S. RAHA AND W. WESEMANNPhysiologisch-Chetnisches Institut II, Philipps-Universitat, D-355 Marburg, Lahnberge,Germany
SUMMARY
The effect of endotoxin treatment (3O;zg/kg body weight) on megakaryocyte development andplatelet production in guinea pigs was investigated. A moderate thrombocytopenia was noted at 24 hafter endotoxin administration. Recovery was observed at 48 h when the platelet count had almostreached the normal level. A significant increase in platelet production was obtained at48h, indicatedby an enhancement in incorporation of labelled sulphate into platelets and also by an increase in thenumber of circulating heavier platelets. Megakaryocytes were classified into different development-al stages - megakaryoblast, promegakaryocyte, megakaryocyte - using standard morphologicalcriteria (size, shape of nucleus, nucleus/cytoplasm ratio). The number of mature megakaryocytesincreased at 24 and 48 h after endotoxin treatment. It was possible to distinguish between twodifferent types of mature megakaryocytes - early mature and mature megakaryocytes - by thepresence of a methanol-sensitive acid phosphatase. This enzyme is found mainly in mature, presum-ably platelet-forming, megakaryocytes and in platelets. The increased number of maturemegakaryocytes at 24 h was due to an enhancement in the number of early mature megakaryocytes,whereas the change at 48 h was due largely to an increased number of mature, platelet-formingmegakaryocytes. Megakaryocytes were isolated from bone marrow of guinea pigs by Percoll density-gradient centrifugation and incubated with radioactively labelled leucine. Compared with theuntreated controls megakaryocytes isolated 24 h after endotoxin treatment took up and incorporatedhigher amounts of leucine into protein, indicating a higher metabolic capacity of the early maturemegakaryocytes. No alteration in the ploidy distribution of megakaryocytes was noted after endo-toxin treatment.
INTRODUCTION
Blood platelets are formed from large, polyploid bone-marrow cells, known asmegakaryocytes. The observation that changes in the circulating platelet levels mayresult in alterations of platelet production, has long suggested a homeostatic mechan-ism for thrombopoiesis (Harker & Finch, 1969; Harker, 1970). Investigators haveused several parameters such as an increase in platelet count and size and an enhancedincorporation of radioisotopes into platelets as indicators of stimulated throm-bopoiesis (Harker, 1970; Weiner & Karpatkin, 1972; Weintraub & Karpatkin, 1974;McDonald, 1980). The alterations in platelet production are widely accepted to be theresult of changes in megakaryocyte development (Ebbe, 1976). As a response to athrombopoietic stimulus, megakaryocytes are known to undergo several changes,such as increased ploidy, size and number, and an enhanced maturation rate (Ebbeet al. 19686; Ebbe, Stohlman, Donovan & Overcash, 1968a; Odell, Jackson, Friday& Charsha, 1969; Odell, Murphy & Jackson, 1976).
68 S. Raha and W. Wesemann
Endotoxin, a product of gram-negative bacteria, is known to stimulate throm-bopoiesis as measured by increased platelet production in laboratory animals (Alvinget al. 1979). In patients with blood-stream infections endotoxin causes intravascularcoagulation and shock (Wolff & Bennett, 1974). Circulating platelet levels in thesepatients are also depressed. Endotoxin-like activity has been detected in both experi-mental endotoxaemia and clinical gram-negative sepsis (Das, Schwarz & Folkman,1973). Studies in vitro have demonstrated that endotoxin could adhere to the surfaceof the platelet membrane and produce a change in shape and release of serotonin,adenine nucleotides and platelet factor 3 (Des Perez, Horowitz & Hook, 1961;Maclntyre et al. 1977; Ausprunk & Das, 1978). In vivo, the interaction betweenplatelets and endotoxin is indicated by thrombocytopenia and alterations in circulat-ing platelets, resembling that undergone during release reaction (Horowitz, DesPerez & Hook, 1962; Muller-Berghaus & Lohmann, 1974; Alvinge* al. 1979). Sinceadministration of endotoxin to laboratory animals increased platelet production onlyafter a minimum interval of 18 h, the involvement of a regulatory mechanism effectiveat the level of the megakaryocytes is assumed (Alving etal. 1979; Ramsey et al. 1980).Hence, in this study we investigated, in parallel, the influence of endotoxin treatmenton megakaryocyte differentiation and platelet production of guinea pigs.
MATERIALS AND METHODS
Chemicals and solutions
Endotoxin from Escherichia coli 005..B5 was kindly provided by Professor Dr Radsak,Medizinisches Zentrum fur Hygiene und Mikrobiologie, Philipps-Universitat, Marburg. En-dotoxin was dissolved in sterile 0-9% NaCl solution to give a final endotoxin concentration oflOOjUg/ml. L-[4,5-3H]leucine (52 Ci/mmol) and sodium ["S]sulphate (5 Ci/mg S) in physiologicalsaline, were obtained from Amersham Buchler, Braunschweig, West Germany. The sulphatesolution was diluted with sterile 0-9% NaCl to give a final concentration of 400^Ci/ml.
Modified Gaintner's solution (Gaintner, Jackson & Maynert, 1962) contained 0-089 M-NaCl,0-040 M-NaH2PO4-H2O, 0-0047 M - K H 2 P O 4 , 0-005 M-glucose, 0-0258 M-Na3citrate-2H2O and wastitrated with citrate or NaOH to pH 6-4 and 7-4, respectively. To obtain modified Gaintner's BSAsolution (Hagen-Aukamp, Wesemann & Aumuller, 1980) 0010 M-theophylline and 0-25 % bovineserum albumin (BSA) were added.
Percoll was purchased from Pharmacia, Uppsala, Sweden. Reagents for histochemicaldemonstration of acid phosphatase were obtained from Sigma Chemie GmbH, Munchen (Sigmaresearch kit no. 386A) and the incubation medium was prepared according to the specified direc-tions. The incubation medium contained cr-napthol-AS-BI-phosphate in 0-lM-acetate buffer(pH 50) and Fast Garnet GBC salt. The solution was freshly filtered before use.
Biochemical determinationsProtein was assayed according to Lowry, Rosebrough, Farr & Randall (1951). Radioactivity was
measured in a Beckman scintillation counter LS-3150 T with external standardization.
Animals
Male guinea pigs of mixed breeds, 350—500g, were obtained from a local supplier at least 1 weekprior to experimentation. They were housed under standard laboratory conditions (21 °C, 55%humidity) and given water and diet ad libitum (Sniff, Soest/Westfalen: Meerschweinchendiat).Animals were killed by cervical dislocation.
Endotoxin-stimulated thrombopoiesis 69
Endotoxin administrationEndotoxin from E. coli was applied as a solution of 100/ig/ml. Each guinea pig received a dose
of 30^g endotoxin/kg body weight as a single intravenous injection in the front paw. NaCl controlanimals received equal volumes of 0-9% NaCl administered by the same route and the controlanimals did not receive any treatment. The maximum volume of endotoxin or 0-9 % NaCl solutioninjected was 015 ml.
