Biotechnol. J. 2013, 8 DOI 10.1002/biot.201300068 www.biotechnology-journal.com
1 Introduction
Seeds provide a useful and versatile platform for the pro-duction of recombinant proteins because they are physi-ologically and morphologically adapted for protein stor-age. Desiccated seeds offer a stable and protease-freeenvironment in which high concentrations of protein (up
to 36.5% of total soluble protein, TSP [1]) can accumulatein a small tissue volume. Developing seeds also contain arich mix of molecular chaperones and disulfide isomeras-es that facilitate protein folding [2, 3]. Although vegeta-tive tissues have a greater biomass, and a flowering cycleis unnecessary for the production of recombinant proteins[4, 5], these advantages are offset by the higher metabol-ic activity and greater water content of vegetative tissues,so that immediate processing, freezing or desiccation isnecessary in order to avoid protein degradation [2, 6, 7].Recombinant proteins expressed in vegetative tissuesmay also interfere with plant growth and development,whereas even toxic proteins can accumulate in seeds.
Cereal crops (particularly maize, rice, barley, andwheat) are favored platforms for the production of recom-binant proteins because of their high yield and well-established agricultural infrastructure, allowing produc-tion to be scaled up and down rapidly in response todemand. The cereal endosperm is a suitable target forrecombinant proteins because it is naturally adapted forprotein accumulation and the number of transgene copies
Research Article
Efficient recovery of recombinant proteins from cerealendosperm is affected by interaction with endogenous storage proteins
Jenny Peters1, Maite Sabalza2, Koreen Ramessar2, Paul Christou2, Teresa Capell2, Eva Stöger1 and Elsa Arcalís1
1 Department of Applied Genetics and Cell Biology, University of Natural Resources and Life Sciences, Vienna, Austria2 Department of Plant Production and Forestry Science, ETSEA, University of Lleida-Agrotecnio Center, Lleida, Spain
Cereal seeds are versatile platforms for the production of recombinant proteins because they pro-vide a stable environment for protein accumulation. Endogenous seed storage proteins, however,include several prolamin-type polypeptides that aggregate and crosslink via intermolecular disul-fide bridges, which could potentially interact with multimeric recombinant proteins such as anti-bodies, which assemble in the same manner. We investigated this possibility by sequentiallyextracting a human antibody expressed in maize endosperm, followed by precipitation in vitro withzein. We provide evidence that a significant proportion of the antibody pool interacts with zein andtherefore cannot be extracted using non-reducing buffers. Immunolocalization experimentsdemonstrated that antibodies targeted for secretion were instead retained within zein bodiesbecause of such covalent interactions. Our findings suggest that the production of soluble recom-binant antibodies in maize could be enhanced by eliminating or minimizing interactions withendogenous storage proteins.
Correspondence: Dr. Elsa Arcalis, Department of Applied Genetics and Cell Biology, University of Natural Resources and Life Sciences, Muthgasse 18, Vienna, AustriaE-mail: [email protected]
Current address: Maite Sabalza, Section of Cell and Development Biology,University of California, San Diego, La Jolla, CA, USAKoreen Ramessar, Molecular Targets Laboratory, Center for CancerResearch, National Cancer Institute, Frederick, MD, USAPaul Christou, Institució Catalana de Recerca i Estudis Avançats,Barcelona, Spain
Abbreviations: ER, endoplasmic reticulum; HIV, human immunodeficiencyvirus; Ig, immunoglobulin; IL-10, interleukin 10; PSV, protein storage vacuole
Received 27 MAY 2013Revised 02 AUG 2013Accepted 16 AUG 2013Accepted article online 20 AUG 2013
is boosted because the tissue is triploid and undergoesseveral rounds of endoreduplication during maturation[8]. Cereal seeds also benefit from a relatively simple seedproteome and low levels of phenolics, alkaloids, and oxal-ic acid, which are known to interfere with protein extrac-tion and downstream processing [9]. Many recombinantproteins have been produced successfully in cereal seeds,including current commercial products such as lactofer-rin, lysozyme, and human serum albumin for medical useproduced in rice by Ventria Bioscience (Fort Collins, CO,USA) [10] and human growth factor for cosmetic use pro-duced in barley by ORF Genetics (Iceland) [11].
Maize has the highest annual grain yield and a largerkernel size than other cereals, with 82% of the seed madeup of endosperm [12]. Maize offers additional advantagesover other cereal crops such as a C4 photosynthetic cycle[3], an easy transformation procedure and several endo -sperm-specific and other seed-restricted promoter systemsthat are well characterized and available for the control oftransgene expression [2, 13]. The first commercial plant-derived recombinant proteins (avidin and β-glucuronidase)were produced in maize by Prodigene Inc. (College Station,TX, USA) [14, 15]. Cellobiohydrolase, an enzyme used toproduce biofuels from plant biomass, has been expressedwith a yield of up to 30% of TSP in maize seeds [16].
Given the advantages discussed above, maize isfavored for the production of pharmaceutical proteinssuch as the human monoclonal human immunodeficien-cy virus (HIV)-neutralizing antibody 2G12 [9]. This recog-nizes oligomannose-type N-glycans on the gp120 enve-lope protein of HIV-1 and is considered a promising micro-bicide [17]. Cocktails of neutralizing antibodies appear toconfer better protection than single antibodies, but multi-ple high doses are required which means that efficientand scalable production platforms are necessary [18].Although correctly-modified full-size antibodies cannotbe produced in bacteria, plant cells can assemble thepolypeptide chains and link them via disulfide bridges ifthe protein is targeted to the secretory pathway [19, 20].Cereal seeds therefore offer one of the most promisingplatforms for the large-scale production of functional anti-bodies, as successfully demonstrated with a series of IgG,IgA, and hybrid molecules, including 2G12 [9, 19–24].
