Effects of ingested ivermectin on body mass J A N E G 0 V E R and L E S S T R 0 N G School of Biological Sciences, University of Bristol Abstract. Adults of Neomyia comicina' (F.) given continuous access to cow dung con- taining ivermectin at concentrations of 0.5.0.25 and 0.125 pg g-', the range found in the dung of injected cattle (Sommer et al., 1992), were monitored every 12 h for changes in mass and excretory rates over a period of 3-5 days. Compared with the control insects, adult females that had ingested ivermectin showed disrupted patterns of daily Inass change, an increase in dawn mass, and greatly reduced rates of excretion. Flies fed continuously with dung containing ivermectin at 0.25 pg g-' and 0.125 pg g-' in a regime identical to the above showed significantly increased abdominal mass compared with the controls. The observations are discussed in relation to other known effects of ivermectin on insect physiology. Introduction Ivermectin is a broad-spectrum antiparasitic drug derived from fermentation products of Srrepromyces avrrmifilis. It is used ex- tensively for the treatment of cattle to reduce nematode burdens and arthropod ectoparasites (Campbell, 1989). Approximately 90% of the drug administered to cattle by subcutaneous injec- tion is excreted in the faeces over a period of about 35 days fol- lowing treatment. The presence of ivermectin in the dung has deleterious effects on dung-breeding pests including the bush fly, Musca vetustissima Walker (Ridsdill-Smith, 1988), and the face fly, Musca autumnalis Degeer (Meyer et nl., 1980). How- ever, Wall & Strong (1987) have drawn attention to the potential damage caused by ivermectin residues which exert a toxic effect on many non-pest insects that utilize dung for breeding, and whose larval activity contributes greatly to dung breakdown and recycling. Most studies of the non-specific effects of ivermectin have focused on its impact on larvae: much less is known of the ef- fects on adults caused by ingesting dung containing ivermectin residues. Mahon et al. (1993) observed increased mortality and reduced reproductive potential in Lucilia cuprina that had fed on dung deposited by sheep after treatment with an ivermectin drench. The survival and reproduction of adults of the fly Neomyia cornicina (F.), which feed on the liquid portion of freshly depos- ited pats (Hammer. 1941), are also significantly affected by 'This insect has been referred to as Cgprolucilia caesarion (Ham- mer, 1941) and Orthelia cornicinn (E) (Wardhaugh & Rodriguez- Menendez, 1988) Correspondence: Dr J. Gover, School of Biological Sciences, Univer- sity of Bristol, Woodland Road, Bristol BS8 1 UG. ivermectin residues (Gover & Strong, 1995). The ingestion of ivermectin causes bloating of the abdomen in adults of two spe- cies of Glossina (Distelmans et d.. 1983; Langley & Roe, 1984), and in Lucilia cuprina (Cook. 199 1). This bloating may be caused by water retention and thus affect mass changes. The aim of this study was to investigate the impact of ivermectin on fluid reten- tion and mass changes in adult females of N.cornicina fed on cattle dung containing ivermectin at concentrations similar to those found after injecting cattle with the drug. Such a study would add significantly to the growing knowledge of the impact of ivermectin on insects of pasture land. Materials and Methods Experirnenlul insecfs. Adult flies were maintained in a labora- tory colony at 25"C, 75% r.h. and LD 175 h photoperiodic re- gime. They were housed in cages (30 x 30 x 30 cm) and fed ad libitum on a mixture of egg powder, icing sugar, and skimmed milk powder in equal parts by weight: fresh liver was offered once weekly. Non-contaminated cattle dung (see below) was provided weekly for feeding and oviposition. 24 h after egg hatch. larvae were transferred to 200 g aliquots of fresh dung contained in a 15 cm diameter plastic dishes in which larval development was completed. Under the above conditions, the third-stage lar- vae wandered after about 6 days and were allowed to pupariate in clean sawdust. Adults emerged about 7 days later. Dung snn7pple.s. Non-contammatcd cattlc dung was collcctcd from the dairy herd at thc Univcrsity of- Bristol farm at Langford (nr Bristol, Avon. U.K.), mixed thoroughly and stored frozen at -20°C until required. The dung was known to be non-contam- inated, because the cattle received no chemical treatments dur-
Transcript
Effects of ingested ivermectin on body mass
J A N E G 0 V E R and L E S S T R 0 N G School of Biological Sciences, University of Bristol
Abstract. Adults of Neomyia comicina' (F . ) given continuous access to cow dung con- taining ivermectin a t concentrations of 0.5.0.25 and 0.125 pg g-', the range found in the dung of injected cattle (Sommer et al., 1992), were monitored every 12 h for changes in mass and excretory rates over a period of 3-5 days. Compared with the control insects, adult females that had ingested ivermectin showed disrupted patterns of daily Inass change, an increase in dawn mass, and greatly reduced rates of excretion. Flies fed continuously with dung containing ivermectin a t 0.25 pg g-' and 0.125 pg g-' in a regime identical to the above showed significantly increased abdominal mass compared with the controls. The observations are discussed in relation to other known effects of ivermectin on insect physiology.