Isolation of platelets from human and guinea pig bloodBlood from healthy human donors was obtained by venipuncture and collected in 0-1 vol. of 3-8%
sodium citrate solution. The blood samples were centrifuged at SSO^mu at room temperature for15 min to obtain platelet-rich plasma (PRP). The PRP was further centrifuged at 4°C for 20minat 1 500 gmtx . A platelet button was obtained by carefully discarding the platelet-poor plasma (PPP).The platelet button was resuspended in 10 ml of 0-9 % NaCl solution and centrifuged again at 1500gm*x for 20 min, 4°C. The wash procedure was repeated twice. A platelet button containing about7-5—8-0 (X 109) platelets was used for acid phosphatase identification by gel electrophoresis.
For isolation of guinea pig platelets, blood was withdrawn from the heart of anaesthetisized guineapigs with a 10 ml plastic syringe fitted with a hypodermic needle 18 G (1-2 mm X 388 mm, B. Braun,Melsungen AG) into 0-125 vol. of 3-8% sodium citrate. The citrated blood was centrifuged at375jfniM at room temperature, for 15 min. The PRP was carefully drawn off and centrifuged at20 min at 4°C, at 1500#,n«x. The platelet button was washed as described for human platelets.Platelet buttons containing about 15 X 109 platelets per sample were used for the identification ofacid phosphatase after separation by disc gel electrophoresis.
Separation of guinea pig platelets into subfractionsCitrated blood was obtained from the heart of two anaesthesized guinea pigs, either untreated
(control) or treated with endotoxin 48 or 72 h prior to blood collection. After 15 min centrifugationat 375gmui at 4°C, plasma was withdrawn and kept at 4°C. To obtain a better yield of platelets, theremaining blood cells were mixed with modified Gaintner's medium (pH 6-4) and recentrifuged at375 giu,, The plasma and buffer fractions were pooled and 2 ml of this mixture were carefullylayered over Percoll gradients containing 1-5 ml each of the following densities: 1040, 1-055, 1-060,1-065, 1-070, 1075 and l-080g/cm3 (Fig. 1). After centrifugation at 5900#„„ (Beckman rotorSW40) at 4°C for 15 min, the different layers were carefully withdrawn. Platelets with adensity^ l-O55g/cm3 (subtraction 1), a density between 1055 and 1-065g/cm3 (subfraction 2),and a density between 1-065 and 1-075 g/cm3 (subfraction 3) were combined, respectively, andwashed twice by recentrifugation at 1500g^ for 15 min and resuspension in modified Gaintner'smedium (pH 6-4). Each subfraction was then suspended in 500^1 of modified Gaintner's medium(pH 7-4). The last density layer, 1-080 g/cm3, was found to contain mainly other blood cells and wasdiscarded.
The subfractions 1, 2 and 3 were processed further for protein analysis and ultrastructuralexamination.
Platelet countBlood collections were made by venipuncture of the marginal ear vein. The blood was diluted
immediately by addition of 10^1 of blood to 990/zl of 1-0 % ammonium oxalate solution. Blood wascollected from each guinea pig immediately before administration of 0-9 % NaCl or endotoxin andat definite intervals during the 72 h following treatment. However, a maximum number of threeblood samples was obtained from each guinea pig in order to avoid unwanted stress on the haema-topoietic system. Platelets were counted in a Neubauer chamber by phase-contrast microscopy.
In vivo [35S]sulphate incorporation into plateletsGuinea pigs received endotoxin intravenously (i.v.) (30/ig/kg of body weight) 18 h (group A) and
24 h (group B) prior to a single intraperitoneal (i.p.) injection of 0-25/iCi sodium [35S]sulphate/kgof body weight. Control animals received no other treatment than the i.p. injection of [3SS]sulphate,
S. Raha and W. Wesemann
Plasma
Lightplatelets
Intermediateplatelets
Heavyplatelets
Other bloodcells
Density
1040
1055
1060
1065
1070
1075
1080
Fig. 1. Subfractionation of guinea pig platelets on Percoll density gradient. PRP wascentrifuged at 5900gm^ at 4°C for 15min in a swinging-bucket rotor. Density is in g/cm3.
since in preliminary studies no difference was found between the two control groups. Blood wascollected either 24 or 48h after [3SS]sulphate injection; lOmin before blood was drawn, 2mlheparin, i.p. (lOOi.u./ml) were administered to each guinea pig. About 7 ml of blood was drawnfrom the heart of each animal and mixed with 1 ml of 1 % Na2EDTA solution in 0-7 % NaCl. Thismixture was diluted further with 4 ml of 0-9% NaCl solution and centrifuged at 1 \0gnit* for 40minat 4°C. The PRP-saline mixture was withdrawn carefully and centrifuged at 760gm±x at 4°C for30min. The platelet button was washed twice by resuspension with 4 ml 0'9% NaCl solution andcentrifugation at 760£mai at 4°C. After the final wash the platelet button was suspended in 0-5 ml0 9 % NaCl solution. A sample of 200^1 of this suspension was solubilized in 10ml of Ready-Solv1M HP scintillation cocktail prior to measurement of radioactivity in a liquid scintillationcounter.
Isolation of megakaryocytesMegakaryocytes were isolated from guinea pigs by a combination of density gradient and
elutriator centrifugation as previously described (Hagen-Aukampef a/. 1980). The guinea pigs wereeither untreated (control) or were administered endotoxin (30//g/kg) 18, 24, 48 or 72 h prior to
Endotoxin-stimulated thrombopoiesis 71
killing. Briefly, the isolation procedure included the following steps: the bone marrow of femora,tibiae and humeri of two untreated or endotoxin-treated guinea pigs was suspended in modifiedGaintner's BSA solution by repeated pipetting. The suspension was carefully layered over a discon-tinuous Percoll gradient consisting of the following densities: 1-030, 1-035, 1-040 and l-O5Og/cm3.The gradients were then centrifuged at 5 9 0 0 ^ for lOmin at 4°C. After centifugation, the upperthree density layers were withdrawn, diluted with modified Gaintner's medium (pH6-4) andcentrifuged at 5401^^ for lOminat 4°C. From the cell pellet obtained the supernatant was carefullywithdrawn. The pellet was washed once by resuspension with modified Gaintner's solution (pH 6-4)and centrifugation to remove Percoll. The final pellet was suspended in 5 ml modified Gaintner'smedium (pH 6-4) and fractionated further by elutriator centrifugation in a Beckman rotor JA6 at470 g W with continuous flow of modified Gaintner's medium (pH 6-4) at 25 ml/min. All cells witha sedimentation velocity of more than 10 mm h~'g"1 were washed out by the medium. The rest ofthe cells, containing about 40 % megakaryocytes, were processed further for DNA determination.
DNA determination in megakaryocytes
The megakaryocyte-enriched fraction obtained by Percoll gradient and elutriator centrifugationwas used for DNA determination by either of the two following methods. (1) Smears of the cell pelletwere stained by the Feulgen method. The DNA content of individual cells was determined with aLeitz Cytofluorometer at 550 nm using the filter 657/11. A total number of 115 cells was measuredfrom each group of the differentially treated animals. Comparison was made with the DNA values ofguinea pigs neutrophils, taken as 2C standard. Ploidy classes were delineated by the midpoint bet-ween two adjacent ploidy peaks. (2) The cells were fixed in ice-cold 80 % ethanol for at least 2 h at— 30°Cand then treated with RNaseat 37 CC for2h. Cells were stained with ethidium bromide (10Hg/ml) for at least 2h at 4°C. DNA content of the cells was measured by impulse cytophotometry(IPC 11, Phywe) with a filter combination PG38/BG3 (excitation) and GG550 (emission).
