Ann. appl. Biol. (1966), 58, 145-157 Printed in Great Britain ‘45 Effects of temperature and nitrogen supply on the growth of timothy (Phleum prutense L.) BY DALE SMITH* AND 0. R. JEWISS The Grassland Research Institute, Hurley, Berkshire (Received 26 October 1965) SUMMARY Timothy was grown in controlled environments with day/night tempera- tures of 18.5/10° C. and 29*5/21O C., with and without extra nitrogen. High temperatures hastened all stages of development after inflorescence initia- tion. Anthesis occurred after 48 days when N was added and after 52 days without N. Anthesis did not occur until after 68 days at the lower tempera- tures, with or without N. Largest yields of leaves, stems and stubble were produced at all growth stages at the lower temperatures with N. This treatment gave the highest crop growth rate, which increased almost linearly with time. The lower temperatures with N gave many more and longer stems and more inflorescences than any other treatment. With N and high temperatures, there was no increase in crop growth rate after inflorescence emergence. Temperature had little effect on growth when no N was provided. With no added N, crop growth rate was retarded and increased very little after inflorescence initiation. Herbage produced at high temperatures had the lower percentages of water-soluble carbohydrates and digestible dry matter, but had the higher percentages of most minerals. N increased the percentage content of most minerals. Weights of stubble and of water-soluble carbohydrates in the stubble, and the numbers of stem bases and tillers, were larger at the lower temperatures, and were usually larger with added N in both temperature regimes. Nitrogen decreased the percentage content of carbohydrates in the stubble. Large haplocorms were produced at the low temperatures, but none developed at the high temperatures, indicating less carbohydrate storage. The results partly explain the vigorous growth of timothy in the cool, temperate regions of Europe and North America, and the small dry matter production, lower nutritional value of the herbage, and lack of persistence in the warm southern regions where rainfall is adequate. INTRODUCTION Timothy (Phleum pratense L.) is grown widely for forage in the temperate regions of Europe and North America. It appears to grow best in cool-temperature areas where rainfall is adequate and well distributed during the growing season. Little is known about the effect of high temperatures on the growth responses or the chemical U.S.A. * Permanent address : Department of Agronomy, University of Wisconsin, Madison, Wisconsin, I0 App. Did. 58
Transcript
Ann. appl. Biol. (1966), 58, 145-157 Printed in Great Britain
‘45
Effects of temperature and nitrogen supply on the growth of timothy (Phleum prutense L.)
BY DALE SMITH* AND 0. R. JEWISS The Grassland Research Institute, Hurley, Berkshire
(Received 26 October 1965)
SUMMARY
Timothy was grown in controlled environments with day/night tempera- tures of 18.5/10° C. and 29*5/21O C., with and without extra nitrogen. High temperatures hastened all stages of development after inflorescence initia- tion. Anthesis occurred after 48 days when N was added and after 52 days without N. Anthesis did not occur until after 68 days at the lower tempera- tures, with or without N.
Largest yields of leaves, stems and stubble were produced at all growth stages at the lower temperatures with N. This treatment gave the highest crop growth rate, which increased almost linearly with time. The lower temperatures with N gave many more and longer stems and more inflorescences than any other treatment. With N and high temperatures, there was no increase in crop growth rate after inflorescence emergence. Temperature had little effect on growth when no N was provided. With no added N, crop growth rate was retarded and increased very little after inflorescence initiation.
Herbage produced at high temperatures had the lower percentages of water-soluble carbohydrates and digestible dry matter, but had the higher percentages of most minerals. N increased the percentage content of most minerals.
Weights of stubble and of water-soluble carbohydrates in the stubble, and the numbers of stem bases and tillers, were larger at the lower temperatures, and were usually larger with added N in both temperature regimes. Nitrogen decreased the percentage content of carbohydrates in the stubble. Large haplocorms were produced at the low temperatures, but none developed at the high temperatures, indicating less carbohydrate storage.
The results partly explain the vigorous growth of timothy in the cool, temperate regions of Europe and North America, and the small dry matter production, lower nutritional value of the herbage, and lack of persistence in the warm southern regions where rainfall is adequate.
I N T R O D U C T I O N
Timothy (Phleum pratense L.) is grown widely for forage in the temperate regions of Europe and North America. It appears to grow best in cool-temperature areas where rainfall is adequate and well distributed during the growing season. Little is known about the effect of high temperatures on the growth responses or the chemical
U.S.A. * Permanent address : Department of Agronomy, University of Wisconsin, Madison, Wisconsin,
I 0 App. D i d . 58
146 DALE SMITH AND 0. R. JEWISS
composition of the herbage of timothy and other temperate-adapted perennial forages. Also, little information is available on the effect of high temperatures on the storage of carbohydrate reserves, but if the accumulation of these reserves is restricted, regrowth after cutting may be retarded, especially if stress conditions prevail.