Cereals are exceptional among seed crops in that pro-teins not only accumulate in storage vacuoles but also inprotein bodies derived directly from the endoplasmicreticulum (ER). Maize prolamins (α-, δ-, and γ-zeins) aresynthesized 10–40 days after pollination (DAP) and makeup approximately 50% of the total protein content of themature seed [25]. They assemble within the ER lumen toform zein bodies, which comprise a central core of α- andδ-zeins and a peripheral layer of γ-zeins [26]. The zeinsinteract non-covalently [27] but are also cross-linked byintermolecular disulfide bridges, particularly the γ-zeinswhich are cysteine rich and form covalently linked multi-meric aggregates that initiate the formation of protein
bodies. The heterologous expression of γ-zein or its cys-teine-rich N-terminal portion is sufficient to induce ER-derived protein bodies in leaves and other non-seed tis-sues [28, 29].
We hypothesized that the abundance of zeins in maizeendosperm and the tendency (particularly of γ-zeins) toform intermolecular disulfide bridges would promoteinteractions between zeins and recombinant proteinsthat also form intermolecular disulfide bridges, such asantibodies. We tested the hypothesis by extracting therecombinant antibody 2G12 from maize seeds under dif-ferent conditions, and found that a portion of the recom-binant protein was indeed insoluble, localized in zein bod-ies, and covalently linked to γ-zein. Our data suggest thatthe yield of 2G12 antibody previously reported followingsaline extraction [9] does not accurately reflect the fullpotential of maize seeds as an expression platform, andthat higher yields of soluble antibody could be achievedin maize varieties with lower levels of endogenous γ-zeinand other storage proteins.
2 Materials and methods
2.1 Transgenic seeds
The transgenic seeds expressing 2G12 were describedpreviously [9], the high yield line producing ~75 μg2G12/g of dry seed weight and the low yield line produc-ing approximately half this amount. The 2G12 heavy andlight chains were expressed under the control of theendosperm-specific rice glutelin-1 promoter and eachcarried an N-terminal signal sequence targeting thesecretory pathway.
2.2 Light and electron microscopy
Developing transgenic seeds at three developmentalstages were harvested and bisected transversally allowingthe embryo to be removed. Thin slices were cut from theendosperm with a razor blade under 0.1 M phosphatebuffer (pH 7.4), and were fixed in 4% w/v paraformalde-hyde and 0.5% v/v glutaraldehyde in 0.1 M phosphatebuffer (pH 7.4) overnight at 4°C and further processed asdescribed previously [30]. For light microscopy, 1-μm sec-tions were stained with toluidine blue. One micrometersections mounted on glass slides for fluorescence micros -copy and 90 nm sections mounted on gold grids for elec-tron microscopy were pre-incubated incubated in 5% w/vbovine serum albumin (BSA Fraction V) in 0.1 M phos-phate buffer (pH 7.4) and then incubated with polyclonalgoat anti-human IgG (Sigma, St. Louis), polyclonal rat anti-α-zein (kindly provided by Jung and co-workers [31]) orpolyclonal rabbit anti-γ-zein (kindly provided by A. Vitale,Milan). Sections were then treated with the secondaryantibody diluted in 0.1 M phosphate buffer (pH 7.4), which
was either a donkey anti-goat IgG Alexa Fluor® 546, don-key anti-rat IgG Alexa Fluor® 488 or donkey anti-rabbit IgGAlexa Fluor® 546 for fluorescence microscopy, and rabbitanti goat IgG coupled to 10-nm colloidal gold for transmis-sion electron microscopy. Observations were made usinga Leica DM5500B fluorescence microscope and a FEI Tec-nai G2 electron microscope.
2.3 Solubility assay
Finely ground flour from mature seeds corresponding toour high expresser line [9] was extracted overnight atroom temperature in three volumes of phosphate salinebuffer containing 0.5 mM EDTA. The pellet was extractedfive times for 1 h with the same buffer to remove all tracesof soluble antibody. Residual proteins were extracted withequal amounts of the same buffer containing 5% 2-mer-captoethanol for 1 h at room temperature. The fractionswere separated by SDS-PAGE (12% AA) and analyzed byimmunoblot. 2G12 was detected with an alkaline phos-phatase labeled antibody against human IgG (Promega).The relative quantities of antibody in the different frac-tions were calculated by comparing the intensity of eachband using ImageJ software. Experiment was repeated atleast three times.
2.4 Renaturation assay
Purified 2G12 (Polymun Scientific, Inc., Vienna, Austria),maize zein (Sigma) and a mixture of the two proteins (1:1),each at a concentration of 50 or 100 μg/mL were dena-tured and reduced in 35 mM KPi-buffer (pH 7.6) contain-ing 1 mM EDTA, 8 M urea and 5% v/v 2-mercaptoethanol.The denatured samples were renatured by three-stepdialysis [32]. Samples were initially dialyzed against35 mM KPi-buffer containing 0.1 M NaCl, 40% w/v glyc-erol and 0.1 M 2-mercaptoethanol for 12 h at 4°C, followedby 24 h at 4°C against the same buffer without 2-mercap-toethanol, and finally 35 mM KPi-buffer containing 0.1 MNaCl for 24 h at 4°C. The samples were then centrifugedand the supernatants were collected while the pelletswere dissolved in 5× SDS loading buffer. Five microlitersof supernatants and pellets were loaded in an SDS-PAGEgel (12% AA) and analyzed by immunoblot. 2G12 wasdetected with an alkaline phosphatase antibody againsthuman IgG (Promega) and zein was detected with a polyclonal rabbit serum anti-γ-zein (kindly provided by A. Vitale, Milan) followed by the corresponding antiseralabeled with alkaline phosphatase (Promega). The blotwas sequentially incubated to detect both zein and IgG.Experiment was repeated at least three times.
