DEVELOPMENTAL BIOLOGY 136.264-272 (1989) Endo16, a Lineage-Specific Protein of the Sea Urchin Embryo, Is First Expressed Just prior to Gastrulation CATHERINENOCENTE-MCGRATH,CAROL A. BRENNER,'AND SUSANG.ERNST Department of Biology, Tufts University, Me&d, Massachusetts 02155 Accepted July 6, 1989 We have isolated and characterized a new endoderm-specific gene, designated Endol6, from a sea urchin gastrula stage cDNA library. Northern blot analysis and in situ hybridization experiments indicate that this gene is first expressed in the vegetal plate, a group of endodermal and mesenchymal precursor cells that are poised to invaginate in the first movement of gastrulation. Expression becomes progressively restricted to a subset of endodermal cells as development proceeds. To study the End016 gene product, a polyclonal antiserum was raised against bacterially expressed End016 protein. Indirect immunofluorescence experiments in midgastrula stage embryos reveal that the End016 protein is localized to the surface of endoderm and secondary mesenchyme cells. In Western blot experiments, the antiserum detects a small set of high molecular weight proteins ranging from 180 to >300 kDa. Analysis of the nucleotide-derived amino acid sequence from a partial End016 cDNA clone reveals a highly repetitive, extremely acidic protein segment that includes the Arg-Gly-Asp (RGD) tripeptide known to be important in cell binding domains of a number of extracellular proteins. Taken together, these data suggest that the End016 protein may be an adhesion molecule involved in gastrulation of the sea urchin embryo. o 1989 Academic PRSS, IX INTRODUCTION The use of cloned gene probes and monoclonal anti- bodies (McAb) has facilitated the identification of an expanding set of lineage-specific molecules in the sea urchin embryo (reviewed in Davidson, 1986; McClay and Wessel, 1985). Knowledge of how expression of these molecules is regulated as well as what role the gene products play in development is crucial to understand- ing how this complex multicellular organism arises from a single cell, the fertilized egg. In mesenchyme blastula stage sea urchin embryos five distinct lineages can be identified on the basis of morphological criteria and lineage-specific markers: the oral and aboral ecto- derm, the primary mesenchyme, the small micromeres, and the vegetal plate, which later gives rise to endo- derm and secondary mesenchyme. It is the vegetal plate cells that perform the morphogenetic movements that accomplish gastrulation in these embryos. These move- ments are accompanied by changes in cell shape, pulsa- tile behavior, filopodial activity, and extensive cell re- packing (reviewed by Keller and Hardin, 1987). In a detailed analysis of sea urchin morphogenesis, Gustaf- son and Wolpert (1967) proposed that all of the cell rearrangements which occur in the course of embryo- genesis could be explained in terms of dynamic changes in cell adhesions and cell migrations. Since the time of their proposal, a growing body of evidence has been Sequence data from this article have been deposited with the EMBL/GenBank/DDBJ Data Libraries under Accession No. M27296. 1 Present address: Genelabs, Inc., Redwood City, CA 94063. accumulating in support of the role of cell adhesion in sea urchin morphogenesis. Recently, several lines of ex- perimental evidence implicate cell-cell and cell-matrix adhesions in sea urchin gastrulation. Inhibitors of N- linked glycosylation (Heifetz and Lennarz, 1979), pro- teoglycan synthesis (Karp and Solursh, 1974;’ Katow and Solursh, 1979), and collagen synthesis or processing (Butler et aZ., 1987; Wessel and McClay, 1987), as well as McAb fragments directed against the apical extracellu- lar matrix protein hyalin (Adelson and Humphries, 1988), are all capable of preventing gastrulation. How- ever, little is known about specific molecules expressed by vegetal plate cells and their descendants that partic- ipate in adhesive interactions involved in gastrulation. In this report we describe the isolation and character- ization of a new endoderm-specific gene designated Endol6. Using in situ hybridization and indirect immu- nofluorescence, we have investigated the spatial and temporal expressions of the End016 transcript as well as localization of the End016 gene product. The results of these investigations, along with analysis of the nu- cleotide-derived amino acid sequence of the End016 cDNA clone, lead us to propose that the End016 protein may play a role in cell adhesion and gastrulation in the sea urchin embryo. MATERIALS AND METHODS Animals. Adult Strongylocentrotus purpuratus were purchased from Alacrity Marine Biological Services (Redondo Beach, CA). Maintenance of animals, spawn- 0012-1606/89$3.00 Copyright 0 1989 by Academic Press, Inc. All rights of reproduction in any form reserved. 264
Transcript
DEVELOPMENTAL BIOLOGY 136.264-272 (1989)
Endo16, a Lineage-Specific Protein of the Sea Urchin Embryo, Is First Expressed Just prior to Gastrulation
CATHERINENOCENTE-MCGRATH,CAROL A. BRENNER,'AND SUSANG.ERNST
Department of Biology, Tufts University, Me&d, Massachusetts 02155
Accepted July 6, 1989
We have isolated and characterized a new endoderm-specific gene, designated Endol6, from a sea urchin gastrula stage cDNA library. Northern blot analysis and in situ hybridization experiments indicate that this gene is first expressed in the vegetal plate, a group of endodermal and mesenchymal precursor cells that are poised to invaginate in the first movement of gastrulation. Expression becomes progressively restricted to a subset of endodermal cells as development proceeds. To study the End016 gene product, a polyclonal antiserum was raised against bacterially expressed End016 protein. Indirect immunofluorescence experiments in midgastrula stage embryos reveal that the End016 protein is localized to the surface of endoderm and secondary mesenchyme cells. In Western blot experiments, the antiserum detects a small set of high molecular weight proteins ranging from 180 to >300 kDa. Analysis of the nucleotide-derived amino acid sequence from a partial End016 cDNA clone reveals a highly repetitive, extremely acidic protein segment that includes the Arg-Gly-Asp (RGD) tripeptide known to be important in cell binding domains of a number of extracellular proteins. Taken together, these data suggest that the End016 protein may be an adhesion molecule involved in gastrulation of the sea urchin embryo. o 1989 Academic PRSS, IX
INTRODUCTION
The use of cloned gene probes and monoclonal anti- bodies (McAb) has facilitated the identification of an expanding set of lineage-specific molecules in the sea urchin embryo (reviewed in Davidson, 1986; McClay and Wessel, 1985). Knowledge of how expression of these molecules is regulated as well as what role the gene products play in development is crucial to understand- ing how this complex multicellular organism arises from a single cell, the fertilized egg. In mesenchyme blastula stage sea urchin embryos five distinct lineages can be identified on the basis of morphological criteria and lineage-specific markers: the oral and aboral ecto- derm, the primary mesenchyme, the small micromeres, and the vegetal plate, which later gives rise to endo- derm and secondary mesenchyme. It is the vegetal plate cells that perform the morphogenetic movements that accomplish gastrulation in these embryos. These move- ments are accompanied by changes in cell shape, pulsa- tile behavior, filopodial activity, and extensive cell re- packing (reviewed by Keller and Hardin, 1987). In a detailed analysis of sea urchin morphogenesis, Gustaf- son and Wolpert (1967) proposed that all of the cell rearrangements which occur in the course of embryo- genesis could be explained in terms of dynamic changes in cell adhesions and cell migrations. Since the time of their proposal, a growing body of evidence has been
Sequence data from this article have been deposited with the EMBL/GenBank/DDBJ Data Libraries under Accession No. M27296.