The values obtained by both methods were compared and the ploidy distribution was found to besimilar. Results from both methods were used for the final calculation of ploidy values.
Histochemical demonstration of acid phosphatase in platelets and megakaryocytes:classification of megakaryocytic cells
Bone marrow smears obtained from the two control groups and from guinea pigs 18,24,48 and 72 hafter administration of endotoxin were fixed on glass slides in 90% methanol at 4°C for 20-30 s,rinsed in distilled water, and air-dried. The dried slides were incubated for 2 h at 37 °C with a freshlyprepared staining solution for acid phosphatase, rinsed in water, and air-dried. Platelet acidphosphatase activity was demonstrated in smears obtained from a suspension of guinea pig plateletsin 0 9 % NaCl. Air-dried smears were either fixed in 90% methanol or used unfixed for acidphosphatase staining reaction. Counter staining was carried out by exposure to acid haematoxylin.
Slides were observed under the microscope at a magnification of X400. The enzyme activity wasestimated on a scale of 0-4+ by the size, the density, and the intensity of the colour of the granulesof the reaction product. 1 + : few, small, fine granules are located on one part of the cytoplasm. 2+ :few, small granules are scattered around the cytoplasm. 3+ : many, large, coarse, dark granules arescattered around the cytoplasm. 4+ : description of granules is same as 3 +, but the granules increasedin number with the tendency to clumping in places.
From each animal 100 megakaryocytic cells were observed under the microscope for classificationaccording to the following criteria:
Stage 1 (megakaryoblast). These cells are the earliest recognizable megakaryocytic cells with adiameter about 12-15 fttn. The nucleus/cytoplasm ratio is usually 1: 1 or slightly more. The nucleusis round or oval with a thin rim of cytoplasm around it. Acid phosphatase activity is sometimes presentand when observed in 90% methanol-fixed smears, it is usually rated as 1+ on the scale describedabove.
Stage 2 (promegakaryocyte). The cells belonging to this category are larger than stage 1 with adiameter between 20 and 35 fim. The nucleus/cytoplasm ratio is between 1: 1 and 1: 1-5. Nucleiusually contain more than one lobe. Acid phosphatase reaction products are almost always seen withan activity rating of 2+ on 90% methanol-fixed smears.
72 5. Raha and W. Wesemann
Stage 3A (early mature megakaryocyte). Cells in this stage are somewhat larger than stage 2 witha diameter between 40 and 70fun. The nucleus/cytoplasm ratio is about 1:2. The position ofnucleus is often eccentric. Intense acid phosphatase activity is observed in the cytoplasm with arating of 3+ or 4+ on 90% methanol-fixed smears.
Stage 3B (mature megakaryocyte). These cells are similar in size to the cells in stage 3A. Theyusually contain an eccentrically located pyknotic nucleus and large amounts of cytoplasm. Thenucleus/cytoplasm ratio is usually less than 1:2. On 90 % methanol-stained smears these cellsexhibit almost no acid phosphatase activity. As a result, these cells are easily distinguishable fromother cells by their large amount of acid-phosphatase-negative, lightly stained cytoplasm. Some cellsthat have already undergone platelet release and have been described in the literature as 'nakednuclei' are also seen and classified together with the stage 3B megakaryocytes.
Identification of acid phosphatase in human and guinea pig platelets by discelectrophoresis
A platelet button obtained from human platelets was suspended in 1-25 ml 0-9 % NaCI solutionand incubated with 1-25 ml of 10% Triton X-100 solution at 4°C for 30min. After 30mincentrifugation at 100000gmtx, 10/J of the platelet extract were mixed with 140/il stacking gel(2-5% polyacrylamide) for the preparation of the sample gel. The protein concentration perelectrophoresis tube was between 60 and 80 /ig. Electrophoresis was performed for 2h at 4°C with6-8mA/tube in a cathodic disc polyacrylamide gel system with 7-5% polyacrylamide separationgel. A solution of 0-35 M-/S-alanine in 0'14M-acetic acid buffer (pH4-5) was used as theelectrophoresis buffer. After electrophoresis the gels were kept at 4 °C for 45 min in 90 % methanolor in 0-1 M-acetate buffer (pHS-0). The gels were rinsed in distilled water and subjected for 3 h tocytochemical reaction for acid phosphatase as described above. After that the gels were rinsed indistilled water and stored in 7 % acetic acid.
For the identification of acid phosphatase in guinea pig platelets the platelet button wassolubilized in Triton X-100 and separated by gel electrophoresis as described before, except for thesample application procedure. Instead of using a sample gel, the sample was prepared by adding75 jul platelet extract to 25 )A of 40% sucrose solution. The sample in 10% sucrose was layereddirectly over the stacking gel. After electrophoresis, acid phosphatase staining was carried out withgels either treated with 90% methanol or untreated.
[3H]leucine uptake and incorporation into protein by guinea pig megakaryocytes
Megakaryocytes were isolated from normal (untreated) and endotoxin-treated guinea pigs(30/ig/kg, 24h prior to killing) by Percoll density-gradient centrifugation, as described earlier inthis section. The cell pellet containing 15—20% megakaryocytes was suspended in 14 ml modifiedGaintner's medium (pH7-4). One ml of this suspension was used for megakaryocyte count byphase-contrast microscopy and for protein determination. A sample (6'5 ml) of the cell suspensionwas incubated at 37°C for 5 min prior to the addition of 2-5/iCi of [4H]leucine to provide a finalconcentration of 7-5 nin. After incubation times of 10, 15, 22'5 and 30 min, 200-^1 samples werepipetted into 1 ml ice-cold 1-56% formalin and immediately mixed thoroughly. Cell pellets wereobtained by centrifugation at 4°C for 4 min at 12 000 g. Parallel experiments at 4°C were perfor-med to eliminate the effect of non-specific uptake. The cell pellets were washed twice by theaddition of 1 ml modified Gaintner's medium (pH6-4) followed by centrifugation at 12 000 £ for4 min.
For the measurement of [3H] leucine uptake by megakaryocytes the cell pellet was lysed overnightin800/ilO-2M-NaOHat40°C. After neutralization with 160/il 1 M-HC1, the cell extract was mixedvigorously with 10 ml of Ready-Solv™ HP scintillation cocktail for measurement of radioactivity.
For the assay of [3H]leucine incorporation into protein, the washed cell pellet was treated with1 M-KOH at room temperature for 30 min in order to hydrolyse RNA. After neutralization with1 M-HCI , 25/il of BSA solution (1 mg/ml) was added as carrier albumin. Finally, 30 % trichloracet-ic acid (final concn 10 %) was added to precipitate the protein. The samples were filtered on a fibre-glass filter (Whatman GF/B) in a Titertek cell harvester (Skatron AS, Norway). The proteinprecipitate was dissolved by shaking of the filters in 5 ml of Ready-Solv™ HP for 1 h.