Mitchell (1956), using constant temperatures, showed that the fastest rate of shoot growth per day of perennial rye-grass (Lolium perenne L.), cocksfoot (Dactylis glomerata L.), Yorkshire fog (Holcus lanatus L.) and subterranean clover (Trifolium subterraneum L.) occurred between 18 and 21" C., of white clover ( T . repens L.) and big trefoil (Lotus uliginosus Schkuhr.) at 24" C., of the tropical species Paspalum dilatatum Poir. at 29" C. Growth was slower at temperatures above these optima, particularly for the temperate-adapted species. Nearly the same optimal temperatures have been found for regrowth after cutting. Harrison (1934) showed that bluegrass (Poa prutensis L.) grew best at 16" C., but little regrowth occurred at 38" C. and the plants died in about 4 weeks. Sullivan & Sprague (1949) showed that new topgrowth of Lolium perenne L. was rapid at 21" C. day and 16" C. night temperature and the most carbohydrates were stored at the lowest temperatures used (16/10" C.). After 40 days at 32/27" C. plants were stunted, with discoloured roots, apparently near death, and carbohydrates in the roots and stubble were almost exhausted.
Brown (1939) similarly showed that the optimal temperatures for herbage growth was 21" C. for Dactylis glomerata L., between 27 and 32' C. for Poa pratensis L. and P. compressa L., and 38" C., the highest temperature used, for Cynodon dactylon (L.) Pers. Root and rhizome growth was best at lower temperatures: 10" C. for Poa compressa, 16" C . for Poa pratensis, and 38" C . for Cynodon dactylon.
Kendall (1958) found maximal growth of red clover (Trifolium pratense L.) at 24/18" C, the same range as the legumes noted above. At the highest temperatures (35/27" C.) tested by Kendall, 90% of the plants died after cutting, and regrowth was poor, apparently because not enough carbohydrate was stored.
This paper deals with the growth of timothy in controlled environment chambers at cool and warm temperatures with two rates of soil N.
MATERIALS A N D M E T H O D S
Circular samples of S48 timothy turf of about 17 cm. in diameter were cut between 18 and 29 March 1964 from a sward of uniform plant density sown during 1963 at Hurley. The turves, trimmed to leave 6.5 cm. depth of soil, were placed in metal cans, 17 cm. in diameter and 23 cm. deep and with a I cm. central drainage hole in the bottom, partly filled with sandy-loam soil from the same site as the turf. The top of each turf was 2 cm. below the rim of the can.
One-hundred cans were randomly selected on 8 April; 4 were used for the initial sample and 48 were placed in each of two growth chambers at two different tempera- ture regimes: ( I ) warm--29*5" C. in the day and 21" C. at night, and (2) cool-18.5" C. in the day and 10" C. at night. Both chambers had an 18 hr. photoperiod of 2300 ft.- candles (6-8 g. cal./cm.2/hr.) at can height from warm-white fluorescent tubes. Temperature was lowered with the change from the light to the dark period.
Half the cans in each chamber received 56 kg. of N per ha. as NH,NO, in solution
Temperature and nitrogen eflects on timothy "7 when placed in the chamber and every time they were sampled except the last. A total of 336 kg. of N per ha. was applied. The other cans received no nitrogen. Treatment combinations were designated as CN (cool temperatures with added N), C (cool temperatures with no added N), WN (warm temperatures plus N), and W (warm temperatures without N).
Aluminium shields were wired around the outside of the cans to simulate lateral shading in mass populations. These were moved upward at short intervals so that their upper edges were within 18 cm. of the ends of the top leaves. This continued until the shields were 19 cm. above the top rim of the cans, when no further extension was possible.
Four cans of plants (A) were sampled on the day before the remaining ones were placed in the growth chambers. Thereafter, plants were sampled from each tempera- ture-N combination at specific growth stages, rather than on a time basis, to simulate stages at which they might be harvested in the field. Plants were sampled at (B) in- florescence initiation, (C) beginning of stem elongation, (D) inflorescence emergence, and (E) anthesis. Initiation of inflorescences was determined every 3-4 days by apical dissection of tillers randomly selected from spare cans in each treatment. At anthesis, remaining plants were cut to leave a 4.5 cm. stubble, and were sampled after 14 days (F) or 28 days regrowth (G).
Each can was sampled for herbage yield. For the initial sample, the small leaves dose to the soil surface were cut at the Iigules, leaving the remainder as stubble. Sub- sequent samples were clipped to leave a 4.5 cm. stubble and the clippings (herbage yield) were separated into leaf blades and stems, leaving the leaf sheaths and in- florescences attached to the stems. Leaf blades were separated into yellow and green tissue. Each fraction was dried separately. Total herbage yields were the sum of the leaf and stem weights. Classical growth analysis was followed. The changes in leaf area were similar to increases in leaf dry weight and the results are not presented; the results of many of the derived parameters were too variable for sound conclusions and are also omitted.
The stubble (4.5 cm.) was removed from the cans by cutting tillers from their roots just below the soil surface. Tillers were cleaned of dead tissue and roots, washed and dried. Total topgrowth yields were the sum of the herbage and stubble weights.
All tissues were dried for 20 min. at 100" C. and then at 75" C. in forced-draught ovens, The dried tissue was ground to pass a 40-mesh sieve. Total nitrogen was estimated by the method of Horwitz (1960). Water-soluble carbohydrates (WS-CHO) were extracted with cold water and determined colorimetrically with anthrone reagent by the method of Deriaz (1961).