Introduction
Ivermectin is a broad-spectrum antiparasitic drug derived from fermentation products of Srrepromyces avrrmifilis. It is used ex- tensively for the treatment of cattle to reduce nematode burdens and arthropod ectoparasites (Campbell, 1989). Approximately 90% of the drug administered to cattle by subcutaneous injec- tion is excreted in the faeces over a period of about 35 days fol- lowing treatment. The presence of ivermectin in the dung has deleterious effects on dung-breeding pests including the bush fly, Musca vetustissima Walker (Ridsdill-Smith, 1988), and the face fly, Musca autumnalis Degeer (Meyer et nl., 1980). How- ever, Wall & Strong (1987) have drawn attention to the potential damage caused by ivermectin residues which exert a toxic effect on many non-pest insects that utilize dung for breeding, and whose larval activity contributes greatly to dung breakdown and recycling.
Most studies of the non-specific effects of ivermectin have focused on its impact on larvae: much less is known of the ef- fects on adults caused by ingesting dung containing ivermectin residues. Mahon et al. (1993) observed increased mortality and reduced reproductive potential in Lucilia cuprina that had fed on dung deposited by sheep after treatment with an ivermectin drench. The survival and reproduction of adults of the fly Neomyia cornicina (F.), which feed on the liquid portion of freshly depos- ited pats (Hammer. 1941), are also significantly affected by
'This insect has been referred to as Cgprolucilia caesarion (Ham- mer, 1941) and Orthelia cornicinn (E) (Wardhaugh & Rodriguez- Menendez, 1988)
Correspondence: Dr J. Gover, School of Biological Sciences, Univer- sity of Bristol, Woodland Road, Bristol BS8 1 UG.
ivermectin residues (Gover & Strong, 1995). The ingestion of ivermectin causes bloating of the abdomen in adults of two spe- cies of Glossina (Distelmans et d.. 1983; Langley & Roe, 1984), and in Lucilia cuprina (Cook. 199 1). This bloating may be caused by water retention and thus affect mass changes. The aim of this study was to investigate the impact of ivermectin on fluid reten- tion and mass changes in adult females of N.cornicina fed on cattle dung containing ivermectin at concentrations similar to those found after injecting cattle with the drug. Such a study would add significantly to the growing knowledge of the impact of ivermectin on insects of pasture land.
Materials and Methods
Experirnenlul insecfs. Adult flies were maintained in a labora- tory colony at 25"C, 75% r.h. and LD 175 h photoperiodic re- gime. They were housed in cages (30 x 30 x 30 cm) and fed ad libitum on a mixture of egg powder, icing sugar, and skimmed milk powder in equal parts by weight: fresh liver was offered once weekly. Non-contaminated cattle dung (see below) was provided weekly for feeding and oviposition. 24 h after egg hatch. larvae were transferred to 200 g aliquots of fresh dung contained in a 15 cm diameter plastic dishes in which larval development was completed. Under the above conditions, the third-stage lar- vae wandered after about 6 days and were allowed to pupariate in clean sawdust. Adults emerged about 7 days later.
Dung snn7pple.s. Non-contammatcd cattlc dung was collcctcd from the dairy herd at thc Univcrsity of- Bristol farm at Langford (nr Bristol, Avon. U.K.), mixed thoroughly and stored frozen at -20°C until required. The dung was known to be non-contam- inated, because the cattle received no chemical treatments dur-
94 June Col.er nnd Les Strong
ing the period of collection. Experimental batches of dung were prepared using samples collected on a single day; this was to obviate problems arising from variations in dung quality.