1 Present address: Genelabs, Inc., Redwood City, CA 94063.
accumulating in support of the role of cell adhesion in sea urchin morphogenesis. Recently, several lines of ex- perimental evidence implicate cell-cell and cell-matrix adhesions in sea urchin gastrulation. Inhibitors of N- linked glycosylation (Heifetz and Lennarz, 1979), pro- teoglycan synthesis (Karp and Solursh, 1974;’ Katow and Solursh, 1979), and collagen synthesis or processing (Butler et aZ., 1987; Wessel and McClay, 1987), as well as McAb fragments directed against the apical extracellu- lar matrix protein hyalin (Adelson and Humphries, 1988), are all capable of preventing gastrulation. How- ever, little is known about specific molecules expressed by vegetal plate cells and their descendants that partic- ipate in adhesive interactions involved in gastrulation.
In this report we describe the isolation and character- ization of a new endoderm-specific gene designated Endol6. Using in situ hybridization and indirect immu- nofluorescence, we have investigated the spatial and temporal expressions of the End016 transcript as well as localization of the End016 gene product. The results of these investigations, along with analysis of the nu- cleotide-derived amino acid sequence of the End016 cDNA clone, lead us to propose that the End016 protein may play a role in cell adhesion and gastrulation in the sea urchin embryo.
MATERIALS AND METHODS
Animals. Adult Strongylocentrotus purpuratus were purchased from Alacrity Marine Biological Services (Redondo Beach, CA). Maintenance of animals, spawn-
0012-1606/89$3.00 Copyright 0 1989 by Academic Press, Inc. All rights of reproduction in any form reserved.
264
NOCENTE-MCGRATH, BRENNER, AND ERNST Sea Urchin Lineage-Spe&ic Protein 265
ing, fertilization, and embryo culture were as previously described (Pittman and Ernst, 1984).
Isolation and characterization of SpGl6 cDNA clone. All enzymes were purchased from Promega Biotech or New England Biolabs except as indicated. All isotopes were purchased from Amersham. Standard molecular biology techniques were used (Maniatis et al, 1982). The 1.3-kb HindIII-BamHI fragment of the Caenorhabditis elegans ~01-1 gene (Kramer et al., 1982) (~01-1 gene, gift of J. Kramer and D. Hirsch; subclone, gift of Dr. H. Boedtker) was nick-translated and used to screen a S. purpuratus gastrula stage XgtlO cDNA library (gift of T. Thomas and E. Davidson) containing 10,000 recombi- nant clones. Hybridization conditions were 5X SSC, 5X Denhardt’s, 0.1% SDS at 55°C. Washing was performed under the same conditions. Positive clones were plaque-purified and phage DNA was isolated and di- gested with EcoRI to determine insert size. One clone, XSpG16, contained the largest cDNA insert and this insert was subcloned into pUC8 to create the plasmid pEndol6 and into m13mp18 for restriction mapping and sequence analysis. Sequencing was performed using the Sequenase kit (US Biochemicals) according to the man- ufacturer’s instructions. Asymmetric probes were pre- pared using standard sequencing reactions on Northern blot strips which were hybridized, washed, and exposed as described below.
Genomic Southern blot. Sperm was collected from a single spawned animal and DNA isolated by the method of Kedes et al. (1975). Ten micrograms of genomic DNA was digested with EcoRI or HindIII, electrophoresed on a 0.65% agarose gel, and blotted to nitrocellulose. The filter was hybridized overnight with nick-translated End016 at 65°C in 5X SSC, 5X Denhardt’s, 0.1% SDS. Washing was done at 65°C in 1X SSC, 0.1% SDS. Auto- radiography was for 24 hr using Kodak XAR-5 film and a DuPont Cronex intensifying screen.
Northern blot analysis. RNA from egg, 16-cell, 128- cell, 20-hr hatched blastula, 26-hr mesenchyme blas- tula, 36-hr gastrula, and 80-hr pluteus embryonic stages and adult tissues was prepared by the procedure of Chirgwin et al. (1979). Ten micrograms total RNA was electrophoresed on 1% agarose/formaldehyde gels and blotted to nitrocellulose. Nick-translated End016 and a control probe were used to probe filters. Hybridization and washing conditions were as for the genomic blot. Autoradiography was for 48 hr using Kodak XAR-5 film and a DuPont Cronex intensifying screen.
In situ hybridizations. The 550-bp EcoRl-HpaII frag- ment from pEndol6 was subcloned into the Sp6/T7 transcription vector pGem-7Zf (Promega Biotech). This construct was used to synthesize either sense or anti- sense RNA probes with the Riboprobe kit (Promega Biotech) according to the manufacturer’s instructions
with [35S]CTP (800 CVmmole, Amersham). Probes were synthesized to a specific activity of -1 X lo* cpm/pg. The final concentration of probe in the hybridization solution was 3.3 X lo6 cpm/ml.
Embryos were fixed, embedded in paraplast, sec- tioned, and spread onto polylysine-coated slides as de- scribed by Cox et al. (1984). Hybridization and washing were performed according to the modifications of Har- din et al. (1988) for 35S probes except that no dehydra- tion through a graded ethanol series was done preced- ing the formamide wash and 2X SSC was substituted for 0.3 M NaC1/20 mM Tris-HCl (pH 8.0). Slides were processed for autoradiography as described (Angerer and Angerer, 1981). Exposure time was 3 days for all sections shown, except pluteus stage which was for 9 days.
