investigate the effects of growth-promoting factors involved in the process of prolife- ration-differentiation. Up to now, no reports have appeared concerning the effects of growth-promoting factors on pro- tein and DNA syntheses in primary cell cultures of marine invertebrates. The purpose of this present study was thus to
Transcript
Biol Cell (I 996) 86,67-12 f3 Elsevier, Paris
67
Original article
Effects of different vertebrate growth factors on primary cultures of hemocytes
from the gastropod mollusc, HaZiotk tubercukzta
Jean-Marc Lebel, Wilfrid Giard, Pascal Favrel, Eve Boucaud-Camou
L&orato&e de Biobgie et Biotechnobgies Marines, IBBA, IFREMER URM 14, Universit6 de Caen, Esplanade de la Paix, 14032 Caen cedex, France
(Received 6 January 1996; accepted 12 March 1996)
Summary - A useful experimental system from primary cultures of hemocytes from H&Otis tuberculutu has been established. Six days after initiation of the culture, the viability of hemocytes remained constant as measured by the M’IT assay. In addition, hemocytes showed physiological responses as judged by protein and DNA syntheses in response to treatment with vertebrate growth factors. Por- cine insulin and human epidermal growth factor (EGF) stimulated [jH]-leucine and [3H]-thymidine incorporation in hemocytes in a dose-dependent manner. No additive effect of insulin and EGF is observed either for PHI-leucine or for [3H]-thymidine incorporation. The response of primary cultures of abalone hemocytes to vertebrate growth factors confirms their growth potential in vitro and provides a suitable model for further studies on regulation of the control of cellular processes such as cell growth, differentiation and migration in invertebrate cells.
In vertebrates as well as in insects, primary culture metho- dology has opened up new approaches for studying cellular and molecular events occurring during control of cell pro- liferation, cell differentiation or cell metabolism. Since the sixties many attempts have been made to establish long-term primary cultures of marine mollwcan cells [35]. However, despite numerous investigations testing a wide variety of growth media, supplements and cultural conditions, very lit- tle progress has been made [ 191. A major difficulty concerns the low rate of cell proliferation in vitro, even for tissues with high mitotic potential such as embryonic or larval tis- sues [24]. The lack of appropriate culture media, especially the absence of growth promoting substances, is the main reason of fails to obtain continuous cell multiplication in vitro [22]. Supplementing the basal medium culture with homologous hemolymph or with extracts from cerebral gan- glia or genital glands, perhaps a source of mitotic promoting factors, has not significantly improved results [7, 10, 191. Interestingly, the presence of fetal calf serum as well as ver- tebrate growth factors in the culture medium increased the viability of a wide variety of marine invertebrate cells including marine molluscan cells [1, 7, 241, suggesting that vertebrate growth-promoting factors may be efficient for these cells as is the case for cultured insect cells [ 171.
In invertebrates, the presence of substances homologous to vertebrate growth-promoting factors has been strongly suggested, as a result of immunocytochemical and biochem- ical investigations. Some of these growth factors belong to the family of the insulin-like peptides (ILPs) [33]. ILPs, which have been detected in the digestive system and the brain of different rnolluscs, present similarities with mam- malian insulin at both the structural and functional levels [8]. Furthermore, in the pulmonate mollusc Lymnaea stugnuks,
cDNAs encoding a protein with characteristics of vertebrate preproinsulins and called molluscan insulin-related peptides (MIPS), have been identified in growth-controlling neurons suggesting a possible role of MIPS in body growth [29, 301. According to these results, the involvement of ILPs in body and shell growth and energy metabolism has been estab- lished in molluscs [28]. Another interesting growth-factor family has also been found in invertebrates. This family is composed of members analogous to vertebrate polypeptidic growth factors such as epidem& growth factor (EGF). The presence of EGF-like peptides in invertebrates was first documented with the identification of specific nucleotide sequences closely homologous to vertebrate EGF and/or EGF-receptor gene in Drosophikz [l& 27, 371, Caenorhub- ditis [13], different sea urchin species [5, 14, 39, 401 and more recently My?& [ 151. In agreement with this hypothe- sis, an EGF-like factor has been found in the tissues of the mussel Mytiks edulis [25]. In addition, Cancre et ul [3], recently demonstrated the presence of an EGF-like sub- stance in hemocytes of the crustacean Palaemon serrutus. Invertebrate hemocytes constitute a population with mor- phological and functional heterogeneity which has been ascribed to their degree of diffe=ntiation [36]. Among their diverse functions is a major role in the defence mechanisms, though they are also involved in wound healing and have the ability to secrete components of the extracellular matrix (EMC) in vitro [32]. Another characteristic of these cells concerns their capacity to divide in viva [6]. Taking into account these characteristics, it seems likely that hemocytes could provide a suitable model to investigate the effects of growth-promoting factors involved in the process of prolife- ration-differentiation. Up to now, no reports have appeared concerning the effects of growth-promoting factors on pro- tein and DNA syntheses in primary cell cultures of marine invertebrates. The purpose of this present study was thus to
establish a suitable primary cell culture of hemocytes and subsequeptIy to investigate the effects of growth promoting factors such as insulin and EGF on the incorporation of [3H]-leucine and of [3H)-thymidine in order to estimate pro- tein and DNA syntheses in these cultured cells.