Freshly thawed dung was weighed into 300 g samples and the required amount of IvomeP ( 1 R w/v ivermectin: MSD Agvet) added in 3 ml of ethanol, followed by 5 min vigorous stirring to ensure the even distribution of drug throughout the dung. The concentrations tested were 0.5,0.25 and 0.125 pg ivermectin per g fresh dung (pg g-I): control dung was prepared using 3 ml of ethanol alone. The above samples were prepared, divided into SO g batches, and frozen at -20°C until required, when they were thawed immediately prior to use.
Effects on mass and excretory rates. Adult flies that emerged over a 6 h period were sexed and segregated to prevent pre-trial mating; the females were maintained as above but in a labora- tory under a natural photoperiodic regime LD 1S:9 h. Females were used because abdominal bloating was clearer in this sex (personal observations). At dawn (6 a.m.), 5 days after emer- gence, forty females were removed at random from the holding cage, placed singly in a glass tube, and weighed on a Mettler AE200 balance. They were then allocated in turn to a control or experimental group. Each insect was isolated in a numbered plas- tic pot (SO0 ml) containing a water-soaked cotton pad and a weighing boat (3.5 cm9 containing about 8 g of control dung or dung containing 0.5 pg g ~ ' ivermectin. 12 h later. each female was removed from the container and weighed again. Also, the number of faecal spots deposited on the sides of the holding pot were counted; as each spot was recorded it was marked with ink to prevent re-counting later in the experiment. Before returning the females to their respective pots, the dung was stirred to retain a moist feeding surface. Data were always collected from the fernales in the same order as the initial allocations (e.g. first con- trol. fir\t treated. second control, second treated, etc.) to reduce time of day effects. At dawn each day, the dung was replaced with freshly thawed dung of the same drug concentration. The whole process of weighing and counting faecal spots was re- peated every 12 h until the sample size of any group fell below ten.
Thib experiment was repeated using concentrations of ivermectin of 0.25 yg g- ' and 0.125 pg g-': in the latter case, dawn time was adjusted to 5 a.m. and the evening data were collected at 5 p.m.
To te>t further whether ingestion of dung containing ivermectin was detrimental, flies alive at the end of the experimental period in tests using the two lower doses were transferred to a diet of sugar and water and their mortalities recorded for one week.
MU.F.S distriburion. Forty 5-day-old flies were randomly allo- cated t(> three treatment groups and given access ad libitum to water and either control dung, dung containing 0.125 pg g-' ivermectin. or dung containing 0.25 pg g-' ivermectin. Dung was replaced daily to maintain a moist surface for feeding. 4 days after the onset of dung feeding, surviving flies were frozen at -20°C. Once frozen, the legs were removed and the body divided into head, thorax and abdomen. The mass of each was recorded while still frozen. The wings were then detached and the length of the posterior cross vein measured as an indication of size of the in- sect. This nieasurement would not be affected by the experimental manipulations and could therefore be used to test whether there were differences in size of insects between treatments when they
were initially allocated to experimental groups. The cross vein was measured using a microscope equipped with an eyepiece micrometer.
Statistical analyses. Body mass and faecal spot data were ana- lysed using repeated-measures analyses of variance (Hand & Taylor, 1987) on SPSS (1988). The analysis comprised a between- subject factor ivermectin concentration and a within-subject factor time. Body mass analyses were performed on actual mass data. Faecal spot values were transformed using (x + 0.9)0.3 (Swaddle et al., 1994) to normalize the data. Normality was determined from normal probability plots. Analyses were performed using flies which survived until the end of each trial. For repeated- measures analyses of variance the parameter of particular inter- est is the dose x time interaction, not the effect of dose or time per se. Significance of this indicates that the effect of dose is different at different times, i.e. the treatment groups show differ- ent patterns of mass change or number of faecal spots produced.
Masses of head, thorax and abdomens were analysed using analysis of variance with ivermectin concentration as the factor. Multiple range tests were performed to identify differences be- tween treatment groups.