Endotoxin-stimulated thrombopoiesis 73
Electron microscopyFor ultrastructural examination, pellets containing megakaryocytic cells or platelets were fixed
in ice-cold Ito's fixative (Ito & Karnovsky, 1968). The pellets were post-fixed in equal parts of 2%osmium tetroxide and 3 % potassium ferricyanide, dehydrated and embedded in Epon. Ultrathinsections were stained with uranyl acetate and lead citrate and examined in a Zeiss EM9.
RESULTS
Effect of endotoxin on platelet count
Platelet counts decreased gradually during the 24 h following administration ofendotoxin (Fig. 2). From a normal level of 1-051 ± 0-206(s.D.)(Xl06)//il of blood,platelet counts declined at 24 h after endotoxin treatment to the lowest value of0-640 ± O-144(xlO6)//il (P< 0-001). This corresponds to a decrease of about 40%.After that time, platelet counts increased and climbed to 0-947 ± 0-157(Xl06)/^lcorresponding to about 97 % by 48 h when they were no longer different from thepretreatment levels. Platelet counts remained unchanged in the animals that received0-9 % NaCl solution. At 24 h after treatment, platelet counts from guinea pigs treatedwith 0-9 % NaCl solution were not distinguishable from the pretreatment levels butwere significantly higher than those from endotoxin-treated guinea pigs (P<0-01).
Distribution of guinea pig platelets on the Percoll density-gradient before and aftertreatment with endotoxin
After density-gradient centrifugation of blood platelets obtained from controlanimals the percentage distribution of the three density subtractions on the Percoll
12
Fig. 2. Effect of endotoxin administration on platelet levels in guinea pigs. Platelet countsare expressed per [A of blood. Physiological NaCl solution ( • • ) and 30 fig endo-toxin/kg of body weight ( • • ) were administered i.v. The numbers of experi-ments are given in parentheses; the bars indicate the S.D.
CEI. 60
74 S. Raha and W. Wesemann
Table 1. Effect of endotoxin administration on the density distribution of guinea pigplatelets in a Percoll gradient
Time after endotoxintreatment (h)
0 (control)
48 (group V)
72 (group VI)
Subtraction 1(ps£ 1-055)
8-9 ±2-6 (6)
**(a)4-1 ±0-7 (4)
•(b)7-5 ±2-0 (3)
Platelet distribution (%)A
Subtraction 2(p 1-055-1-065)
72-3 ±1-6 (5)
••(a)48-3 ± 6-6 (4)
"(a)43-2 ±5-3 (8)
•\Subtraction 3
(p 1-065-1-075)
19-1 ±2-9 (6)
••(a)47-9 ±5-6 (4)
"(a)49-3 ±4-6 (3)
Guinea pigs were treated with 30^g endotoxin/kg body weight prior to heart puncture. PRP wasfractionated by Percoll density centrifugation as described in the text and shown in Fig. 1. Data arepresented as the mean percentage ± S.D. The number of experiments is given in parenthesis. Totalplatelet count: 5X 10* platelets/1 ml PRP-buffer mixture. ••P<0-01, *P<0-05, significantlydifferent from the corresponding value in control (a) and in group V (b), respectively, p, densityin g/cm3.
gradient (Fig. 1) are 8-9 ±2-6 (subtraction 1), 72-3 ±1-6 (subtraction 2), and19-1 ±2-9 (subfraction 3), respectively (Table 1). At 48h after endotoxin a sig-nificant change was observed in all of the three subfractions, demonstrated by asignificant decrease (i3 < 0 • 01) in subfractions 1 and 2 and a significant increase (P< 0-001) in subfraction 3. Some of these changes persisted at 72h, when subfraction3 was still significantly enhanced. However, at this time subfraction 1 did not differfrom control values.
Electron micrographs showed that subfraction 1 (Fig. 3) contained platelets smallerin size with fewer granules when compared with that of subfraction 3 (Fig. 4).Platelets from subfraction 2 showed characteristics intermediate between subfractions1 and 3. Compared with the respective subfractions of the controls, no morphologicalchanges are observed in the three platelet subfractions 48 and 72 h after endotoxintreatment.
[3S S] sulphate incorporation into platelets
The [35S]sulphate uptake was assayed 24 h after administration of Na235SO4.Compared with the controls, a significant increase in [3SS]sulphate incorporation intothe platelets was found if guinea pigs were treated with endotoxin either 18 or 24 hprior to [ Sjsulphate injection (Fig. 5).
Figs 3, 4. Guinea pig platelets isolated from platelet-rich plasma by Percoll density-gradient centrifugation. For details see legend to Fig. 1. Double fixation with Ito's fixativeand osmium tetroxide, lead citrate staining. X6600.
Fig. 3. Platelets with density^ 1-055g/cm3 (subfraction 1).Fig. 4. Platelets with density between 1-065 and 1-075 g/cm3
(subfraction 3).
Endotoxin-stimulated thrombopoiesis 75
Figs 3 and 4
76 S. Raha and W. Wesemann
5000-:
a 4000-
Eci
0)TO
03
SL 1000
3000-
2000-
24Time (h)
48
Fig. 5. Platelet production measured by [3SS] sulphate incorporation into newly formedplatelets. Guinea pigs were untreated (control; • • ) or received 30fig endotoxin/kg body weight (i.v.) at 18h ( • • ) and 24h (A • ) prior to Na2
35SO4 injec-tion (i.p.) at Oh. The means ± S.D. of three experiments are given. # P < 0 - 0 5 , **P<0 0 1 , significantly different from the corresponding data in the control group.
Separation of proteins with acid phosphatase activity in human and guinea pig bloodplatelets
After 2 h of electrophoresis of soluble human platelet proteins, two adjacent proteinbands with acid phosphatase activity were observed. Both of these bands were sen-sitive to methanol, as was demonstrated by the marked reduction of activity aftertreatment with 90% methanol. If soluble proteins from guinea pig platelets wereseparated under the same electrophoresis conditions, only one band of acid
Figs 6—9. Light micrographs (differential interference contrast) of guinea pigmegakaryocytes and platelets. Staining of bone marrow and platelet smears for acidphosphatase with a-naphthol-AS-BI-phosphate/Fast Garnet GBC salt, counter stainingwith acid haematoxylin.
Fig. 7. Methanol-fixed mature megakaryocyte (stage 3B) with only a faint acidphosphatase (methanol-sensitive) reaction. X360.
Fig. 8. Strong reaction of acid phosphatase in unfixed guinea pig platelets. X420.Fig. 9. Marked reduction of acid phosphatase reaction in guinea pig platelets pretreated
with methanol. X420.
Endotoxin-stimulated thrombopoiesis 77
Figs 6-9
78 5. Raha and W. Wesemann
phosphatase activity could be seen. Apparently the acid phosphatase activity of theband was methanol-sensitive since a pronounced reduction of staining intensity wasobtained after treatment with 90% methanol.
Histochemical demonstration of acid phosphatase in megakaryocytes and platelets ofthe guinea pig .
Light micrographs of two mature megakaryocytes from bone marrow smearstreated with 90% methanol are shown in Figs 6 and 7. Although the twomegakaryocytes were similar to each other, the megakaryocytes in Fig. 6 demonstratea positive acid phosphatase reaction in contrast to the megakaryocyte seen in Fig. 7that shows almost no acid phosphatase stain. In unfixed platelet smears strong acidphosphatase activity was observed (Fig. 8). The acid phosphatase staining reactionwas markedly reduced if platelets were treated with 90 % methanol prior to thestaining reaction (Fig. 9).
