Electrostatic shape control of a charged molecular membrane from ribbon to scroll Changrui Gao a,1 , Sumit Kewalramani a,1 , Dulce Maria Valencia a , Honghao Li a , Joseph M. McCourt b , Monica Olvera de la Cruz a,b,c,2 , and Michael J. Bedzyk a,b,2 a Department of Materials Science and Engineering, Northwestern University, Evanston, IL 60208; b Department of Physics and Astronomy, Northwestern University, Evanston, IL 60208; and c Department of Chemistry, Northwestern University, Evanston, IL 60208 Edited by Lia Addadi, Weizmann Institute of Science, Rehovot, Israel, and approved September 22, 2019 (received for review August 12, 2019) Bilayers of amphiphiles can organize into spherical vesicles, nano- tubes, planar, undulating, and helical nanoribbons, and scroll-like cochleates. These bilayer-related architectures interconvert under suitable conditions. Here, a charged, chiral amphiphile (palmitoyl- lysine, C 16 -K 1 ) is used to elucidate the pathway for planar nano- ribbon to cochleate transition induced by salt (NaCl) concentration. In situ small- and wide-angle X-ray scattering (SAXS/WAXS), atomic force and cryogenic transmission electron microscopies (AFM and cryo-TEM) tracked these transformations over angstrom to microme- ter length scales. AFM reveals that the large length (L) to width (W) ratio nanoribbons (L/W > 10) convert to sheets (L/W → 1) before rolling into cochleates. A theoretical model based on electrostatic and surface energies shows that the nanoribbons convert to sheets via a first-order transition, at a critical Debye length, with 2 shallow minima of the order of thermal energy at L/W >> 1 and at L/W = 1. SAXS shows that interbilayer spacing (D) in the cochleates scales linearly with the Debye length, and ranges from 13 to 35 nm for NaCl concentrations from 100 to 5 mM. Theoretical arguments that include electrostatic and elastic energies explain the membrane roll- ing and the bilayer separation–Debye length relationship. These models suggest that the salt-induced ribbon to cochleate transi- tion should be common to all charged bilayers possessing an intrinsic curvature, which in the present case originates from molecular chirality. Our studies show how electrostatic interac- tions can be tuned to attain and control cochleate structures, which have potential for encapsulating, and releasing macromol- ecules in a size-selective manner. bilayer assembly | electrostatics | nanoribbon | cochleate A mphiphilic molecules can self-assemble into a variety of 3D, 2D, and 1D nano- and mesoscale structures. These struc- tures serve as simplified models for understanding biological assemblies and their functions and have applications in drug delivery (1–5), regenerative medicine (6, 7), biosensing (8), hy- drogen production (9, 10), and clean water technologies (11). An interesting assembly structure is the nanoribbon, which is a high aspect ratio (10:1 or greater) bilayer. Nanoribbons are a gateway to a number of other morphologies with distinct functionalities. For example, nanoribbons of a charged chromophore amphiphile can transform to a scroll-like (cochleate) morphology when the solu- tion ionic strength is increased (9). These cochleates serve as ef- ficient charge-transfer agents for photocatalysts in hydrogen production. Cochleate formation from liposomes of negatively charged phospholipids in the presence of multivalent cations also involves a nanoribbon intermediate (3, 12, 13). Biocompatible phospholipid cochleates are being explored as drug-delivery agents because they can trap macromolecules, such as proteins, and DNA, and provide protection against degradation due to their multilayer geometry. Nanoribbons have also been observed in peptide amphiphiles (PAs), which consist of a sequence of amino acids covalently linked to an alkyl tail (14, 15). For example, a peptide amphiphile that stimulates collagen production has been found to self-assemble into nanotapes with an internal bilayer structure (16). In a PA with alternating charged and neutral amino acids, nanoribbons were found to transform into helical ribbons as the PA concentration was reduced (17) and into helical and twisted nanoribbons when the amino acid sequence was permuted (18). Helical assemblies have been previously used to template semiconductor nanohelices (19). Despite the progress, the corre- lation between experimental conditions such as molecular design, ionic strength, pH, amphiphile concentration, and the attained nanoribbon-related morphology are not fully established. There- fore, precise control of nanoribbon-related architecture requires further understanding of the delicate interplay between intermo- lecular interactions and elastic and interfacial energies. A recent theoretical study showed that for charged molecules, tuning the range of electrostatic interactions could induce tran- sitions between different nanoribbon-related morphologies (20). Specifically, a phase diagram was deduced for a 2D lattice of charged points, which interacted via long-range repulsive elec- trostatic interactions and short-range attractive interactions. Planar nanoribbon to wavy ribbon with periodic undulations to helical ribbon transitions were predicted as the range of the electrostatic interactions is increased. This study suggests a facile method for accessing distinct nanoribbon architectures by vary- ing the ionic strength (μ) of the solution because the range of electrostatic interactions as parametrized by Debye length (λ d ) scales as μ −1/2 . Recent experiments also attest that tuning the Significance Controlling the shape and internal architecture of assemblies of amphiphiles is critical for many technologies. The structure, and thus the function, of these assemblies reconfigures in re- sponse to stimuli, via mechanisms that are often elusive. Here, we observe and explain how molecular reordering driven by variations in electrostatic screening length induce micrometer- scale structural changes in crystalline membranes of charged, chiral molecules: The transformation of high aspect ratio, pla- nar bilayers into scroll-like cochleates by increasing the solution salt content is described and explained. Our study suggests that this transformation should be general to charged bilayers pos- sessing a spontaneous curvature. Author contributions: S.K., M.O.d.l.C., and M.J.B. designed research; C.G., S.K., D.M.V., H.L., and J.M.M. performed research; C.G., S.K., D.M.V., H.L., J.M.M., M.O.d.l.C., and M.J.B. analyzed data; and C.G., S.K., D.M.V., M.O.d.l.C., and M.J.B. wrote the paper. The authors declare no competing interest. This article is a PNAS Direct Submission. Published under the PNAS license. Data deposition: All the X-ray data shown in the manuscript and the SI Appendix, as well as the code for simulating the WAXS intensity along with the data from theory calcula- tions, have been deposited at Bitbucket (https://bitbucket.org/NUaztec/gao_et_al_pnas_ 2019_charged_membrane/src/master/). 1 C.G. and S.K. contributed equally to this work. 2 To whom correspondence may be addressed. Email: [email protected] or [email protected]. This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10. 1073/pnas.1913632116/-/DCSupplemental. First published October 14, 2019. 22030–22036 | PNAS | October 29, 2019 | vol. 116 | no. 44 www.pnas.org/cgi/doi/10.1073/pnas.1913632116 Downloaded by guest on December 4, 2021
Transcript
Electrostatic shape control of a charged molecularmembrane from ribbon to scrollChangrui Gaoa,1, Sumit Kewalramania,1, Dulce Maria Valenciaa, Honghao Lia, Joseph M. McCourtb,Monica Olvera de la Cruza,b,c,2, and Michael J. Bedzyka,b,2
aDepartment of Materials Science and Engineering, Northwestern University, Evanston, IL 60208; bDepartment of Physics and Astronomy, NorthwesternUniversity, Evanston, IL 60208; and cDepartment of Chemistry, Northwestern University, Evanston, IL 60208
Edited by Lia Addadi, Weizmann Institute of Science, Rehovot, Israel, and approved September 22, 2019 (received for review August 12, 2019)
Bilayers of amphiphiles can organize into spherical vesicles, nano-tubes, planar, undulating, and helical nanoribbons, and scroll-likecochleates. These bilayer-related architectures interconvert undersuitable conditions. Here, a charged, chiral amphiphile (palmitoyl-lysine, C16-K1) is used to elucidate the pathway for planar nano-ribbon to cochleate transition induced by salt (NaCl) concentration.In situ small- and wide-angle X-ray scattering (SAXS/WAXS), atomicforce and cryogenic transmission electron microscopies (AFM andcryo-TEM) tracked these transformations over angstrom tomicrome-ter length scales. AFM reveals that the large length (L) to width (W)ratio nanoribbons (L/W > 10) convert to sheets (L/W → 1) beforerolling into cochleates. A theoretical model based on electrostaticand surface energies shows that the nanoribbons convert to sheetsvia a first-order transition, at a critical Debye length, with 2 shallowminima of the order of thermal energy at L/W >> 1 and at L/W = 1.SAXS shows that interbilayer spacing (D) in the cochleates scaleslinearly with the Debye length, and ranges from 13 to 35 nm forNaCl concentrations from 100 to 5 mM. Theoretical arguments thatinclude electrostatic and elastic energies explain the membrane roll-ing and the bilayer separation–Debye length relationship. Thesemodels suggest that the salt-induced ribbon to cochleate transi-tion should be common to all charged bilayers possessing anintrinsic curvature, which in the present case originates frommolecular chirality. Our studies show how electrostatic interac-tions can be tuned to attain and control cochleate structures,which have potential for encapsulating, and releasing macromol-ecules in a size-selective manner.