Results
Effect of ivermectin on muss changes
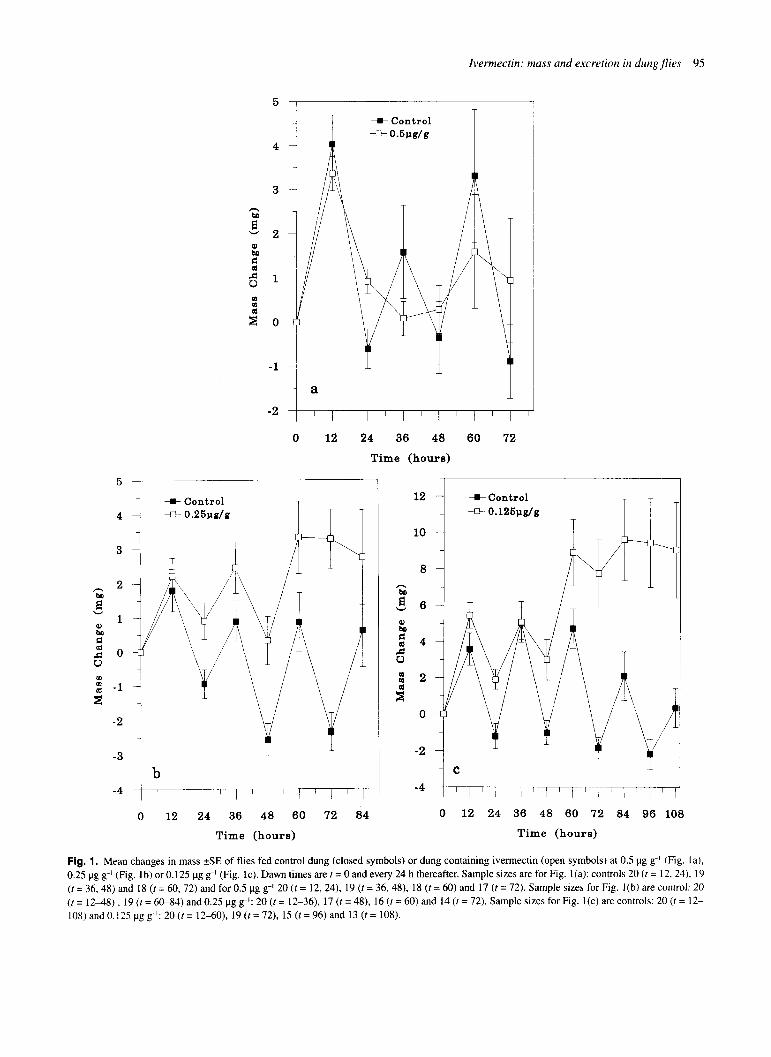
Figs l(a-c) show mean mass changes (kSE) of flies fed con- trol dung and dung containing 0.5 pg g-' 0.25 pg g-' or 0.125 yg g-' ivermectin respectively. In each case dawn occurred at time 0 and every 24 h thereafter. The effects of ivermectin ingestion were to increase dawn mass and to cause disruption of the daily oscillations in mass change which were seen in the controls. Flies fed ivermectin at 0.5 pg g-', 0.25 pg g-' and 0.125 pg g-' weighed on average 1.68, 5.4 and 12.01 mg more than their respective controls by the last dawn-weighing for each trial. These values correspond to an average of 5.73%, 22.51% and 38.93% increase in mass compared with controls.
Results for the analysis of trial l a (control versus 0.5 pg g-' ivermectin) are as follows: there was a significant dose x time interaction (F,,,,= 3.14, P = 0.018), a significant effect of time (F6,?,= 13.57, P < 0.001) but no significant effect of dose (FL,? ,= 0.01, P = 0.934). For trial l b (control versus 0.25 pg g-') there was a significant dose x time interaction (F,,2,= 19.17, P < 0.001), a significant effect of time (F,,,, = 16.64, P < 0.001), but no significant effect of dose (F , 2r= 2.83, P = 0.103). For the final analysis (control versus 0. I 25 bg 8-11 all effects were sig- nificant (dose x time (F,,22 = 4.92, P < 0.001), time (F,,, = 26.76, P < 0.001). dose (F,,>,= 5.84, P = 0.022)).
