Effects of neural androgen receptor disruption on aggressive behavior, arginine vasopressin and galanin systems in the bed nucleus of stria terminalis and lateral septum Clarisse Marie-Luce a,b,c,1 , Kalina Raskin a,b,c,1 , Matei Bolborea d,2 , Marion Monin a,b,c , Marie Picot a,b,c , Sakina Mhaouty-Kodja a,b,c,⇑ a Centre National de la Recherche Scientifique (CNRS) Unité Mixte de Recherche (UMR) 7224, 9 quai St Bernard, Paris CEDEX 05, France b Institut National de la Santé et de la Recherche Médicale (INSERM) U952, 9 quai St Bernard, Paris CEDEX 05, France c Université Pierre & Marie Curie, 9 quai St Bernard, Paris CEDEX 05, France d Institut des Neurosciences Cellulaires et Intégratives, CNRS UPR 3212, Département de Neurobiologie des Rythmes, 5 Rue Blaise Pascal, 67084 Strasbourg CEDEX, France article info Article history: Available online 10 April 2013 Keywords: Testosterone Aggressive behavior Arginine vasopressin Galanin Nervous system Androgen receptor abstract In the present study, we investigated the role of the androgen receptor (AR) in the nervous system in the regulation of aggressive behavior and arginine vasopressin and galanin systems by testosterone. For this purpose, we used a conditional mouse line selectively lacking AR gene in the nervous system, backcrossed onto the C57BL/6J strain. Adult males were gonadectomized and supplemented with similar amounts of testosterone. When tested on two consecutive days in the resident intruder paradigm, fewer males of the mutant group exhibited aggressive behavior compared to their control littermates. In addition, a high latency to the first offensive attack was observed for the few animals that exhibited fighting behavior. This alteration was associated with a normal anogenital chemoinvestigation of intruder males. In olfac- tory discrimination tasks, sexual experience enhanced preference towards female-soiled bedding rather than male-soiled bedding and estrus females rather than intact males, regardless of genotype. This indi- cated that the behavioral alteration induced by neural AR mutation occurs in brain areas located down- stream from the olfactory bulb. Quantification of the sexually dimorphic cell populations expressing preprovasopressin and galanin mRNAs in the bed nucleus of stria terminalis (BNST) and vasopressin-neu- rophysin 2 and galanin immunoreactivity in the lateral septum showed no significant differences between the two genotypes. The present findings indicate that the neural AR is required in the expression of aggressive behavior but not in the sexual differentiation of AVP and galanin cell number in the BNST and fiber immunoreactivity in the lateral septum. They also suggest that AR in the nervous system could mediate activational effects of testosterone in the regulation of aggressive behavior during adulthood. Ó 2013 Elsevier Inc. All rights reserved. 1. Introduction Testosterone has a central role in the regulation of male behav- iors ranging from partner preference and mating to aggression and territoriality. In male rodents, testosterone liberated by fetal and neonatal testes permanently potentiates male (masculinization) and inhibits female (defeminization) behavioral and neuroana- tomical characteristics. These organizational effects of testosterone result in sex differences in brain anatomy at the structural, neurochemical and molecular levels. Sex differences in cell number and morphology or fiber density have been widely described in rat brain nuclei that regulate aggressive behavior like the bed nucleus of stria terminalis (BNST) and the medial amygdala (MeA). These brain structures contain cells expressing arginine-vasopressin (AVP), a key neuromodulator of social behaviors. Cells expressing AVP are more numerous in the male than in the female (van Leeuwen et al., 1985; De Vries and Al-Shamma, 1990). Further- more, density of AVP fibers in the lateral septum is also more important in rat males than in females (van Leeuwen et al., 1985; De Vries and Al-Shamma, 1990). The sexual dimorphism of AVP system is organized perinatally (De Vries and Al-Shamma, 1990; Wang et al., 1993) and involves estradiol (Han and De Vries, 2003). However, it seems also maintained by activational effects of testosterone during adulthood (De Vries et al., 1994). In fact, adult castration of rats results in decreased expression of AVP mRNA and reduced AVP immunoreactivity in the BNST and the MeA, whereas testosterone supplementation restores AVP to levels noted in in- 0016-6480/$ - see front matter Ó 2013 Elsevier Inc. All rights reserved. http://dx.doi.org/10.1016/j.ygcen.2013.03.031 ⇑ Corresponding author. Address: CNRS UMR 7224, Université P. & M. Curie, 9 quai St Bernard Bât B 2ème étage, Paris CEDEX 05, France. Fax: +33 1 44 27 25 08. E-mail address: [email protected](S. Mhaouty-Kodja). 1 These authors contributed equally to this work. 2 Present address: School of Life Sciences, University of Warwick, Gibbet Hill Road, Coventry CV4 7AL, United Kingdom. General and Comparative Endocrinology 188 (2013) 218–225 Contents lists available at SciVerse ScienceDirect General and Comparative Endocrinology journal homepage: www.elsevier.com/locate/ygcen
Transcript
General and Comparative Endocrinology 188 (2013) 218–225
Contents lists available at SciVerse ScienceDirect
General and Comparative Endocrinology
journal homepage: www.elsevier .com/locate /ygcen
Effects of neural androgen receptor disruption on aggressive behavior, argininevasopressin and galanin systems in the bed nucleus of stria terminalis andlateral septum
Clarisse Marie-Luce a,b,c,1, Kalina Raskin a,b,c,1, Matei Bolborea d,2, Marion Monin a,b,c, Marie Picot a,b,c,Sakina Mhaouty-Kodja a,b,c,⇑a Centre National de la Recherche Scientifique (CNRS) Unité Mixte de Recherche (UMR) 7224, 9 quai St Bernard, Paris CEDEX 05, Franceb Institut National de la Santé et de la Recherche Médicale (INSERM) U952, 9 quai St Bernard, Paris CEDEX 05, Francec Université Pierre & Marie Curie, 9 quai St Bernard, Paris CEDEX 05, Franced Institut des Neurosciences Cellulaires et Intégratives, CNRS UPR 3212, Département de Neurobiologie des Rythmes, 5 Rue Blaise Pascal, 67084 Strasbourg CEDEX, France
0016-6480/$ - see front matter � 2013 Elsevier Inc. Ahttp://dx.doi.org/10.1016/j.ygcen.2013.03.031
⇑ Corresponding author. Address: CNRS UMR 7224quai St Bernard Bât B 2ème étage, Paris CEDEX 05, Fr
E-mail address: [email protected] (S1 These authors contributed equally to this work.2 Present address: School of Life Sciences, University
Coventry CV4 7AL, United Kingdom.