Herbage from replicates was bulked by treatments. Total nitrogen and water- soluble carbohydrates were determined by the same methods used for the stubble. The samples were also analysed for digestible dry matter (DDM) by the method of Tilley & Terry (1963), for crude fibre by the method of Horwitz (1960), for total ash after 16 hr. ashing at 550" C, and for Na, K, P, Ca, and Mg photospectrometrically.
10-2
148 DALE SMITH AND 0. R. JEWISS
R E S U L T S Time to growth stages
The time to inflorescence initiation (stage B) was the same for all treatments. Plants in the high temperatures subsequently developed more rapidly (Table I ) but the influence of N was small. Anthesis (stage E) occurred at the high temperatures after 48 days with N and after 52 days without N, and at the low temperatures after 68 days with or without N.
24
. M
.6, 16 3 e M 12 w 8
i"
0- A B C D E F G
h x
Y E
50 V . 5 40 X ab 30 *
L
5 3 20 e M
g 10 U
0
L 1-ZN
j-1 '\, ,' ,/ 'tf"
A-B B-C C-D D-E I I
A-B B-C C-D D-E - E-F F-G
Stages of growth Stages of growth
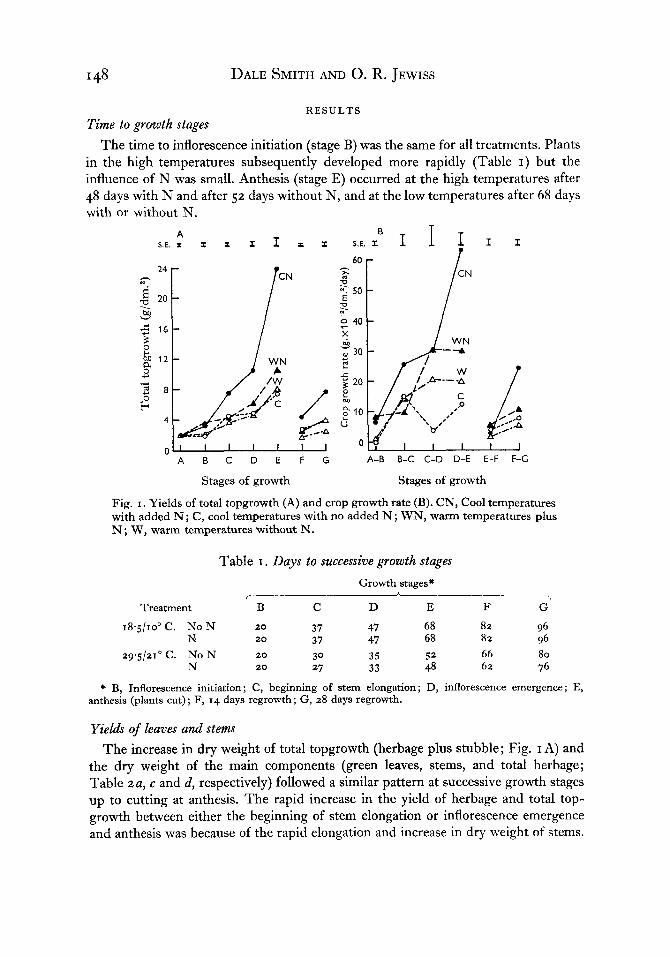
Fig. I . Yields of total topgrowth (A) and crop growth rate (B). CN, Cool temperatures with added N ; C, cool temperatures with no added N ; WN, warm temperatures plus N; W, warm temperatures without N.
Table I . Days to successive growth stages Growth stages*
r 7 - I _ _ ~
Treatment B C D E F G
18*5/1o" C. No N 20 37 47 68 82 96 N 20 37 47 68 82 96
29.5/21° C. No N 20 30 35 52 66 80 N 20 27 33 48 62 76
* B, Inflorescence initiation ; C, beginning of stem elongation; D, inflorescence emergence ; E, anthesis (plants cut); F, 14 days regrowth; G, 28 days regrowth.
Yields of leaves and stems The increase in dry weight of total topgrowth (herbage plus stubble; Fig. I A) and
the dry weight of the main components (green leaves, stems, and total herbage; Table 2 a , c and d, respectively) followed a similar pattern at successive growth stages up to cutting at anthesis. The rapid increase in the yield of herbage and total top- growth between either the beginning of stem elongation or inflorescence emergence and anthesis was because of the rapid elongation and increase in dry weight of stems.
Temperature and nitrogen effects on timothy I49 Nitrogen increased yield and the largest yield of all these components at anthesis was obtained with CN. Yields under the other environmental conditions were in the decreasing order of WN, W and C. The yield of yellow leaves (Table z b ) was small and was not much affected by the treatments except at anthesis (stage E), when the lower leaves were considerably shaded, especially with CN.
The dry weight of green leaves 14 and 28 days after cutting was greatest under CN and then decreased in the order of WN, C, and W. No stems were produced during the 28-day period of regrowth. The yields of total herbage and of total topgrowth were generally in the same order of rank as those for green leaves.