Materials and methods
Source and maintenance of anitnuis
Adult abalones, Haiiotk tuberculata, 8-12 cm in shell length, were collected monthly from the West Coast of the Cotentin peninsula (Manche, France) and acclimated to iaboraiory condi- tions for at least 2 weeks before experimentation. Animals were Faintained in natural and continuoutiy aerated seawater at sea.. sonal ambient temperature. They were fed daily with a mixed algal diet but starved 2 days prior to being killed.
Primaty cell cultures
After an incision in the foot, hemolymph was collecte4. (S-10 ml per animal) using a 5-ml st.eriIe syringe fitted with a Sgauge hypo- dermic needle. Hemolymph was transferred to a sterile? tube and simultane5usly diluted 1:3 in cooled sterile anti-coagulmt m5&
fied Alsever’s solution [2] (115 &- glucose; 27 i&l @iurn iit- rate; 11.5 n&l EDTA; 382 mM NaCl). Ijemocytes were rapidly plated at 0.8 Iti cells per well in &well culture plate3 to which three volumes of sterile artiticiaI seawater were added. Cultures were maintained at 15°C in a humidified incubator (~COX-free). After 90 min of incubation, cells were washed with Hanks’-199 medium modified by addition of 250 mM NaCl; 10 mM KCl; 25
mM MgSOA; 2.S mh4 .CaCJ; 10 mM Hepes: final pH was 7.4 im& osmolarity 1100 mosmol/l. Then cells were covered with fresh medium supplemented with antifungal and antibacterial substance5 t 100 pg/ml streptomycin sulfate; 60 wml~penicillin G: -5O-$g/mt gentamycin &fate; 0.20 .ug/ml amphotericin By and nystatin (8 pg/ml), L-glutamine (2 mM), concahavalin A (2 mM) and w&c incubated (CO?-free) at 1 S’C.
MIT reductioti ussay
The M’M red?ction assay is an enzymabc test based on the aetcr. mination *f the activity of mitochtindrial deshydrogenasc enzymes- This test, developed by Mosmann [23], h&been adapted to evaluate the viability of marine invertebrate cells i?? 24i. F5r assays> I?0 ~1 of stock MTT solution (5 mg Ml-r/ml artificial seawater filtered throygh a O-22. pm filter) were addee to each dish to be tested.. ~Plates were incuiated at i5YJ. Reaction? were stopped by addition of an equal volume of isopropanoi COH.
taining 0.04 N HCl. The plates were shaken for 30 min at room temperature. Absorbances at wavelength of 570 nm were meas.. ured with a reference of 630 nm (OD 570 nm/ref 630 nm) within 30 tin of adding the isopropanol.