In the present study, we investigated the role of the androgen receptor (AR) in the nervous system in theregulation of aggressive behavior and arginine vasopressin and galanin systems by testosterone. For thispurpose, we used a conditional mouse line selectively lacking AR gene in the nervous system, backcrossedonto the C57BL/6J strain. Adult males were gonadectomized and supplemented with similar amounts oftestosterone. When tested on two consecutive days in the resident intruder paradigm, fewer males of themutant group exhibited aggressive behavior compared to their control littermates. In addition, a highlatency to the first offensive attack was observed for the few animals that exhibited fighting behavior.This alteration was associated with a normal anogenital chemoinvestigation of intruder males. In olfac-tory discrimination tasks, sexual experience enhanced preference towards female-soiled bedding ratherthan male-soiled bedding and estrus females rather than intact males, regardless of genotype. This indi-cated that the behavioral alteration induced by neural AR mutation occurs in brain areas located down-stream from the olfactory bulb. Quantification of the sexually dimorphic cell populations expressingpreprovasopressin and galanin mRNAs in the bed nucleus of stria terminalis (BNST) and vasopressin-neu-rophysin 2 and galanin immunoreactivity in the lateral septum showed no significant differencesbetween the two genotypes.
The present findings indicate that the neural AR is required in the expression of aggressive behavior butnot in the sexual differentiation of AVP and galanin cell number in the BNST and fiber immunoreactivityin the lateral septum. They also suggest that AR in the nervous system could mediate activational effectsof testosterone in the regulation of aggressive behavior during adulthood.
� 2013 Elsevier Inc. All rights reserved.
1. Introduction and morphology or fiber density have been widely described in rat
Testosterone has a central role in the regulation of male behav-iors ranging from partner preference and mating to aggression andterritoriality. In male rodents, testosterone liberated by fetal andneonatal testes permanently potentiates male (masculinization)and inhibits female (defeminization) behavioral and neuroana-tomical characteristics. These organizational effects of testosteroneresult in sex differences in brain anatomy at the structural,neurochemical and molecular levels. Sex differences in cell number
ll rights reserved.
, Université P. & M. Curie, 9ance. Fax: +33 1 44 27 25 08.. Mhaouty-Kodja).
of Warwick, Gibbet Hill Road,
brain nuclei that regulate aggressive behavior like the bed nucleusof stria terminalis (BNST) and the medial amygdala (MeA). Thesebrain structures contain cells expressing arginine-vasopressin(AVP), a key neuromodulator of social behaviors. Cells expressingAVP are more numerous in the male than in the female (vanLeeuwen et al., 1985; De Vries and Al-Shamma, 1990). Further-more, density of AVP fibers in the lateral septum is also moreimportant in rat males than in females (van Leeuwen et al.,1985; De Vries and Al-Shamma, 1990). The sexual dimorphism ofAVP system is organized perinatally (De Vries and Al-Shamma,1990; Wang et al., 1993) and involves estradiol (Han and De Vries,2003). However, it seems also maintained by activational effects oftestosterone during adulthood (De Vries et al., 1994). In fact, adultcastration of rats results in decreased expression of AVP mRNA andreduced AVP immunoreactivity in the BNST and the MeA, whereastestosterone supplementation restores AVP to levels noted in in-
C. Marie-Luce et al. / General and Comparative Endocrinology 188 (2013) 218–225 219
tact males (Brot et al., 1993; De Vries et al., 1994). In this context,both the androgenic and the estrogenic metabolites of testosteronewere needed to fully restore AVP expression in the rat BNST (Brotet al., 1993; De Vries et al., 1994). In the central nervous system,testosterone acts directly by stimulating the androgen receptor(AR) or can be metabolized in situ by the cytochrome P450 aroma-tase into 17-b estradiol, which then activates its receptors ERa andERb. AR and ERs belong to the nuclear receptor superfamily.
The mechanisms underlying the effects of testosterone in theregulation of aggression and AVP system have been addressed byusing transgenic mice ubiquitously lacking Cyp19 aromatase orERs. Males with Cyp19 aromatase deletion exhibit deficient socialrecognition ability and reduced AVP-immunoreactivity in the BNSTand MeA (Pierman et al., 2008; Plumari et al., 2002). Adult treat-ment of these mice with estradiol restored social recognition andAVP-immunoreactivity in the lateral septum, suggesting that theandrogenic rather than the estrogenic pathway plays a role inthe organizational effects of testosterone (Pierman et al., 2008).Nevertheless, data obtained in males lacking ERa gene showed asevere aggressive behavioral deficit (Ogawa et al., 1997) and adulttreatment of these males with estradiol did not restore the behav-ioral deficiency (Scordalakes and Rissman, 2004). In these mice,AVP-immunoreactivity was found reduced only when both ARand ERa were lacking, suggesting that AR may contribute withERa in the organization of AVP-immunoreactivity during develop-ment (Scordalakes and Rissman, 2004).
The direct contribution of AR in aggressive behavior has beenstudied in mice carrying spontaneously mutated AR gene (Testicu-lar Feminization Mutation) or ubiquitously invalidated for AR gene(ARKO). Genetic (XY) males with Tfm or AR mutation have femaleexternal genitalia, undifferentiated urogenital tract and low levelsof testosterone during adulthood. Aggressive behavior was foundreduced in Tfm mice (Ohno et al., 1974) and ablated in ARKO ani-mals (Sato et al., 2004). However, a comparable behavior and AVPimmunoreactivity system was described in Tfm and wild type micesubjected to adult gonadectomy and estradiol supplementation(Scordalakes and Rissman, 2004).