Amphiphilic molecules can self-assemble into a variety of 3D,2D, and 1D nano- and mesoscale structures. These struc-
tures serve as simplified models for understanding biologicalassemblies and their functions and have applications in drugdelivery (1–5), regenerative medicine (6, 7), biosensing (8), hy-drogen production (9, 10), and clean water technologies (11). Aninteresting assembly structure is the nanoribbon, which is a highaspect ratio (10:1 or greater) bilayer. Nanoribbons are a gateway toa number of other morphologies with distinct functionalities. Forexample, nanoribbons of a charged chromophore amphiphile cantransform to a scroll-like (cochleate) morphology when the solu-tion ionic strength is increased (9). These cochleates serve as ef-ficient charge-transfer agents for photocatalysts in hydrogenproduction. Cochleate formation from liposomes of negativelycharged phospholipids in the presence of multivalent cations alsoinvolves a nanoribbon intermediate (3, 12, 13). Biocompatiblephospholipid cochleates are being explored as drug-delivery agentsbecause they can trap macromolecules, such as proteins, andDNA, and provide protection against degradation due to theirmultilayer geometry. Nanoribbons have also been observed inpeptide amphiphiles (PAs), which consist of a sequence of aminoacids covalently linked to an alkyl tail (14, 15). For example, apeptide amphiphile that stimulates collagen production has beenfound to self-assemble into nanotapes with an internal bilayerstructure (16). In a PA with alternating charged and neutral amino
acids, nanoribbons were found to transform into helical ribbons asthe PA concentration was reduced (17) and into helical andtwisted nanoribbons when the amino acid sequence was permuted(18). Helical assemblies have been previously used to templatesemiconductor nanohelices (19). Despite the progress, the corre-lation between experimental conditions such as molecular design,ionic strength, pH, amphiphile concentration, and the attainednanoribbon-related morphology are not fully established. There-fore, precise control of nanoribbon-related architecture requiresfurther understanding of the delicate interplay between intermo-lecular interactions and elastic and interfacial energies.A recent theoretical study showed that for charged molecules,
tuning the range of electrostatic interactions could induce tran-sitions between different nanoribbon-related morphologies (20).Specifically, a phase diagram was deduced for a 2D lattice ofcharged points, which interacted via long-range repulsive elec-trostatic interactions and short-range attractive interactions.Planar nanoribbon to wavy ribbon with periodic undulations tohelical ribbon transitions were predicted as the range of theelectrostatic interactions is increased. This study suggests a facilemethod for accessing distinct nanoribbon architectures by vary-ing the ionic strength (μ) of the solution because the range ofelectrostatic interactions as parametrized by Debye length (λd)scales as μ−1/2. Recent experiments also attest that tuning the
Significance
Controlling the shape and internal architecture of assemblies ofamphiphiles is critical for many technologies. The structure,and thus the function, of these assemblies reconfigures in re-sponse to stimuli, via mechanisms that are often elusive. Here,we observe and explain how molecular reordering driven byvariations in electrostatic screening length induce micrometer-scale structural changes in crystalline membranes of charged,chiral molecules: The transformation of high aspect ratio, pla-nar bilayers into scroll-like cochleates by increasing the solutionsalt content is described and explained. Our study suggests thatthis transformation should be general to charged bilayers pos-sessing a spontaneous curvature.
Author contributions: S.K., M.O.d.l.C., and M.J.B. designed research; C.G., S.K., D.M.V.,H.L., and J.M.M. performed research; C.G., S.K., D.M.V., H.L., J.M.M., M.O.d.l.C., and M.J.B.analyzed data; and C.G., S.K., D.M.V., M.O.d.l.C., and M.J.B. wrote the paper.
The authors declare no competing interest.
This article is a PNAS Direct Submission.
Published under the PNAS license.
Data deposition: All the X-ray data shown in the manuscript and the SI Appendix, as wellas the code for simulating the WAXS intensity along with the data from theory calcula-tions, have been deposited at Bitbucket (https://bitbucket.org/NUaztec/gao_et_al_pnas_2019_charged_membrane/src/master/).1C.G. and S.K. contributed equally to this work.2To whom correspondence may be addressed. Email: [email protected][email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1913632116/-/DCSupplemental.