Growth factors assq
Cells were plated at 0.8 106 cells per well in &well culture plates. 24 b. after the beginning of c.ulture the mediu& was renewed (l-2 ml p& dish) without loss of dells. Then 10 p$ (I PCi) of f3H]-thymidine (sp act: 35 Ci/mmol, ICN r?di.ochemi- cal diluted in culture mechum) or [sH]-leucine (sp act: 12! Ci/mmol, ICN radiochemical .diluted in culture medium) was added to each dish. Cultures were perfomed in-absence or ia presence of the different growth factors to be tested for 24 h
Fig I99
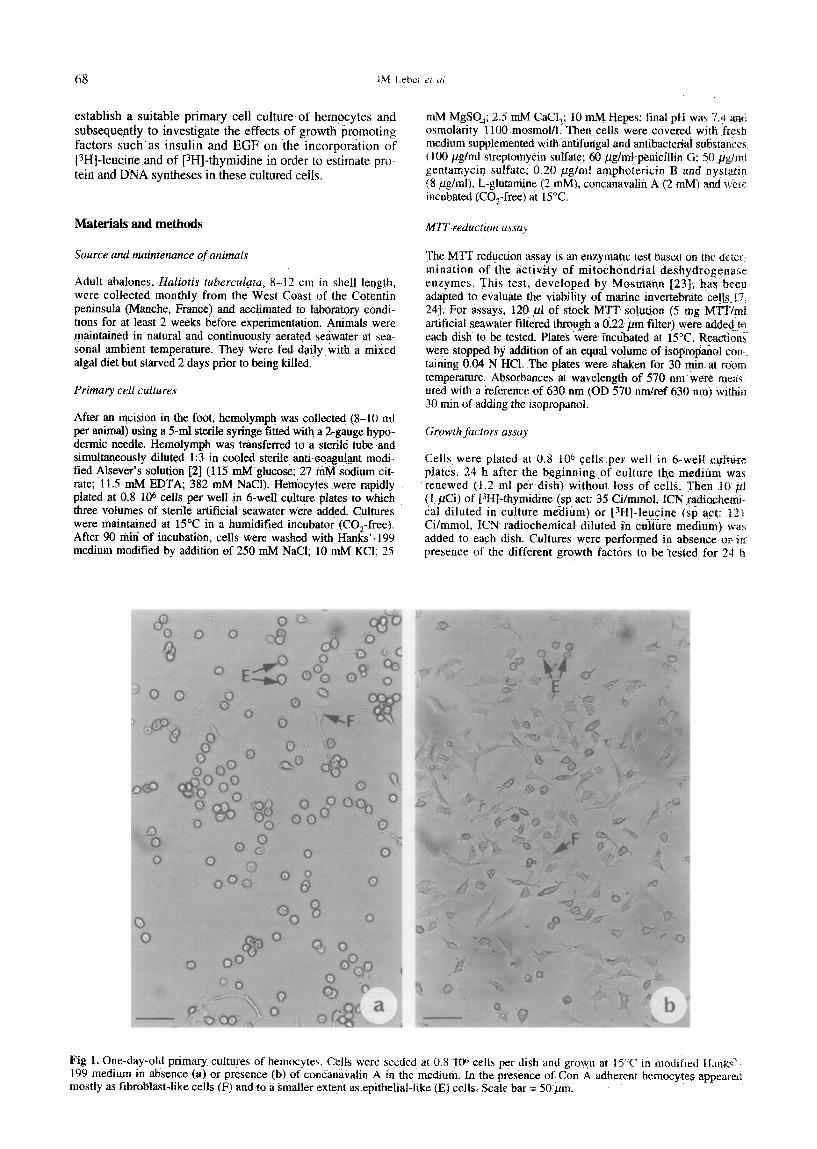
1. One-day-old primary cultures of bemocytes. Cells were seeded at 0.8 I@ cells per dish and grown at 1S”C in modified H&:s’ medium in absence (a) or presence (b) of conc%navalin A in the medium. In tie presence of Con A adherent hemocytes appcAr+.?d tly as fibroblast-like cells (F) and to a smaller extetit as epitbelial-like (E) cells. Scale bar = 5O~prn.
A test system from H tuberdata 69
([3H]-leucine) or 48 h (f3H]-thymidine). Bovine insulin and human epidermal growth factor (EGQ were dissolved in Hanks’- 199 supplemented medium and 10 ~1 of stock solutions to be tested were added to the wells. Incorporation was stopped by adding an equal volume of ice-cold trichloroacetic acid (10%). After total precipitation (overnight at 4°C) cells were scrapped and transferred in tubes which were centrifuged at 3000 g for 10 min. The supematant was discarded and the pellets were washed twice with 10% trichloroacetic acid. Then 500 ~1 of KOH (0.3 M) were added and tubes were incubated for 30 min at 60°C in order to dissolve the pellet. The total fraction was transferred in scintillation vial and the radioactivity was counted after addition of 4 ml of liquid scintillation to each vial. Blank controls were performed with cells that were precipitated with trichloroacetic acid prior to the addition of r3H]-leucine or [3H]-thymidine.