We recently generated a conditional knockout mouse line lack-ing AR in the nervous system (Raskin et al., 2009). Unlike Tfm andARKO mice, the conditional mutant males develop male externalgenitalia and functional testes producing testosterone. Only asmall proportion of mutants displayed aggressive behavior (Raskinet al., 2009). However, this first study was performed on mice ofmixed genetic background and did not investigate in parallel theAVP system. In fact, the effects of AR gene disruption on aggressivebehaviors may be modified by background strain, as reported forother genes (Dominguez-Salazar et al., 2004; Le Roy et al., 2000).
The first aim of this study was the analysis of aggressive behav-ior in mice with a nervous system knockout of AR backcrossed ontoC57BL/6J, a strain widely used in reproductive behavior studies(Burns-Cusato et al., 2004). As aggressive behavior is induced byolfactory cues in rodents, we investigated whether the disruptedbehaviors of mutant males could be accounted for by deficienciesin olfactory preference. We also quantified mRNAs positive cellsfor AVP in the BNST and AVP fiber immunoreactivity in the lateralseptum. Similar quantifications were performed for galaninexpressing cell population known to be also sexually dimorphicin the BNST of mice (Rajendren et al., 2000).
2. Materials and methods
2.1. Animals
ARNesCre mice were initially obtained in a mixed genetic back-ground (C57BL6/J and 129SvEv), by crossing floxed AR mice and
transgenic mice expressing Cre recombinase under the control ofrat nestin (Nes), as previously described (Raskin et al., 2009). Thismouse line was backcrossed for at least 9 generations onto theC57BL/6J strain. Mice were weaned at 24–26 days of age andhoused in groups in conditions of controlled photoperiod (12 hlight/12 h dark cycle – lights on at 7 am) and temperature(22 �C), with free access to food and water. Genotyping was per-formed as previously described (Raskin et al., 2009). All animalstudies were performed in accordance with the NIH guidelinesfor care and use of Laboratory animals (NIH Guide) and Frenchand European legal requirements (Decree 87-848, 86/609/ECC).
2.2. Treatments
Adult males (2–3 months of age) were anesthetized with xyla-zine/ketamine, gonadectomized and implanted with Silastic�
tubes containing 10 mg of testosterone (Sigma–Aldrich, St.-Quen-tin Fallavier, France) as previously described (Raskin et al., 2012).Behavioral studies were conducted on males between two and fourweeks after treatment (Groups 1 and 2). In situ hybridization andimmunohistochemistry were performed four weeks after treat-ment (Group 3). In these conditions, circulating levels of testoster-one are similar between control and mutant males (Raskin et al.,2012). Indeed, in the present study, the weight of the androgen-sensitive seminal vesicles was comparable between the two geno-types (0.0068 ± 0.0007% of body weight for controls versus0.0064 ± 0.0004% of body weight for mutants).
Estrus C57BL/6J females (Janvier, Le Genest, France) were ovari-ectomized, implanted with Silastic� implants filled with estradiol-benzoate and subcutaneously treated with progesterone (Sigma–Aldrich) 4–5 h before the tests as previously reported (Raskinet al., 2009, 2012).
2.3. Resident-intruder test
Naive males of Group 1 were individually housed for two weekswithout bedding changes two weeks after treatment. They werethen tested in their home cages for two consecutive days by usingA/J mice (The Jackson Laboratory, USA) as intruders as previouslydescribed (Raskin et al., 2009). The tests were conducted underred-light illumination 2 h after lights off and lasted 10 min. Thelatencies to the first anogenital chemoinvestigation and first offen-sive attack (defined by biting and wrestling) displayed toward theintruder mice were scored for each resident animal. When no at-tack occurred, the latency was 600 s.
2.4. Olfactory preference
Two weeks after treatment, naive males of Group 2 were testedfor olfactory preference (Test 1). One week later, males were matedwith receptive females and let to reach ejaculation as previouslydescribed (Raskin et al., 2009), before tested again one week laterfor their olfactory preference (Test 2). All mice exhibited a com-plete sexual behavior, but mutant males took longer to show thefirst mount, intromission and to reach ejaculation compared totheir control littermates (63 ± 23 min vs 7 ± 1 min, 72.4 ± 24 minvs 10.4 ± 2 min, 94.3 ± 23 min vs 49.5 ± 6.3; respectively).
Olfactory preference was conducted in an enclosed PlexiglasY-maze, under red-light illumination 2 h after lights off. Mice wereinitially allowed to become familiar, for 5 min on each of two con-secutive days, with the experimental paradigm, but with no stim-ulus in the goal boxes. On the day of the test, mice were offered thechoice between bedding soiled by males or estrus females and be-tween an estrus female and a gonadally intact male. Male-soiledbedding was obtained from animals placed in a cage with clean
220 C. Marie-Luce et al. / General and Comparative Endocrinology 188 (2013) 218–225
bedding 24 h before the test. Female-soiled bedding was obtainedfrom a group of five estrus females.
The time spent sniffing each goal box was scored over the five-minute test. The maze was cleaned with 10% ethanol between trials.
2.5. Non radioactive in situ hybridization
Four weeks after treatment, males of Group 3 were transcardial-ly perfused with 4% paraformaldehyde. The brains were postfixedfor 8 h at 4 �C, then rinsed in 50% ethanol and embedded in polyeth-ylene glycol (Klosen et al., 1993). Coronal brain sections (12 lm)were processed as previously described (Bolborea et al., 2010)and labeled with rat vasopressin-neurophysin (472 pb) or rat gala-nin (431 bp) riboprobes. Briefly, the hybridization was performedwith a digoxigenin-labeled riboprobe. Digoxigenin-labeled boundprobes were detected with alkaline phosphatase-labeled antidigox-igenin antibodies and alkaline phosphatase activity was detectedwith bromo-chloro-indolyl phosphate and nitroblue tetrazolium.
Positive cells were counted throughout the whole BNST (leftand right) and multiplied by 7 (1 in 7 serial sections) to estimatethe total number of cells.