ionic strength leads to predictable changes in the nanoribbon-related assembly morphology. For example, the period of thetwists in amyloid fibril aggregates monotonically decreases withdecreasing ionic strength (21).In this study, we analyze morphological changes in charged
planar nanoribbons as a function of increasing ionic strength. Inthis regime, nanoribbon to cochleate transformations have beenobserved in phospholipids (12) and chromophore amphiphiles (9,10, 22, 23). However, the generality and the mechanistic details ofthis transition are still unknown. In particular, the correlationbetween the ionic strength induced changes in the molecularpacking and the mesoscopic morphology transformations areelusive. The principal aim of this study is to start with a nano-ribbon structure and experimentally trace the micrometer toangstrom length-scale transformations in the membranestructure as a function of ionic strength by using a combinationof cryotransmission electron microscopy (cryo-TEM), liquid-atomic force microscopy (liquid-AFM), and in situ small- andwide-angle X-ray scattering (SAXS/WAXS). The experiments arecoupled with theoretical models that qualitatively explain theobserved morphological transitions.We chose a simple peptide amphiphile (PA), C16K1, with a
single ionizable amino acid lysine (K) covalently linked to apalmitoyl (C16) alkyl tail (Fig. 1A). This PA was chosen becauseour recent study (24) on C16K2 found spherical micelle to cylin-drical micelle to a mixture of cylindrical micelle and nanoribbontransformations as the molecular charge was reduced by increasingthe solution pH. Therefore, we hypothesized that removing one ofthe charged lysines from the headgroup could yield a macroscopicstate consisting purely of nanoribbons. Second, the choice of thisPA ensures that the interheadgroup interactions are Coulombic,unlike the case of PAs with multiple amino acids, where the as-sembly is strongly modulated by intermolecular hydrogen bonding.We first describe the C16K1 assembly in the absence of added
salt. For this, L-C16K1 was dispersed in pure water at 4 mMconcentration. Unless otherwise stated, the enantiomeric form ofthe amino acid [left-handed (L)] and the PA concentration(4 mM) are the same in all of the samples. The pH of this C16K1dispersion was ∼4.6, which is much lower than the dissociationconstant pK ∼ 7.4 for C16K1 in their aggregates (SI Appendix,
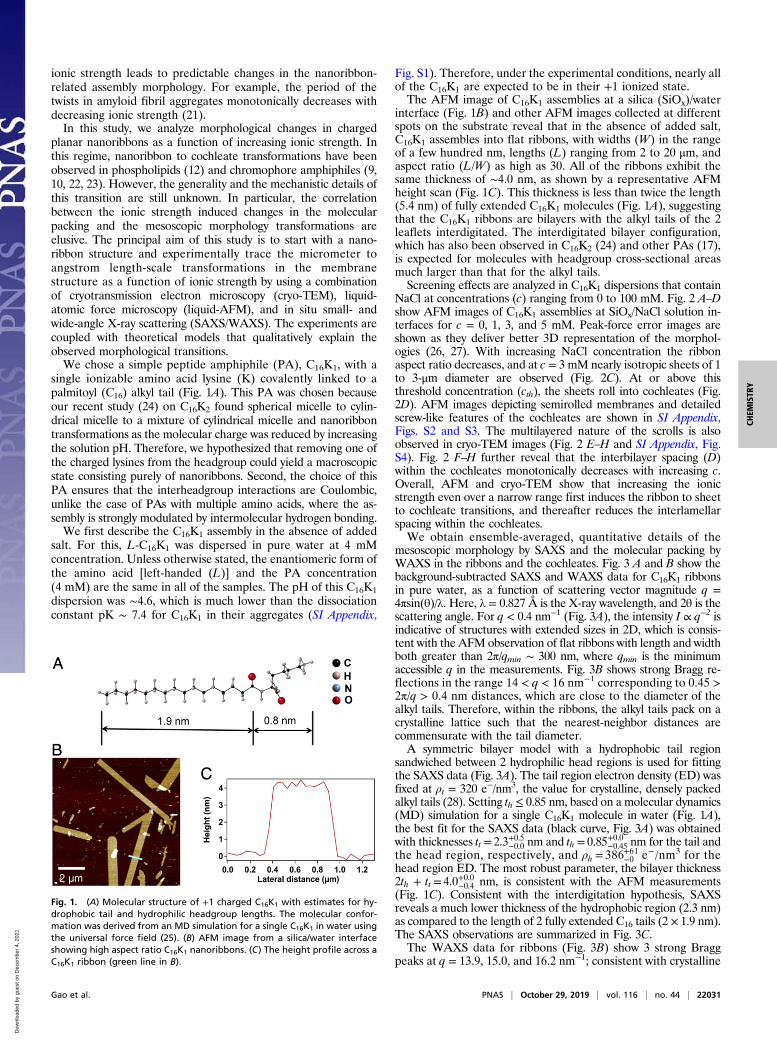
Fig. S1). Therefore, under the experimental conditions, nearly allof the C16K1 are expected to be in their +1 ionized state.The AFM image of C16K1 assemblies at a silica (SiOx)/water
interface (Fig. 1B) and other AFM images collected at differentspots on the substrate reveal that in the absence of added salt,C16K1 assembles into flat ribbons, with widths (W) in the rangeof a few hundred nm, lengths (L) ranging from 2 to 20 μm, andaspect ratio (L/W) as high as 30. All of the ribbons exhibit thesame thickness of ∼4.0 nm, as shown by a representative AFMheight scan (Fig. 1C). This thickness is less than twice the length(5.4 nm) of fully extended C16K1 molecules (Fig. 1A), suggestingthat the C16K1 ribbons are bilayers with the alkyl tails of the 2leaflets interdigitated. The interdigitated bilayer configuration,which has also been observed in C16K2 (24) and other PAs (17),is expected for molecules with headgroup cross-sectional areasmuch larger than that for the alkyl tails.Screening effects are analyzed in C16K1 dispersions that contain
NaCl at concentrations (c) ranging from 0 to 100 mM. Fig. 2 A–Dshow AFM images of C16K1 assemblies at SiOx/NaCl solution in-terfaces for c = 0, 1, 3, and 5 mM. Peak-force error images areshown as they deliver better 3D representation of the morphol-ogies (26, 27). With increasing NaCl concentration the ribbonaspect ratio decreases, and at c = 3 mM nearly isotropic sheets of 1to 3-μm diameter are observed (Fig. 2C). At or above thisthreshold concentration (cth), the sheets roll into cochleates (Fig.2D). AFM images depicting semirolled membranes and detailedscrew-like features of the cochleates are shown in SI Appendix,Figs. S2 and S3. The multilayered nature of the scrolls is alsoobserved in cryo-TEM images (Fig. 2 E–H and SI Appendix, Fig.S4). Fig. 2 F–H further reveal that the interbilayer spacing (D)within the cochleates monotonically decreases with increasing c.Overall, AFM and cryo-TEM show that increasing the ionicstrength even over a narrow range first induces the ribbon to sheetto cochleate transitions, and thereafter reduces the interlamellarspacing within the cochleates.We obtain ensemble-averaged, quantitative details of the
mesoscopic morphology by SAXS and the molecular packing byWAXS in the ribbons and the cochleates. Fig. 3 A and B show thebackground-subtracted SAXS and WAXS data for C16K1 ribbonsin pure water, as a function of scattering vector magnitude q =4πsin(θ)/λ. Here, λ = 0.827 Å is the X-ray wavelength, and 2θ is thescattering angle. For q < 0.4 nm−1 (Fig. 3A), the intensity I ∝ q−2 isindicative of structures with extended sizes in 2D, which is consis-tent with the AFM observation of flat ribbons with length and widthboth greater than 2π/qmin ∼ 300 nm, where qmin is the minimumaccessible q in the measurements. Fig. 3B shows strong Bragg re-flections in the range 14 < q < 16 nm−1 corresponding to 0.45 >2π/q > 0.4 nm distances, which are close to the diameter of thealkyl tails. Therefore, within the ribbons, the alkyl tails pack on acrystalline lattice such that the nearest-neighbor distances arecommensurate with the tail diameter.A symmetric bilayer model with a hydrophobic tail region
sandwiched between 2 hydrophilic head regions is used for fittingthe SAXS data (Fig. 3A). The tail region electron density (ED) wasfixed at ρt = 320 e−/nm3, the value for crystalline, densely packedalkyl tails (28). Setting th ≤ 0.85 nm, based on a molecular dynamics(MD) simulation for a single C16K1 molecule in water (Fig. 1A),the best fit for the SAXS data (black curve, Fig. 3A) was obtainedwith thicknesses tt = 2.3+0.5−0.0 nm and th = 0.85+0.0−0.45 nm for the tail andthe head region, respectively, and ρh = 386+61−0 e−/nm3 for thehead region ED. The most robust parameter, the bilayer thickness2th + tt = 4.0+0.0−0.4 nm, is consistent with the AFM measurements(Fig. 1C). Consistent with the interdigitation hypothesis, SAXSreveals a much lower thickness of the hydrophobic region (2.3 nm)as compared to the length of 2 fully extended C16 tails (2 × 1.9 nm).The SAXS observations are summarized in Fig. 3C.The WAXS data for ribbons (Fig. 3B) show 3 strong Bragg
peaks at q = 13.9, 15.0, and 16.2 nm−1; consistent with crystalline
Fig. 1. (A) Molecular structure of +1 charged C16K1 with estimates for hy-drophobic tail and hydrophilic headgroup lengths. The molecular confor-mation was derived from an MD simulation for a single C16K1 in water usingthe universal force field (25). (B) AFM image from a silica/water interfaceshowing high aspect ratio C16K1 nanoribbons. (C) The height profile across aC16K1 ribbon (green line in B).