Fusion protein production and puri~cation. To con- struct the Trp-End016 fusion protein, the 550-bp EcoRI-HpaII fragment of the End016 cDNA was li- gated into EcoRI-CZaI digested pATH1 (Dieckmann and Tzagoloff, 1985) and transformed into Escherichia coli strain RRI. This arrangement fuses the End016 frag- ment in frame with the truncated E. coli TrpE gene under the control of the Trp promoter/operator se- quences. Induction of the fusion protein was accom- plished by growing cells overnight in M9 media supple- mented with 0.5% casamino acids, 20 pg/ml L-trypto- phan, and 50 pg/ml ampicillin. The cells were then diluted 1:lO in the same media minus tryptophan and grown for 1 hr at 30°C with vigorous aeration. At this time 5 pg/ml indoleacrylic acid (Sigma) was added to the cultures and incubation continued for 2 hr. For so- dium dodecyl sulfate-polyacrylamide gel (SDS-PAGE) analysis, 1 ml of culture was pelleted and resuspended in 50 ~1 of sample buffer (Laemmli, 1970). Ten micro- liters of sample was loaded per lane. The fusion protein was purified by preparative gel electrophoresis. Gels were stained briefly in 4 M Na acetate (Higgins and Dahmus, 1979) and bands were excised and eluted by crushing and soaking overnight in 3 vol of 1 mM Tris- HCl, pH 7.5,0.1 mMEDTA, 0.05% SDS. The sample was dialyzed against 10 mMTris-HCl, pH 7.5,150 mMNaC1, 1 mM EDTA, precipitated, and resuspended in phos- phate-buffered saline (PBS). Protein concentrations were determined using the Bio-Rad protein assay kit.
The P-gal-End016 fusion protein was constructed by ligating the 794-bp End016 cDNA into EcoRI cut pUR288 expression plasmid (Ruther and Mtiller-Hill, 1983) and transformed into E. coli strain JMlOl. Induc- tion of the fusion protein was accomplished by adding 0.5 mM isopropylthio-fi-galactoside and growing at 37°C for l-3 hr, and resulted in over production of a protein at the expected -150 kDa size (data not shown).
Polyclonal antiserum. Preimmune serum was col-
266 DEVELOPMENTALBIOLOGY V0~~~~136,1989
lected from the rabbit before antigen injections began. The primary immunization consisted of 200 pg TrpE- End016 fusion protein emulsified in Freund’s complete adjuvant injected intradermally. The animal was given two booster injections at 3-week intervals consisting of 150 pg TrpE-End016 fusion protein emulsified in Freund’s incomplete adjuvant.
Western blot analysis. Total embryo protein lysates were prepared as described (Wessel and McClay, 1987); 25 pg of lysate was loaded per lane of 10% SDS-PAGE and electroblotted to nitrocellulose (Towbin et ah, 1979). Filters were blocked in 5% dry milk/PBS for 1 hr, in- cubated in 1:lOOO dilution of preimmune or immune serum in PBS for 1 hr, washed in 5% dry milk/PBS for 1 hr, incubated in 1:‘7500 dilution of goat anti-rabbit:al- kaline phosphatase conjugate (Promega Biotech) for 1 hr, and washed again in 5% dry milk/PBS for 1 hr. The reaction was visualized by incubating in alkaline phos- phatase substrate according to the manufacturer’s in- structions (Promega Biotech).
Indirect immunoJuorescence. Embryos were settled onto polylysine-coated coverslips and fixed in 90% methanol/seawater on ice for 30 min. Embryos were then rehydrated to PBS through a graded methanol series. Embryos were incubated in 3% BSA/PBS for 15 min, 1:lOO dilution of immune serum in PBS for 1 hr, 3% BSA/PBS 1 hr, 1:lOO dilution of fluorescein-conjugated goat anti-rabbit antiserum (Polysciences) 1 hr, and 3% BSA/PBS 1 hr. The reaction was viewed and photo- graphed under a Zeiss microscope equipped for epi- fluorescence using Kodak T-Max film at an ASA setting of 800.
EXPERIMENTAL RESULTS
Cloning and Sequence Analysis of the Endo cDNA
The 794-bp End016 cDNA clone was selected from a sea urchin gastrula stage XgtlO cDNA library on the basis of low stringency hybridization to a C. elegans collagen type 1 probe. The restriction map and genomic representation of this clone are shown in Figs. la and lb. The End016 cDNA probe hybridizes to a single DNA fragment in both EcoRI and HindIII-digested sea ur- chin genomic DNA, indicating that the End016 mRNA is transcribed from a single copy gene (Fig. lb). The transcriptional orientation of this gene was determined by hybridizing strand-specific End016 probes to North- ern blot strips of gastrula stage RNA. Only one of these two probes reacts with an mRNA transcript, 6.6 kb in length (Fig. lc). Sequence analysis of the cDNA clone revealed that the entire 794-bp insert is an open reading frame which could encode 265 amino acids (Fig. 2A). Although the DNA sequence of the sea urchin clone does contain short regions of up to 65% similarity to the
C. elegans gene (data not shown), the predicted amino acid sequence of the sea urchin clone does not contain any collagen-related sequence, including that of re- cently reported sea urchin collagens (Venkatesan et al., 1986; Saitta et al., 1989). However, the deduced amino acid sequence does display several noteworthy features. The sequence includes an Arg-Gly-Asp (RGD; boxed in Fig. 2A), which has been shown to be important in the cell binding domain of many extracellular proteins (re- viewed in Ruoslahti and Pierschbacher, 1987). Two po- tential N-linked glycosylation sites are present (under- lined in Fig. 2A), although the proline containing site is probably not utilized (Hubbard and Ivat, 1981). In ad- dition the sequence contains six clustered copies of a 20-amino acid repeat (Fig. 2B), two of which are in tan- dem. Overall, this protein segment is very hydrophilic and extremely acidic, comprised of 30% glutamic and aspartic acid residues. A computer-assisted search, using the algorithm of Lipman and Pearson (1985), has not identified significant similarity of End016 with any sequences contained in the GenBank or EMBL nucleic acid databases, nor is the deduced amino acid sequence related to any protein sequence compiled in the NBRF or Institut Pasteur databases.