2.5 BiP extraction and immunoblot analysis
The expression levels of the immunoglobulin (Ig) bindingprotein (BiP) were compared in the mature seeds of wild-
type plants and those expressing 2G12 at high and lowlevels. Finely ground flour was extracted in three volumesof PBS for 1 h at room temperature, and protein extractswere separated by SDS-PAGE, transferred to a nitrocellu-lose membrane and probed with a polyclonal anti-BiP rab-bit antiserum kindly provided by Ludevid and co-workers[33]. The antigen-antibody reaction was visualized withthe correspondent secondary antisera labeled with alka-line phosphatase. Experiment was repeated at least threetimes.
3 Results
3.1 The subcellular localization of 2G12 changesduring seed development
We investigated the localization of 2G12 during seeddevelopment in transgenic maize plants expressing theantibody in the seed endosperm by focusing on threewell-characterized developmental stages [30]. Stage 1corresponds to young cells that contain small starchgrains, ER-derived zein bodies, and protein storage vac-uoles (PSVs). In stage 2, the starch grains and PSVsincrease in size and number as shown in Fig. 1A. Cellscontain several PSVs ~5 μm in diameter and abundantzein bodies throughout the cytoplasm. In stage 3, as theseed approaches maturity, the endosperm cells arepacked with starch and zein bodies but PSVs can nolonger be identified [30].
The localization of 2G12 was found to change duringseed development. In stage 1, the antibody was detectedwithin the PSVs but a substantial fraction was also pres-ent in the zein bodies (Fig. 1B). In stage 2, the antibodywas more prominently localized in the zein bodies, typi-cally as ring-like peripheral deposits, additional to the ful-ly labeled zein bodies present in stage 1 (Fig. 1C). In stage3, the zein bodies formed clusters as previously reported[30] with a peripheral layer of 2G12 (Fig. 1D). Electronmicroscopy confirmed that 2G12 was deposited in thePSVs of young cells (stage 1), and was present within Gol-gi-associated vesicles of such cells confirming that theantibody passed through the Golgi body en route to thePSVs (Fig. 1E–F).
3.2 The morphology of zein bodies changes in the presence of 2G12
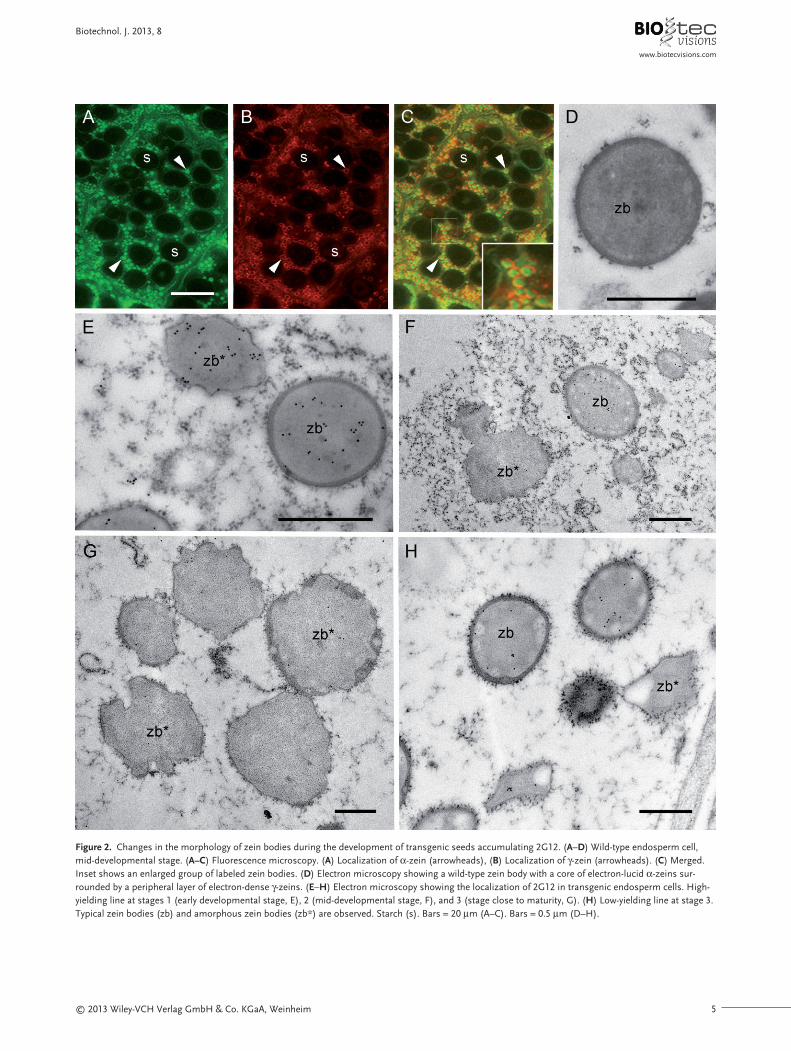
The three zein subfamilies are organized within themature protein body in a very specific manner, to form acentral electron-lucent core of α- and δ-zeins (Fig. 2A, C,D, inset in D) [26] and a peripheral electron-dense ring ofγ-zeins (Fig. 2B–D, inset in D) [26]. Early zein bodies con-tained 2G12 in the inner core, but in older seeds antibodylabeling was particularly strong around the periphery(Figs. 1B–D and 2E). We also found that the zein bodies
became progressively more abnormal during develop-ment in the presence of 2G12 (Fig. 2E–H). In stage 1, thezein bodies were approximately spherical, although somewere amorphous with a thin γ-zein layer (Fig. 2E). In stage 2, normal zein bodies were still observed but amor-
phous zein bodies were more common, featuring wavymargins and atypical distribution of the zein subfamilies(Fig. 2F). In stage 3, normal zein bodies were rare and theγ-zein layer was identified in a few protein bodies but onlyas a non-continuous layer with variable thickness. Amor-
www.biotechnology-journal.com
Figure 1. Subcellular localization of 2G12during seed development. (A) Lightmicroscopy, stained with toluidine blue.Representative maize endosperm cell,mid developmental stage showing twoprotein storage compartments: PSVs(arrows) and zein bodies (arrowheads).(B–D) Localization of 2G12 at stages 1(early developmental stage, B), 2 (mid-developmental stage, C), and 3 (stageclose to maturity, D). (B) Strong signalwithin the PSVs (arrows) and within thezein bodies (arrowheads). (C) Ring-likesignal around the zein bodies (arrow-heads) and labeling within the zein bod-ies (arrows). (D) Signal in between theclustered zein bodies (arrowheads).Nucleus (n), starch (s). (E, F) Electronmicroscopy, localization of 2G12 withinthe PSVs (arrow, E) and in Golgi-derivedvesicles (arrows, F). Golgi (g), mitochon-dria (m), and rough endoplasmic reticu-lum (rER). Bars = 20 μm (A–D).Bars = 0.25 μm (E–F).