Effect of ivermectin on excretion
Figs 2(a-c) show the mean numbers of faecal spots produced by insects at intervals of 12 h. The effects of ivermectin ingestion were to reduce the levels of excretion, as measured by the amount of faecal spotting, and to disrupt the cyclic nature of excretion observed in the controls. Flies ingesting dung containing ivermectin produced an average of 2.00 2 0.657 (0.5 pg g-'), 2.21 k 1.196 (0.25 yg g-I) and 2.77 k 0.863 (0.125 pg g-I) faecal
Fig. 1. Mean changes in mass +SE of flies fed control dung (closed symbols) or dung containing ivermectin (open symbols) at 0.5 pg g-l (Fig. la), 0.25 pg g-' (Fig. lb) or 0.125 pg g-' (Fig. lc). Dawn times are t = 0 and every 24 h thereafter. Sample sizes are for Fig. l(a): controls 20 ( t = 12, 24). 19 ( t = 36.48) and 18 ( t= 60, 72) and for 0.5 pg g-l 20 ( t = 12, 24). 19 (t = 36,48), 18 (t = 60) and 17 ( r = 72). Sample sizes for Fig. I(b) are control: 20 (t = 1248) , 19 ( t = 60-84) and 0.25 pg g-I: 20 ( t = 12-36), 17 ( t = 48), 16 (t = 60) and 14 (t = 72). Sample sizes for Fig. l(c) are controls: 20 ( t = 12- 108) and 0.125 pg g-I: 20 ( t = 12-60), 19 ( t = 72). 15 (t = 96) and 13 (t = 108).
Fig. 2. Mean number of faecal spots +SE produced per 12 h time period for females fed water and either control dung (closed symbols) or dung containing iverniecrin (open symbols) at 0.5 pg g-' (Fig. 2a), 0.25 pg g-' (Fig. 2b) or 0.125 pg g-'. Dawn times are t = 0 and every 24 h thereafter. Sample w c s fu r each of the comparisons are as for the corresponding figures of mass change.
Ivermectin: mass and excretion in dung Pies 91
spots per final 12 h day period of each trial. Respective controls produced an average of 22.05 & 5.277, 28.47 f 5.96 and 72.63 f 9.74 spots.
Analysis of results obtained from the trial in which flies were fed either control dung or dung containing ivermectin of 0.5 pg g-' show there to be significant effects of dose (F,,,,= 49.44, P < 0.001) and time (F5,,,= 28.7, P < 0.001) on the number of faecal spots produced. The dose x time interaction approaches significance (F,,,, = 2.38, P = 0.064). The trial in which compari- sons were made between flies fed control dung and dung con- taining 0.25 pg g-' revealed significant effects of dose (F1,26= 83.02, P < 0.001) and time (F,,,,= 58.60, P < 0.001) and dose x time interaction (F,,z,= 9.43, P < 0.001). Similar results were found for the final comparison of control versus 0.125 pg g-I with dose (F, , , ,= 70.09, P < 0.001), time (F,,,,= 35.49, P < 0.001) and dose x time interaction (F,,,, = 13.03, P < 0.001), all being highly significant.
Mortalities
One week after the final mass data were collected, only one female out of twenty fed dung containing 0.25 pg g-I ivermectin was alive, whereas there were eighteen alive in the control group. A11 flies fed dung containing 0.125 pg g-' ivermectin had died within I week of the end of the experiment, with no additional deaths occurring during that time in the controls.
Mass distribution
Fig. 3 shows the ventral view of two flies one of which (a) was fed control dung and one (b) fed dung containing 0.125 pg g-I ivermectin. The increase in abdomen size can be seen clearly.
Fig. 4 shows the mean mass of the head, thoracic and abdomi- nal regions ( S E ) of the flies fed control dung (n = 32) dung containing 0.25 pg g-' (n = I 5) or 0.125 pg g-I (n = 29, except for head region where n = 28) ivermectin.
Fig. 3. Ventral view of a fly fed control dung (a) and a fly fed dung containing 0.125 pg g-I ivermectin (b) to show differences in abdominal sweiiing. The legs have been removed.
Control
20
18 -
16 - -
n
W iY1"
4 10 12 - PD
8 -
6 -
4 -
2 -
0 -
-
-
T
Head Thorax Abdomen
Fig. 4. Mean masses (mg kSE) of head, thoracic and abdominal re- gions of flies fed water and either control dung or dung containing 0.25 or 0.125 pg g-I ivermectin. Sample sizes for thoracic and abdominal re- gions are 32 (control), 29 (0.125 pg g-') and 15 (0.25 pg g-'). Sample sizes for the head region are the same except the 0.125 pg g-' sample which contained 28.
There were no significant differences between treatment groups for fly size, as measured by wing vein length (F , , , ,= 1.8, P = 0.173). There was a significant effect of ivermectin on the mass of all other body sections. For the head region the results of the ANOVA were: F2.72= 4.193, P = 0.019. A multiplerange test revealed the controls to be significantly heavier than those fed dung containing 0.25 pg g-I. Ivermectin had a significant effect on thoracic mass (F,,7, = 5.154, P = 0.008) with those of flies fed 0.25 pg g-' ivermectin dung being significantly lighter than con- trols and those fed 0.125 pg g-' ivermectin dung. Ivermectin con- centration had a highly significant effect on abdominal mass (F,,,,= 11.511, P < 0.001), with both ivermectin treatments being significantly heavier than controls but not significantly different from one another.