Table 2. Dry weight of leaves, stems and total herbage above cutting height (g . x I O - ~ per dm.2)
Growth stages
Treatment A B
18.5/10°C. No N 57 106 (a) Green leaves
N 57 I97
N 57 245
18.5/1o"C. No N 13 9 N 13 7
29.5/2I0C. N o N 13 4 N I3 9
18.5/10°C. No N - -
29.5/21°C. N o N - -
29.5121" C. NO N 57 143
S.E. 4 I9 (6 ) Yellow leaves
S.E. I I
(c) Stems
- - N
N - - - - S.E.
(d ) Total herbage above cutting height" r8.5/ro0 C. No N 69 115
N 69 204 29'5/21°C. N o N 69 147
N 69 254 S.E. 4 19
C
196 339 200
246 '5
19 15 9
I 2 2
65 162 53 68 12
279 5'6 263 326 20
D
I95 414 229 317 14
23 34
14 3
77 305
I 2
I I 0
184 29
295 753 352 515 41
I.:
214 527 259 351 33
88 228
96 I00
19
226 I243 300 454 75
528 I998 658
89 901
* Values at stages F and G include pseudostems and elongated leaf sheaths in addition to green leaves.
Crop growth rate Crop growth rate, estimated as (W2- W,) / ( t2 - t l ) , where W, and W, were dry
weights of the total topgrowth at times t , and t,, was largest with CN and increased nearly linearly until anthesis (Fig. I B). Under the other conditions, crop growth rate increased in the order of WN, W and C, but after the initial increase it changed very little at the more mature stages.
1 50 DALE SMITH AND 0. R. JEWISS
Number of stem bases and tillers The total number of stem bases and tillers per unit area decreased at first (Table 3 a),
but then increased after inflorescence initiation or the beginning of stem elongation. Langer, Ryle & Jewiss (1964) also found similar changes in simulated field conditions. The largest increases generally occurred between inflorescence emergence and anthesis (stages D and E) and during the first 14 days after cutting at anthesis (stages E and F). Little or no increase occurred during the second 14 days of regrowth. The largest numbers always occurred under CN. At anthesis and thereafter, the number of stem bases and tillers and the weight of stubble (Table 3 b) were in the decreasing order of CN, C, WN and W.
Table 3. Number and dry weight of stem bases and tillers (stubble) per dm.2
Growth stages
Treatment A B C D E F G
(a) Number 185/1o" C. No N
N
N 29'5/21° C. NO N
S.E.
(b) Dry weight (g. x 10' 18-5/10" C. No N
N
N 29*5/21° C. NO N
S.E.
48 48 48 48
-a per dma) I18 118 118 118
I
I 0
37 33 36 38 43 54 32 36 39 38 37 38 2 3 2
70 163 I 86 109 23 5 301 65 96 I11
I 06 104 I02 5 I1 I2
51 75 39 5 0
5
213 3 87 I45 168 15
78 79 105 105 63 66 66 78 6 4
222 263 310 403 121 142 167 190 I1 I2
Table 4. Stem measurements at anthesis (stage E ) Stem bases
and Elongated Elongated Height Inflorescences Treatment tillers/dm.a ~ tems/dm.~ stems* of stems /dm.8
(no. 1 (no.) ( %) (cm.1 (no.)
18.5/10" C. No N 51 23 70 83 1'5
29'5/21" C. NO N 39 25 69 94 2'2
29 78 I22 6.7
+ Percentages based on number of tillers present at beginning of stem elongation (stage C).
N 75 44 I02 1.51 13'7
5 1'7 - N 5 0 S.E. 5 3
Stem numbers and lengths at anthesis The effect of treatments on the number and height of stems longer than 4.5 cm.
and on the number of inflorescences (Table 4) was like that for the yield of total herbage (Table z d ) and of total topgrowth (Fig. IA). They were in the decreasing order of treatments CN, WN, W and C.
The percentage of stems longer than the cutting height at anthesis was calculated
Temperature and nitrogen eflects on timothy 1.51
from the number of tillers present at the beginning of stem elongation (stage C), because new tillers were continually being produced between these two growth stages. Many did not elongate, or did not begin to do so until after cutting at anthesis.
Yield of stubble Stubble dry weight decreased between growth stages A and B (Table 3 b), probably
because carbohydrates in the stubble were used for new growth. The decrease was greater when no N was given. After inflorescence initiation (stage B), stubble weight increased steadily at the low temperatures until the herbage was cut at anthesis (stage E), and the increase was greater when N was added. Between these stages, N also caused a larger increase in tiller numbers (Table 3 u). Only a small increase in stubble weight occurred under the high temperatures between inflorescence initiation and anthesis, and the increase was greater with added N.