2.6. Immunohistochemistry
Immunodetection of AVP and galanin was performed on malesas previously described (Bolborea et al., 2010) by using the rabbitanti-vasopressin-neurophysin 2 (Sigma N0744, St. Quentin Falla-vier, France) and rabbit anti-galanin (Peninsula-Bachem T4330,San Carlos, CA, USA). Briefly, 12 lm sections were blocked for 1 hwith 3% dry skimmed milk in TBS containing 0.05% Tween 20 thenincubated with the primary antibody overnight. The primary anti-body was detected with a biotinylated donkey anti-rabbit antise-rum (Jackson Immunoresearch Laboratories, West Grove PA, USA)for 1 h. Bound antibodies were visualized by incubation with thebiotin-streptavidin complex reagent (ABC kit; Vector Laboratories),followed by a color development with 3,30-diaminobenzidine tetr-ahydrochloride chromogenic substrate from Sigma–Aldrich.
The quantification of the immunoreactivity in the lateral sep-tum was done using ImageJ software. A background image of eachslide was subtracted from the corresponding sample image, thenthis one was threshold to delineate the stained lateral septum.The same threshold level was used. A region of interest, includingthe whole lateral septum, was then selected and the integrateddensity measured.
2.7. Statistics
The percentages of animals showing behaviors were comparedby Fisher’s Exact Probability Test. All other data are expressed asmeans ± S.E.M. Analyses were performed by two-way ANOVA forrepeated measurements for the main effects of genotype, testday, and their interaction followed by Bonferroni tests for posthoc comparisons. When variances were not homogeneous betweengroups, data were analyzed by non-parametric tests (Kruskal–Wal-lis followed by post hoc Mann–Whitney U test for independentsamples, or Wilcoxon matched pairs signed ranks test for repeatedmeasurements). One-way ANOVA or Student’s t tests were used forunpaired data. P values less than 0.05 were considered to besignificant.
3. Results
3.1. Reduced aggressive behavior of ARNesCre males
In the resident-intruder paradigm over two consecutive days,all resident males, regardless of genotype, exhibited anogenital
chemo-investigation towards intruder mice. Two-way ANOVAshowed no effect of genotype (F(1,10) = 0.91, P = 0.39) or experience(F(1,10) = 0.77, P = 0.40) on the latency to the first anogenital chemo-investigation (Fig. 1A). By contrast, fewer males of the mutantgenotype compared to their control littermates showed an aggres-sive behavior during the two tests (P < 0.05) (Fig. 1B). Although theproportion of mutant males exhibiting a fighting behavior slightlyincreased during the second test, it remained lower than that ob-served for the control group. Analysis of the latency to the first at-tack revealed an effect of genotype (P < 0.001) (Fig. 1C). Post hocanalysis showed a longer latency to attack for mutant males thancontrol mice. A significant effect of experience was also foundwhen the latency to the first attack was compared between thetwo tests (P < 0.05).
3.2. Sexual experience enhanced olfactory preference in both controland mutant males
We investigated whether the disrupted behavior of ARNesCre
mice could be explained by a deficiency in their olfactory prefer-ence. For this purpose, preferences for male-soiled or female-soiledbedding and for an intact male or an estrus female were comparedbetween control and mutant males either naive or sexually experi-enced. Time spent sniffing the bedding or stimulus was determinedand expressed as a percentage of the total time devoted to chemo-investigation. The total time devoted to chemoinvestigation didnot differ significantly between males, regardless of genotype orsexual experience state (�90 s).
In Test 1, naive males displayed no behavioral discrimination asthey spent similar amounts of time investigating male- and fe-male-soiled bedding (Fig. 2A). Indeed, two-way ANOVA showedno effect of genotype (F(1,20) = 0.00, P = 1) or bedding stimulus(F(1,20) = 0.16, P = 0.69). Similar data were obtained for the chemo-investigation of animal stimuli (F(1,20) = 0.00, P = 1 for genotype andF(1,20) = 0.00, P = 0.97 for stimulus) (Fig. 2B).
Males were then mated with estrus females and one week latertested again for olfactory preference (Test 2). In these conditions,there was an effect of bedding (F(1,14) = 7.57, P = 0.0156) and animalstimulus (F(1,18) = 20.41, P = 0.0003) on male chemoinvestigation(Fig. 2A and B). No effect of genotype was found on the time spentinvestigating bedding (F(1,14) = 0.00, P = 1) or animal stimuli(F(1,18) = 1.33, P = 0.2633).
3.3. Prepro-vasopressin and prepro–galanin mRNAs in the BNST andvasopressin-neurophysin 2 and galanin immunoreactivity in thelateral septum
The unaffected olfactory preference of ARNesCre mice stronglysuggested that the behavioral alteration might occur downstreamfrom the olfactory in brain areas underlying aggressive behavior.In these brain areas, AVP is a key neuromodulator of such behav-iors. The number of AVP-positive cells located in BNST and project-ing AVP-immunoreactive fibers to the lateral septum are sexuallydimorphic, under the influence of testosterone. We thus comparedthe number of preprovasopressin mRNAs positive cells in the BNSTand AVP-immunoreactivity in the lateral septum.