Gao et al. PNAS | October 29, 2019 | vol. 116 | no. 44 | 22031
packing of the alkyl tails into an oblique unit cell with latticeconstants at = 0.49 nm, bt = 0.53 nm, and γt = 127°. To distin-guish between the tails from the 2 leaflets, the unit cell is recastwith a = 0.49 nm, b = 0.85 nm, and γ = 100° (Fig. 3D). In this unitcell, the tails at the vertices and the center belong, respectively,to the 2 opposing leaflets. By using these 2D unit cell parametersand an untilted parallelepiped to model each tail (SI Appendix,section 2), we are able to simulate the positions and the intensities
of the Bragg reflections for q > 10 nm−1 (Fig. 3B, black curve).Complementary grazing incidence X-ray scattering (GIXS)measurement on C16K1 ribbons drop-casted onto a Si substraterevealed diffraction peaks at qxy positions, which were identicalto the q positions of the diffraction peaks observed in solutionWAXS (SI Appendix, Fig. S5). Here, qxy is the component of thescattering vector in the bilayer plane. The above observationvalidates the WAXS-derived oblique lattice (Fig. 3D), and thenear-zero tilt of the tails with respect to the bilayer normal.Specifically, GIXS yields the maximum tilt of the alkyl tails ∼6°(SI Appendix, Fig. S5). Line-shape analysis of WAXS peaks (Fig.3B) reveals an average 2D crystal domain size of ∼15 nm, whichis significantly smaller than the ribbon size. This implies thatthe ribbons are polycrystalline.The SAXS/WAXS data from the ribbons also exhibit weak, but
sharp Bragg reflections for q ≤ 10 nm−1. The most prominent ofthese reflections is at q = 2.5 nm−1 (Fig. 3A). A much weakerreflection is also observed at q = 7.5 nm−1 (Fig. 3B). In fact, GIXSmeasurements of ribbons revealed qz-extended intensity rods atqxy = 2.5, 5.0, 7.5, and 10.0 nm−1 (SI Appendix, Fig. S6). Here, qz isthe scattering vector component along the bilayer normal. ThusGIXS shows that the low-q Bragg reflections originate from an in-plane ordering within the bilayer, which we attribute to a preferredorientational ordering of C16K1 headgroups. This headgroup or-dering is commensurate with the alkyl tail lattice, and can be de-fined by a unit cell that is a 1 × 3 supercell of the alkyl tail unit cellin Fig. 3D because the position of the first Bragg reflection (q1 =2.5 nm−1) equals one-third of the magnitude of the (0 1) reciprocallattice vector for the alkyl tail lattice [i.e., q1 = b*/3 = 2π /(3b sinγ)].Perhaps headgroups of neighboring C16K1 along the b axis in eachleaflet are rotated 120° clockwise (or anticlockwise) relative toeach other about the bilayer normal. However, proving this hy-pothesis is beyond the scope of the current work. Overall, theSAXS/WAXS analysis of C16K1 ribbons in pure water proves thatribbons are bilayers with interdigitated leaflets and that thepacking of alkyl tails and headgroups exhibits crystalline ordering.
Fig. 2. (A–D) AFM peakforce error images of drop-cast C16K1 membranes at SiOx/NaCl solution interface. As NaCl concentration increases, structuraltransformations are observed from nanoribbon to isotropic sheet and to rolled-up cochleates, which exhibit a screw-like pitch. (E–H) Cryogenic TEM images ofcochleates exhibiting scroll morphology and the internal multilayer features. The interbilayer spacing D within the cochleate structure decreases as NaClconcentration increases.
Fig. 3. (A) Background-subtracted in situ SAXS intensity profile for C16K1
nanoribbon. The solid black curve is the best fit over the range of 0.1 < q <6 nm−1 based on a symmetric bilayer model. (B) Background subtracted insitu WAXS intensity profile of C16K1 nanoribbon shows diffraction peaksover the range of 10 < q < 30 nm−1. The solid black curve is a simulationbased on a parallelepiped model for alkyl tails and the unit cell in D. (C) Theinterdigitated C16K1 bilayer structure and electron density profile deter-mined from analysis of SAXS data. (D) Two-dimensional unit cell and latticeparameters for alkyl tail packing, derived from WAXS.
22032 | www.pnas.org/cgi/doi/10.1073/pnas.1913632116 Gao et al.
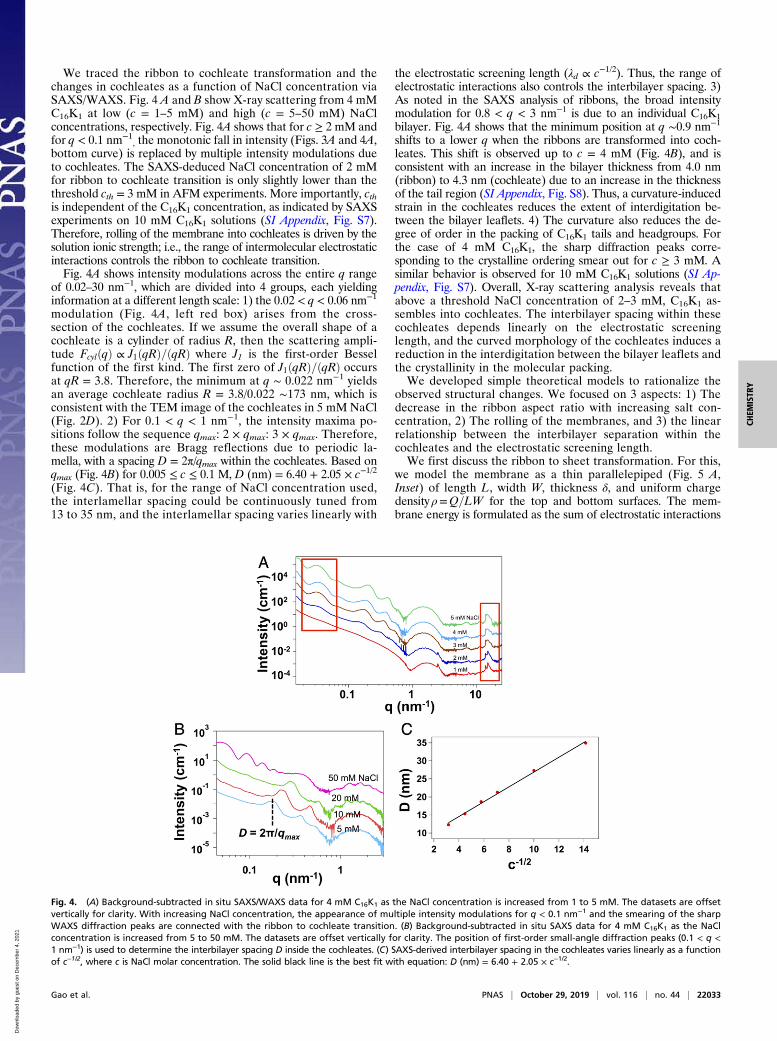
We traced the ribbon to cochleate transformation and thechanges in cochleates as a function of NaCl concentration viaSAXS/WAXS. Fig. 4 A and B show X-ray scattering from 4 mMC16K1 at low (c = 1–5 mM) and high (c = 5–50 mM) NaClconcentrations, respectively. Fig. 4A shows that for c ≥ 2 mM andfor q < 0.1 nm−1