Figure 2. Changes in the morphology of zein bodies during the development of transgenic seeds accumulating 2G12. (A–D) Wild-type endosperm cell,mid-developmental stage. (A–C) Fluorescence microscopy. (A) Localization of α-zein (arrowheads), (B) Localization of γ-zein (arrowheads). (C) Merged.Inset shows an enlarged group of labeled zein bodies. (D) Electron microscopy showing a wild-type zein body with a core of electron-lucid α-zeins sur-rounded by a peripheral layer of electron-dense γ-zeins. (E–H) Electron microscopy showing the localization of 2G12 in transgenic endosperm cells. High-yielding line at stages 1 (early developmental stage, E), 2 (mid-developmental stage, F), and 3 (stage close to maturity, G). (H) Low-yielding line at stage 3.Typical zein bodies (zb) and amorphous zein bodies (zb*) are observed. Starch (s). Bars = 20 μm (A–C). Bars = 0.5 μm (D–H).
Zein/2G12 heavy chain hybrids should be recognized asunfolded proteins and should therefore induce the unfold-ed protein response (UPR) and ER stress. We investigatedthis hypothesis by comparing the levels of the chaperoneBiP in seeds from wild-type plants and those producinghigh and low yields of 2G12. Preliminary data seem toindicate an increase in BiP in both transgenic lines (seeSupporting information). The seeds were also pho-tographed under transmitted light to screen for a poten-tial opaque phenotype, which is found in several naturalmaize mutants with abnormal zein bodies, but we foundno significant difference between the wild-type andtransgenic seeds (Fig. 4).
4 Discussion
The HIV-neutralizing antibody 2G12 has previously beenexpressed in maize seeds and extensive structural andfunctional characterization has confirmed that its anti-gen-binding and HIV-neutralizing properties are equal orsuperior to those of the same antibody produced in Chi-nese hamster ovary cells, the “gold standard” industryplatform [9]. We carried out a detailed analysis of the local-ization of this antibody in developing endosperm cells andalso studied its extractability from seeds. As anticipatedfor a recombinant protein carrying a signal peptide but nofurther targeting information, 2G12 was predominantlylocalized within the PSVs of young endosperm cells [30,34, 35], but the intracellular localization of the antibodyprogressively switched to ER-derived protein bodies asthe seed matured, as previously observed for recombinantfungal phytase expressed in the same tissue [30].
In contrast to phytase, which was solely located in theperipheral zone, we found that 2G12 was also present inzein body core, as previously observed for 2G12 append-ed with a C-terminal KDEL tag for retrieval to the ER
www.biotechnology-journal.com
phous zein bodies with wavy margins and lobular struc-tures were prevalent in these mature cells (Fig. 2G). How-ever, a stage-matched sample from another transgenicmaize line producing lower yields of the antibody con-tained a mixture of these amorphous, disorganized zeinbodies and spherical bodies with the typical distributionof zeins, suggesting the degree of abnormality correlatedwith the quantity of antibody (Fig. 2H).
3.3 Insoluble 2G12 accumulates due to covalentinteractions with zeins
We investigated the potential for covalent interactionsbetween 2G12 and endogenous zeins because both pro-teins assemble into multimeric structures via the forma-tion of intermolecular disulfide bonds. Mature seedsexpressing 2G12 were therefore extracted repeatedlywith a saline buffer, resulting in the complete extractionof soluble antibody chains after six rounds. When theresidual protein fraction was extracted under reducingconditions, an essential part of the total amount wasreleased (Fig. 3A). These data indicate that the antibodyinteracts covalently with endogenous seed proteins,resulting in the formation of insoluble oligomers withinthe zein network.