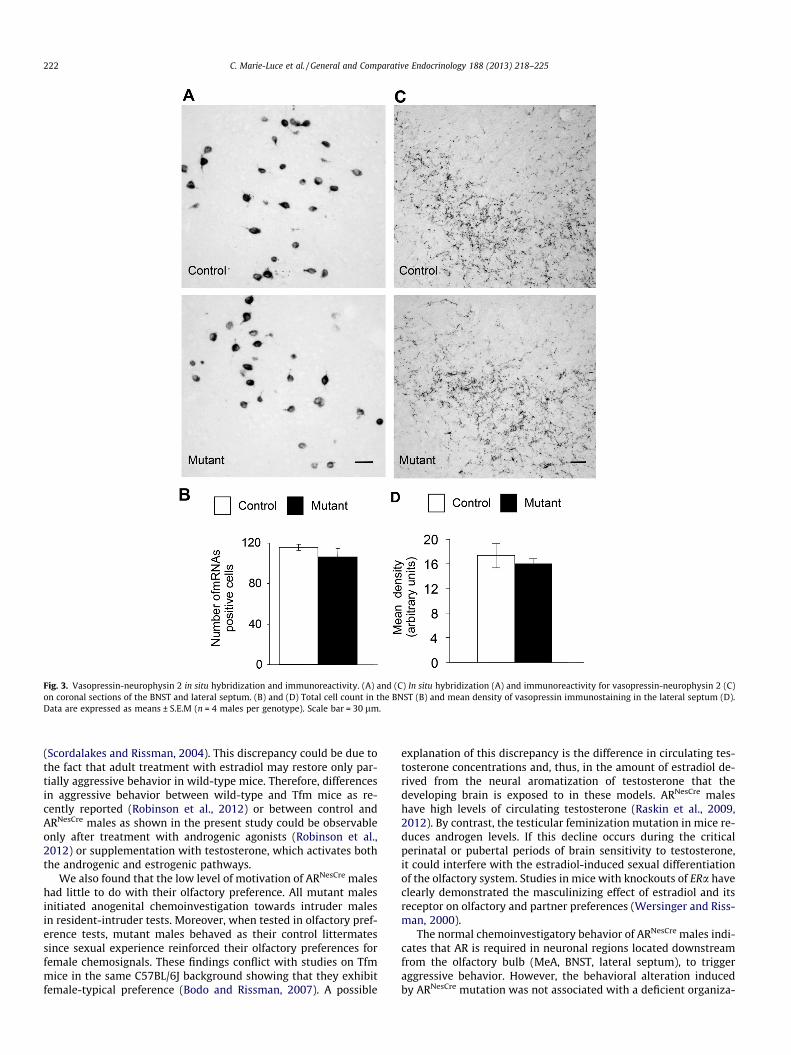
In situ hybridization for preprovasopressin mRNA labeled neu-ronal cell bodies in the BNST of both genotypes (Fig. 3A). Quantifi-cation of the number of mRNAs positive cells was not statisticallydifferent between control males and their mutant littermates(Fig. 3B). The vasopressin antisera stained a plexus of vasopres-sin-neurophysin 2 positive fibers in the lateral septum (Fig. 3C).Quantification of vasopressin-neurophysin 2 fiber density did notshow any significant differences between the two genotypes(Fig. 3D).
Fig. 1. Aggressive behavior of mice in the resident-intruder paradigm over twoconsecutive days. (A) Latency to the first anogenital chemo-investigation ofintruders. (B) Percentage of control and mutant males showing or not andaggressive behavior in Tests 1 and 2. (C) Latency to the first attack of intrudermales. Data are expressed as means ± S.E.M of n = 6 males per genotype. (a),P < 0.001 compared to control males; (b), P < 0.05 compared to Test 1.
Fig. 2. Olfactory preference of control and mutant males. (A) Time spent chemo-investigating male-soiled versus female-soiled-bedding in Tests 1 and 2. (B) Timespent chemoinvestigating gonad-intact male versus estrus female in Tests 1 and 2.Data are expressed as means ± S.E.M (n = 12 males per genotype). (a), P < 0.05compared to male stimulus.
C. Marie-Luce et al. / General and Comparative Endocrinology 188 (2013) 218–225 221
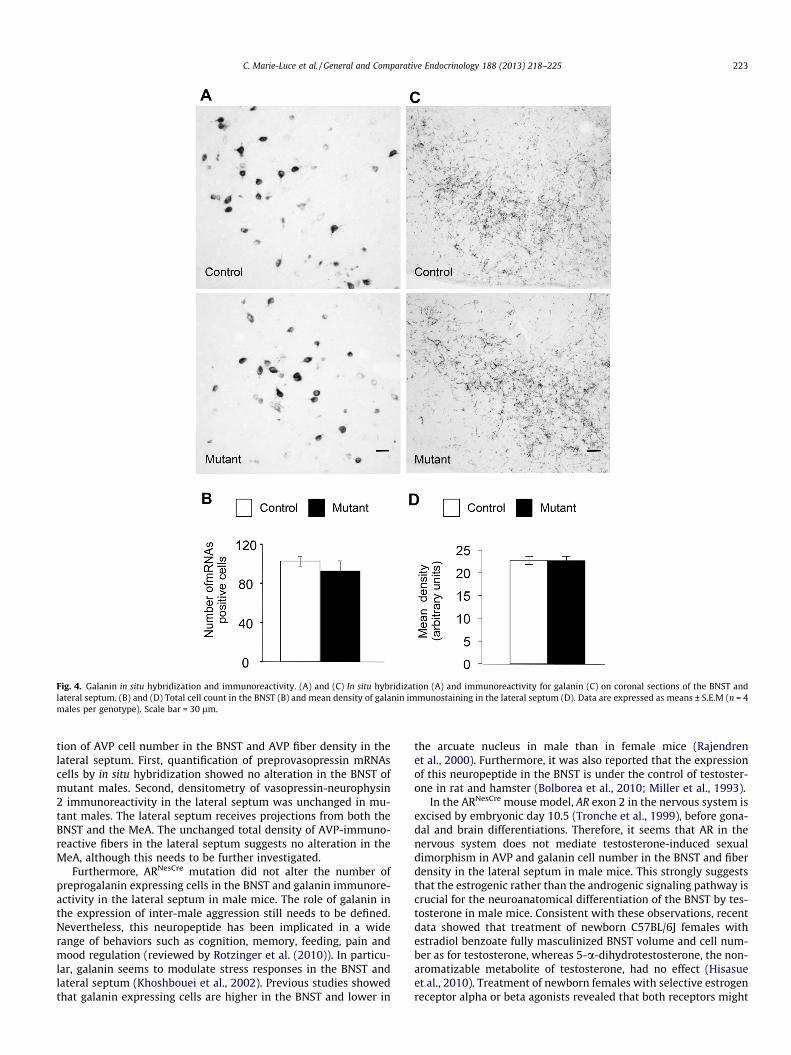
We also compared the number of mRNAs positive cells for an-other sexually dimorphic neuronal population in the BNST. Indeed,the number of preprogalanin expressing cells is higher in malesthan in females in mice (Rajendren et al., 2000). Similar to resultsobtained for AVP, we found a comparable number of mRNAs prep-rogalanin cells in the BNST (Fig. 4A and B) and density of projectinggalanin-immunoreactive fibers in the lateral septum between con-trol and mutant males (Fig. 4C and D).