, the monotonic fall in intensity (Figs. 3A and 4A,bottom curve) is replaced by multiple intensity modulations dueto cochleates. The SAXS-deduced NaCl concentration of 2 mMfor ribbon to cochleate transition is only slightly lower than thethreshold cth = 3 mM in AFM experiments. More importantly, cthis independent of the C16K1 concentration, as indicated by SAXSexperiments on 10 mM C16K1 solutions (SI Appendix, Fig. S7).Therefore, rolling of the membrane into cochleates is driven by thesolution ionic strength; i.e., the range of intermolecular electrostaticinteractions controls the ribbon to cochleate transition.Fig. 4A shows intensity modulations across the entire q range
of 0.02–30 nm−1, which are divided into 4 groups, each yieldinginformation at a different length scale: 1) the 0.02 < q < 0.06 nm−1
modulation (Fig. 4A, left red box) arises from the cross-section of the cochleates. If we assume the overall shape of acochleate is a cylinder of radius R, then the scattering ampli-tude FcylðqÞ ∝ J1ðqRÞ=ðqRÞ where J1 is the first-order Besselfunction of the first kind. The first zero of J1ðqRÞ=ðqRÞ occursat qR = 3.8. Therefore, the minimum at q ∼ 0.022 nm−1 yieldsan average cochleate radius R = 3.8/0.022 ∼173 nm, which isconsistent with the TEM image of the cochleates in 5 mM NaCl(Fig. 2D). 2) For 0.1 < q < 1 nm−1, the intensity maxima po-sitions follow the sequence qmax: 2 × qmax: 3 × qmax. Therefore,these modulations are Bragg reflections due to periodic la-mella, with a spacing D = 2π/qmax within the cochleates. Based onqmax (Fig. 4B) for 0.005 ≤ c ≤ 0.1 M, D (nm) = 6.40 + 2.05 × c−1/2
(Fig. 4C). That is, for the range of NaCl concentration used,the interlamellar spacing could be continuously tuned from13 to 35 nm, and the interlamellar spacing varies linearly with
the electrostatic screening length (λd ∝ c−1/2). Thus, the range ofelectrostatic interactions also controls the interbilayer spacing. 3)As noted in the SAXS analysis of ribbons, the broad intensitymodulation for 0.8 < q < 3 nm−1 is due to an individual C16K1bilayer. Fig. 4A shows that the minimum position at q ∼0.9 nm−1
shifts to a lower q when the ribbons are transformed into coch-leates. This shift is observed up to c = 4 mM (Fig. 4B), and isconsistent with an increase in the bilayer thickness from 4.0 nm(ribbon) to 4.3 nm (cochleate) due to an increase in the thicknessof the tail region (SI Appendix, Fig. S8). Thus, a curvature-inducedstrain in the cochleates reduces the extent of interdigitation be-tween the bilayer leaflets. 4) The curvature also reduces the de-gree of order in the packing of C16K1 tails and headgroups. Forthe case of 4 mM C16K1, the sharp diffraction peaks corre-sponding to the crystalline ordering smear out for c ≥ 3 mM. Asimilar behavior is observed for 10 mM C16K1 solutions (SI Ap-pendix, Fig. S7). Overall, X-ray scattering analysis reveals thatabove a threshold NaCl concentration of 2–3 mM, C16K1 as-sembles into cochleates. The interbilayer spacing within thesecochleates depends linearly on the electrostatic screeninglength, and the curved morphology of the cochleates induces areduction in the interdigitation between the bilayer leaflets andthe crystallinity in the molecular packing.We developed simple theoretical models to rationalize the
observed structural changes. We focused on 3 aspects: 1) Thedecrease in the ribbon aspect ratio with increasing salt con-centration, 2) The rolling of the membranes, and 3) the linearrelationship between the interbilayer separation within thecochleates and the electrostatic screening length.We first discuss the ribbon to sheet transformation. For this,
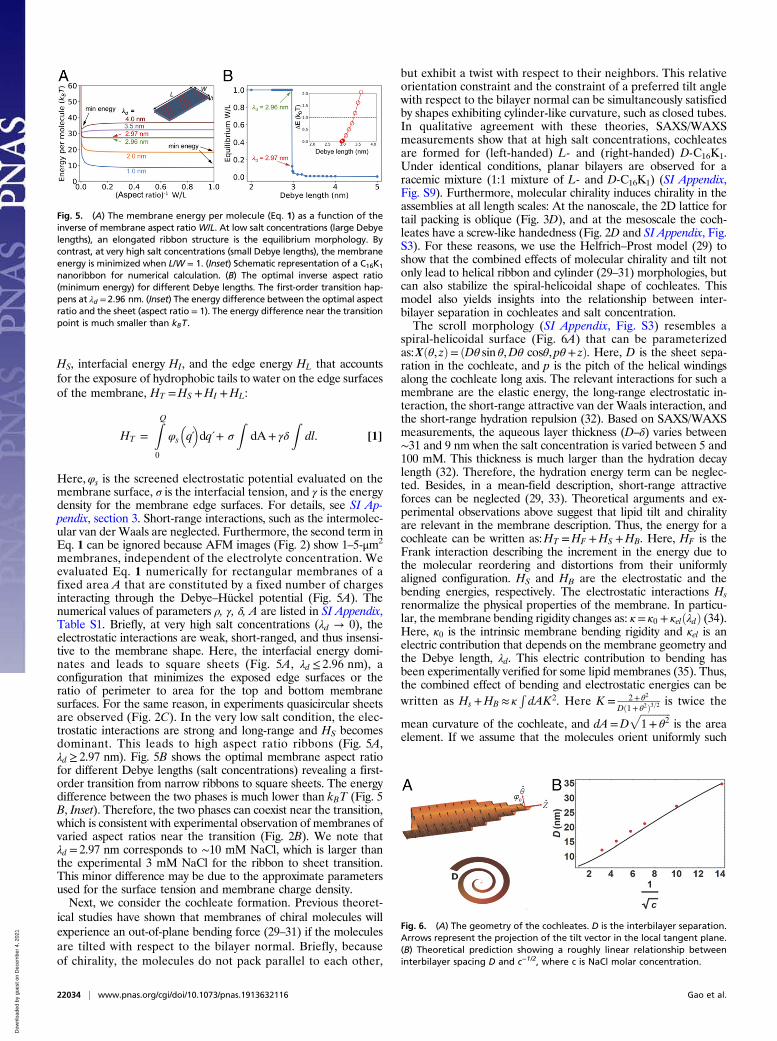
we model the membrane as a thin parallelepiped (Fig. 5 A,Inset) of length L, width W, thickness δ, and uniform chargedensity ρ=Q=LW for the top and bottom surfaces. The mem-brane energy is formulated as the sum of electrostatic interactions
Fig. 4. (A) Background-subtracted in situ SAXS/WAXS data for 4 mM C16K1 as the NaCl concentration is increased from 1 to 5 mM. The datasets are offsetvertically for clarity. With increasing NaCl concentration, the appearance of multiple intensity modulations for q < 0.1 nm−1 and the smearing of the sharpWAXS diffraction peaks are connected with the ribbon to cochleate transition. (B) Background-subtracted in situ SAXS data for 4 mM C16K1 as the NaClconcentration is increased from 5 to 50 mM. The datasets are offset vertically for clarity. The position of first-order small-angle diffraction peaks (0.1 < q <1 nm−1) is used to determine the interbilayer spacing D inside the cochleates. (C) SAXS-derived interbilayer spacing in the cochleates varies linearly as a functionof c−1/2, where c is NaCl molar concentration. The solid black line is the best fit with equation: D (nm) = 6.40 + 2.05 × c−1/2.