We gained further evidence for this phenomenon bydenaturing and reducing zein, 2G12, and mixtures of thetwo proteins in a buffer containing urea and 2-mercap-toethanol. Upon stepwise dialysis [32], the individual pro-tein samples regained their normal behavior, i.e. most ofthe antibody became soluble in the supernatant whereasmost of the zein was precipitated and was found in thepellet (Fig. 3B; lanes 1 and 2, respectively). However,when the mixture of 2G12 and zein was dialyzed, only asmall portion of the antibody was resolubilized and mostremained in the precipitate along with the zeins (Fig. 3B;lanes 3 and 4, respectively).
Figure 3. Solubility and renaturation assays followed by SDS-PAGE and immunoblot analysis. (A) Solubility assay (see Section 2): Salt extract (lane 1),wash steps (lanes 2–6), re-extraction of the pellet under reducing conditions (lane 7). The extract loaded per lane corresponds to ~5 mg of ground tissue.(B) Renaturation of 2G12 and zein either alone or in combination (1 = 100 μg/mL, 2 = 50 μg/mL) by stepwise dialysis: 2G12 (supernatant), zein (pellet),and 2G12-zein mixture (S, supernatant; P, pellet). Five microliters of sample were loaded per lane. M, size markers; HC, heavy chain; LC, light chain.
lumen [24, 30]. Recombinant proteins targeted for secre-tion from the cereal endosperm are often found partiallydistributed within the ER- and ER-derived protein bodies,especially in rice [35–40]. As previously shown, the fourpolypeptide components of a secretory IgA accumulateexclusively within ER-derived protein bodies (and not inthe PSV) of rice endosperm cells when expressed individ-ually, whereas the assembled dimeric IgA accumulatesspecifically within the PSVs when the components arecombined in the same cell [41]. From these data, wehypothesized that the individual polypeptides may havebeen influenced by interactions with endogenous riceprolamins through the formation of intermolecular disul-fide bonds, whereas the lack of available free cysteineresidues on the dimeric IgA prevented such interactions.Here, we similarly found evidence for a covalent interac-tion between the 2G12 heavy chain and endogenous zeinproteins, resulting in a substantial insoluble antibodyfraction that could only be extracted using a reducingbuffer, and the presence of distorted zein bodies in trans-genic seeds suggesting that the formation of 2G12/zeincomplexes interferes with normal zein packing and pre-vents the formation of bodies with typical layered struc-tures.
The prevalence of abnormal zein bodies appeared tobe dependent on the quantity of 2G12 because the disor-dered bodies were more common and more disorganizedin seeds with the highest antibody yields. It is unlikelythat passive trapping of antibody chains in the zein bod-ies would distort the organelle morphology because thenative interactions between zeins would be preserved.We propose that a more likely explanation is the formationof aberrant aggregates, which would disrupt the balancebetween the endogenous zein subfamilies and lead to dis-ordered packing. Previous studies investigating the het-erologous expression of zein subgroups alone or in com-bination have shown that the presence and balancedinteraction of all zein subfamilies is necessary to stabilizethe zein bodies and form typical spherical structures.Thus, although δ-zein [42], β-zein (re-assigned to the γ-zein subfamily [31]) [43], and γ-zein [28] can form protein
aggregates by themselves, these do not form typical zeinbodies. For example, α-zein forms stable protein bodieswhen expressed in Xenopus laevis oocytes but not intobacco leaves, but typical protein body structures areformed ectopically when different zein subtypes are coex-pressed [44]. In the final stages of protein body formation,the typical zein body consists of a central core of α- andβ-zeins surrounded by a thin layer of γ-zeins [26]. In con-trast, rosette-shaped protein bodies form when β-zein isexpressed alone [43] or in combination with δ-zein [42],but small, round or amorphous protein bodies form whenβ-zein is expressed in combination with a-zein [45].
Mutations in zein genes also result in the developmentof endosperm cells with abnormal zein bodies in terms ofshape, size, and/or number, e.g. the high-lysine mutantsfloury2 (fl2), Mucuronate (Mc), and Defective endosperm-B30* (DeB30*) are associated with amorphous proteinbodies that appear similar to those we observed. Wetherefore looked for phenotypic correlations betweenthese three mutants and our transgenic lines, includingthe opaque phenotype and higher levels of the ER-resi-dent chaperone BiP [46]. Neither of the 2G12 transgeniclines had an opaque phenotype but we obtained prelimi-nary results that indicate an increase in the abundance ofBiP (Supporting information, Fig. S1), suggesting theonset of ER stress and, indirectly, the activation of theUPR cascade. The UPR cascade was triggered in Ara-bidopsis thaliana seeds expressing a recombinant scFv-Fc antibody, reflecting the disruption of ER home-ostasis [47]. Furthermore, the interaction between freeantibody chains and endogenous storage proteins couldalso induce the expression of chaperones, as previouslyreported in rice seeds expressing a recombinant versionof human interleukin 10 (IL-10) that also interacted withcysteine-rich prolamins [38]. We found that the putativeER stress did not affect the germination of our maizeseeds or seedling growth (data not shown) similar to theobservations in Arabidopsis [47].
The interaction with endogenous storage preventedthe extraction of more than 50% of the total antibody pro-duced in maize seeds, due to the formation of insoluble
Biotechnol. J. 2013, 8
www.biotecvisions.com
Figure 4. Phenotype analysis. (A) Kernel phenotype of wild-type seeds, (B) seeds from a line with high yields of 2G12, and (C) seeds from a line with lowyields of 2G12. Photographs were taken under transmitted light.
teine-containing peptide 7Crp peptide accumulates inER-derived protein bodies in rice regardless of other sort-ing signals that may be present due to the formation ofdisulfide bonds with endogenous prolamins [56]. The judi-cious manipulation of the storage protein machinery inmaize seeds can therefore be used on a case-by-casebasis to a void interactions and increase the yield of solu-ble recombinant proteins, or to encourage those interac-tions and reduce the rate of protein release in harsh envi-ronments.