Discussion
Distelmans et al. (1983) and Langley & Roe (1984) observed abdominal bloating in two species of tsetse fly which had fed on blood containing ivermectin. The latter study included dissec- tions of the flies which failed to show that bloating was as a result of gut distension. It was concluded that fluid transporta- tion across the gut wall was unaffected but that Malpighian tu- bule function may have been impaired. In 1991, Cook also observed abdominal bloating in the sheep blowfly Lucilia cuprina, but found that the gut appeared highly swollen and tilled
98 June Gor*er rrnd Les Strong
with quantities of olive-green fluid. In neither study was this bloat- ing investigated quantitatively to clarify effects in the treated flies,
Females fed control dung in each of the present trials showed a regular daily cycling of mass change, with a rise during the day and fall overnight. In contrast, insects which had ingested dung containing ivermectin showed a gradual loss of this cyclic activ- ity and gained mass by the final dawn reading of each trial. The time taken for insects to show this disruption depended on the concentration of ivermectin. with those fed the lowest dose (0.125 pg g') showing patterns most similar to the controls for the longest time. In each of the three comparisons, analyses showed there to be differences in mass change patterns over the course of the experiments. The mass gains detected appeared to coincide with observed bloating, seen only in the ivermectin groups. The most extreme bloating was observed in flies given dung containing 0.125 pg g-' ivermectin. This might be explained in terms of lengthened time for feeding due to reduced toxicity effects, and greater ability to orientate towards and to ingest dung and water compared with those more disabled by higher concen- trations of the drug. At the end of the trial, flies from the group fed dung containing iverniectin at 0. I25 pg g ' were on average 1 1.6 mg heavier than their respective controls. Before dung feed- ing began, female mass ranged from 29.2 to 37.7 mg, therefore for even the heaviest adults the effect would be to increase mass by apprnximately 30%. As shown in the Results section, this effect was even more dramatic at the dawn reading (1 2 h prior to the end of the experiment), where flies which had fed on the dung containing iverniectin at 0.125 pg g-' were on average 38.9% heavier than their respective controls. These mass changes may. in part, explain effects of ivermectin ingestion on locomotory behaviour and ease of capture (Gover & Strong, unpublished).
In all the trials. mass gain over the first 12 h period was simi- lar for controls and those fed dung containing ivermectin. This suggests that ivermectin does not show anti-feedant properties, at least in the short term. If the mass changes observed in flies that had ingested dung containing ivermectin were due to the laying down of energetic reserves, then they could perhaps be considered to be beneficial. However, the high mortalities seen in these flies both during and after the experimental period fail to support this interpretation. Although it cannot be concluded that the mass gains per se are causing the increases in mortality, ivermectin is clearly causing severe disruption of normal diurnal mass change patterns. and increasing mortality.
Excretion rate, as estimated by the number of faecal spots pro- duced by inviduals within each 12 h time period, showed a simi- lar cyclical pattern to that of mass change in females fed control dung. Ivermectin ingestion caused significant changes in these patterns of excretion and significantly reduced the levels of ex- cretion >I \ measured by the number of faecal spots produced. The reduction in excretion was most rapid and most marked in flies fed dung containing 0.5 pg g ', but showed a more gradual change in flies fed at the lowest concentration.
Ivermectin I S thought to act by disrupting neural transmission at GABA-mediated synapses which are found at neuromuscular junctions in insects (Campbell, 1989), leading to paralysis. One possibility, therefore. that might explain these effects on mass change and excretion is rbat ivermectin directly affects the mus- culature of the gut wall and prevents defaecation. If this were the
case it would be expected that dissection would reveal a highly distended gut as found by Cook (1991). Dissection of insects which appeared very bloated, however, did not reveal any gut distension. Additionally, even highly bloated flies were observed to defaecate during the course of the experiment. As found by Langley & Roe (1984), dissection revealed that the haemocoel contained large quantities of fluid. One further observation noted during the course of the experiment was that a number of fe- males in which the abdomen was swollen showed the ovipositor and proboscis to be extended. This is likely to have been due to high haemocoelic pressure.