3h 4 A B C D E F G
Stages of growth A B C D E F G
Stages of growth
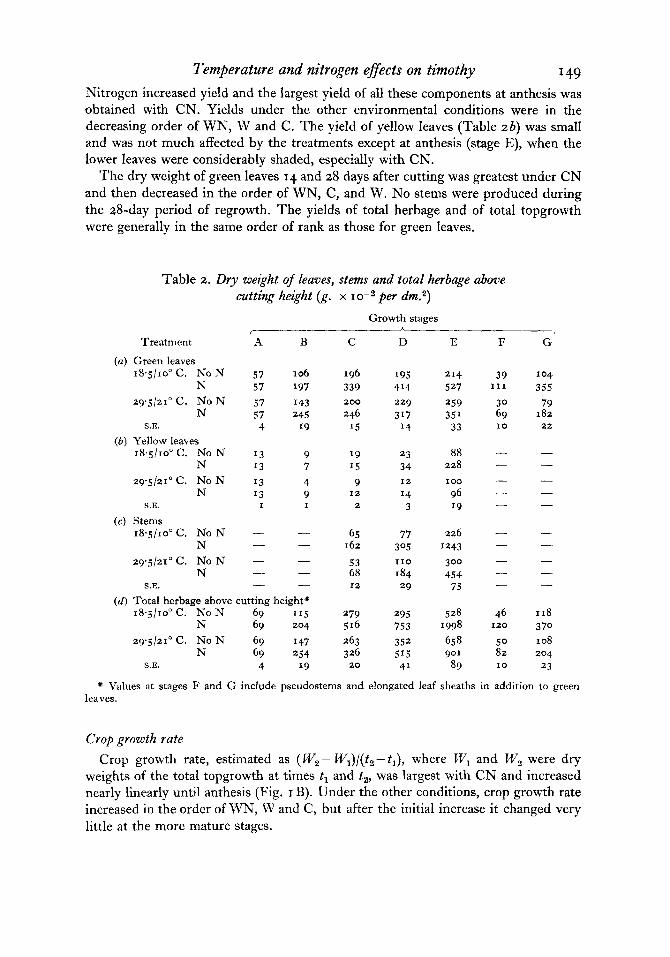
Fig. 2. The percentage of water-soluble carbohydrates (A) and the total amount of water-soluble carbohydrates (B) in the stubble. Symbols as in Fig. I .
After cutting at anthesis, stubble weight generally decreased during the first 14 days of regrowth (stages E and F), but tiller numbers increased faster than at any other time and green leaves were few. Stubble weight again increased during the second 14 days of regrowth.
Water-soluble carbohydrates in stubble The percentage of water-soluble carbohydrates (WS-CHO) in the stubble was
much decreased by the high temperatures and usually to a smaller extent by adding N (Fig. 2A).
Percentages decreased rapidly in all treatments to a minimum at inflorescence initiation (stage B); the reduction was greater under high temperatures. No doubt carbohydrates in the stubble were used to produce at least the first new roots and topgrowth and account in part for the decrease in stubble weight at this time (Table 3 b).
n
cn
N
Tab
le 5
. Che
mic
al co
mposition
of t
he h
erba
ge a
vera
ged
over
W
ater
-sol
. D
iges
tibl
e C
rude
T
otal
T
otal
C
HO
D
M
fibre
f-
N
ash
Na
Tre
atm
ent
(%)
( %)
(%)
(%I
(%I
(%I
18.5
/10"
C.
No
N
10.3
71
.9
23'3
I .6
6 8.
88
0'01
5 N
12'1
72.6
25.2
2.04
7.
62
0.0
18
29'5/21° C
. N
O N
6.
2 68
.5
25.1
2'24
9'
3 1
0.01
5
1.1
1
'0
01
4
04
3
00
04
N
5.2
707
24'5
3.
08
8.99
0'025
S.E.
0.7
stage
s of
gro
wth"
K
P (%
) ( %
) 2'22
0.29
I .6
4 0.
26
2.28
0.29
2'05
029
0'1
5
002
0.70
0.15
tl
0.98
0'21
m
0.08
0'01
P
*:
Ave
rage
of
anal
yses
mad
e at
gro
wth
sta
ges
B (i
nflo
resc
ence
ini
tiatio
n), C
(be
ginn
ing
of s
tem
elo
ngat
ion)
, D (i
nflo
resc
ence
emer
genc
e), E
(ant
hesi
s (pl
ants
t A
vera
ge o
f an
alys
es m
ade
at g
row
th s
tage
s C
, D a
nd E
. ?
cut)
), F
(1
4 da
ys r
egro
wth
), an
d G
(28
day
s re
grow
th).
;; u1
Temperature and nitrogen effects on timothy '53 A high percentage of WS-CHO was found in the stubble when stem elongation
began (stage C) under the low temperatures, and remained so except for a decrease after cutting at anthesis. Under the high temperatures, it increased slowly after ear initiation and reached a maximum at anthesis.
The total amount of WS-CHO per unit area of stubble (Fig. zB) changed with time similarly to the dry weight of stubble (Table 3 6 ) . The effects of temperature were relatively large because both percentage of WS-CHO and dry weight of stubble were decreased by the high temperatures.