Stutistical analysis
Significance of the difference between mean values was estimated using the Student’s t-test. Each experiment was repeated at least three times and for one experiment the means were calculated from triplicates.
ResUlt.5
Primary cultures qf hemocytes
In absence of concanavalin A (Con A) a large part of hemo- cytes remained round like freshly bled cells (fig la). A con- sistent feature of hemocytes cultures grown in Con A-free medium was a significant increase in non-adherent floating cells in the culture supematants during experiments (data not shown). The presence of Con A in the medium (1 pg/ml) enhanced the adherence of hemocytes on the plates. After the initiation of cultures, hemocytes adhered on glass and formed clusters. Rapidly cells migrated out of the aggregates and flattened on the surface of dishes. Hemocytes appeared more and more isolated and then formed a uniform monolayer (fig lb). Based on their mor- phological characteristics, two cell categories could be dis- tinguished. A large majority of hemocytes is represented by the fibroblast-like cells. The remainder is constituted by the epithelial-like cells (fig la, b).
MT assay and viability of hemocytes
Figure 2 represents the validity of the MTT assay test for hemocytes. The absorbance was directly proportional to the number of cells cultured per dish and this linear function (y = 0.009 x + 0.009, r* = 0.975) included the greater value tested, 0.9 106 cells per dish. The viability of hemocytes remained constant during the fast 6 days of culture (data not shown).
Efects of insulin or EGF on [3H]-leucine incorporation
Insulin and EGF significantly stimulated PHI-leucine incor- poration in hemocytes in a dose-dependent manner from 10-T M to 10-5 M for insulin (fig 3) and from lO-7 M to 10-6 M for EGF (fig 4). The maximal stimulation induced an increase of 196 k 4% for insulin and 207 & 12% for EGF with respect to 100% of control. The combined effect of both insulin and EGF is illustrated in figure 5. No additive effect on [3H]-leucine incorporation is observed for insulin (104 M) and EGF (lO-7 M). For these concentrations the combined response (227 & 15% relative to the 100% meas- ured for control) is not significantly higher than the
OS 10
1 O5 cells / well
Fig 2. Validity of the MTT reduction assay. One day after the beginning of cultures the medium was renewed and M’IT test was performed. Hemocytes were plated at a density from 0.15 106 to 0.9 l@ cells per well in 6-well culture plates and cultured at WC in modified Hanks’-199 medium. Each data point (OD 570 nrn/ref 630 nm) represents the mean 2 standard deviation of triplicate cultures.
response of insulin (104 M) or EGF (lo-7 M) tested alone, respectively 210 ? 9% and 198 k 10%. [3H]-leucine incor- poration measured for control represented 1283 * 125 cpm/l@ cells.
Effects of insulin or EGF on [3H]-thymidine incorporation
Addition of insulin and EGF significantly stimulated [3H]- thymidine incorporation in hemocyte culture (fig 6). How- ever, no additive effect on the incorporation of [YHJ-thymi- dine is observed when a combined treatment with insulin (10 -6 M) and EGF (10-T M) is performed. The combined response (224 k 8% with respect to the 100% measured for control) is not significantly higher than the response of insu- lin (lo-6 M) or EGF (10-T M) tested alone, respectively 194 * 9% and 213 k 10%. [3H]-thymidine incorporation meas- ured for control represented 602 ? 15 cpm/l@ cells.
Discussion
The first aim of this work was to establish a suitable in vitro model for studying the cellular metabolism of hemocytes. As previously observed for sea-urchin embryonic cells, the presence of Con A in the medium enhanced hemocyte adherence on the plates [21]. In our study, cells attached to the surface of the plates after about 1 h of incubation and remained fully spread during the 6-day experiments. Thus hemocytes may be seeded without cell loss and in a most reproducible manner resulting in a highly quantifiable
CO -10 -9 -8 -7 -6 .-5
k!@
Fig 3. Dose-response effect of insulin on the incorporation of [SHJ-Ieucine in.heinocytes in culture. Ceils were seeded at 0.8 106 cells per dish and grown in absence or presence tif insulin (from l@-*‘J M to I@5 M) for 24 h. Cells were cultur.%d at WC! in modi- fied Hanks’-199 medium. Each data point repssents the mean percentage * standard deviation (with respect to the 100% of incorporation- measured for control) of &$&ate cuItures. Such a typical experiment was repeatedat least three times. Sign&ant difference from control cells (Co) at P < W301 (**).