Effect of endotoxin on maturation and ploidy of megakaryocytes
Bone marrow smears were used to classify the megakaryocytic cells according tomaturation and ploidy. Figs 10 and 11 illustrate the effect of endotoxin administrationon the distribution of megakaryocytes in different stages of maturation, classifiedaccording to the criteria described in Materials and Methods. The percentagedistribution of the megakaryocytic cells between the different stages was similar in theuntreated (group I) and NaCl-treated (group II) controls. The percentage of maturemegakaryocytes (stage 3A + 3B) in the two control groups (I and II) is about 66, whilesignificantly higher percentages - 81 and 88, respectively - were found 24 h (groupIV) and 48 h (group V) after administration of endotoxin. A slight but not statisticallysignificant increase in the percentage of stage 3A + 3B megakaryocytes had beenobserved already at 18h after administration of endotoxin (group III). The firstsignificant difference was noted at 24 h after administration of endotoxin and it waseven greater at 48 h, declining towards control values at 72 h. The percentage ofyounger megakaryocytes showed corresponding alterations in the reverse direction at24 and 48 h after endotoxin administration. When the mature megakaryocytes were
Fig. 10. Percentage distribution of guinea pig megakaryocytes in different stages ofmaturation as observed in bone marrow smears. The criteria described in the text wereused for classification of the megakaryocytes into megakaryoblasts (stage 1,^B),promegakaryocytes (stage 2, JJj ), early mature megakaryocytes (stage 3A,IM), andmature megakaryocytes (stage 3B, ̂ u ) . The total number of megakaryocytic cells countedwere taken as 100%. Megakaryocytes from at least three animals (100 megakaryocytesfrom each animal) were counted. The bars indicate the s.D. * P < 0 ' 0 5 , • * P < 0 - 0 1 ,***P<0-001, significantly different from the corresponding data in control (II). I,untreated control; II, saline control; III, 18h after 30/ig endotoxin/kg (i.v.); IV, 24hafter endotoxin; V, 48h after endotoxin; VI, 72h after endotoxin.
Fig. 11. Classification of guinea pig megakaryocytes into two developmental stages.Immature megakaryoctyes (stages 1 + 2g3 ) ; early mature and mature megakaryocytea(stages 3 A + 3 B tmX). Classification and experimental conditions as described in the legendto Fig. 10.
Endotoxin-stimulated thrombopoiesis 79
100 H
80 H
uu
cyti
o
CO
60
40
20 H
100 i
80
60 ^
oCD
atv
40
20
III IVTreatment
Fig. 10
V VI
III IV V
Treatment
Fig. 11
80 5. Raha and W. Wesemann
compared separately, according to the subdivision into 3A and 3B, a difference wasobserved between 24 and 48 h after endotoxin. At 24 h after endotoxin, the increasein the percentage of mature megakaryocytes could be attributed to a significant in-crease in stage 3A megakaryocytes while the stage 3B megakaryocytes were onlyslightly increased in number. At 48 h after endotoxin administration, this pattern wasreversed, as stage 3A remained the same and stage 3B was significantly increased.
Fig. 12 demonstrates a linear relationship between the amount of labelled sulphateincorporated into the platelets and the percentage of stage 3A megakaryocytes. Anincrease in the number of stage 3A megakaryocytic cells at the time of [35S] sulphateadministration, produced a corresponding increase in the incorporation of sulphateinto newly formed platelets 24 h later.
Ploidy distribution of endotoxin-treated guinea pigs and controls are given in Fig.13 and Table 2. No significant differences were found between the ploidy distributionpattern of control animals and those of guinea pigs 18, 24, 48 or 72 h after endotoxintreatment. The distribution pattern corresponds well with previously published data(Hagen-Aukamp et al. 1980; Penington & Oleson, 1970).
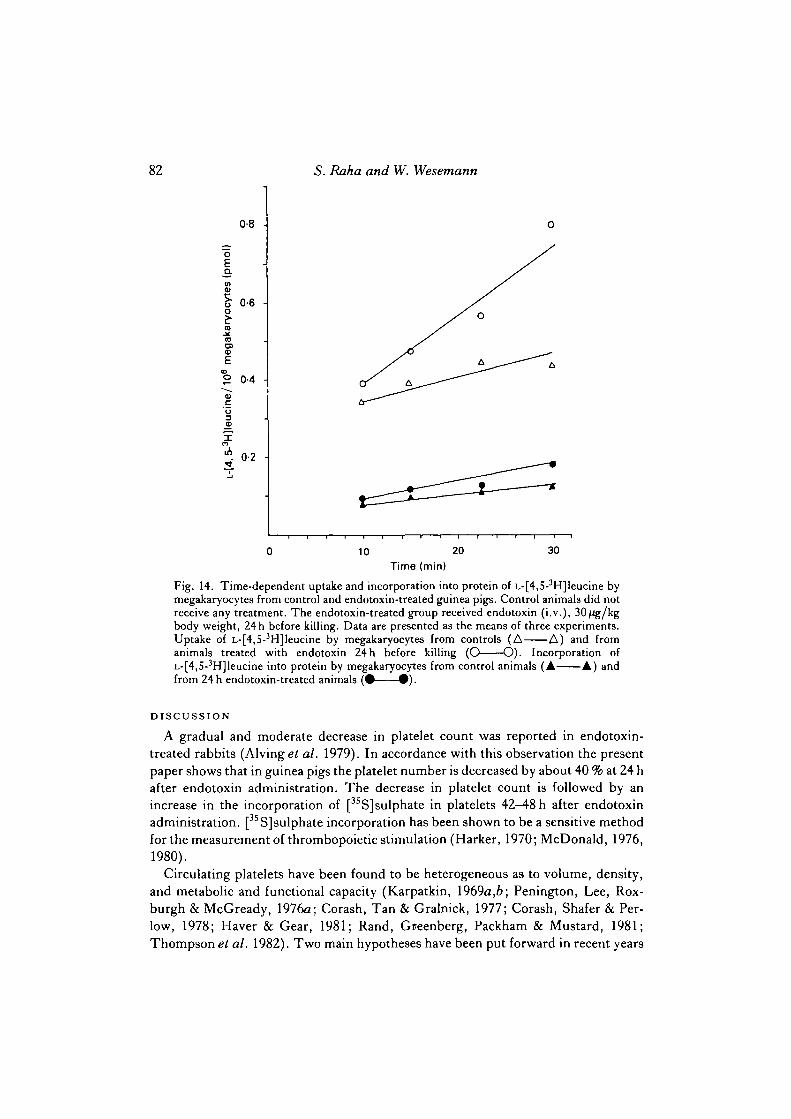
Leucine uptake and incorporation into protein by guinea pig megakaryocytes
As shown in Fig. 14, 0-430 ± 0-023 pmol of [3H]leucine/106 megakaryocytes weretaken up by megakaryocytes within 30 min. About 28 % of the labelled leucine corres-ponding to 0-122 ±0-016 pmol/106 megakaryocytes are incorporated within this timeinto trichloroacetic acid-precipitable protein. When expressed in term9 of proteincontent, the uptake of labelled leucine/mg megakaryocyte protein was 0-210 ± 0-016pmol and the incorporation into protein was 0-066 ±0-018 pmol after 30 min ofincubation. When the same experiments were performed with megakaryocytesisolated from bone marrow of guinea pigs treated 24 h earlier with endotoxin, the
2 4000 -|
a^ 3000
Ed
~ 2000
a
a 1000 -a
40 60Stage 3A megakaryocytes (%)
80
Fig. 12. Relationship between the incorporation of [35S]sulphate into guinea pig plateletsand the percentage of stage 3A megakaryocytes at the time of [35S]sulphate injection. Forexperimental details see legends to Figs 5 and 10.