92r A 24
56 0 A B C D E F G
Stages of growth
J l l l l l l A B C D E F G
Stages of growth
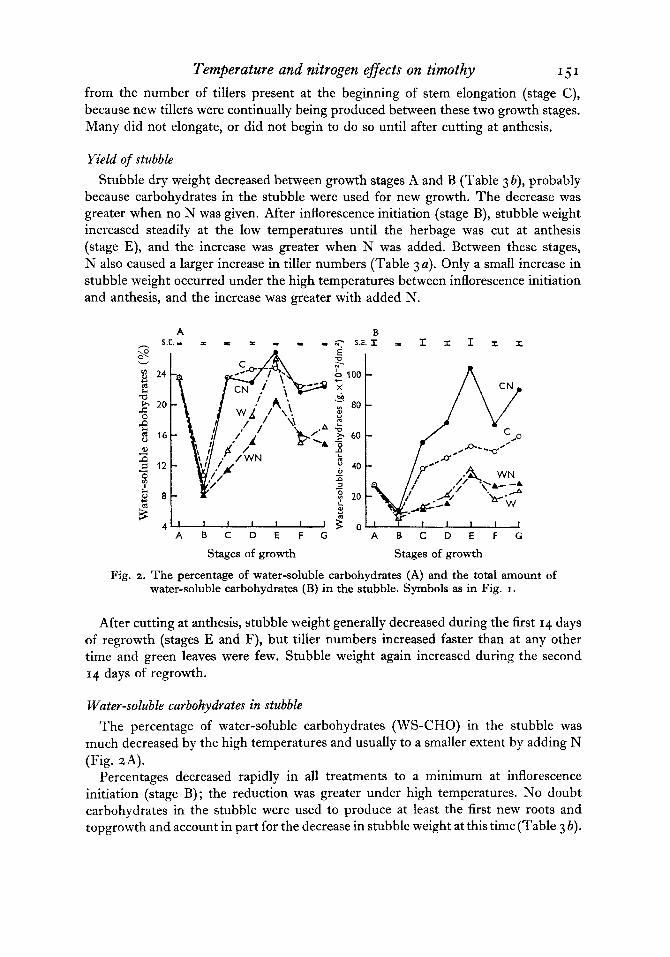
Fig. 3. The percentage of digestible dry matter (A) and the percentage of water- soluble carbohydrates (B) in the herbage above cutting height at each stage of growth. Symbols as in Fig. I.
Chemical composition of the herbage Herbage of plants grown at the high temperatures had less WS-CHO and digestible
dry matter (DDM) percent of dry matter, but more total N, total ash, Na, K, Ca and Mg than those grown at the lower temperatures (Table 5) . Temperatures had little effect on the percentages of crude fibre and P. Smaller percentages of mineral elements in the herbage produced under the lower temperatures may be attributable to dilution by the higher concentrations of WS-CHO.
Applied N affected primarily the mineral constituents of the herbage. Percentages of total N, Na, Ca and Mg were increased by added N, but those of total ash and K were decreased.
Changes in DDM and WS-CHO in the herbage with time The percentage of DDM in the herbage at each stage of growth (Fig. 3 A) decreased
steadily with time. The percentage of WS-CHO decreased rapidly to inflorescence initiation, but subsequently changed very little (Fig. 3 B).
Herbage grown at the lower temperatures had the highest percentages of both constituents at all growth stages, except that at anthesis there was more DDM in the herbage from the high temperatures. The low DDM values at anthesis under CN
154 DALE SMITH AND 0. R. JEWISS
could be associated with the large increase in weight of stems (Table 2c) and of yellowed lower leaves (Table 26) between stages D and E, but it also occurred under treatment C, where growth was retarded and the weight of yellowed leaves was least. The longer period of time between inflorescence emergence and anthesis at low temperatures may have resulted in a greater maturation of stem tissue.
Nitrogen increased the percentage of DDM in the herbage, except under the low temperatures at anthesis. It also increased the percentage of WS-CHO under the low temperatures, but decreased the percentages under the high temperatures.
D I S C U S S I O N
The largest yields of green leaves, stems and total top growth at successive stages of growth were obtained with treatment CN ; differences between treatments became apparent after the beginning of inflorescence initiation (Table 2 4 c; Fig. IA). The higher temperatures shortened the time intervals between successive stages of growth (Table I) , so that when the results were plotted on a time scale, over a common period of time the curves for WN and CN were coincident and lay above the points for treatments without N, and the curve for W was steeper than for C. When N was applied, the faster reproductive development at high temperatures was associated with a slower crop growth rate between successive stages of development and particularly between inflorescence emergence and anthesis. This shows that the high temperatures were above the optimum for growth, and this is confirmed by the much lower crop growth rate of treatment WN compared with CN during the final 14 days of regrowth after cutting at anthesis, and is supported by the various stem measurements at anthesis (Table 4).
Part of the reduction in crop growth rate in treatment WN before anthesis may be attributed to a two-stage effect on reproductive development, by a decrease, first in the number of elongated stems and secondly in the proportion of these that produced inflorescences. Similarly Ryle & Langer (1963a, 6) showed, with the same variety of timothy, that high temperatures increased the rate of inflorescence development but decreased the number of fertile tillers. They found that the magnitude of the latter effect diminished with increase in photoperiod and disappeared at shorter daylengths than in the present experiment. However, they worked at a lower temperature regime (23.5" C.) only on the response of the main stems which are influenced less than subsidiary tillers by changes in the environment, so it is reasonable to extend their findings to the present results. Similarly Ryle & Langer (1963 b) showed that increased temperatures decreased the size of inflorescences, and presumably the weight of the inflorescence was also decreased, further accounting for differences in dry weight at anthesis between treatments WN and CN. Certainly much of the high crop growth rate between inflorescence emergence and anthesis in treatment CN may be attributed to the large contribution to dry weight by the stem fraction (Fig. I B; Table 2c, d ) .