Research at the University of Natural Resources and LifeSciences in Vienna was supported by the ERA net projectFKZ0313994. Research at the Universitat de Lleida is sup-ported by MICINN, Spain BIO2012-35359; and CentreCONSOLIDER on Agrigenomics funded by MICINN,Spain. Collaboration between the authors was supportedby COST Action FA0804: Molecular farming: plants as aproduction platform for high value proteins.
The authors declare no financial or commercial conflict ofinterest.
5 References
[1] De Jaeger, G., Scheffer, S., Jacobs, A., Zambre, M. et al., Boostingheterologous protein production in transgenic dicotyledonous seedsusing Phaseolus vulgaris regulatory sequences. Nat. Biotechnol.2002, 20, 1265–1268.
[2] Stoger, E., Ma, J. K., Fischer, R., Christou, P., Sowing the seeds ofsuccess: Pharmaceutical proteins from plants. Curr. Opin. Biotech-nol. 2005, 16, 167–173.
[3] Naqvi, S., Ramessar, K., Farre, G., Sabalza, M. et al., High-value prod-ucts from transgenic maize. Biotechnol. Adv. 2011, 29, 40–53.
[4] Twyman, R. M., Stoger, E., Schillberg, S., Christou, P., Fischer, R.,Molecular farming in plants: Host systems and expression technolo-gy. Trends Biotechnol. 2003, 21, 570–578.
[5] Abranches, R., Marcel, S., Arcalis, E., Altmann, F. et al., Plants asbioreactors: A comparative study suggests that Medicago truncat-ula is a promising production system. J. Biotechnol. 2005, 120, 121–134.
[6] Fischer, R., Stoger, E., Schillberg, S., Christou, P., Twyman, R. M.,Plant-based production of biopharmaceuticals. Curr. Opin. PlantBiol. 2004, 7, 152–158.
[7] Twyman, R. M., Schillberg, S., Fischer, R., Transgenic plants in thebiopharmaceutical market. Exp. Opin. Emerg. Drugs 2005, 10,185–218.
[8] Sabelli, P. A., Larkins, B. A., The development of endosperm in grass-es. Plant Physiol. 2009, 149, 14–26.
[9] Ramessar, K., Rademacher, T., Sack, M., Stadlmann, J. et al., Cost-effective production of a vaginal protein microbicide to prevent HIVtransmission. Proc. Natl. Acad. Sci. USA 2008, 105, 3727–3732.
[10] Ramessar, K., Sabalza, M., Capell, T., Christou, P., Maize plants: Anideal production platform for effective and safe molecular pharming.Plant Sci. 2008, 174, 409–419.
[11] Erlendsson, L. S., Muench, M. O., Hellman, U., Hrafnkelsdottir, S. M.et al., Barley as a green factory for the production of functional Flt3ligand. Biotechnol. J. 2010, 5, 163–171.
www.biotechnology-journal.com
oligomers. This has a clear impact on the economic valueof seeds as an expression platform for recombinant anti-bodies because it halves the efficiency of downstreamprocessing. The maximum yield of soluble 2G12 reportedin transgenic maize seeds is 75 μg/g, with a recovery rateof 60–90% using a bench-scale purification method [9, 24].Although such yields are comparable to those achieved intransgenic tobacco leaves per unit of biomass (T. Rade -macher, personal communication) our data suggest thatthe intrinsic capability of maize seeds could be doubled ifthe antibody could be persuaded to accumulate in a solu-ble form. Similar considerations apply for the productionof other recombinant proteins with free cysteine residues,e.g. the major house dust mite allergen Der p 1, the Japan-ese cedar pollen allergen Cry j 1, and the human cytokineIL-10, all required a reducing agent to achieve completeextraction from rice seeds because a significant propor-tion of each protein interacted with rice prolamins [36, 38,48].
One potential strategy to minimize interactionsbetween cysteine-rich recombinant proteins and endoge-nous zeins is the use of maize mutants with lower prolaminlevels. The silencing of key zein genes could increase thesoluble fraction of recombinant proteins with free cysteineresidues, and could also increase the overall yield ofrecombinant proteins by freeing up more of the proteinsynthesis and storage capacity. The suppression ofendogenous prolamins (but not glutelins) increased theyields of human IL-10 by three-fold in rice seeds, perhapsby reducing competition for the filling of ER-derived com-partments [38]. The suppression of endogenous storageprotein genes is also known to increase the expression ofother seed proteins at the transcriptional and translationallevels in order to rebalance seed protein levels and main-tain homeostasis, thus it may be possible to introduce arecombinant protein as a surrogate storage protein in thisbalanced environment [49–51]. Although the suppressionof zeins appears as a promising strategy to increase thesoluble fraction of recombinant antibodies in maize, otherproteins with free cysteine residues have an inherent tendency to self-aggregate and form ER-derived proteinbodies that remain separate from prolamin bodies [36–38,52]. In these cases the insoluble fraction is less likely to beaffected by the presence or absence of prolamins.
Interactions with endogenous prolamins may be ben-eficial in some cases and should therefore be encouraged,e.g. for the production of pharmaceuticals for oral delivery.The complete purification of a recombinant protein des-tined for oral delivery is often unnecessary, and encapsu-lation of the recombinant protein within seed storageorganelles is often desirable because thus offers partialprotection from digestive enzymes and the acidic gut.Prolamin bodies are particularly insensitive to pepsindigestion, allowing the slow release of recombinant pro-teins from seeds which is advantageous for the delivery of vaccines, enzymes and tolerogens [53-55]. The cys-
[12] Watson, S. A., Structure and composition, in: Watson, S. A., Ram-stad, P. T. (Ed.), Corn: Chemistry and Technology, American Associ-ation of Cereal Chemists, St. Paul, MN 1987, pp. 53–82.