Gao et al. PNAS | October 29, 2019 | vol. 116 | no. 44 | 22033
HS, interfacial energy HI, and the edge energy HL that accountsfor the exposure of hydrophobic tails to water on the edge surfacesof the membrane, HT =HS +HI +HL:
HT =ZQ
0
φs
�q��dq�+ σ
ZdA+ γδ
Zdl. [1]
Here,φs is the screened electrostatic potential evaluated on themembrane surface, σ is the interfacial tension, and γ is the energydensity for the membrane edge surfaces. For details, see SI Ap-pendix, section 3. Short-range interactions, such as the intermolec-ular van der Waals are neglected. Furthermore, the second term inEq. 1 can be ignored because AFM images (Fig. 2) show 1–5-μm2
membranes, independent of the electrolyte concentration. Weevaluated Eq. 1 numerically for rectangular membranes of afixed area A that are constituted by a fixed number of chargesinteracting through the Debye–Hückel potential (Fig. 5A). Thenumerical values of parameters ρ, γ, δ, A are listed in SI Appendix,Table S1. Briefly, at very high salt concentrations (λd → 0), theelectrostatic interactions are weak, short-ranged, and thus insensi-tive to the membrane shape. Here, the interfacial energy domi-nates and leads to square sheets (Fig. 5A, λd ≤ 2.96 nm), aconfiguration that minimizes the exposed edge surfaces or theratio of perimeter to area for the top and bottom membranesurfaces. For the same reason, in experiments quasicircular sheetsare observed (Fig. 2C). In the very low salt condition, the elec-trostatic interactions are strong and long-range and HS becomesdominant. This leads to high aspect ratio ribbons (Fig. 5A,λd ≥ 2.97 nm). Fig. 5B shows the optimal membrane aspect ratiofor different Debye lengths (salt concentrations) revealing a first-order transition from narrow ribbons to square sheets. The energydifference between the two phases is much lower than kBT (Fig. 5B, Inset). Therefore, the two phases can coexist near the transition,which is consistent with experimental observation of membranes ofvaried aspect ratios near the transition (Fig. 2B). We note thatλd = 2.97 nm corresponds to ∼10 mM NaCl, which is larger thanthe experimental 3 mM NaCl for the ribbon to sheet transition.This minor difference may be due to the approximate parametersused for the surface tension and membrane charge density.Next, we consider the cochleate formation. Previous theoret-
ical studies have shown that membranes of chiral molecules willexperience an out-of-plane bending force (29–31) if the moleculesare tilted with respect to the bilayer normal. Briefly, becauseof chirality, the molecules do not pack parallel to each other,
but exhibit a twist with respect to their neighbors. This relativeorientation constraint and the constraint of a preferred tilt anglewith respect to the bilayer normal can be simultaneously satisfiedby shapes exhibiting cylinder-like curvature, such as closed tubes.In qualitative agreement with these theories, SAXS/WAXSmeasurements show that at high salt concentrations, cochleatesare formed for (left-handed) L- and (right-handed) D-C16K1.Under identical conditions, planar bilayers are observed for aracemic mixture (1:1 mixture of L- and D-C16K1) (SI Appendix,Fig. S9). Furthermore, molecular chirality induces chirality in theassemblies at all length scales: At the nanoscale, the 2D lattice fortail packing is oblique (Fig. 3D), and at the mesoscale the coch-leates have a screw-like handedness (Fig. 2D and SI Appendix, Fig.S3). For these reasons, we use the Helfrich–Prost model (29) toshow that the combined effects of molecular chirality and tilt notonly lead to helical ribbon and cylinder (29–31) morphologies, butcan also stabilize the spiral-helicoidal shape of cochleates. Thismodel also yields insights into the relationship between inter-bilayer separation in cochleates and salt concentration.The scroll morphology (SI Appendix, Fig. S3) resembles a
spiral-helicoidal surface (Fig. 6A) that can be parameterizedas:Xðθ, zÞ= ðDθ sin θ,Dθ cosθ, pθ+ zÞ. Here, D is the sheet sepa-ration in the cochleate, and p is the pitch of the helical windingsalong the cochleate long axis. The relevant interactions for such amembrane are the elastic energy, the long-range electrostatic in-teraction, the short-range attractive van der Waals interaction, andthe short-range hydration repulsion (32). Based on SAXS/WAXSmeasurements, the aqueous layer thickness (D–δ) varies between∼31 and 9 nm when the salt concentration is varied between 5 and100 mM. This thickness is much larger than the hydration decaylength (32). Therefore, the hydration energy term can be neglec-ted. Besides, in a mean-field description, short-range attractiveforces can be neglected (29, 33). Theoretical arguments and ex-perimental observations above suggest that lipid tilt and chiralityare relevant in the membrane description. Thus, the energy for acochleate can be written as:HT =HF +HS +HB. Here, HF is theFrank interaction describing the increment in the energy due tothe molecular reordering and distortions from their uniformlyaligned configuration. HS and HB are the electrostatic and thebending energies, respectively. The electrostatic interactions Hsrenormalize the physical properties of the membrane. In particu-lar, the membrane bending rigidity changes as: κ= κ0 + κelðλdÞ (34).Here, κ0 is the intrinsic membrane bending rigidity and κel is anelectric contribution that depends on the membrane geometry andthe Debye length, λd. This electric contribution to bending hasbeen experimentally verified for some lipid membranes (35). Thus,the combined effect of bending and electrostatic energies can bewritten as Hs +HB ≈ κ
RdAK2. Here K = 2+ θ2
Dð1+ θ2Þ3=2 is twice the
mean curvature of the cochleate, and dA=Dffiffiffiffiffiffiffiffiffiffiffiffi1+ θ2
pis the area
element. If we assume that the molecules orient uniformly such
Fig. 5. (A) The membrane energy per molecule (Eq. 1) as a function of theinverse of membrane aspect ratioW/L. At low salt concentrations (large Debyelengths), an elongated ribbon structure is the equilibrium morphology. Bycontrast, at very high salt concentrations (small Debye lengths), the membraneenergy is minimized when L/W = 1. (Inset) Schematic representation of a C16K1
nanoribbon for numerical calculation. (B) The optimal inverse aspect ratio(minimum energy) for different Debye lengths. The first-order transition hap-pens at λd = 2.96 nm. (Inset) The energy difference between the optimal aspectratio and the sheet (aspect ratio = 1). The energy difference near the transitionpoint is much smaller than kBT.