system. This first point is an original aspect of our model. Con A, which tiects cell attachment, has b&n also shown to act on cell proliferation for different cell types. Because aI cell cultures were carried out in the preseti@ of Con A, this possible influence has not been investigated. In addition, for marine molluscan cells, Domart-Coulon er uZ [7J did not observe any significant influence of Con A on the viability of cell cultures either after 2 or after 6 days of incubation for concentrations corresponding to 1 pg/ml or 25 @@ml. How- ever, we call not rule out an @feet of Con A on thyddine incorporation .in cultured hemocytes. The viability of henio- cytes cultur& in a medium without growth fac@rs remaibe4l constant during the 6 fast diys of culture. In &&ion, the cells showed physiological responses as jtidg&d by protein and DNA syntheses in response to treatment witbinsulin or EGF, confirming their gruwth potential in V&RX
Treatment with porcine in&in stim&ated incorporation of labelled leucine and thymidine in cultured hemocytes, reflecting to a certain extent an increase in the protein and DNA syntheses. These effects are significant for about l @/ml and maximal for 100 fig/ml. EffecEs of vertebrate insulin on other cell types from .molluscs have .been reported. In Htdisomu &ryi, treatment with porcine insulin (0.1 @ml during 48 h) increased amino acid incorporation.in the man- tle collar jn vi?ro [28]. Recently, Domwd-Coulon et ul [‘7] reported that 6 days of treatment with bovine insulin (XI pg/ml) induced a 25% rise in celIulztr viabiliti:in heart cells from Crussos~rea g&as in vitr.. 1~~expetimen.t~ conducted with dissociited cells we showed that insuW (50 pg/ml)
co -9 -8 “7 “4
Log M
Fig 4. Dose-response effect of EGF on the incorporation of [JH]* leucine in hemocytes in culture. Cells were seeded &t 0.8 W cells pr dish and grown ig absence or p=sence of.EGF (from IV M to lv-M) for 24 h. Cells were cultured at WC in rqdi- fie$ Hanks’-l9~~m~d&m. Each data point rqzsents the-&can percentage * standard deviation (with respect to the. l~OU% of incorporation m&sured for control) of triplicate-cultures. &ch a typical expzriment was rested at least three times: &gnificmt~ difference fr&ti controf ceils (Co) at P < 0.001 (**).
incr?ases incorpQration of labelled leucine in mantle cells, but also in di@tive bells from the molIusc Pec~en-muximy (Giard et ai, subm@ed). These results dem~strated that ver- tebrate insulins have biological effects on a large va&ty of invertebrate cell types but the precise functiti ofthis growth factor on these different ce& remains to be- elucida&. Usirig porcine or human insulin antisera, immunoreactive in&in7 like peptide (l&P) was detected in the neurosecretory medio- dorsal cells and hemolymph of the snail, He&om~&q+ [ 16,281, I&Ps appear to arise from &&rent soqces, such -as the digestive gland or the central wwous system, $zvala et u2 [28] suggested that lLP from the digestive gland could IX invalved in sugar met&&sm, whereas ILP from the central nervous system.could be a shelXgro* f&tor. In the case of hemocytes no eiid&nce is available to date p&&t&g a &s- tinction betwe& insulin being a .general met&o& &n&a- tor or having a. more specif?c fumztion.