The area fraction covered by vasopressin-neurophysin 2- andgalanin-immunoreactive fibers was also similar between control(0.067 ± 0.009 and 0.089 ± 0.003, respectively) and mutant males(0.060 ± 0.003 and 0.088 ± 0.004, respectively).
4. Discussion
AR disruption in the nervous system clearly interferes with theaggressive behavior of male mice of the C57BL/6J strain. Only a lowpercentage of gonadectomized ARNesCre males supplemented withtestosterone exhibited an aggressive behavior towards intrudermice compared to similarly treated control littermates. Further-more, an effect of genotype was found for the latency to the firstattack. Our results confirm our previous data for mice of mixed ge-netic background (Raskin et al., 2009). They are also in agreementwith a very recent study where gonadally intact Tfm mice givenvehicle or anabolic androgenic steroids were slower to attack andexhibited fewer aggressive events than wild type males (Robinsonet al., 2012). These data differ from the results showing no differ-ences in aggressive behavior between gonadectomized wild-typeand Tfm mice implanted with estradiol-containing implants
Fig. 3. Vasopressin-neurophysin 2 in situ hybridization and immunoreactivity. (A) and (C) In situ hybridization (A) and immunoreactivity for vasopressin-neurophysin 2 (C)on coronal sections of the BNST and lateral septum. (B) and (D) Total cell count in the BNST (B) and mean density of vasopressin immunostaining in the lateral septum (D).Data are expressed as means ± S.E.M (n = 4 males per genotype). Scale bar = 30 lm.
222 C. Marie-Luce et al. / General and Comparative Endocrinology 188 (2013) 218–225
(Scordalakes and Rissman, 2004). This discrepancy could be due tothe fact that adult treatment with estradiol may restore only par-tially aggressive behavior in wild-type mice. Therefore, differencesin aggressive behavior between wild-type and Tfm mice as re-cently reported (Robinson et al., 2012) or between control andARNesCre males as shown in the present study could be observableonly after treatment with androgenic agonists (Robinson et al.,2012) or supplementation with testosterone, which activates boththe androgenic and estrogenic pathways.
We also found that the low level of motivation of ARNesCre maleshad little to do with their olfactory preference. All mutant malesinitiated anogenital chemoinvestigation towards intruder malesin resident-intruder tests. Moreover, when tested in olfactory pref-erence tests, mutant males behaved as their control littermatessince sexual experience reinforced their olfactory preferences forfemale chemosignals. These findings conflict with studies on Tfmmice in the same C57BL/6J background showing that they exhibitfemale-typical preference (Bodo and Rissman, 2007). A possible
explanation of this discrepancy is the difference in circulating tes-tosterone concentrations and, thus, in the amount of estradiol de-rived from the neural aromatization of testosterone that thedeveloping brain is exposed to in these models. ARNesCre maleshave high levels of circulating testosterone (Raskin et al., 2009,2012). By contrast, the testicular feminization mutation in mice re-duces androgen levels. If this decline occurs during the criticalperinatal or pubertal periods of brain sensitivity to testosterone,it could interfere with the estradiol-induced sexual differentiationof the olfactory system. Studies in mice with knockouts of ERa haveclearly demonstrated the masculinizing effect of estradiol and itsreceptor on olfactory and partner preferences (Wersinger and Riss-man, 2000).
The normal chemoinvestigatory behavior of ARNesCre males indi-cates that AR is required in neuronal regions located downstreamfrom the olfactory bulb (MeA, BNST, lateral septum), to triggeraggressive behavior. However, the behavioral alteration inducedby ARNesCre mutation was not associated with a deficient organiza-
Fig. 4. Galanin in situ hybridization and immunoreactivity. (A) and (C) In situ hybridization (A) and immunoreactivity for galanin (C) on coronal sections of the BNST andlateral septum. (B) and (D) Total cell count in the BNST (B) and mean density of galanin immunostaining in the lateral septum (D). Data are expressed as means ± S.E.M (n = 4males per genotype). Scale bar = 30 lm.
C. Marie-Luce et al. / General and Comparative Endocrinology 188 (2013) 218–225 223
tion of AVP cell number in the BNST and AVP fiber density in thelateral septum. First, quantification of preprovasopressin mRNAscells by in situ hybridization showed no alteration in the BNST ofmutant males. Second, densitometry of vasopressin-neurophysin2 immunoreactivity in the lateral septum was unchanged in mu-tant males. The lateral septum receives projections from both theBNST and the MeA. The unchanged total density of AVP-immuno-reactive fibers in the lateral septum suggests no alteration in theMeA, although this needs to be further investigated.
Furthermore, ARNesCre mutation did not alter the number ofpreprogalanin expressing cells in the BNST and galanin immunore-activity in the lateral septum in male mice. The role of galanin inthe expression of inter-male aggression still needs to be defined.Nevertheless, this neuropeptide has been implicated in a widerange of behaviors such as cognition, memory, feeding, pain andmood regulation (reviewed by Rotzinger et al. (2010)). In particu-lar, galanin seems to modulate stress responses in the BNST andlateral septum (Khoshbouei et al., 2002). Previous studies showedthat galanin expressing cells are higher in the BNST and lower in
the arcuate nucleus in male than in female mice (Rajendrenet al., 2000). Furthermore, it was also reported that the expressionof this neuropeptide in the BNST is under the control of testoster-one in rat and hamster (Bolborea et al., 2010; Miller et al., 1993).