The effect of N on reproductive development can be explained in the same way as Some of the differences in yield and crop growth rate between treatments WN and CN. On swards of timothy grown in natural conditions closely similar to the low tempera- ture regime of this experiment, Lambert (1962) found effects of nitrogen on yield at
Temperature and nitrogen ejjfects on timothy I55 anthesis of the same magnitude. With high rates of N, about twice as many inflor- escences were present per unit area as with low rates (Lambert, private communica- tion), indicating that nitrogen had a large effect on inflorescence production. With treatment W the number of inflorescences (Table 4), and part of the total herbage and total topgrowth yields at anthesis (Table 2d; Fig. IA), were the result of effects of available N and high temperature on inflorescence development. As treatment W outyielded C, the relative availability of N with these two treatments was probably more important than effects of temperature on inflorescence production.
When treatments W and C are compared on a time-scale instead of a growth-stage basis, the results agree well with those of Beevers & Cooper (1964a), who showed that the dry matter production of Lolium multiflorum and L. perenne seedlings (apparently grown in soil without added N) was greater over a 10-week period at 25/25' C. than at 12/12' C. At warm temperatures soil N may be mineralized and become available to the plant more rapidly than at low temperatures. When N was applied in solution, as in the WN and CN treatments, effects of temperature on mineralization of N were of little significance. After cutting at anthesis, treatment C showed a higher dry-matter production and crop growth rate than W (Fig. IA, B). At this stage regrowth was undoubtedly influenced by the number of new tillers in the sward and the level of carbohydrate reserves in the stem bases of the old tillers; these were always greater in the low-temperature treatments (Table 3a; Fig. 2).
Chemical composition of the herbage was affected more by temperature than by N (Table 5) . Herbage produced at the higher temperatures usually had less WS-CHO and DDM, but more minerals, per cent of dry matter. Within each temperature regime, the percentage of most minerals was higher with added N. The WS-CHO in herbage produced at warm temperatures averaged 5 - 5 % less than at cool temperatures; this probably accounted for the generally lower content of DDM in the herbage pro- duced at warm temperatures, and, in the absence of dilution by carbohydrate, a larger concentration of minerals. The percentage of WS-CHO was usually less with added N as Beevers & Cooper (19646), Colby, Drake, Field & Kreowski (1965), and others found. The greatest percentage and weight of WS-CHO in the stubble was generally found at anthesis and the least at inflorescence initiation (Fig. 2), although, in field studies of another variety of timothy, Okajima & Smith (1964) found the least con- centration at the early stages of stem elongation. The weight of carbohydrates in the stubble is more important than the percentage content, because the weight indicates the total energy available to the plants during periods of stress, such as for regrowth after cutting. The amount of carbohydrate, tiller numbers and stubble weight were always greater in the lower-temperature treatment (Fig. 2; Table 3), which gave more total topgrowth at anthesis with each of the N regimes. Tiller numbers and stubble weights were usually higher with added N and gave more new growth and higher crop growth rates after cutting at anthesis.
The warmer temperatures were apparently above the optimum for vegetative growth, presumably because photosynthesis was decreased and respiration increased. Carbohydrates synthesized were largely used in growth or lost by respiration so that little was stored, and at anthesis the increase in stubble weight and the storage of carbohydrates were small, haplocorms were not evident, and the percentage of
156 DALE SMITH AND 0. R. JEWISS
WS-CHO in the herbage was small (Fig. 2 ; Tables 4, 5). Murata & Iyama (1963) studied the effect of temperature between o and 45' C. on Lolium perenne, L. multi- JEorum and Dactylis glomerata, and found that photosynthesis was faster around 10-15' C. and decreased slowly at lower temperatures and rapidly at temperatures above 35' C. On the other hand, respiration rate increased exponentially with increase in temperature throughout. Also Davidson & Milthorpe (1965) suggested that photo- synthetic capacity of the leaf surface of Dactylisglomerata decreased at high temperature (26" C.) as the plant aged. In the present experiment the effects of warm temperatures no doubt contributed to the small weight of total topgrowth and accounted for the low stubble weights and the absence of haplocorm formation.
Regrowth occurred at the warm temperatures after cutting at anthesis even though the amount of carbohydrates in the stubble was small and there were no green leaves to synthesize carbohydrates. Regrowth might not have been possible had the plants been cut at inflorescence emergence, when the amount of reserve carbohydrates was even less and there were fewer tillers per unit area than at anthesis. Had the experi- ment continued, the plants at the high temperatures probably would not have survived further cutting.