Temporal and Spatial Expression of Endo Transcript during Embryogenesis
Northern blot analysis was used to examine the de- velopmental expression pattern of the End016 mRNA transcript. The results of this experiment are shown in Fig. 3. The 794-bp cDNA hybridizes to a single 6.6-kb transcript that is not detected in egg or morula stages, but is first detected at the hatched blastula stage. The transcript increases in abundance by the mesenchyme blastula stage, continues to increase during gastrula- tion, and declines in abundance by the time the forma- tion of the pluteus larva is complete. The End016 tran- script was not detected in any adult tissue examined, including testis, ovary, intestine, coelomocyte, and tube foot RNAs. A reprobing of this blot with a sea urchin actin probe demonstrated the integrity of all the RNA samples (data not shown).
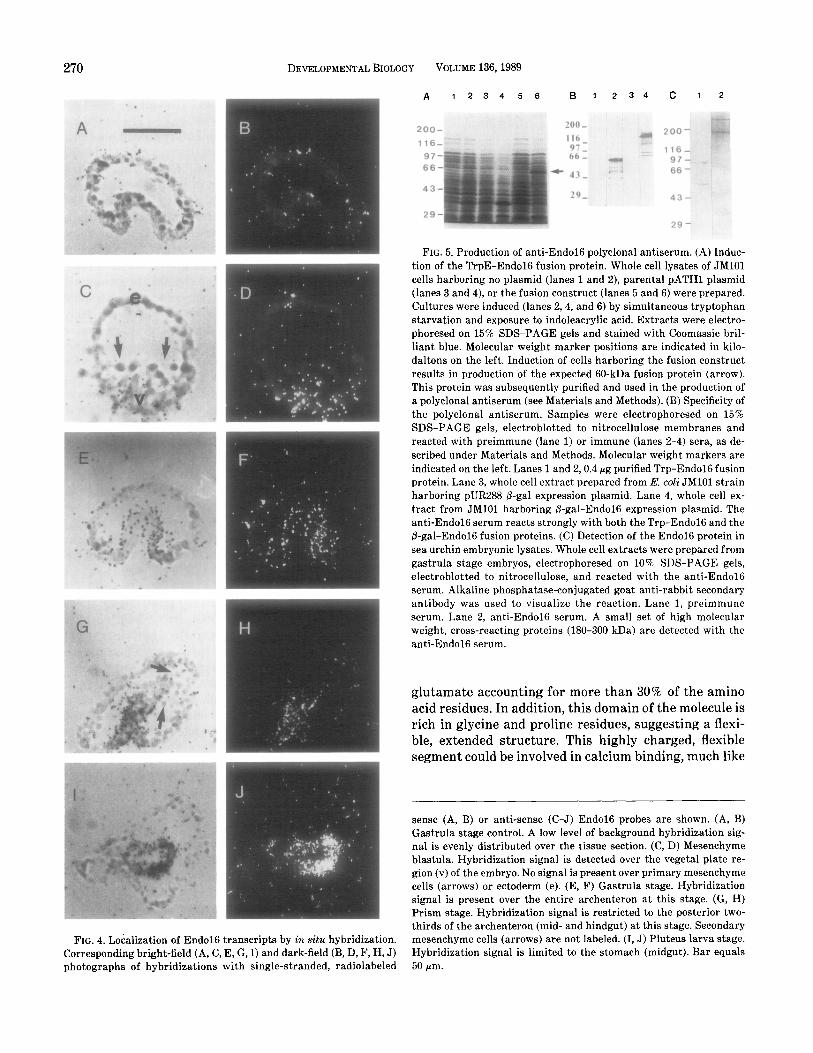
The embryonic distribution of End016 transcripts during development was determined by in situ hybrid- ization. The results of these in situ hybridizations with radioactively labeled End016 antisense probes and de- velopmentally staged embryo sections are shown in Fig. 4. In 28-hr mesenchyme blastula embryos, the autora- diographic signal is concentrated over the vegetal plate region. These cells are poised to invaginate in the first movement of gastrulation to form the archenteron, which later gives rise to the endoderm and secondary mesenchyme tissues. No signal above background is de-
NOCENTE-MCGRATH, BRENNER, AND
a
ERNST Sea Urchin Lineage-Specific Protein 267
b 1 2 c 1 2
~CoRl pieI
I
?glll Awl ECORI
-1 I
pal ptfhll
I I
/ \
Taql Taql Hincll Taql Taql
toobp
2.3 - 2.0- : *
FIG. 1. Characterization of the End016 cDNA clone. (a) Restriction map of pEndo16. Open box corresponds to the sea urchin cDNA insert; flanking plasmid sequences are represented as thin lines. Arrow indicates direction of transcription. (b) Genomic Southern blot. Ten micro- grams of sea urchin sperm DNA was digested with either EcoRI (lane 1) or Hind111 (lane 2), electrophoresed, transferred to nitrocellulose, and probed with radiolabeled pEndo16. A single hybridizing fragment is detected in each digest. (c) Determination of transcriptional orientation. Strand-specific probes were prepared and hybridized to Northern blot strips containing 10 pg of total gastrula stage RNA as described under Materials and Methods. Only one of the probes (lane 1) detected a transcript, 6.6 kb in length.
tected over primary mesenchyme or ectodermal cells. In 36-hr gastrula stage embryos the End016 mRNA is de- tected in cells in all regions of the archenteron. By the 72-hr prism stage, the archenteron has begun constric- tion to form a tripartite larval gut. At this stage, hy- bridization is restricted to the posterior two-thirds of the gut (mid- and hindgut) which will become the larval stomach and intestine. No signal is detected over sec- ondary mesenchyme cells at this stage. Interestingly, hybridization to 90-hr pluteus stage larvae shows label- ing of stomach only. We did not obtain any tissue sec- tions in which all three digestive tract regions were clearly identifiable. However, sections which included esophagus and stomach or stomach and intestine all showed labeling of stomach tissue exclusively. A repre- sentative example of control hybridizations with End016 sense probes is shown in Figs. 4A and 4B. In these controls, a low level of background signal is pres- ent evenly distributed over the tissue sections. These results demonstrate that the End016 gene is expressed in a spatially regulated, lineage-specific manner and that this expression becomes restricted during differ- entiation.
Production and Specificity of Polyclonal Antiserum
To facilitate study of the End016 gene product, poly- clonal antibodies were generated. A bacterial fusion
protein was produced using one of the PATH series of plasmid vectors and employed as the source of antigen. The plasmids contain a truncated E. coli trpE gene with a synthetic polylinker at the 3’ end. A 550-bp restriction fragment of the End016 cDNA was subcloned into the polylinker of pATH1 so that the correct translational reading frame was preserved. Induction of the trp pro- moter by simultaneous tryptophan starvation and ex- posure to indoleacrylic acid results in the production of large quantities of the trpE fusion protein in cultures harboring the expression plasmid. Protein extracts from induced and uninduced cultures were analyzed by SDS-PAGE, and the appropriately sized fusion protein band was observed after staining with Coomassie blue (Fig. 5A). The fusion protein was purified by prepara- tive gel electrophoresis and used for immunization of a Dutch Belted rabbit.