The final part of this study confirmed that the most dramatic differences in the masses of the three body parts were between abdominal regions with control flies having significantly lighter abdomens than flies receiving ivermectin. Higher masses of head and thorax regions of flies fed control dung may be due to hav- ing fuller digestive tracts with this effect being masked in the abdomen by the increase in volume of haemolymph in flies fed dung containing ivermectin. The abdomen is the most likely area to respond to this increase due to the greater flexibility of the exoskclcton coinpared with that of the head and thorax.
In conclusion, this study shows that the presence of ivermectin in the diet at concentrations normally found in the dung of treated cattle causes severe disruption of normal mass change patterns and excretory processes in the fly Neomyia cornicina. As sug- gested by Langley & Roe (1984), this may occur due to an in- crease in the volume of haemolymph as a result of failure in Malpighian tubule function, although the actual mechanism is not known.
Acknowledgments
The authors thank Dr Mark Witter for his help and statistical advice. The project was funded by a grant from the BBSRC.
References
Campbell, W.C. (1989) Zvermectin andAbamectin. Springer, New York. Cook, D.F. (1991) Ovarian development in females of the Australian
sheep blow fly Lucilia cuprina (Diptera: Calliphoridae) fed on sheep faeces and the effect of ivermectin residues. Bulletin ojEntomologi- cul Research, 81,249-256.
Distelmans, W., D'Haeseleer, E & Mortelmans, J. (1983) Efficacy of systemic administration of ivermectin against tsetse flies. Annales de la Socie'tk Belge de Medecine Tropicale, 63, 119-125.
Gover, J. & Strong, L. (1995) The effects of ivermectin in ingested cow dung on the mortality and oviposition of the dung fly Neomyia cornicina (Diptera: Muscidae). Bulletin of Entomological Research (in press).
Hammer, 0. (1941) Biological and ecological investigations of flies as- sociated yith cattle and their excrement. Videnskabelige Meddelelser fra Dansk Naturhistorisk Forening, 105, 141-393.
Hand, D.J. &Taylor, C.C. (1987) Multivariate Analyses ofvariance and Repeated Measures: a practical approach for the behavioural sciences. Chapman & Hall, London.
Langley, P.A. & Roe, J.M. (1984) lvermectin as a possible control agent for the tsetse fly, Glossina morsitans. Entomologia Experimentalis et Applicnta, 36, 137-143.
Ivermectin: mass and excretion in dung flies 99
Mahon, R.J., Wardhaugh, K.G., van Genven, A.C.M. & Whitby, W.A. (1993) Reproduction development and survival of Lucilia cuprina Weidermann when fed sheep dung containing ivermectin. Veterinary Parasitology, 48, 205-214.
Meyer, J.A., Simco, J.S. & Lancaster, J.L. (1980) Control of face fly development with ivermectin, MK-933. Southwesrern Entomologisr,
Ridsdill-Smith, T.J. (1988) Survival and reproduction of Musca vetustissima Walker (Diptera: Muscidae) and a Scarabaeine dung bee- tle in dung of cattle treated with avermectin B,. Journal of the Aus- tralian Entomological Socieg, 27, 175-178.
Sommer, C., Steffansen, B., Overgaard Nielsen, B., Gronvold, J., Vagn Jensen, K.-M., Brochner Jesperson, J., Springborg, J. & Nansen, P. (1992) Ivermectin excreted in cattle dung after subcutaneous injec-
5,201-209.
tion or pour-on treatment: concentrations and impact on dung fauna. Bulletin of Entomological Research, 82,251-264.
SPSS, (1988) SPSSx Users'Guide, 3rd edn. SPSS Inc., Chicago. Swaddle, J.P., Witter, M.S. & Cuthill, LC. (1994) The analysis of fluctu-
ating asymmetry. Animal Behaviour; 48,986989. Wall, R. & Strong, L. (1987) Environmental consequences of treating
cattle with the antiparasitic drug ivermectin. Nature, 327.4 18-42 1, Wardaugh, K.G. & Rodriguez-Menendez, H. (1988) The effects of the
antiparasitic drug, ivermectin, on the development and survival of the dung breeding fly Orthelia cornicina (F.) and the scarabaeine dung beetles, Copris hispanus L., Bubas bubalus (Oliver) and Onitis belial F. Journal ofApplied Entornolog): 106,381-389.