Evidently, timothy will grow most vigorously in the cool, temperate regions and will grow less vigorously, or may not persist, in more southerly areas, even though rain- fall and soil fertility may be adequate. However, temperatures may be unfavourable in temperate areas. During summer in the north-central and north-eastern areas of the United States, where timothy is widely grown, temperatures may be as high as 27-35' C. Little dry matter is produced during the warm summer period; most is produced during the cool spring and autumn periods. Soil moisture has generally been considered the major factor involved. The present results indicate that warm tempera- tures before cutting for hay could decrease storage of assimilates. This could be further aggravated by large soil N supply limiting the substrate available for regrowth after cutting. Warm temperatures immediately after cutting could dissipate the carbo- hydrates by high respiration loss to the point of exhaustion and death of the plant. This condition would be intensified if carbohydrate reserves were small at the time of cutting; thus Colby et al. (1965) found that cocksfoot, previously heavily fertilized with N, recovered poorly when warm temperatures and low soil moisture followed cutting. The microclimate of a plant may also be altered by differences in cutting height. Soil temperatures under a short stubble are higher than under a tall stubble; Mitchell (1958) recorded a difference of as much as 17' C. at a soil depth of $in. beneath a short pasture with an open, dried-up cover (48" C.) and a long pasture with a 3 in. cover of green herbage (31" C.).
The senior author is indebted to the Wisconsin Agricultural Experiment Station, the Research Committee of the Graduate School of the University of Wisconsin, and the North Atlantic Treaty Organization (NATO) for financial support.
Both authors thank Dr G. J. A. Ryle, M r C. E. Powell and, Mrs C. A. Cowling and other members of the Plant Physiology Department for advice and help, Mr E. C. Jones for the chemical analyses, and M r P. J. Radford at Hurley and the Statistical Department at Rothamsted for the statistical analyses.
Temperature and nitrogen effects on timothy I57
R E F E R E N C E S
BEEVERS, L. & COOPER, J. P. (1964~) . Influence of temperature on growth and metabolism of ryegrass seedlings. I. Seedling growth and yield components. Crop Sci., Madison, 4, 139.
BEEVERS, L. & COPPER, J. P. (19646). Influence of temperature on growth and metabolism of ryegrass seedlings. 11. Variation in metabolites. Crop Sci., Madison, 4, 143.
BROWN, E. M. (1939). Some effects of temperature on the growth and chemical composition of certain pasture grasses. Bull. Mo. agric. Exp. Stn., no. 299.
COLBY, W. G., DRAKE, M., FIELD, D. L. & KREOWSKI, G. (1965). Seasonal pattern of fructosan in orchard grass stubble as influenced by nitrogen and harvest management. Agron. J. 57, 169.
DAVIDSON, J. L. & MILTHORPE, F. L. (1965). The effect of temperature on the growth of cocks- foot (Dactylis glomerata L.). Ann. Bot., Lond., N.S. 29, 407.
DERIAZ, R. E. (1961). Routine analysis of carbohydrates and lignin in herbage. J. Sci. Fd Agric. 12, 152.
HARRISON, C. M. (1934). Responses of Kentucky bluegrass to variations in temperature, light, cutting and fertilizing. PI. Physiol., Lancaster, 9, 83.
HORWITZ, W. (1960). Oficial methods of analysis of the Association of Oficial Agricultural Chemists. 9th ed., 832 pp. Assoc. of Official Agric. Chemists, Washington D.C.
KENDALL, W. A. (1958). The persistence of red clover and carbohydrate concentration in the roots at various temperatures. Agron. J. 50, 657.
LAMBERT, D. A. (1962). A study of growth in swards of timothy and meadow fescue. 111. The effect of two levels of nitrogen under two cutting treatments. J. agric. Sci., Camb., 59, 25.
LANCER, R. H. M., RYLE, S. M. & JEWISS, 0. R. (1964). The changing plant and tiller popula- tions of timothy and meadow fescue swards. I. Plant survival and the pattern of tillering. J. appl. Ecol. I, 197.
MITCHELL, K. J. (1956). Growth of pasture species under controlled environment. I. Growth at various levels of constant temperature. N.Z. Jl Sci. Technol. A, 38, 203.
MITCHELL, K. J. (1958). The influence of temperature on the growth of pasture plants. UNESCO Arid Zone Res. 11, 175.
MURATA, Y. & IYAMA, J. (1963). Studies on the photosynthesis of forage crops. 11. Influence of air-temperature upon the photosynthesis of some forage and grain crops. Proc. Crop Sci. SOC. Japan, 31, 315.
OKAJIMA, H. & SMITH, DALE. (1964). Available carbohydrate fractions in the stem bases and seed of timothy, smooth bromegrass and several other northern grasses. Crop Sci., Madison, 4,3 17.
RYLE, G. J. A. & LANCER, R. H. M. (1963a). Studies on the physiology of flowering of timothy (Phleum pratense L.). I. Influence of daylength and temperature on initiation and differentia- tion of the inflorescence. Ann. Bot., Lond., N.S. 27, 213.
RYLE, G. J. A. & LANCER, R. H. M. (1963 b). Studies on the physiology of flowering of timothy (Phleumpratense L.). 11. Influence of daylength and temperature on size of the inflorescence. Ann. Bot., Lond., N.S. 27, 233.
SULLIVAN, J. T. & SPRACUE, V. G. (1949). The effect of temperature on the growth and compo- sition of the stubble and roots of perennial ryegrass. PI. Physiol., Lancaster, 24, 706.
TILLEY, J. M. A. & TERRY, R. A. (1963). A two-stage technique for the in vitro digestion of forage crops. J. BY. Grassld SOC. 18, 104.