To determine the reactivity of the resulting anti- serum against the sea urchin portion of the fusion pro- tein, we produced a second fusion protein using the pUR series of plasmid vectors. These vectors allow the ex- pression of protein coding sequence as C-terminal ad- dition to E. coli ,&galactosidase. Induction of the P-gal- End016 fusion protein resulted in overproduction of a protein of the expected -150-kDa size as judged by Coomassie blue staining of whole cell extracts electro- phoresed on SDS-PAGE (data not shown). Figure 5B shows the results of Western blot experiments reacting
FIG. 2. Sequence analysis of Endol6. (A) Nucleotide sequence and derived amino acid sequence of the End016 cDNA. The nucleotide sequence of the coding strand (as determined by the strand-specific hybridization, Fig. lc) is shown. The entire 794-bp cDNA is an open reading frame encoding 265 amino acids. The sequence includes a potential cell attachment site (RGD, boxed) as well as two potential N-linked glycosylation sites (N x S/T, underlined). (B) End016 repeats. Six copies of a 20-amino acid repeat are present in the End016 sequence. Exact matches that are present in at least three repeats are boxed.
the anti-End016 serum with both fusion proteins. As anticipated, the serum reacts strongly with the Trp- End016 protein used in the immunization protocol. The serum also reacts very strongly with the P-gal-End016 fusion protein, indicating that the sea urchin portion of the Trp-End016 fusion protein elicited a strong immune response. The lower molecular weight bands present must be due to breakdown products of the fusion pro- tein since they are not present in the lane containing the parent plasmid. Therefore, the majority of the im- mune response was elicited against the Trp-End016 fu- sion protein and little response was directed at any con- taminants present in the preparation used for immuni- zation.
To investigate the expression of the End016 gene product in sea urchin embryos, the polyclonal anti- serum was reacted with total sea urchin gastrula stage protein extracts in Western blot experiments. Preim- mune serum, collected from the animal before immuni- zation, was used as a control. As shown in Fig. 5C, the anti-End016 serum reacts with a small set of high mo-
lecular weight proteins, ranging from 180 to >300 kDa. These bands are not observed in the lane reacted with preimmune serum. It remains to be determined whether the multiple bands are the result of degrada- tion, post-translational modification, or common epi- topes shared by different proteins. The 6.6-kb End016 mRNA can potentially encode a primary translation product with a maximum size of 240-250 kDa. There- fore, degradation in the protein sample cannot account for the higher molecular weight bands observed.
Immunolocalixation of the End016 Gene Product in Gastrula Stage Embryos
We reacted the anti-End016 serum with fixed, whole, midgastrula stage embryos in an effort to determine the spatial and cellular distribution of the End016 gene product. The reaction was visualized with fluorescein- conjugated goat anti-rabbit secondary antibody and observed with a fluorescence microscope. The results of this experiment, shown in Fig. 6, reveal that the End016
NOCENTE-MCGRATH, BRENNER, AND ERNST Sea Urchin Lineage-Speci$c Protein 269
E16MHBG POT I FC
FIG. 3. Northern blot analysis of End016 expression. Ten micro- grams of total RNA prepared from various developmental stages and adult tissues was eleetrophoresed, blotted onto nitrocellulose, and hybridized to radiolabeled pEndol6 as described under Materials and Methods. E, egg; 16, 5-hr 16-cell stage; M, 128-cell morula; H, 20-hr hatched blastula; B, 26-hr mesenchyme blastula; G, 36-hr gastrula; P, 80-hr pluteus larva; 0, ovary; T, testis; I, intestine; F, tube foot; C, coelomocyte. The 6.6-kb End016 transcript is first detected at the hatched blastula stage, increases dramatically in abundance through the gastrula stage, and declines in abundance by the pluteus larva stage. An additional higher molecular weight RNA transcript is oc- casionally observed in some RNA preparations (lane B). The End016 transcript is not detected in any of the adult tissue RNAs.
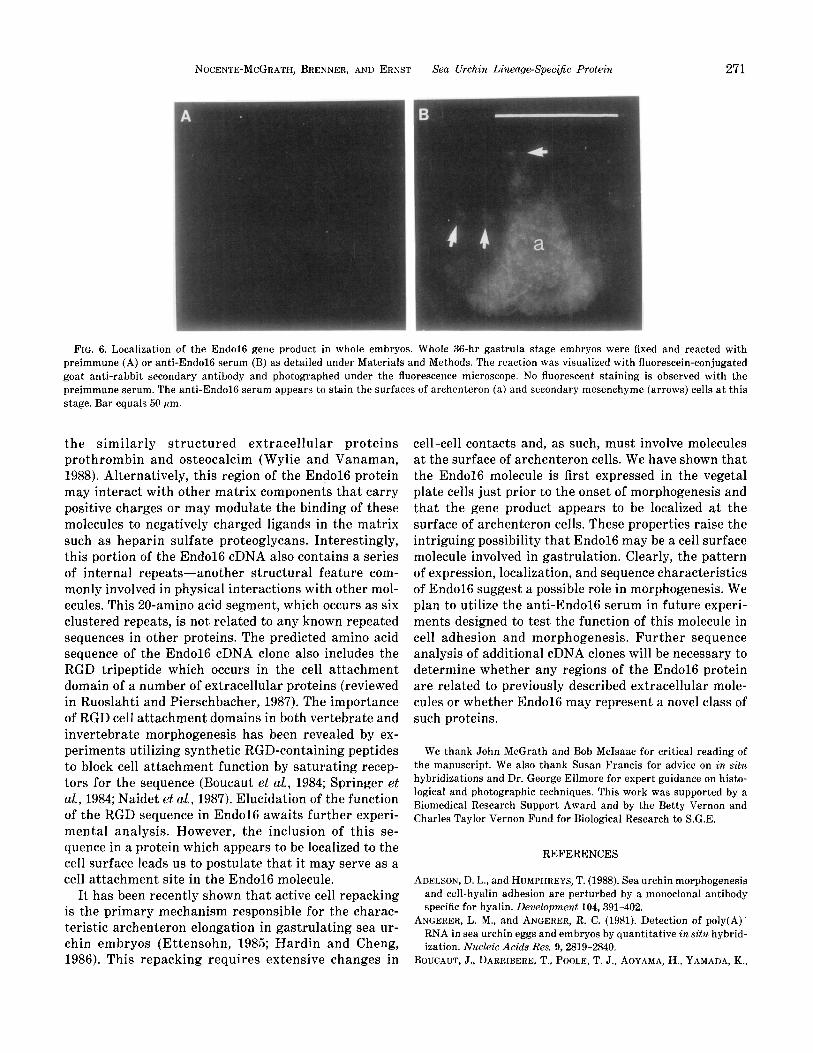
protein is ,localized to the surface of endodermal cells of the archenteron and secondary mesenchyme cells. The immunolocalization of the End016 protein to the sur- face of endodermal cells is corroborated by the in situ hybridization experiments, which detected significant levels of End016 transcripts in these cells. However, although the End016 protein appears to be present on the surface of secondary mesenchyme cells at midgas- trula stage (arrows in Fig. 6), these cells do not contain detectable End016 transcripts (Figs. 4E and 4F). There are several possible explanations to account for the ob- servation of End016 protein in association with these cells. At earlier stages of development, our in situ hy- bridization experiments have shown that the End016 gene is actively transcribed in vegetal plate cells and the archenteron, which includes the precursors of the secondary mesenchyme. Long-lived End016 protein, synthesized by the precursor cells, may persist in the cells through later stages of development. Alterna- tively, endodermal cells may be secreting the End016 protein, which subsequently binds to the surface of sec- ondary mesenchyme cells by means of specific receptors expressed by the secondary mesenchyme.