In the ARNesCre mouse model, AR exon 2 in the nervous system isexcised by embryonic day 10.5 (Tronche et al., 1999), before gona-dal and brain differentiations. Therefore, it seems that AR in thenervous system does not mediate testosterone-induced sexualdimorphism in AVP and galanin cell number in the BNST and fiberdensity in the lateral septum in male mice. This strongly suggeststhat the estrogenic rather than the androgenic signaling pathway iscrucial for the neuroanatomical differentiation of the BNST by tes-tosterone in male mice. Consistent with these observations, recentdata showed that treatment of newborn C57BL/6J females withestradiol benzoate fully masculinized BNST volume and cell num-ber as for testosterone, whereas 5-a-dihydrotestosterone, the non-aromatizable metabolite of testosterone, had no effect (Hisasueet al., 2010). Treatment of newborn females with selective estrogenreceptor alpha or beta agonists revealed that both receptors might
224 C. Marie-Luce et al. / General and Comparative Endocrinology 188 (2013) 218–225
be required in estradiol-induced effects (Hisasue et al., 2010). Fur-thermore, analysis of the ontogeny of AR expression in the devel-oping mouse brain showed that AR is lately expressed in theBNST, preoptic area and medial amygdala, suggesting that it mightnot play a major role in the prenatal and neonatal masculinizationof male-typical behaviors (Juntti et al., 2010).
It is possible that the ARNesCre mutation triggered activationalmodifications at the level of vasopressin receptors and/or down-stream components, thereby interfering with AVP signaling andconsequently with the expression of aggressive behavior. Two cen-trally expressed V1a and V1b receptors can mediate the effects ofAVP on aggressive behavior. Pharmacological studies in rodentsand genetic invalidation of genes encoding these receptors in miceclearly showed their involvement in the regulation of aggressivebehavior and social recognition (Bielsky et al., 2004; Ferris and Pot-egal, 1988; Wersinger et al., 2002, 2007). Furthermore, V1 recep-tors have been shown to be modulated by androgens (Delvilleet al., 1996; Young et al., 2000). Alternatively, ARNesCre mutationcould indirectly reduce aggressive behavior by affecting the estro-genic signaling pathway. Our previous studies showed no changesin the number of ERa-immunoreactive cells in the BNST of mutantmales (Raskin et al., 2009). Future studies will investigate whetherARNesCre mutation reduces neural aromatase expression (Yamadaet al., 1993).
In conclusion, the data presented here provide evidence that theAR signaling pathway in the nervous system is essential for theexpression of aggressive behavior but not for the sexual dimor-phism in cell number in the BNST and fiber density in the lateralseptum for AVP and galanin. In addition, AR signaling pathwaydoes not seem to be required for the masculinization and/or activa-tion of olfactory/partner preference. It is possible that the neuralAR is involved in the perinatal organization of neuroanatomicalprocesses others than those assessed in the present paper. Alterna-tively, AR could rather mediate the activation by testosterone ofbrain areas underlying the expression of aggressive behavior dur-ing adulthood. In this context, the conditional genetic model stud-ied here provides a unique opportunity to explore the contributionof AR in testosterone-induced regulation of male behaviors.
Acknowledgments
We thank Prof. Guido Verhoeven (Laboratory of ExperimentalMedicine and Endocrinology, Katholieke Universiteit Leuven, Leu-ven, Belgium) for providing the floxed AR mouse line. We thankthe UPMC platform for taking care of the animals.
This work was supported by the ‘‘Contaminants-Ecosystèmes-Santé’’ program of the ‘‘Agence Nationale de la Recherche’’,‘‘Agence Nationale de Sécurité Sanitaire’’, the ‘‘Réseau Santé Envi-ronnement Toxicologie’’ of the ‘‘Région Ile de France’’, and the Mar-ie Curie PhD-Program (MEST-CT-2005-021014).
References
Bielsky, I.F., Hu, S.-B., Szegda, K.L., Westphal, H., Young, L.J., 2004. Profoundimpairment in social recognition and reduction in anxiety-like behavior invasopressin V1a receptor knockout mice. Neuropsychopharmacology 29, 483–493.
Bodo, C., Rissman, E.F., 2007. Androgen receptor is essential for sexualdifferentiation of responses to olfactory cues in mice. Eur. J. Neurosci. 25,2182–2190.
Bolborea, M., Ansel, L., Weinert, D., Steinlechner, S., Pévet, P., Klosen, P., 2010. Thebed nucleus of the stria terminalis in the Syrian hamster (Mesocricetus auratus):absence of vasopressin expression in standard and wild-derived hamsters andgalanin regulation by seasonal changes in circulating sex steroids. Neuroscience165, 819–830.
Brot, M.D., De Vries, G.J., Dorsa, D.M., 1993. Local implants of testosteronemetabolites regulate vasopressin mRNA in sexually dimorphic nuclei of therat brain. Peptides 14, 933–940.
Burns-Cusato, M., Scordalakes, E.M., Rissman, E.F., 2004. Of mice and missing data:what we know (and need to learn) about male sexual behavior. Physiol. Behav.83, 217–232.
De Vries, G.J., Al-Shamma, H.A., 1990. Sex differences in hormonal responses ofvasopressin pathways in the rat brain. J. Neurobiol. 21, 686–693.
De Vries, G.J., Wang, Z., Bullock, N.A., Numan, S., 1994. Sex differences in the effectsof testosterone and its metabolites on vasopressin messenger RNA levels in thebed nucleus of the stria terminalis of rats. J. Neurosci. 14, 1789–1794.
Delville, Y., Mansour, K.M., Ferris, C.F., 1996. Testosterone facilitates aggression bymodulating vasopressin receptors in the hypothalamus. Physiol. Behav. 60, 25–29.
Dominguez-Salazar, E., Bateman, H.L., Rissman, E.F., 2004. Background matters: theeffects of estrogen receptor alpha gene disruption on male sexual behavior aremodified by background strain. Horm. Behav. 46, 482–490.
Ferris, C.F., Potegal, M., 1988. Vasopressin receptor blockade in the anteriorhypothalamus suppresses aggression in hamsters. Physiol. Behav. 44,235–239.
Han, T.M., De Vries, G.J., 2003. Organizational effects of testosterone, estradiol, anddihydrotestosterone on vasopressin mRNA expression in the bed nucleus of thestria terminalis. J. Neurobiol. 54, 502–510.
Hisasue, S., Seney, M.L., Immerman, E., Forger, N.G., 2010. Control of cell number inthe bed nucleus of the stria terminalis of mice: role of testosterone metabolitesand estrogen receptor subtypes. J. Sex. Med. 7, 1401–1409.