The other stimulating growth factor tested, EGF”: increased significantly (at a cmcentition of 0.6 &ml) the. .metab& a&@-of ct.&red hemocytes. In the same way? Domart-Coulon et ui [7] showed that treatment with this poIypeptidic -gFowth factor has a significant posit& effect on the viability of Crossasfrea~ gigtis heart cell cultures+ A biological Effect of EGF on maririe invertebrate cells is in agreemerit with results of Odinstova et ul [25]. Th&e authors have extracted a substance which may belong t@ the fatiily of EGF&ke factors from Myfz’Zus edulzk .‘l%is facto1 showed a mitogqk activity in dissociated mussel m&k cells as w&tis in mouse fibro.bktsts. A mitogenic factor
.4 test system from H tuberdata 71
co E I E+I
Fig 5. Combined effect of insulin and EGF on the incorporation of [sH]-leucine in hemocytes in culture. Cells were seeded at 0.8 106 cells per dish and grown in absence or presence of EGF (10-7 M) or insulin (1V M) alone; or with EGF (10-T M) and insulin (10-6 M) simultaneously, for 48 h. Cells were cultured at 15T in modilied Hanks’-199 medium. Each data point represents the mean percentage k standard deviation (with respect to the 100% of incorporation measured for control) of triplicate cultures. Such a typical experiment was repeated at least three times. Signi- ficant difference from control cells (Co) at P < 0.001 (**).
able to regulate the metabolism of mantle cells has also been extracted from cerebral ganglion and hemolymph of the mussel Mytilus eduZis [20, 341. The presence of a mito- genie factor in hemolymph is in agreement with an endo- crine function of these growth factors. Other possibilities, however may be postulated. Recently, a protein, mainly consisting of EGF-like repeats, has been detected in the foot of the mussel MytiZus guZZoprovincialis [ 151. Interestingly, it seems that this protein functions as a matrix protein. In vertebrates, several extracellular matrix proteins contain domains with homology to EGF and exhibit growth pro- moting activities including mitogenic activity [ 111. It has been suggested that diffusible EGF-like peptides may be released by processing of the extracellular domain. These proteins of the ECM may express their growth factor-like function on neighbouring cells by paracrine stimulation. They may also act on the cells that secrete it by autocrine stimulation [1 11. Taking into account the fact that hemocy- tes secrete a lot of components of the EMC [31], the in vivo effects of some growth promoting factors on hemocytes may result from a paracrine as well as autocrine stimula- tion.
In this study, despite an effect of insulin and EGF on the stimulation of DNA synthesis, the induction of cell prolife- ration is not demonstrated. However, the capacity of hemo- cytes to divide in vitro has been demonstrated by Dikke- boom er aZ [6]. In the same way, division in vitro of cells from larvae of the abalone, HuZiotis rufescens was evi- denced [24]. No additive effects have been evidenced
CO E I E+I
Fig 6. Effects of EGF and insulin on the incorporation of [sH]- thymidine in hemmytes in culture. Cells went seeded at 0.8 106 cells per dish and grown in absence or presence of EGF (10-T M) or insulin (1tV M) alone; or with EGF (10-T M) and insulin (10-6 M) simultaneously, for 48 h. Cells were cultured at 15°C in modi- fied Hanks’-199 medium. Each data point represents the mean percentage i standard deviation (with respect to the 100% of incorporation measured for control) of triplicate cultures. Such a typical experiment was repeated at least three times. Significant difference from control cells (Co) at P c 0.001 (**).
between the two tested growth factors either for protein or DNA syntheses. In vertebrates, it is now well established that insulin and EGF induce their biological effects through interactions with their cell surface receptors that contain intrinsic tyrosine kinase activities [4, 381. Moreover, in vivo stimulation by insulin and EGF may overlap in term of their intracellular signalling pathways [12, 381. F’rotein tyrosine kinases signalling pathways have been conserved throughout evolution [9, 261. In addition, in DrosophiZu a receptor that shows a dual binding specificity for both insulin and EGF has been identified [32]. The lack of addi- tive effects observed in this study may result from the expression of a such protein in marine molluscan cells.
In conchtsion, we have established a routine experimental system of primary hemocyte cultures. Hemocytes are target cells for vertebrate growth factors such as insulin or EGF which increased hemocyte protein as well as DNA synthe- ses. This model appears to be particularly useful for investi- gating the in vitro effects of growth factors on the control of cell growth, differentiation and migration at the cellular level in invertebrate cells.
Acknowledgments
We are very grateful to Professor E Lopez for encouragement and helpful advice on the manuscript. This study was supported by a special grant of the University of Caen (Action specifique recherche). A doctoral fellowship (W Giard) from the Dielen society is gratefully acknowledged.