Endotoxin-stimulated thrombopoiesis 81
50% 15%
32
Fig. 13. Ploidy distribution of megakaryocytes isolated from bone marrow of untreatedguinea pigs by density-gradient plus elutriator centrifugation. The percentage ploidydistribution is given at the top of the figure. The DNA content of 115 cells was determinedwith a Leitz cytofluorometer at 550 nm, filter 657/11, and compared with results obtainedby impulse cytophotometry. Guinea pig neutrophils were used as 2C standard.
Table 2. Ploidy distribution in megakaryocytes of normal and endotoxin-treatedguinea pigs
Guinea pigs were treated with 30 /Jg endotoxin/kg body weight (i. v.). Cells stained with ethidiumbromide were measured by impulse cytophotometry. The distribution was verified bycytofluorometry of Feulgen-stained cells. Guinea pig neutrophils were taken as 2C standard.
uptake and incorporation of labelled leucine into protein were significantly (P < 0-05)increased to 0-797 ± 0-234 and 0-177 ± 0-024 pmol/106 megakaryocytes, respective-ly. The same values when expressed in terms of protein content (0-373 ±0-113 and0-083 ± 0-012 pmol/mg protein, respectively) were not significantly different fromthose obtained with megakaryocytes from untreated guinea pigs. This finding in-dicates that the morphologically observed increase in the percentage of maturemegakaryocytes is paralleled by an increase in protein synthesis.
82 5. Raha and W. Wesemann
0-8 •
oQ.
0-6 -o
ID
E° 0-4 -
10 20 30
Time (min)
Fig. 14. Time-dependent uptake and incorporation into protein of L-[4,5-3H]leucine bymegakaryocytes from control and endotoxin-treated guinea pigs. Control animals did notreceive any treatment. The endotoxin-treated group received endotoxin (i.v.), 30/Jg/kgbody weight, 24 h before killing. Data are presented as the means of three experiments.Uptake of L-[4,5-3H]leucine by megakaryocytes from controls (A A) and fromanimals treated with endotoxin 24 h before killing (O O). Incorporation ofL-[4,S-3H]leucine into protein by megakaryocytes from control animals (A A) andfrom 24 h endotoxin-treated animals ( • • ) .
DISCUSSION
A gradual and moderate decrease in platelet count was reported in endotoxin-treated rabbits (Alvinget al. 1979). In accordance with this observation the presentpaper shows that in guinea pigs the platelet number is decreased by about 40 % at 24 hafter endotoxin administration. The decrease in platelet count is followed by anincrease in the incorporation of [35S] sulphate in platelets 42—48 h after endotoxinadministration. [35S]sulphate incorporation has been shown to be a sensitive methodfor the measurement of thrombopoietic stimulation (Harker, 1970; McDonald, 1976,1980).
Circulating platelets have been found to be heterogeneous as to volume, density,and metabolic and functional capacity (Karpatkin, 1969a,b\ Penington, Lee, Rox-burgh & McGready, 1976a; Corash, Tan & Gralnick, 1977; Corash, Shafer & Per-low, 1978; Haver & Gear, 1981; Rand, Greenberg, Packham & Mustard, 1981;Thompson et al. 1982). Two main hypotheses have been put forward in recent years
Endotoxin-stimulated thrombopoiesis 83
to explain this. Some investigators suggested that platelet heterogeneity could beexplained by the production of platelets from megakaryocytes of different ploidyclasses and that it was independent of platelet age (Peningtonef a/. 1976a; Penington,Streatfield & Roxburgh, 19766). In contrast to this interpretation it was proposed thatfrom the total platelet population the larger, heavier and metabolically more activeplatelets could be identified as young platelets, and that these platelets becameprogressively smaller and lighter as they aged in the circulation (Karpatkin, 1969a,6;Charmatz & Karpatkin, 1974; Corashei al. 1978; Karpatkin, 1978; Blajchmane* al.1981; Rande* al. 1981; Corash & Shafer, 1982).
In the present study, concomitantly with the increase of [3sS]sulphate incorpora-tion into platelets, the number of heavy platelets in the circulation was also found tobe elevated. Since after endotoxin treatment no change in megakaryocyte ploidy wasfound, it is unlikely that the presence of a higher number of heavier platelets in ourstudies is related to alterations in megakaryocyte ploidy. In recent years, the presenceof a higher number of larger platelets in the circulation has been used as an indicatorof increased platelet production (Weiner & Karpatkin, 1972; Weintraub & Karpatkin,1974; McDonald, 1980). The two methods - [35S]sulphate incorporation anddensity-gradient fractionation into light and heavy platelet populations — are used inthe present studies, together with that of the conventional one of platelet counting,to provide an accurate measure of the thrombopoietic stimulus. The data obtainedshow that after treatment of guinea pigs with endotoxin, a stimulus for plateletproduction is detected by all of the three methods mentioned above.
Compared with the small—light platelets, the heavy—large platelets aggregate morereadily and completely when treated with ADP, thrombin or epinephrine (Karpatkin,19696; Thompson et al. 1982). In view of this finding, the presence of a significantlyhigher number of heavier platelets cannot be accounted for by the possibility that thelighter platelets are preferentially degranulated and removed by endotoxin treatment.
Alving et al. (1979) and Ramsey et al. (1980) measured the increased plateletproduction 42 h after treatment with endotoxin or lipid A of rabbits by[75Se]methionine incorporation. The increase in platelet production was accom-panied by moderate thrombocytopenia only in the case of endotoxin treatment but notif lipid A was administered to the rabbits. From these findings, the authors concludedthat the thrombopoietic stimulus might have been independent of thrombocytopenia.However, the time sequence of the application of the thrombopoietic stimulus and theadministration of the radioactive label appears to be critical. In both of the studiesmentioned above, the radioactive platelet label was administered 18 h after the ad-ministration of endotoxin and lipid A, resulting in an increased incorporation of thelabel into the platelets 24 h later. In our studies also an increase in platelet[35S]sulphate incorporation was observed when the platelet label was administered18 h after endotoxin treatment, although there was no significant decrease in circula-ting platelet level at that time. Furthermore, the elevation of [35S]sulphate incorpora-tion was of short duration, as demonstrated by the return to normal level 66—72 h afterendotoxin treatment. This is in contrast to the effect of severe and sustained throm-bocytopenia, in which case the [3sS]sulphate incorporation was elevated for a longer
84 5. Raha and W. Wesemann
period of time (Harker, 1970). The time sequence and the duration of incorporationof radioactive label into platelets are similar to the results obtained after stimulationof thrombocytopoiesis by the thrombopoiesis-stimulating factor, which is unac-companied by thrombocytopenia (McDonald, 1980). The presence of a significantlyhigher number of circulating heavier platelets at the time when [35S]sulphate incor-poration is also significantly enhanced provides additional evidence that the measure-ment of platelet size or density may reflect a measure of thrombopoietic stimulus.