Juntti, S.A., Tollkuhn, J., Wu, M.V., Fraser, E.J., Soderborg, T., Tan, S., et al., 2010. Theandrogen receptor governs the execution, but not programming, of male sexualand territorial behaviors. Neuron 66, 260–272.
Khoshbouei, H., Cecchi, M., Morilak, D.A., 2002. Modulatory effects of galanin in thelateral bed nucleus of the stria terminalis on behavioral and neuroendocrineresponses to acute stress. Neuropsychopharmacology 27, 25–34.
Klosen, P., Maessen, X., van den Bosch, P., de Aguilar,, 1993. PEG embedding forimmunocytochemistry: application to the analysis of immunoreactivity lossduring histological processing. J. Histochem. Cytochem. 41, 455–463.
Le Roy, I., Pothion, S., Mortaud, S., Chabert, C., Nicolas, L., Cherfouh, A., et al., 2000.Loss of aggression, after transfer onto a C57BL/6J background, in mice carrying atargeted disruption of the neuronal nitric oxide synthase gene. Behav. Genet.30, 367–373.
Miller, M.A., Kolb, P.E., Raskind, M.A., 1993. Testosterone regulates galanin geneexpression in the bed nucleus of the stria terminalis. Brain Res. 611, 338–341.
Ogawa, S., Lubahn, D.B., Korach, K.S., Pfaff, D.W., 1997. Behavioral effects of estrogenreceptor gene disruption in male mice. Proc. Natl. Acad. Sci. USA 94, 1476–1481.
Ohno, S., Geller, L.N., Young Lai, E.V., 1974. Tfm mutation and masculinizationversus feminization of the mouse central nervous system. Cell 3, 235–242.
Pierman, S., Sica, M., Allieri, F., Viglietti-Panzica, C., Panzica, G.C., Bakker, J., 2008.Activational effects of estradiol and dihydrotestosterone on social recognitionand the arginine-vasopressin immunoreactive system in male mice lacking afunctional aromatase gene. Horm. Behav. 54, 98–106.
Plumari, L., Viglietti-Panzica, C., Allieri, F., Honda, S., Harada, N., Absil, P., et al., 2002.Changes in the arginine-vasopressin immunoreactive systems in male micelacking a functional aromatase gene. J. Neuroendocrinol. 14, 971–978.
Rajendren, G., Levenkova, N., Gibson, M.J., 2000. Galanin immunoreactivity inmouse basal forebrain: sex differences and discrete projections of galanin-containing cells beyond the blood-brain barrier. Neuroendocrinology 71, 27–33.
Raskin, K., de Gendt, K., Duittoz, A., Liere, P., Verhoeven, G., Tronche, F., et al., 2009.Conditional inactivation of androgen receptor gene in the nervous system:effects on male behavioral and neuroendocrine responses. J. Neurosci. 29,4461–4470.
Raskin, K., Marie-Luce, C., Picot, M., Bernard, V., Mailly, P., Hardin-Pouzet, H., et al.,2012. Characterization of the spinal nucleus of the bulbocavernosusneuromuscular system in male mice lacking androgen receptor in the nervoussystem. Endocrinology 153, 3376–3385.
Robinson, S., Penatti, C.A.A., Clark, A.S., 2012. The role of the androgen receptor inanabolic androgenic steroid-induced aggressive behavior in C57BL/6J and Tfmmice. Horm. Behav. 61, 67–75.
Rotzinger, S., Lovejoy, D.A., Tan, L.A., 2010. Behavioral effects of neuropeptides inrodent models of depression and anxiety. Peptides 31, 736–756.
Sato, T., Matsumoto, T., Kawano, H., Watanabe, T., Uematsu, Y., Sekine, K., et al.,2004. Brain masculinization requires androgen receptor function. Proc. Natl.Acad. Sci. USA 101, 1673–1678.
Scordalakes, E.M., Rissman, E.F., 2004. Aggression and arginine vasopressinimmunoreactivity regulation by androgen receptor and estrogen receptoralpha. Genes Brain Behav. 3, 20–26.
Tronche, F., Kellendonk, C., Kretz, O., Gass, P., Anlag, K., Orban, P.C., et al., 1999.Disruption of the glucocorticoid receptor gene in the nervous system results inreduced anxiety. Nat. Genet. 23, 99–103.
van Leeuwen, F.W., Caffe, A.R., De Vries, G.J., 1985. Vasopressin cells in the bednucleus of the stria terminalis of the rat: sex differences and the influence ofandrogens. Brain Res. 325, 391–394.
Wang, Z., Bullock, N.A., De Vries, G.J., 1993. Sexual differentiation of vasopressinprojections of the bed nucleus of the stria terminals and medial amygdaloidnucleus in rats. Endocrinology 132, 2299–2306.
Wersinger, S.R., Rissman, E.F., 2000. Oestrogen receptor alpha is essential forfemale-directed chemo-investigatory behaviour but is not required for thepheromone-induced luteinizing hormone surge in male mice. J.Neuroendocrinol. 12, 103–110.
C. Marie-Luce et al. / General and Comparative Endocrinology 188 (2013) 218–225 225
Wersinger, S.R., Ginns, E.I., O’Carroll, A.-M., Lolait, S.J., Young 3rd, W.S., 2002.Vasopressin V1b receptor knockout reduces aggressive behavior in male mice.Mol. Psychiatry 7, 975–984.
Wersinger, S.R., Caldwell, H.K., Christiansen, M., Young 3rd, W.S., 2007. Disruptionof the vasopressin 1b receptor gene impairs the attack component of aggressivebehavior in mice. Genes Brain Behav. 6, 653–660.
Yamada, K., Harada, N., Tamaru, M., Takagi, Y., 1993. Effects of changes in gonadalhormones on the amount of aromatase messenger RNA in mouse braindiencephalon. Biochem. Biophys. Res. Commun. 195, 462–468.
Young, L.J., Wang, Z., Cooper, T.T., Albers, H.E., 2000. Vasopressin (V1a) receptorbinding, mRNA expression and transcriptional regulation by androgen in theSyrian hamster brain. J. Neuroendocrinol. 12, 1179–1185.