[13] Kawakatsu, T., Takaiwa, F., Cereal seed storage protein synthesis:Fundamental processes for recombinant protein production in cere-al grains. Plant Biotechnol. J. 2010, 8, 939–953.
[14] Hood, E. E., Witcher, D. R., Maddock, S., Meyer, T. et al., Commer-cial production of avidin from transgenic maize: Characterization oftransformant, production, processing, extraction and purification.Mol. Breeding 1997, 3, 291–306.
[15] Witcher, D. R., Hood, E. E., Peterson, D., Bailey, M. et al., Commer-cial production of beta-glucuronidase (GUS): A model system for theproduction of proteins in plants. Mol. Breeding 1998, 4, 301–312.
[16] Devaiah, S. P., Requesens, D. V., Chang, Y. K., Hood, K. R. et al., Het-erologous expression of cellobiohydrolase II (Cel6A) in maizeendosperm. Transgenic Res. 2013, 22, 477-488.
[17] Armitage, A. E., McMichael, A. J., Drakesmith, H., Reflecting on aquarter century of HIV research. Nat. Immunol. 2008, 9, 823–826.
[18] Trkola, A., Kuster, H., Rusert, P., Joos, B. et al., Delay of HIV-1rebound after cessation of antiretroviral therapy through passivetransfer of human neutralizing antibodies. Nat. Med. 2005, 11, 615–622.
[19] Ma, J. K., Hiatt, A., Hein, M., Vine, N. D. et al., Generation andassembly of secretory antibodies in plants. Science 1995, 268, 716–719.
[20] Frigerio, L., Vine, N. D., Pedrazzini, E., Hein, M. B. et al., Assembly,secretion, and vacuolar delivery of a hybrid immunoglobulin inplants. Plant Physiol. 2000, 123, 1483–1494.
[21] Nicholson, L., Gonzalez-Melendi, P., van Dolleweerd, C., Tuck, H.et al., A recombinant multimeric immunoglobulin expressed in riceshows assembly-dependent subcellular localization in endospermcells. Plant Biotechno. J. 2005, 3, 115–127.
[22] Van Droogenbroeck, B., Cao, J., Stadlmann, J., Altmann, F. et al.,Aberrant localization and underglycosylation of highly accumulat-ing single-chain Fv-Fc antibodies in transgenic Arabidopsis seeds.Proc. Natl. Acad. Sci. USA 2007, 104, 1430–1435.
[23] Floss, D. M., Sack, M., Arcalis, E., Stadlmann, J. et al., Influence ofelastin-like peptide fusions on the quantity and quality of a tobacco-derived human immunodeficiency virus-neutralizing antibody.Plant Biotechnol. J. 2009, 7, 899–913.
[24] Rademacher, T., Sack, M., Arcalis, E., Stadlmann, J. et al., Recombi-nant antibody 2G12 produced in maize endosperm efficiently neu-tralizes HIV-1 and contains predominantly single-GlcNAc N-gly-cans. Plant Biotechnol. J. 2008, 6, 189–201.
[25] Lee, K. H., Jones, R. A., Dalby, A., Tsai, C. Y., Genetic-regulation ofstorage protein-content in maize endosperm. Biochem. Genet. 1976,14, 641–650.
[26] Lending, C. R., Larkins, B. A., Changes in the zein composition ofprotein bodies during maize endosperm development. Plant Cell1989, 1, 1011–1023.
[27] Kim, C. S., Woo Ym, Y. M., Clore, A. M., Burnett, R. J. et al., Zein pro-tein interactions, rather than the asymmetric distribution of zeinmRNAs on endoplasmic reticulum membranes, influence proteinbody formation in maize endosperm. Plant Cell 2002, 14, 655–672.
[28] Geli, M. I., Torrent, M., Ludevid, D., Two structural domains mediatetwo sequential events in [gamma]-zein targeting: Protein endoplas-mic reticulum retention and protein body formation. Plant Cell 1994,6, 1911–1922.
[29] Mainieri, D., Rossi, M., Archinti, M., Bellucci, M. et al., Zeolin. A newrecombinant storage protein constructed using maize gamma-zeinand bean phaseolin. Plant Physiol. 2004, 136, 3447–3456.
[30] Arcalis, E., Stadlmann, J., Marcel, S., Drakakaki, G. et al., The chang-ing fate of a secretory glycoprotein in developing maize endosperm.Plant Physiol. 2010, 153, 693–702.
[31] Woo, Y. M., Hu, D. W., Larkins, B. A., Jung, R., Genomics analysis ofgenes expressed in maize endosperm identifies novel seed proteinsand clarifies patterns of zein gene expression. Plant Cell 2001, 13,2297–2317.
[32] Katsube, T., Kurisaka, N., Ogawa, M., Maruyama, N. et al., Accu-mulation of soybean glycinin and its assembly with the glutelins inrice. Plant physiology 1999, 120, 1063–1074.
[33] Joseph, M., Ludevid, M. D., Torrent, M., Rofidal, V. et al., Proteomiccharacterisation of endoplasmic reticulum-derived protein bodies intobacco leaves. BMC Plant Biol. 2012, 12, 36.
[34] Arcalis, E., Marcel, S., Altmann, F., Kolarich, D. et al., Unexpecteddeposition patterns of recombinant proteins in post-endoplasmicreticulum compartments of wheat endosperm. Plant Physiol. 2004,136, 3457–3466.