DISCUSSION
The results of Northern blot experiments demon- strate that the End016 gene is a member of the class of
so-called “embryonic late genes” (Davidson, 1986) de- fined as not being significantly expressed in oocytes but activated during early development. It is estimated that not more than 10% of all sequences utilized during early sea urchin development fall into this class (Flyt- zanis et ah, 1982). This pattern of expression is charac- teristic of spatially regulated genes (Davidson, 1986; Sargent et al, 1986) and in situ hybridization results confirm that End016 is expressed in a spatially regu- lated manner. Moreover, the gene is first transcribed in a group of endodermal and secondary mesenchymal precursor cells and transcription becomes progressively restricted to a subset of endoderm cells as development proceeds. Within the limits of resolution of our tech- nique, we do not observe any differential expression of End016 among vegetal plate cells at early stages; how- ever, we cannot completely rule out this possibility. In- terestingly, the transcript has not been detected in any adult tissues examined, indicating that the function of the End016 gene product may be specific to embryo- genesis.
Several endoderm-specific markers have been identi- fied previously by M&lay and colleagues using mono- clonal antibodies generated against the insoluble frac- tion of sea urchin plutei and indirect immunofluores- cence. Some of these antigens, for example, those recognized by McAbAAla3 and McAbLLEif7 (McClay and Wessel, 1984), are first detected in all regions of the egg and early embryo and only become localized to the endoderm later in development. Others, such as those recognized by McAb7B3, McAb4BlO (M&lay and Et- tensohn, 1987), and the antigens designated Meso 2 and Endo 1 (Wessel and M&lay, 1985), are already localized to the vegetal plate or endoderm when first detected at mesenchyme blastula or very early gastrula stages. A recently isolated endoderm-specific cDNA clone, desig- nated Lv1.4 (G. Wessel, personal communication), is first detected in early gastrula stage embryos and is present only in the mid- and hindgut regions of the embryo at all stages of development. The pattern of End016 expression that we have observed is unique among endoderm-specific molecules; nevertheless it does share a feature common to the majority of endo- dermal markers described to date. With the exceptions of Endo 1 and Lv1.4, expression of endoderm-specific markers becomes progressively restricted to a subset of vegetal plate derivatives as the archenteron differen- tiates into a tripartite gut and secondary mesenchyme.
The portion of the End016 gene product encoded by the 794-bp cDNA displays several unique structural features which may provide clues to the biological function of the molecule. Overall, this region of the predicted End016 protein contains a very high density of negatively charged amino acids, with aspartate and
270 DEVELOPMENTAL BIOLOGY VOLUME 136,1989
FIG. 4. Localization of End016 transcripts by in situ hybridization. Corresponding bright-field (A, C, E, G, I) and dark-field (B, D, F, H, J) photographs of hybridizations with single-stranded, radiolabeled
A 1 23456 B 1 234 C 12
200-
116-
97-
66-
43- 43-
29-
29-
FIG. 5. Production of anti-End016 polyclonal antiserum. (A) Induc- tion of the TrpE-End016 fusion protein. Whole cell lysates of JMlOl cells harboring no plasmid (lanes 1 and 2), parental pATH1 plasmid (lanes 3 and 4), or the fusion construct (lanes 5 and 6) were prepared. Cultures were induced (lanes 2,4, and 6) by simultaneous tryptophan starvation and exposure to indoleacrylic acid. Extracts were electro- phoresed on 15% SDS-PAGE gels and stained with Coomassie bril- liant blue. Molecular weight marker positions are indicated in kilo- daltons on the left. Induction of cells harboring the fusion construct results in production of the expected 60-kDa fusion protein (arrow). This protein was subsequently purified and used in the production of a polyclonal antiserum (see Materials and Methods). (B) Specificity of the polyclonal antiserum. Samples were electrophoresed on 15% SDS-PAGE gels, electroblotted to nitrocellulose membranes and reacted with preimmune (lane 1) or immune (lanes 2-4) sera, as de- scribed under Materials and Methods. Molecular weight markers are indicated on the left. Lanes 1 and 2,0.4 pg purified Trp-End016 fusion protein. Lane 3, whole cell extract prepared from E. coli JMlOl strain harboring pUR288 P-gal expression plasmid. Lane 4, whole cell ex- tract from JMlOl harboring P-gal-End016 expression plasmid. The anti-End016 serum reacts strongly with both the Trp-End016 and the &gal-End016 fusion proteins. (C) Detection of the End016 protein in sea urchin embryonic lysates. Whole cell extracts were prepared from gastrula stage embryos, electrophoresed on 10% SDS-PAGE gels, electroblotted to nitrocellulose, and reacted with the anti-End016 serum. Alkaline phosphatase-conjugated goat anti-rabbit secondary antibody was used to visualize the reaction. Lane 1, preimmune serum. Lane 2, anti-End016 serum. A small set of high molecular weight, cross-reacting proteins (180-300 kDa) are detected with the anti-End016 serum.