Fig. 6. (A) The geometry of the cochleates. D is the interbilayer separation.Arrows represent the projection of the tilt vector in the local tangent plane.(B) Theoretical prediction showing a roughly linear relationship betweeninterbilayer spacing D and c−1/2, where c is NaCl molar concentration.
22034 | www.pnas.org/cgi/doi/10.1073/pnas.1913632116 Gao et al.
that the tilt projection m in the local tangent plane forms an angleφ0 with the azimuthal direction, then m= cosφ0θ+ sinφ0z, whereθ, z are the unit vectors in the azimuthal and axial directions (Fig.6A). Therefore, the Frank energy takes the simple form
HF
W=κ′2K2 cos2φ0 − λHP K sinφ0 cosφ0. [2]
Here, κ′ is the difference between elastic constants for bend-ing the membrane in the parallel and perpendicular directionsto the tilt vector, and λHP measures the strength of the inter-molecular chiral interactions (29, 30). The minimization of thetotal energy HT with respect to φ0 predicts a critical tilt anglesec 2φ0 =−ð2κ+ κ′Þ=κ′. If the energetic costs of bending the mem-brane parallel or perpendicular to the tilt direction are almost thesame, (i.e., κ′≈ 0), then φ0 ≈ 45°. This is roughly equal to the angleof the helical windings with the cochleate’s principal axis (SI Ap-pendix, Fig. S3) suggesting that molecular tilt direction coincideswith the membrane folding direction. This hypothesis needs fur-ther exploration. While the molecular tilt orientation is relatedonly to the membrane elastic parameters, the interbilayer separa-tion D depends on κ/λHP, i.e., the ratio of the membrane bendingrigidity and the molecular chiral interaction parameter (SI Appen-dix, Eq. S16). In particular, D decreases nearly linearly with theDebye length for R � k=λHP, where R is the cochleate externalradius. Furthermore, a theoretical curve that approximately repro-duces the D vs. λd experimental data (Fig. 6B) yields κ0/λHP ∼200 m. (See SI Appendix, section 3 for details.) The slight quanti-tative deviation between the experiment and theory (Fig. 6B) islikely due to neglecting the stretching degrees of freedom and thethickness of the membrane. Nevertheless, the accuracy of thequalitative predictions of the model clearly highlights the collec-tive effect of molecular tilt and chirality in inducing the spontane-ous membrane curvature. This combined with the electrostaticeffects, which rigidifies the membrane, enable us to deduce qual-itatively the key structural features of the cochleates. In particular,this simplified theoretical model suggests that the linear relation-ship between the interbilayer separation and the electrostaticscreening length is not a result of system-specific design, but ofthe interplay between electrostatic energy and the membrane in-ternal degrees of freedom. Therefore, it is not surprising thatsimilar linear relationships have been observed in other chargedlayered systems, such as clay mineral montmorillonite (36, 37).We note that the linear relationship is not valid in the presencemultivalent ions. For example, negatively charged phospholipidcochleates show little or no dependence of interbilayer spacingon the CaCl2 concentration (38). It is possible that the multi-valent cations are tightly bound to the molecules resulting ininterbilayer electrostatic interactions that cannot be parame-terized by the screening length λd alone. By contrast, the use ofmonovalent salts to induce the C16K1 cochleate structure leadsto tunable interbilayer spacing over ∼10–40 nm. This structuralfeature may have application for controlled encapsulation andrelease of drug particles within a specific size range.Finally, we note that while the experimental results for
cochleates are in qualitative agreement with the predictions oftheoretical models for assembly of chiral molecules, there are stillunresolved questions. First, these continuum models (29–31) arestrictly applicable to fluid-like membranes or membranes withhexatic order. That is for cases where there are no long-rangeintermolecular positional correlations. However, the WAXS data
from L- and D-C16K1 ribbons clearly show sharp diffraction peaksindicating crystalline bilayers (Fig. 3C and SI Appendix, Figs. S5and S9). Second, membrane curvature is expected for cases wherethe molecules are tilted with respect to the bilayer normal. OurWAXS and GIXS data are currently inconclusive in this regard: Inthe planar ribbon phase, the molecules may have a slight (<6°) tilt.Whether, the molecules in bilayer have a small tilt or the mole-cules undergo a tilting transition just prior to the sheet to cochleatetransformation will be a subject of future studies.
ConclusionsWe designed a peptide amphiphile C16K1 to investigate theelectrolyte-induced transformation of planar bilayers to scroll-like cochleates. We show that with the addition of NaCl, thehigh aspect ratio C16K1 ribbons formed in zero salt conditionstransform to isotropic sheets, prior to rolling up to form coch-leates. This ribbon to cochleate transformation also induces areduction in the crystallinity in the molecular packing. Furtheraddition of salt reduces, within the cochleates, the interbilayerseparation, which scales linearly with the Debye length. A sim-plified model demonstrates that the ribbon to sheet trans-formation is a first-order transition induced by the reduction inthe range of electrostatic interactions. Theoretical models showthat rolling of membranes into cochleates is the combined effectof molecular chirality and tilt. The linear relationship betweenthe interbilayer separation and the screening length in cochleatesis qualitatively explained by the competition between electro-static and the effective elastic interactions that include the in-ternal degrees of freedom of tilt and chirality. These resultssuggest that the salt-induced structural transitions in the C16K1system should be observed in other charged bilayer mem-branes. Our combined experimental and theoretical studyyields insight into attaining the cochleate structures and con-trolling their internal architecture. These results should beuseful for optimizing the structure and function of cochleatesin many applications, including drug delivery and photocatalyticproduction of hydrogen.
Materials and MethodsPeptides synthesis and SAXS/WAXS, Cryo-TEM, and liquid-AFM measure-ments are described in SI Appendix.
Data AvailabilityAll the X-ray data shown in the manuscript and the SI Appendix,as well as the code for simulating the WAXS intensity along withthe data from theory calculations, have been deposited at Bitbucket(39). The files are in folders labeled with corresponding figurenumbers.
ACKNOWLEDGMENTS. This research was primarily supported by the De-partment of Energy (DOE), Office of Basic Energy Sciences under ContractDE-FG02-08ER46539. Peptide synthesis was performed in the PeptideSynthesis Core Facility of the Simpson Querrey Institute at NorthwesternUniversity. The SAXS/WAXS experiments were performed at the DuPont-Northwestern-Dow Collaborative Access Team (DND-CAT) located at Sector 5of the Advanced Photon Source (APS) and at APS Sector 12. The APS, anOffice of Science User Facility operated for DOE by Argonne NationalLaboratory, is supported by DOE under Contract DE-AC02-06CH11357. GIXSwas performed at the XRD Facility and TEM used the EPIC facility atNorthwestern University. The authors thank M. Karver for peptide synthesis,Dr. Liam Palmer for discussions and for suggesting cryo-TEM, and Drs. S. Weigand(DND-CAT) and B. Lee (APS, sector 12) for the assistance with the X-rayscattering measurements.
1. T. Bramer, N. Dew, K. Edsman, Pharmaceutical applications for catanionic mixtures.
J. Pharm. Pharmacol. 59, 1319–1334 (2007).2. V. P. Torchilin, Recent advances with liposomes as pharmaceutical carriers. Nat. Rev.