Markovic & Shulman (1977) first described a histochemical method to distinguishbetween different developmental stages of megakaryocytes in human bone marrowsmears. Megakaryocyte acid phosphatase shows different sensitivities to methanoltreatment depending on the maturational stage of the megakaryocytes. The acidphosphatase of some mature megakaryocytes in bone marrow smears possesses thesame methanol sensitivity as blood platelets. This observation suggests that thesemegakaryocytes might be at the stage of final maturation and platelet release, and thatthe presence of this methanol-sensitive isoenzyme of acid phosphatase might providean index of platelet formation by megakaryocytes. The results presented in this paperconfirm the finding in human platelets and megakaryocytes, by demonstrating thepresence of an acid phosphatase isoenzyme sensitive to methanol treatment in guineapig platelet and megakaryocytes. As compared with early mature megakaryocytes(stage 3A), the acid phosphatase activity is drastically decreased after pretreatmentwith methanol in mature megakaryocytes (stage 3B) when platelet release occurs. Theacid phosphatase staining is combined with standard morphological criteria toelucidate the effect of endotoxin treatment on megakaryocytic development. At 24 hafter endotoxin treatment, a significant increase in the number of early maturemegakaryocytes was noted. These early mature megakaryocytes contain an acidphosphatase insensitive to methanol treatment, while the number of platelet-formingmegakaryocytes has not significantly changed. However, 24h later, the number ofmature megakaryocytes with methanol-sensitive acid phosphatase (presumably theplatelet-forming megakaryocytes) is significantly increased. This observation can beinterpreted as an enhanced maturation rate in megakaryocytes (Ebbe et al. 1968a).The increased [35 S] sulphate incorporation into platelets and the occurrence of a largernumber of heavier platelets in circulation coincide with the presence of a largernumber of platelet-forming megakaryocytes in the bone marrow 48 h after endotoxintreatment. In contrast to the effects observed after acute thrombocytopenia inducedby anti-platelet serum (Penington & Oleson, 1970; Odell et al. 1976; Levin, Levin,Penington & Metcalf, 1981), no alterations in the ploidy of megakaryocytes towardshigher ploidy levels are observed if thrombocytopenia is obtained by endotoxin ad-ministration. The different nature of the thrombopoietic stimulus might be proposedas a tentative explanation for this difference.
Though mammalian platelets do not contain nuclei, a low synthesis of protein invitro has been described (Agam et al. 1976; Bessler, Agam & Djaldetti, 1976; Plow,1979). Compared with human platelets (Warshaw, Laster & Shulman, 1966; Nowak,1981), rather high uptake of radioactively labelled leucine into normal guinea pigmegakaryocytes and incorporation into protein are obtained in vitro. Both parameters
Endotoxin-stimulated thrombopoiesis 85
(uptake and incorporation into protein) are significantly increased if themegakaryocyte fraction contains an increased number of early maturemegakaryocytes, which is obtained 24h after endotoxin treatment. The elevation ofamino acid uptake and protein synthesis rate seems to be due to the larger cytoplasmicmass in these cells, as indicated by the observation that the increase in theseparameters is only observed in respect of the cell number not the protein content. Thehigher metabolic capacity of these cells is also demonstrated by the evidence that theadministration of radioactively labelled sulphate 24 h after endotoxin treatmentresults in a 2'3-fold increase in platelet incorporation of radioactive [35S]sulphateanother 24 h later.
The data presented here indicate that endotoxin treatment results in a stimulationof platelet production as evidenced by the presence of an increased number of heavyplatelets, increased [3SS]sulphate incorporation into platelets, and an enhancedmaturation rate of megakaryocytes. Although these observations describe some ef-fects of endotoxin-stimulated thrombopoiesis, the exact mechanism still remains tobe elucidated.
The authors would like to express their gratitude to Professor Dr G. Aumiiller and Professor DrP. Koch, Marburg, and Professor Dr G. Cleffmann, GiefJen, for stimulating discussions and helpwith the histological examination, microscopy and cytofluorometry, respectively, of megakaryocytesand platelets. They also wish to thank Mrs U. Lehr, Mr N. Fey and Mr H. Gerlach for expert technicalassistance. The help of Dr R. Merz in performing the impulse cytophotometric DNA measure-ments is gratefully acknowledged. This work has been supported by grants from the DeutscheForschungsgemeinschaft, Sonderforschungsbereich 103 'Zellenergetik und Zelldifferenzierung'.
REFERENCESAGAM, G., GASNER, S., BESSLER, H., FISHMAN, P. & DJALDETTI, M. (1976). Chloramphenicol
induced inhibition of platelet protein synthesis: in vitro and in vivo studies. Br.J. Haemal. 33,53-59.
ALVING, B. M., EVATT, B. L., LEVIN, J., BELL, W. R., RAMSEY, R. B. & LEVIN, F. C. (1979).
Platelet and fibrinogen production: relative sensitivities to endotoxin. J . Lab. din. Med. 93,437-448.
AUSPRUNK, D. & DAS, J. (1978). Endotoxin-induced changes in human platelet membranes:morphologic evidence. Blood 51, 487—495.
BESSLER, H., AGAM, G. & DJALDETTI, M. (1976). Increased protein synthesis by human plateletsduring phagocytosis of latex particles in vitm. Thrombos. Haemostas. 35, 350-357.
BLAJCHMAN, M. A., SENYL, A. F., HIRSH, J., GENTON, E. & GEORGE, J. N. (1981). Hemostatic
function, survival and membrane glycoprotein changes in young versus old rabbit platelets. J.din. Invest. 68, 1289-1294.
CHARMATZ, A. & KARPATKIN, S. (1974). Heterogeneity of rabbit platelets. I. Employment of analbumin density gradient for separation of a young platelet population identified with Se75-selenomethionine. Thrombos. Diathes. Haemorrh. 31, 485—492.
CORASH, L. & SHAFER, B. (1982). Use of asplenic rabbits to demonstrate that platelet age anddensity are related. Blood 60, 166-171.
CORASH, L., SHAFER, B. & PERLOW, M. (1978). Heterogeneity of human whole blood plateletsubpopulations. II. Use of a subhuman primate model to analyze the relationship between densityand platelet age. Blood 52, 726-734.
CORASH, L., TAN, H. & GRALNICK, H. R. (1977). Heterogeneity of human whole blood plateletsubpopulations. I. Relationship between buoyant density, cell volume, and ultrastructure. Blood49, 71-87.
86 5. Raha and W. Wesemann
DAS, J., SCHWARZ, A. A. & FOLKMAN, J. (1973). Clearance of endotoxin by platelets: role inincreasing the accuracy of the Limulus gelatin test and combating experimental endotoxemia.Surgery 74, 235-240.
DES PEREZ, R. N., HOROWITZ, H. I. &HOOK, E. W. (1961). Effect of bacterial endotoxin on rabbitplatelets. I. Platelet aggregation and release of platelet factors in vitro. J. exp.Med. 114,857-874.