glutamate accounting for more than 30% of the amino acid residues. In addition, this domain of the molecule is rich in glycine and proline residues, suggesting a flexi- ble, extended structure. This highly charged, flexible segment could be involved in calcium binding, much like
-
sense (A, B) or anti-sense (C-J) End016 probes are shown. (A, B) Gastrula stage control. A low level of background hybridization sig- nal is evenly distributed over the tissue section. (C, D) Mesenchyme blastula. Hybridization signal is detected over the vegetal plate re- gion (v) of the embryo. No signal is present over primary mesenchyme cells (arrows) or ectoderm (e). (E, F) Gastrula stage. Hybridization signal is present over the entire archenteron at this stage. (G, H) Prism stage. Hybridization signal is restricted to the posterior two- thirds of the archenteron (mid- and hindgut) at this stage. Secondary mesenchyme cells (arrows) are not labeled. (I, J) Pluteus larva stage. Hybridization signal is limited to the stomach (midgut). Bar equals 50 pm.
NOCENTE-MCGRATH, BRENNER, AND ERNST Sea Urchin Lineage-Speci& Protein 271
FIG. 6. Localization of the End016 gene product in whole embryos. Whole 36-hr gastrula stage embryos were fixed and reacted with preimmune (A) or anti-End016 serum (B) as detailed under Materials and Methods. The reaction was visualized with fluorescein-conjugated goat anti-rabbit secondary antibody and photographed under the fluorescence microscope. No fluorescent staining is observed with the preimmune serum. The anti-End016 serum appears to stain the surfaces of archenteron (a) and secondary mesenchyme (arrows) cells at this stage. Bar equals 50 pm.
the similarly structured extracellular proteins prothrombin and osteocalcim (Wylie and Vanaman, 1988). Alternatively, this region of the End016 protein may interact with other matrix components that carry positive charges or may modulate the binding of these molecules to negatively charged ligands in the matrix such as heparin sulfate proteoglycans. Interestingly, this portion of the End016 cDNA also contains a series of internal repeats-another structural feature com- monly involved in physical interactions with other mol- ecules. This 20-amino acid segment, which occurs as six clustered repeats, is not related to any known repeated sequences in other proteins. The predicted amino acid sequence of the End016 cDNA clone also includes the RGD tripeptide which occurs in the cell attachment domain of a number of extracellular proteins (reviewed in Ruoslahti and Pierschbacher, 1987). The importance of RGD cell attachment domains in both vertebrate and invertebrate morphogenesis has been revealed by ex- periments utilizing synthetic RGD-containing peptides to block cell attachment function by saturating recep- tors for the sequence (Boucaut et al., 1984; Springer et al., 1984; Naidet et al, 1987). Elucidation of the function of the RGD sequence in End016 awaits further experi- mental analysis. However, the inclusion of this se- quence in a protein which appears to be localized to the cell surface leads us to postulate that it may serve as a cell attachment site in the End016 molecule.
It has been recently shown that active cell repacking is the primary mechanism responsible for the charac- teristic archenteron elongation in gastrulating sea ur- chin embryos (Ettensohn, 1985; Hardin and Cheng, 1986). This repacking requires extensive changes in
cell-cell contacts and, as such, must involve molecules at the surface of archenteron cells. We have shown that the End016 molecule is first expressed in the vegetal plate cells just prior to the onset of morphogenesis and that the gene product appears to be localized at the surface of archenteron cells. These properties raise the intriguing possibility that End016 may be a cell surface molecule involved in gastrulation. Clearly, the pattern of expression, localization, and sequence characteristics of End016 suggest a possible role in morphogenesis. We plan to utilize the anti-End016 serum in future experi- ments designed to test the function of this molecule in cell adhesion and morphogenesis. Further sequence analysis of additional cDNA clones will be necessary to determine whether any regions of the End016 protein are related to previously described extracellular mole- cules or whether End016 may represent a novel class of such proteins.
We thank John McGrath and Bob McIsaac for critical reading of the manuscript. We also thank Susan Francis for advice on in situ hybridizations and Dr. George Ellmore for expert guidance on histo- logical and photographic techniques. This work was supported by a Biomedical Research Support Award and by the Betty Vernon and Charles Taylor Vernon Fund for Biological Research to S.G.E.
REFERENCES
ADELSON, D. L., and HUMPHREYS, T. (1988). Sea urchin morphogenesis and cell-hyalin adhesion are perturbed by a monoclonal antibody specific for hyalin. Development 104, 391-402.
ANGERER, L. M., and ANGERER, R. C. (1981). Detection of poly(A)+ RNA in sea urchin eggs and embryos by quantitative in situ hybrid- ization. Nucleic Acids Res. 9,2819-2840.
BOIJCAUT, J., DARRIBERE, T., POOLE, T. J., AOYAMA, H., YAMADA, K.,
272 DEVELOPMENTAL BIOLOGY VOLUME 136,1989
and THIERY, J. P. (1984). Biologically active peptides as probes of embryonic development: A competitive peptide inhibitor of fibro- nectin function inhibits gastrulation in amphibian embryos and neural crest cell migration in avian embryos. J. Cell Biol. 99,
1822-1830.
BUTLER, E., HARDIN, J., and BENSON, S. (1987). The role of lysyl oxi- dase and collagen crosslinking during sea urchin development. Exp. Cell Res. 173,174-182.
CHIRGWIN, J. M., PRZYBYLA, A. E., MACDONALD, R. J., and RUTTER, W. J. (1979). Isolation of biologically active RNA from sources enriched in ribonucleases. Biochemistry l&5294-5299.
Cox, K. H., DELEON, D. V., and ANGERER, R. C. (1984). Detection of mRNAs in sea urchin embryos by in situ hybridization using asym- metric RNA probes. Dev. Biol. 101,485-502.
DAVIDSON, E. (1986). “Gene Activity in Early Development.” 3rd ed. Academic Press, Orlando, FL.
DIECKMANN, C. L., and TZAGOLOFF, A. (1985). Assembly of the mito- chondrial membrane system. CBPG, a yeast nuclear gene necessary for synthesis of cytochrome b. J. Biol. Chem. 260,1513-1520.