Drug Discov. 4, 145–160 (2005).3. A. Pawar, C. Bothiraja, K. Shaikh, A. Mali, An insight into cochleates, a potential drug
delivery system. Rsc Adv. 5, 81188–81202 (2015).
4. A. Rosler, G. W. M. Vandermeulen, H. A. Klok, Advanced drug delivery devices via self-
assembly of amphiphilic block copolymers. Adv. Drug Deliv. Rev. 64, 270–279 (2012).5. M. Goldberg, R. Langer, X. Jia, Nanostructured materials for applications in drug
delivery and tissue engineering. J. Biomater. Sci. Polym. Ed. 18, 241–268 (2007).6. J. B. Matson, S. I. Stupp, Self-assembling peptide scaffolds for regenerative medicine.
Chem. Commun. (Camb.) 48, 26–33 (2012).
Gao et al. PNAS | October 29, 2019 | vol. 116 | no. 44 | 22035
7. H. Hosseinkhani, P. D. Hong, D. S. Yu, Self-assembled proteins and peptides for re-generative medicine. Chem. Rev. 113, 4837–4861 (2013).
8. M. Yemini, M. Reches, J. Rishpon, E. Gazit, Novel electrochemical biosensing platformusing self-assembled peptide nanotubes. Nano Lett. 5, 183–186 (2005).
9. R. V. Kazantsev et al., Crystal-phase transitions and photocatalysis in supramolecularscaffolds. J. Am. Chem. Soc. 139, 6120–6127 (2017).
10. A. S. Weingarten et al., Self-assembling hydrogel scaffolds for photocatalytic hydro-gen production. Nat. Chem. 6, 964–970 (2014).
11. Y. Kaufman, A. Berman, V. Freger, Supported lipid bilayer membranes for waterpurification by reverse osmosis. Langmuir 26, 7388–7395 (2010).
12. K. Nagarsekar et al., Understanding cochleate formation: Insights into structuraldevelopment. Soft Matter 12, 3797–3809 (2016).
13. D. Papahadjopoulos,W. J. Vail, K. Jacobson, G. Poste, Cochleate lipid cylinders: Formationby fusion of unilamellar lipid vesicles. Biochim. Biophys. Acta 394, 483–491 (1975).
14. I. W. Hamley, Self-assembly of amphiphilic peptides. Soft Matter 7, 4122–4138 (2011).15. H. Cui, M. J. Webber, S. I. Stupp, Self-assembly of peptide amphiphiles: From mole-
cules to nanostructures to biomaterials. Biopolymers 94, 1–18 (2010).16. V. Castelletto, I. W. Hamley, J. Perez, L. Abezgauz, D. Danino, Fibrillar superstructure
from extended nanotapes formed by a collagen-stimulating peptide. Chem. Com-mun. (Camb.) 46, 9185–9187 (2010).
17. H. Cui, T. Muraoka, A. G. Cheetham, S. I. Stupp, Self-assembly of giant peptidenanobelts. Nano Lett. 9, 945–951 (2009).
18. H. Cui, A. G. Cheetham, E. T. Pashuck, S. I. Stupp, Amino acid sequence in constitu-tionally isomeric tetrapeptide amphiphiles dictates architecture of one-dimensionalnanostructures. J. Am. Chem. Soc. 136, 12461–12468 (2014).
19. E. D. Sone, E. R. Zubarev, S. I. Stupp, Semiconductor nanohelices templated by su-pramolecular ribbons. Angew. Chem. Int. Ed. Engl. 41, 1706–1709 (2002).
20. Z. Yao, M. Olvera de la Cruz, Electrostatics-driven hierarchical buckling of chargedflexible ribbons. Phys. Rev. Lett. 116, 148101 (2016).
21. J. Adamcik, R. Mezzenga, Adjustable twisting periodic pitch of amyloid fibrils. SoftMatter 7, 5437–5443 (2011).
22. A. S. Weingarten et al., Supramolecular packing controls H2 photocatalysis in chro-mophore amphiphile hydrogels. J. Am. Chem. Soc. 137, 15241–15246 (2015).
23. J. A. Lehrman, H. Cui, W. W. Tsai, T. J. Moyer, S. I. Stupp, Supramolecular control ofself-assembling terthiophene-peptide conjugates through the amino acid side chain.Chem. Commun. (Camb.) 48, 9711–9713 (2012).
24. C. Gao et al., Electrostatic control of polymorphism in charged amphiphile assemblies.J. Phys. Chem. B 121, 1623–1628 (2017).
25. A. K. Rappe, C. J. Casewit, K. S. Colwell, W. A. Goddard, W. M. Skiff, Uff, a fullperiodic-table force-field for molecular mechanics and molecular-dynamics simula-tions. J. Am. Chem. Soc. 114, 10024–10035 (1992).
26. J. M. Wallace, Applications of atomic force microscopy for the assessment of nano-scale morphological and mechanical properties of bone. Bone 50, 420–427 (2012).
27. Q. Y. Huang, H. Y. Wu, P. Cai, J. B. Fein, W. L. Chen, Atomic force microscopy mea-surements of bacterial adhesion and biofilm formation onto clay-sized particles. Sci.Rep. 5, 16857 (2015).
28. J. F. Nagle, S. Tristram-Nagle, Structure of lipid bilayers. Biochim. Biophys. Acta 1469,159–195 (2000).
29. W. Helfrich, J. Prost, Intrinsic bending force in anisotropic membranes made of chiralmolecules. Phys. Rev. A Gen. Phys. 38, 3065–3068 (1988).
30. J. V. Selinger, J. M. Schnur, Theory of chiral lipid tubules. Phys. Rev. Lett. 71, 4091–4094 (1993).
31. J. V. Selinger, M. S. Spector, J. M. Schnur, Theory of self-assembled tubules and helicalribbons. J. Phys. Chem. B 105, 7157–7169 (2001).
32. S. Komura, H. Shirotori, T. Kato, Phase behavior of charged lipid bilayer membraneswith added electrolyte. J. Chem. Phys. 119, 1157–1164 (2003).
33. A. Onuki, Surface tension of electrolytes: Hydrophilic and hydrophobic ions near aninterface. J. Chem. Phys. 128, 224704 (2008).
34. J. L. Harden, C. Marques, J. F. Joanny, D. Andelman, Membrane curvature elasticity inweakly charged lamellar phases. Langmuir 8, 1170–1175 (1992).
35. A. C. Rowat, P. L. Hansen, J. H. Ipsen, Experimental evidence of the electrostaticcontribution to membrane bending rigidity. Europhys. Lett. 67, 144–149 (2004).
36. K. Norrish, The swelling of montmorillonite. Discuss. Faraday Soc. 18, 120–134 (1954).37. C. C. Tester, S. Aloni, B. Gilbert, J. F. Banfield, Short- and long-range attractive forces
that influence the structure of montmorillonite Osmotic hydrates. Langmuir 32,12039–12046 (2016).
38. K. Nagarsekar et al., Electron microscopy and theoretical modeling of cochleates.Langmuir 30, 13143–13151 (2014).
39. C. Gao et al., Data from “Gao_et_al_PNAS_2019_charged_membrane.” Bitbucket.https://bitbucket.org/NUaztec/gao_et_al_pnas_2019_charged_membrane/src/master/.Deposited 3 October 2019.
22036 | www.pnas.org/cgi/doi/10.1073/pnas.1913632116 Gao et al.