EBBE, S. (1976). Biology of megakaryocytes. \n Progress in Hemostasis and Thrombosis, vol. 3 (ed.Th. Spaet), pp. 211-229. New York: Grune and Stratton Verlag.
EBBE, S., STOHLMAN, F. JR, DONOVAN, J. & OVERCASH, J. (1968a). Megakaryocyte maturationrate in thrombocytopenic rats. Blood 32, 787-795.
EBBE, S., STOHLMAN, F. JR, OVERCASH, J., DONOVAN, J. & HOWARD, D. (1968A).
Megakaryocyte size in thrombocytopenic and normal rats. Blood 32, 383-392.GAINTNER, J. R., JACKSON, D. P. & MAYNERT, E. W. (1962). The action of thrombin on platelet
5-hydroxytryptamine. Bull. Johns Hopkins Hosp. I l l , 185-197.HAGEN-AUKAMP, C , WESEMANN, W. & AUMOLLER, G. (1980). Intracellular distribution of
adenine and 5-hydroxytryptamine in megakaryocytes isolated by density gradient and velocitysedimentation from bone marrow. Eur.J. Cell Biol. 23, 149—156.
HARKER, L. A. (1970). Regulation of thrombopoiesis. Atn. J. Physiol. 218, 1376-1380.HARKER, L. A. & FINCH, C. A. (1969). Thrombokinetics in man. J. din. Invest. 48, 963-974.HAVER, V. M. & GEAR, A. R. (1981). Functional fractionation of platelets. J. Lab. din. Med. 97,
187-204.HOROWITZ, H. I., DES PEREZ, R. M. &HOOK, E. W. (1962). Effect of bacterial endotoxin on rabbit
platelets. II. Enhancement of platelet factor 3 activity in vitro and in vivo.J. exp. Med. 116,619-633.
ITO, S. & KARNOVSKY, M. J. (1968). Formaldehyde-glutaraldehyde fixatives containing tri-nitrocompounds. J. Cell Biol. 39, 168a-169a.
KARPATKIN, S. (1969a). Heterogeneity of human platelets. I. Metabolic and kinetic evidencesuggestive of young and old platelets. J. din. Invest. 48, 1073—1082.
KARPATKIN, S. (19696). Heterogeneity of human platelets. II. Functional evidence suggestive ofyoung and old platelets. J. din. Invest. 48, 1083-1087.
KARPATKIN, S. (1978). Heterogeneity of rabbit platelets. VI. Further resolution of changes inplatelet density, volume and radioactivity following cohort labelling with 75Se-selenomethionine.Br.J. Haemat. 39, 459-469.
LEVIN, J., LEVIN, F. C , PENINGTON, D. G. & METCALF, D. (1981). Measurement of ploidydistribution in megakaryocyte colonies obtained from culture: with studies of the effects ofthrombocytopenia. Blood 57, 287-297.
LOWRY, O. H., ROSEBROUGH, N. J., FARR, A. L. & RANDALL, R. J. (1951). Protein measurementwith the Folin phenol reagent. J. biol. Chem. 193, 265-275.
MACINTYRE, D. E., ALLEN, A. P., THORNE, K. J. I., GLAUERT, A. M. & GORDON, J. L. (1977).Endotoxin-induced platelet aggregation and secretion. I. Morphological changes andpharmacological effects. J . Cell Sd. 28, 211-223.
MCDONALD, T. P. (1976). A comparison of platelet size, platelet count and platelet 35S incorpora-tion as assays of thrombopoietin. Br.J. Haemat. 34, 257-267.
MCDONALD, T. P. (1980). Effect of thrombopoietin on platelet size of mice. Expl Hemat. 8,527-532.
MARKOVIC, O. S. & SHULMAN, N. R. (1977). Megakaryocyte maturation indicated by methanolinhibition of an acid phosphatase shared by megakaryocytes and platelets. Blood 50, 905-914.
MOLLER-BERGHAUS, G. & LOHMANN, E. (1974). The role of complement in endotoxin-induceddisseminated intravascular coagulation: studies in congenitally C^-deficient rabbits. Br. J.Haemat. 28, 403-418.
NOWAK, P. O. (1981). Zur Proteinbiosynthese in isolierten Megakaryozyten und Thrombozyten.Diplomarbeit, Philipps-Universitat, Marburg.
ODELL, T. T. JR, JACKSON, C. W., FRIDAY, T. J. & CHARSHA, D. E. (1969). Effects of throm-bocytopenia on megakaryocytopoiesis. Br.J. Haemat. 17, 91-101.
ODELL, T. T. JR, MURPHY, J. R. & JACKSON, C. W. (1976). Stimulation of megakaryocytopoiesisby acute thrombocytopenia in rats. Blood 48, 765-775.
PENINGTON, D. G., LEE, N. L. Y., ROXBURGH, A. E. & MCGREADY, J. R. (1976a). Plateletdensity and size: the interpretation of heterogeneity. Br.J. Haemat. 34, 365-376.
Endotoxin-stimulated thrombopoiesis 87
PENINGTON, D. G. & OLESON, T. E. (1970). Megakaryocytes in states of altered platelet produc-tion: cell number, size and DNA content. Br.J. Haemat. 18, 447-463.
PENINGTON, D. G., STREATFIELD, K. & ROXBURGH, A. E. (19766). Megakaryocytes and theheterogeneity of circulating platelets. Br.J. Haemat. 34, 639-653.
PLOW, E. F. (1979). Extracellular factors influencing the in vitro protein synthesis of platelets.Thrombos. Haemostas. 42, 666-678.
RAMSEY, R. B., HAMNER, M. B., ALVING, B. M., FINLAYSON, J. S., ALVING, C. R. & EVATT, B.L. (1980). Effects of lipid A and liposomes containing lipid A on platelet and fibrinogen produc-tion in rabbits. Blood 56, 307-310.
RAND, M. L., GREENBERG, J. P., PACKHAM, M. A. & MUSTARD, J. F. (1981). Density subpopula-tions of rabbit platelets: size, protein, and sialic acid content, and specific radioactivity changesfollowing labelling with 3sS-sulfate in vivo. Blood 57, 741-746.
THOMPSON, C. B., EATON, K. A., PRINCIOTTA, S. M., RUSHIN, C. A. & VALERI, C. R. (1982).Size dependent platelet subpopulations: relationship of platelet volume to ultrastructure,enzymatic activity, and function. Br.J. Haemat. 50, 509—519.
WARSHAW, A. L., LASTER, L. & SHULMAN, R. (1967). Protein synthesis by human platelets. J.biol. Chem. 24, 2094-2097.
WEINER, M. & KARPATKIN, S. (1972). Use of the megathrombocyte to demonstrate throm-bopoietin. Thrombos. Diathes. Haemorrh. 28, 24—30.
WEINTRAUB, A. H. & KARPATKIN, S. (1974). Heterogeneity of rabbit platelets. II. Use of themegathrombocyte to demonstrate a thrombopoietic stimulus. J . Lab. din. Med. 83, 896-901.
WOLFF, S. M. & BENNETT, J. V. (1974). Gram-negative rod bacteremia. N. Engl.J. Med. 291,733-734.