ETTENSOHN, C. A. (1985). Gastrulation in the sea urchin embryo is accompanied by the rearrangement of invaginating epithelial cells. Dev. Biol. 112,383-390.
FLYTZANIS, C. N., BRANDHORST, B. R., BRITTEN, R. J., and DAVIDSON, E. H. (1982). Developmental patterns of cytoplasmic transcript prevalence in sea urchin embryos. Dev. Biol. 91,27-37.
GUSTAFSON, G., and WOLPERT, L. (1967). Cellular movement and con- tact in sea urchin morphogenesis. Biol. Rev. 42,442-498.
HARDIN, J. D., and CHENG, L. Y. (1986). The mechanisms and me- chanics of archenteron elongation during sea urchin gastrulation. Dev. Biol. 115,490-501.
HARDIN, P. E., ANGERER, L. M., HARDIN, S. H., ANGERER, R. C., and KLEIN, W. H. (1988). Spec 2 genes of Strongylocentrotus purpuratus. Structure and differential expression in embryonic aboral ecto- derm cells. J. Mol. Biol. 202, 417-431.
HEIFEITZ, A., and LENNARZ, W. J. (1979). Biosynthesis of N-glucosidi- tally linked glycoproteins during gastrulation of sea urchin em- bryos. J. Biol. Chem. 254,6119-6127.
HIGGINS, R. C., and DAHMUS, M. E. (1979). Rapid visualization of protein bands in preparative SDS-polyacrylamide gels. Anal. Bia them. 93,257-260.
HUBBARD, S. C., and IVATT, R. J. (1981). Synthesis and processing of asparagine-linked oligosaccharides. Annu. Rev. Biochem. 50, 555-583.
KARP, G. C., and SOLURSH, M. (1974). Acid mucopolysaccharide me- tabolism, the cell surface, and primary mesenchyme cell activity in the sea urchin embryo. Dev. Biol. 41,110-123.
KATOW, H., and SOLURSH, M. (1979). Ultrastructure of blastocoel ma- terial in blastulae and gastrulae of the sea urchin Lytechinus pictus. J. Exp. 2001. 210, 561-567.
KEDES, L. H., COHN, R. H., LOWRY, J. C., CHANG, A. C. Y., and COHEN, S. N. (1975). The organization of sea urchin histone genes. Cell 6, 359-369.
KELLER, R., and HARDIN, J. (1987). Cell behavior during active cell rearrangement: Evidence and speculations. In “Cell Behavior: Shape, Adhesion and Motility.” J. Cell Sci., S,(Suppl.).
KRAMER, J. M., Cox, G. N., and HIRSCH, D. (1982). Comparisons of the complete sequences of two collagen genes from Caenorhabditis ele- gans. Cell 30, 599-606.
LAEMMLI, U. K. (1970). Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature (London) 227, 680-685.
LIPMAN, D. J., and PEARSON, W. R. (1985). Rapid and sensitive protein similarity searches. Science 227,1435-1441.
MANIATIS, T., FRITSCH, E., and SAMBROOK, J. (1982). “Molecular Clon- ing: A Laboratory Manual.” Cold Spring Harbor Laboratory, Cold Spring Harbor, NY.
MCCLAY, D. R., and ETTENSOHN, C. A. (1987). Cell recognition during sea urchin gastrulation. In “Genetic Regulation of Development,” (William F. Loomis, Ed.), 111-128. A. R. Liss, New York.
MCCLAY, D. R., and MARCHASE, R. B. (1979). Separation of ectoderm and endoderm from sea urchin pluteus larvae and demonstration of germ layer specific antigens. Dev. Biol. 71,289-296.
MCCLAY, D. R., and WESSEL, G. M. (1984). Spatial and temporal ap- pearance and redistribution of cell surface antigens during sea ur- chin development. In “Molecular Biology of Development” (R. Fir- tel and E. Davidson, Eds.), pp. 165-184. A. R. Liss, New York.
MCCLAY, D. R., and WESSEL, G. M. (1985). The surface of the sea urchin embryo at gastrulation: A molecular mosaic. TIGS 1.12-16.
NAIDET, C., SEMERIVA, M., YAMADA, K., and THIERY, J. P. (1987). Peptides containing the cell-attachment recognition signal Arg- Gly-Asp prevent gastrulation in Drosophila embryos. Nature (Lan- don) 325,348-350.
PITTMAN, D., and ERNST, S. G. (1984). Developmental time, cell lin- eage, and environment regulate the newly synthesized proteins in sea urchin embryos. Dev. Biol. 106,236-242.
RUOSLAHTI, E., and PIERSCHBACHER, M. D. (1987). New perspectives in cell adhesion: RGD and integrins. Science 238,491-497.
RIJTHER, C., and MULLER-HILL, B. (1983). Easy identification of cDNA clones. EMBO J. 2,1791-1794.
SAITTA, B., BUTTICE, G., and GAMBINO, R. (1989). Isolation of a puta- tive collagen-like gene from the sea urchin Paracentrotus lividus. Biochem. Biophys. Res. Commun. 158,633-639.
SARGENT, T. D., JAMRICH, M., and DAWID, I. B. (1986). Cell interac- tions and the control of gene activity during early development of Xenopus laevis. Dev. Biol. 114, 238-246.
SPRINGER, W. R., COOPER, D. N. W., and BARONDES, S. H. (1984). Discoidin 1 is implicated in cell-substratum attachment and or- dered cell migration in Dictyostelium discoideum and resembles fibronectin. Cell 39, 557-564.
TOWBIN, H., STAEHELIN, T., and GORDON, J. (1979). Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: Procedure and some applications. Proc. Natl. Acad. Sci. USA 76,4350-4354.
VENKATESAN, M., DEPABLO, F., VOGELI, G., and SIMPSON, R. T. (1986). Structure and developmentally regulated expression of a Strongy- locentrotus purpuratus collagen gene. Proc. Natl. Acad. Sci. USA 83, 3351-3355.
WESSEL, G. M., and MCCLAY, D. R. (1985). Sequential expression of germ-layer specific molecules in the sea urchin embryo. Dev. Biol. 111,451-463.
WESSEL, G., and MCCLAY, D. (1987). Gastrulation in the sea urchin embryo requires the deposition of crosslinked collagen within the extracellular matrix. Dev. Biol. 121,149-165.
WYLIE, D. C., and VANAMAN, T. C. (1988). Structure and function of the calmodulin family of calcium regulatory proteins. In “Calmo- dulin” (P. Cohen and C. Biklee, Eds.). Elsevier, New York.