References
I
2
3
4
5
6
7
8
9
10
11
12
13
14
IS
16
17
18
19
20
Auzoux S, Domart-Coulon 1, Doumenc D ( I9931 G111 cell cultures of the butter-fish clam Ruditapes decussatus. .I Mar BiotechnoI 1,79-81 Bach&e E, Chagot D, Grizel H.(l988) Separation of Crawostrw gigas hemocytes by density gradient centtifugation and counter. flow centrimgal ehnriation. & Camp Immune 112,549-5s9 Cancre I, Van-Wormhoudt A, Legal Y (1995) Effects of ccl- lular growth factors on crustacean hepatopancreas ceil sus pensions. J Mar Biotechnol2, 83387 Carpenter G, Cohen S (1990) Epidermai growth factor. ./ &<J/ Chem 265,7709-77 12 Delgadillo-Reynoso MG, Rollo DR, Hursh DA, Gaff RA (1989) Structural analysis of the pEGF gene in the sea urchin Stongylocentrotus purpurus reveals more similarity to verte- brate than to invertebrate genes with EGF-like repeats. .r h&)1 Evol29,3 14-327 Dikkeboom R, van der Knaap WPW, Meuleman EA, Sminia T (1984) Differences between blood cells of juvenile and adult specimens of the pond snail Lymnaea stagnalis. Cc+ Tissue Res 238,43-47 Domart-Coulon I, Doumenc D, Auzoux-Bordenave S, IX Fichant Y (1994) Identification of media supplements that improve the viability of primarily ceil cultures of Crassos- trea gigas oysters. Cytotechnologv 16. 109-120 Ebb&r& RH, Smit AB, Van Mmnen J (1989’1 The insulin family: evolution of structure and function in vertebrates and invertebrates. Biol3ull 177. 176-182 Egan SE, Weinberg RA ‘(1993) The pathway to signal achievement. Nature 365,781-783 Ellis LL, Bishop, SH (1989) Isolation of cell lines with ltm. ited growth potential from marine bivalves. In: btvertebrare cell system applications, ~012 (Mitsuhashi J, ed) CRC Press Boca Raton, FL, 243-25 I Engel J (1989) EGF-like domains in extracellular matrix pro- teins: localized signals for growth and differentiation? FEBS Lett 251, l-7 Gingras AC, Donze 0 (1995) Regulation par l’insuhne de l’initiation de la synthese proteique. Medecine/Sci I j. 866-872 Greenwald I (1985) lin 12, a nematode homeotic gene* is homologous to a set of mammalian proteins that includes epi- dermal growth factor. Cell 43,583-590 Hursh DA, Andrews ME, Raff RA (1987) A sea urchin gene encodes a polypeptide homologous to epidermal growth fac- tors. Scjertce 237,1487-1490 Inoue K, Takeuchi Y, Miki D, Odo S (1995) Mussel adhesive plaque protein gene is a novel member of epidermal growth factor-like gene family. J Biol Chem 270,6698-6701 Khan HR, Griffond B, Saleuddin ASM (1992) Irnsulin-like peptide in the central nervous system of tie snail Helisoma duryi. Brain Res 580, 111-l 14 Landureau JC, Lenoir-Rousseaux JJ (1988} New culture media for insect cells. In: Invertebrate andfish tissue culture (Kuroda Y, Kurstak E, Maramorosch K, eds) Japan Scientific Societies Press and Springer-Verlag, 23-27 Livner E, Glazer L, Segal D, Schlessinger J, Shilo BZ (1985) The Drosophila EGF-receptor homologue: Conserva- tion of both hormone binding and kinase domains. Cell 40. 599-607 Machii A, Wada KT (1989) Some marine invertebrates tissue culture. In: Invertebrate cell system applications, ~012 (Mit- suhashi, J, ed) CRC Press, Boca Raton, FL, 225.233 Mathieu M, Lenoir F, Robbins i (1988) A genial mitosis- stimulating factor in cerebral ganglia and hemolymph of the marine mussel Mytilus edulis L. Gen Camp Endocrinol 72. 2S7-263
21
22
2.3
I?4
2s
26
.Y?