[35] Drakakaki, G., Marcel, S., Arcalis, E., Altmann, F. et al., The intracel-lular fate of a recombinant protein is tissue dependent. Plant Physi-ol. 2006, 141, 578–586.
[36] Yang, L., Hirose, S., Suzuki, K., Hiroi, T., Takaiwa, F., Expression ofhypoallergenic Der f 2 derivatives with altered intramolecular disul-phide bonds induces the formation of novel ER-derived protein bod-ies in transgenic rice seeds. J. Exp. Bot. 2012, 63, 2947–2959.
[37] Wang, S., Takahashi, H., Kajiura, H., Kawakatsu, T. et al., Transgenicrice seeds accumulating recombinant hypoallergenic birch pollenallergen bet v 1 generate giant protein bodies. Plant Cell Physiol.2013, 54, 914-933.
[38] Yang, L., Hirose, S., Takahashi, H., Kawakatsu, T., Takaiwa, F.,Recombinant protein yield in rice seed is enhanced by specific sup-pression of endogenous seed proteins at the same deposit site. PlantBiotechnol. J. 2012, 10, 1035–1045.
[39] Stoger, E., Vaquero, C., Torres, E., Sack, M. et al., Cereal crops asviable production and storage systems for pharmaceutical scFv anti-bodies. Plant Mol. Biol. 2000, 42, 583–590.
[40] Torres, E., Gonzalez-Melendi, P., Stoger, E., Shaw, P. et al., Native andartificial reticuloplasmins co-accumulate in distinct domains of theendoplasmic reticulum and in post-endoplasmic reticulum com-partments. Plant Physiol. 2001, 127, 1212–1223.
[41] Nicholson, L., Gonzalez-Melendi, P., van Dolleweerd, C., Tuck, H. etal., A recombinant multimeric immunoglobulin expressed in riceshows assembly-dependent subcellular localization in endospermcells. Plant Biotechnol. J. 2005, 3, 115–127.
[42] Bagga, S., Adams, H. P., Rodriguez, F. D., Kemp, J. D., Sengupta-Gopalan, C., Coexpression of the maize delta-zein and beta-zeingenes results in stable accumulation of delta-zein in endoplasmicreticulum-derived protein bodies formed by beta-zein. Plant Cell1997, 9, 1683–1696.
[43] Bagga, S., Adams, H., Kemp, J. D., Sengupta-Gopalan, C., Accumu-lation of 15-kilodalton zein in novel protein bodies in transgenictobacco. Plant Physiol 1995, 107, 13–23.
[44] Wallace, J. C., Galili, G., Kawata, E. E., Cuellar, R. E. et al., Aggrega-tion of lysine-containing zeins into protein bodies in Xenopusoocytes. Science 1988, 240, 662–664.
[45] Coleman, C. E., Yoho, P. R., Escobar, S., Ogawa, M., The accumula-tion of alpha-zein in transgenic tobacco endosperm is stabilized byco-expression of beta-zein. Plant Cell Physiol. 2004, 45, 864–871.
[46] Zhang, F., Boston, R. S., Increases in binding-protein (Bip) accom-pany changes in protein body morphology in 3 high-lysine mutantsof maize. Protoplasma 1992, 171, 142–152.
[47] De Wilde, K., De Buck, S., Vanneste, K., Depicker, A., Recombinantantibody production in Arabidopsis seeds triggers an unfolded pro-tein response. Plant Physiol. 2013, 161, 1021–1033.
[48] Yang, L., Suzuki, K., Hirose, S., Wakasa, Y., Takaiwa, F., Developmentof transgenic rice seed accumulating a major Japanese cedar pollenallergen (Cry j 1) structurally disrupted for oral immunotherapy. PlantBiotechnol. J. 2007, 5, 815–826.
[49] Kawakatsu, T., Hirose, S., Yasuda, H., Takaiwa, F., Reducing riceseed storage protein accumulation leads to changes in nutrientquality and storage organelle formation. Plant Physiol. 2010, 154,1842–1854.
[50] Schmidt, M. A., Barbazuk, W. B., Sandford, M., May, G. et al., Silenc-ing of soybean seed storage proteins results in a rebalanced proteincomposition preserving seed protein content without major collat-eral changes in the metabolome and transcriptome. Plant Physiol.2011, 156, 330–345.
[51] Tada, Y., Utsumi, S., Takaiwa, F., Foreign gene products can beenhanced by introduction into low storage protein mutants. PlantBiotechnol. J. 2003, 1, 411–422.
[52] Yang, L., Hirose, S., Takahashi, H., Kawakatsu, T., Takaiwa, F.,Recombinant protein yield in rice seed is enhanced by specific sup-pression of endogenous seed proteins at the same deposit site. PlantBiotechnol. J. 2012, 10, 1035-1045.
[53] Takagi, H., Hiroi, T., Hirose, S., Yang, L., Takaiwa, F., Rice seed ER-derived protein body as an efficient deliver vehicle for oral tolero-genic peptides. Peptides 2010, 31, 1421–1425.
[55] Wakasa, Y., Takaiwa, F., The use of rice seeds to produce humanpharmaceuticals for oral therapy. Biotechnol. J. 2013, 8, DOI:10.1002/biot.201200065.
[56] Takaiwa, F., Hirose, S., Takagi, H., Yang, L., Wakasa, Y., Depositionof a recombinant peptide in ER-derived protein bodies by retentionwith cysteine-rich prolamins in transgenic rice seed. Planta 2009,229, 1147–1158.