28
2Y
30
31
32
33
.34
3s
36
37
38
39
40
; l991) Adhesion of sea-urchin embryonic cells to substraL<r coated with cell adhesion molecules. Biol CelZ 7 1, 289-29 1 Mialhc E, Boulo V. Grizel H (1988) Bivalve mollusc cs:il Lbulture. American t~isheries Society Speciai Pubhcation I $. 31 I-315 Mosmann ‘f (iW.3~ Rapid colorimettie assay for cellular growth and survivah application to proliferation and cytotox icity assays. J lmmunol Methods 65, SS-63 Naganuma T. Degnan BM, Horikoshi K, Morse DE: (1994i Myogenesis in primary cell cultures from larvae of the aba lone* Haliotis rufescens. Mol Mar Rio1 Biotechnol3, 13 I- 14# Odintsova NA, Nesterov AM, .Korchagina DA (I 993-j ,A growth factor from tissues of the mussel MyIiius edut?is, Cornp Biochem Physiol 105A, 667-67 1 Perrimon N (1994) Signalliflg pathways inmated by receptor protein tyrosine kinases in Drosophila. Curr 0pin CeKBio/ 6.260-266 Rutiedge BJ, Zhang K, Bier l$ Jan YN, Perrmon N (l!>%> The drosophila spitz gene encodes a putative EGF-I&c growth factor involved in dorsal-ventral axis formations and nemogenesis. Genes Dev 6, 1503-I 5 f 7 Sevala VM, Sevala VL, Saleuddin ASM (1993) Hemolymph insulin-like peptides (ILP) titers .and the influence of KP and mammalian insulin on the amino acid incorporation in& mantle collar in vitro in Helisoma (Mollusca). &oI !&It 18.5, 140-148 Smit AB, Vreugdenhil E, Ebbetink RHM, Geraerts WPM. Klootwijk J, Joosse J ( 1988) Growth-controlling molluscan neurons produce the precursor of an insulin-related peptide Nature 331s S35--538 Smit AB, van Marie A, van ElkR, Bogerd J, van Heerikbu. izen H, Geraerts WPM (J993) Evolutionary conservation of the insulin gene structure in invertebrates: cloning of-tile gene encoding moIluscan insulin-related peptide 111 from l*ymnaeu sfagnahs. J Mel Endocrinot I I, 103-I 13 Suzuki T. Yoshinaka R, Mizma S, Funakoshi S, Wada h. t 1991) Extracellular matrix formation by amebocytes during epithelial regeneration in the pearl oyster Pinctada ,fucato Cell Tissue Res 266. 7582 Thompson KL, Decker SJ, Rosner MR (1985) ldentificatiori of’ a novel receptor in Drosophila for both epidermal growth factor and insulin. Proc Nat1 Acad Sci USA 82,8443:8447 Thorpe A, Duve H {1984) Insulin- and glucagon-like pep. tides in insects and molluscs. Mol Physiol5,2352260 Toullec JY, Lenoir F. Van-Wormhoudt A, -Mathieu M (1988f Nonspecies-specific growth factor from cerebral ganglia of Mytilus edulis. J Exp Mar Biol EcoI 119, 11 l-l 17 Vago C. Chastang S (1960) Cuhure de tissus d’huitr~~ C/t Acad Sci Paris 250, 275 l-27S3 Van der Knaap WPW, Adema CM, Sminia T (1993) invertc brate blood.cells: morphological and ftmctiorml aspects of the haemocytes in the pond snail Lymnaea .stagnalis. romp Hae- matol Jnt 3, 20-26 Wharton KA, Johansen KM, Xu T, Artavanis-Tsakonas !z; ( 1985) Nucleotide sequence from the neurogenic locus h&+r implies a gene product that shares homology with protein? containing EGF-like repeats. Cell 43, 567-581 White MF, Kahn CR (1994) The insulin signaling system, J Biol Chem 269, l-4 Yamasu K, Watanabe H, Kohchi C, Soma Gl, Mizuno Dl, Akasaka K. Shimada H, Suyemitsu T- Ishihara K (1993; Molecular cloning of a cDNA that encodes the precutsot- to several exogastrula-inducing peptides, epidermal- grow-th-factor-reIated polypeptides of.the sea .urchih Anthocidaris crassispina. Eur J 3iochem 228 5 1 S -523 Yang Q- Angerer LM, Angerer RC (1989) Unusual patterns of accumulation of mRNA encoding EGF-related protein m tea-m&in embryos. Science 246,-8&k-807